Область техники
Настоящее изобретение относится к новому способу детоксикации материала, зараженного трихотеценом. Более конкретно, настоящее изобретение относится к способу биотрансформации трихотецена путем приведения материала, зараженного трихотеценами, в контакт с экзогенной глутатион-S-трансферазой (GST) неживотного происхождения, обладающей субстратной специфичностью к эпоксидному кольцу трихотецена. Настоящее изобретение также относится к рекомбинантным GST и трансгенным растениям и животным, экспрессирующим указанные GST.
Уровень техники
Грибы производят большое количество метаболитов, которые не являются необходимыми для жизни, но могут обеспечить грибу экологическое преимущество в определенных условиях. Такие метаболиты называются вторичными метаболитами. Вторичные метаболиты грибов включают регуляторы роста растений (например, гиббереллины), фармацевтически полезные соединения (например, пенициллин, ловастатин), пигменты (например, каротиноиды) и микотоксины (например, трихотецены, фумонизины, афлатоксины, охратоксины).
Трихотеценовые микотоксины представляют собой большое семейство химически родственных микотоксинов с общей структурой трициклических 12, 13-эпокситрихотец-9-ен (ЕРТ) ядер, которые токсичны для людей, животных, растений и эукариотических клеток в целом. Фузариум и другие грибы и плесени, продуцирующие трихотецен, такие как Fusarium, Myrochecium, Trichoderma, Trichothecium, Cephalosporium, Verticimonosporium и Stachybotrys, заражают важные культурные растения, например, зерна пшеницы, ячменя или маиса. В результате они стали ассоциироваться с интоксикациями людей и животных во всем мире (Arunachalam С, Doohan FM., Toxicol Lett. 2013 Feb 27; 217(2): 149-58. doi: 10.1016/j.toxlet.2012.12.003. Epub 2012 Dec 26. Review). Трихотецены обладают сильной цитотоксичностью, а также провоспалительными и эметогенными свойствами, и наносят вред кроветворным органам и иммунной функции. Трихотецены ингибируют синтез эукариотического белка, в частности, предотвращая образование пептидных связей в пептидилтрансферазном центре субъединицы рибосомы 60S, ингибируя синтез митохондриального белка и взаимодействуя с сульфгидрильными группами белков. Токсины фузариума могут присутствовать во многих типах продуктов питания человека и животных, включая зерновые культуры, такие как ячмень, овес, рис, рожь, теф, тритикале, пшеница, дикий рис, просо пальчатое, фонио, просо итальянское, просо Кодо, японское просо, Иовлевы слезы, маис (кукуруза), жемчужное просо, просо обыкновенное и сорго. Токсины также были обнаружены в сене, льне, горохе, сое, рапсе и семенах других масличных культур, таких как подсолнечник, конопля и мак. Трихотеценовые токсины могут встречаться и в других видах пищи, например, в свекле, выращенной на поле, где в почву были запаханы остатки предыдущего урожая.
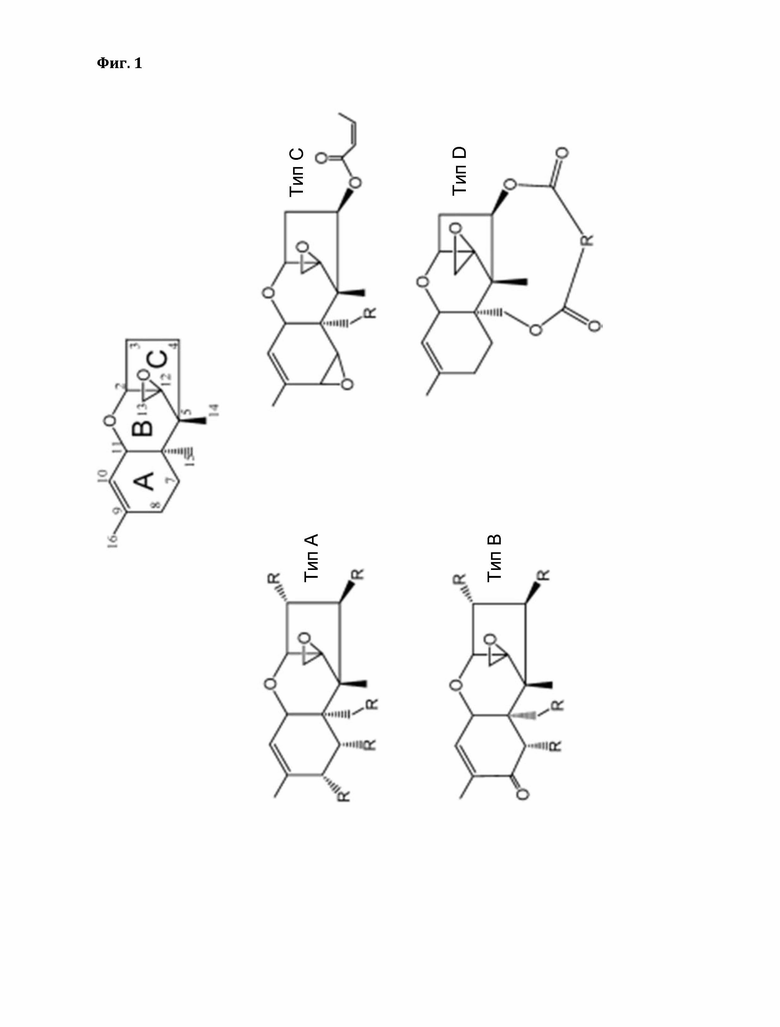
Трихотецены представляют собой сесквитерпеновые соединения, небольшие амфипатические молекулы, которые состоят из трихотеценового ядра с эпоксидными кольцами в положениях С-12 и С-13. Трихотецены классифицируют на четыре типа (A-D) на основе карбонильной группы в положении С-8, макролидных колец в положениях 4 и 15 и количества эпоксидных колец. Среди этих типов трихотецены типа А и трихотецены типа В имеют большое значение в сельском хозяйстве. Одним из наиболее подробно изученных трихотеценов является токсин Т-2 типа А-трихотецена. Важные трихотецены типа В включают дезоксиниваленол (ДОН, вомитоксин) и ниваленол (NIV).
Существуют также макроциклические трихотецены (трихотецены THnaD). Примеры макроциклических трихотеценов включают веррукарины, роридины и сатратоксин. Трихотеценовые микотоксины представляют собой нелетучие низкомолекулярные соединения, которые обычно относительно растворимы в воде, а также во многих органических растворителях, таких как ацетон, этилацетат, хлороформ, диметилсульфоксид и этанол.
В литературе описаны различные ферментативные способы, связанные с детоксикацией дезоксиниваленола и других трихотеценов. Большинство этих реакций детоксикации нацелены на побочные группы (например, восстановление и эпимеризация группы С3-ОН, ацетилирование С3-ОН, гликозилирование С3-ОН, гидроксилирование С16).
Было описано восстановление эпоксидадо олефина (С=С) анаэробными бактериями, но гипотетическая «эпоксид-редуктаза» пока неуловима, и ни один ген не был клонирован. Был проведен анализ состояния ферментативной (микробной) детоксикации (Karlovsky Р., 2011, Appl.Microbiol.Biotechnol., 91(3), 491-504; McCormick SP., 2013, J.Chem.Ecol., 39(7), 907-18). Эпоксид трихотеценов удивительно нереакционноспособен. Не известно ни одной эпоксид-гидролазы (раскрытие эпоксида путем добавления воды), способной инактивировать трихотецены. Известно, что в некоторых случаях происходит раскрытие эпоксида, катализируемое глутатион-8-трансферазой (GST) (de Vries EJ. & Janssen DB, 2003, Curr.Opin.Biotechnol., 14(4), 414-429).
Gardiner S.А. с соавторами (Molecular Plant-Microbe Interactions, 2010, 23 (7):962-976) сообщили, что глутатион может спонтанно (неферментативно) реагировать с ДОН, образуя так называемый аддукт Михаэля (фиг. 2, левая структура). Конъюгаты ДОН-глутатиона и продукты переработки были обнаружены in planta (Kluger В. et al., 2013, Analytical and Bioanalytical Chemistry, 405, 15, 5031-5036). Было показано, что добавление небольшой S-метильной группы (предполагаемый продукт деградации) к двойной связи значительно снижает токсичность (FruhmannP. et al., 2014, Org.Biomol.Chem., 12(28), 5144-5150), что указывает на то, что добавление громоздкого глутатиона должно также предотвратить взаимодействие с рибосомной мишенью. Тем не менее, аддукт Михаэля довольно неустойчив, особенно в щелочных условиях, и при удалении глутатиона из равновесия исходный токсин может быть восстановлен.
Совсем недавно было описано, что в очень медленной реакции (после нескольких недель в щелочных условиях) эпоксид также может быть раскрыт спонтанной реакцией с глутатионом (Stanic A. et al., 2016, J. Agric. Food Chem, 64(36), 6903-6910). Эта реакция (фиг. 2 А и В) считается необратимой (однако данных о стабильности продукта пока мало). Дальнейшая переработка эпоксид-глутатионового аддукта in vivo вполне вероятна, на данный момент (Kluger В. et al., 2015, PLOS One 10 (3):e0119656.) существуют доказательства только для производного, в котором 2 атома водорода отсутствуют в составе ДОН.
Аддукт к С13 (эпоксид) был обнаружен в (интенсивно хранившейся) пшенице, и было предположено (Uhlig S. et al., 2016, Toxins 8 (11):329), что это результат спонтанного (неферментативного) переноса глутатиона с двойной связи (аддукт Михаэля) на эпоксид.
В WO 2015/169847A1 сообщают о неферментативной детоксикации трихотецена, включающей раскрытие эпоксидного кольца трихотецена путем реакции с тиолом в щелочных условиях.
В WO 2000/20573A2 раскрыто применение нуклеиновых кислот, полученных путем перетасовки ДНК, для детоксикации микотоксинов, например, нуклеиновых кислот, кодирующих фермент с активностью глутатион-S-трансферазы.
Wahibah N.N. с соавторами сообщают о способе детоксикации реактивных форм кислорода и токсичных соединений с помощью экзогенной глутатион S-трансферазы неживотного происхождения (Plant Biotechnology 2018, 35(1), 71-79).
He J. с соавторами представили анализ биотрансформаций для детоксикации трихотецена в пищевых цепях (Trends in Food Science and Technology 2010, 21(2), 67-76).
He Z. с соавторами сообщают о потребности в увеличении производства зерновых в Китае (Journal of Cereal Science 2014, 59(3), 235-244).
Borisjuk N. с соавторами описывают генетическую модификацию для увеличения производства пшеницы (Biomed Research International 2019, 2019, 1-18).
Pan Y. с соавторами раскрывают анализ транскриптома и выявление дифференциально экспрессирующихся генов, таких как гены, кодирующие глутатион-S-трансферазу, мембранные белки и отдельные LRR-RK, которые связаны с устойчивостью к фузариуму (ВМС Genomics 2018, 19(1), 1-26).
Theodoulou F. с соавторами сообщают о совместной индукции глутатион-S-трансферазы и белка, связанного с множественной лекарственной устойчивостью, под действием ксенобиотиков, таких как гербициды (Pest Management Science 2003, 59 (2), 202-214).
Заражение трихотеценовыми микотоксинами является мировой проблемой, требующей ежегодной утилизации и уничтожения огромного количества сельскохозяйственной продукции. Поскольку превентивные меры пока не привели к успеху в предотвращении заражения микотоксинами, существует необходимость найти способ получения детоксифицированного и обеззараженного материала, а также способы обеззараживания зараженных веществ.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Таким образом, задачей настоящего изобретения является создание способа биотрансформации трихотеценов.
Указанную задачу решают с помощью объекта согласно настоящему изобретению.
Настоящее изобретение предоставляет способ биотрансформации трихотецена путем приведения материала, зараженного трихотеценами, в контакт с экзогенной глутатион-8-трансферазой (GST) неживотного происхождения, обладающей субстратной специфичностью к эпоксидному кольцу трихотецена, включающий этапы:
a) приведение материала в контакт с GST,
b) добавление глутатиона к указанному материалу, и
c) инкубирование смеси в водном растворе в диапазоне рН примерно от 6 до 9 в условиях, при которых глутатион реагирует с эпоксидным фрагментом, тем самым образуя эпоксидный аддукт.
Инкубацию продолжают в течение периода времени, достаточного для образования эпоксидного аддукта, чтобы снизить количество трихотеценового токсина в материале до приемлемого уровня, такого как уровень, приемлемый для потребления людьми и/или животными.
Согласно дополнительному варианту осуществления трихотецен выбран из группы трихотеценов типа А, трихотеценов типа В, трихотеценов типа С и трихотеценов типа D, в частности, трихотецен выбран из группы, состоящей из триходермола, триходермина, 4,15-диацетоксисцирпенола (ДАС), неозоланиола, токсина Т-2, токсина НТ-2, изотриходермола, калонектрина, 7,8-дигидроксикалонектрина, гарциана А, нивалонола (NIV), дезоксиниваленола (ДОН), 3- и 15-ацетилдезоксиниваленола, фузаренона-Х трихотецина, трихотеколона, трихотецинола А, кротоцина, сатратоксина Н, роридина А, бакарина, веррукарина А.
Более конкретно, трихотецен выбран из токсина Т-2, токсина НТ-2, неозоланиола, дезоксиниваленола (ДОН), ниваленола (NIV), 3- и 15-ацетилдезоксиниваленола, роридина А и веррукарина А.
В общем, способ, описанный в настоящем документе, можно применять для любого материала или вещества, которые могут быть заражены трихотеценовыми микотоксинами.
Согласно конкретному варианту осуществления настоящего изобретения способ, описанный в настоящем документе, применяют для получения обеззараженной кормовой добавки, кормового материала, пищевой добавки или пищевого материала.
Согласно дополнительному варианту осуществления способ, описанный в настоящем документе, используют для обеззараживания аэрозоля, жидкого или твердого материала или поверхности материала, в частности, текстильного материала, фильтрующего материала, противогазов, систем кондиционирования воздуха, для очистки поверхности материала, поверхности животных или человека, например, кожи и т.д.
В дополнительном варианте осуществления GST, используемая для описанного в настоящем документе способа, представляет собой рекомбинантную GST.
Согласно конкретному варианту осуществления GST имеет аминокислотную последовательность SEQ ID NO. 2 или SEQ ID NO. 4 или последовательность, имеющую по меньшей мере 80%, 85%, 90%, 95%, в частности, по меньшей мере 99% идентичности последовательности с любой из SEQ ID NO. 2 или SEQ ID NO. 4 и обладающую субстратной специфичностью к эпоксидному кольцу трихотецена.
Согласно дополнительному конкретному варианту осуществления GST содержит 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 аминокислотных модификаций SEQ ID NO. 2 или SEQ ID NO. 4.
В еще одном дополнительном варианте осуществления GST кодируется полинуклеотидной последовательностью, выбранной из группы, состоящей из a) SEQ ID NO. 1 или SEQ ID NO. 3 или последовательности, имеющей по меньшей мере 80%, 85%, 90%, 95%, в частности, по меньшей мере 99% идентичности последовательности с любой из SEQ ID NO. 1 или SEQ ID NO. 3 или b) выделенной молекулы нуклеиновой кислоты, комплементарной полинуклеотидной последовательности согласно а).
Согласно дополнительному конкретному варианту осуществления GST кодируется полинуклеотидной последовательностью, выбранной из группы, состоящей из a) SEQ ID NO. 1, или SEQ ID NO. 3, имеющих 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 20 и до 50 нуклеотидных модификаций или б) выделенной молекулы нуклеиновой кислоты, комплементарной полинуклеотидной последовательности согласно а).
Согласно варианту осуществления GST экспрессируется в клетке-хозяине.
Настоящее изобретение также обеспечивает кормовую добавку или кормовой материал, содержащий экзогенную GST неживотного происхождения, обладающую субстратной специфичностью к эпоксидному кольцу трихотецена и, необязательно, глутатион.
Согласно конкретному варианту осуществления кормовая добавка или кормовой материал содержит часть трансгенного растения, ткань трансгенного растения, клетку трансгенного растения, семя или его потомство, в частности, лист, стебель, корень, семядолю или гипокотиль, каждый из которых содержит экзогенную GST, обладающую субстратной специфичностью к эпоксидному кольцу трихотецена.
Согласно конкретному варианту осуществления способ, описанный в настоящем документе, применяют для ферментативной деградации трихотецена в кормах для животных или в пищеварительном тракте животных.
Согласно альтернативному варианту осуществления описанный способ применяют для производства кормовой или пищевой добавки.
Настоящее изобретение также обеспечивает кормовой или пищевой материал, содержащий обеззараженный растительный материал, полученный описанным в настоящем документе способом.
Кроме того, в настоящем документе предложен хозяин, сверхэкспрессирующий эндогенную GST или трансформированный вектором, экспрессирующим экзогенную GST, при этом указанная GST обладает субстратной специфичностью к эпоксидному кольцу трихотецена. В частности, хозяин является прокариотом или эукариотом, в частности, это растительная клетка, животная клетка, грибковая клетка или бактериальная клетка.
Согласно дополнительному варианту осуществления клетка-хозяин содержит GST, где GST кодируется
a) полинуклеотидной последовательностью, выбранной из группы, состоящей из SEQ ID NO. 1, SEQ ID NO. 3 или последовательности, имеющей по меньшей мере 80%, 85%, 90%, 95%, в частности, по меньшей мере 99% идентичности последовательности с любой из SEQ ID NO. 1, SEQ ID NO. 3, или
b) выделенной молекулой нуклеиновой кислоты, комплементарной а).
Согласно дополнительному варианту осуществления клетка-хозяин включает экспрессионную кассету. В частности, экспрессионная кассета содержит полинуклеотидную последовательность, кодирующую экзогенную GST, функционально связанную с регуляторной последовательностью, такой как, но не ограниченной указанными, CaMV 35S, NOS, OCS, AdhI, AdhII и Ubi-1, malE, функциональной в указанном хозяине.
В дополнительном варианте осуществления экспрессионная кассета дополнительно содержит считываемый маркерный полинуклеотид, такой как, но не ограничиваясь указанными, GUS, GFP, CAT, LUC, сиалидаза, функционально связанный с регуляторной последовательностью, функционирующей в растении. Кроме того, полинуклеотид селективного маркера, такой как, но не ограничиваясь указанными, nptII, hptII, pat и bar, функционально связан с регуляторной последовательностью, функционирующей в указанном хозяине.
В дополнительном варианте осуществления предложен хозяин, экспрессирующий описанную в настоящем документе GST, обладающую субстратной специфичностью к трихотецену, выбранному из группы трихотеценов типа А, трихотеценов типа В, трихотеценов типа С, трихотеценов типа D.
В настоящем изобретении также предложен хозяин, экспрессирующий GST и обладающий субстратной специфичностью к трихотецену, выбранному из группы трихотеценов типа А, трихотеценов типа В, трихотеценов типа С и трихотеценов типа D, в частности трихотецен выбран из группы, состоящей из триходермола, триходермина, 4,15-диацетоксисциртоксина (ДАС), неозоланиола, токсина Т-2, токсина НТ-2, изотриходермола, калонектрина, 7,8-дигидроксикалонектрина, гарциана А, нивалонола (NIV), дезоксиниваленола (ДОН), 3- и 15-ацетилдезоксиниваленола, фузаренона-Х трихотецина, трихотеколона, трихотецина А, кротоцина, сатратоксина Н, роридина А, бакарина и веррукарина А.
Более конкретно, трихотецен выбран из токсина Т-2, токсина НТ-2, неозоланиола, дезоксиниваленола (ДОН), ниваленола (NIV), 3- и 15-ацетилдезоксиниваленола, роридина А и веррукарина А.
Согласно конкретному варианту осуществления экзогенная GST хозяина содержит аминокислотную последовательность, выбранную из группы SEQ ID NO. 2, SEQ ID NO. 4 или последовательности, имеющей по меньшей мере 80%, 85%, 90%, 95%, в частности, по меньшей мере 99% идентичности последовательности с любой из SEQ ID NO. 2, SEQ ID NO. 4 и обладающую субстратной специфичностью к эпоксидному кольцу трихотецена.
Также предложено трансгенное растение с повышенной устойчивостью к трихотецену, содержащее экзогенную GST, обладающую субстратной специфичностью к эпоксидному кольцу трихотецена.
Также предложена часть трансгенного растения, ткань трансгенного растения, клетка трансгенного растения, семя или его потомство, в частности, лист, стебель, корень, семядолю и гипокотиль, содержащие экзогенную GST, обладающую субстратной специфичностью к эпоксидному кольцу трихотецена.
Согласно одному варианту осуществления в настоящем документе также предложено трансгенное животное с повышенной устойчивостью к трихотецену, содержащее экзогенную GST, обладающую субстратной специфичностью к эпоксидному кольцу трихотецена.
В настоящем изобретении также предложен способ получения трансгенного растения или животного, включающий трансформацию растения или животного молекулой нуклеиновой кислоты, кодирующей GST, и экспрессию молекулы нуклеиновой кислоты в указанном растении, при этом молекула нуклеиновой кислоты содержит полинуклеотидную последовательность, выбранную из группы, состоящей из полинуклеотидной последовательности, содержащей SEQ ID NO. 1 или SEQ ID NO.3 или последовательности, имеющей по меньшей мере 80%, 85%, 90%, 95%, в частности, по меньшей мере 99% идентичности последовательности с любой из SEQ ID NO. 1 или SEQ ID NO. 3.
Согласно дополнительному варианту осуществления растение может быть трансформировано способом, выбранным из группы, состоящей из Agrobacterium-опосредованной трансформации, биобаллистической бомбардировки, вакуумной инфильтрации, трансформации in planta и химического метода.
В настоящем изобретении также предложен способ экспрессии экзогенной GST, обладающей субстратной специфичностью к эпоксидному кольцу трихотецена в организме хозяина, включающий:
a) трансформацию клетки-хозяина вектором, содержащим последовательность гена, кодирующего GST, и
b) выращивание трансформированной клетки в условиях, подходящих для экспрессии гена, кодирующего экзогенную GST.
Трансгенное растение, описанное в настоящем документе, может быть однодольным или двудольным растением.
В частности, однодольное растение представляет собой траву, в частности, выбранную из группы, состоящей из риса, маиса, пшеницы, ячменя, сорго, ржи и овса.
В частности, двудольное растение выбрано из группы, состоящей из табака, томата, гороха, сои, капусты, нута, арабидопсиса и моркови.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
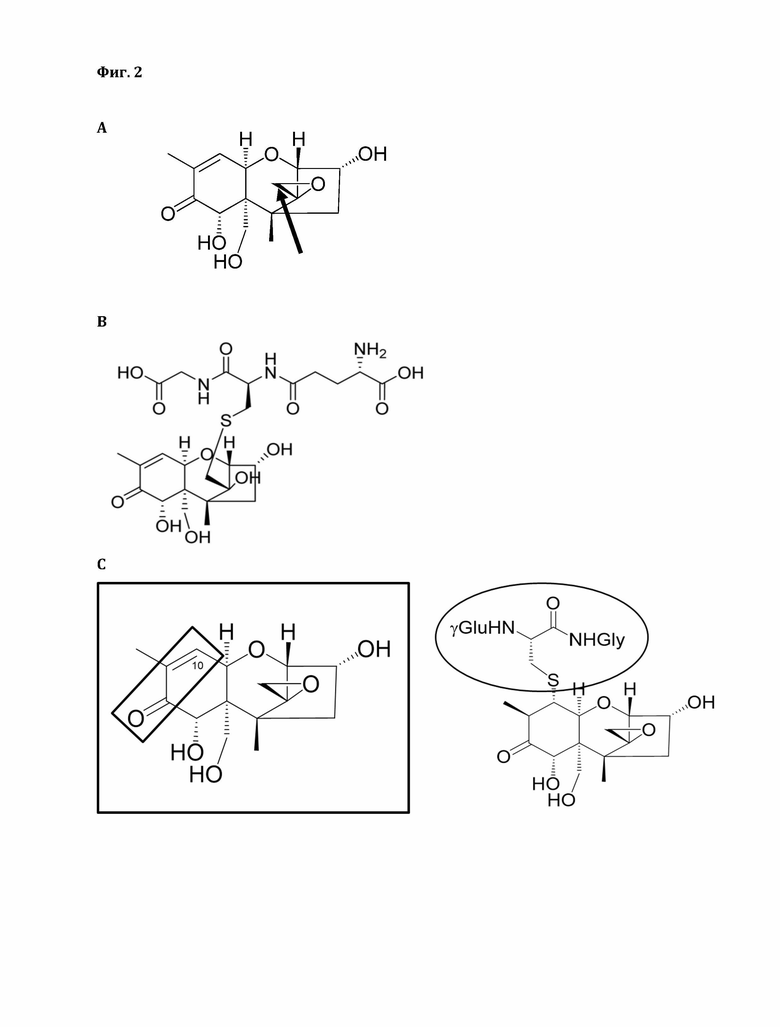
Фигура 1: Система нумерации углеродов трихотецена и структуры различных типов трихотеценов. Структуры трихотецена и трихотеценов типов А, В, С и D.
Фигура 2: А) Дезоксиниваленол (ДОН), раскрытие эпоксида. Стрелка указывает на атом углерода, который атакуется SH-группой глутатиона. В) Аддукт ДОН-глутатион. Показана структура образующегося эпоксидного аддукта ДОН-глутатион (вторично С15-ОН может также реагировать с С8-кето с образованием гемикеталя). С) Структурные требования для образования аддукта Михаэля, в результате чего образуется аддукт ДОН-глутатион (ДОН-GSH). Образование аддукта Михаэля возможно только при наличии сопряженной двойной связи и кето-группы (трихотецены типа В).
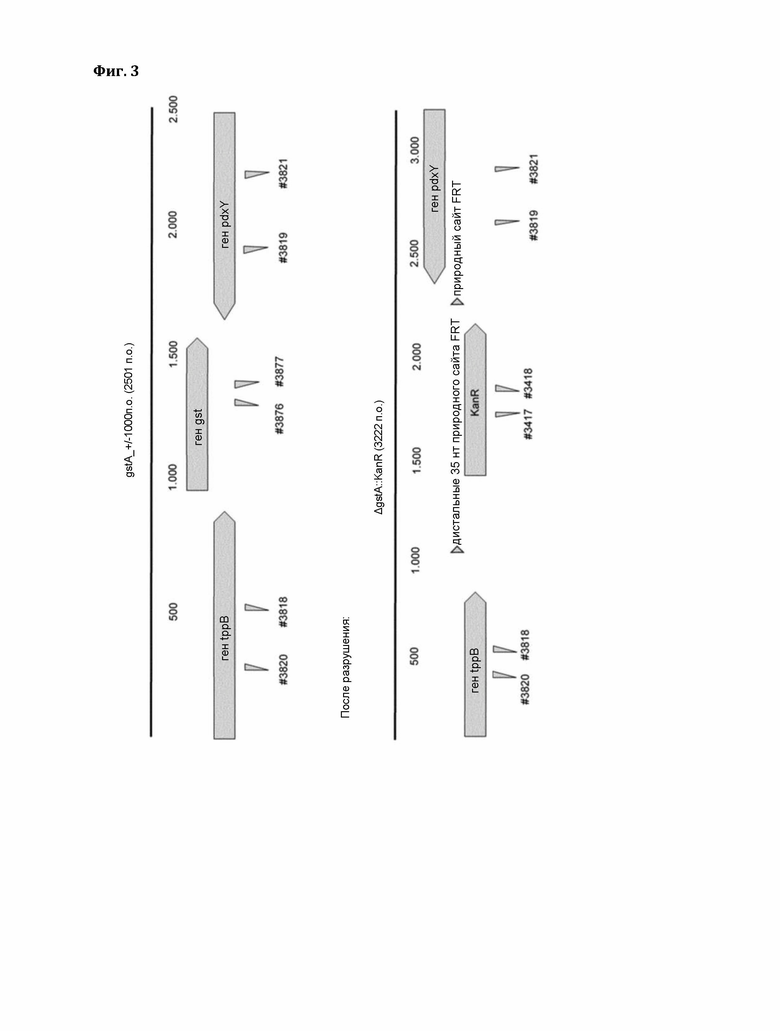
Фигура 3: Конструирование штамма-хозяина в системе «Т-7 Express». Схематическое сравнение дикого типа (А) и штамма после разрушения (В).
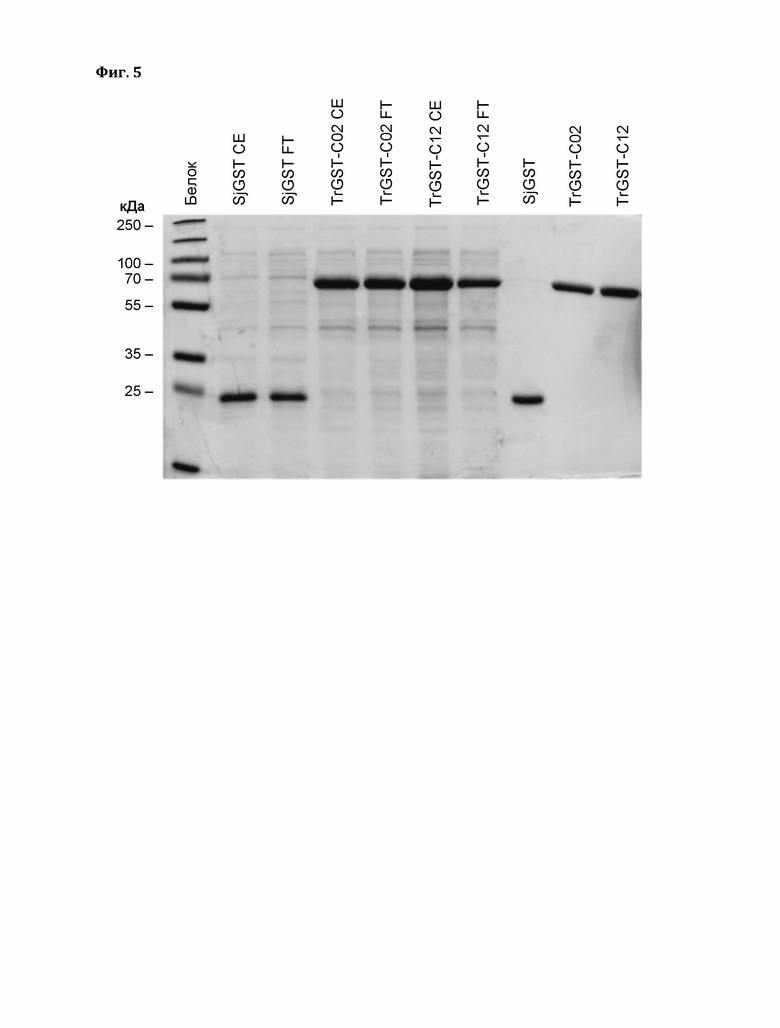
Фигура 4: SDS-ПААГ слитых белков 6xHis-malE-(TEV)-GST. Результаты SDS-ПААГ в 12% полиакриламиде, использованного для оценки экспрессии слитых белков 6xHis-malE-(TEV)-GST. От каждого кандидата на гели загружали два образца: общий белок слева и клеточный экстракт (очищенный лизат) справа.
Фигура 5: Очистка белка в SDS-ПААГ. Применяемые концентрации: СЕ (клеточный экстракт) и FT (проточная фракция): 0,75 мг/мл, Белок: 0,23 мг/мл
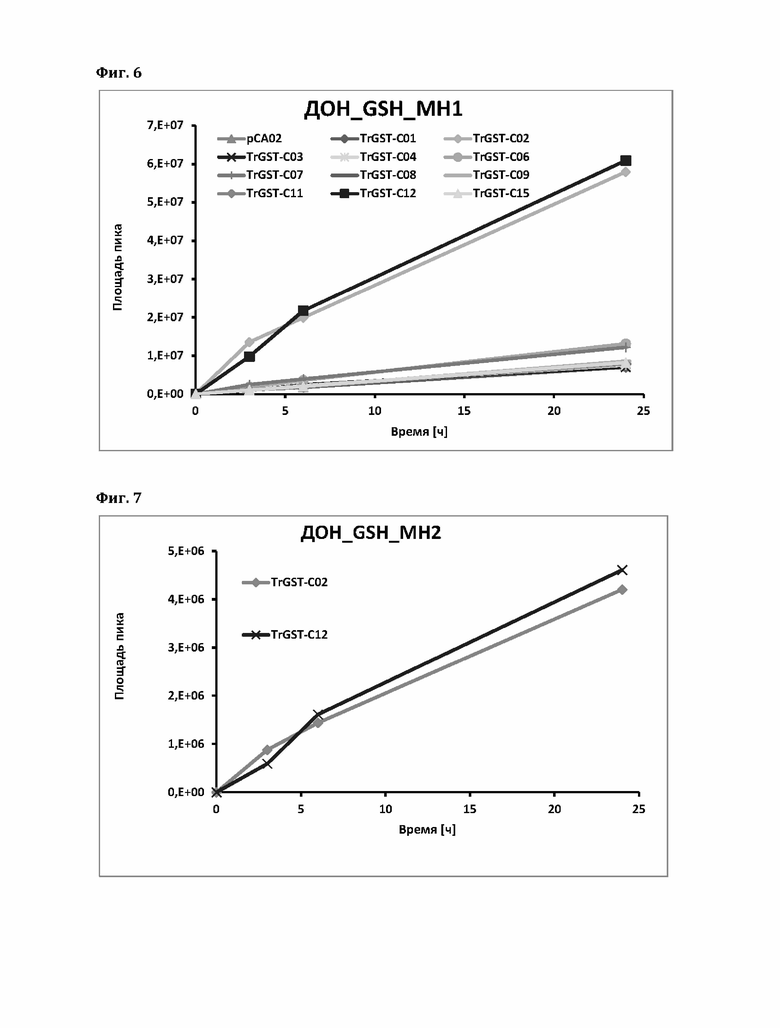
Фигура 6: Ферментативное образование аддукта Михаэля ДОН (МН1) с применением различных GST.
Фигура 7: Ферментативное образование эпоксидного аддукта ДОН (МН2), катализируемое GST С02 и С12. Применяли те же GST, что и для анализа ДОН-GSH_MH1, показанного на фиг. 6, но GST кроме GST С02 и С12 не показали обнаруживаемого образования эпоксидных аддуктов.
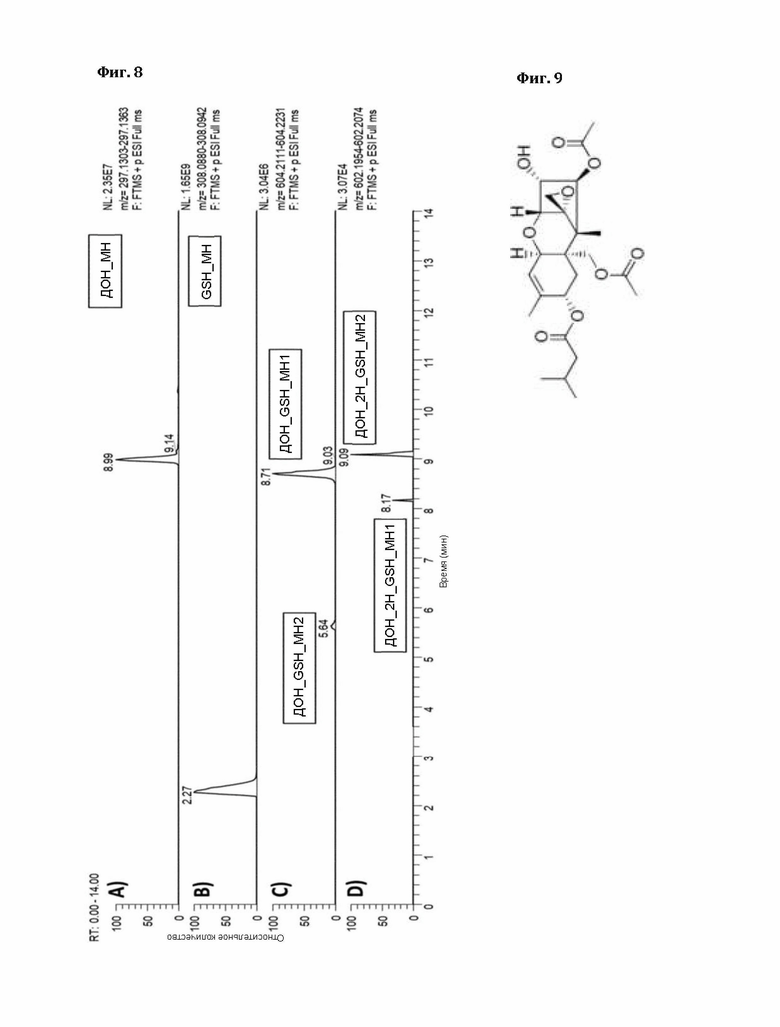
Фигура 8: Экстракционные ионные хроматограммы для ДОН (реакция с пшеничным GST С02).
Фигура 9: Структура токсина Т-2.
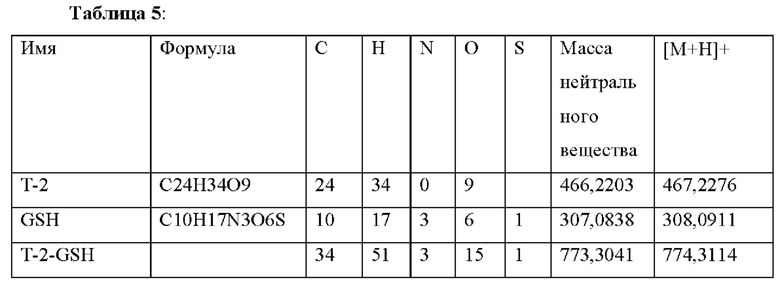
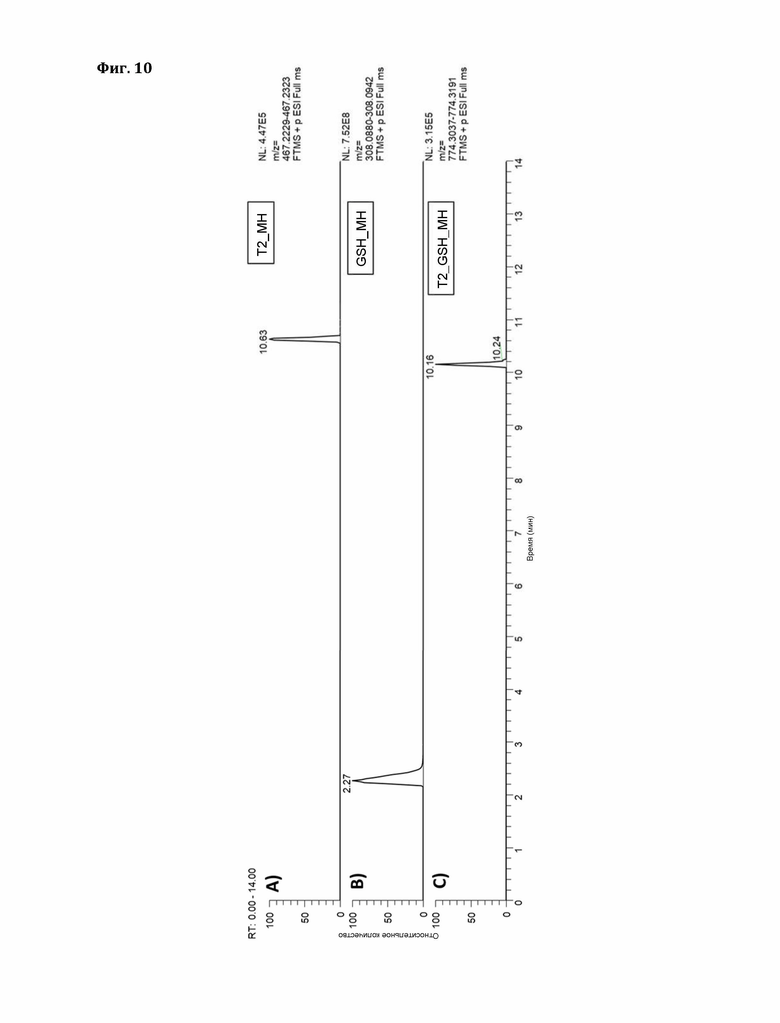
Фигура 10: Экстракционные ионные хроматограммы ионов [М+Н]+ А) Т2, В) GSH, С) Т2_GSH (окно масс:±5ppm).
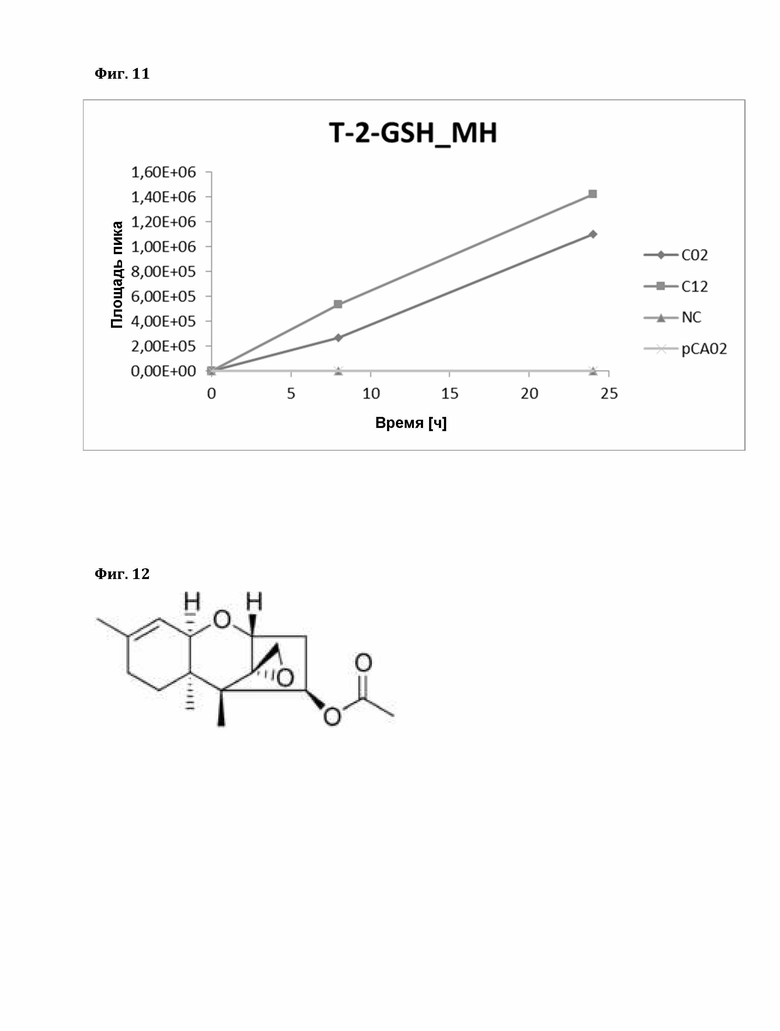
Фигура 11: Фермент-зависимое образование аддукта глутатиона (GSH) с Т-2 токсином. T-2-GSH_МН; рСА02 - пустой вектор, NC - отрицательный контроль (без белка), TaGST - C02/C12.
Фигура 12: Структура триходермина.
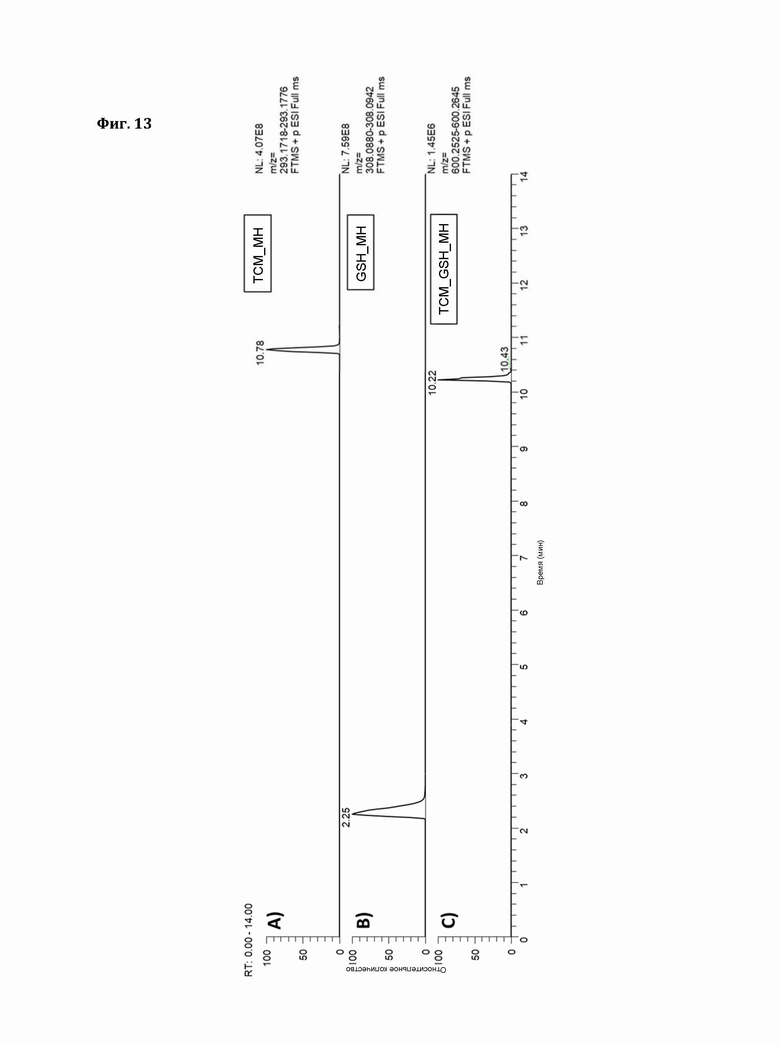
Фигура 13: Экстракционные ионные хроматограммы ионов [М+Н]+ А) ТСМ, В) GSH, С) ТСМ_GSH (окно масс:±5ppm).
Фигура 14: Фермент-зависимое образование аддукта глутатиона (GSH) с триходермином. Триходермин-GSH_MH; рСА02 - пустой вектор, NC - отрицательный контроль (без белка), TaGST - С02/С12.
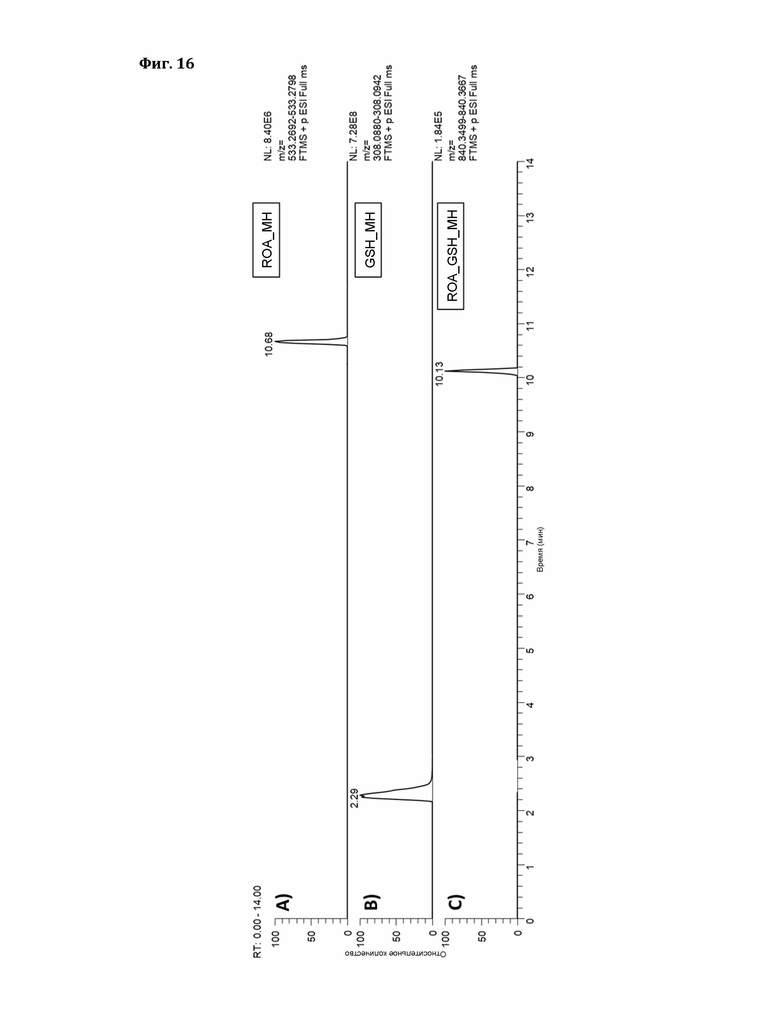
Фигура 15: Структура роридина A (ROA).
Фигура 16: Экстракционные ионные хроматограммы ионов [М+Н]+ A) ROA, В) GSH, С) ROA_GSH (окно масс:±5ppm).
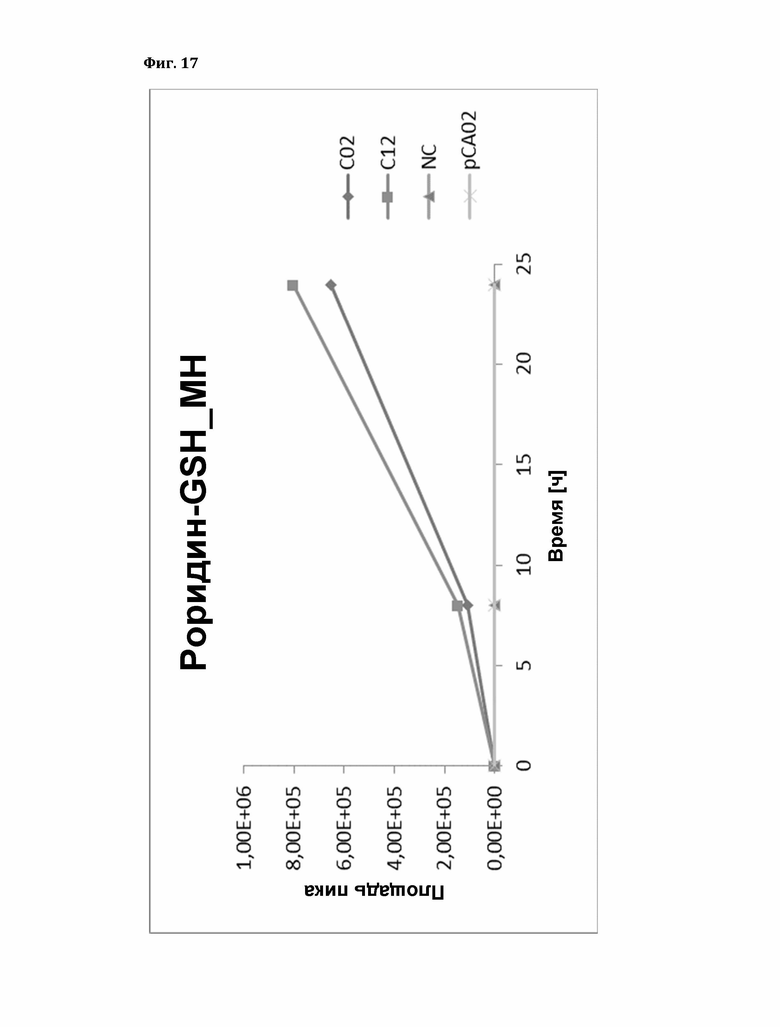
Фигура 17: Фермент-зависимое образование аддукта глутатиона (GSH) с роридином А. Роридин-GSH_МН; рСА02 - пустой вектор, NC - отрицательный контроль (без белка), TaGST - С02/С12.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Если не указано или не определено иное, все термины, используемые в настоящем документе, имеют свое обычное значение в данной области техники, которое будет понятно специалисту. Например, приведены ссылки на стандартные справочники, такие как Sambrook et al., "Molecular Cloning: A Laboratory Manual" (2-е изд.), тома 1-3, лаборатория издательства Cold Spring Harbor (1989); Lewin, "Genes IV", издательство Oxford University, New York, (1990), и Janeway et al., "Immunobiology" (5-е изд., или более поздние издания, Garland Science, New York, 2001).
Объект согласно формуле изобретения в частности относится к искусственным продуктам или способам, использующим или производящим такие искусственные продукты, которые могут быть вариантами нативных продуктов (продуктов дикого типа). Хотя может быть определенная степень идентичности последовательности нативной структуре, хорошо понятно, что материалы, способы и применения настоящего изобретения, например, конкретно относящиеся к выделенным последовательностям нуклеиновых кислот, последовательностям аминокислот, слитым конструкциям, экспрессионным конструкциям, трансформированным клеткам-хозяевам и модифицированным белкам, включая ферменты, являются «рукотворными» или синтетическими и поэтому не рассматриваются как результат «законов природы».
Термины «состоять», «содержать», «иметь» и «включать», используемые в настоящем документе, могут использоваться как синонимы и должны пониматься как открытое определение, допускающее дополнительные члены или части, или элементы. «Состоящий» рассматривается как наиболее близкое определение без дополнительных элементов признака составного определения. Таким образом, «включающий» является более широким и содержит определение «состоящий».
Термин «примерно», используемый в настоящем документе, относится к одному и тому же значению или значению, отличающемуся на +/-5% от данного значения.
В настоящем документе представлен новый способ биотрансфомации трихотеценов с применением неживотных GST, обладающих субстратной специфичностью к эпоксидному кольцу трихотеценов.
Глутатион S-трансферазы (GST) представляют собой семейство эукариотических и прокариотических изоферментов метаболизма фазы II, известных своей способностью катализировать конъюгацию восстановленной формы глутатиона (GSH) с субстратами ксенобиотиков с целью детоксикации. Семейство GST состоит из трех суперсемейств: цитозольных, митохондриальных, и микросомных белков. Члены суперсемейства GST чрезвычайно разнообразны по аминокислотной последовательности, и большая часть последовательностей, депонированных в открытых базах данных, имеет неизвестную функцию.
«GST неживотного происхождения» относится к GST любого происхождения, кроме животного происхождения, такого как крыса или любого другого вида животных.
Трихотецены типа А включают соединения, которые имеют гидроксильную группу у С-8 (например, неозоланиол), сложноэфирную группу у С-8 (например, токсин Т-2) или без кислородного замещения у С-8 (например, триходермин, 4, 15-диацетоксисцирпенол и гарциан А).
Трихотецены типа А: триходермол, триходермин, 4,15-диацетоксицирпенол (ДАС), неосоланиол, токсин Т-2, токсин НТ-2, изотриходермол, калонектрин, 7,8-дигидроксикалонектрин, гарциан А.
Трихотецены типа В имеют кето (карбонильную) группу при С-8 (например, ниваленол, дезоксиниваленол и трихотецин).
У Fusarium трихотецены типа В обычно имеют гидроксильную группу С-7, но эта структурная особенность не присутствует у других родов.
Трихотецены типа В: нивалонол (NIV), дезоксиниваленол (ДОН), 3- и 15-ацетилдезоксиниваленол, фузаренон-Х, трихотецин, трихотецинол А.
Трихотецены типа С имеют эпоксид С-7/С-8 (например, кротоцин). Трихотецен типа С: кротоцин.
Трихотецены типа D имеют дополнительное кольцо, связывающее положения С-4 и С-15 (например, роридин А, веррукарин А, сатратоксин Н).
Трихотецены типа D: сатратоксин Н, роридин А, бакарин, веррукарин А.
Все трихотецены Fusarium (включая тип А и тип В) имеют кислородную группу (т.е. гидроксильную или ацетильную группу) при С-3. Трихотецены, производимые Trichoderma, Trichothecium, Myrothecium или Stachybotrys (включая типы А, В, С и D), не имеют кислородной группы в положении С-3.
В описанном в настоящем документе способе биотрансформации зараженный материал подвергают контакту с GST и глутатионом и инкубируют в течение периода времени, достаточного для протекания реакции детоксикации, чтобы снизить количество трихотеценового токсина путем образования эпоксидного аддукта в образце до приемлемого уровня, например, уровня, приемлемого для потребления человеком и/или животными. В частности, трихотецен полностью превращается в эпоксидный аддукт с помощью описанного в настоящем изобретении способа.
Например, реакция биотрансформации может протекать от одного часа до 24 часов, в частности, материал инкубируют с GST в течение 1, 2, 3, 4, 5, 10, 15, 20, 24 часов. В частности, реакция биотрансформации прекращается в течение 24 часов.
Реакция может протекать при температуре окружающей среды, например, при комнатной температуре. Кроме того, реакцию можно проводить при повышенных температурах, таких как температуры, обычно используемые при производстве продуктов питания и кормов. Они могут составлять, например, от примерно 30°С до примерно 40°С.
Реакцию биотрансформации можно, например, проводить при рН от примерно 6 до 9, например, в диапазоне рН от примерно 6,5 до 8,5.
Способ биотрансформации, раскрытый в настоящем документе, можно применять для детоксикации любого вида материала, зараженного трихотеценом. Материал может содержать один вид трихотеценового токсина типа А, В, С или типа D, или два или более видов токсинов типа А, В, С и/или типа D. Как правило, материал представляет собой продукт, предназначенный для использования в качестве пищи или корма, кормовой добавки или пищевой добавки, как таковой или после обработки, например, сельскохозяйственный продукт.В дополнительном примере образец представляет собой пищевой продукт или кормовой продукт.Примерный материал для обработки включает, но не ограничивается указанными, сено или солому, зерно или семена, муку и другие продукты перемола, корм для скота или рыбы. Материал может быть продуктом, полученным из зерна или содержащим зерно, таким как зерно или семена, предназначенные для производства продуктов питания или кормов. Типичные зерна включают, но не ограничиваются указанными, овес, ячмень, маис, рожь, рис, сорго, пшеницу, теф, тритикале, дикий рис, пальчатое просо, фонио, просо итальянское, просо Кодо, японское просо, Иовлевы слезы, жемчужное просо и просо обыкновенное. Другие примеры материалов, которые могут быть заражены трихотеценами, включают лен, горох, сою, рапс и семена других масличных культур, таких как подсолнечник, конопля и мак. Трихотеценовые токсины могут содержаться и в других видах пищи, например, в свекле. Продукты, полученные из зерна, включают, но не ограничиваются указанными, сырое зерно, муку и крупы. Кроме того, трава и корма для животных подходят для детоксикации в соответствии с настоящим документом. Способ также можно применять для обеззараживания непищевых материалов или их поверхности, таких как текстильный материал, например, одежда, фильтрующий материал, противогазы, системы кондиционирования воздуха и т.д.
Описанный в настоящем документе способ также можно применять для очистки материалов и поверхностей материалов от трихотеценов, а также для очистки или обеззараживания поверхностей животных и человека, таких как кожа, волосы, мех и т.д.
В частности, GST представляет собой рекомбинантную GST.
Термин «рекомбинантный», используемый в настоящем документе, относится к молекуле или конструкции, которая не встречается естественным образом в клетке-хозяине. В некоторых вариантах осуществления рекомбинантные молекулы нуклеиновых кислот содержат две или более последовательностей, встречающихся в природе, которые связаны друг с другом таким образом, который не встречается в природе. Рекомбинантный белок относится к белку, который кодируется и/или экспрессируется рекомбинантной нуклеиновой кислотой. В некоторых вариантах осуществления «рекомбинантные клетки» экспрессируют гены, которые не встречаются в идентичной форме в нативной (т.е. нерекомбинантной) форме клетки и/или экспрессируют нативные гены, которые в противном случае аномально сверхэкспрессированы, недоэкспрессированы и/или вообще не экспрессированы вследствие преднамеренного вмешательства человека. Рекомбинантные клетки содержат по меньшей мере один рекомбинантный полинуклеотид или полипептид. «Рекомбинация», «рекомбинирование» и создание «рекомбинированной» нуклеиновой кислоты в целом охватывают сборку по меньшей мере двух фрагментов нуклеиновой кислоты. В некоторых вариантах осуществления рекомбинантные белки и рекомбинантные нуклеиновые кислоты остаются функциональными, т.е. сохраняют свою активность или проявляют повышенную активность в клетке-хозяине.
В частности, рекомбинантная GST содержит одну из следующих нуклеотидных или аминокислотных последовательностей:




GST может иметь аминокислотную последовательность SEQ ID NO. 2 или SEQ ID NO. 4 или последовательность, имеющую по меньшей мере 80%, 85%, 90%, 95%, в частности, по меньшей мере 99% идентичности последовательности с SEQ ID NO. 2 или SEQ ID NO. 4 и обладающую субстратной специфичностью к эпоксидному кольцу трихотецена. В качестве альтернативы аминокислотные последовательности SEQ ID NO. 2 или SEQ ID NO. 4 могут включать 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 модификаций, при условии, что GST, состоящая из SEQ ID NO 2 или SEQ ID NO 4, сохраняет свою субстратную специфичность к эпоксидному кольцу трихотецена.
GST может быть закодирована последовательностью нуклеиновой кислоты SEQ ID NO. 1 или SEQ ID NO. 3 или последовательностью, имеющей по меньшей мере 80%, 85%, 90%, 95%, в частности, по меньшей мере 99% идентичности последовательности с SEQ ID NO. 1 или SEQ ID NO. 3 и обладающей субстратной специфичностью к эпоксидному кольцу трихотецена. В качестве альтернативы последовательности нуклеиновых кислот SEQ ID NO. 1 или SEQ ID NO. 3 могут включать 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20 или более модификаций, при условии, что GST, кодируемая SEQ ID NO. 1 или SEQ ID NO. 3, сохраняет свою субстратную специфичность к эпоксидному кольцу трихотецена.
Аминокислотные или нуклеотидные модификации в настоящем документе относятся к любым модификациям, известным в данной области техники, таким как точечные мутации, делеции или вставки аминокислотных остатков или нуклеотидов.
«Процент (%) идентичности» в отношении аминокислотной последовательности, гомологов и ортологов, описанных в настоящем документе, определяется как процент аминокислотных остатков в кандидатной последовательности, которые идентичны аминокислотным остаткам в конкретной полипептидной последовательности, после выравнивания последовательности и введения пробелов, если необходимо, для достижения максимального процента идентичности последовательностей, и без учета любых консервативных замен как части идентичности последовательности. Специалисты в данной области техники могут определить соответствующие параметры для измерения выравнивания, включая любые алгоритмы, необходимые для достижения максимального выравнивания по всей длине сравниваемых последовательностей.
Для целей, описанных в настоящем документе, идентичность последовательностей между двумя аминокислотными последовательностями определяется с использованием программы NCBI BLAST версии 2.2.29 (06.01.2014) со следующими типичными параметрами blastp: программа: blastp, размер слова: 6, ожидаемое значение: 10, размер списка совпадений: 100, цены разрывов: 11,1, матрица: BLOSUM62, фильтрующая строка: F, генетический код: 1, размер окна: 40, порог: 21, статистика по составу: 2.
«Процент (%) идентичности» в отношении нуклеотидной последовательности, например, молекулы нуклеиновой кислоты или ее части, в частности, кодирующей последовательности ДНК, определяется как процент нуклеотидов в последовательности ДНК-кандидата, которая идентична нуклеотидам в последовательности ДНК после выравнивания последовательностей и введения пробелов, если необходимо, для достижения максимального процента идентичности последовательностей и без учета каких-либо консервативных замен как части идентичности последовательности. Выравнивание для целей определения процента идентичности нуклеотидных последовательностей может быть достигнуто различными способами, доступными специалистам в данной области техники, например, с помощью общедоступного компьютерного программного обеспечения. Специалисты в данной области техники могут определить соответствующие параметры для измерения выравнивания, включая любые алгоритмы, необходимые для достижения максимального выравнивания по всей длине сравниваемых последовательностей.
Оптимальное выравнивание может быть определено с помощью любого подходящего алгоритма для выравнивания последовательностей, неограничивающие примеры которых включают алгоритм Смита-Ватермана, алгоритм Нидлмана-Вунша, алгоритмы, основанные на преобразовании Барроуза-Уиллера (например, Burrows Wheeler Aligner), ClustalW, Clustal X, BLAT, Novoalign (Novocraft Technologies; доступен на novocraft.com), ELAND (Illumina, San Diego, CA), SOAP (доступен Hasoap.genomies.org.cn) и Maq (доступен на maq.sourceforge.net).
Для экспрессии GST, описанных в настоящем документе, можно использовать любую клетку-хозяина, подходящую для экспрессии рекомбинантной экзогенной функциональной GST, обладающей субстратной специфичностью к эпоксидному кольцу трихотецена. Клетки-хозяева могут быть прокариотическими или эукариотическими клетками или клеточными культурами. Такие клетки-хозяева могут быть, но не ограничиваются указанными, клетками грибов, такими как Saccharomyces sp., бактериальными клетками, такими как Е. coli, растительными клетками или клетками животных. Благодаря рекомбинантной экспрессии GST, описанной в настоящем документе, трансформированные таким образом клетки-хозяева могут обладать субстратной специфичностью в отношении трихотеценов, в частности, трихотеценов типа А, трихотеценов типа В, трихотеценов типа С, и трихотеценов типа D, в частности дезоксиниваленола (ДОН), 3- и 15-ацетилдезоксиниваленола, ниваленола (NIV), токсина Т2, токсина НТ-2, неозоланиола, диацетоксицирпенола, трихотецина, роридина А и веррукарина А. Благодаря субстратной специфичности клетки-хозяева могут лучше переносить заражение трихотеценами.
В частности, предложены трансгенные растения или их части, такие как ткани, семена, листья, стебли, корни, семядоли или гипокотили, экспрессирующие экзогенную GST, обладающую субстратной специфичностью к эпоксидному кольцу трихотецена. Такие растения могут быть однодольными или двудольными растениями, такими как, но не ограничиваясь указанными, рис, маис, пшеница, ячмень и сорго, рожь, овес или табак, томат, горох, соя, капуста, нут, арабидопсис и морковь.
В частности, в настоящем документе также представлены трансгенные животные, не являющиеся человеком, с повышенной устойчивостью к трихотецену, содержащие экзогенную GST, обладающую субстратной специфичностью к эпоксидному кольцу трихотецена. В частности, трансгенные виды, такие как свиньи, рыбы или насекомые, могут таким образом лучше переносить зараженный токсинами корм.
Термин «экзогенная глутатион-S-трансфераза неживотного происхождения», используемый в настоящем документе, относится к GST неживотного происхождения, которая не присутствует в естественном виде в материале, зараженном трихотеценом, или не экспрессируется в материале изначально. Термин «экзогенный» может использоваться взаимозаменяемо с термином «чужеродный». GST неживотного происхождения присутствует в материале в результате добавления указанного белка GST или экспрессии кодирующей последовательности GST в трансгенной последовательности ДНК клетки-хозяина или организма, тем самым кодируя экзогенную GST. Экзогенная GST представляет собой любую GST неживотного происхождения, которая добавляется к материалу и/или экспрессируется в организме или клетке-хозяине, возникший вне этого материала или организма, в отличие от эндогенного фактора.
Термин «экспрессия» понимается следующим образом. Молекулы нуклеиновой кислоты, содержащие желаемую кодирующую последовательность продукта экспрессии, такого как GST, описанная в настоящем документе, как описано в настоящем документе, могут быть использованы для целей экспрессии. Такие молекулы нуклеиновой кислоты конкретно называются «выделенная молекула нуклеиновой кислоты» или «выделенная нуклеотидная последовательность». Хозяева, трансформированные или транс фицированные этими последовательностями, способны продуцировать закодированные белки. Для осуществления трансформации система экспрессии может быть включена в вектор; однако соответствующая ДНК также может быть интегрирована в хромосому хозяина. В частности, этот термин относится к клетке-хозяину и совместимому вектору в подходящих условиях, например, для экспрессии белка, кодируемого чужеродной ДНК, переносимой вектором и введенной в клетку-хозяина.
Кодирующая ДНК представляет собой последовательность ДНК, которая кодирует определенную аминокислотную последовательность для конкретного полипептида или белка. Промоторная ДНК представляет собой последовательность ДНК, которая инициирует, регулирует или иным образом опосредует или контролирует экспрессию кодирующей ДНК. ДНК промотора и кодирующая ДНК могут быть из одного и того же гена или из разных генов, и могут быть из одного и того же или разных организмов. Рекомбинантные клонирующие векторы часто включают одну или более систем репликации для клонирования или экспрессии, один или более маркеров для селекции в хозяине, например, устойчивость к антибиотикам, один или более сигналов ядерной локализации (NLS) и одну или более экспрессионных кассет.
«Векторы экспрессии» или «векторы», используемые в настоящем документе, определены как последовательности ДНК, необходимые для транскрипции клонированных рекомбинантных нуклеотидных последовательностей, т.е. рекомбинантных генов, и трансляции их мРНК в подходящем организме-хозяине. Для достижения экспрессии последовательность, кодирующая желаемый продукт экспрессии, такой как GST, описанная в настоящем документе, обычно клонируется в вектор экспрессии, который содержит промотор для направления транскрипции. Подходящие бактериальные и эукариотические промоторы хорошо известны в данной области техники. Промотор, используемый для направления экспрессии нуклеиновой кислоты, зависит от конкретного применения. Например, сильный конститутивный промотор обычно используется для экспрессии и очистки рекомбинантных белков. Напротив, когда продукт экспрессии должен вводиться in vivo для регуляции генов, можно использовать конститутивный или индуцибельный промотор, в зависимости от конкретного применения продукта экспрессии. Кроме того, предпочтительным промотором для введения может быть слабый промотор. Векторы экспрессии включают экспрессионную кассету и дополнительно обычно включают источник для автономной репликации в клетках-хозяев ах или сайт интеграции в геном, один или более селектируемых маркеров (например, ген синтеза аминокислот или ген, придающий устойчивость к антибиотикам, таким как зеоцин, канамицин, G418 или гигромицин; или nptI, hptII, pat и bar), несколько сайтов расщепления ферментами рестрикции, подходящую последовательность промотора и терминатор транскрипции, компоненты которых функционально связаны друг с другом. Векторы также могут содержать регуляторные элементы, такие как, но не ограничиваясь указанными, CaMV, 35S, NOS, AdhI, Adhll, Ubi-1 и mal E.
«Экспрессионная кассета» относится к кодирующей последовательности ДНК или сегменту ДНК, кодирующему продукт экспрессии, который может быть вставлен в вектор в определенных сайтах рестрикции. Сайты рестрикции кассеты предназначены для обеспечения вставки кассеты в соответствующую рамку считывания. Как правило, чужеродная ДНК вставляется в один или более сайтов рестрикции векторной ДНК, а затем переносится вектором в клетку-хозяина вместе с трансмиссивной векторной ДНК. Сегмент или последовательность ДНК со встроенной или добавленной ДНК, например, вектор экспрессии, также можно назвать «конструкцией ДНК».
Термин «вектор», используемый в настоящем документе, включает автономно реплицирующиеся нуклеотидные последовательности, а также нуклеотидные последовательности, интегрирующиеся в геном. Распространенным типом вектора является «плазмида», которая обычно представляет собой автономную молекулу двухцепочечной ДНК, которая может легко принимать дополнительную (чужеродную) ДНК и может быть легко введена в подходящую клетку-хозяина. Плазмидный вектор часто содержит кодирующую ДНК и промоторную ДНК и имеет один или более сайтов рестрикции, подходящих для вставки чужеродной ДНК. В частности, термин «вектор» или «плазмида» относится к носителю, с помощью которого последовательность ДНК или РНК (например, чужеродный ген) может быть введена в клетку-хозяина, чтобы трансформировать хозяина и способствовать экспрессии (например, транскрипции и трансляции) введенной последовательности.
Можно использовать любую из известных процедур для введения чужеродных нуклеотидных последовательностей в клетки-хозяева. Они включают использование трансфекции с использованием фосфата кальция, полибрена, слияние протопластов, электропорацию, нуклеофекцию, липосомы, микроинъекцию, голую ДНК, плазмидные векторы, вирусные векторы, как эписомные, так и интегративные, и любые другие известные методы введения клонированной геномной ДНК, кДНК, синтетическую ДНК или другой чужеродный генетический материал в хозяйскую клетку (см., например, в источнике Sambrook et al). Растения могут быть трансформированы с помощью Agrobacterium-опосредоъшиой трансформации, бомбардировки частицами, вакуумной инфильтрации, трансформации in planta и химических методов, известных специалистам в данной области техники.
Следующие пункты являются конкретными вариантами осуществления изобретения, представленного в настоящем документе.
1. Способ биотрансформации трихотецена путем приведения материала, зараженного трихотеценами, в контакт с экзогенной глутатион-8-трансферазой (GST) неживотного происхождения, обладающей субстратной специфичностью к эпоксидному кольцу трихотецена, включающий этапы:
a) приведение материала в контакт с GST,
b) необязательно добавление глутатиона к указанному материалу, и
c) инкубирование смеси в водном растворе в диапазоне рН от 6 до 9 в условиях, при которых глутатион реагирует с эпоксидным фрагментом, тем самым образуя эпоксидный аддукт.
2. Способ по п. 1, в котором трихотецен выбран из группы трихотеценов типа А, трихотеценов типа В, трихотеценов типа С и трихотеценов типа D.
3. Способ по пп. 1 или 2, где трихотецен выбран группы, состоящей из токсина Т-2, токсина НТ-2, неозоланиола, дезоксиниваленола (ДОН), ниваленола (NIV), трихотецина, 3- и 15-ацетилдезоксиниваленола, роридина А и веррукарина А.
4. Способ по любому из пп. 1-3 для получения обеззараженной кормовой добавки, кормового материала, пищевой добавки или пищевого материала.
5. Способ по любому из пп. 1-3 для обеззараживания жидкого или твердого материала или поверхности материала, в частности, текстильного материала, фильтрующего материала, противогазов, систем кондиционирования воздуха.
6. Способ по любому из пп. 1-3 для очистки поверхности материала, животного или человека.
7. Способ по любому из пп. 1-6, где GST представляет собой рекомбинантную GST.
8. Способ по любому из пп. 1-7, где GST имеет аминокислотную последовательность SEQ ID NO. 2 или SEQ ID NO. 4 или последовательность, имеющую по меньшей мере 80%, 85%, 90%, 95%, в частности, по меньшей мере 99% идентичности последовательности с любой из SEQ ID NO. 2 или SEQ ID NO. 4 и обладающую субстратной специфичностью к эпоксидному кольцу трихотецена.
9. Способ по любому из пп. 1-8, где GST кодируется
a) полинуклеотидной последовательностью, выбранной из группы, состоящей из SEQ ID NO. 1 или SEQ ID NO. 3 или последовательности, имеющей по меньшей мере 80%, 85%, 90%, 95%, в частности, по меньшей мере 99% идентичности последовательности с любой из SEQ ID NO. 1 или SEQ ID NO. 3, или
b) выделенной молекулой нуклеиновой кислоты, комплементарной полинуклеотидной последовательности согласно а).
10. Способ по любому из пп. 1-9, где GST экспрессируется в клетке-хозяине.
11. Кормовая добавка или кормовой материал, содержащий экзогенную GST неживотного происхождения, обладающую субстратной специфичностью к эпоксидному кольцу трихотецена.
12. Кормовая добавка или кормовой материал по п. 11, содержащий часть трансгенного растения, ткань трансгенного растения, клетку трансгенного растения, семя или его потомство, в частности, лист, стебель, корень, семядолю или гипокотиль, каждый из которых содержит экзогенную GST, обладающую субстратной специфичностью к эпоксидному кольцу трихотецена.
13. Применение способа по любому из пп. 1-10 для ферментативной деградации трихотецена в кормах для животных или в пищеварительном тракте животных.
14. Применение способа по любому из пп. 1-10 для производства корма или пищевой добавки.
15. Кормовой или пищевой материал, содержащий обеззараженный растительный материал, полученный способом по любому из пп. 1-10.
16. Кормовая добавка по п. 15, в которой GST неживотного происхождения имеет аминокислотную последовательность SEQ ID NO. 2, или SEQ ID NO. 4 или последовательность, имеющую по меньшей мере 80%, 85%, 90%, 95%, в частности, по меньшей мере 99% идентичности последовательности с любой из SEQ ID NO. 2 или SEQ ID NO. 4 и обладающую субстратной специфичностью к эпоксидному кольцу трихотецена.
17. Клетка-хозяин, сверхэкспрессирующий эндогенную GST или трансформированный вектором, экспрессирующим экзогенную GST, при этом указанная GST обладает субстратной специфичностью к эпоксидному кольцу трихотеценов.
18. Клетка-хозяин согласно п. 17, где экзогенная GST содержит аминокислотную последовательность, выбранную из группы SEQ ID NO. 2, SEQ ID NO. 4, или последовательность, имеющую по меньшей мере 80%, 85%, 90%, 95%, в частности, по меньшей мере 99% идентичности последовательности с любой из SEQ ID NO. 2, SEQ ID NO. 4 и обладающую субстратной специфичностью к эпоксидному кольцу трихотецена.
19. Клетка-хозяин по пп. 17 или 18, который является прокариотом или эукариотом, в частности, представляет собой растительную клетку, животную клетку, грибковую клетку или бактериальную клетку.
20. Способ по любому из пп. 17-19, где GST кодируется
a) полинуклеотидной последовательностью, выбранной из группы, состоящей из SEQ ID NO. 1, SEQ ID NO. 3, или последовательностью, имеющей по меньшей мере 80%, 85%, 90%, 95%, в частности, по меньшей мере 99% идентичности последовательности с любой из SEQ ID NO. 1, SEQ ID NO. 3, или
b) выделенной молекулой нуклеиновой кислоты, комплементарной а).
21. Хозяин по любому из пп. 17-20, где вектор включает экспрессионную кассету, в частности, содержащую полинуклеотидную последовательность, кодирующую экзогенную GST, функционально связанную с регуляторной последовательностью, функционирующей в данном хозяине.
22. Хозяин по п. 21, где указанная кассета дополнительно содержит оцениваемый маркерный полинуклеотид, функционально связанный с регуляторной последовательностью, функциональной в растении.
23. Хозяин по пп. 21 или 22, где указанная кассета дополнительно содержит полинуклеотид селективного маркера, функционально связанный с регуляторной последовательностью, функциональной в указанном хозяине.
24. Хозяин по любому из пп. 21-23, обладающий субстратной специфичностью к трихотецену, выбранному из группы, состоящей из трихотеценов типа А, трихотеценов типа В, трихотеценов типа С и трихотеценов типа D.
25. Хозяин по любому из пп. 21-24, где трихотецен выбран из группы, состоящей из дезоксиниваленола (ДОН), 3- и 15-ацетилдезоксиниваленола, ниваленола (NIV), токсина Т2, токсина НТ-2, неозоланиола, диацетоксицирпенола, трихотецина, роридина А и веррукарина А.
26. Хозяин по любому из пп. 21-25, где экзогенная GST содержит аминокислотную последовательность, выбранную из группы SEQ ID NO. 2 или SEQ ID NO. 4 или последовательности, имеющей по меньшей мере 80%, 85%, 90%, 95%, в частности, по меньшей мере 99% идентичности последовательности с любой из SEQ ID NO. 2 или SEQ ID NO. 4 и обладающую субстратной специфичностью к эпоксидному кольцу трихотеценов.
27. Трансгенное растение с повышенной устойчивостью к трихотецену, содержащее экзогенную GST, обладающую субстратной специфичностью к эпоксидному кольцу трихотеценов.
28. Часть трансгенного растения, ткань трансгенного растения, клетка трансгенного растения, семя или его потомство, в частности, лист, стебель, корень, семядолю и гипокотиль, включающие экзогенную GST, обладающую субстратной специфичностью к эпоксидному кольцу трихотеценов.
29. Трансгенное животное с повышенной устойчивостью к трихотецену, содержащее экзогенную GST, обладающую субстратной специфичностью к эпоксидному кольцу трихотеценов.
30. Способ получения трансгенного растения или животного согласно пп. 27-29, включающий трансформацию растения или животного молекулой нуклеиновой кислоты, кодирующей GST, и экспрессию молекулы нуклеиновой кислоты в указанном растении, при этом молекула нуклеиновой кислоты включает полинуклеотидную последовательность, выбранную из группы, состоящей из полинуклеотидной последовательности, содержащей SEQ ID NO: 1 или SEQ ID NO. 3 или последовательности, имеющей по меньшей мере 80%, 85%, 90%, 95%, в частности, по меньшей мере 99% идентичности последовательности с любой из SEQ ID NO. 1 или SEQ ID NO. 3.
31. Способ по п. 30, в котором растение трансформируют методом, выбранным из группы, состоящей из Agrobacterium-опосредованной трансформации, биобаллистической бомбардировки, вакуумной инфильтрации, трансформации in planta и химического способа.
32. Способ экспрессии экзогенной GST, обладающей субстратной специфичностью к эпоксидному кольцу трихотецена в организме хозяина, включающий:
a) трансформацию клетки-хозяина вектором, содержащим последовательность гена, кодирующего GST,
b) выращивание трансформированной клетки в условиях, подходящих для экспрессии гена, кодирующего экзогенную GST.
33. Способ получения трансгенного растения по любому из пп. 28-30, где указанное растение является однодольным или двудольным растением.
34. Способ по п. 31, в котором однодольное растение представляет собой траву, в частности, выбранную из группы, состоящей из риса, маиса, пшеницы, ячменя и сорго, ржи, овса.
35. Способ по п. 34, в котором двудольное растение выбрано из группы, состоящей из табака, томата, гороха, сои, капусты, нута, арабидопсиса и моркови.
Следующие примеры приведены для облегчения понимания изобретения, но не предназначены и не должны быть истолкованы как ограничивающие каким-либо образом объем изобретения. Примеры не включают подробное описание традиционных методов. Такие методы хорошо известны специалистам в данной области техники.
ПРИМЕРЫ
Пример 1:
Создание экспрессионного хозяина:
Для детекции активности растительной GST в стандартном колориметрическом анализе был создан подходящий штамм-хозяин Е. coli, в котором инактивировали эндогенный ген GST (gstA), проявляющий активность со стандартным субстратом (2,4-динитрохлорбензолом (ДНХБ)) колориметрического анализа. Ген gstA в экспрессионном хозяине Е. coli «Т7 Express», основанном на полимеразе Т7, был разрушен следующим образом: Сначала плазмида pDK46 (с термочувствительным ориджином репликации и индуцируемой арабинозой системой рекомбинации фага лямбда (γ β ехо), полученная из штамма BW25113/pKD46 (http://cgsc2.biology.yale.edu/Strain.php?ID=68099), была введена в экспрессионный штамм Т7 Express. Затем индуцированные арабинозой компетентные клетки трансформировали продуктом ПЦР, полученным из мутантного штамма gstA JW1627-1 из коллекции систематических нокаутов Е. coli (http://cgsc2.biology.yale.edu/Strain.php?ID= 107667). ПЦР-продукт, полученный с помощью фланкирующих праймеров (Del_gstA_fw #3818 и Del_gstA_rv #3819), содержит ген устойчивости к канамицину. Нокаут-мутанты отбрали на среде LB-KAN при 37°С, что также приводит к потере pKD46. Замену гена подтверждали с помощью ПЦР. Сравнение штамма дикого типа и штамма после разрушения показано на фиг. 3 (А: дикий тип, В: после разрушения эндогенного гена gstA).

После разрушения полученный штамм «Т7 Express ΔgstA::KanR» имеет следующий генотип:
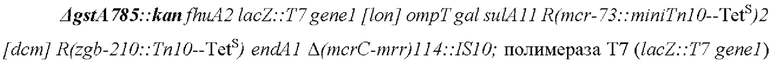 в хромосомном1 lac-опероне (без фага λDE3).
в хромосомном1 lac-опероне (без фага λDE3).
Указанный хозяин не имеет значительного фона в анализе на основе ДНХБ и был использован для тестирования многочисленных (конститутивно и индуцированных ДОН) кандидатных генов GST (из ячменя, риса, Brachypodium и пшеницы), большинство из которых показали четкую активность с ДНХБ, но не активность выше фона с ДОН.
Пример 2:
Клонирование и экспрессия кандидатов GST:
Кандидаты клонировали в вектор экспрессии Е. coli рСА02, который кодирует N-концевую 6х гистидиновую метку и домен связывания мальтозы. Вектор несет ориджин ColEl, маркер ампициллина, копию репрессора lac и последовательность промотора Т7 и оператора lac.
Пустой вектор рСА02 использовали в качестве отрицательного контроля.
Клонирование TrGST-C02:
Глутатион-S-трансфераза пшеницы
TRIAE_CS42_lDL_TGACvl_062916_АА0221650 амплифицировали из геномной ДНК китайской яровой пшеницы с праймерами TrGST-C02_upstr_fw и TrGST-C02_3'UTR_rv и клонировали в pMiniT (NEB® PCR Cloning Kit). Экзон 1 амплифицировали с праймерами TrGST-C02_GA-E1-fw и TrGST-C02_GA-E1-rv, экзон 2 амплифицировали с праймерами TrGST-C02_GA-E2-fw и TrGST-C02_GA-E2-rv из клона pMiniT. ПЦР с перекрывающимися праймерами проводили с праймерами TrGST-C02_GA-E1-fw и TrGST-C02_GA-E2-rv. Продукт ПЦР обработали Ndel/EcoRI и лигировали в рСА02, обработанный теми же ферментами.
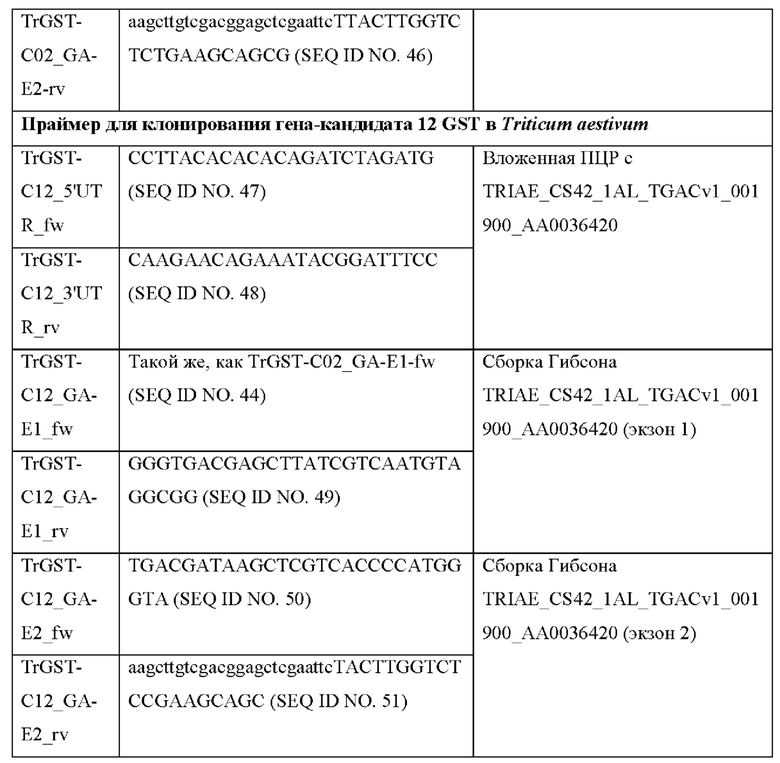
Клонирование TrGST-C12:
Глутатион-S-трансфераза пшеницы TRIAE_CS42_1AL_TGACvl_001900_AA0036420 амплифицировали из геномной ДНК китайской яровой пшеницы с праймерами TrGST-C12_5'UTR fw и TrGST-C12_3'UTR_rv и клонировали в pMiniT. Экзон 1 амплифицировали с праймерами TrGST-C12_GA-E1_fw и TrGST-C12_GA-E1_rv, экзон 2 амплифицировали с праймерами TrGST-C12_GA-E2_fw и TrGST-C12_GA-E2_rv из клона pMiniT. Сборку Гибсона выполняли с обработанной NdeI/EcoRI рСА02 и обоими продуктами ПЦР.


Нуклеотидные и аминокислотные последовательности экспрессированных GST, используемых в настоящем документе, приведены ниже:


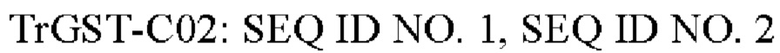

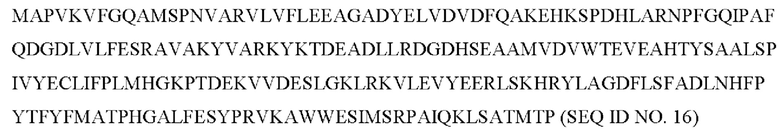





















Экспрессию проводили в 50 мл среды Terrific Broth с ампициллином (ТВ+100 ppm Amp), индуцированным 1 мМ изопропил-β-d-1-тиогалактопиранозидом (ИПТГ), 20°С в течение ночи. Осадки промывали в 1х фосфатно-солевом буфере (ФСБ: 50 мМ фосфат натрия, 150 мМ хлорид натрия, рН 7) и ресуспендировали в 2,5 мл 100 мМ фосфатно-солевого буфера рН 6, содержащего 10% глицерина. Клетки разрушали путем соникации (ультразвуковой лизис с помощью сонификатора Brason W250D: большой наконечник, амплитуда 40%, общее время импульса 20 сек).
Для оценки экспрессии слитых белков 6xHis-malE-(TEV)-GST использовали электрофорез в полиакриламидном геле с додецилсульфатом натрия (SDS-ПААГ) с 12% полиакриламидным гелем. Результаты показаны на фиг. 4.
2 мкл образцов разбавляли водой до 15 мкл + 5 мкл 4х SDS загрузочного красителя, инкубировали в течение 5 мин при 95°С и центрифугировали. Загружали 12 мкл (кроме: 5 мкл PageRuler™ Prestained Protein Ladder, ThermoFisher Scientific).
Белки с меткой 6xHis аффинно очищали методом метал-аффинной хроматографии (MAX, IMAC) и использовали для ферментативных анализов. Для сравнения также очищали GST Schistosoma japonicum, который не производит Дон-13-GSH (он показал примерно в 10 раз более высокую специфическую активность в отношении ДНХБ, но никакой активности в отношении эпоксида ДОН). Этот фермент экспрессировали из вектора pGEX-4T3, в котором отсутствует последовательность гистидиновой метки, и очищали с помощью аффинной хроматографии Glutathione Sepharose 4 В.
SDS-ПААГ: (Ожидаемый размер: TrGST-C02: 69,2 кДа; TrGST-C12: 69,2 кДа; SjGST: 26 кДа). Как описано выше, кандидатные GST клонировали в вектор рСА02 и экспрессировали с N-концевой меткой 6xHis и мальтозосвязывающим доменом.
На фиг. 5 показан SDS-ПААГ, применяемые концентрации: СЕ (клеточный экстракт) и FT (проточная фракция): 0,75 мг/мл, Белок: 0,23 мг/мл.
Ферментативные анализы конъюгации ДОН-GSH и аналитические измерения:
Конечные концентрации в анализе: Концентрация токсина в анализе: 100 ppm ДОН (исходная концентрация 1000 ppm ДОН в H2O), 5 мМ GSH, ОД М фосфатно-солевой буфер
рН 6, 7,5% глицерин.
Временные точки: 0, 0, 3, 24 ч.
Анализ орбитрэп (орбитальная ионная ловушка):
Аликвоты по 20 мкл образцов из анализа конъюгации ДОН-GSH смешивали с 20 мкл ацетонитрила в пробирке типа эппендорф и центрифугировали при 20 000 об/мин в течение 7,5 мин. 15 мкл супернатанта отбирали и смешивали с 135 мкл Н2О в пробирках, чтобы получить в целом 20-кратное разведение, при содержании 5% ацетонитрила и 5 ppm ДОН в этом разведении. Образцы измеряли в течение нескольких часов, а до измерения хранили при 4°С.
Для анализов детоксикации ДОН использовали HPLC-MS/MS (высокоэффективная жидкостная хромотография с тандемной масс-спектрометрией, ВЭЖХ-МС/МС). Образцы измеряли методом LC-HRMS(/MS) (жидкостная хроматография с масс-спектрометрией высокого разрешения) с использованием системы Vanquish (Thermo Scientific), соединенной с прибором QExactive HF Orbitrap (Thermo Scientific). Для этого 5 мкл охлажденного разбавленного супернатанта (10°С) вводили для градиентного элюирования на колонку Atlantis® (dC18, 3 мкм, 2,1×1500 мм, Waters) со скоростью потока 0,3 мл/мин. Двухступенчатое градиентное элюирование (элюент А: Н2О+0,1% муравьиной кислоты (FA); элюент В: МеОН+0,1% FA) начало увеличивать элюент В с 3 до 20% в течение 5,5 минут после одной минуты равновесия, затем последовало увеличение В с 20 до 100% в течение 2,5 минут. После 1,5 минут элюирования 100% В систему повторно калибровали в течение 3,5 минут (общее время работы 14 минут). Масс-спектры регистрировали с разрешением 120000 (при m/z, равном 200) в режиме полного сканирования с применением быстрого переключения полярности и диапазона сканирования от m/z 100 до 1000. Спектры фрагментации MS/MS регистрировали в позитивном режиме с разрешением 30 000 (при m/z, равном 200) и ступенчатой энергией столкновения (25, 35, 45) с применением зависящего от данных метода с использованием списка включений.
В принципе, два типа аддуктов ДОН с глутатионом образуются неферментативно. Образование аддукта Михаэля (к С10) происходит в течение нескольких дней, и, следовательно, в течение 24-часового периода наблюдения виден фон. Большинство ферментов не усиливают образование аддукта Михаэля выше этого фона (см. ниже). Плазмида pCA02 является пустым вектором экспрессии и кластеризуется с большинством (неактивных) GST. Исключение составляют два пшеничных фермента С02 и С12, которые явно усиливают образование аддуктов Михаэля.
Образование аддукта Михаэля «МН1» показано на фиг. 6.
Напротив, образование эпоксидного аддукта («МН2» с другим временем удерживания) не происходит в значительной степени в течение 24 часов. Пустой вектор и другие протестированные выше ферменты не дают измеримого фона, в то время как GST С02 и С12 пшеницы явно катализируют образование аддукта эпоксида (фиг. 8).
Идентичность пиков определяли путем сравнения со стандартом, полученным путем длительной инкубации ДОН и GSH (в течение нескольких недель), как описано Stanic с соавторами (https://www.ncbi.nlm.nih.gov/pubmed/27548277). В отличие от ферментов GST человека, протестированных этими авторами, С02 и С12 явно катализируют как образование аддукта Михаэля, так и раскрытие эпоксида.
Хроматограмма образца показана на фиг. 9 (реакция с пшеничной GST С02).
Кроме аддуктов глутатиона, фермент в меньших количествах образует продукты, не содержащие 2 атома водорода в части ДОН, которые, скорее всего, являются аддуктами GSH С3-кетопроизводных ДОН.
Пример 3:
Было показано, что TrGST-C02 и TrGST-C12 также могут раскрывать эпоксид трихотеценов типа А и макроциклических (тип D) трихотеценов. В следующих соединениях отсутствует сопряженная С8 кето-группа трихотеценов типа В (необходимая для образования аддукта Михаэля), поэтому образование аддукта происходит за счет раскрытия эпоксида. Расчеты ожидаемых масс аддуктов с соответствующими соединениями приведены в следующих таблицах.
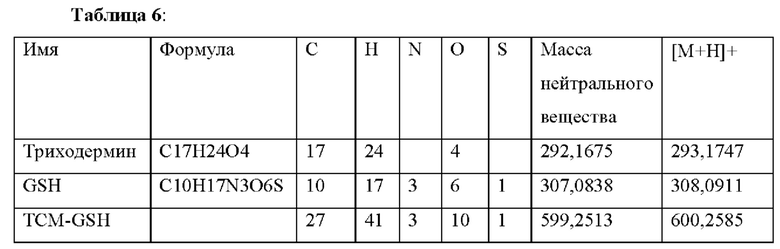
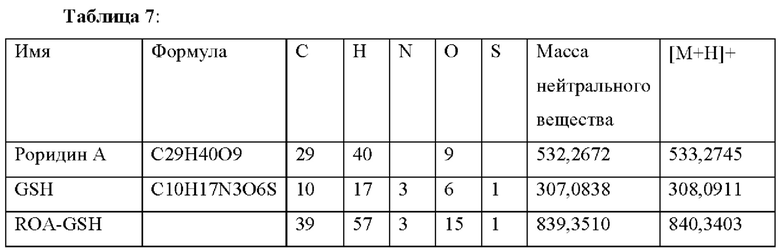
| название | год | авторы | номер документа |
|---|---|---|---|
| ФУНКЦИОНАЛЬНЫЕ ЛОКУСЫ FAD2 И СООТВЕТСТВУЮЩИЕ СПЕЦИФИЧНЫЕ ДЛЯ САЙТА-МИШЕНИ СВЯЗЫВАЮЩИЕСЯ БЕЛКИ, СПОСОБНЫЕ ИНДУЦИРОВАТЬ НАПРАВЛЕННЫЕ РАЗРЫВЫ | 2013 |
|
RU2656158C2 |
| ВЫДЕЛЕННАЯ МОЛЕКУЛА НУКЛЕИНОВОЙ КИСЛОТЫ, ХИМЕРНЫЙ ГЕН, РЕКОМБИНАНТНЫЙ ВЕКТОР, ШТАММ БАКТЕРИЙ, ТОКСИН, ИНСЕКТИЦИДНАЯ КОМПОЗИЦИЯ, СПОСОБ ПОЛУЧЕНИЯ ТОКСИНА, СПОСОБ ПОЛУЧЕНИЯ УСТОЙЧИВОГО К НАСЕКОМЫМ РАСТЕНИЯ И СПОСОБ БОРЬБЫ С НАСЕКОМЫМИ | 1999 |
|
RU2240003C2 |
| СПОСОБЫ И ВЕЩЕСТВА ДЛЯ ОСНОВАННОГО НА РЕКОМБИНАЦИИ ПОЛУЧЕНИЯ СОЕДИНЕНИЙ ШАФРАНА | 2012 |
|
RU2676730C2 |
| ПЕСТИЦИДНЫЕ КОМПОЗИЦИИ И ОТНОСЯЩИЕСЯ К НИМ СПОСОБЫ | 2013 |
|
RU2662672C2 |
| ИНСЕКТИЦИДНЫЕ БЕЛКИ | 2010 |
|
RU2613778C2 |
| ЛОКУСЫ ФУНКЦИОНАЛЬНОСТИ FAD2 И СООТВЕТСТВУЮЩИЕ СПЕЦИФИЧНЫЕ К УЧАСТКУ-МИШЕНИ СВЯЗЫВАЮЩИЕ БЕЛКИ, СПОСОБНЫЕ ИНДУЦИРОВАТЬ НАПРАВЛЕННЫЕ РАЗРЫВЫ | 2013 |
|
RU2656159C2 |
| ЛОКУСЫ FAD3 ДЛЯ ВЫПОЛНЕНИЯ ОПЕРАЦИЙ И СООТВЕТСТВУЮЩИЕ СВЯЗЫВАЮЩИЕСЯ СО СПЕЦИФИЧЕСКИМИ САЙТАМИ-МИШЕНЯМИ БЕЛКИ, СПОСОБНЫЕ К ВЫЗОВУ НАПРАВЛЕННЫХ РАЗРЫВОВ | 2013 |
|
RU2665811C2 |
| СПОСОБ ДЕТОКСИКАЦИИ МИКОТОКСИНОВ | 2004 |
|
RU2347810C2 |
| ПОСЛЕДОВАТЕЛЬНОСТИ СИНТЕТИЧЕСКИХ НУКЛЕОТИДОВ, КОДИРУЮЩИХ ИНСЕКТИЦИДНЫЙ КРИСТАЛЛИЧЕСКИЙ БЕЛОК, И ИХ ПРИМЕНЕНИЯ | 2020 |
|
RU2820699C2 |
| УСТОЙЧИВЫЕ К БАКТЕРИЯМ ТРАНСГЕННЫЕ РАСТЕНИЯ | 2012 |
|
RU2640246C2 |

Изобретение относится к биотехнологии, в частности к способу детоксикации материала, зараженного трихотеценом. Более конкретно, настоящее изобретение относится к способу биотрансформации трихотецена путем приведения материала, зараженного трихотеценами, в контакт с экзогенной глутатион-S-трансферазой (GST) неживотного происхождения, обладающей субстратной специфичностью к эпоксидному кольцу трихотецена. Настоящее изобретение также относится к рекомбинантным GST и трансгенным растениям и животным, экспрессирующим указанные GST. Изобретение позволяет проводить биотрансформацию трихотецена с высокой степенью эффективности. 15 н. и 13 з.п. ф-лы, 17 ил., 7 табл., 3 пр.
1. Способ биотрансформации трихотецена путем приведения материала, зараженного трихотеценами, в контакт с экзогенной глутатион-S-трансферазой (GST) неживотного происхождения, обладающей субстратной специфичностью к эпоксидному кольцу трихотецена, где GST представляет собой рекомбинантную GST, имеющую аминокислотную последовательность SEQ ID NO. 2 или SEQ ID NO. 4 или последовательность, имеющую по меньшей мере 80%, 85%, 90%, 95% или 99% идентичности последовательности с любой из SEQ ID NO. 2 или SEQ ID NO. 4, включающий этапы:
a) приведение материала в контакт с GST,
b) необязательно добавление глутатиона к указанному материалу и
c) инкубирование смеси в водном растворе в диапазоне pH от 6 до 9 в условиях, при которых глутатион реагирует с эпоксидным фрагментом, тем самым образуя эпоксидный аддукт.
2. Способ по п. 1, в котором трихотецен выбран из группы, состоящей из трихотеценов типа А, трихотеценов типа В, трихотеценов типа С и трихотеценов типа D, или трихотецен выбран из группы, состоящей из токсина T-2, токсина HT-2, неозоланиола, дезоксиниваленола (ДОН), ниваленола (NIV), трихотецина, 3- и 15-ацетилдезоксиниваленола, роридина A и веррукарина A.
3. Способ по п. 1 или 2 для применения в производстве обеззараженной кормовой добавки, кормового материала, пищевой добавки или пищевого материала, для обеззараживания жидкого или твердого материала или поверхности материала, или текстильного материала, фильтрующего материала, противогазов, систем кондиционирования воздуха, или для очистки поверхности материала, поверхности животного или человека.
4. Способ по любому из пп. 1-3, где GST кодируется
a) полинуклеотидной последовательностью, выбранной из группы, состоящей из SEQ ID NO. 1 или SEQ ID NO. 3 или последовательности, имеющей по меньшей мере 80%, 85%, 90%, 95% или 99% идентичности последовательности с любой из SEQ ID NO. 1 или SEQ ID NO. 3, или
b) выделенной молекулой нуклеиновой кислоты, комплементарной полинуклеотидной последовательности согласно a).
5. Способ по любому из пп. 1-4, где GST экспрессируется в клетке-хозяине.
6. Кормовая добавка, содержащая экзогенную неживотную GST, которая обладает субстратной специфичностью к эпоксидному кольцу трихотеценов, где GST неживотного происхождения имеет аминокислотную последовательность SEQ ID NO. 2 или SEQ ID NO. 4 или последовательность, имеющую по меньшей мере 80%, 85%, 90%, 95% или 99% идентичности последовательности с любой из SEQ ID NO. 2 или SEQ ID NO. 4.
7. Кормовой материал, содержащий экзогенную неживотную GST, которая обладает субстратной специфичностью к эпоксидному кольцу трихотеценов, где GST неживотного происхождения имеет аминокислотную последовательность SEQ ID NO. 2 или SEQ ID NO. 4 или последовательность, имеющую по меньшей мере 80%, 85%, 90%, 95% или 99% идентичности последовательности с любой из SEQ ID NO. 2 или SEQ ID NO. 4.
8. Кормовая добавка по п. 6, содержащая часть трансгенного растения, ткань трансгенного растения, клетку трансгенного растения, семя или его потомство или лист, стебель, корень, семядолю или гипокотиль, каждый из которых содержит экзогенную GST, обладающую субстратной специфичностью к эпоксидному кольцу трихотеценов.
9. Кормовой материал по п. 7, содержащий часть трансгенного растения, ткань трансгенного растения, клетку трансгенного растения, семя или его потомство или лист, стебель, корень, семядолю или гипокотиль, каждый из которых содержит экзогенную GST, обладающую субстратной специфичностью к эпоксидному кольцу трихотеценов.
10. Применение способа по любому из пп. 1-5 для ферментативной деградации трихотецена в кормах для животных или в пищеварительном тракте животных.
11. Применение способа по любому из пп. 1-5 для производства кормовой добавки.
12. Применение способа по любому из пп. 1-5 для производства пищевой добавки.
13. Клетка-хозяин для экспрессии эндогенной GST, где клетка-хозяин содержит эндогенную GST или трансформирована вектором, экспрессирующим экзогенную GST, при этом указанная GST обладает субстратной специфичностью к эпоксидному кольцу трихотеценов, где GST имеет аминокислотную последовательность SEQ ID NO. 2 или SEQ ID NO. 4 или последовательность, имеющую по меньшей мере 80%, 85%, 90%, 95% или 99% идентичности последовательности с любой из SEQ ID NO. 2 или SEQ ID NO. 4.
14. Клетка-хозяин по п. 13, которая является прокариотом или эукариотом или это растительная клетка, животная клетка, грибковая клетка или бактериальная клетка.
15. Клетка-хозяин по любому из пп. 13 или 14, где GST кодируется
a) полинуклеотидной последовательностью, выбранной из группы, состоящей из SEQ ID NO. 1, SEQ ID NO. 3 или последовательности, имеющей по меньшей мере 80%, 85%, 90%, 95% или 99% идентичности последовательности с любой из SEQ ID NO. 1, SEQ ID NO. 3, или
b) выделенной молекулой нуклеиновой кислоты, комплементарной a).
16. Клетка-хозяин по любому из пп. 13, 14, содержащая GST, обладающую субстратной специфичностью к трихотецену, выбранному из группы, состоящей из трихотеценов типа A, трихотеценов типа B, трихотеценов типа C и трихотеценов типа D, или выбранного из группы, состоящей из дезоксиниваленола (ДОН), 3- и 15-ацетилдезоксиниваленола, ниваленола (NIV), токсина T-2, токсина HT-2, неозоланиола, диацетоксицирпенола, трихотецина, роридина A и веррукарина A.
17. Трансгенное растение, имеющее повышенную устойчивость к трихотецену, содержащее экзогенную GST, обладающую субстратной специфичностью к эпоксидному кольцу трихотеценов, где GST представляет собой рекомбинантную GST, имеющую аминокислотную последовательность SEQ ID NO. 2 или SEQ ID NO. 4 или последовательность, имеющую по меньшей мере 80%, 85%, 90%, 95% или 99% идентичности последовательности с любой из SEQ ID NO. 2 или SEQ ID NO. 4.
18. Ткань трансгенного растения, имеющая повышенную устойчивость к трихотецену, содержащая экзогенную GST, обладающую субстратной специфичностью к эпоксидному кольцу трихотеценов, где GST представляет собой рекомбинантную GST, имеющую аминокислотную последовательность SEQ ID NO. 2 или SEQ ID NO. 4 или последовательность, имеющую по меньшей мере 80%, 85%, 90%, 95% или 99% идентичности последовательности с любой из SEQ ID NO. 2 или SEQ ID NO. 4.
19. Клетка трансгенного растения, имеющая повышенную устойчивость к трихотецену, содержащее экзогенную GST, обладающую субстратной специфичностью к эпоксидному кольцу трихотеценов, где GST представляет собой рекомбинантную GST, имеющую аминокислотную последовательность SEQ ID NO. 2 или SEQ ID NO. 4 или последовательность, имеющую по меньшей мере 80%, 85%, 90%, 95% или 99% идентичности последовательности с любой из SEQ ID NO. 2 или SEQ ID NO. 4.
20. Семя трансгенного растения, имеющее повышенную устойчивость к трихотецену, содержащее экзогенную GST, обладающую субстратной специфичностью к эпоксидному кольцу трихотеценов, где GST представляет собой рекомбинантную GST, имеющую аминокислотную последовательность SEQ ID NO. 2 или SEQ ID NO. 4 или последовательность, имеющую по меньшей мере 80%, 85%, 90%, 95% или 99% идентичности последовательности с любой из SEQ ID NO. 2 или SEQ ID NO. 4.
21. Трансгенное животное, не являющееся человеком, с повышенной устойчивостью к трихотецену, содержащее экзогенную GST неживотного происхождения, обладающую субстратной специфичностью к эпоксидному кольцу трихотеценов, где GST представляет собой рекомбинантную GST, имеющую аминокислотную последовательность SEQ ID NO. 2 или SEQ ID NO. 4 или последовательность, имеющую по меньшей мере 80%, 85%, 90%, 95% или 99% идентичности последовательности с любой из SEQ ID NO. 2 или SEQ ID NO. 4.
22. Способ получения трансгенного растения по любому из пп. 17-20, включающий трансформацию растения молекулой нуклеиновой кислоты, кодирующей GST, обладающую субстратной специфичностью к эпоксидному кольцу трихотецена, и экспрессию молекулы нуклеиновой кислоты в указанном растении, при этом молекула нуклеиновой кислоты содержит полинуклеотидную последовательность, выбранную из группы, состоящей из полинуклеотидной последовательности, содержащей SEQ ID NO. 1, SEQ ID NO. 3, или последовательности, имеющей по меньшей мере 80%, 85%, 90%, 95% или 99% идентичности последовательности с любой из SEQ ID NO. 1 или SEQ ID NO. 3.
23. Способ получения трансгенного животного по п. 21, включающий трансформацию животного молекулой нуклеиновой кислоты, кодирующей GST, обладающую субстратной специфичностью к эпоксидному кольцу трихотецена, и экспрессию молекулы нуклеиновой кислоты в указанном животном, при этом молекула нуклеиновой кислоты содержит полинуклеотидную последовательность, выбранную из группы, состоящей из полинуклеотидной последовательности, содержащей SEQ ID NO. 1, SEQ ID NO. 3, или последовательности, имеющей по меньшей мере 80%, 85%, 90%, 95% или 99% идентичности последовательности с любой из SEQ ID NO. 1 или SEQ ID NO. 3.
24. Способ по п. 22, при котором растение трансформируют методом, выбранным из группы, состоящей из Agrobacterium-опосредованной трансформации, биобаллистической бомбардировки, вакуумной инфильтрации, трансформации in planta и химического метода.
25. Способ экспрессии экзогенной GST, обладающей субстратной специфичностью к эпоксидному кольцу трихотеценов, в организме хозяина, где хозяин является таким, как он определен в п. 13, включающий:
a) трансформацию указанной клетки-хозяина вектором, содержащим последовательность гена, кодирующего GST,
b) выращивание трансформированной клетки в условиях, подходящих для экспрессии гена, кодирующего экзогенную GST.
26. Способ получения трансгенного растения по п. 22, где указанное растение является однодольным или двудольным растением.
27. Способ по п. 26, в котором однодольное растение представляет собой траву, в частности, выбранную из группы, состоящей из риса, маиса, пшеницы, ячменя и сорго, ржи, овса.
28. Способ по п. 27, в котором двудольное растение выбирают из группы, состоящей из табака, томата, гороха, сои, капусты, нута, арабидопсиса и моркови.
| WO 2015169847 A1, 12.11.2015 | |||
| US 6500639 B2, 31.12.2002 | |||
| Прочищалка для примуса и т.п. горелок | 1926 |
|
SU6737A1 |

