ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Данное изобретение относится к области биотехнологии, в частности к разработке белковых гормонов с помощью технологии рекомбинантной ДНК и их получению в клетках млекопитающих.
УРОВЕНЬ ТЕХНИКИ
Хорионический гонадотропин лошади (eCG, Equine Chorionic Gonadotropin) является членом семейства гликопротеиновых гормонов вместе с лютеинизирующим гормоном (LH, Luteinizing Hormone), фолликулостимулирующим гормоном (FSH, Follicle-Stimulating Hormone) и тиреотропным гормоном (TSH, Thyroid Stimulating Hormone) (Murphy and Martinuk, 1991). Поскольку eCG продуцируется трофобластными клетками эндометриальных чаш жеребых кобыл, первоначально он назывался гонадотропином сыворотки жеребых кобыл (PMSG, Pregnant Mare Serum Gonadotropin). eCG играет важную роль в поддержании ранней беременности (в течение первых трех месяцев), косвенно стимулируя выработку прогестерона желтым телом до тех пор, пока плацента не сможет секретировать его самостоятельно. Концентрация eCG, секретируемого трофобластами, достигает максимальной концентрации на около 50-й день беременности, а затем начинает прогрессивно снижаться (Allen and Moor, 1972).
eCG обладает двумя отличительными характеристиками по сравнению с другими гликопротеиновыми гормонами. С одной стороны, у видов, не относящихся к лошадиным, eCG проявляет высокую FSH- и LH-подобную активность и обладает высокой аффинностью к рецепторам этих гормонов (Combamous et al., 1984). С другой стороны, он отличается высоким содержанием углеводов, которое составляет 45% от его общей молекулярной массы. Данное последнее свойство определяет длительный период полувыведения eCG из кровотока, составляющий около шести дней. Благодаря обеим характеристикам eCG используется в ветеринарии для контроля репродуктивной активности различных видов скота, в том числе крупного рогатого скота, овец, коз и свиней (Rensis and Lopez-Gatius, 2014).
Как и другие члены семейства гликопротеиновых гормонов, eCG представляет собой гетеродимерный белок, состоящий из двух разных нековалентно-связанных субъединиц, называемых α и β. Субъединица α является общей для всех членов семейства и кодируется одним геном, тогда как субъединицы β, которые придают гетеродимерам специфичность, кодируются разными генами (Stewart and Allen, 1976).
Субъединица а состоит из 96 аминокислот и имеет два сайта N-гликозилирования, расположенных в Asn56 и Asn82, в то время как субъединица β состоит из 149 аминокислот и имеет единственный сайт N-гликозилирования в Asn13. Кроме того, субъединица β имеет карбоксиконцевой пептид (СТР), состоящий из 28 аминокислот (122-149), который содержит 12 сайтов О-гликозилирования в остатках Ser или Thr (Bousfield and Butnev, 2001). Обе субъединицы содержат множественные внутримолекулярные дисульфидные связи, и их сборка происходит главным образом в эндоплазматическом ретикулуме, что представляет собой лимитирующий этап в процессе секреции димеров (Hoshina and Boime, 1982). У лошадей β-субъединицы плацентарного CG и гипофизарного LH кодируются одним и тем же геном (Sherman et al., 1992). Однако eCG состоит из более высокого и более разветвленного содержания углеводов, чем eLH. Оба гормона значительно различаются по терминальным частям N-гликана. eCG содержит гликаны, кэпированные N-ацетилнейраминовой кислотой (сиаловой кислотой), в то время как eLII имеют сульфатированные N-ацетилгалактозаминовые (SO44--GalNAc) гликаны. Заметная разница в их молекулярной массе в основном обусловлена наличием более длинных дисиалированных поли-N-ацетиламинов O-гликановых структур в eCG (Smith et al., 1993). Высокое содержание сиаловой кислоты в eCG является причиной его исключительного периода полувыведения из кровотока, поскольку этот остаток снижает как клубочковую фильтрацию в почках, так и метаболизм в печени.
В настоящее время продукты, доступные на рынке, представляют собой частично очищенные составы eCG из крови жеребых кобыл (PMSG), обладающие многими недостатками. С одной стороны, они демонстрируют различия между сериями, поскольку профиль гликозилирования различается между животными и между сыворотками на разных стадиях беременности. С другой стороны, PMSG также может содержать загрязняющие вещества, потенциально опасные для здоровья. Это идет вразрез с текущей тенденцией регулирующих органов получать более безопасные ветеринарные продукты, очищенные от вирусов, прионов и других загрязняющих белков. Наконец, что не менее важно, практика получения сыворотки, содержащей eCG, от жеребой кобылы включает еженедельный забор 10 литров крови и последующую индукцию искусственного аборта путем проникновения в матку и разрыва амниотического мешка. Практика, которой подвергается животное, ставит под угрозу благополучие животного и совершенно сомнительна с биоэтической точки зрения: это кровавый процесс, который может вызвать серьезные анемии и в некоторых случаях заканчивается смертью животного.
По этой причине целесообразна разработка рекомбинантного eCG (reCG) в качестве заменителя PMSG. Сообщалось о нескольких попытках получения рекомбинантного eCG на разных хозяевах. С одной стороны, Legardinier et al. (2005) описали получение eCG в двух линиях клеток насекомых, Sf9 и Mimic™, при этом последняя представляет собой клеточную линию, полученную из первой, которая была модифицирована для экспрессии различных генов гликозилтрансфераз млекопитающих.
Тем не менее, продуцируемый гормон не проявлял FSH/LH-подобной активности на крысиных моделях in vivo, что авторы связывают с его чрезвычайно коротким периодом полувыведения из кровотока, обусловленным отсутствием терминальной сиаловой кислоты в цепях олигосахаридов. С другой стороны, Ubach et al. (2009) и Ingles et al. (2012) описали экспрессию и очистку eCG у Pichia pastohs; тем не менее, рекомбинантный гормон не проявлял биологической активности в биологическом анализе in vivo на самках крыс. Опять же, эти результаты коррелировали с коротким периодом полувыведения рекомбинантного гормона из кровотока, поскольку только 1% введенного белка можно было обнаружить в сыворотке через 90 минут. Такой быстрый клиренс можно объяснить активацией маннан-связывающего лектинового пути, который происходит после инъекции белка с высоким содержанием маннозы. Эти результаты демонстрируют важность правильно профиля гликозилирования для проявления биоактивности eCG in vivo и, следовательно, важность правильного выбора хозяина для его рекомбинантной экспрессии.
О разработке рекомбинантных версий eCG в клетках СНО, включая клеточную линию СНО DG44, сообщалось в публикациях и патентах. В заявке на патент WO 2017112987 А1 описано использование данной линии клеток СНО и получение reCG с ожидаемым профилем гликозилирования в зависимости от используемого хозяина. Несмотря на это, коммерческие версии reCG еще не доступны на рынке. Данный результат свидетельствует о проблеме, требующей решения, заключающейся в том, что большие количества рекомбинантного eCG (или вообще рекомбинантных гонадотропинов) не могут быть получены «эффективным способом» или с профилем гликозилирования, как у природной eCG, что гарантирует его биологическую активность in vivo (Hesser, 2011). Таким образом, задача состоит в том, чтобы разработать систему производства, которая демонстрирует высокую производительность для получения достаточного количества reCG, удовлетворяющего высокий спрос (поскольку гормон используется в различных видах домашнего скота) и, следовательно, приводит к снижению производственных затрат, что является ограничивающим фактором для успеха рекомбинантного гормона на рынке. WO2017112987A1_2017 описывает использование системы амплификации гена DHFR-MTX, позволяющей получить продукцию 18 МЕ/мл (в бессывороточной среде). Это значение не обеспечивает рентабельность процесса по сравнению с текущим производственным процессом PMSG.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
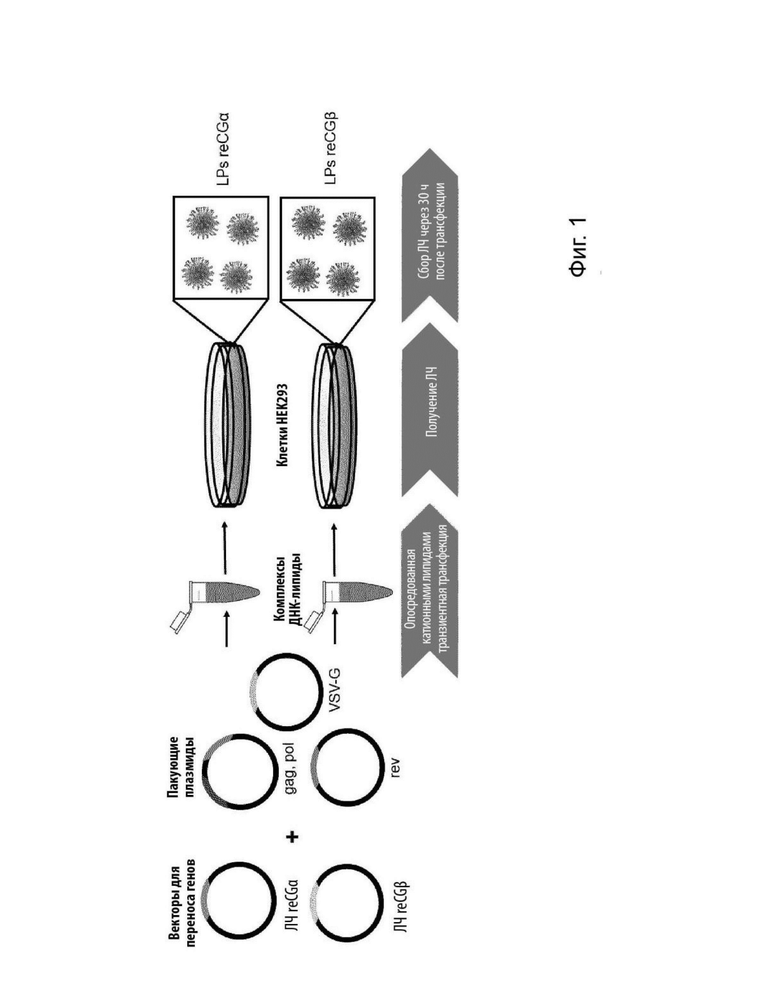
На Фиг. 1 представлено схематическое изображение сборки лентивирусных частиц третьего поколения для получения линий рекомбинантных клеток СНО-K1, продуцирующих reCG.
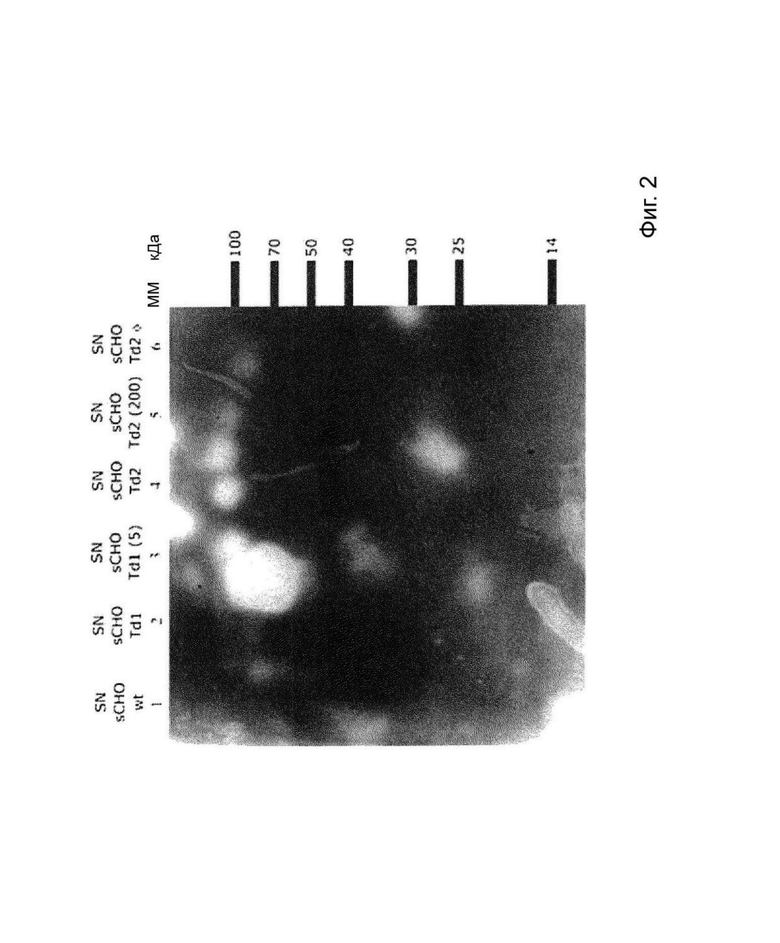
На Фиг. 2 изображен ДСН-ПААГ-электрофорез с последующим вестерн-блоттингом, при котором концентрацию reCG в супернатантах, полученных из линий, продуцирующих reCG, оценивают с использованием специфических антител против reCG.
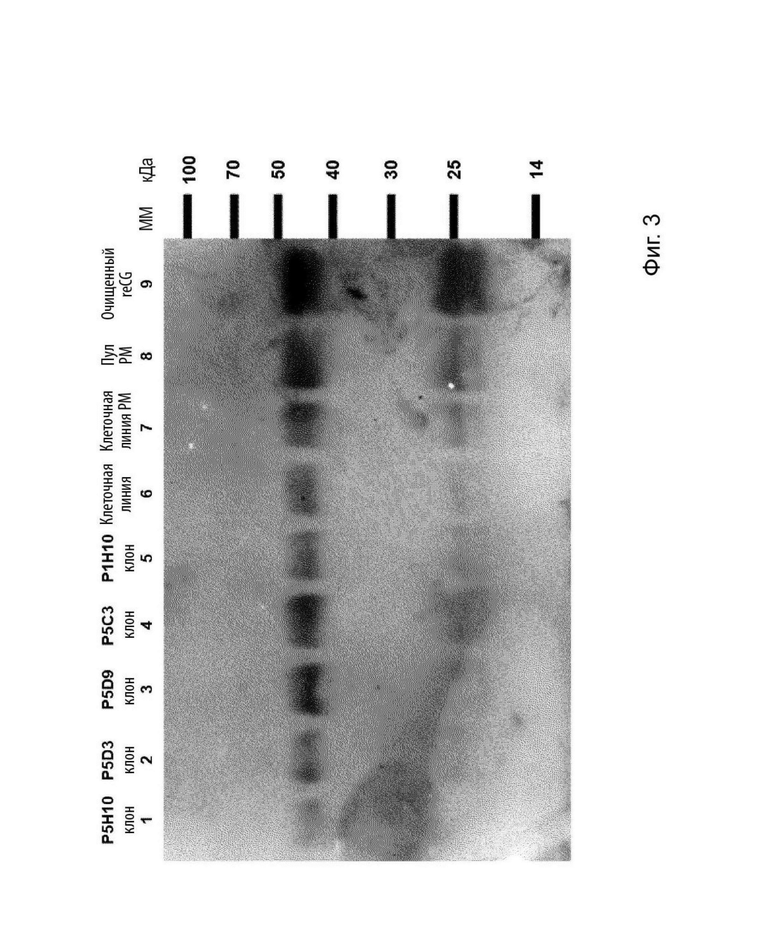
На Фиг. 3 изображен ДСН-ПААГ-электрофорез с последующим вестерн-блоттингом, при котором оценивается «наблюдаемая» продуктивность предварительно отобранных клонов, продуцирующих reCG.
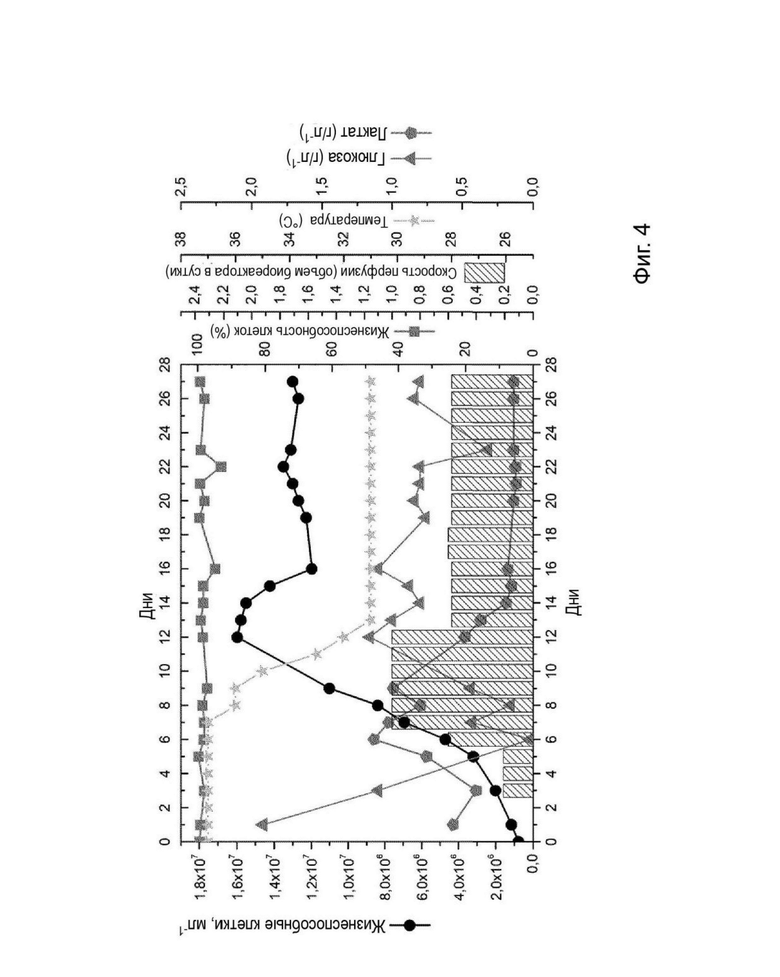
На Фиг. 4 представлен график, сравнивающий изменение концентрации жизнеспособных клеток, жизнеспособности, концентрации лактата и глюкозы во времени в культуре клона Р5С3 в биореакторе объемом 1 л. Показаны температура и скорость перфузии в сутки, (↓) представляет слив из биореактора.
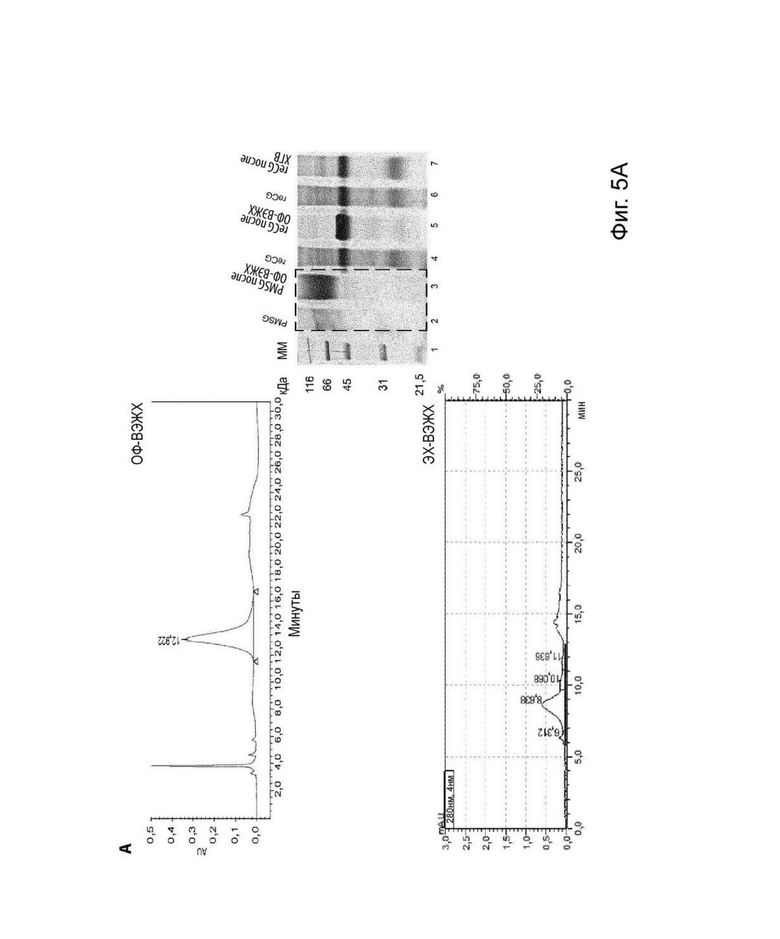
На Фиг. 5 представлена структурная характеризация и анализ чистоты (А) молекулы PMSG после ОФ-ВЭЖХ, (В) reCG ОФ-ВЭЖХ и (С) reCG ХГВ с помощью ДСН-ПААГ, ОФ-ВЭЖХ и ЭХ-ВЭЖХ.
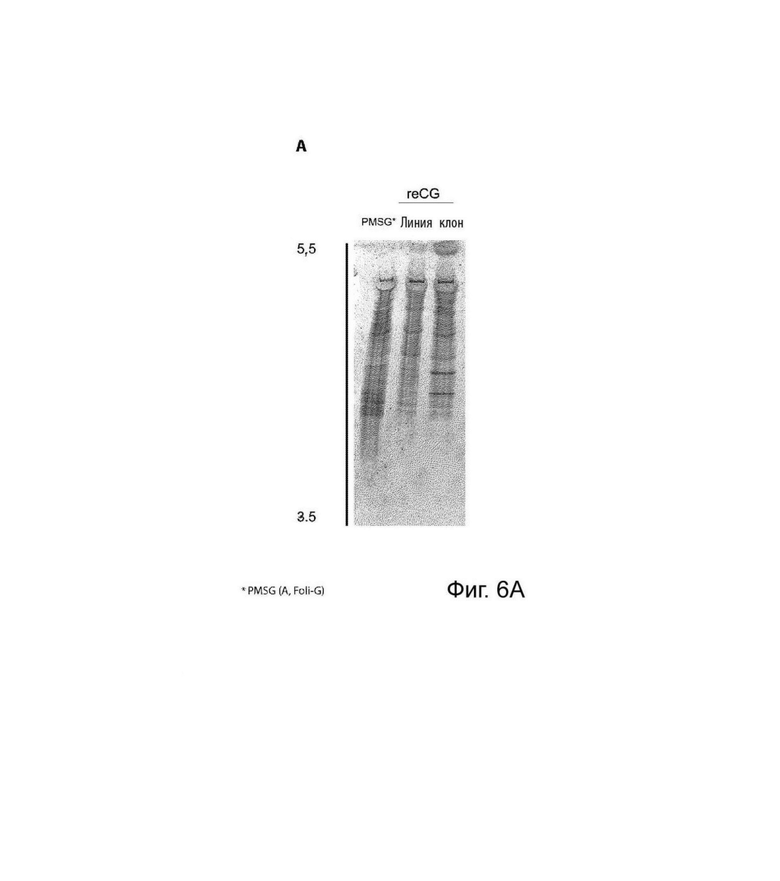
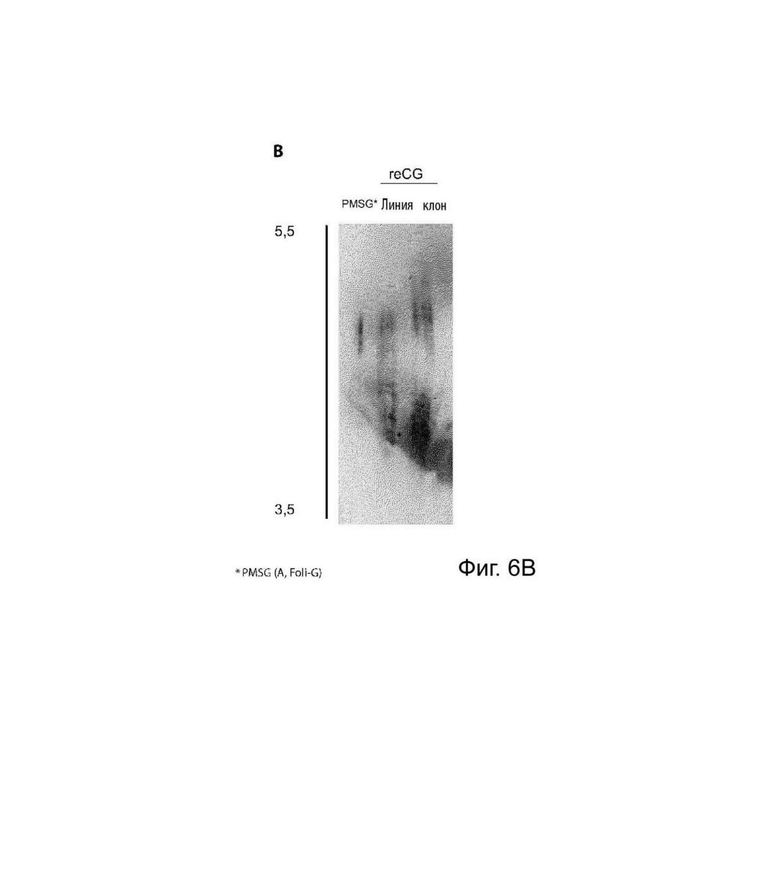
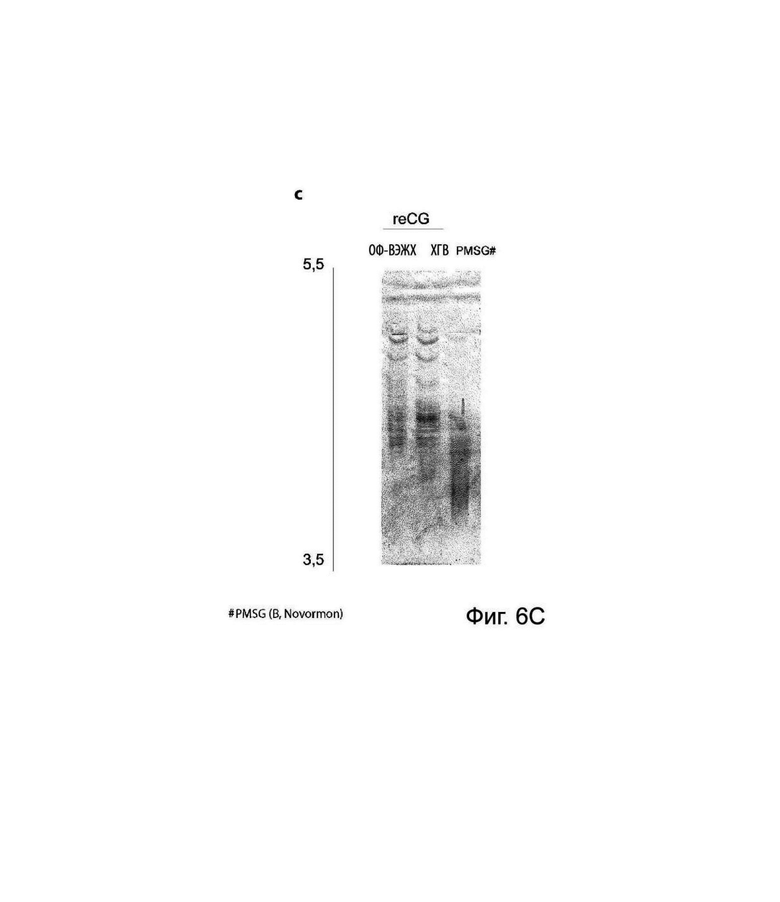
На Фиг. 6 представлен анализ профиля изоформ. (А) Анализ ИЭФ с последующим окрашиванием коллоидным кумасси синим. Сравнивали изоформный профиль варианта PMSG (коммерческий препарат A, Foli-G) и элюата reCG CaptoB из клеточной линии и клона, соответственно. (В) Анализ ИЭФ с последующим вестерн-блоттингом PMSG (коммерческий препарат A, Foli-G) и элюата reCG CaptoB из клеточной линии и клона, соответственно. (С) Анализ ИЭФ с последующим окрашиванием коллоидным кумасси синим. Сравнивали изоформный профиль PMSG (коммерческий препарат В, Novormon), вариантов reCG ОФ-ВЭЖХ и reCG ХГВ, соответственно.
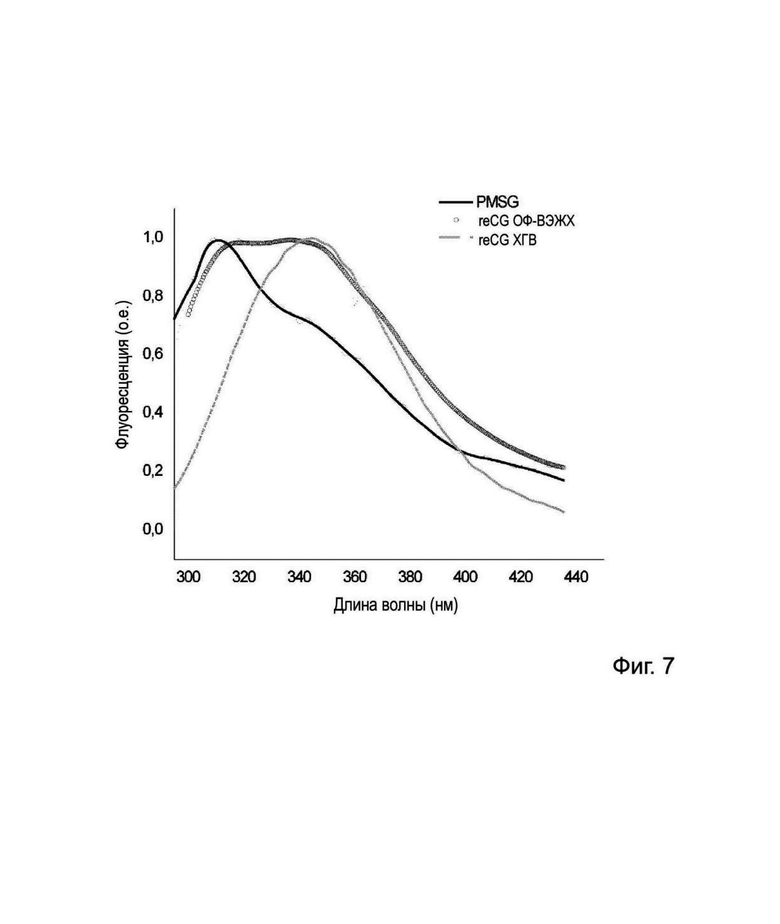
На Фиг. 7 представлен спектр излучения флуоресценции PMSG, reCG ОФ-ВЭЖХ и reCG ХГВ в буферном растворе фосфата натрия.
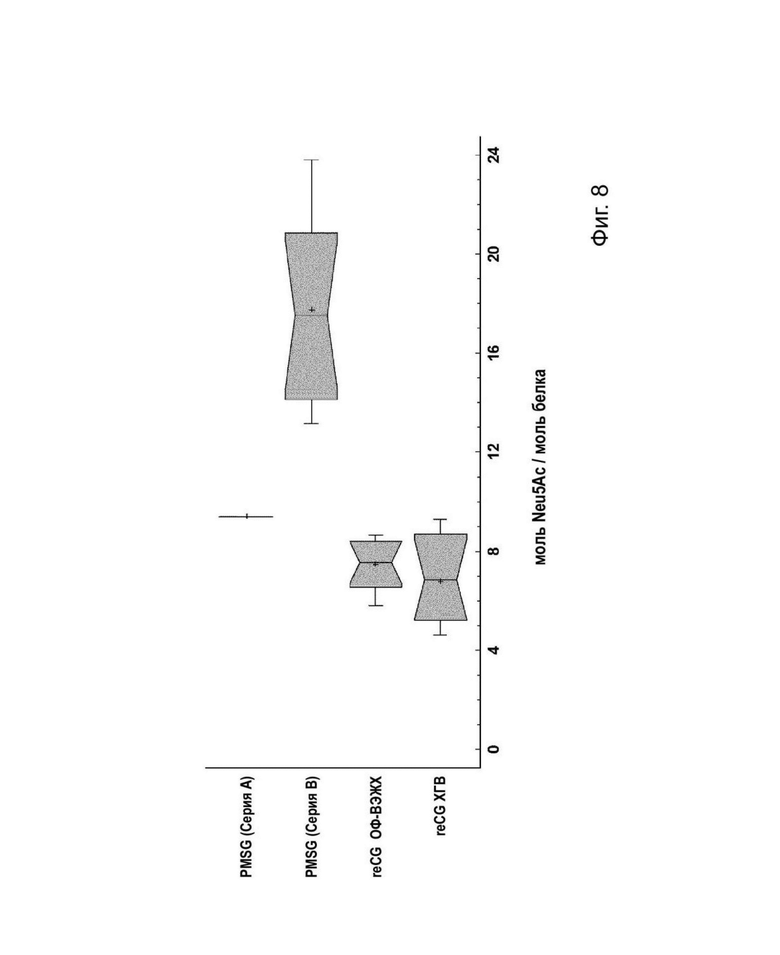
На Фис. 8 представлены коробчатые диаграммы с доверительными интервалами («зарубками»), на которых изображено значимое различие в содержании Neu5Ac (моль/моль белка).
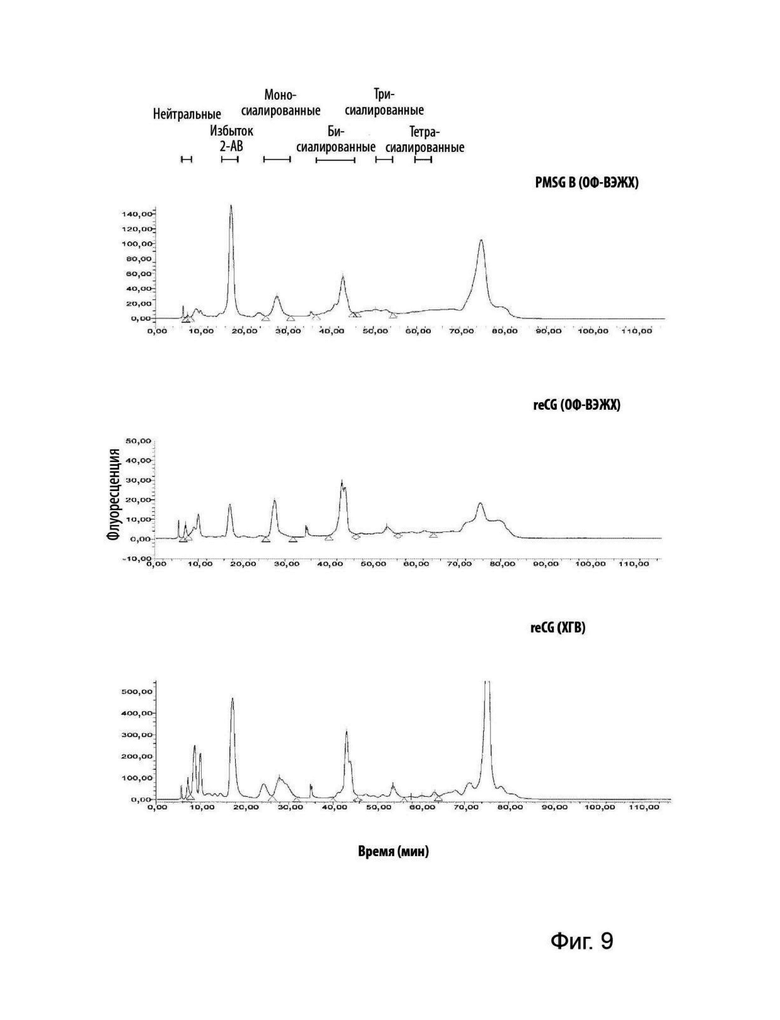
На Фиг. 9 представлен анализ 2-АВ-меченого N-гликана PMSG, reCG ОФ-ВЭЖХ и reCG ХГВ с помощью WAX-ВЭЖХ. N-гликаны разделяли в соответствии с их зарядом.
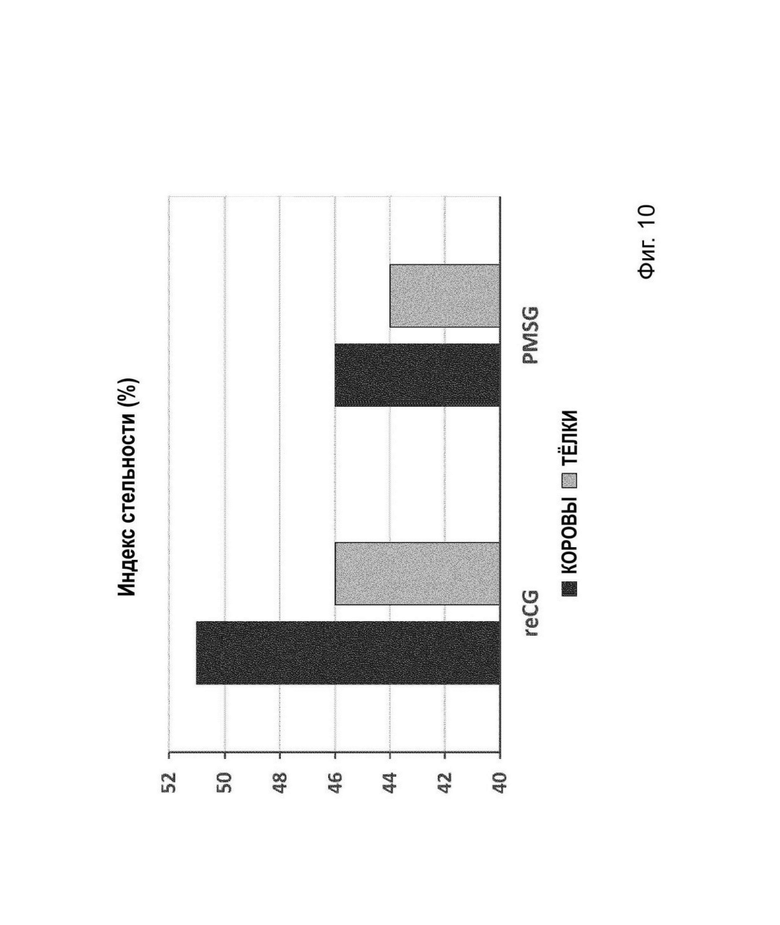
На Фиг. 10 представлен сравнительный индекс стельности.
КРАТКОЕ ОПИСАНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
В данном изобретении описан способ получения линии клеток млекопитающих, экспрессирующих рекомбинантный хорионический гонадотропин лошади (reCG), включающий следующие этапы:
a. получение кодирующих последовательностей α- и β-субъединиц reCG, оптимизированных для их экспрессии в клетках млекопитающих;
b. введение указанных кодирующих последовательностей в лентивирусные векторы экспрессии;
c. получение лентивирусов, содержащих кодирующую последовательность reCG;
d. трансдукция клеток млекопитающих указанными лентивирусами;
e. отбор наиболее подходящего клона клеток млекопитающих для получения reCG.
Указанная линия рекомбинантных клеток демонстрирует продуцирование reCG, по меньшей мере, 100 МЕ/мл в бессывороточной среде.
В предпочтительном варианте реализации данного изобретения на этапе а. данного способа используются последовательности субъединиц, по существу, сходные с последовательностями α и β - SEQ ID №1 и SEQ ID №2, соответственно. На этапе b. лентивирусные векторы состоят из вектора pLV, содержащего промотор EF-1α. Этап с. включает транзиентную трансфекцию клеток HEK293 плазмидами pREV, pVSVG, pMDL, pLV-reCG α и pLV-reCG β с использованием катионных липидов в качестве носителей. Этап d. включает трансдукцию клеток СНО-K1.
В качестве альтернативы способ по данному изобретению включает выполнение двух последовательных событий трансдукции.
Другая цель данного изобретения представляет собой создание линии клеток млекопитающих, полученной предыдущим способом и содержащей нуклеиновую кислоту, кодирующую гормон рекомбинантный хорионический гонадотропин лошади (reCG), при этом последовательности, кодирующие α- и β-субъединицы reCG, содержат последовательности, по существу, сходные с последовательностями SEQ ID №1 и SEQ ID №2. Предпочтительно указанная линия клеток млекопитающих представляет собой СНО-K1 и демонстрирует продуцирование reCG, по меньшей мере, 100 МЕ/мл.
Другая цель данного изобретения представляет собой создание способа получения гормона рекомбинантного хорионического гонадотропина лошади (reCG), который включает следующие этапы:
a. культивирование указанной линии клеток млекопитающих в биореакторе в среде, не содержащей фетальную телячью сыворотку, для крупномасштабного производства указанного reCG;
b. сбор супернатанта; и
c. очистка.
Данный способ производства обеспечивает продуктивность reCG, по меньшей мере, 100 МЕ/мл в бессывороточной среде.
В предпочтительном варианте реализации данного изобретения указанный этап а. включает культивирование в бессывороточной среде, содержащей 50% коммерческой среды Ехсе1302 и 50% фосфатно-солевого буфера. На указанном этапе с. очистка включает псевдоаффинную хроматографию с использованием красителя. Предпочтительно в указанной псевдоаффинной хроматографии с использованием красителя используется матрица CaptoBlue-Sepharose. Альтернативно указанный этап очистки с. включает тангенциальную проточную фильтрацию и последующее концентрирование reCG.
Необязательно указанный способ очистки также включает этап очистки с помощью ВЭЖХ с использованием колонки С4.
reCG, полученный указанным способом производства, обладает удельной активностью (в единицах эффективности in vivo по отношению к массе белка, определенной с помощью ИФА), по меньшей мере, 6000 МЕ/мг.
Другая цель данного изобретения включает получение нуклеиновой кислоты, кодирующей альфа-субъединицу reCG, которую можно получить описанным способом получения, имеющей последовательность, по существу, сходную с последовательностью SEQ ID №1. Объект также содержит нуклеиновую кислоту, кодирующую бета-субъединицу reCG с последовательностью, по существу, сходную с последовательностью SEQ ID №2.
Другая цель данного изобретения представляет собой получение гормона reCG, получаемого описанным способом производства, который имеет профиль гликозилирования с, по меньшей мере, 3% нейтральных структур и, по меньшей мере, 3% тетрасиалированных структур. Предпочтительно указанный reCG имеет профиль гликозилирования с, по меньшей мере, 3% нейтральных структур, от 26 до 30% моносиалированных структур, от 50 до 55% бисиалированных структур, от 8 до 15% трисиалированных структур и, по меньшей мере, 3% тетрасиалированных структур.
Еще одна цель данного изобретения представляет собой получение фармацевтического состава, содержащего терапевтически эффективное количество описанного в данном документе reCG. В предпочтительной форме реализации данного изобретения указанный состав является жидким и хранится охлажденным, предпочтительно при 5°С, без необходимости замораживания для коммерческого использования. В альтернативной форме реализации данного изобретения указанный состав является лиофилизированным. В предпочтительной форме реализации данного изобретения указанный состав дополнительно содержит сахар, консервант, антиоксидант, маннит и антиагрегационный агент. Предпочтительно указанный состав содержит дигидрат цитрата натрия, моногидрат лимонной кислоты, аргинин, сахарозу, маннит, L-метионин, полоксамер 188, m-крезол и воду.
Другая цель данного изобретения представляет собой разработку способа индукции овуляции у животных, включающий введение, по меньшей мере, 140 МЕ/животное reCG. При этом введение указанной дозы вызывает овуляцию через 48 часов после ее применения.
ПОДРОБНОЕ ОПИСАНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
В данном изобретении описан способ получения линии (клона) клеток млекопитающих, экспрессирующих рекомбинантный хорионический гонадотропин лошади (reCG), включающий следующие этапы:
a. получение кодирующих последовательностей α- и β-субъединиц reCG, оптимизированных для их экспрессии в клетках млекопитающих;
b. введение указанных кодирующих последовательностей в лентивирусные векторы экспрессии с. получение лентивирусов, содержащих кодирующую reCG последовательность;
d. трансдукция клеток млекопитающих указанными лентивирусами;
e. отбор наиболее подходящего клона клеток млекопитающих для получения reCG.
Клоны клеток, демонстрирующие продуцирование, по меньшей мере, 100 МЕ/мл в бессывороточной среде в биореакторах при крупномасштабном производстве, получают способом, описанным в данном изобретении.
Одной из основных характеристик способа получения линии клеток млекопитающих, продуцирующих reCG, является то, что в нем используется инновационная и оптимизированная последовательность ДНК, кодирующая reCG. Последовательность была модифицирована и оптимизирована для экспрессии в клетках СНО млекопитающих. Предпочтительно клетках СНО-К1. reCG содержит, по меньшей мере, одну альфа-субъединицу и одну бета-субъединицу, поэтому кодирующие последовательности ДНК для обеих субъединиц были оптимизированы. Указанные кодирующие последовательности для каждой субъединицы, по существу, аналогичны или идентичны последовательностям SEQ ID №1 (α) и SEQ ID №2 (β).
Данные кодирующие последовательности для каждой субъединицы содержат небольшие модификации их нуклеотидов, которые делают их оптимальными для транскрипционных, трансляционных и посттрансляционных механизмов в клетках СНО-K1. После оптимизации было замечено, что рекомбинантная последовательность бета-субъединицы eCG демонстрирует гомологию 82,2% с неоптимизированной последовательностью бета-субъединицы («природной» бета-субъединицей eCG), тогда как в отношении других рекомбинантных последовательностей, опубликованных в различных патентах, гомология была ниже (78%). Было замечено, что те нуклеотидные положения нашей оптимизированной рекомбинантной последовательности, которые отличались как от остатков нативной последовательности, так и от остатков оптимизированных последовательностей, опубликованных в других патентах, соответствовали 4% от общего количества. Этого процента было достаточно, чтобы действовать как определяющий фактор в получении более высоких уровней экспрессии reCG по сравнению с теми, о которых сообщалось в данном уровне техники.
Способ получения линии клеток млекопитающих, продуцирующих reCG, по данному изобретению обеспечивает получение стабильных клеточных линий, продуцирующих reCG, путем использования лентивирусных векторов третьего поколения в качестве инструмента для переноса генетического материала. Используемый лентивирусный вектор представляет собой pLV, содержащий промотор EF-1α. Векторы, несущие каждую кодирующую последовательность для каждой субъединицы, называются pLV-reCGα и pLV-reCGβ. Кроме того, указанный pLV содержит кодирующую область гена устойчивости к пуромицину в качестве маркера селекции.
Для получения лентивирусных частиц в данном изобретении описаны транзиентные трансфекции клеток HEK293 плазмидами pREV, pVSVG, pMDL, pLV-reCGα и pLV-reCGβ с использованием катионных липидов в качестве носителей. После получения лентивирусных частиц их используют для трансдукции клеток млекопитающих (этап d). Предпочтительно указанные клетки млекопитающих представляют собой клетки СНО-K1. Предпочтительно способ включает две последовательные трансдукции.
Другая цель данного изобретения включает получение линии клеток млекопитающих, часть генома которых содержит нуклеиновую кислоту, кодирующую гормон рекомбинантный хорионический гонадотропный лошади (reCG), при этом последовательности, кодирующие альфа- и бета-субъединицы указанного reCG, содержат последовательности, по существу, сходные с последовательностями SEQ ID №1 и SEQ ID №2. Данные клетки получают способом получения reCG-продуцирующей клеточной линии млекопитающих по данному изобретению. Предпочтительно линия клеток представляет собой СНО-K1. Данная линия (клон) клеток продуцирует, по меньшей мере, 100 ME reCG/мл в бессывороточной среде.
Другая цель данного изобретения представляет собой создание способа получения гормона рекомбинантного хорионического гонадотропина лошади (reCG), отличающегося тем, что он включает следующие этапы:
d. культивирование указанной линии (клона) клеток млекопитающих в биореакторе в бессывороточной среде для крупномасштабного производства указанного reCG;
e. сбор супернатанта; и
f. очистка.
Способ получения рекомбинантного хорионического гонадотропина лошади (reCG) по данному изобретению обеспечивает получение, по меньшей мере, 100 МЕ/мл в бессывороточной среде. Такой высокий уровень производства обусловлен несколькими факторами. Во-первых, высокая продуктивность reCG обусловлена линией клеток, трансформированных последовательностями SEQ ID №1 и SEQ ID №2 по данному изобретению, которая была дополнительно адаптирована и оптимизирована для культивирования в среде, не содержащей бычью сыворотку. Среда, используемая для получения, представляет собой среду МС02, которая содержит 50% коммерческой среды Excel302 и 50% комбинации солей, аминокислот, углеводов и т.д., как описано в Таблице 2.
Еще одним важным фактором является этап очистки. Уровень техники описывает сложные и дорогие процессы очистки, которые делают конечный продукт более дорогим. Способ получения рекомбинантного хорионического гонадотропина лошади (reCG) по данному изобретению включает этап очистки, включающий псевдоаффинную хроматографию с использованием красителя. В предпочтительном варианте реализации данного изобретения матрица, используемая для проведения указанной хроматографии, представляет собой матрицу CaptoBlue-Sepharose. Необязательно можно добавить дополнительный этап очистки ВЭЖХ с использованием колонки С4.
В качестве альтернативы очистке с помощью псевдоаффинной хроматографии с использованием красителя указанный этап очистки включает этап тангенциальной проточной фильтрации и последующий этап концентрирования reCG.
С помощью способа получения гормона рекомбинантного хорионического гонадотропного лошади (reCG) по данному изобретению можно получить reCG с удельной активностью (в единицах эффективности in vivo по отношению к массе белка, определенной с помощью ИФА), по меньшей мере, 6000 МЕ/мг.
Другая цель данного изобретения включает получение нуклеиновой кислоты, кодирующей альфа-субъединицу reCG, которую можно получить способами, описанными в данном изобретении. Указанная альфа-субъединица указанного reCG содержит последовательность, по существу, сходную с последовательностью SEQ ID №1.
Другая цель данного изобретения включает получение нуклеиновой кислоты, кодирующей бета-субъединицу reCG, которую можно получить способами, описанными в данном изобретении. Указанная бета-субъединица указанного reCG содержит последовательность, по существу, сходную с последовательностью SEQ ID №2.
В предпочтительном варианте реализации данного изобретения указанная последовательность, по существу, сходная с последовательностью SEQ ID №1, означает, что указанная последовательность, по меньшей мере, на 90% идентична последовательности SEQ ID №1.
В предпочтительном варианте реализации данного изобретения указанная последовательность, по существу, сходная с последовательностью SEQ ID №1, означает, что указанная последовательность, по меньшей мере, на 95% идентична последовательности SEQ ID №1.
В предпочтительном варианте реализации данного изобретения указанная последовательность, по существу, сходная с последовательностью SEQ ID №1, означает, что указанная последовательность, по меньшей мере, на 98% идентична последовательности SEQ ID №1.
В предпочтительном варианте реализации данного изобретения указанная последовательность, по существу, сходная с последовательностью SEQ ID №2, означает, что указанная последовательность, по меньшей мере, на 90% идентична последовательности SEQ ID №2.
В предпочтительном варианте реализации данного изобретения указанная последовательность, по существу, сходная с последовательностью SEQ ID №2, означает, что указанная последовательность, по меньшей мере, на 95% идентична последовательности SEQ ID №2.
В предпочтительном варианте реализации данного изобретения указанная последовательность, по существу, сходная с последовательностью SEQ ID №2, означает, что указанная последовательность, по меньшей мере, на 98% идентична последовательности SEQ ID №2.
Процентная идентичность рассчитывается путем деления количества совпавших частей в окне сравнения на общее количество позиций в окне сравнения и умножения на 100. Идентификацию выполняют с использованием алгоритмов BLAST и BLAST 2.0 (см., например, Altschul et al., 1990, J. Mol. Biol. 215:403-410 and Altschul et al., 1997, Nucleic Acids Res. 25(17):3389-3402).
Другая цель данного изобретения включает получение гормона reCG, получаемого способом получения гормона рекомбинантного хорионического гонадотропина лошади (reCG) по данному изобретению. Указанный гормон reCG имеет профиль гликозилирования с, по меньшей мере, 3% нейтральных структур и, по меньшей мере, 3% тетрасиалированных структур. Предпочтительно указанный гормон имеет профиль гликозилирования с, по меньшей мере, 3% нейтральных структур, от 26 до 30% моносиалированных структур, от 50 до 55% бисиалированных структур, от 8 до 15% трисиалированных структур и, по меньшей мере, 3% тетрасиалированных структур.
Другая цель данного изобретения включает получение фармацевтического состава, характеризующегося тем, что оно включает терапевтически эффективное количество reCG по данному изобретению. В одном варианте реализации данного изобретения указанный состав является лиофилизированным. В другом варианте реализации данного изобретения указанный фармацевтический состав является жидким. Фармацевтический состав по данному изобретению дополнительно содержит сахар, консервант, антиоксидант, маннит и антиагрегационный агент. Указанный жидкий фармацевтический состав содержит дигидрат цитрата натрия, моногидрат лимонной кислоты, аргинин, сахарозу, маннит, L-метионин, полоксамер 188, m-крезол и воду.
Другая цель данного изобретения включает разработку способа индукции овуляции у животных, включающего введение, по меньшей мере, 140 МЕ/животное reCG, получаемого с помощью способа получения гормона рекомбинантного хорионического гонадотропина лошади (reCG) по данному изобретению. Данный способ позволяет вызвать овуляцию через 48 часов после его применения.
Данное изобретение для получения reCG устраняет все недостатки, связанные с использованием гормона, полученного из крови жеребых кобыл (PMSG), а также недостатки, представленные всеми рекомбинантными вариантами, описанными до сих пор, ни один из которых еще не вышел на ветеринарный рынок:
- способ получения reCG, описанный в данном изобретении, заменяет использование животных для получения гормона, исключая жестокие практики, которым они подвергаются, что полностью противоречит биоэтическим стандартам и стандартам безопасности, которых требует общество;
- reCG, полученный согласно данному изобретению и описанный в данном изобретении, представляет собой продукт более высокого качества, полученный в результате культивирования клеток животных в биореакторах в бессывороточной среде, что позволяет стандартизировать процессы производства. Таким образом, как параметры культивирования, так и процедуры очистки являются легко контролируемыми и воспроизводимыми (в отличие от использования животного-хозяина), что приводит к получению продукта (reCG) с большей сопоставимостью между сериями, свободного от загрязняющих веществ из плазмы животных и, следовательно, более безопасного с санитарной точки зрения;
- способ, описанный в данном изобретении, позволяет получить гормон reCG, проявляющий FSH/LH-активность in vivo, в отличие от остальных описанных в литературе технологий, использующих другие типы клеточных хозяев, не позволяющих получить рекомбинантный гормон, проявляющий активность in vivo;
- для лечения животных требуется меньшее количество Международных Единиц для reCG, полученного и описанного в данном изобретении. Поскольку reCG по данному изобретению более эффективен, для FTAI требуется всего 100 МЕ/животное для Bos indicus и 140 МЕ/животное для Bos taurus вместо 300 ME и 400 ME соответственно, которые требуются при работе с PMSG, полученного из крови жеребых кобыл. Также требуется всего 1000 ME для Bos indicus и 2000 ME для Bos taurus для SOV, вместо 2000 ME и 4000 ME, соответственно, которые необходимы при работе с PMSG, полученным от кобыл;
- после воздействия 140 ME reCG на животное циклическое состояние у скота во время анэструса восстанавливается в 85% случаев, по сравнению с 65% успеха после воздействия PMSG. Это очень важно, так как эти животные с очень низкой упитанностью восстанавливают циклическое состояние только благодаря действию гормона на яичники. Если гормона нет, животное остается в ациклическом состоянии (в анэструсе) и, следовательно, представляет собой непродуктивное животное;
- достигается более эффективная синхронизация животных. Это значительно облегчает их использование в протоколах синхронизированного искусственного осеменения (FTAI) и суперовуляции (SOV). Все коровы, получавшие reCG по данному изобретению согласно FTAI, овулируют через 48 часов после введения, тогда как коровы, получавшие PMSG, овулируют через 50-60 часов после введения. Наличие фиксированного времени для эффективной и целесообразной процедуры осеменения-беременности имеет важное значение для содержания скота, в котором проходят лечение сотни коров. Таким образом, коров, которых подвергли воздействию гормона «утром», можно осеменять в то же время через два дня;
- по сравнению с другими заменителями PMSG, полученными в клетках животных, например, СНО DG44, которые, по-видимому, демонстрируют подходящие профили гликозилирования, данное изобретение позволяет получить reCG с самой высокой продуктивностью, описанной на сегодняшний день. Таким образом, данное изобретение преодолевает основной ограничивающий недостаток, препятствовавший появлению на рынке рекомбинантной версии гормона: в данном изобретении предложена первая технология, способная генерировать большие количества reCG с биологической активностью при низких затратах. Высокая производительность вместе с использованием недорогой бессывороточной культуральной среды способствует снижению общей стоимости способа по данному изобретению. reCG, полученный как с использованием высокоэффективного одноэтапного способа очистки, так и в результате однократной тангенциальной проточной фильтрации и процедуры концентрирования исходного образца, демонстрирует подходящие профили гликозилирования и, следовательно, проявляет биологическую активность in vivo на крысах, коровах и свиньях;
- получен жидкий состав reCG, который является стабильным во времени и, таким образом, превосходит существующие на рынке продукты, состоящие из лиофилизированных или замороженных жидких составов, со всеми вытекающими отсюда недостатками.
Далее будут описаны примеры анализов, выполненных для исполнения и реализации каждой цели данного изобретения. Следует отметить, что данные примеры приведены в качестве иллюстрации без намерения ограничить спектр охраны прав данного изобретения.
ПРИМЕРЫ
Пример 1 - Разработка и оптимизация кодирующей последовательности
Технология, разработанная в данном изобретении, сначала включала разработку линий клеток млекопитающих, в частности, суспензионных клеток СНО-K1 (клеток яичника китайского хомячка), которые продуцируют рекомбинантный хорионический гонадотропин лошади (reCG). Кодирующие последовательности α- и β-субъединиц reCG (reCGα и reCGβ) были оптимизированы для экспрессии в клетках СНО-K1, чтобы получить высокие уровни мРНК и, таким образом, максимизировать экспрессию кодируемого белка. Оптимизация генов использует преимущества вырожденности генетического кода, благодаря чему белок может кодироваться различными альтернативными генетическими последовательностями. Поскольку использование кодонов различается в каждом организме, это может создавать проблемы при экспрессии рекомбинантных белков в гетерологичных хозяевах, что приводит к очень низкой экспрессии. Таким образом, алгоритмы оптимизации генов позволяют проводить многопараметрическую оптимизацию последовательностей ДНК, охватывающую несколько аспектов экспрессии генов: транскрипцию, сплайсинг, трансляцию и деградацию мРНК, для достижения наиболее эффективной экспрессии данного белка.
После оптимизации было обнаружено, что рекомбинантная последовательность eCG-бета продемонстрировала 82,2% гомологии с последовательностью неоптимизированной бета-субъединицы ("природная" eCG-бета), в то время как с другими рекомбинантными последовательностями, упомянутыми в разделе «Уровень техники", гомология была ниже (78%). Было обнаружено, что те нуклеотидные положения кодирующих последовательностей в оптимизированном рекомбинантном гормоне по данному изобретению, которые отличались как от остатков нативной последовательности, так и от остатков оптимизированных последовательностей, опубликованных в заявке, соответствовали 4% от общего количества, этого было достаточно, чтобы стать определяющим фактором в получении более высоких уровней экспрессии reCG по сравнению с теми, о которых сообщалось в других патентах.
Синтетические последовательности были получены в виде ДНК и затем клонированы в векторы p-alpha_eCG (AmpR) и p-beta_eCG (AmpR), соответственно. Эти векторы содержат бактериальную точку начала репликации (ориджин Col E1), которая позволяет амплифицировать плазмиду в Е. coli и ген устойчивости к антибиотику ампициллину (AmpR). Вектор p-eCGα имеет длину 3000 п.н. и содержит кодирующую последовательность eCGα длиной 360 п.н., при этом первые 72 п.н. кодируют природный сигнальный пептид eCGα. Вектор p-eCGβ имеет длину 3200 п.н. и содержит кодирующую последовательность eCGβ длиной 507 п.н., содержащую сигнальный пептид длиной 60 п.н. в начале этой последовательности.
Пример 2 - Конструирование векторов экспрессии
Для получения стабильных клеточных линий, продуцирующих reCG, в качестве способа доставки генетического материала были сконструированы лентивирусные векторы третьего поколения. Для этого необходимо было сначала клонировать кодирующие последовательности каждой субъединицы в лентивирусных векторах-переносчиках, а затем провести сборку лентивирусных частиц (ЛЧ) и их последующее титрование.
Были сконструированы лентивирусные векторы экспрессии, кодирующие α- и β-субъединицы reCG. Таким образом, вектор, содержащий a-субъединицу, расщепляли ферментами Xbal/EcoRV с высвобождением кодирующей последовательности reCGα, которую клонировали в сайты Nhel/EcoRV лентивирусного плазмидного вектора pLVenhCEF. Вектор, содержащий β-субъединицу, расщепляли ферментами BamHI/EcoRV с высвобождением кодирующей последовательности reCGβ, которую клонировали в сайты BamHI/Smal вектора pLVenhCEF. Этот вектор, разработанный в нашей лаборатории, содержит промотор EF-1α в качестве регуляторного элемента экспрессии, характеризующегося высокими уровнями экспрессии в самых разных клетках животных. Он также содержит стимулирующий экспрессию фрагмент, полученный из последовательности энхансера CMV, и кодирующую область для гена устойчивости к пуромицину в качестве маркера селекции. Полученные плазмиды размножали с помощью прокариотических клеток (Е. coli), культивировали в условиях встряхивания и очищали путем экстракции органическим растворителем и осаждения. Секвенирование отобранных клонов E. coli подтвердило идентичность фрагментов ДНК, клонированных в плазмиды pL VenhCEF-reCGα и pL VenhCEF-reCGβ, продемонстрировав 100%-ную гомологию с последовательностями синтетических генов eCGα и eCGβ, соответственно.
Для получения лентивирусных частиц третьего поколения проводили транзиентные трансфекции клеток HEK293 (пакующих клеток) четырьмя плазмидами: pREV, pGlyco-G, pMDL (пакующими плазмидами) и векторами переноса pL VenhCEF-reCGα и pL VenhCEF-reCGβ (кодирующими каждую из субъединиц reCG). Для этого в качестве носителя ДНК использовали катионные липиды. Супернатанты, содержащие лентивирусные частицы, собирали через 30 ч после трансфекции, центрифугировали при 65,000 g для концентрирования и хранили при -80°С до момента использования (Фиг. 1).
Титрование лентивирусных частиц проводили с использованием набора QuickTiter™ Lentivirus Titer Kit (Cell Biolabs Inc.). Этот набор был разработан для обнаружения только капсидного белка р24 ВИЧ-1, связанного с лентивирусом, поэтому свободный белок, который остается в супернатанте, не мешает анализу. Таким образом, для reCGα и reCGβ был получен физический титр 2,7×109 ЛЧ/мл и 2,1×109 ЛЧ/мл, что может быть приблизительно тождественным титру трансдукции 5,0×106 и 3,9×106 TU/мл, соответственно, что приводит к высоким титрам для проведения трансдукции клеток СНО-K1.
Пример 3 - Получение клеточных линий
Полученные лентивирусные частицы использовали для получения линий рекомбинантных клеток СНО-K1, продуцирующих reCG, в суспензии. Всего было выполнено два последовательных события трансдукции (Td1 и Td2), поскольку клетки не выживали до третьего события трансдукции. Линии трансдуцированных клеток подвергали селективному давлению путем инкубации с увеличивающимися концентрациями пуромицина. Эта стратегия позволила обогатить популяцию клетками, устойчивыми к более высоким концентрациям антибиотика, что привело к увеличению продуктивности всей клеточной линии. На Фиг. 2 представлен анализ ДСН-ПААГ с последующим вестерн-блоттингом с использованием специфических поликлональных антител против reCG. Как видно, при одинаковой плотности клеток наибольшая концентрация reCG была получена в супернатанте клеточной линии sCHO Td2 (200) (резистентной к 200 мкг пуромицина).
В последующем оценивали reCG-продуктивность полученных клеточных линий, определяя концентрацию накопленного гормона в супернатанте, а также начальную и конечную плотность клеток через определенный период. Количественную оценку reCG проводили с помощью конкурентного ИФА, разработанного в нашей лаборатории. Анализ включал конкуренцию между иммобилизованным в твердой фазе антигеном (reCG) и тем же антигеном (референтным reCG или неизвестным образцом) в растворе за связывание со специфическими кроличьими антителами против reCG (pAb anti-reCG). Эти антитела ранее были получены в нашей лаборатории. Затем добавляли вторичные антитела, конъюгированные с пероксидазой, для обнаружения оставшихся комплексов, связанных твердой фазой. Продуктивность различных клеточных линий представлена в Таблице 1.
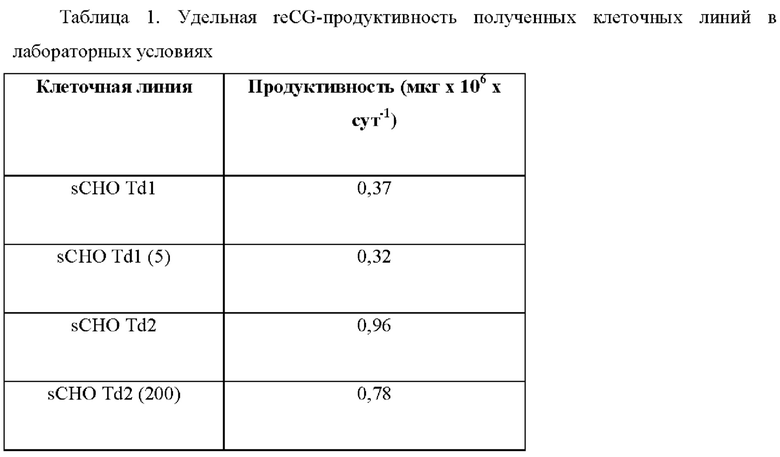
Принимая во внимание данные результаты, для клонирования была выбрана клеточная линия с наивысшим уровнем продуцирования reCG (клеточная линия reCG Td2).
Пример 4 - Выделение клонов
reCG-продуктивность полученных клеточных линий позволила выбрать одну из них, получить клон одиночных клеток с подходящим профилем роста и высокой reCG-продуктивностью.
Чтобы выбрать клоны с самым высоким уровнем экспрессии reCG, более 400 клонов были оценены в первоначальном скрининге с помощью дот-блоттинга с использованием специфических антител против reCG. Отобранные клоны подвергали криоконсервации. После этапа предварительной селекции для уменьшения числа анализируемых клонов «наблюдаемую» продуктивность (определяемую как концентрация reCG, полученная для одной и той же плотности клеток для каждого клона) выбранных клонов оценивали с помощью ДСН-ПААГ с последующим вестерн-блоттингом (Фиг. 3). Данный анализ продемонстрировал, что клоны P5D9 (дорожка 3) и Р5СЗ (дорожка 4) демонстрируют самый высокий уровень экспрессии reCG. В последующем оценивали reCG-продуктивность отобранных клонов, определяя концентрацию накопленного гормона в супернатанте, а также начальную и конечную плотность клеток через определенный период. Концентрацию reCG определяли с помощью конкурентного ИФА, описанного выше. Расчетная продуктивность составила 0,80 и 0,81 пг × 106 клеток-1 × сут.-1 для клонов P5D9 и Р5С3, соответственно. Наконец, был выбран клон Р5С3, так как он демонстрировал лучшие характеристики роста, чем клон P5D9.
Пример 5 - Оптимизация условий культивирования для достижения оптимального гликозилирования
5.1 Культура высокой плотности клона Р5С3 в бессывороточной среде в однолитровом биореакторе
Клон Р5С3 культивировали в питательной среде для культивирования, не содержащей фетальную бычью сыворотку, (МС01) в однолитровом биореакторе в перфузионном режиме в течение 27 дней. Культивирование начинали при плотности клеток 7,8×105 клеток/мл-1, при этом культура демонстрировала экспоненциальный рост без лаг-фазы до достижения максимальной плотности клеток 1,6×107 клеток/мл-1. Жизнеспособность клеток составляла выше 94%. Скорость перфузии, которая была начата на третий день культивирования, варьировали от 0,21 до 1,00 объема реактора в сутки. Концентрация лактата оставалась ниже 1,1 г/л-1 (12,2 мМ) (Фиг. 4). Удельная скорость роста клона составила 0,013 ч-1.
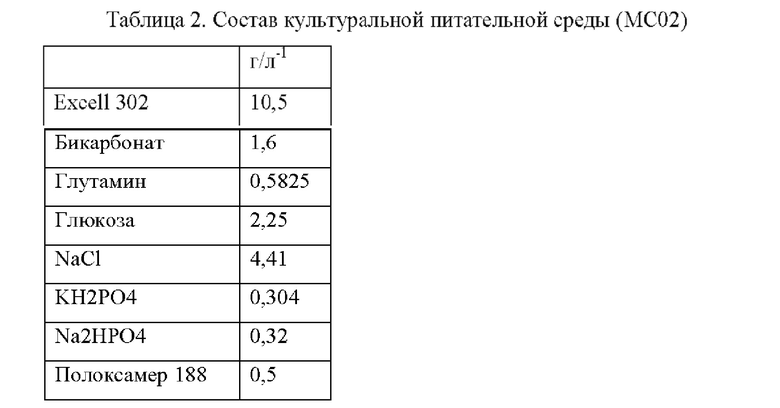
Впоследствии была разработана новая питательная среда (МС02). Важно: состав данной питательной среды является результатом объединения коммерческой питательной среды с солями, аминокислотами, углеводами и т.д. (Таблица 2). Это было сделано для оптимизации стоимости питательной среды, снижая ее стоимость на 50%.
Данный новый состав питательной среды не изменил ни продуктивность клеток, ни какие-либо характеристики молекулы.
5.2 Культура высокой плотности клона Р5С3 в бессывороточной среде в 50-литровом биореакторе
Культура в промышленных масштабах. После культивирования клеток в однолитровом биореакторе выполняли масштабирование до 50-литрового биореактора, воспроизводя параметры культуры клеток, исследованные в лабораторных масштабах. Поскольку скорость перфузии составляла один объем реактора в сутки, в этом процессе ферментации было получено 50 л собранного материала, каждый из которых содержал 157 МЕ/мл reCG. Учитывая, что одна доза reCG по данному изобретению составляет 140 ME и что было собрано 50000 мл супернатанта, содержащего 157 МЕ/мл, всего за 24 часа было получено 7850000 ME. Таким образом, способ производства по данному изобретению предоставляет около 56000 доз reCG в день при промышленном масштабе (50 л). Таким образом, в промышленных масштабах получают более одной дозы/мл собранного материала. Сравнивая данные результаты с текущим способом производства (выделение крови из жеребых кобыл), условия культивирования, разработанные в биореакторе объемом 50 л, соответствуют около 600 кобылам. Другими словами, технология, описанная в данном изобретении, может произвести за 25-30 дней биопроцесса (включая культивирование, очистку, получение состава, упаковку) такое же количество доз, что и 600 жеребых кобыл за 200 дней [при условии, что все кобылы имеют одинаковую концентрацию eCG в их крови, и что eCG, полученный от каждой кобылы, имеет одинаковое качество (что невозможно)].
Кроме того, reCG-продуктивность клона Р5С3 в промышленном масштабе (биореакторе 50 л) рассчитывали при использовании различных культуральных питательных сред разной стоимости (Таблица 3). Данные культуральные питательные среды содержали исходную коммерческую среду (МС01, EX-CELL 302), оптимизированную питательную среду МС02 (описанную ранее) и питательную среду МС05. В этой последней питательной среде была получена еще более высокая продуктивность, что привело к большей разнице по сравнению с результатами, опубликованными другими авторами, а также при меньших затратах.
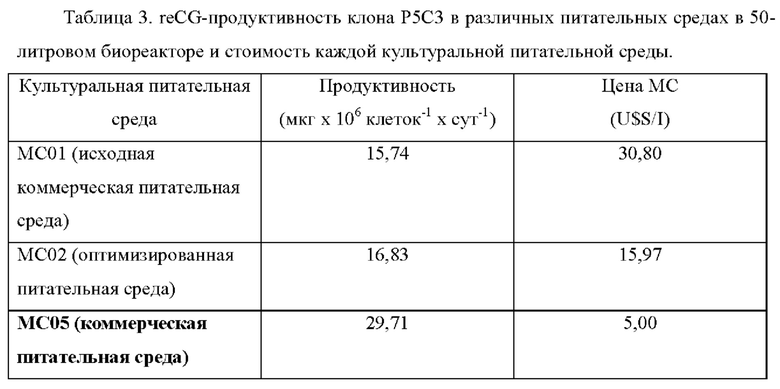
Культуральная питательная среда 01 (МС01): Ex-Cell СНО 302
Культуральная питательная среда 02 (МС02): МР01 / P2G 50/50 (MP 02) оптимизированная питательная среда
Культуральная питательная среда 05 (МС05): Производственная питательная среда CD BHK-21
В патентной заявке WO2017112987 используют систему амплификации генов DHFR-MTX. Они не сообщают о продуктивности, а вместо этого сообщают о кинетике экспрессии reCGα после адаптации к росту в отсутствие МТХ. Из прочтения документа получено приблизительное кумулятивное значение в МЕ/мл reCG для клеточной линии, культивируемой в присутствии фетальной бычьей сыворотки: 10 МЕ/мл (24 ч), 20 МЕ/мл (48 ч) и 28 МЕ/мл (72 ч), и для линии, культивируемой без фетальной бычьей сыворотки, 5 МЕ/мл (24 ч), 10 МЕ/мл (48 ч) и 18 МЕ/мл (72 ч).
По сравнению с этими результатами, клеточные клоны, полученные в данном изобретении, продуцируют 45,6 МЕ/мл (P5D9) и 50,2 МЕ/мл (Р5С3) в небольшом масштабе, в отсутствие фетальной бычьей сыворотки, за 72 ч, в то время как в режиме непрерывной перфузии в биореакторах уровни продуктивности достигали более 15 ME × 106 клеток-1 × сут.-1. Это соответствует 157 МЕ/мл за 24 часа, что представляет собой значительно более высокое значение, чем значения, указанные в патенте WO 2017112987 (они сообщают о 5 МЕ/мл в отсутствие фетальной бычьей сыворотки за 24 часа). Это означает производство более 7000000 ME в день, что составляет более 50000 доз reCG по данному изобретению в день.
Таким образом, технология по данному изобретению позволила получить самые высокие показатели производительности и продуктивности, о которых сообщалось на сегодняшний день, в результате сочетания уникальных факторов нашей технологии: оптимизация последовательностей для экспрессии в клетках СНО-K1 (виды Cricetulus griseus), использование собственных лентивирусных векторов третьего поколения и использование линии клеток СНО-K1, которая экспрессирует широкий спектр гликозилтрансфераз, способных добавлять би-, три- и тетрасиалированные N-гликаны комплексного типа в полипептиды, в дополнение к образованию моно- и бисиалированных О-гликанов муцинового типа, которые являются ключевыми факторами для проявления биоактивности eCG in vivo.
Действительно, эксклюзивный процесс оптимизации последовательности ДНК был эффективен для достижения высокой экспрессии гормона, о чем свидетельствуют 4% нуклеотидов, которые были модифицированы в нативной последовательности ДНК белка, и тот факт, что они не были изменены в синтетических последовательностях, о которых сообщалось в других патентах.
Пример 6 - Очистка
6.1 - Первый этап очистки
После культивирования клеток СНО в биореакторе в бессывороточной среде Excell 302 (Sigma) собранный материал использовали для разработки первого этапа очистки отобранного материала. Таким образом, была выбрана псевдоаффинная хроматография с использованием красителя с применением смолы CaptoBlue-Sepharose, упакованной в колонку XK (GE, Healthcare) и уравновешенной в 20 мМ буфере Трис-HCl рН 7. Осветленный собранный материал без предварительного кондиционирования загружали на смолу при скорости потока 153,06 см/ч и общем времени удерживания пять мин. После этапа промывки тем же уравновешивающим раствором белок элюировали с использованием изократического градиента (буфер Трис-HCl рН 8, 2 М NaCl, 20% (об./об.) этанол). Во время этапов загрузки и промывки этой первой хроматографии просачивания гормона не наблюдалось, таким образом, условия загрузки были подходящими. Интактный гормон был выделен с заметно более высоким уровнем чистоты, чем полученный при частичной очистке сыворотки жеребых кобыл (PMSG). Таким образом, этап отбора reCG из супернатанта клеточной культуры был оптимизирован с использованием смолы для псевдоаффинной хроматографии с применением красителя. Был достигнут высокий выход без какой-либо потери белка, так как степень извлечения составила 98% (оценка как с помощью ИФА, так и с помощью ОФ-ВЭЖХ).
6.2 - Второй этап очистки
В качестве второго этапа очистки была выбрана хроматография гидрофобного взаимодействия, поскольку частично очищенный гормон с первого этапа очистки отобранного материала (названный post-Blue) элюировали в условиях высокой ионной силы (2 М NaCl). Для уменьшения числа отдельных операций и стоимости процесса глобальной очистки были предложены следующие стратегии: 1) загрузка «неочищенного элюата», т.е. фракции post-Blue без предварительного кондиционирования, исключая этап диафильтрации (поскольку эта фракция находится в состоянии с высокой ионной силой); 2) скрининг двух типов гидрофобных лигандов, доступных в нашей лаборатории: Фенила и Бутила; 3) оценка элюата, диафильтрованного против лимоннокислого/ цитратного буфера с рН 6,0, поскольку это условия, в которых получают АФИ post-Blue (названный FD1 RECG), и 4) оценка эффективности очистки с использованием различных солей: во-первых, NaCl (поскольку он присутствует в буфере post-Blue), затем Na2SO4 и, наконец, ((NH4)2SO4 (поскольку эта соль с самым высоким гидрофобным эффектом). С учетом эффективности очистки (степени извлечения и чистоты) всех стратегий наилучшие условия были следующими: DF1 REG загружали на смолу Butyl Sepharose 4FF при скорости потока 15 см/ч и общем времени удерживания три мин. Для улучшения гидрофобного взаимодействия между белком и лигандом смолу уравновешивали 50 мМ лимоннокислым/нитратным буфером, рН 6,0, 2 М (NH4)2SO4, и образец (DF1 REG) кондиционировали с использованием того же уравновешивающего буфера. После этого были выполнены два этапа промывки: первый с использованием того же буфера, что и на этапе уравновешивания, и второй с использованием буфера с более низкой ионной силой (50 мМ лимоннокислого/нитратного буфера, рН 6,0, 1,5 М (NH4)2SO4), чтобы удалить примеси. Наконец, проводили изократическую элюцию с использованием 50 мМ лимоннокислого/цитратного буфера, рН 6,0, 0,5 М (NH4)2SO4.
Пример 7 - Состав
7.1 - Жидкий состав конечного продукта
7.1.1 - Разработка жидкого состава reCG с использованием инструментов QbD
Благодаря данному изобретению был разработан жидкий состав reCG, который позволил получить стабильный и, таким образом, активный гормон в жидкой форме. Таким образом, можно было бы избежать процедуры лиофилизации, которая представляет собой более истощающую и более дорогостоящую операцию. Следовательно, получение жидкого состава вместо лиофилизированного гарантирует не только более низкие затраты, но и сокращение производственного цикла, позволяя избежать этапа восстановления лиофилизированного продукта. Кроме того, в случае работы с многодозовой формой выпуска необходимое количество состава можно разделить на части, обеспечивая его длительную стабильность при применении.
7.1.1.1 - Анализы предварительного состава - установление значимых факторов, влияющих на стабильность reCG в жидком составе
Для исследований термической форсированной деградации reCG, очищенного с помощью хроматографии на CaptoBlue-Sepharose, температуру изменяли в диапазоне от 20 до 70°C с использованием диапазона рН от 3,0 до 8,0. Образцы нагревали в течение десяти минут при каждом условии в амплификаторе и хранили при - 70°С до проведения анализа. Затем аликвоту reCG при в каждом условии оценивали в невосстанавливающем ДСН-ПААГ с последующим окрашиванием кумасси бриллиантовым синим для визуализации степени диссоциации reCG. Различия в профилях подвижности в ДСН-ПААГ указывают на то, что рН оказывает влияние на стабильность гетеродимера reCG. Образцы, соответствующие низкому диапазону рН (рН от 3,0 до 5,0), инкубированные при высоких температурах, демонстрировали другой паттерн фрагментов по сравнению с образцами, соответствующими более щелочному диапазону рН (рН от 6,0 до 8,0). Параллельно оценивали профиль излучения (флуоресценции) (у возбуждение: 274 нм) для анализа возможных конформационных изменений. Наблюдался сдвиг в длинноволновую область в тех образцах, которые подвергались воздействию более высоких температур и более низких значений рН. Сдвиг в длинноволновую область может быть связан с более высокой денатурацией белка или потерей нативной конформации. С учетом этих результатов и с целью уменьшения процессов химической деградации (главным образом, дезаминирования и окисления), оказывающих меньшее влияние при нейтральном рН, был установлен оптимальный диапазон рН от 5 до 7. Точно так же, поскольку pi (изоэлектрическая точка) белка была близка к 3,5-5,5 (согласно анализу ИЭФ), работа в диапазоне рН от 5 до 7 гарантирует, что белок будет демонстрировать отрицательный суммарный заряд, уменьшая случаи физической деградации в виде агрегации (в отличие от того, что может произойти при значениях рН, близких к pi reCG).
7.1.1.2 - Анализ состава
Одной из основных задач в процессе производства биотерапевтических белков является получение состава, гарантирующего высокое качество и стабильность белка. Комбинируя дизайн экспериментов (DoE) с простыми аналитическими способами и ускоренными анализами стабильности, был получен жидкий состав, позволяющий поддерживать 98% биологической активности reCG (интактного, активного reCG) до шести месяцев в ускоренных условиях (25°С, относительная влажность (ОВ) 60%) (CAMEVET, 2012).
Дизайн Placket-Burman design (PBD) использовали для определения влияния нескольких факторов на стабильность reCG в ускоренных условиях (25°С, ОВ 60%, семь дней). Было проведено двенадцать экспериментов с трехкратной повторностью в центральной точке для изучения стандартного отклонения эффектов в N=15 (количество экспериментов). Оценивали влияние восьми реальных факторов и трех фиктивных переменных (виртуальных переменных) на стабильность reCG. Анализируемыми факторами были: количество стабилизаторов (сахарозы, маннита, Arg, L-met) и поверхностно-активного вещества (полоксамера 188), молярность и рН буфера, концентрация АФИ (доза reCG). Отклик на воздействие заключался в анализе количества reCG (%) после семи дней хранения при 25°С и ОВ 60% путем определения площади под кривой интактного reCG, оцениваемой способом ОФ-ВЭЖХ (tR: 13,58 мин).
Диаграмма Парето использовалась для определения факторов, влияющих на характеристики. После этого был применен тест ANOVA для изучения влияния факторов на ответ и для подтверждения значимого влияния факторов. Полученная модель удовлетворяла предположениям о нормальности, гомоскедастичности и независимости переменных. Анализ данных продемонстрировал, что значимыми факторами, влияющими на стабильность reCG в ускоренных условиях, были молярность буфера (р: 0,0013), количество L-met (р: 0,0171), сахарозы (р: 0,0044) и поверхностно-активного вещества (р: 0,0097). Кроме того, R-квадрат (0,8929) и скорректированный R-квадрат (0,8393) указывали на хорошую связь между экспериментальными данными и подобранными данными.
На этапе оптимизации жидкого состава reCG был выполнен четырехфакторный (молярность буфера, количество сахарозы, L-met и Плюроник F-68) пятиуровневое центральное композиционное планирование (CCD). Было проведено двадцать семь пробегов, и ответ снова заключался в анализе количества reCG (%) через 0, 6, 12, 60, 90, 125 и 150 дней хранения при 25°С, 60% ОВ и 40°С, ОВ 75% с помощью способа ОФ-ВЭЖХ. Анализируемыми определяющими факторами были молярность буфера, количество сахарозы, поверхностно-активного вещества и антиоксиданта. Факторы, которые, как было установлено, не оказывают существенного влияния на стабильность reCG, поддерживали на постоянном уровне концентрации во всех исследованиях состава.
Через 90 дней наблюдались значимые различия между составами. Таким образом, стабильность reCG в 27 составах соответствовала квадратичной модели.
Полученная иерархическая модель удовлетворяла предположениям о нормальности, гомоскедастичности и независимости переменных. Кроме того, скорректированный R-квадрат (0,7238) указывает на достаточную связь между экспериментальными данными и подобранными данными: Rскор.: 0,8122; R2скop.: 0,7238; CV% 1,53, отсутствие согласия: 0,1107 (р<0,05 является значимым).
Было получено надежное поле проектных параметров и выбраны оптимальные условия состава, включающие 70 мМ лимоннокислого/цитратного буфера (рН 6,0), 161 мМ сахарозы, 1,0 мг/мл L-met и 1,0 мг/мл поверхностно-активного вещества (плюс 5 мМ L-Arg и 5 мг/мл маннита).
Данный теоретически оцененный жидкий состав reCG был проверен путем изготовления трех отдельных партий препарата и оценки стабильности после 0, 15, 45 и 90 дней хранения при 4°С, 25°С, 60% ОВ и 40°С, 75% ОВ. Через 90 дней интактное количество reCG (оцененное как аис с помощью ОФ-ВЭЖХ) 125±3, 125±1 и 94,6±0,2% при 4°С, 25°С/60% ОВ и 40°С/75% ОВ, соответственно, было получено, демонстрируя надежность разработанного жидкого состава. Валидированный жидкий состав анализировали до 150 дней, получив 98±13% интактного reCG в ускоренных условиях (25°С/60% ОВ).
7.2 - Состав лиофилизированного препарата
Стабильный раствор или состав представляет собой такой раствор, который гарантирует, что степень деградации, изменения, агрегации или потери биологической активности является приемлемой или управляемой. В идеале состав должен сохранять, по меньшей мере, 80% исходной активности белка в течение периода хранения в течение шести месяцев при 2-8°С (US 7,740,884 В2). Таким образом, был разработан состав твердого эксципиента, содержащий соли в качестве буферных агентов, такие как соль лимонной кислоты/цитрат, с низкой молярностью (10 мМ) и рН 6,5.
Кроме того, успех лиофилизации в твердом состоянии зависит от баланса двух конкурирующих требований: образование твердого осадка, который не разлагается при первичной сушке, и существование аморфного состояния, обеспечивающего взаимодействие между эксципиентами и белком (Jhonson et. al., 2001). Эксципиенты, которые действуют как стабилизаторы белка, такие как сахароза или трегалоза, ведут себя как аморфные твердые вещества, тогда как эксципиенты, такие как маннит, действуют как кристаллические твердые вещества, которые уменьшают разложение осадка. Таким образом, в данном изобретении использовали соотношение маннит:сахароза 4:1 при концентрациях 40 г/л и 10 г/л, соответственно.
Белки, такие как гликопротеиновые гормоны, подвержены окислению; поэтому желательно использование соединений с антиоксидантными свойствами, таких как некоторые аминокислоты, такие как метионин, хелатирующие агенты (например, ЭДТА) или бисульфит натрия. Кроме того, для предотвращения адсорбции reCG на поверхности флакона и уменьшения взаимодействия белков на границе воздух-вода использовали неионогенное поверхностно-активное вещество (Плюроник F68 или Полоксамер Р188).
В данном изобретении использовали 0,1 мг/мл и 0,25 мг/мл метионина и Полоксамера Р188 соответственно.
В идеале количество reCG в восстановленном осадке должно быть близко к 1500 МЕ/мл, при этом конечный объем должен быть таким же, как и первоначальный (3 мл).
Процесс включал смешивание эксципиентов состава с АФИ, фильтрацию с использованием 0,2-мм фильтров ПЭС (полиэфирсульфон), заполнение соответствующим образом промытых и стерилизованных боросиликатных флаконов (закрытых резиновыми пробками) и лиофилизации в соответствии с уровнем техники.
Пример 8 - Биохимическая и физико-химическая характеризация
8.1 Способ исследования
reCG получали путем культивирования суспензионного клона Р5С3 в однолитровом биореакторе в режиме перфузии (Biostat Q Plus, Sartorius) в бессывороточной среде. Затем осветленный супернатант (Sartobran-P 0,45 мкм, Sartorius) очищали с использованием хроматографии CaptoBlue-Sepharose в качестве этапа захвата. После этого для получения аликвот белка с более высокой степенью чистоты оценивали два альтернативных этапа очистки:
a) Обращенно-фазовая высоко эффективная жидкостная хроматография (ОФ-ВЭЖХ);
b) Хроматография гидрофобного взаимодействия (ХГВ).
Молекула reCG, очищенная с помощью ОФ-ВЭЖХ, была названа reCG ОФ-ВЭЖХ, тогда как молекула reCG, очищенная с помощью ХГВ, была названа reCG ХГВ.
Коммерческие препараты eCG от Foli-G, Zoovet SA (Аргентина) и Novormon, Syntex (Аргентина) приобретали в региональных ветеринарных аптеках и использовали в качестве внутренних референтных стандартов. Коды А и В присвоены Foli-G и Novormon, соответственно.
8.1.1 - ОФ-ВЭЖХ (RP-HPLC) для структурного анализа. ОФ-ВЭЖХ для оценки биологической активности
Качественные и количественные исследования проводили на колонке С4 с градиентным элюированием и УФ-детектированием (210 нм).
Была продемонстрирована хорошая корреляция между анализом эффективности у крыс и интактным reCG, измеренным как AUC с помощью ОФ-ВЭЖХ, с применением теста EJCR (эллиптической совместной доверительной области). В данном исследовании применялись тест EJCR и способ билинейной регрессии наименьших квадратов (BLS). Если бы идеальная точка (1,0) была включена в эллипс, можно было бы предположить, что способ точен. Данная эллиптическая область описывается математическими уравнениями, которые изображаются в двумерных графиках. Размер эллипса связан с другими аналитическими параметрами, такими как точность анализа.
8.1.2 - ДСН-ПААГ (SDS-PAGE)
Во время проведения всего данного анализа чистоту и наблюдаемую молекулярную массу анализировали в невосстанавливающих условиях. Выполняли колориметрическую детекцию (Кумасси бриллиантовый синий) или иммунохимическую детекцию (вестерн-блоттинг). Для вестерн-блоттинга использовали кроличью поликлональную сыворотку против reCG, полученную в нашей лаборатории.
8.1.3 - Изоэлектрофокусирование (ИЭФ, IEF)
Чтобы разделить изоформы белковых вариантов, выполняли ИЭФ с использованием оборудования Pharmacia®, состоящего из резервуара для электрофореза (Multiphor II), охлаждающей бани (Multitemp III) и источника напряжения (EPS3500XL). Диапазон рН был установлен с использованием 75% (масса/объем) 3-5 амфолитов и 25% (масса/объем) 5-7 амфолитов (GE Healthcare). Детекцию проводили с помощью окрашивания коллоидным кумасси синим или вестерн-блоттинга.
8.1.4 - Эксклюзионная хроматография (ЭХ)-ВЭЖХ ((SEC)-HPLC)
Чистоту и идентичность вариантов reCG и PMSG определяли с помощью эксклюзионной хроматографии (ЭХ)-ВЭЖХ, проведенной на TSKgel G3000SW с размером частиц 10 мкм и УФ-детектированием.
8.1.5 - Спектрофлуориметрический анализ
Спектрофлуориметрические измерения проводились с использованием люминесцентного спектрометра Perkin-Elmer LS-55, оснащенного ксеноновой газоразрядной лампой, монохроматорами типа Монка-Гиллисона и стробируемым фотоумножителем, подключенным к ПК AMD Sempron с использованием Windows Хр.
8.1.6 - Высокоэффективная анионообменная хроматография с импульсным амперометрическим детектированием (ВЭАОХ-ИАД, HPAEC-PAD)
Содержание сиаловой кислоты определяли кислотным гидролизом образцов с последующей высокоэффективной анионообменной хроматографии с импульсным амперометрическим детектированием (ВЭАОХ-ИАД) с использованием системы DIONEX ICS-5000, оснащенной колонкой CarboPac™ РА20 (Thermo Fisher Scientific Dionex). Стандарт N-ацетилнейраминовой (Neu5Ac) кислоты (Calbiochem, Франция) использовали в качестве референтного сравнения.
Кроме того, тип и количество нейтральных моносахаридов, присутствующих в очищенных гликанах reCG и PMSG, определяли кислотным гидролизом образцов с последующей ВЭАОХ-ИАД с использованием системы DIONEX ICS-5000, оснащенной колонкой CarboPacTM РА20. Стандартные растворы смеси моносахаридов (СМ-Mono-Mix-10, Людгер, Великобритания) подвергали такому же воздействию как и растворы образцов, и использовали для идентификации и количественного определения пиков, полученных от образцов гликопротеинов.
8.1.7 - Анализ N-гликанов
8.1.7.1 - Ферментативное N-дегликозилирование в денатурирующих условиях
Для удаления N-гликанов из очищенных образцов проводили ферментативное расщепление в денатурирующих условиях с использованием набора PNGAse F (Biolabs Inc.).
8.1.7.2 - Слабая анионообменная хроматография (WAX) для анализа заряженных меченых N-гликанов
Высвобожденные N-гликаны очищали путем осаждения этанолом и метили флуорофором 2-АВ. Наконец, для анализа относительного количества нейтральных, моно-, би-, три- и тетрасиалированных структур белков была проведена слабая анионообменная (WAX) хроматография.
8.2 - Результаты
8.2.1 - Подготовка проб
Осветленный супернатант клеточной культуры клона клеток-продуцентов Р5С3, культивируемого в однолитровом биореакторе (режим перфузии), очищали с использованием псевдоаффинную хроматографию с использованием красителя (CaptoBlue-Sepharose, GE, Healthcare) в качестве первого этапа очистки. После этого были выполнены два альтернативных этапа хроматографии: ОФ-ВЭЖХ или ХГВ (как было указано ранее).
Биохимическую и физико-химическую характеристирацию проводили в сравнении с препаратами PMSG двух коммерческих направлений.
Во время (ЭХ)-ВЭЖХ анализировали степень чистоты различных молекул. Чистота 90% была получена для reCG ОФ-ВЭЖХ, оставшийся процент, вероятно, соответствует субъединицам аир гетеродимера, которые диссоциируют во время проведения способ очистки ОФ-ВЭЖХ. Напротив, молекула reCG, очищенная с помощью ХГВ, имела чистоту 55%, поскольку основной примесью был избыток свободной альфа-субъединицы (43%). Препарат PMSG, очищенный с помощью ОФ-ВЭЖХ, демонстрировал чистоту 73% (Фиг. 5).
С помощью ДСН-ПААГ, ЭС-ВЭЖХ (Фиг. 5) и ИЭФ (Фиг. 6) можно было определить наблюдаемую молекулярную массу и профиль изоформ различных препаратов. Молекулярная масса коммерческого PMSG составляла приблизительно 67 и 66 кДа при анализе с помощью ДСН-ПААГ и ЭС-ВЭЖХ, соответственно; тогда как рекомбинантный вариант имел молекулярную массу 45 кДа и 46 кДа при анализе с помощью ДСН-ПААГ и ЭС-ВЭЖХ, соответственно. Как рекомбинантные, так и коммерческие препараты PMSG демонстрировали сложные паттерны изоэлектрической фокусировки с большим разнообразием гликоформ с низкой изоэлектрической точкой. Однако, несмотря на то, что оба гормона имеют значительное количество общих изоформ, PMSG продемонстрировал более высокую долю гликоформ, сконцентрированных в более кислой зоне рН (этот эффект более выражен для Novormon), тогда как рекомбинантные варианты представляли более широкий спектр изоформ, распределенных по всему диапазон рН.
Очищенный препарат PMSG продемонстрировал tR, равное 12,954 мин, тогда как очищенные рекомбинантные варианты продемонстрировали tR, равное 13,583 и 13,747 мин для reCG, очищенного с помощью ОФ-ВЭЖХ и ХГВ, соответственно (Фиг. 5). Данные различия в гидрофобности трех молекул могут быть связаны с различиями в структурах гликанов, таких как содержание сиаловой кислоты, поскольку они являются одной из основных структур, придающих заряд белкам.
8.2.2 - Спектрофлуориметрический анализ
Поскольку профиль флуоресценции чрезвычайно чувствителен к возмущениям в локальной структурной среде, он предоставляет простые и убедительные доказательства, подтверждающие высокую степень структурного сходства между различными партиями данного белка. Кроме того, он может предоставить полезную информацию о сопоставимости и биоподобии продуктов (Houde et al., 2015). Структурная конформация, оцененная по эмиссионному профилю различных препаратов, выявила различия не только в их максимальных пиках, но и в профиле их спектров. Данные результаты должны указывать на различия в конформации различных препаратов (Фиг. 7).
8.2.3 - ВЭАОХ-ИАД
С помощью ВЭАОХ-ИАД (высокоэффективной анионообменной хроматографии с импульсным амперометрическим детектированием) с использованием системы DIONEX оценивали содержание сиаловой кислоты (Neu5Ac). PMSG (A, Foli-G), PMSG (В, Novormon), reCG ОФ-ВЭЖХ и reCG ХГВ продемонстрировали содержание Neu5Ac 9,4 (n=1); 18±4 (n=12); 7±1 (n=11) у 7±2 (n=11) моль Neu5 Ас/моль белка, соответственно. Эти результаты коррелируют с результатами, полученными с помощью анализов ОФ-ВЭЖХ и ИЭФ. Непараметрический статистический тест (медианный критерий Муда) позволил определить значимые различия между содержанием сиаловой кислоты в разных препаратах (р: 0,00048). Как видно на Фиг. 8, на диаграммах с доверительными интервалами («зарубками») представлены значимые различия между рекомбинантными гормонами и PMSG (В, Novormon) при доверительном уровне 95%.
Несмотря на то, что препарат PMSG демонстрировал более высокое содержание сиаловой кислоты, чем рекомбинантные варианты, как PMSG, так и рекомбинантные формы демонстрировали почти одинаковое соотношение сиаловая кислотаталактоза, поскольку соотношение остатков сиаловой кислоты и галактозы трех гормонов было сходным (PMSG (В, Novormon), reCG ОФ-ВЭЖХ и reCG ХГВ продемонстрировали содержание Gal 18±0,9 (n=2), 4,7±0,1 (n=2) и 6±0,6 (n=2) моль Gal/моль белка, соответственно). Следовательно, это один из атрибутов, который, вероятно, способствует более низкому удалению reCG из кровотока, снижению клубочковой фильтрации в почках и, таким образом, позволяет reCG проявлять свое биологическое действие у целевых видов животных. Кроме того, количество остатков маннозы между гормонами было сходным, хотя и меньшим, чем ожидалось, поскольку, если бы гетеродимер обладал тремя сайтами N-гликозилирования, количество Man (моль на моль белка) должно было бы составлять около девяти моль/моль. Таким образом, это меньшее содержание следует отнести к экспериментальной ошибке. Содержание фукозы было одинаковым между PMSG и рекомбинантными молекулами.
8.2.4 - Слабая анионообменная хроматография (WAX) для анализа заряженных меченых N-гликанов
Для оценки характера сиалирования очищенного коммерческого препарата PMSG (В) в сравнении с рекомбинантными вариантами из каждого гормона выделяли N-гликаны и метили их с помощью 2-АВ. Затем 2-АВ-меченные гликаны наносили на колонку WAX-ВЭЖХ и разделяли в соответствии с их зарядом и, в некоторой степени, в соответствии со структурой N-гликанов. Гликаны были идентифицированы как нейтральные (асиало-), моно-, би-, три- и тетрасиалированные структуры с использованием соответствующих стандартов.
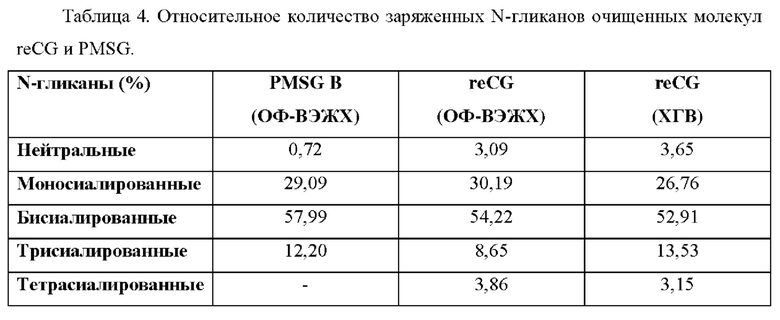
Как представлено на Фиг. 9, профили N-гликанов были одинаковыми для разных препаратов. Однако рекомбинантные препараты продемонстрировали более высокое содержание нейтральных гликанов и тетрасиалированных гликанов по сравнению с PMSG. Кроме того, в препаратах PMSG обнаружено большее количество бисиалированных гликанов, полностью замещенных остатками сиаловой кислоты (пик, соответствующий последнему положению, слева направо каждой группы), в отличие от рекомбинантных вариантов, которые демонстрировали неполные бисиалированные структуры (и, вероятно, более сложные).
Молекула reCG ОФ-ВЭЖХ содержала 3,09% нейтральных структур, 30,19% моносиалированных, 54,22% бисиалированных, 8,65% трисиалированных и 3,86% тетрасиалированных структур. reCG ХГВ содержал 3,65% нейтральных структур, 26,76% моносиалированных, 52,91% бисиалированных, 13,53% трисиалированных и 3,86% тетрасиалированных структур. Наконец, препарат PMSG продемонстрировал следующие проценты: 0,72% нейтральных структур, 29,02% моносиалированных, 57,99% бисиалированных и 12,20% трисиалированных структур (Таблица 4).
ПРИМЕР 9 - АНАЛИЗ ЭФФЕКТИВНОСТИ НА ЦЕЛЕВЫХ ЖИВОТНЫХ
9.1 - reCG-ИНДУЦИРОВАННАЯ СУПЕРОВУЛЯЦИЯ У ТЕЛОК
Использование однократной дозы eCG в протоколах суперовуляции и получения эмбрионов продемонстрировало эффективность, аналогичную той, которая достигается при применении многократных доз FSH. Цель данного исследования состояла в том, чтобы оценить эффективность reCG в индукции суперовуляции. Восемнадцать телок весом от 350 до 370 кг были синхронизированы с использованием следующего протокола: В День -10 телкам вводили 150 мкг PGF2α (Ciclar, ZOOVET); В день 0 им установили интравагинальное устройство с 1200 мг Р4 (ИВУ, Diprogest 1200, ZOOVET) плюс ввели инъекцию 2 мг ЕВ (бензоата эстрадиола, ZOOVET); На День 4 животных случайным образом делили на 4 группы и вводили: Группа 1 (n=4): 1000 ME reCG; Группа 2 (n-4): 1500 ME reCG; Группа 3 (n=5): 2000 ME reCG и Группа 4 (n=4): 2500 ME PMSG; На День 6 вводили 150 мкг PGF2α (Ciclar, ZOOVET); На День 7-й день ИВУ удаляли и повторяли инъекцию дозы PGF2α; На День 8 вводили 0,02 мг бусерелина ацетата (ZOOVET). Ультразвуковое исследование (УЗИ) проводили в Дни -10 и 0, чтобы определить стадию эстрального цикла в начале протокола. Кроме того, День 8 выполняли УЗИ для оценки количества фолликулов больше и меньше 8 мм (ФОЛ <8; ФОЛ> 8, соответственно) и количества желтых тел (ЖТ), а также допплерометрию (Mindray Z6Vet) проводили для оценки фолликулярной ирригации >8 мм.
Все полученные данные были проанализированы с помощью ANOVA с последующим апостериорным тестом Дункана. Переменные ирригации, Фол> 8 и ЖТ затем коррелировали с использованием корреляционного теста Пирсона (SPSS Statistics 23, IBM). Результаты обобщены в Таблице 5. Статистические различия наблюдались между группами 1 и 3 в отношении количества ФОЛ> 8 и ЖТ (Р <0,05). Фолликулярная ирригация положительно коррелировала с ФОЛ> 8 и ЖТ (Р <0,05). Как следует из результатов, reCG продемонстрировал дозозависимый эффект в отношении продуцирования преовуляторных фолликулов и ЖТ при одновременном увеличении дозы. Доза 2000 ME reCG оказалась столь же эффективной, как и PMSG, в индукции суперовуляции у телок. Наконец, было продемонстрировано, что более высокая фолликулярная ирригация коррелировала с большим количеством преовуляторных фолликулов и большим количеством ЖТ.
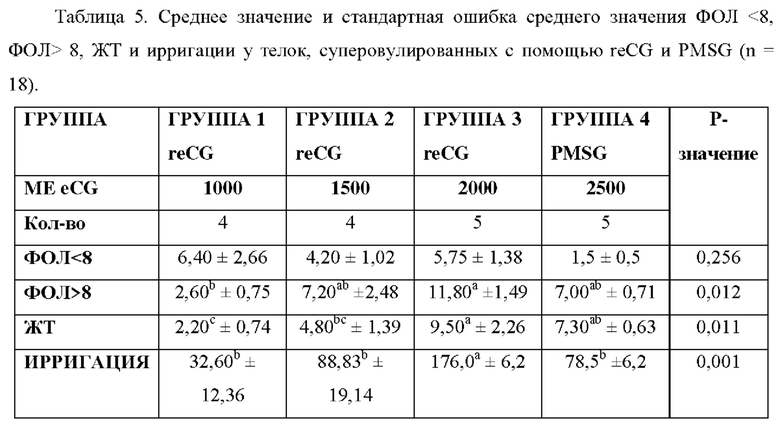
Значения с разными надстрочными буквами в столбце значимо различаются (Р<0,05).
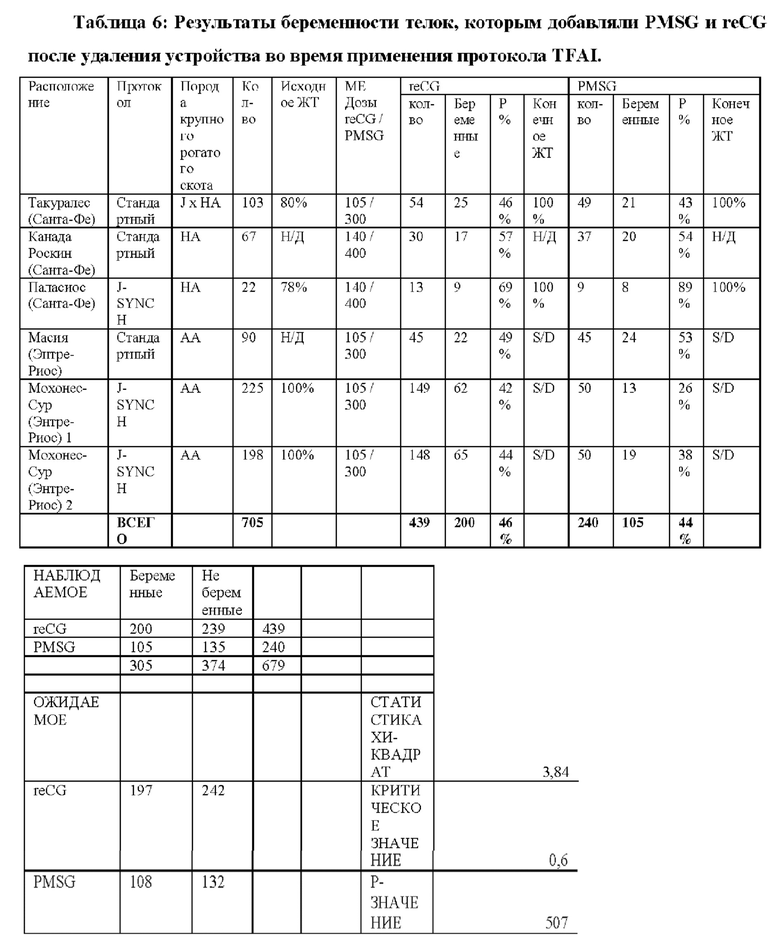
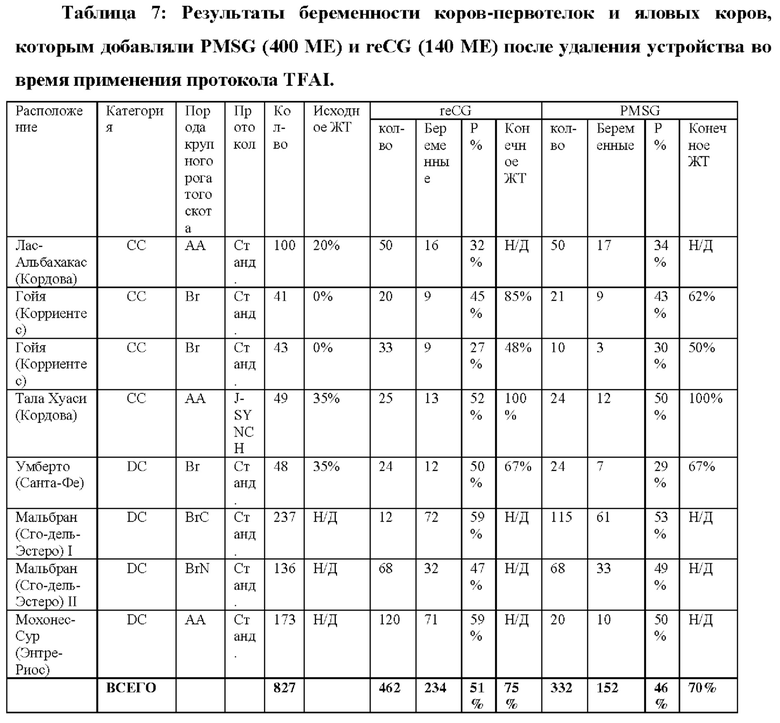
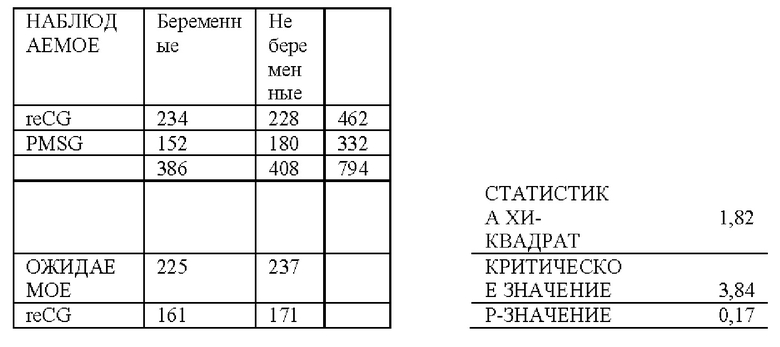
9.2- АНАЛИЗЫ ТЕСТОВ НА БЕРЕМЕННОСТЬ С ИСПОЛЬЗОВАНИЕМ reCG
9.2.1 - TFAI: Традиционный протокол
Традиционный протокол 8 последовательных дней использования интравагинального устройства, высвобождающего Р4.
День 0: Диагностическое УЗИ овариального статуса животных и конформации исследуемых групп. Применение интравагинального устройства с прогестероном 750 мг (Prociclar, Zoovet) плюс инъекция ES 2 мг (Zoovet).
День 8: Удаление устройства с последующими инъекциями 150 мкг D + Клопростенола (Ciclar, Zoovet), 1 мг эстрадиола ципионата (Zoovet) и 140 ME reCG или 400 ME PMSG, в зависимости от группы. Детектор половой охоты размещали на основание хвоста всех коров, чтобы определить наличие или отсутствие садки (половой охоты) перед синхронизированным искусственным осеменением (FTAI).
День 10 (48 ч после удаления устройства): Синхронизированное искусственное осеменение.
День 40 (День 30 от TFAI): Проведение ультразвуковой диагностики беременности или цикличности.
График ультразвукового исследования: Трансректальное УЗИ выполняли в момент времени 0 для определения состояния яичников у исследуемых самок. Затем на День 40 (30-й после осеменения) было проведено ультразвуковое исследование для установления диагноза беременности.
Сперма и осеменитель: Чтобы избежать возможных различий в фертильности вне протокола, которые могли повлиять на результат, все самки были осеменены одной и той же спермой от одного и того же быка и одной и той же серии пайет, и осеменение проводилось одним и тем же специалистом.
ПОЛУЧЕННЫЕ РЕЗУЛЬТАТЫ: Приводится сводка результатов, полученных во время процесса, и ультразвукового исследования на День 30 день после синхронизированного искусственного осеменения (FTAI).
9.2.2- Протокол FTAI - J-Syne
День 0: Диагностическое УЗИ овариального статуса животных и конформации исследуемых групп. Применение интравагинального устройства с прогестероном 600 мг (Diprogest, Zoovet) плюс инъекция 2 мг бензоата эстрадиола (ЕВ) (Zoovet).
День 6: Удаление устройства с последующими инъекциями 150 мкг D + клопростенола (Ciclar, Zoovet) и 105 ME reCG или 300 ME PMSG, в зависимости от группы. Детектор половой охоты размещали на основание хвоста всех коров, чтобы определить наличие или отсутствие садки (половой охоты) перед синхронизированным искусственным осеменением (FTAI).
День 9 (72 ч после удаления устройства): Всем животным вводили 0,010 мг бусерелина ацетата (Zoovet) с последующим синхронизированным искусственным осеменением.
День 43 (День 34 от TFAI): Ультразвуковое исследование для диагностирования беременности.
График ультразвукового исследования: Трансректальное УЗИ выполняли в момент времени 0 для определения состояния яичников у исследуемых самок. Затем на День 43 (34-й после осеменения) было проведено ультразвуковое исследование для установления диагноза беременности.
Сперма и осеменитель: Чтобы избежать возможных различий в фертильности вне протокола, которые могли повлиять на результат, все самки были осеменены одной и той же спермой от одного и того же быка и одной и той же серии пайет, и осеменение проводилось одним и тем же специалистом.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> UNIVERSIDAD NACIONAL DEL LITORAL
CONSEJO NACIONAL DE INVESTIGACIONES CIENTIFICAS Y TÉCNICAS (CONICET)
BIOTECNOFE S.A.
CEAGLIO Natalia
<120> СПОСОБ ПОЛУЧЕНИЯ ЛИНИИ КЛЕТОК МЛЕКОПИТАЮЩИХ, ЭКСПРЕССИРУЮЩИХ
РЕКОМБИНАНТНЫЙ ХОРИОНИЧЕСКИЙ ГОНАДОТРОПИН ЛОШАДИ (reCG),
ЛИНИИ РЕКОМБИНАНТНЫХ КЛЕТОК, ПРОДУЦИРУЮЩИХ reCG, СПОСОБ
КРУПНОМАСШТАБНОГО ПРОИЗВОДСТВА reCG, reCG, СОСТАВЫ, СОДЕРЖАЩИЕ reCG,
НУКЛЕИНОВЫЕ КИСЛОТЫ, КОДИРУЮЩИЕ reCG, И ИХ ПРИМЕНЕНИЕ
<130> -
<160> 2
<170> PatentIn версия 3.5
<210> 1
<211> 366
<212> DNA
<213> -
<400> 1
atggactact acagaaagca cgccgccgtg atcctggcta ccctgtccgt gttcctgcac 60
atcctgcata gcttccccga cggcgagttc acaacccagg actgccctga gtgcaagctg 120
agagagaaca agtacttctt caagctgggc gtgcccatct accagtgcaa gggctgctgc 180
ttctcccggg cctatcctac ccctgcccgg tccagaaaga ccatgctggt gcccaagaac 240
atcacctccg agtctacctg ctgcgtggcc aaggccttca tcagagtgac cgtgatgggc 300
aacatcaagc tggaaaacca cacccagtgc tactgctcca cctgttacca ccacaagatc 360
tgatga 366
<210> 2
<211> 513
<212> DNA
<213> -
<400> 2
atggaaacac tgcagggcct gctgctgtgg atgctgctgt ctgtgggcgg cgtgtgggct 60
tctagaggac ctctgaggcc cctgtgccgg cctatcaatg ctaccctggc cgccgagaaa 120
gaggcctgcc ctatctgcat caccttcacc acctccatct gcgccggcta ctgcccctcc 180
atggtgcgag tgatgccagc cgctctgcct gccattcctc agcccgtgtg cacctacaga 240
gagctgagat tcgcctccat ccggctgcct ggatgtcctc ctggcgtgga ccctatggtg 300
tccttccctg tggccctgtc ttgccactgc ggcccctgtc agatcaagac caccgactgc 360
ggcgtgttcc gggatcagcc tctggcatgt gcacctcagg cctccagctc ctccaaggac 420
cctccatctc agcccctgac ctccacctct acccctacac ctggcgcctc tcggagatcc 480
tctcaccccc tgcctatcaa gacctcctga tga 513
<---


Группа изобретений относится к биотехнологии. Представлены: способ получения линии клеток млекопитающих, которые экспрессируют гормон рекомбинантный хорионический гонадотропин лошади (reCG). Также описаны линия клеток, экспрессирующих reCG, крупномасштабный способ получения reCG, reCG с более высокой биологической активностью по сравнению с PMSG, состав, содержащий reCG, нуклеиновые кислоты, кодирующие reCG, и их применение. Изобретение может использоваться для индукции овуляции у самок животных. 9 н. и 18 з.п. ф-лы, 14 ил., 5 табл., 9 пр.
1. Способ получения линии клеток млекопитающих, экспрессирующих рекомбинантный хорионический гонадотропин лошади гормон (reCG), включающий следующие этапы:
a. получение кодирующих последовательностей α- и β-субъединиц reCG, которые имеют по крайней мере 90% идентичности с SEQ ID №1 и SEQ ID №2 соответственно, оптимизированных для их экспрессии в клетках млекопитающих;
b. введение указанных кодирующих последовательностей в лентивирусные векторы экспрессии;
c. получение лентивирусов, содержащих кодирующую reCG последовательность;
d. трансдукция клеток млекопитающих указанными выше лентивирусами;
e. отбор наиболее подходящего клона клеток млекопитающих для продуцирования reCG.
2. Способ по п. 1, отличающий тем, что указанная линия клеток млекопитающих обеспечивает продуцирование reCG не менее 100 МЕ/мл в бессывороточной среде.
3. Способ по п. 1, отличающийся тем, что на указанном этапе b. указанные лентивирусные векторы содержат лентивирусный вектор pLV, который содержит промотор EF-1α.
4. Способ по п. 1, отличающийся тем, что указанный этап с. включает транзиентную трансфекцию клеток HEK293 с плазмидами pREV, pVSVG, pMDL, pLV-reCG α и pLV-reCG β с использованием катионных липидов в качестве носителей.
5. Способ по п. 1, отличающийся тем, что указанный этап d. включает трансдукцию клеток СНО-К1.
6. Способ по п. 5, отличающийся тем, что он включает две последовательные трансдукции.
7. Линия клеток млекопитающих СНО, экспрессирующая рекомбинантный хорионический гонадотропин лошади гормон (reCG) для индукцирования овуляции у лошадей, полученная способом по п. 1, которая содержит нуклеиновую кислоту, кодирующую рекомбинантный хорионический гонадотропин лошади гормон (reCG), при этом последовательности, кодирующие альфа- и бета-субъединицы указанного reCG, содержат последовательности, имеющие по меньшей мере 90% идентичности последовательностям SEQ ID №1 и SEQ ID №2 соответственно, при этом линия клеток млекопитающих СНО представляет собой линию клеток СНО-К1.
8. Линия клеток по п. 7, отличающаяся тем, что она продуцирует по меньшей мере 100 МЕ/мл reCG.
9. Способ получения рекомбинантного хорионического гонадотропина лошади гормона (reCG), включающий следующие этапы:
a. культивирование указанной линии клеток млекопитающих по п. 7 в биореакторе в бессывороточной среде для крупномасштабного производства reCG;
b. сбор супернатанта; и
c. очистку суперстанта с получением reCG.
10. Способ по п. 9, отличающийся тем, что он характеризуется продуктивностью по меньшей мере 100 МЕ/мл reCG в бессывороточной среде.
11. Способ по п. 9, отличающийся тем, что указанный этап а. включает культивирование в бессывороточной питательной среде, содержащей 50% коммерческой питательной среды Ехсе1302 и 50% фосфатно-солевого буфера.
12. Способ по п. 9, отличающийся тем, что указанный этап очистки с. включает псевдоаффинную хроматографию с использованием красителя.
13. Способ по п. 12, отличающийся тем, что в указанной псевдоаффинной хроматографии с использованием красителя применяется матрица CaptoBlue-Sepharose.
14. Способ по п. 9, отличающийся тем, что он также включает этап очистки ВЭЖХ с использованием колонки С4.
15. Способ по п. 9, отличающийся тем, что указанный этап очистки с. включает тангенциальную проточную фильтрацию и последующее концентрирование reCG.
16. Способ по п. 9, отличающийся тем, что указанный reCG обладает удельной активностью в единицах эффективности in vivo по отношению к массе белка, определенной с помощью ИФА, по меньшей мере 6000 МЕ/мг.
17. Нуклеиновая кислота, кодирующая альфа-субъединицу reCG, полученную способом по п. 9, при этом указанная нуклеиновая кислота содержит последовательность, имеющую по меньшей мере 90% идентичности с последовательностью SEQ ID №1.
18. Нуклеиновая кислота, кодирующая бета-субъединицу reCG, полученную способом по п. 9, при этом указанная нуклеиновая кислота содержит последовательность, имеющую по меньшей мере 90% идентичности с последовательностью SEQ ID №2.
19. reCG, полученный способом по п. 9, для индуцирования овуляции у лошадей, отличающийся тем, что кодирующие последовательности для альфа- и бета-субъединиц указанного reCG содержат последовательности, имеющие по меньшей мере 90% идентичности с последовательностями SEQ ID №1 и SEQ ID №2 соответственно, причем указанный reCG включает профиль гликозилирования с по меньшей мере 3% нейтральных структур и по меньшей мере 3% тетрасиалированных структур.
20. reCG, полученный способом по п. 9, для индуцирования овуляции у лошадей, отличающийся тем, что кодирующие последовательности для альфа- и бета-субъединиц указанного reCG содержат последовательности, имеющие по меньшей мере 90% идентичности с последовательностями SEQ ID №1 и SEQ ID №2 соответственно, причем указанный reCG содержит профиль гликозилирования с по меньшей мере 3% нейтральных структур, от 26 до 30% моносиалированных структур, от 50 до 55% бисиалированных структур, от 8 до 15% трисиалированных структур и по меньшей мере 3% тетрасиалированных структур.
21. Фармацевтический состав для индукции овуляции у лошадей, содержащий терапевтически эффективное количество reCG по п. 19.
22. Фармацевтический состав по п. 21, отличающийся тем, что он является лиофилизированным.
23. Фармацевтический состав по п. 21, отличающийся тем, что он является жидким.
24. Фармацевтический состав по п. 23, отличающийся тем, что он дополнительно содержит сахар, консервант, антиоксидант, маннит и антиагрегационный агент, и состав хранится охлажденным при 5°С без необходимости замораживания.
25. Фармацевтический состав по п. 23, отличающийся тем, что он содержит дигидрат цитрата натрия, моногидрат лимонной кислоты, аргинин, сахарозу, маннит, L-метионин, полоксамер 188, m-крезол и воду.
26. Способ индукцирования овуляции у самок животных, включающий введение по меньшей мере 140 МЕ/животного reCG, полученного способом по любому из пп. 19, 20.
27. Способ по п. 26, отличающийся тем, что введение указанной дозы вызывает овуляцию через 48 часов после ее применения.
| EP 3434686 A1, 30.01.2019 | |||
| WO 2016178087 A1, 10.11.2016 | |||
| EP 3398609 A1, 07.11.2018 | |||
| СУПЕРАГОНИСТЫ ГЛИКОПРОТЕИНОВОГО ГОРМОНА ДЛИТЕЛЬНОГО ДЕЙСТВИЯ | 2013 |
|
RU2668174C2 |

