ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Данное изобретение относится в целом к модифицированным гликопротеиновым гормонам, имеющим активность суперагонистов, и их применению при лечении состояний, связанных с активностью гликопротеинового гормона. Более конкретно, настоящее изобретение относится к модифицированным молекулам гликопротеинов, содержащих аминокислотные замещения и один или более вставленных пептидов в альфа-субъединицах, по сравнению с альфа-субъединицами дикого типа, причем такие модифицированные молекулы обладают улучшенными фармакологическими свойствами по сравнению с гликопротеинами дикого типа.
УРОВЕНЬ ТЕХНИКИ
Гонадотропины фоллитропин (фолликул-стимулирующий гормон, FSH) и хорионический гонадотропин (CG), лютропин (лютеинизирующий гормон, LH) и тиреотропный гормон (тиреотропный гормон, TSH) составляют семейство гликопротеиновых гормонов. Каждый гормон является гетеродимером из двух нековалентно связанных субъединиц: альфа и бета. В тех же видах аминокислотная последовательность альфа-субъединицы идентична во всех гормонах, в то время как последовательность бета-субъединицы является гормон-специфичной (Pierce, Ann Rev. Biochem 50: 465-495 (1981)). Тот факт, что последовательности субъединиц высоко сохранны от рыб до млекопитающих, означает, что эти гормоны эволюционировали от общего белка-предка (Фонтейн, Gen. Comp Endocrinol 32: 341-347. (1977)).
Предыдущие исследования с измененными гликопротеиновыми гормонами показали обнадеживающие данные. Например, в дополнение к обеспечению модифицированных гликопротеиновых гормонов с повышенной активностью, дальнейшие мутации показали увеличение рецепторов сродством связывания (см., например, WO 2005/089445 и WO 2005/101000). Тем не менее, в то время как аффинность была увеличена, исследования показали, что модифицированные гликопротеиновые гормоны были очищены настолько же быстро, если не быстрее, чем их аналоги дикого типа. Для того, чтобы генерировать клинически полезный суперагонист с повышенной активностью, модифицированный гликопротеиновый суперагонист должны иметь повышенный период биологического полураспада в дополнение к улучшению аффинности связывания рецептора. Тем не менее, предыдущие попытки дальнейших модификаций гликопротеиновых гормонов увеличить период полураспада и повысить биодоступность были менее чем удовлетворительными, и вместо этого модифицированные гликопротеиновые гормоны продемонстрировали только ослабленный ответ.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Данное изобретение касается модифицированного гликопротеинового гормона, содержащего аминокислотную последовательность, по меньшей мере одно консервативное замещение основной аминокислоты в Q13, EL4, Р16 или Q20 и инсерцию VNVTINVT (SEQ ID NO: 20) между D3 и Q5 альфа-субъединицами в гликопротеиновом гормоне.
В некоторых вариантах воплощения изобретения модифицированный гликопротеиновый гормон содержит по меньшей мере два или по крайней мере три основных аминокислотных замещения в Q13, Р16 и Q20. В некоторых вариантах воплощения изобретения модифицированный гликопротеиновый гормон дополнительно содержит основное аминокислотное замещение в EL4. В некоторых вариантах воплощения изобретения основная аминокислота представляет собой аргинин.
В некоторых вариантах воплощения изобретения альфа-субъединица содержит аминокислотную последовательность с по меньшей мере 85% идентичностью относительно SEQ ID NO: 11 и дополнительно содержит бета-субъединицу лютенизирующего гормона (LH), хорионического гонадотропина (CG), фолликулостимулирующего гормона (FSH) или тиреотропного гормона ((TSH). В некоторых вариантах воплощения изобретения альфа-субъединица происходит от человеческой альфа-субъединицы (SEQ ID NO: 6).
Настоящее изобретение включает модифицированный гликопротеиновый гормон, содержащий аминокислотную последовательность с по меньшей мере одним основным консервативным аминокислотным замещением на K15, K17, K20 или K24 и инсерцию NVTINV (SEQ ID NO: 1) между F6 и Т7 альфа-субъединицей гликопротеинового гормона.
В некоторых вариантах воплощения изобретения модифицированный гликопротеиновый гормон содержит по меньшей мере два, или по крайней мере три, или по крайней мере четыре основных аминокислотных замещения на K15, K17, K20 и K24. В некоторых вариантах воплощения изобретения модифицированный гликопротеиновый гормон дополнительно содержит основное аминокислотное замещение на Е18. В некоторых вариантах воплощения изобретения основная аминокислота представляет собой аргинин.
В некоторых вариантах воплощения изобретения альфа-субъединица содержит аминокислотную последовательность с по меньшей мере 85% идентичностью относительно SEQ ID NO: 7 и дополнительно содержит бета-субъединицу лютеинизирующего гормона (LH), хорионического гонадотропина (CG), фолликулостимулирующего гормона (FSH) или тиреотропного гормона ((TSH). В некоторых вариантах воплощения изобретения альфа-субъединицы получают из крупного рогатого скота, свиней, овец или альфа-субъединицы (SEQ ID NO: 2, SEQ ID NO: 5, SEQ ID NO: 3, соответственно).
Настоящее изобретение включает модифицированный гликопротеиновый гормон, содержащий аминокислотную последовательность с по меньшей мере одним консервативным замещением основной аминокислоты на K15, K20, Е18 или K24, и инсерцию NVTINV (SEQ ID NO: 1) между F6 и Т7 или, альтернативно, инсерцию NV между F6 и Т7 плюс инсерцию INV между Т7 и Т8 альфа субъединицы гликопротеинового гормона.
В некоторых вариантах воплощения изобретения модифицированный гликопротеиновый гормон содержит по меньшей мере два, или по крайней мере три, или по крайней мере четыре основные аминокислотных замещения в K15, Е18, K20 и K24. В некоторых вариантах воплощения изобретения модифицированный гликопротеиновый гормон включает инсерцию NVTINV (SEQ ID NO: 1) между F6 и Т7 альфа-субъединицей. В некоторых вариантах воплощения изобретения модифицированный гликопротеиновый гормон включает инсерцию NV между F6 и Т7 плюс инсерцию INV между Т7 и Т8 альфа-субъединицей. В некоторых вариантах воплощения изобретения основная аминокислота представляет собой аргинин или гистидин. В некоторых вариантах воплощения изобретения основная аминокислота представляет собой аргинин.
В некоторых вариантах воплощения изобретения альфа-субъединица содержит аминокислотную последовательность с по меньшей мере 85% идентичностью относительно SEQ ID NO: 4 и дополнительно содержит бета-субъединицу лютеинизирующего гормона (LH), хорионического гонадотропина (CG), фолликулостимулирующего гормона (FSH) или тиреотропного гормона ((TSH). В некоторых вариантах воплощения изобретения альфа-субъединица происходит из конской альфа-субъединицы (SEQ ID NO: 4).
Настоящее изобретение также относится к способу стимуляции гликопротеинового рецептора у животного, включающему введение указанных выше модифицированных гликопротеиновых гормонов данному животному. Настоящее изобретение также относится к способу стимуляции овуляции у животного, включающему введение любого из вышеперечисленных модифицированных гликопротеиновых гормонов данному животному. В некоторых вариантах воплощения изобретения животным является человек, корова, овца, свинья или конь.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На Фиг. 1 показано сАМР стимуляции в CHO-FSHR клетках с выбранными bFSH аналогами, производимыми временной трансфекцией. На Фиг. 1А показано сравнение FolltropIn®-V (pFSH), bFSH-WT (дикого типа) с bFSH-5R аналогом. На Фиг. 1B показано ослабление биологической активности в in vitro hFSH-TR4402 аналога (переходной 4402) на два N-концевые расширения (ANITV, NITV) и одной внутренней неогликолизации (V78N) (SPA сАМР). На Фиг. 1С показано сравнение DFSH-5R аналогов с инсерцией 1 (5R + инсерция 1) и инсерцией 2 (5R + инсерция 2).
На Фиг. 2А и 2В показано РK скрининг различных аналогов bFSH после однократной подкожной инъекции мышам. В каждом эксперименте 5 мышей использовали для каждого препарата. Образцы крови брали через 24, 32 и 48 часов после инъекции, плазменные уровни были вычтены и были выражены данные в % от введенной дозы (% ID). FSH в образцах плазмы анализировали с использованием FSH ELISA (Endodrine Technologies).
На Фиг. 3A-D проиллюстрирован анализ производства различных партий TR55601. Фиг. 3А иллюстрирует анализ гетерогенности заряда с помощью IEF и затем вестерн-блоттингтинга. Субоптимальное сиалилирование Партии 3 (дорожки 2 и 3) находится в резком контрасте с оптимальными сильно кислыми изоформами, обнаруженными в партии 4 (полосы 5 и 6). Дорожка 1, IEF 3-10 маркер; Дорожка 2, TR55601/Партии 3 (8 мкг); Дорожка 3, TR55601/Партии 3 (4 мкг); Дорожка 4 и 8, TR4401 (LUG); Дорожка 5, TR55601/Партии 4 (8 мкг); Дорожка 6, TR55601 /Партии 4 (4 мкг); Дорожка 8, IEF 3-10 маркер. Фиг. 3В иллюстрирует анализ заряженных изоформ с помощью нейраминидазы (холерный вибрион), IEF и Вестерн-блоттинг.Необработанные TR55601/Партия 4 образца (дорожки 2 и 3) и TR55601 /Партия 4 образец, обработанные нейраминидазой перед нанесением 3-10 IEF гель (полосы 4-6). Дорожка 1, IEF 3-10 маркер; Дорожка 2, необработанная TR55601/Партия 4 (8 мкг); Дорожка 3, необработанная TR55601/Партия 4 (4 мкг); Дорожка 4, обработанная TR55601/Партия 4 (4 мкг); Дорожка 5, обработанная TR55601 /Партия 4 (2 мкг); Дорожка 6, обработанная TR55601/Партия 4 (1 мкг). Профиль IEF для нейраминидаза дигестированных изоформ сместился в диапазоне pI от 7,8 до 10,0. Средний сдвиг pI составляет приблизительно 5 единиц рН и множественные полосы (близко к 10 полосам) превращаются в одну основную полосу (pI ~9,5) и три слабые полосы (pI 7,8-10,0), указывая, что большинство наблюдаемых гетерогенностей заряда (Фиг. 3А - Партия 4) зависит от концевых остатков сиаловой кислоты, с небольшими другими модификациями, такими как деамидирование и/или протеолитическая деградация. Остаточные основные группы (PI 4.8-5.5), приведенные на Фиг. 3В, являются неспецифическими, полученных от подготовки нейраминидазы. На Фиг. 3С проиллюстрировано анализ заряженных изоформ TR55601-Партия 5 по IEF 3-10 в пи градиентном геле (IEF 3-10 с последующим вестерн-блоттингтингом. Дорожка 1, IEF 3-10 маркер; Дорожка 2, TR55601-Партия 5 (4 мкг); Дорожка 3, TR55601-Партия 5 (4 мкг); Дорожка 4, TR55601-Партия 4 (4 мкг); Дорожка 5, TR55601-Партия 4 (8 мкг); Дорожка 6, TR55601-Партия 3 (4 мкг); дорожка 7, TR55601-Партия 3 (8 мкг). На Фиг. 3D показан SDS-Вестерн-блоттинг анализ TR55601 Партии 5 по сравнению с TR55601 Партией 4 и Fol-V. Дорожка 1, маркерный белок; Дорожка 2: Партия 4, 500 мкг; Дорожка 3: Партии 5, LUL; Дорожка 4: пустая дорожка; Дорожка 5: Партии 4, 4 мкг; Дорожка 6: Партия 5, 1 мкг; Полоса 7: Fol-V, 673 мкг; Дорожка 8: маркерный белок.
На Фиг. 4 приведены результаты классического биотестирования Steelman-Pohley с hCG увеличением яичников вес у незрелых (22 дня) самок крыс Спрэга-Доули. Массу яичников определяли через 72 часа после приема препарата. Данные представлены в виде среднего общего веса яичников двух яичников + SEM (n=5 на дозу, в группе). Крысы были стимулированы одной инъекцией испытуемого изделия или носителя, с добавлением 40 ME hCG. Были использованы следующие лекарственные группы: 1 группа получала hCG только (не FSH), группы 2-5 получали TR55601 партия 4 (0, 0,33 мкг, 1,0 мкг, 3,33 мкг, и 10 мкг, соответственно слева направо), группы 6-8, получали FolltropIn-V® (3,333 мкг, 10,000 мкг, и 30,000 мкг соответственно слева направо), и группа 9-10 получала TR4401 (1,0 мкг и 3,33 мкг).
На Фиг. 5 приведен протокол синхронизации фолликулярной волны для суперовуляции, индукции овуляции и фиксированным временем искусственного осеменения. В 8 инъекциях FolltropIn-VR (Bioniche) заменяли одной или двумя инъекциями TR55601.
На Фиг. 6 показано среднее число фолликулов (от 3 до 5 мм в диаметре) во время суперстимуляционной обработки мясных коров, получавших 60 мкг rFSH заданной одним им инъекции или 300 мкг FolltropIn-V (контроль) проведенной дважды в день IM инъекции в течение 4 дней (3 эксперимента комбинированные).
На Фиг. 7 показано среднее число фолликул 6 до 8 мм в диаметре при суперстимуляционной обработке мясных коров, обработанных 60 мкг rFSH заданной одной внутримышечной инъекцией или 300 мг FolltropIn-V (контроль), проведенной дважды в день IM ежедневных инъекций в течение 4 дней (3 эксперимента комбинированные).
На Фиг. 8 показано среднее число фолликулов > 9 мм в диаметре во время суперстимуляционной обработки мясных коров, получавших 60 мкг rFSH заданной одной внутримышечной инъекцией или 300 мкг FolltropIn-V (контроль), проведенной дважды ежедневной внутримышечной инъекцией более 4 дней (3 эксперимента объединены).
На Фиг. 9 показаны профили среднего диаметра всех фолликулов > 3 мм в диаметре во время суперстимуляционной обработки мясных коров, получавших 60 мкг rFSH заданной одной внутримышечной инъекцией или 300 мг FolltropIn-V (контроль), проведнной дважды ежедневной внутримышечной инъекцией более 4 дней (3 эксперимента объединены).
На Фиг. 10А приведено сравнение производства сАМР для инсерции в альфа-субъединицы человека (А2), инсерции без амино-концевого валина (Инсерция 2), 5 аргинина замещений только без инсерции (5R) и контроля только среды. Фиг. 10В показывает ЕС50 для трех испытанных конструкций.
На Фиг. 11А приведено сравнение производства сАМР в ответ на человеческую модифицированную альфа-субъединицц с инсерцией SEQ ID NO: 1 и модифицированную альфа-субъединицу крупного рогатого скота, которые не имеют инсерции. На Фиг. 11В приведено сравнение производства сАМР в ответ на человеческую модифицированную альфа-субъединицу и субъединицы крупного рогатого скота с и без различных инсерций.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение обеспечивает молекулы модифицированного суперактивного гликопротеинового гормона, показывающие удивительно повышенную эффективность и повышенный период биологического полураспада по сравнению с их аналогами дикого типа. Модифицированные означает, что, в то время как белок содержит аминокислотную последовательность, которая отличается от гликопротеиновых гормонов дикого типа, последовательность была изменена так, что она не совпадает с известной гликопротеиновой последовательностью другого вида гормонов. Суперактивность может быть оценена в соответствии с различными параметрами, в том числе активностью и эффективностью. Эффективность является параметром биологической активности, которая определяется путем измерения половины максимального ответа. Различия в эффективности определяются путем сравнения значения ответа гликопротеиновых гормонов аналога на полпути между базовым и максимальным значением (ЕС50) по сравнению гликопротеиновым гормоном дикого типа. Ответы гликопротеинового гормона могут быть измерены in vitro с использованием очищенных белков, или могут быть оценены следующей переходной трансфекцией нуклеиновой кислоты, кодирующей модифицированный белок. Ответы гликопротеинового гормона также могут быть измерены in vivo, то есть у животного, реагирующего на аналоги гликопротеинового гормона. Такие ответы охватывают любой известный клеточный или биологический и количественный или качественный ответ гликопротеинового гормона с его рецептором, например, производство сАМР, синтез белков, таких как прогестерон, способность к оплодотворению, скорость образования бластоцистов, развитие эмбриона в оплодотворенной яйцеклетке, и т.д. Эффективность (Vмакс) или максимальная реакция - это еще один параметр биологической активности. Как обсуждалось в данной заявке, параметры биологической активности могут изменяться в зависимости от количества рецепторов и связывания рецептора в анализе клеточной линии. В системах с меньшим количеством рецепторов или нарушенной связью, различия более заметны в плане Vmax (эффективность). В системах, где происходит надэкспрессия рецепторов, различия в эффективности более заметны.
Например, в тех случаях, когда модифицированный гликопротеиновый гормон является модифицированной FSH или CG молекулой, in vivo количественные и качественные параметры, такие как количество ооцитов, скорость оплодотворения и бластоцисты и скорости образования зародышей, могут быть измерены в максимально эффективной дозе для числа ооцитов. Максимально эффективной дозой для числа ооцитов является оптимальное количество суперактивного FSH для качества ооцитов и их количества. Максимально эффективная доза для числа ооцитов зависит от массы и скорости метаболизма животного. Например, максимально эффективная доза для большего животного с более медленной скоростью метаболизма, больше, чем максимально эффективная доза для меньшего животного с более высокой скоростью метаболизма. Максимально эффективная доза определяется эмпирически для каждого животного.
Однако, независимо от используемой системы, белки модифицированного суперактивого гликопротеинового гормона в соответствии с данным изобретением могут демонстрировать по меньшей мере, приблизительно от 2 до 10 кратное увеличение эффективности или по меньшей мере приблизительно 20-кратное, 30-кратное, 40-кратное, 50-кратное, 60-кратное, 70-кратное, 80-кратное, 90-кратное или даже 100-кратное увеличение активности по сравнению с диким типом, или увеличение максимальной эффективности от 2 до 10%, или по меньшей мере 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, или даже 100%-ное увеличение максимальной эффективности по сравнению с аналогом дикого типа. Суперактивные аналоги в соответствии с данным изобретением могут также обеспечивать приблизительно 5-10 кратное увеличение активности или от 5% до 10% увеличение максимальной эффективности по сравнению с FSH дикого типа. Некоторые из модифицированных белков в соответствии с данным изобретением могут демонстрировать по меньшей мере приблизительно от тридцати до пятидесяти кратное увеличение активности или от 30% до 50% увеличение максимальной эффективности по сравнению с диким типом. Таким образом, белки модифицированного гликопротеинового гормона в соответствии с данным изобретением могут быть полезны для лечения субъектов с низким количеством рецепторов или недостатками в ответе рецептора, так как модифицированный белок в соответствии с данным изобретением может поддерживать по меньшей мере на 10-кратное увеличение эффективности или 10%-ное увеличение максимальной эффективности даже в системах с низким количеством рецепторов или ответом.
Скорость всасывания модифицированного суперактивного гликопротеинового гормона может привести к увеличению продолжительности действия. Аналог модифицированного гликопротеинового гормона с уменьшением скорости поглощения и увеличения продолжительности действия может быть полезен для гипосенситивных субъектов, таких как тех, кто страдает от бесплодия. Скорость поглощения измеряют при помощи Ka. Скорость выведения измеряют при помощи Ke.
Молекулы модифицированного гликопротеинового гормона в соответствии с данным изобретением включают модифицированные белки видов, выбранных из группы, состоящей из человека, крупного рогатого скота, лошадей, свиней, овец, мышей, крыс, кроликов, приматов и др. Гликопротеиновый гормон рыб (также известный как GTH-1) может использоваться в аквакультуре, то есть, для того, чтобы помочь росту находящихся под угрозой исчезновения или других видов рыб в неволе. Другие виды модифицированных гликопротеиновых гормонов могут быть использованы в сельском хозяйстве для разведения, и далее в лабораторных условиях для тестирования влияния различных комбинированных мутаций на различные мужские и женские состояния, связанные с гликопротеиновым гормоном.
Молекулы модифицированного гликопротеинового гормона других видов имеют замещения в положениях, соответствующих положениям, в модифицированных молекулах гликопротеинового гормона человека (например, Таблица 1), крупного рогатого скота (см. Таблица 2), овец, лошадей и свиней, описанные в данной заявке, которые могут быть идентифицированы с помощью любой программы выравнивания, включая, но не ограничиваясь, применение программ DNASIS, ALIONment, SIM и GCG, таких как Gap, BestFit, FrameAlign и Compare.
Молекулы модифицированного гликопротеинового гормона в соответствии с данным изобретением включают, по меньшей мере, модифицированную альфа-субъединицу, где альфа-субъединица содержит по меньшей мере две основные аминокислоты, например на лизиновых остатках. В альфа-субъединицах человека, основные аминокислоты могут быть введены в положениях 13, 14, 16 и 20 дикого типа человеческой альфа-субъединицы (SEQ ID NO: 6). В других видах, основные аминокислоты могут быть введены в положениях, соответствующих положениям 15, 17, 20 и 24 дикого типа бычьей альфа (SEQ ID NO: 2), дикого типа свиньи альфа (SEQ ID NO: 5) и дикого типа овец альфа (SEQ ID NO: 3), и положениях 15, 20 и 24 дикого типа лошадей альфа (SEQ ID NO: 4). Глутаматный остаток в положении 18 (бычий, свиной, овечий и лошадиный) может быть также замещен основной аминокислотой. В некоторых вариантах воплощения изобретения основная аминокислота может представлять собой аргинин или гистидин. В некоторых вариантах воплощения изобретения основная аминокислота может быть аргинином.
Пептид с последовательностью NVTINV (SEQ ID NO: 1) или TNVTINV (SEQ ID NJ: 12), или VNVTINVT (SEQ ID NO: 20) может быть вставлен между аминокислотами D3 и Q5 человеческой альфа-субъединицы (SEQ ID NO: 6), а также между F6 и Т7 альфа-субъединицы крупного рогатого скота, свиней, овец и лошадей.
Альтернативно, альфа-субъединица модифицированного гликопротеинового гормона быков, свиней, овец или коней может включать инсерцию NV между F6 и Т7 плюс инсерцию INV между Т7 и Т8. Модифицированные белки в соответствии с данным изобретением могут также содержать дополнительные замещения, особенно консервативные замещения, которые не изменяют улучшенные свойства белка. Обычно, однако, такие модифицированные белки содержат менее чем пять замещений в положениях, иных, чем те, которые перечислены выше, и могут проявлять полную идентичность аминокислотной последовательности с соответствующим дикого типа гликопротеиновым гормоном альфа в положениях, иных, чем указанные выше положения.
Основные аминокислоты включают аминокислоты лизин, аргинин и гистидин, а также любые другие основные аминокилоты, которые могут быть модификацией любой из этих трех аминокислот, синтетическими основными аминокислотами, которые обычно не встречаются в природе, или любой другой аминокислотой, которая положительно заряжена при нейтральном рН. Основные аминокислоты, в частности, выбраны из группы, состоящей из лизина и аргинина.
Примеры модифицированных альфа молекул, имеющих основные аминокислотные замещения и пептидную инсерцию, показаны в SEQ ID NO: 11 (человека), SEQ ID NO: 7 (быка), SEQ ID NO: 8 (овцы), SEQ ID NO: 10 (свиньи), и SEQ ID NO: 9 (лошадей). Настоящее изобретение относится к модифицированным гликопротеинам с аминокилотными последовательностями с по меньшей мере 80%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или выше идентичностью с любой из SEQ ID NO: 7 до 11.
Модифицированные альфа-субъединицы белков модифицированного гликопротеинового гормона в соответствии с данным изобретением могут также иметь альфа-субъединицу, содержащую два, три, четыре или пять основных аминокислотных замещений. Замещенные аминокислоты могут быть остатками лизина, глутаматными остатками, остатками пролина или глутаминовыми остатками. Например, бычьи альфа-субъединицы дикого типа, один или более из лизинов в положениях 15, 17, 20 и 24 могут быть замещены, а также глутамат в положении 18, основной аминокислотой, такой как аргинин и гистидин. В дикого типа человеческих альфа-субъединицах, один или более из глутаминов в положениях 13 и 20 могут быть замещены, а также глутамат в положении 14 и пролин в положении 16 основной аминокислотой, такой как аргинин и гистидин. В дикого типа свиной альфа-субъединице, один или более из лизинов в положениях 15, 17, 20 и 24 могут быть замещены, а также глутамат в положении 18, основной аминокислотой, такой как аргинин и гистидин. В дикого типа овечьей альфа-субъединице, один или более из лизинов в положениях 15, 17, 20 и 24 могут быть замещены, а также глутамат в положении 18, основной аминокислотой, такой как аргинин и гистидин. В дикого типа лошадиной альфа-субъединице, один или более из лизинов в положениях 15, 20 и 24 могут быть замещены, а также глутамат в положении 18, основной аминокислотой, такой как аргинин и гистидин.
В качестве примера, дополнительно модифицированные бычьи альфа-субъединицы представлены в последовательностях, изложенных в SEQ ID NO: 13 до 19 и 22. Настоящее изобретение относится к модифицированным гликопротеинам с аминокислотными последовательностями с по меньшей мере 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или выше идентичностью с любой из SEQ ID NO: 13 с 19 и 22.
В качестве примера, дополнительно модифицированные лошадиные альфа-субъединицы представлены в последовательностях, изложенных в SEQ ID NO: 38 до 42. Настоящее изобретение обеспечивает модифицированные гликопротеины с аминокислотными последовательностями с по меньшей мере 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или выше идентичностью с любой из SEQ ID NO: 43 до 45.
Дополнительно модифицированные альфа-субъединицы могут быть сконструированы путем сравнения аминокислотных последовательностей рассматриваемой альфа-субъединицы с последовательностями других видов, чтобы определить соответствующие основные остатки в белках других видов. Такие способы описаны в патенте США 6,361,992, который включен в данную заявку в качестве ссылки в полном объеме. Рассмотрение может быть также предоставлено относительной биологической активности гликопротеинового гормона из различных видов, чтобы выбрать виды для сравнения и замещения. Кроме того, гомологическое моделирование на основе структуры смежных гликопротеиновых гормонов полезна, чтобы идентифицировать остатки аминокислот на поверхности. Чтобы изменить дополнительные аминокислотные позиции, гликопротеиновый гормон последовательности людей и не-людей могут быть выровнены с помощью стандартных компьютерных программ, такой как DNASIS (Hitachi Software Engineering) или любой другой программы выравнивания, перечисленных выше, в том числе, но не ограничиваясь приведенным, ALIONment, SIM и GCG программы, таких как Gap, BestFit, FrameAlign и Compare. Аминокислотные остатки, которые отличаются между человеческим и не-человеческим гликопротеиновым гормоном, могут быть замещены с помощью одного из указанных выше методов, и полученный гликопротеиновый гормон анализировали на его эффективности с помощью одного из упомянутых в данной заявке анализов.
Соответственно, данное изобретение также представляет модифицированный FSH белок, имеющий повышенную активность по сравнению с FSH дикого типа тех же видов, содержащие модифицированные альфа-субъединицы описанные в данной заявке.
Данное изобретение также представляет модифицированный LH белок, имеющий повышенную активность по сравнению с дикого типа LH тех же видов, содержащих модифицированные альфа-субъединицы описанные в данной заявке.
Данное изобретение также представляет модифицированный TSH белок, имеющий повышенную активность по сравнению с дикого типа TSH из тех же видов, содержащих модифицированные альфа-субъединицы. описанные в данной заявке. Данное изобретение также представляет модифицированный CG белок, имеющий повышенную активность по сравнению с дикого типа CG из тех же видов, содержащих модифицированные альфа-субъединицы описанные в данной заявке.
Настоящее изобретение также охватывает фрагменты аналогов, описанных в данной заявке, которые имеют суперагонистическую или антагонистическую активность. Например, фрагменты модифицированных альфа-цепей в соответствии с данным изобретением могут быть использованы отдельно или в сочетании с любым фрагментом или полной длиной бета цепи, чтобы создать суперагонистические соединения. В некоторых случаях, фрагменты молекул модифицированной альфа-субъединицы согласно данному изобретению также могут быть использованы в качестве антагонистов, например, чтобы ограничить продолжительность активности терапевтического гликопротеинового гормона, после того как он был введен.
Данное изобретение также представляет последовательности нуклеиновых кислот, кодирующих модифицированные гликопротеиновые гормоны, описанные в данной заявке. Данное изобретение также представляет нуклеиновые кислоты, которые кодируют полипептиды с консервативными аминокислотными замещениями. Нуклеиновые кислоты в соответствии с данным изобретением могут кодировать полипептиды, которые транспортируют сахар. Выделенные нуклеиновые кислоты могут иметь по меньшей мере приблизительно 30%, 40%, 50%, 60%, 70%, 80% 85%, 90%, 95% или 99% идентичность последовательности с указанными выше последовательностями. Выделенные нуклеиновые кислоты могут кодировать полипептид, имеющий аминокислотную последовательность, имеющую идентичность, по меньшей мере приблизительно 30%, 40%, 50%, 60%, 70%, 80%, 85%, 90%, 95%, или последовательность, идентичную 99% аминокислотных последовательностей, кодируемых определенными выше номерами доступа. Выделенная нуклеиновая кислота, кодирующая транспортер, может гибридизоваться с определенными выше последовательностями нуклеиновых кислот.
Нуклеиновая кислота, кодирующая белок модифицированного гликопротеинового гормона, может быть генетически слита с последовательностями, контролирующими экспрессию, для экспрессии. Подходящие последовательности, контролирующие экспрессию, включают промоторы, которые применимы в целевом организме-хозяине. Такие промоторы хорошо известны специалистам в данной области для различных хозяев прокариотических и эукариотических организмов и описаны в литературе. Например, такие промоторы могут быть выделены из природных генов или могут быть синтетическими или химерными промоторами.
Данное изобретение также представляет кассеты экспрессии для инсерции нуклеиновой кислоты, кодирующей белок модифицированного гликопротеинового гормона в целевых молекулах нуклеиновых кислот, таких как векторы. Для этого, кассета экспрессии снабжена нуклеотидными последовательностями в 5'- и 3'-флангах, чтобы облегчить удаление из и инсерции в определенных положениях последовательности, как, например, сайты узнавания ферментов рестрикции или последовательности-мишени для гомологичной рекомбинации, как, например, катализируемой рекомбиназами. В дополнение к кассете нуклеиновой кислоты или молекуле экспрессии в соответствии с данным изобретением, вектор может содержать дополнительные гены, такие как маркерные гены, которые позволяют выбор упомянутого вектора в клетку-хозяин, приемлемую в подходящих условиях. Как правило, вектор также содержит один или несколько источников репликации. Векторы могут также содержать терминаторные последовательности, чтобы ограничить длину за пределами транскрипции нуклеиновой кислоты, кодирующей транспортеры в соответствии сданным изобретением.
Предпочтительно, чтобы молекулы нуклеиновых кислот, содержащихся в векторах, были функционально связаны с последовательностями, контролирующими экспрессию, позволяющими экспрессию, то есть обеспечивающими транскрипцию и синтез транслируемой РНК, в прокариотических или эукариотических клетках.
Термин ''изолированный'' относится к молекулам, отделенным от других компонентов клеток/тканей (например, ДНК или РНК), которые присутствуют в природном источнике макромолекулы. Термин ''изолированные'', используемый в данной заявке, также относится к нуклеиновой кислоте или пептиду, который по существу не содержит клеточного материала, вирусного материала, и культуральной среды, когда их получают способами рекомбинантных ДНК, или, по существу, свободным от химических предшественников или других химических веществ, при химическом синтезе. Кроме того, выделенная нуклеиновая кислота или пептид может включать кислоты или пептидные фрагменты нуклеиновых кислот, которые не встречаются в природе в виде фрагментов, и не будут найдены в естественном состоянии.
Термины ''плазмид'' и ''вектор'' используются взаимозаменяемо, поскольку плазмид является наиболее часто используемой формой вектора. Тем не менее, изобретение предназначено, чтобы включать такие другие формы векторов экспрессии, которые имеют эквивалентные функции и которые станут известны в данной области, относящейся к данной заявке. Вектор может быть любой из нескольких нуклеиновых кислот, в которую желательная последовательность может быть вставлена ограничением и лигированием для транспортировки между различными генетическими средами или для экспрессии в клетке-хозяине. Векторы, как правило, состоят из ДНК, РНК, хотя векторы также доступны. Векторы включают, но не ограничиваются ими, плазмиды и фагемиды. Клонирующий вектор является тем, который способен к репликации в клетке-хозяине, и который дополнительно характеризуется одним или более сайтов эндонуклеазной рестрикции, в которых вектор может быть вырезан определяемым образом и в который желаемая последовательность ДНК может быть лигирована так, чтобы новый рекомбинантный вектор сохранял свою способность к репликации в клетке-хозяине. В случае плазмид репликация требуемой последовательности может происходить много раз, так как плазмид увеличивается числом копий внутри бактерии-хозяине или всего одино раз в хозяине перед воспроизведением хозяина путем митоза. В случае фага репликация может активно происходит во время литической фазы или пассивно во время лизогенной фазы.
Векторы могут дополнительно содержать промоторную последовательность. Промотор может включать нетранслируемую последовательность нуклеиновой кислоты, как правило, расположенную выше по потоку кодирующей области, которая содержит сайт для того, чтобы инициировать транскрипцию нуклеиновой кислоты. Область промотора может также включать другие элементы, которые действуют как регуляторы экспрессии генов. В других вариантах воплощения настоящего изобретения вектор экспрессии содержит дополнительную область, чтобы помочь в выборе клеток, которые имеют вектор экспрессии. Последовательность промотора часто ограничена (включительно) на ее 3'-конце сайтом инициации транскрипции и проходит выше (5' направление), чтобы включить минимальное число оснований или элементов, необходимых для инициации транскрипции на уровне выше фоновых детектируемых значений. В промоторной последовательности можно найти сайт инициации транскрипции, а также домены связывания белка, ответственного за связывание РНК-полимеразы. Эукариотические промоторы часто, но не всегда, содержат TATA боксы и CAT боксы. Активация промоторов может быть специфической для определенных клеток или тканей, например, с помощью транскрипционных факторов, которые экспрессируются только в определенных тканях или промотор может быть повсеместным и способным к экспрессии в большинстве клеток или тканей.
Векторы могут дополнительно содержать один или несколько маркерных последовательностей, пригодных для использования при идентификации и селекции клеток, которые были превращены или трансфицированы вектором. Маркеры включают, например, гены, кодирующие белки, которые увеличивают или уменьшают сопротивление или чувствительность к антибиотикам или другим соединениям, гены, которые кодируют ферменты, деятельность которых детектируется стандартными анализами, известными в данной области (например, (3-галактозидаза или щелочная фосфатаза), и гены, которые заметно влияют на фенотип трансформированных или трансфицированных клеток, хозяев, колоний или бляшек. Векторы могут быть теми, которые способны к автономной репликации и экспрессии структурных генов, присутствующих в продуктах сегментов ДНК, к которому они функционально присоединены. Вектор экспрессии тот, в котором желательная последовательность нуклеиновой кислоты может быть встроена ограничением и лигированием так, что он функционально соединен или функционально связан с регуляторными последовательностями, и может быть экспрессирован как РНК-транскрипт.Экспрессия относится к транскрипции и/или трансляции эндогенного гена, трансгену или кодирующей области в клетке.
Кодирующая последовательность и регуляторная последовательность, функционально соединены, когда они ковалентно связаны таким образом, чтобы поместить экспрессию или транскрипцию кодирующей последовательности под влиянием или контролем регуляторных последовательностей. Если желательно, кодирующие последовательности будут транслированы на функциональный белок, две последовательности ДНК, как говорят, функционально соединены, если индукция промотора в регуляторных последовательностях приводит к 5' транскрипции кодирующей последовательности, и если природа связи между двумя последовательностями ДНК не (1) приводят к введению мутации сдвига рамки, (2) не мешает способности области промотора направлять транскрипцию кодирующих последовательностей, или (3) не мешает способности соответствующего РНК-транскрипта трансляции на белок. Таким образом, область промотора может быть функционально соединена с кодирующей последовательностью, если область промотора была способна осуществлять транскрипцию этой последовательности ДНК таким образом, что в результате транскрипт может быть транслирован на нужный белок или полипептид.
Некоторые аспекты настоящего изобретения включают трансформацию и/или трансфекцию нуклеиновых кислот. Трансформация является введением экзогенной или гетерологичной нуклеиновой кислоты внутрь прокариотической клетки. Трансфекция является введением экзогенной или гетерологичной нуклеиновой кислоты внутрь эукариотической клетки. Трансформация или трансфекция нуклеиновой кислоты может или не может быть интегрирована (ковалентно связана) в хромосомную ДНК, составляющую геном клетки. В прокариотах, например, трансформирующаяся нуклеиновая кислота может быть сохранена на эписомальном элементе, таком как плазмид или вирусный вектор. В отношении эукариотических клеток, стабильно трансфицированными клетками является та, в которой трансфекция нуклеиновой кислоты стала интегрирована в хромосому так, что она наследуется дочерними клетками посредством хромосомной репликации. Эта стабильность демонстрирует способность эукариотической клетки установить клеточные линии или клоны, состоящие из популяции дочерних клеток, содержащих трансфицированную нуклеиновую кислоту.
Существуют многочисленные экспрессирующие векторы Е.coli (кишечная палочка), известные любому специалисту в данной области, которые полезны для экспрессии инсерции нуклеиновой кислоты. Другие микробные хозяева, подходящие для использования, включают бациллы, такие как Bacillus subtilis, и другие энтеробактерии, такие как Salmonella, Serratia, и различные виды Pseudomonas. В этих прокариотических хозяевах можно также сделать векторы экспрессии, которые обычно содержат контролирующие экспрессию последовательности, совместимые с клеткой-хозяином (например, источник репликации). Кроме того, любое количество различных известных промоторов будет присутствовать, например, в системе промотора лактозы, триптофановой (Trp) промоторной системы, системы промоторной бета-лактамазы, или системы промотора из фага лямбда. Промоторы обычно контролируют экспрессию, необязательно, посредством операторной последовательности и имеют последовательности связывания рибосомного сайта, например, для инициации и терминации транскрипции и трансляции. При необходимости, концевая аминокислота метионин может быть обеспечена путем инсерции Met-кодона 5' и в рамку с последующей инсерцией нуклеиновой кислоты. Кроме того, карбоксильное-концевое удлинение инсерции нуклеиновой кислоты может быть удалена с использованием стандартных процедур олигонуклеотидного мутагенеза.
Кроме того, могут быть использованы экспрессии дрожжей. Есть несколько преимуществ систем экспрессии в дрожжах. Во-первых, существуют доказательства того, что белки, производимые в системах секреции дрожжей, проявляют правильное дисульфидное спаривание. Во-вторых, посттрансляционное гликозилирование эффективно осуществляется дрожжевыми секреторными системами. Пре-про-альфа-факторная лидерная область Saccharomyces cerevisiae (кодируемая MF''-1 геном) обычно используется для прямой секреции белка из дрожжей (Brake, Proc. Nat. Acad. Sci., 81: 4642-4646 (1984)). Лидерная область пре-про-альфа-фактора содержит сигнальный пептид и про-сегмент, который включает последовательность узнавания для дрожжевой протеазы, кодируемой геном KEX2: этот фермент расщепляет белок-предшественник на карбоксильной стороне сигнальной дипептидной последовательности расщепления Lys-Arg. Кодирующая последовательность FSH может быть слита в рамке считывания с лидерной областью пре-про-альфа-фактора. Этот конструкт затем помещают под контроль сильной промоторной транскрипции, такой как алкогольдегидрогеназный I промотор или гликолитический промотор. Кодирующая последовательность нуклеиновой кислоты с последующей трансляцией концевого кодона, а затем терминацией транскрипции сигналов. Альтернативно, кодирующие последовательности нуклеиновой кислоты могут быть слиты с кодирующей последовательностью второго белка, например, Sj26 или бета, галактозидазой, которые могут быть использованы для облегчения очистки слитого белка с помощью аффинной хроматографии. Инсерция сайтов расщепления протеазой, чтобы отделить компоненты слитого белка применима к конструктам, используемым для экспрессии в дрожжах. Эффективное посттрансляционное гликозилирование и экспрессия рекомбинантных белков также могут быть достигнуты в системах Baculoviras.
Клетки млекопитающих позволяют экспрессию белков в среде, которая способствует важные посттрансляционные модификации, такие как складывание и спаривание цистеина, добавлением сложных углеводных структур, и секреция активного белка. Векторы, применимые для экспрессии активных белков в клетках млекопитающих, характеризуются инсерцией кодирующей последовательности белка между сильным промотором вируса и сигналом полиаденилирования. Векторы могут содержать гены, придающие устойчивость к гигромицину, сопротивлению к гентамицину, или другим генам или фенотипам, подходящим для использования в качестве маркеров селективности или сопротивления метотрексата для амплификации гена. Химерный белок кодирующей последовательности может быть введен клеточные линии в клетках яичника китайского хомячка (СНО) с использованием устойчивых к метотрексату кодирующему вектору или других клеточных линий с помощью выбора подходящих маркеров. Наличие векторной ДНК в трансформированных клетках может быть подтверждено Саузерн-блоттинг-анализом. Производство РНК, соответствующей инсерция кодирующей последовательности может быть подтверждено с помощью Нозерн-блоттинг-анализа. Ряд других линий клеток-хозяев способных секретировать интактные человеческие белки были разработаны в данной области, и включают клеточные линии СНО, HeLa клетки, клеточные линии миеломы, клетки Jurkat и т.д. Экспрессирующие векторы для этих клеток могут включать последовательности, контролирующие экспрессию, такие как источник репликации, промотор, энхансер, и необходимые сайты обработки информации, такие как сайты связывания рибосом сплайсингом РНК, сайты полиаденилирования, и последовательности терминации транскрипции. Примерами последовательностей, контролирующих экспрессию, являются промоторы, полученные из генов иммуноглобулина, SV 40, аденовируса, бычьего вируса папилломы и т.п. Векторы, содержащие сегменты рассматриваемой нуклеиновой кислоты, могут быть переданы в клетку-хозяина хорошо известными методами, которые изменяются в зависимости от типа клетки-хозяина. Например, преобразование хлорида кальция обычно используют для прокариотических клеток, тогда как фосфат кальция, DEAE декстран, или липофектин опосредованная трансфекция или электропорация могут быть использованы для других клеток-хозяев.
Альтернативные векторы для экспрессии генов в клетках млекопитающих, сходные с разработанными для экспрессии человеческого гамма-интерферона, тканевого активатора плазминогена, фактор свертывания VIII, поверхностного антигена вируса гепатита В, Nexinl протеазы и основного белка основных эозинофилов, могут быть использованы. Кроме того, вектор может включать CMV промоторные последовательности и сигнал полиаденилирования, доступные для экспрессии нуклеиновых кислот, вставленных в клетки млекопитающих (например, COS-7).
Экспрессия гена или гибридного гена может быть in vivo или in vitro. In vivo синтез включает превращение прокариотические или эукариотические клеткки, которые могут служить в качестве клеток-хозяев для вектора. Альтернативно, экспрессия гена может происходить в лабораторной системе экспрессии. Например, in vitro системы транскрипции являются коммерчески доступными, и обычно используются для синтеза относительно больших количеств мРНК. В таких системах in vitro транскрипции, нуклеиновая кислота, кодирующая гликопротеиновый гормон может быть клонирована в вектор экспрессии смежный с промотором транскрипции. Например, векторы клонирования и экспрессии Bluescript II содержат сайты множественного клонирования, которые примыкают к сильным прокариотическим промоторам транскрипции (Stratagene). Доступны наборы, которые содержат все необходимые реагенты для синтеза в пробирке РНК из шаблона ДНК, такие как векторы Bluescript (Stratagene). РНК получают in vitro с помощью системы, так как это может быть транслировано in vitro для получения желаемого гликопротеинового гормона (Stratagene).
Другой способ получения гликопротеинового гормона состоит в том, чтобы связать два пептида или полипептида совместно методами химии белков. Например, пептиды или полипептиды могут быть синтезированы химически с использованием доступного в настоящее время лабораторного оборудования, используя химию Fmoc (9-флуоренилметилоксикарбонил) или Воc (трет-бутилоксикарбонил) (Applied Biosystems). Специалисты в данной области могут легко понять, что пептид или полипептид, соответствующий гибриду гликопротеинового гормона, могут быть синтезированы с помощью стандартных химических реакций. Например, пептид или полипептид могут быть синтезированы, а не отщеплены от его синтетической смолы, в то время как другой фрагмент гибридного пептида может быть синтезирован, а затем его отщепляли от смолы, тем самым открывая концевую группу, которая функционально заблокирована на другом фрагменте. По реакции пептидной конденсации эти два фрагмента могут быть ковалентно соединены через пептидную связь их карбоксильными и амино-концами, соответственно, с образованием гибридного пептида. (Grant, Synthetic Peptides: A User Guide, W.H. Freeman (1992) and Bodansky, Principles of Peptide Synthesis, Springer-Verlag (1993) и Bodansky, Principles of Peptide Synthesis, Springer-Verlag (1993)). Альтернативно, пептид или полипептид могут быть независимо друг от друга синтезированы in vivo, как описано выше. После выделения эти независимые пептиды или полипептиды могут быть связаны с образованием гликопротеинового гормона через аналогичные реакции конденсации пептида. Например, ферментативное или химическое лигирование клонированных или синтетических сегментов пептида может позволить относительно короткие пептидные фрагменты, которые будут присоединены к образованию более крупных пептидных фрагментов, полипептидов или целые белковые домены (Abrahmsen, Biochemistry, 30: 4151 (1991); Dawson, Science, 266: 776-779 (1994)).
Модифицированные гликопротеиновые гормоны в соответствии с данным изобретением могут быть рекомбинантными белками, полученными путем клонирования нуклеиновых кислот, кодирующих полипептид, в системе экспрессии, способной производить их фрагменты полипептида. Например, можно определить активный домен модифицированной альфа-субъединицы, который вместе с бета-субъединицей может взаимодействовать с рецептора гликопротеинового гормона и вызвать биологический эффект, связанный с гликопротеиновым гормоном. В одном примере, найдено, что аминокислоты не способствуют активности или специфичности связывания или сродство гликопротеинового гормона может быть удалена без потерь соответствующей активности. Например, амино или карбоксильные терминальные аминокислоты могут быть удалены из последовательно или нативного или модифицированного гликопротеинового гормона и испытаны в одном из многих доступных анализов, описанных выше для соответствующей активности. В другом примере, модифицированные белки в соответствии с данным изобретением могут иметь части терминальных амино или карбоксильных терминальных аминокислот, или даже внутреннюю область гормона, замещенную на фрагмент полипептида или другой фрагмент, такой как биотин, которые могут способствовать в очистке модифицированного гликопротеинового гормона. Например, модифицированный гликопротеин может быть слит с белком, связывающим мальтозу, либо через пептидную химию клонирования соответствующих нуклеиновых кислот, кодирующих два полипептидных фрагмента в вектор экспрессии таким образом, чтобы экспрессия приводила к кодирующей области в гибридном полипептиде. Гибридный полипептид может быть подвергнут аффинной очистке путем пропускания его через аффинную амилозную колонку и модифицированный гликопротеин может быть отделен от связывающей мальтозу области путем расщепления гибридного полипептида специфическим протеазным фактором Ха.
Активные фрагменты молекул модифицированного гликопротеинового гормона в соответствии с данным изобретением также могут быть синтезированы непосредственно, либо получены путем химического или механического разрушения большего гликопротеинового гормона. Активный фрагмент определяется как аминокислотная последовательность по меньшей мере приблизительно 5 последовательных аминокислот, полученных из природной аминокислотной последовательности, имеющей соответствующую активность, например, связывания или регуляторную. Фрагменты, присоединены ли они к другим последовательностям или нет, могут также включать инсерции, делеции, замещения или другие выбранные модификации конкретных областей или конкретных аминокислотных остатков, при условии, что активность пептида существенно не изменена или нарушена в сравнении с модифицированным гликопротеиновым гормоном. Эти модификации могут предусматривать какое-то дополнительное свойство, таким образом, чтобы удалить/добавить аминокислоты, способные к образованию дисульфидных связей, чтобы повысить его биологическую долговечность и т.д. В любом случае, пептид должен обладать биологически активнымй свойством, таким как связующая активность, регулирование связывания в связывающем домене и т.д. Функциональные или активные области гликопротеинового гормона могут быть идентифицированы путем мутагенеза конкретной области гормона, с последующей экспрессей и испытаниями экспрессированного полипептида. Такие способы очевидны для квалифицированного специалиста в данной области, и могут включать сайт-специфический мутагенез нуклеиновой кислоты, кодирующей рецептор.
Настоящее изобретение также охватывает слитые белки и химерные белки, содержащие мутации, описанные в данной заявке, в том числе, например, слитые с гликопротеином FSH. Такой слитый белок может быть получен путем лигирования соответствующих последовательностей нуклеиновых кислот, кодирующих желательные аминокислотные последовательности, друг с другом с помощью методов, известных в данной области, в собственной системе кодирования, и экспрессируя слитый белок любым из средств, описанных выше. Альтернативно, такой слитый белок может быть получен с помощью методов синтеза белка, например, с использованием пептидного синтезатора. Аналоги с одиночными цепями и химерные белки в соответствии с настоящим изобретением могут включать пептидный линкер между альфа- и бета-субъединицами или между различными участками химерного белка.
Характеристика суперагонистов гликопротеинового гормона
Эффект изменения или модификации дикого типа гликопротеиновых гормонов, описанных в данной заявке, может быть установлен любыми способами. Например, изменения в системах второго мессенджера в клетках, трансфицированных нуклеиновой кислотой, кодирующей модифицированные гликопротеиновые гормоны, могут быть измерены и по сравнению с аналогичными клетками, трансфицированными нуклеиновой кислотой, кодирующей дикого типа гликопротеиновый гормон. Альтернативно, активность модифицированного гликопротеинового гормона может быть определена из анализов связывания рецептора, анализов поглощения тимидина, анализов производства прогестерона, или анализов Т4 секреции. Специалисту в данной области может легко определить любой подходящий анализа и использовать его для определения активности дикого типа или модифицированного гликопротеинового гормона.
В одном из вариантов исполнения настоящего изобретения модифицированный гликопротеиновый гормон имеет эффективность, которая увеличилась по сравнению с эффективностью гликопротеинового гормона дикого типа. Эта повышенная активность может быть оценена с помощью любого из методов, указанных выше, или в любом другом соответствующем анализе, как легко определено специалистом в данной области. Увеличение эффективности не должно быть последовательными от анализа к анализу, или от клеточной линии до клеточной линии, так как они, конечно, будет варьироваться.
В другом варианте исполнения настоящего изобретения модифицированный гликопротеиновый гормон имеет максимальную эффективность, которая увеличивается по сравнению с максимальной эффективностью гликопротеинового гормона дикого типа. Это увеличение максимальной эффективности можно оценить с помощью любой из методик, указанных выше, или в любом другом соответствующем анализе, как легко определено специалистом в данной области. Увеличение максимальной эффективности не должно быть последовательными от анализа к анализу, или от клеточной линии до клеточной линии, так как они, конечно, будет варьироваться.
Другие анализы, подходящие для характеристики аналогов, описанных в данной заявке, описаны в PCT/US99/05908, которая включена в данной заявке в качестве ссылки в полном объеме. Например, различные иммунологические анализы могут быть использованы, включая, но не ограничиваясь ими, конкурентные и неконкурентные тест-системы с использованием таких методов, как радиоиммуноанализы, ELISA, анализы изоэлектрической фокусировки (IEF), иммунологические сэндвич-анализы, иммунорадиометриеские анализы, гель диффузионные реакции осаждения, анализы иммунодиффузии, in situ иммунологические анализы, вестерн-блоттинг, реакции осаждения, анализы агглютинации, анализы связывания комплемента, иммунофлуоресцентные анализы, анализы белка А и иммуноэлектрофоретические анализы и т.д.
Например, когда бета-субъединицы являются такими, что FSH улучшения качества и количества ооцитов могут быть оценены в анализах vitro и in vivo, суперактивные FSH могут быть использованы для улучшения качества и количества ооцитов от животных, в том числе, но не ограничиваясь ими, человека, мыши, крысы, примата, кролика, свиньи, лошади, овцы и собаки. Предпочтительно, суперактивный FSH вводят человеку или любому животному. Он является распространенным для повышения количества ооцитов улучшения и качества, которые должны быть определены, используя различные конечные точки процессе оплодотворения in vitro, таком как образование яйцеклеток, оплодотворение ооцитов, и образование бластоцисты. Эксперименты оплодотворения in vitro могут следовать ''протоколу суперовуляции '', в котором субъектов обрабатывают суперактивным FSH аналогом в соответствии с настоящим изобретением, что приводит к высвобождению и созреванию нескольких ооцитов. В in vitro экспериментам оплодотворения, FSH (суперактивный FSH и рекомбинантный дикого типа FSH) могут быть введены с HCG, чтобы вызвать овуляцию. Контрольные животные могут быть использованы, которые получают только HCG или сыворотку гонадотропина беременной кобылы (PMSG). Качество ооцитов может быть улучшено за счет увеличения скорости оплодотворения ооцитов у животного. Скорость оплодотворения суперактивного фолликулостимулирующего гормона может быть определена в in vivo или in vitro путем сравнения скорости оплодотворения, достигнутой с суперактивным FSH по сравнению со скоростью оплодотворения, достигнутой с таким же количеством рекомбинантного дикого типа FSH. Контрольное животное также может быть использовано, которое принимает hCG. Скорость оплодотворения может быть измерена в процентах двухклеточных эмбрионов, которые развиваются, на общее число ооцитов. Если происходит оплодотворение in vitro, два клеточных эмбриона можно пересчитать в чашках оплодотворения. У мышей, два клеточных эмбрионы развиваются через приблизительно двадцать четыре часа после оплодотворения. Скорость оплодотворения изменяется в зависимости от количества введенного суперактивного FSH. Животное может принимать множество источников суперактивного FSH. Скорость оплодотворения увеличивается по меньшей мере приблизительно на 10 процентов, в результате введения суперактивного FSH при максимально эффективной дозе на количество ооцитов. Скорость оплодотворения может увеличиваться по меньшей мере приблизительно на 20 процентов, предпочтительно по меньшей мере на 30%, 40%, 50%, 60%, 70%, 80%, 90% или 100% в результате введения суперактивного FSH при максимально эффективной дозе на количество ооцитов. Суперактивный фолликулостимулирующий гормон может улучшить качество ооцитов путем повышения скорости образования бластоцистов для оплодотворенной яйцеклетки. Скорость формирования бластоцистов может быть измерена путем определения процентной доли двухклеточных эмбрионов, которые образуют бластоцисты. Скорость формирования бластоцистов увеличивается в зависимости от того, находятся ли формы бластоцистов in vivo или in vitro. Скорость образования бластоцистов зависит от количества суперактивного фолликулостимулирующего гормона, который вводят. Скорость формирования бластоцистов увеличивается по меньшей мере приблизительно на 10 процентов, в результате введения суперактивного фолликулостимулирующего гормона в максимально эффективной дозе для количества ооцитов. Скорость формирования бластоцистов может увеличиваться по меньшей мере приблизительно на 20 процентов, предпочтительно по меньшей мере на 30%, 40%, 50%, 60%, 70%, 80%, 90% или 100% в результате введения суперактивного FSH при максимально эффективной дозе для количества ооцитов.
Суперактивный фолликулостимулирующий гормон может улучшить качество ооцитов путем увеличения общего количества зародышей в оплодотворенной яйцеклетке. Увеличение общего количества зародышей в оплодотворенных ооцитах увеличивается, в зависимости от того, происходит ли оплодотворение in vivo или in vitro. Увеличение общего количества зародышей в оплодотворенной яйцеклетке зависит от количества суперактивного фолликулостимулирующего гормона, который вводят. Общее количество эмбрионов в оплодотворенных ооцитах увеличивается по меньшей мере на приблизительно 10 процентов, в результате введения суперактивного фолликулостимулирующего гормона в максимально эффективной дозе для количества ооцитов. Общее количество эмбрионов в оплодотворенной яйцеклетке может увеличиться на по меньшей мере приблизительно 20 процентов, предпочтительно по меньшей мере на 30%, 40%, 50%, 60%, 70%, 80%, 90% или 100% в результате введения суперактивного FSH в максимально эффективной дозе для количества ооцитов.
Например, когда бета-субъединица является субъединицей CG, эффективный лютеинизирующий гормон с (LH)-подобной активностью может быть оценен in vitro и in vivo биологических анализах. Суперактивный CG индуцирует овуляцию, расширяет срок жизни желтого тела, увеличивает синтез прогестерона и способствует формированию вспомогательных желтых тел у некоторых видов. Такие действия приводят к более эффективному сбору ооцитов, повышению качества ооцитов у некоторых видов, улучшению сохранения беременности и повышению количества беременностей. Аналоги гликопротеинового гормона с повышенным сроком полувыведения сыворотки
Белки модифицированного гликопротеинового гормона в соответствии с данным изобретением могут быть также дополнительно модифицированы таким образом, что полураспад в плазме увеличивается по сравнению с аналогами дикого типа. Белки модифицированного гликопротеинового гормона в соответствии с данным изобретением могут дополнительно содержать потенциальный сайт гликозилирования, в том числе последовательности, содержащие сайты N-гликозилирования и/или О-гликозилирования. Например, размещение пептида NVTINV (SEQ ID NO: 1) или VNVTINVT (SEQ ID NO: 20) в альфа-субъединице обеспечивает потенциальный сайт гликозилирования альфа-субъединицы. Пептиды SEQ ID NO: 20 или SEQ ID NO: 1 могут быть помещены в человеческие дикого типа последовательности между D3 и Q5. Пептиды SEQ ID NO: 20 или SEQ ID NO: 1 могут быть помещены в последовательности дикого типа крупного рогатого скота, лошадей, свиней и овец между F6 и Т7. Вставленный пептид может дополнительно содержать дополнительный остаток треонина на амино-конце. Пептиды для инсерции, чтобы изменить гликозилирование, включают Н.В., INV и TNV пептиды, а также TNVTINV (SEQ ID NO: 12). Например, модифицированная альфа-субъединица из гликогормона может включать инсерцию NV между F6 и Т7 плюс инсерцию INV между Т7 и Т8. Увеличение периода полураспада также может быть предоставлено пегилированием или конъюгацией других соответствующих химических групп или путем конструкта слитых белков, имеющих повышенный период полураспада или любым другим способом. Такие способы известны в данной области, например, как описано в патенте США 5612034, патенте США 6225449 и патенте США 6555660, каждый из которых включен путем ссылки в полном объеме.
Период полураспада также может быть увеличен за счет увеличения количества отрицательно заряженных остатков в молекуле, например, количества глутаматных и/или аспартатных остатков. Такое изменение может быть осуществлено путем сайт-направленного мутагенеза. Такое изменение может быть также достигнуто с помощью инсерции из аминокислотной последовательности, содержащей один или более отрицательно заряженных остатков в белки модифицированного гликопротеинового гормона.
Период полураспада белка является мерой стабильности белка и указывает на время, необходимое для уменьшения половины восстановления концентрации белка. Период полураспада сыворотки белка модифицированного гликопротеинового гормона, описанного в данной заявке, может быть определен с помощью любого способа, приемлемого для измерения уровня гормонов в образцах субъекта в течение времени, например, но не ограничиваясь приведенным, иммунологическим использованием антител для измерения уровня в образцах сыворотки, взятых в течение периода времени после введения белков модифицированного гликопротеинового гормона, или путем обнаружения меченых молекул гормона, то есть, меченых молекул, в образцах, взятых у субъекта после введения меченых гликопротеиновых гормонов.
Способы лечения
Белки модифицированного гликопротеинового гормона в соответствии с данным изобретением могут быть использованы для лечения любого состояния, связанного с активностью гликопротеинового гормона. Белки модифицированного гликопротеинового гормона в соответствии с данным изобретением могут быть использованы для лечения субъекта, нуждающегося в этом. Субъект может быть животным, таким как млекопитающее, рептилии, рыбы, птицы и амфибии. Субъект может быть млекопитающим, таким как человек, корова, овца, свинья или лошадь. Животное включает скот и домашних животных, таких как собаки и кошки. Субъект может быть человеческим пациентом или животным, которое нуждается в активности улучшенного гликопротеинового гормона. Состояниями, связанные с активность гликопротеинового гормона, являются те, которые полностью или частично вызваны реагированием измененного гликопротеинового гормона, или те, которые получают пользу после введения гликопротеинового гормона. Например, такие состояния включают в себя, но не ограничиваются ими, дисфункции овуляции, дефекты лютеиновой фазы, необъяснимое бесплодие, мужское бесплодие, ограниченное по времени зачатие, низкую экспрессиею рецептора FSH, низкую чувствительность рецептора FSH, дефекты связывающего FSH рецептора, дефекты соединительного FSH рецептора, производство низкого уровня тестостерона, мужское облысение, и отказ или травму гипофиза.
Например, количество и качество ооцитов могут быть улучшены путем введения аналога суперактивного FSH, как описано в данной заявке, животному. Например, Заявители неожиданно обнаружили, что путем введения суперактивного FSH, содержащего модифицированный альфа-субъединицу, получают резкое увеличение количества и качества ооцитов. Влияние суперактивного FSH на количество и качество ооцитов могут быть дополнительно повышены за счет увеличения периода полураспада FSH в сыворотке в суперактивном FSH. Период полураспада сыворотки FSH может быть увеличен путем дальнейшего изменения суперактивного FSH. Дальнейшие модификации, в том числе, но не ограничиваясь теми, которые описаны выше, могут быть использованы, чтобы увеличить период полурапсада в сыворотке FSH.
Белки модифицированного FSH, CG, LH или TSH гликопротеинового гормона в соответствии с данным изобретением также могут быть использованы в терапевтических схемах вспомогательной репродукции в любом субъекте мужского или женского пола, и включают введение вспомогательного количества белков модифицированного гликопротеинового гормона субъекту. В таких способах, аналоги могут быть введены отдельно или в сочетании с другими терапевтическими средствами, например, включая, но не ограничиваясь ими, кломифен цитрат и GnRH (гонотропин-высвобождающий гормон). Белки модифицированного гликопротеинового гормона в соответствии с данным изобретением могут быть введены в виде комбинации одного или более гликопротеинов. Например, модифицированные альфа-субъединицы могут быть объединены с бета-субъединицами FSH, бета-субъединицей CG, бета-субъединицей TSH, и/или бета-субъединицей LH, вместе или по отдельности, и модифицированные гликопротеины затем вводят субъекту. Например, у субъекта с изолированным дефицитом гонадотропина (IGD), модифицированные FSH, CG, TSH, LH и могут быть введены субъекту, чтобы восстановить нормальную функцию половых желез. Широко известно в данной области, что гликопротеиновые гормоны, такие как FSH, CG, TSH, LH, являются неотъемлемыми в женской репродуктивной физиологии, и эти гликопротеиновые гормоны можно вводить субъекту, чтобы преодолеть ряд репродуктивных расстройств, и тем самым помочь воспроизведению.
Однократные и многократные режимы дозирования инъекций проверяли для модифицированного гликопротеина CG лошадей. Для суперстимуляции с помощью eCG аналога у лошадей, коров, или свиней, использовали одиночные или 2:1 сплит инъекции eCG аналога. Доза в диапазоне исследования для eCG аналога включала однократную im. инъекцию 30, 45, 60, 75, 90, 105 и 120 мкг. Оптимизация дозы протестирована в соотношении 2:1 и дополнительная вторая сплит-доза будет совпадать с лечением РСР2альфа на 6-й день или другую дату. Лечение модифицированным eCG производит суперовуляцию лошадей, коров и свиней.
Квалифицированному специалисту в данной области легко определить эффективное количество гликопротеинового гормона для введения, и оно будет зависеть от таких факторов, как масса, размер, тяжесть конкретного состояния и от типа самого субъекта. Терапевтически эффективное количество может быть легко определено с помощью обычных процедур оптимизации. Настоящее изобретение относится к гликопротеиновым гормонам с повышенной эффективностью по отношению к дикого типа гликопротеиновому гормону. Эти модифицированные гликопротеиновые гормоны позволят практикующим специалистам вводить более низкую дозу модифицированного гликопротеинового гормона относительно дикого типа гликопротеинового гормона для достижения подобного терапевтического эффекта, или альтернативно, вводить дозу модифицированного гликопротеинового гормона, аналогичную дозе дикого типа гликопротеинового гормона, чтобы достичь повышенного терапевтического эффекта.
В зависимости от того, вводят ли гликопротеиновый гормон перорально, парентерально, или иным образом, введение простагландина может быть в виде твердых, полутвердых или жидких лекарственных форм, таких как, например, таблетки, пилюли, капсулы, порошки, жидкости, кремы, и суспензии или т.п., предпочтительно в единичной дозированной форме, пригодный для доставки точной дозы. Гликопротеиновый гормон может содержать эффективное количество выбранного гликопротеинового гормона в комбинации с фармацевтически приемлемым носителем, и, кроме того, содержать другие медицинские агенты, фармацевтические агенты, носители, адъюванты, разбавители и т.д. Под ''фармацевтически приемлемым'' подразумевают материал, который не является биологически или иным образом нежелательным, т.е. материал можно вводить индивидууму вместе с выбранным гликопротеиновым гормоном, не вызывая неприемлемых биологических эффектов или взаимодействия неприемлемым образом с гликопротеиновым гормоном. Современные способы получения таких лекарственных форм известны или будут очевидны для специалиста в данной области; например, см. Remington's Pharmaceutical Sciences, latest edition (Mack Publishing).
Приведенные ниже примеры предназначены для описания и иллюстрации данного изобретения. Как таковые, они не должны быть истолкованы как ограничивающие объем данного изобретения. Специалисты в данной области оценят, что многие другие варианты воплощения изобретения также входят в объем данного изобретения, как это описано в данной заявке выше и в формуле данного изобретения.
ПРИМЕРЫ
Конструкция аналогов альфа субъединицы
FSH человеческого суперагониста гликопротеина с изменениями в а-субъединице в Q13R + E14R + P16R + Q20R (человеческий 4R) с диким типом бета-субъединицы продемонстрировали значительное превосходство связывания над их аналогами дикого типа.
В Таблице 1 приведено сравнение человеческой альфа дикого типа (WT) и выбранных hFSH суперагонистов первичной аминокислотной структуры. Показаны N-концевые участки человеческой альфа дикого типа (аминокислотные остатки 1-28 из 92 общих остатков) и мутантные формы. Расположение 4 замещений суперагонистов аргинина (R) находятся в затененной области. Выбранные 4 различные инсерции введения одной или двух дополнительных N-связанных углеводных цепей, отмечены между аминокислотной D3 и Q5 последовательностью дикого типа.
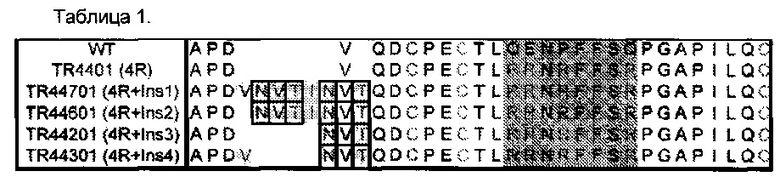
Сегменты в Таблице 1 перечислены в следующем виде: SEQ ID NO: 43: hFSH WT; SEQ ID NO: 33, hFSH альфа (4R); SEQ ID NO: 34, hFSH альфа (4R+Insl); SEQ ID NO: 35, hFSH альфа (4R+INS2); SEQ ID NO: 36, hFSH альфа (4R + в S3); SEQ ID NO: 37, hFSH альфа (4R+Ins4).
Замещения FSH (bFSH) крупного рогатого скота в некоторых вариантах воплощения изобретения весьма аналогичны остаткам ранее мутагенизированных в человеческом FSH альфа-субъединицам и включают сочетание 5 мутаций называемых ''5R'' (K15R+K17R+E18R+K20R+K24R). Например, SEQ ID NO: 7. Чтобы увеличить вероятность того, что введенные последовательности узнавания гликозилирования (NXT или NXS) приводят к присоединению N-связанной углеводной цепи, 18 различных бычьих конструктов альфа-субъединиц были созданы и клонированы в ранее разработанные векторы экспрессии. 12 конструктов содержали N-концевое удлинение пептидных последовательностей ANITV, ANTTA, ANTSA, ANITVNITV, ANTSANTTA и ANTSANTSA.
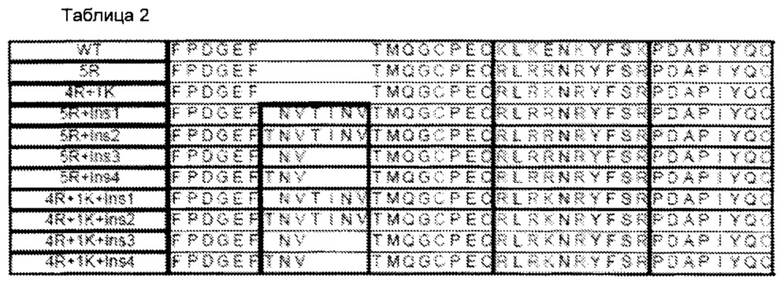
В Таблице 2 приведено сравнение бычьей альфа дикого типа (WT) и структур первичной аминокислоты, выбранных bFSH суперагонистов. Показаны N-концевые участки бычьей альфа дикого типа (аминокислотные остатки 1-32 из 96 общих остатков) и бычьей мутантной альфа. Расположение замещения 5 суперагониста аргинина (R) или лизина (K) отмечены в затененной области между аминокислотами С14 и Р25, кодированы как 5R из 4R + 1K. Выбранные 4 различные инсерции введения одной или двух дополнительных цепей N-связанных углеводных отмечены между F6 аминокислотой и Т8 последовательностью дикого типа.
Сегменты в Таблице 2 приведены следующим образом: SEQ ID NO: 44, WT bFSH; SEQ ID NO: 23, bFSH альфа (5R); SEQ ID NO: 24, bFSH альфа (4R+1K); SEQ ID NO: 25, bFSH альфа (5R+lnsl); SEQ ID NO: 26, bFSH альфа (5R+lns2); SEQ ID NO: 27, bFSH альфа (5R+lns3); SEQ ID NO: 28, bFSH альфа (5R+lns4); SEQ ID NO: 29, bFSH альфа (4R+1K+lnsl); SEQ ID NO: 30, bFSH альфа (4R+1K+lns2); SEQ ID NO: 31, bFSH альфа (4R+1K+lns3); SEQ ID NO: 32, bFSH альфа (4R+1K+lns4).
Гликопротеиновые альфа-субъединицы коней, в том числе мутации, были также подготовлены и некоторые варианты включают замещение K15R, E18R, K20R, и K24R. Например SEQ ID NO: 9; SEQ ID NO: 38-40. Кроме того, гликопротеиновые альфа-субъединицы коней, имеющие K15R, K20R, Е18Н и K24R, также были получены (например, SEQ ID NO: 41; SEQ ID NO: 42). Эти мутантные альфа-субъединицы также содержат инсерцию NVTINT между F6 и Т7 субъединицами (например, SEQ ID NO: 9, 40, и 42) или NV инсерцию между F6 и Т7 плюс INV между Т7 и Т8 (например, SEQ ID NO: 38, 39 и 41).
Переходная трансфекция bFSH аналогов с использованием полиэтиленимина (PEI) приводит в результате к 3,7-4,5 кратному увеличению экспрессии аналога bFSH по сравнению с методами на основе липофектамина (данные не показаны). Уровни экспрессии различных аналогов FSH на основе гетеродимерно-специфичной ELISA не показали большую потерю формирования FSH димера и не поддерживает предыдущие заявления, что одноцепочечный конструкт необходим, чтобы достичь высокого уровня экспрессии.
Выбор временно экспрессированных bFSH аналогов включал сАМР биотестирование in vitro и PK скрининг (см. Фиг. 1 и 2). Имело место значительное 3-4 кратное увеличение активности bFSH с 5R замещениями по сравнению с свиной FSH (pFSH-FolltropIn®-V) и контролей bFSH (Фиг. 1А). Примечательно, что в отличие от многих предыдущих исследований (Фиг. 1В), (Heikopp, Eur J. Biochem 261: 81-84, 1999; Trousdale, Fertil Steril 91: 265-270, 2009) новые инсерции неогликозилирования были обнаружены как не уменьшающие биологическую активность in vitro FSH 5R аналогов крупного рогатого скота (Фиг. 1С). Идентичные сайты неогликозилирования, добавленные в N-конец, понижали in vitro биологическую активность bFSH, аналогично ослабляющему влиянию неогликозилирования на внутреннюю активность эритропоэтина ((Elliott, Exp. Hematol. 32: 1146-1155, 2004; Elliott, Nat. Biotechnol. 21: 4144-421, 2003; Sinclair, J. Pharm. Sci. 94: 1626-1635, 2005) и эффект многих других продолжений а полураспада, включающих сайт-направленное легирование (Fishburn, J. Pharm. Sci. 97: 4167-4183, 2008; Uchiyama, Vet. J. 184: 208-211, 2010). Двухдневный PK скрининг исследование на мышах показали, что все нео-гликолизированные FSH ''5R'' аналоги крупного рогатого скота увеличили период полураспада по сравнению с bFSH-WT и FolltropIn®-V (Фиг. 2А и 2В). Данные РK скрининг-теста показали значительно более длительный период полувыведения из плазмы из-за гликозилирования в одном или двух введенных сайтах неогликозилирования (инсерция 1 и 2) в сравнении с bFSH-WT, FolltropIn®-V и контролем TR4401. Наблюдаемые уровни были сопоставимы с bFSH одноцепочеченой молекулы с линкером 29 аминокислот и 4-5 О-связанными углеводными цепями. Два первоначально испытанных аналога, выполненные в виде комбинированных инсерций суперагониста и неогликозилирования, названные ''Инсерция 1 и 2'' имели биологическую активность in vitro, сравнимую с 5R суперагонистом контроля самим по себе (Фиг. 1С) и все еще длительный период полураспада у мышей (Фиг. 2В). Такие длительно действующие аналоги без снижения активности суперагониста являются беспрецедентными и приводят к ожидаемой впечатляющей производительности in vivo у коров.
Несколько сотен миллиграмм различных FSH и TSH были получены рекомбинантно (rFSH или RTSH) с СНО клетки с помощью колбы, шейкера, роллер-флакона и биореакторов. Во время первоначальной работы дицистронные ретровирусные векторные системы была оптимизированы для высокой экспрессии уровня FSH и TSH аналогов в CHO-DG44 клетках. Препараты человеческого rFSH были протестированы. Одна 60 мкг доза rFSH индуцировала развитие фолликула, в результате чего большое количество эмбрионов хороших качества соответствовало ранее оптимизированнным восьми инъекциям FolltropIn®-V (300 мкг), которые вводили дважды в день в течение 4 дней, в дальнейшем поддерживая свои уникальные свойства, такие как замедленная абсорбция после внутримышечной, инъекции, а также увеличенное время пребывания FSH рецептора. См. Фигуры 3 и 5-8. RFSH суперагонист при 10 мкг дозе показал исключительную способность набора и поддержки в течение 12 дней повышенного пула растущих фолликулов, в частности фолликулов в 3-5 мм диапазоне размеров, которые также известны в организме человека как имеющие небольшое количество рецептора FSH (данные не показаны). Это неожиданное расширение и поддержка мелких фолликулов со стороны rFSH не наблюдается с ранее оптимизированной дозой контроля FolltropIn®-V, и обеспечивает новый способ набора FSH реагирующих фолликулов на случайных этапах цикла и повышает потенциал для успешного ЭКО и суперовуляции для ряда обедненных респондентов, вызванных снижением FSH числа рецепторов или функции (Perez Mayorga, J. Clin. Endocrinology 152: 3268-3369, 2000; Levallet, Arch. Med. Red. 30: 486-494, 1999; Rannikko,  . Hum. Reprod. 8: 311-317, 2002; Cai, Feril. Steril. 87: 1350-1356, 2007). Тем не менее, из-за разницы в 40 аминокислот между бычьим FSH и человеческим FSH, на основе альфа-субъединиц с 4 аргинина замещением, наблюдались некоторые переходные эффекты от предыдущих процедур с rFSH у тех же коров, что согласуется с данными предыдущих исследований, показывающими иммуногенные свойства человеческого FSH у кроликов и макак резус (Cai, Int. J. Toxicol. 30: 153-161, 2011; De Castro, Theriogenology 72: 655-662, 2009).
. Hum. Reprod. 8: 311-317, 2002; Cai, Feril. Steril. 87: 1350-1356, 2007). Тем не менее, из-за разницы в 40 аминокислот между бычьим FSH и человеческим FSH, на основе альфа-субъединиц с 4 аргинина замещением, наблюдались некоторые переходные эффекты от предыдущих процедур с rFSH у тех же коров, что согласуется с данными предыдущих исследований, показывающими иммуногенные свойства человеческого FSH у кроликов и макак резус (Cai, Int. J. Toxicol. 30: 153-161, 2011; De Castro, Theriogenology 72: 655-662, 2009).
Введение аргининовых (R) или лизиновых (K) остатков в выбранные модификации разрешающей области общей альфа-субъединицы, как ранее было показано, модулирует активность гликопротеиновых гормонов в процессе эволюции (Szkudlinski, Nat. Biotechnol. 14: 1257-1263, 1996; Szkudlinski, Physiol. Rev. 82: 473-502, 2002) и играет важную роль в электростатическом взаимодействии с отрицательно заряженным кластером, который находится в шарнирной области рецепторов гликопротеинового гормона (Mueller, Trends Endocrinol. Metab. 21: 111-122, 2010; Mueller, J. Biol. Chem. 284: 16317-16324, 2009; Mueller, Endocrinol. 152: 3268-3278, 2011). Развитие специфичного bFSH суперагониста включает минимальные замещения 4-5 R и/или K, чтобы получить более сильную и эффективную молекулу с возможной задержкой абсорбции для увеличения продолжительности действия, как показано в данных настоящей заявки и других исследованиях для hFSH и других аналогов гликопротеинового гормона (Szkudlinski, Physiol. Rev. 82: 473-502, 2002). Инсерции аминокислот минимальной длины, содержащих один или два углеводных сайтов неогликозирования, чтобы увеличить период полураспада для получения одного аналога инъекции без снижения повышенного потенции/эффективности суперагониста, также была исследована (см. Фиг. 4). Для дальнейшего анализа, 8 конструктов, содержащих пептидные инсерции NVTINV, NVTINVT, NV и NVT, расположенные между аминокислотой 6 и 8 последовательности дикого типа, могут быть использованы. Как показано в данных в данной заявке и в отличие от предыдущих исследований неогликозирования и пегилирования (Trousdale, Feril. Steril. 91: 265-270, 2009; Uchiyama, Vet J. 184: 208-211, 2010; Perlman, J. Clin. Endocrinol. Metab. 88: 3227-3235, 2003), можно спроектировать инсерцию аминокислотны минимальной длины, содержащей сложные углеводы, чтобы увеличить период полураспада без снижения повышенной потенции/эффективности суперагониста. Эти новые аналоги могут быть экспрессированы с помощью временной трансфекции в клетки СНО-K1 с использованием оптимизированного способа системной высокой экспрессии с помощью PEI и роллер-флаконов.
Очистка выбранных 4-6 аналогов, экспрессированных временной трансфекцией, может быть выполнена с использованием стадии захвата с помощью SP Sepharose колонки, а затем выбора аналогов при помощи Mono Q ионообменной хроматографии перед окончательной полировкой с использованием гель-фильтрации. Чистота аналогов bFSH может быть боле чем 98%. Накопительное восстановления может достигать 50% при полной 50-кратной очистке. Все аналоги могут быть охарактеризованы in vitro путем ELISA иммуноанализа, грубо в in vitro сАМР биоанализе с использованием клеточной линии CHO-FSHR, SDS-PAGE электрофореза и анализа геля изоэлектрической фокусировки (IEF). Выбранные очищенные аналоги также могут быть проанализированы с помощью точной количественной ВЭЖХ с обращенной фазой, углеводного композиционного анализа и оценки устойчивости и агрегационных состояний.
Дальнейшие эксперименты приводили к получению бычьего FSH аналога TR55601 (альфа-субъединицы: SEQ ID NO: 7), содержащего в себя замещения аргинина (R) в 15K, 17K, 18K, 20K, 24K и альфа-субъединицы, а также NVTINV (SEQ ID NO:1) инсерции между 6F и 7Т альфа-субъединицами. Несколько партий TR55601 были получены и испытаны с использованием IEF и вестерн-блоттинг анализа. На Фиг. 3А-D показано иллюстративные результаты анализа. На Фиг. 3A-3D показано, в частности, что TR55601/Партия 4 и Партия 5 демонстрируют оптимальное распределение и проверяют эффективность мутаций в этой партии. Партии 4 и 5 и партии, имеющие сходные результаты скрининга, были использованы при лечении животных. Например, анализ IEF--вестерн-блоттинг показывает оптимизированные кислотные изоформы TR56001 Партии 4 и Партии 5 на Фиг. 3А-С. С другой стороны, как показано на Фиг. 3А и 3В, Партия 3 была не полностью проверена и не был использована при лечении животных. Аналоги bFSH и, в частности, образцы TR55601 изучали также при помощи РK скрининга у мышей. Мышам делали подкожные инъекции выбранными образцами bFSH и образцы крови были отобраны через 24, 32 и 48 часов после инъекции. Плазма была выделена и проанализирована при помощи bFSH ELISA (Endocrine Technologies, Inc.). Длительный период полураспада Партии 4 образцов TR55601 был подтвержден при помощи РК скрининга. Данные не показаны.
Полный фармакокинетический профиль выбранных аналогов-кандидатов может быть выполнен с помощью подкожного введения разовой дозы 10 нг на крысу и 10 разных сборов крови (1, 5, 15, 30 мин и 1, 2, 6, 24 и 48 часов), охватывающих фазу распределения и выведения. Уровень FSH в бычьей плазме может быть количественно определен в плазме с помощью bFSH-ELISA. Полный анализ PK может быть выполнен.
Аналоги были выбраны для построения бицистронных векторов экспрессии, и выбора и усиления экспрессии аналога в CHO-DG44 клетках при подготовке для крупномасштабного производства, очистке и исследований суперовуляции крупного рогатого скота. CHO-DHFR(-) DG44 клетки совместно трансфицировали векторами экспрессии и подвергали амплификации гена в культуральной среде, содержащей приращения постадийные инкременты метотрексата (МТХ). Клетки были квалифицированы для следующей стадии амплификации после восстановления их многоугольной морфологии (2-3 недели). Клоны, которые представляли уровень секреции > 2 пг/клетка/день, могут быть подвергнуты второй обработке, направленной на амплификацию гена-маркера GS (MSX). Дополнительное 2-5-кратное увеличение можно получить, достигнув уровня секреции до 10 пг/клетка/день. Получение и эксперименты с аналогами альфа субъединицы
Хотя rFSH вызывали суперовуляторный ответ после однократной внутримышечной инъекции и 60 мкг, очень близко к оптимальной дозе, было значительное снижение суперовуляторного ответа, когда коровы были подвержены rFSH три раза или более. Таким образом, были разработаны эксперименты, чтобы протестировать новую rFSH композицию под названием TR 55601 rFSH, содержащую альфа-субъединицы (SEQ ID NO: 7), имеющую ''5R'' замещение и инсерцию NVTINT между F6 и Т7. Цель заключалась в первую очередь в том, чтобы определить эффект однократной или дробной дозы лечения rFSH, чтобы вызвать суперовуляторную реакцию у мясных коров, а затем дополнительно оценить суперовуляторную реакцию однократной инъекции rFSH и определить, остается ли суперовуляторная реакция высокой, когда коровы подвергают rFSH дважды или трижды.
Образцы TR55601 FSH были проанализированы in vivo у крыс при помощи классического Steelman-Pohley FSH биоанализа (Steelman et al., Endocrinol. 53: 604-616, 1953). Самкам крыс Спрэга-Доули (200-220 грамм) вводили одну дозу испытуемого изделия (например, bFSH) или основы, с добавлением 40 ME hCG. Массу яичников определяли через 72 часа после приема препарата. См. Фиг. 4. Каждую группу обрабатывали hCG, чтобы обеспечить фоновый уровень. TR55601 bFSH, при всех концентрациях, значительно увеличивает массу яичников.
На Фиг. 5 приведен протокол синхронизации фолликулярной волны для суперовуляции, индукции овуляции и фиксированного времени искусственного осеменения, где 8 инъекций FolltropIn-VR (Bioniche) заменяли на одинарное или двойное впрыскивание TR55601. Лечение предусматривало введение прогестерон (Р4) высвобождающего интравагинального устройства и введение эстрадиол бензоата (BE) в день 0. Суперовуляционное лечение было начато на 4 день с TR55601 как однократное или двойное впрыскивание. Вторая инъекция сплит-дозы совпадала с PGF2a лечением на 6-й день. Прогестероновое устройство было удалено с последней FSH инъекцией на день 7. На 8 день доноры получали свиной LH и их осеменяли без обнаружения эструса через 12 и 24 часов, или один раз на день 8 (через 16 часов после pLH). OVA/эмбрионы были собраны без хирургического вмешательства на 15-й день (2, 3). FolltropIn-V® использовали в качестве контроля; всего 300 мг* было дано в 8 внутримышечных инъекциях дважды в день в течение 4 дней (мг * - на основе NIH-FSH-P1 эталона с высоким содержанием примесей).
В частности, 30 некормящих коров породы красный ангус были стратифицированы и блокировали в зависимости от их предыстории производства эмбрионов и случайным образом распределены в одну из трех лечебных групп. Коровы в контрольной группе (n=10) получали 300 мг FolltropIn-V, внутримышечно в протоколе убывающей дозы два раза в день в течение 4-дневного периода. В частности: день 4, 3,0 мл (утро и вечер); день 5, 2,5 мл (утро и вечер); день 6, 1,5 мл (утро и вечер) и день 7, 0,5 мл (утро и вечер). Коровы в rFSH 60 мкг, группа лечения, получали однократную инъекцию внутримышечно 60 нг rFSH и коровы в rFSH 40-20 мкг, группа лечения, получали инъекцию внутримышечно 40 нг, rFSH на 4-й день с последующей еще одной внутримышечной инъекцией 20 мкг, rFSH на 6-й день.
Затем, 25 из 30 коров снова суперстимулировали FolltropIn-V (контроль) или однократным введением 60 нг rFSH. Животные в контрольной группе (n=10) оставались в контрольной группе, и 8 из 10 коров в rFSH 60 нг в эксперименте 1, были обработаны еще раз с помощью одной внутримышечной инъекции rFSH. Кроме того, 7 из 10 коров, ранее получавших сплит-однократные инъекции rFSH, лечили однократной внутримышечной инъекцией rFSH. Интервал между отборами эмбрионов составил 29 дней.
24 из 25 коров, используемых во втором эксперименте, были вновь суперстимулированы FolltropIn-V (контроль) или однократным введением 60 мкг rFSH. Опять же, коровы контроля оставались в контрольной группе, и rFSH коровы оставались в rFSH группе. Интервал между коллекциями эмбрионов составлял 30 дней.
На День 0 (начало эксперимента), все животные получали 5 мг эстрадиола-17β + 50 мг прогестерона и интравагинальное устройство, пропитанное прогестероном (Cue-Mate, Bioniche Animal Health). На день 4 (ожидаемый день возникновения фолликулярной волны), все коровы были суперстимулированы по группам, описанным выше. 60 мкг дозы для одной внутримышечной инъекции rFSH составляли 7,5 мл и FolltropIn-V вводили в 8 м инъекциях в протоколе уменьшения дозы. Все животные получали 500 нг клопростенола внутримышечно (Cyclase, Syntex, Argentina) в день 6 утром и вечером. Cue-mates были удалены вечером на 6-й день. Утром 8-го дня, коровы получили 100 нг гонадогерила (Gonasyn, Синтекс Аргентина) и оплодотворил 12 и 24 часов спустя. Все коровы были осемененены замороженной спермой того же быка. Ova /эмбрионы были собраны без хирургического вмешательства на 15-й день и оценены в соответствии с IETS рекомендациями.
Все коровы были исследованы с помощью ультразвука в дни 0, 4, 6 и 8 на присутствие CL и размер и количество фолликул, чтобы определить профили роста фолликулов. Овуляторная реакция была подтверждена путем подсчета количества CL и фолликул > 10 мм в диаметре с помощью УЗИ и пальпации прямой кишки на 15-й день.
В каждом эксперименте точки данных были оценены сначала на нормальность и однородность отклонений. Поскольку отклонения отличались между группами, данные были преобразованы квадратным корнем и проанализированы однонаправленным анализом ANOVA. Анализ общей реакции после трех экспериментов будет закончен двусторонней ANOVA для определения влияния экспериментального количества и лечения и их взаимодействия. Средние значения были сравнены с помощью теста защищенного LSD. Данные фолликул были проанализированы с помощью смешанной процедуры для выявления эффекта лечения, днем и их взаимодействия на количества фолликул и профили роста. Все анализы были выполнены с помощью программного обеспечения Infostat Analytical Software (Universidad Nacional de Cordoba, Argentina).
Данные суперовуляционного реагирования и яйцеклеток/эмбрионов приведены в Таблице 3. Хотя среднее число CL и коров с < 2 CL в день коллекции яйцеклеток/эмбрионов не отличалось между группами, сплит-инъекция rFSH приводила в результате к большему (Р<0,05) количеству неоплодотворенных фолликул. Среднее общее количество яйцеклеток/эмбрионов, оплодотворенных яйцеклеток и переносимых эмбрионов (классы 1, 2 и 3) также не отличалось.
В Таблице приведены данные суперовуляции с одинарными или сплит-одинарными дозами TR55601 (rFSH).

Суперовуляторный ответ и данные яйцеклеток/эмбрионов для второй дозы приведены в Таблицах 4 и 5. Среднее количество CL, фолликул > 10 мм и коров с < 2 CL в день сбора яйцеклеток/эмбрионов не отличалось между группами. Среднее общее количество яйцеклеток/эмбрионов, оплодотворенных яйцеклеток и переносимых эмбрионов (классы 1, 2 и 3) также не различались между группами.
В Таблицах 4 и 5 приведены данные суперовуляции с одной дозой TR55601 (rFSH).
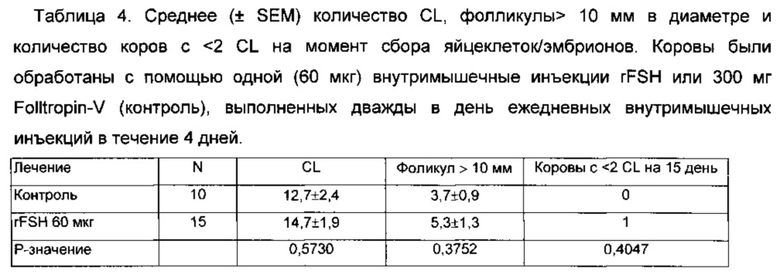
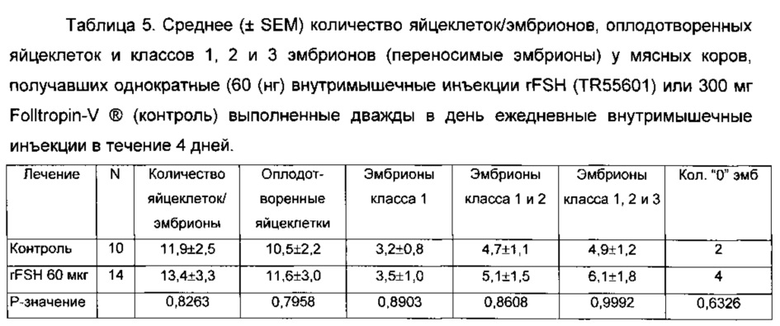
Для окончательного эксперимента по дозированию только данные фолликул доступны в данный момент. Не было никакого существенного различия в профилях роста фолликул и количеством фолликул в день перед осеменением между rFSH и FolltropIn-V группами. Данные овуляции и яйцеклеток/эмбрионов будут доступны в ближайшее время и будут представлены отдельно для эксперимента 3, а затем в сочетании с экспериментом 1 и 2. Данные фолликул в эксперименте 3 доступны и были объединены с данными экспериментов 1 и 2, и представлены в Таблице 6.
В Таблице 6 приведены данные по трем суперовуляциям с TR55601 (rFSH) с 30-дневными интервалами.
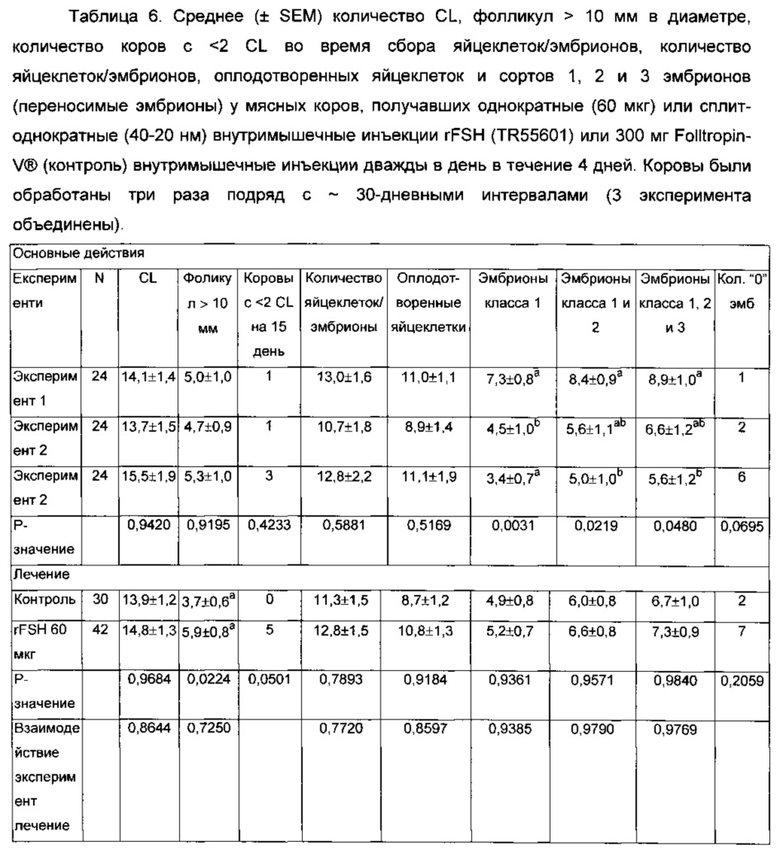
Характеристики фолликул от начала лечения непосредственно до искусственного осеменения показаны в Таблице 7 и на Фиг. 6-9.
В Таблице 7 приведены номера фолликул в модели суперовуляции у некормящих коров. Развитие фолликулов (в среднем ± SEM) был обнаружено при помощи УЗИ мясных коров, получавших 60 мкг rFSH однократной внутримышечной инъекцией или 300 мг FolltropIn®-V внутримышечной инъекцией дважды в день в течение 4 дней.
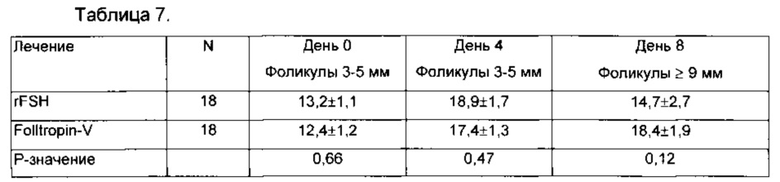
Количества фолликул 3-5 мм в диаметре не различались между группами лечения (Фиг. 6). Также не отличалось количество фолликул от 6 до 8 мм в диаметре (Фиг. 7), фолликул > 9 мм (Фиг. 8) или средний диаметр фолликул в течение дней лечения (Фиг. 9).
Результаты, полученные в этой серии опытов можно интерпретировать таким образом, чтобы предположить, что rFSH продукт индуцирует суперовуляторный ответ у мясных коров, который не отличается от FolltropIn-V. Также нет данных о снижении суперовуляторного ответа по сравнению с FolltropIn-V, когда коров обработали трижды подряд. Суперовуляторный ответ в конечном дозировании похож на первые два, и это будет подтверждено следующим набором OVA/эмбриона. Существует обеспокоенность по поводу увеличения числа неоплодотворенных (>10 мм) фолликул у коров, получавших rFSH (в основном из-за большого количества неоплодотворенных фолликул у двух коров), что должна быть предметом дальнейшего расследования. Другие исследования также необходимы, чтобы определить оптимальную дозировку rFSH супероплодотворенных коров и молочных коров и определить долгосрочные последствия лечения последовательно rFSH более чем в три раза.
Специфичность инсерции для улучшенного периода полувыведения
Чтобы определить, является ли наблюдаемое улучшение полураспада специфическим для инсерции последовательности SEQ ID NO: 1, человеческая альфа-субъединица с инсерцией была изменена, чтобы удалить аминоконцевой валин. Затем оба были испытаны на их способность продуцировать сАМР, вместе с аналогом, который не имеет инсерции. Исследования показали, что инсерция представляет собой конкретную последовательность и дает превосходное связывание и период полураспада в альфа-субъединице (см. Фиг. 9). Условия получения затем были рассмотрены в различных условиях роста для оптимизации максимального продуцирования (см. Таблицу 8).
В Таблице 8 приведены данные оптимизации продуцирования человеческой альфа-субъединицы.
Таблица 8
Инсерцию затем исследовали вместе с бычьим аналогом, и было подтверждено, что увеличение периода полураспада является специфическим для последовательности инсерции (см. Фиг. 11).





















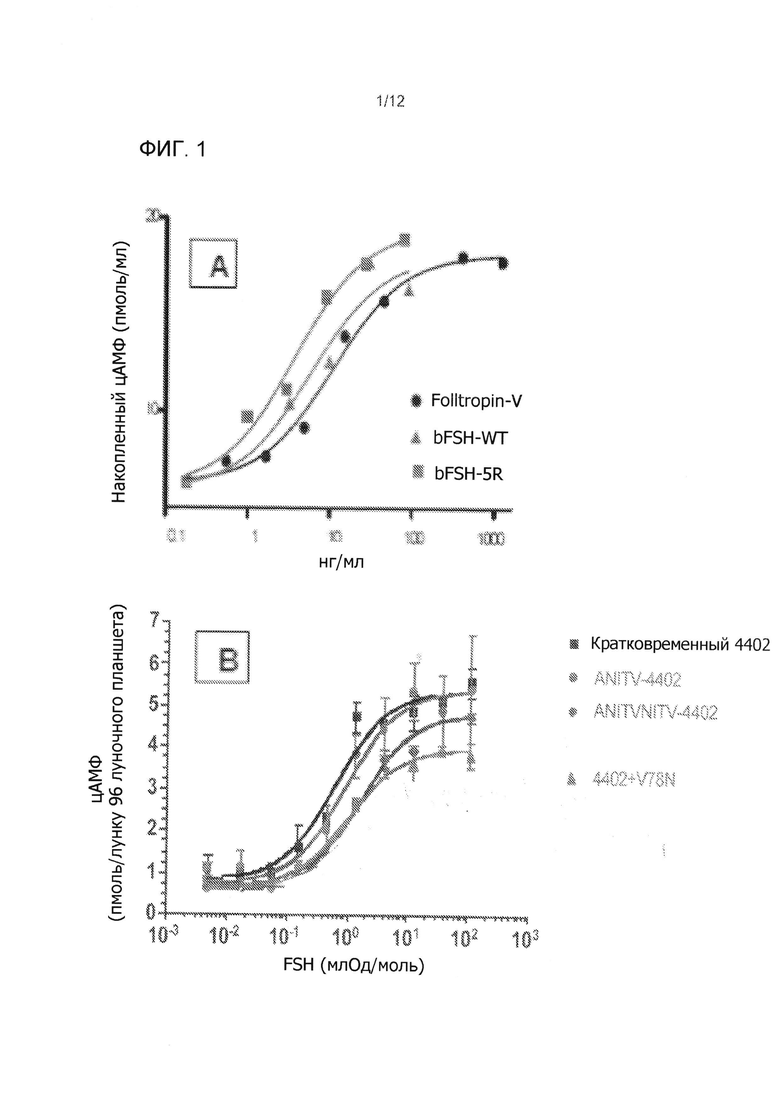
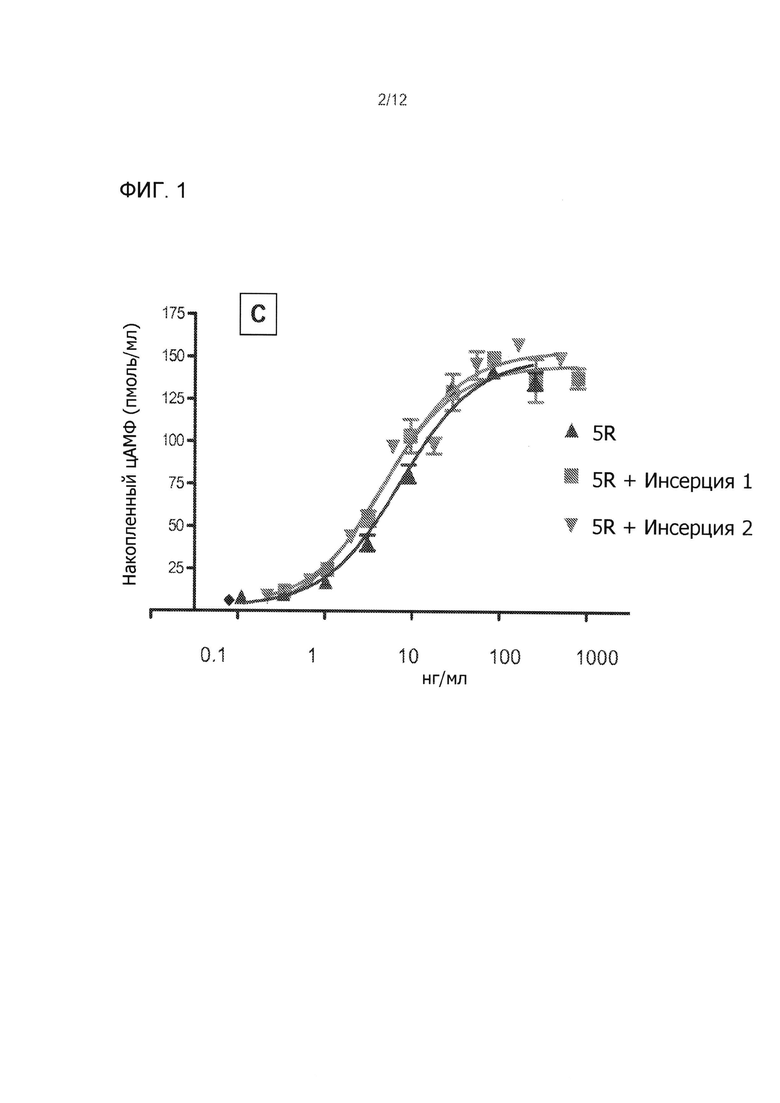
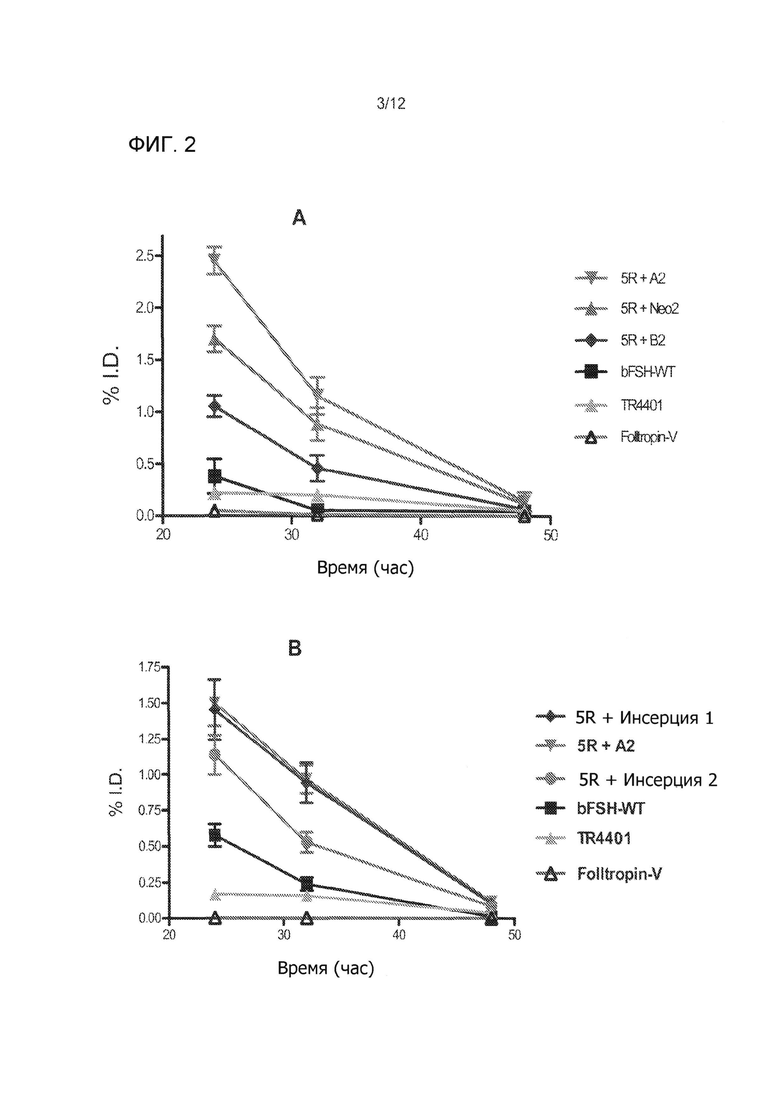
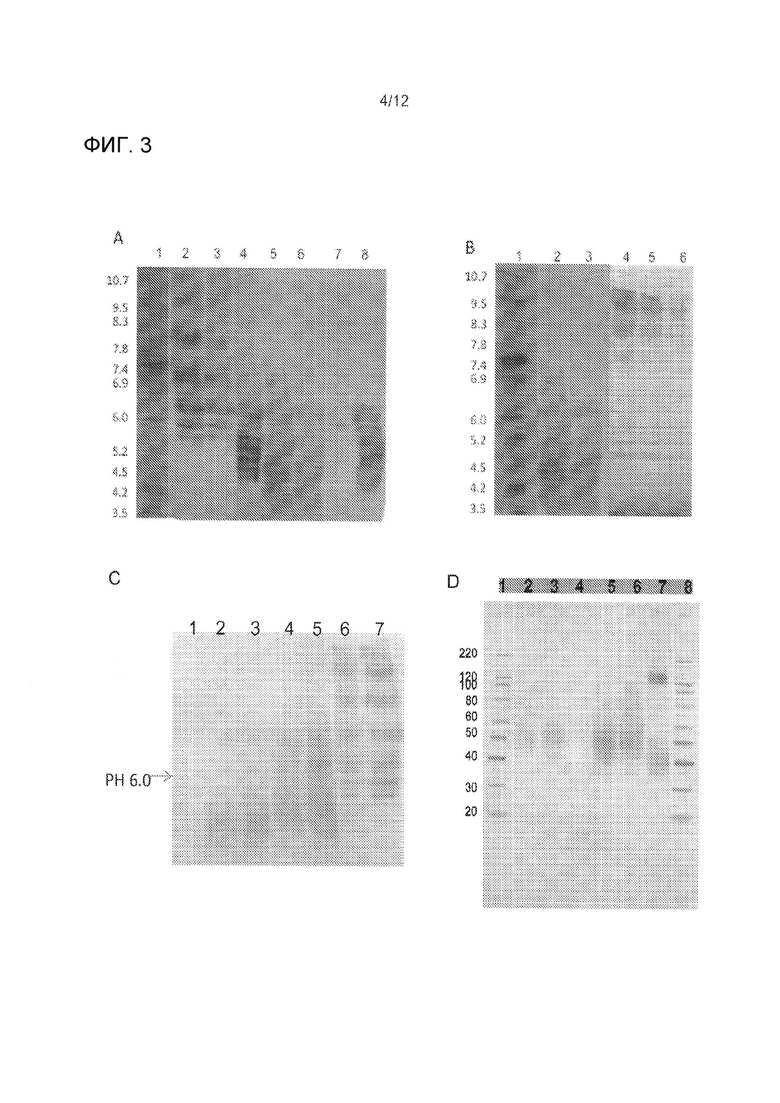
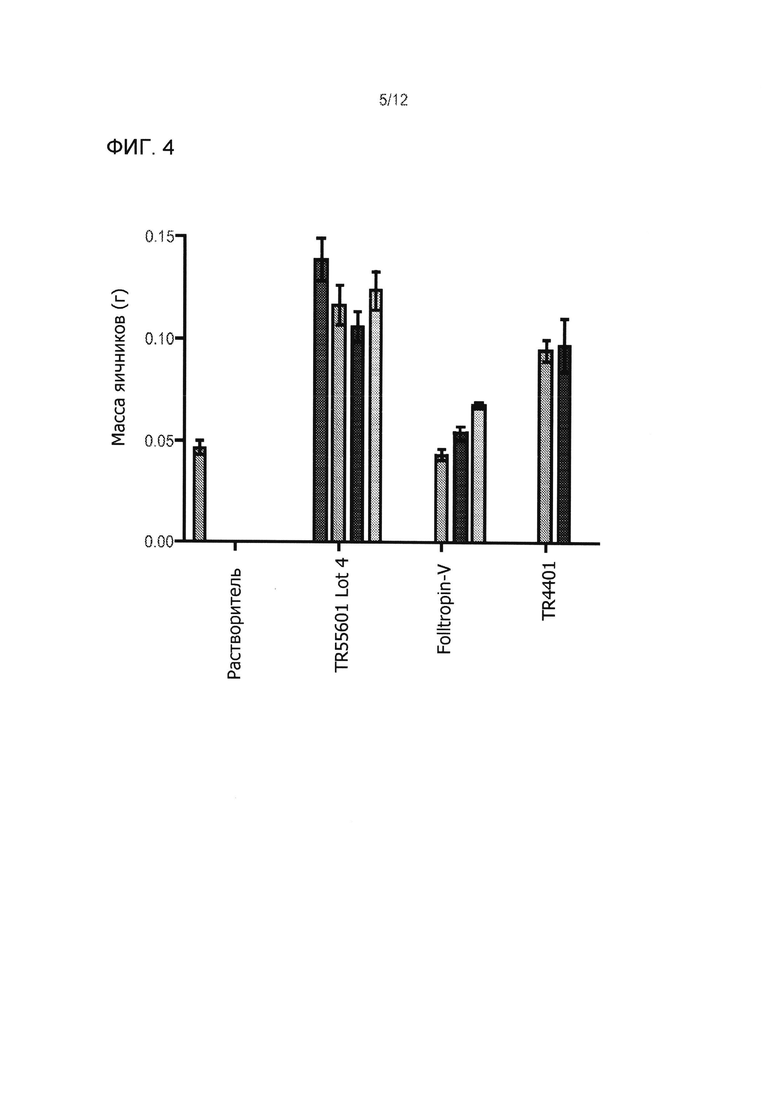
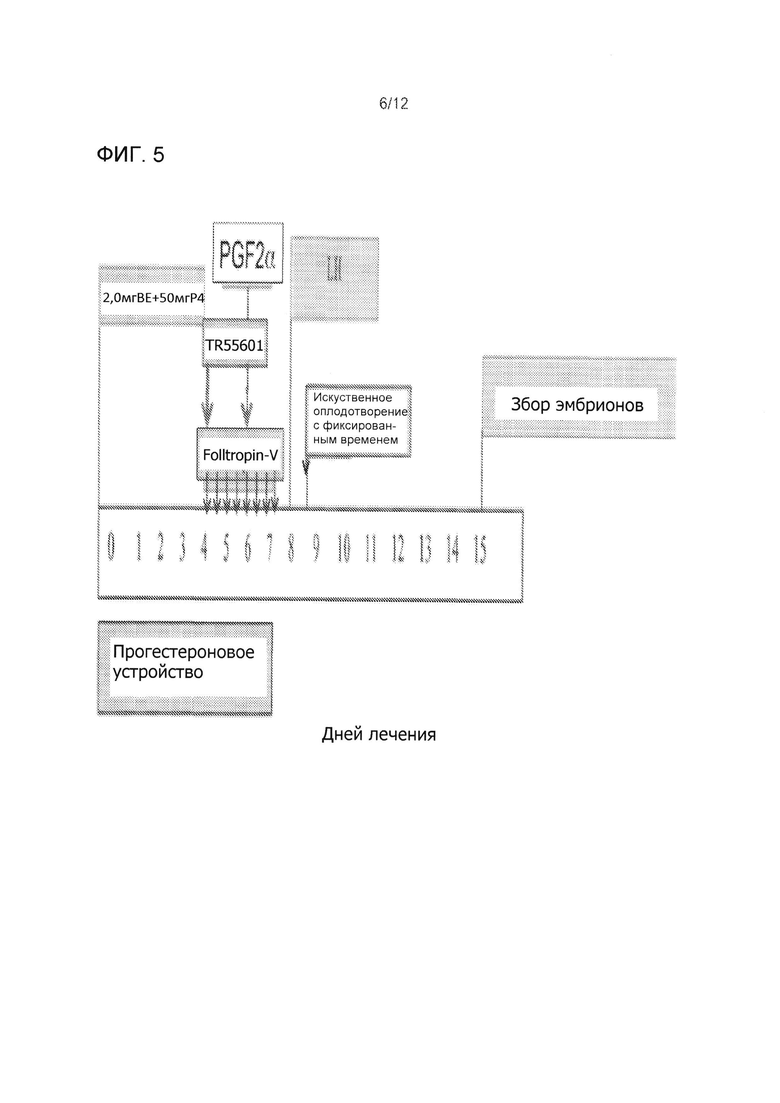
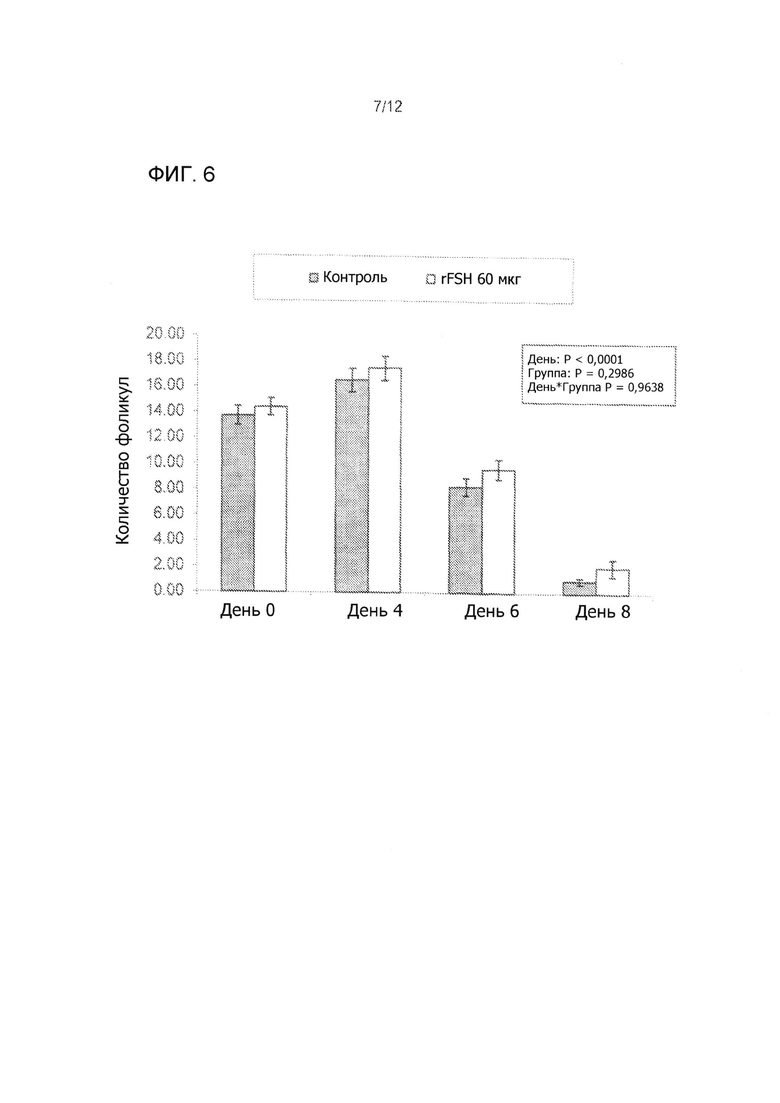
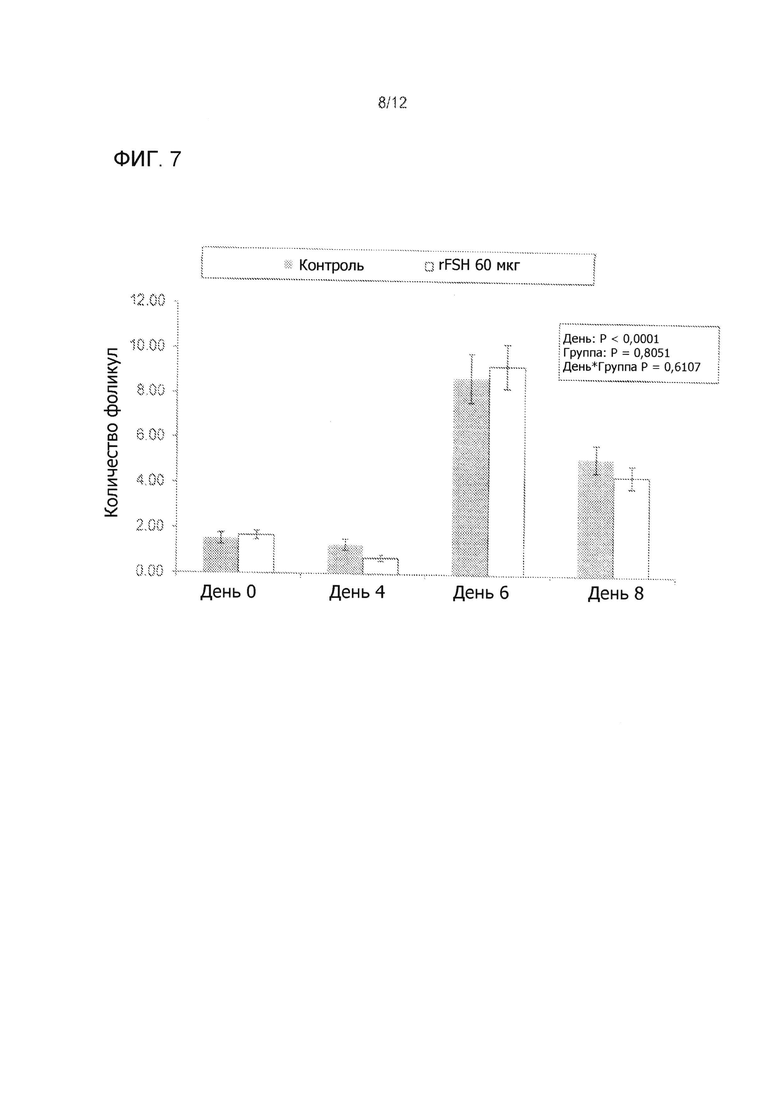
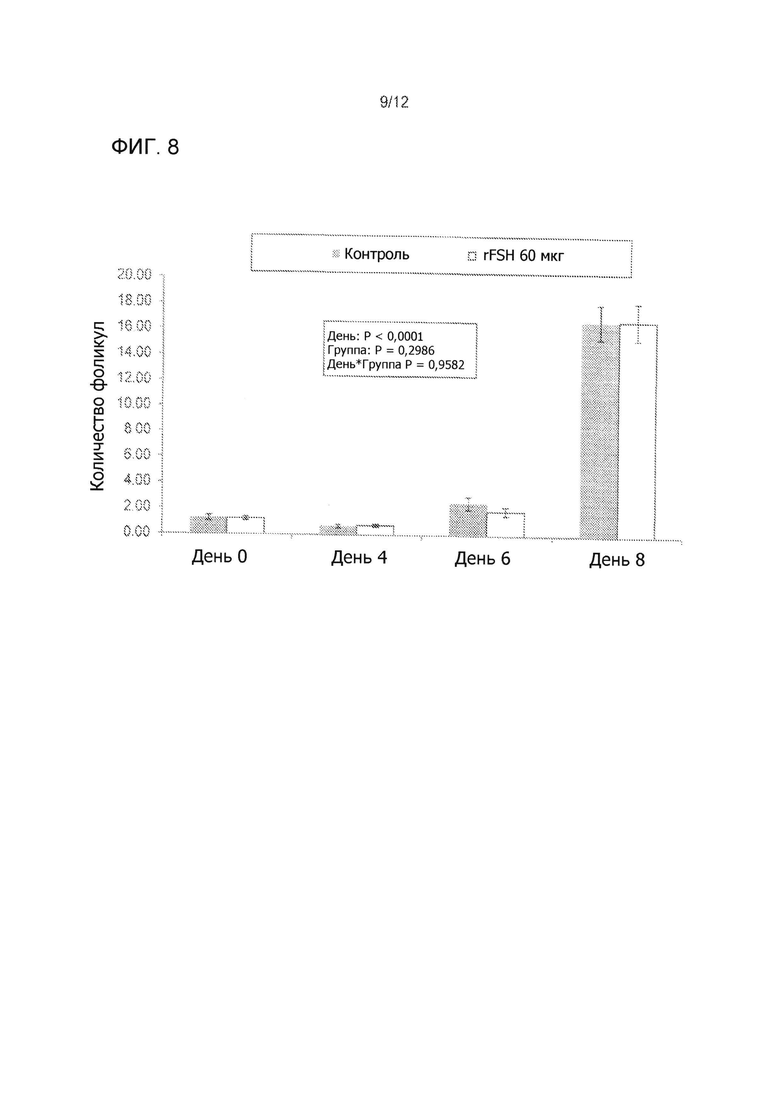
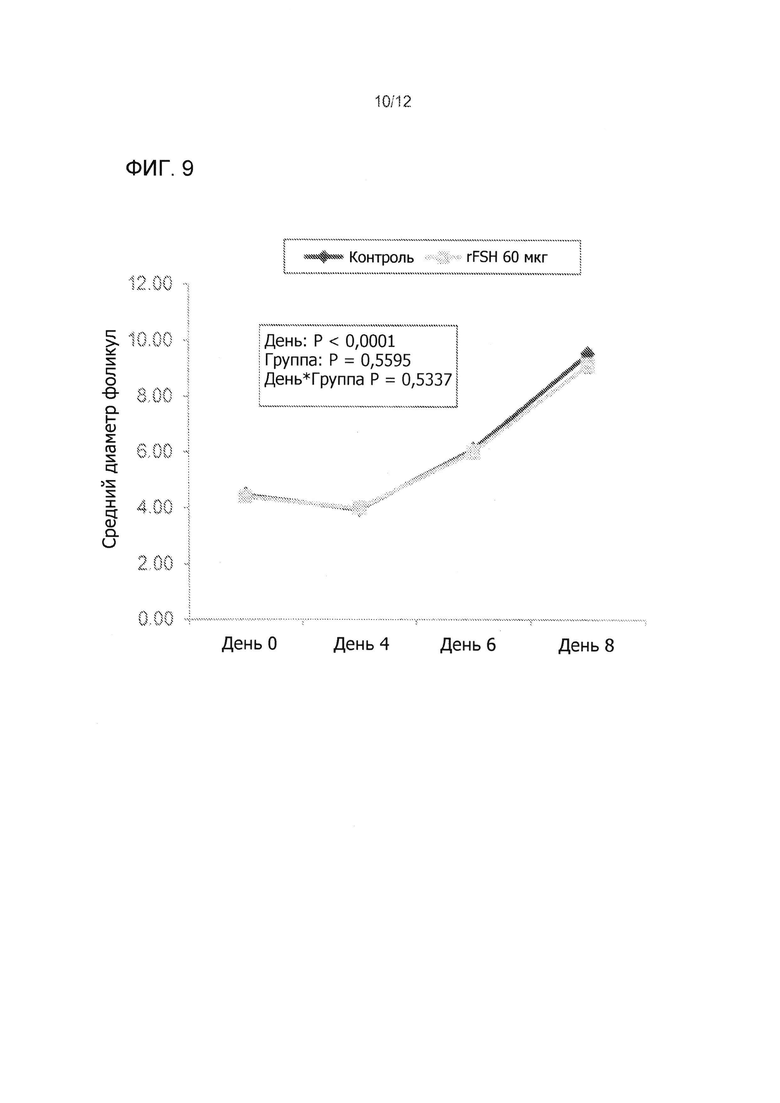
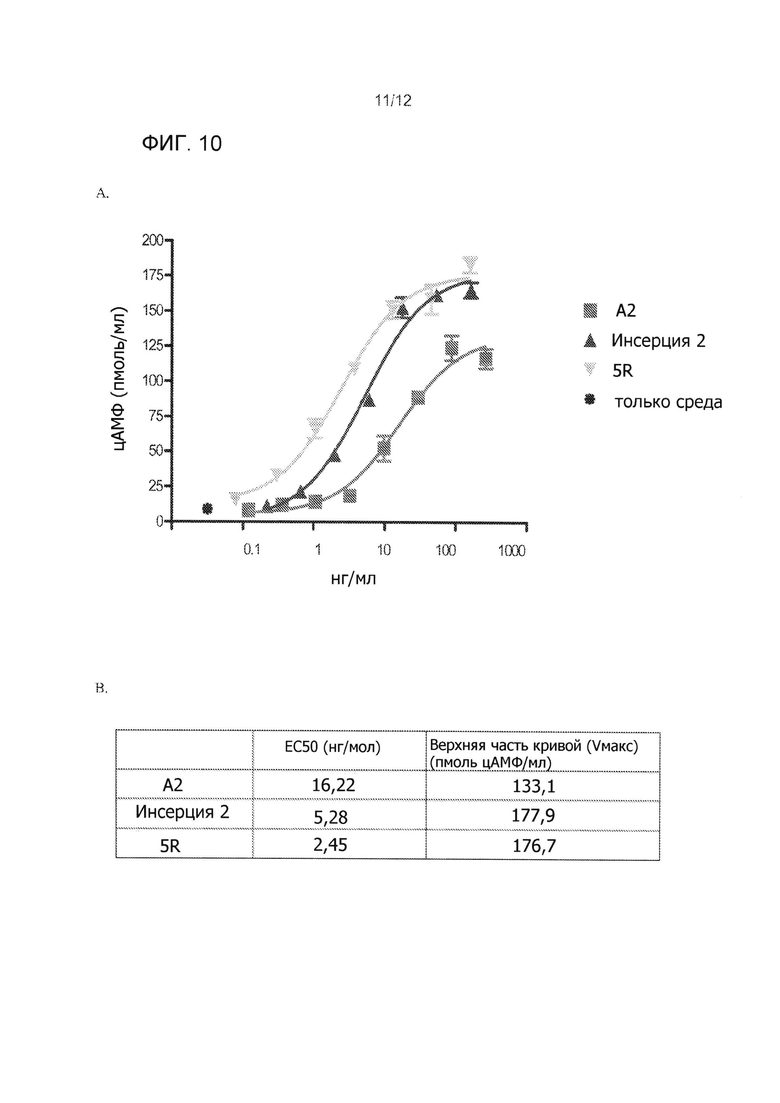
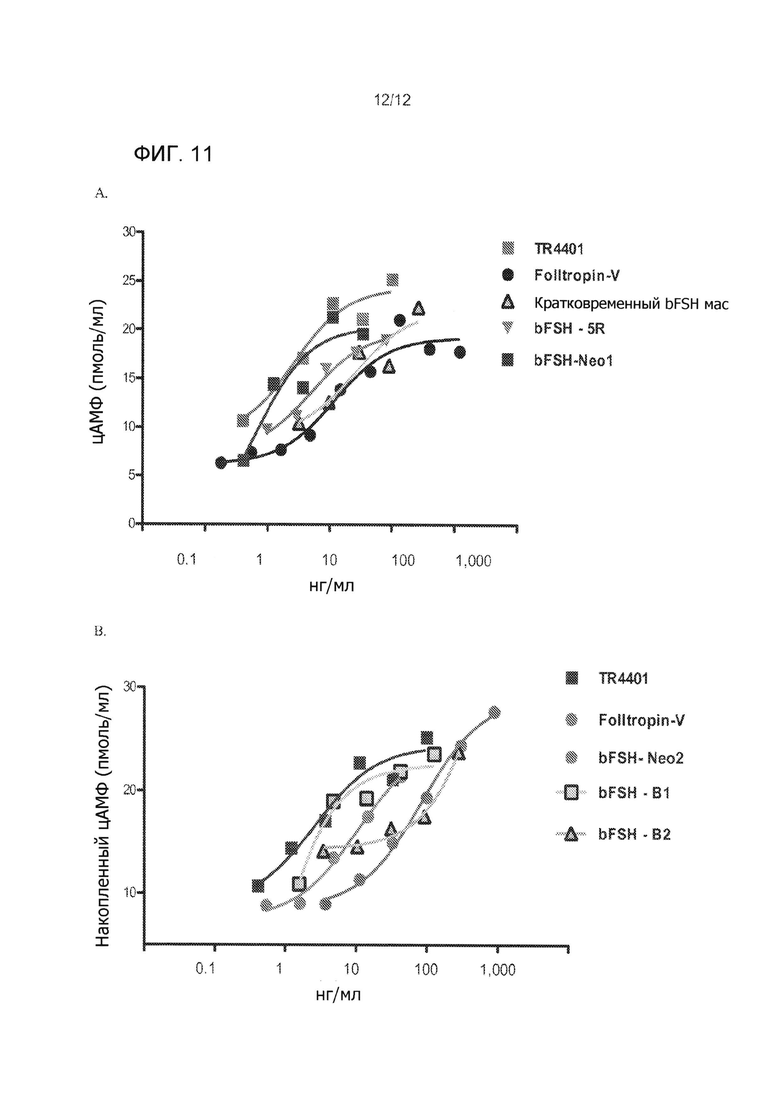
Группа изобретений относится к биотехнологии и медицине, в частности к модифицированным гликопротеиновым гормонам, имеющим активность суперагонистов, и их применению при лечении состояний, связанных с активностью гликопротеинового гормона. Предложен модифицированный полипептид альфа-субъединицы, выбранный из группы, состоящей из дикого типа полипептида альфа-субъединицы крупного рогатого скота (SEQ ID NO: 2), дикого типа полипептида альфа-субъединицы овцы (SEQ ID NO: 3), дикого типа полипептида альфа-субъединицы лошади (SEQ ID NO: 4), дикого типа полипептида альфа-субъединицы свиньи (SEQ ID NO: 5), в который внесены по меньшей мере одно замещение аминокислоты дикого типа на основную аминокислоту в положении 15, 17, 18, 20 и/или 24 и одна, две или три аминокислотные вставки непосредственно после положения 6 или 7. Указанный модифицированный полипептид альфа-субъединицы способен связываться с полипептидом бета-субъединицы с образованием гормона из семейства гликопротеиновых гормонов. Образованный модифицированный гликопротеиновый гормон применяют для стимулирования овуляции. Полученные аналоги гликопротеиновых гормонов длительного действия демонстрируют повышенную биоактивность как in vitro так in vivo по сравнению с аналогом дикого типа, полезны для лечения субъектов, проявляющих низкую экспрессию рецепторов или недостаточный рецепторный ответ, и для лечения любого состояния, связанного с активностью гликопротеинового гормона. 5 н. и 4 з.п. ф-лы, 11 ил., 8 табл.
1. Модифицированный полипептид альфа-субъединицы, способный связываться с полипептидом бета-субъединицы с образованием гормона из семейства гликопротеиновых гормонов, выбранный из группы, состоящей из дикого типа полипептида альфа-субъединицы крупного рогатого скота (SEQ ID NO: 2), дикого типа полипептида альфа-субъединицы овцы (SEQ ID NO: 3), дикого типа полипептида альфа-субъединицы лошади (SEQ ID NO: 4), дикого типа полипептида альфа-субъединицы свиньи (SEQ ID NO: 5), в который внесены следующие модификации:
(A) одна, две или три аминокислотные вставки, что обеспечивает потенциальный сайт гликозилирования, где каждая указанная вставка располагается непосредственно после положения 6 или 7 указанного полипептида альфа-субъединицы, где каждая указанная вставка содержит:
(i) Аспарагин (N);
(ii) Треонин (Т);
(iii) Цистеин (С);
(iv) Аспарагин-Валин (NV);
(v) Треонин-Аспарагин-Валин (TNV);
(vi) Треонин-Изолейцин-Аспарагин-Валин-Треонин (TINVT);
(vii) Аспарагин-Валин-Треонин-Изолейцин-Аспарагин-Валин (NVTINV) (SEQ ID NO:1);
(viii) Аспарагин-Валин-Треонин-Изолейцин-Аспарагин-Валин-Треонин (NVTINVT);
(ix) Треонин-Аспарагин-Валин-Треонин-Изолейцин-Аспарагин-Валин (TNVTINV) (SEQ ID NO: 12); и
(x) Валин-Аспарагин-Валин-Треонин-Изолейцин-Аспарагин-Валин-Треонин (VNVTINVT) (SEQ ID NO: 20);
(B) по меньшей мере одно замещение аминокислоты дикого типа на основную аминокислоту, где указанное замещение выбирают из группы, состоящей из:
(i) аминокислота дикого типа в положении 15 замещается на Аргинин (R),
(ii) аминокислота дикого типа в положении 17 замещается на Аргинин (R),
(iii) аминокислота дикого типа в положении 18 замещается на Аргинин (R), Лизин (K) или Гистидин (Н),
(iv) аминокислота дикого типа в положении 20 замещается на Аргинин (R), и
(v) аминокислота дикого типа в положении 24 замещается на Аргинин (R),
где гормон, образованный указанным модифицированным полипептидом альфа-субъединицы и указанным полипептидом бета-субъединицы, способен связываться и стимулировать рецептор гликопротеиновых гормонов.
2. Модифицированный полипептид альфа-субъединицы по п.1, отличающийся тем, что указанный полипептид альфа-субъединицы содержит одно, два, три, четыре или пять замещений.
3. Модифицированный полипептид альфа-субъединицы по п.1, отличающийся тем, что указанный полипептид альфа-субъединицы содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 7, SEQ ID NO: 8, SEQ ID NO: 9 и SEQ ID NO: 10.
4. Модифицированный гликопротеиновый гормон для стимулирования овуляции, содержащий модифицированный полипептид альфа-субъединицы по п.1 и полипептид бета-субъединицы лютеинизирующего гормона (LH), хорионического гонадотропина (CG), фолликулостимулирующего гормона (FSH) или тироид-стимулирующего гормона (TSH).
5. Модифицированный гликопротеиновый гормон для стимулирования овуляции, содержащий модифицированный полипептид альфа-субъединицы по п.1 и полипептид бета-субъединицы фолликулостимулирующего гормона (FSH).
6. Применение модифицированного гликопротеинового гормона по п.4 или 5 в лечении овуляторной дисфункции, дефектов лютеиновой фазы, необъяснимого бесплодия, мужского бесплодия, ограниченного по времени зачатия, низкой экспрессии рецептора FSH, низкой чувствительности рецептора FSH, дефицита связывания рецептора FSH, дефицита сцепления с рецептором FSH, низкого уровня продуцирования тестостерона, облысения по мужскому типу, и гипофизарной недостаточности или повреждения гипофиза у коровы, овцы, лошади или свиньи.
7. Способ стимулирования овуляции у коровы, овцы, лошади или свиньи, включающий введение указанному животному модифицированного гликопротеинового гормона по п.4 или 5 в эффективном количестве.
8. Модифицированный полипептид альфа-субъединицы по п.1, отличающийся тем, что указанный полипептид альфа-субъединицы представляет собой дикого типа полипептид альфа-субъединицы крупного рогатого скота (SEQ ID NO: 2), содержащий:
(A) полипептидную вставку Аспарагин-Валин-Треонин-Изолейцин-Аспарагин-Валин (NVTINV) (SEQ ID NO: 1) непосредственно после положения 6 указанного полипептида альфа-субъединицы; и
(B) замещения на Аргинин (R) в положениях 15, 17, 18, 20 и 24.
9. Модифицированный полипептид альфа-субъединицы по п.1, отличающийся тем, что указанный полипептид альфа-субъединицы представляет собой последовательность, представленную в SEQ ID NO: 7.
| WO 2005089445 A2, 29.09.2005 | |||
| WO 2012016576 A1, 09.02.2012 | |||
| WO 2011075606 A2, 23.06.2011 | |||
| RU 2006138235 A, 10.05.2008 | |||
| ОДНОЦЕПОЧЕЧНЫЙ БЕЛОК, МОДУЛЯТОР ГОРМОНОВ ЛЮТЕИНИЗИРУЮЩЕГО, ФОЛЛИКУЛОСТИМУЛИРУЮЩЕГО, ТИРЕОСТИМУЛИРУЮЩЕГО И ХОРИОНИЧЕСКОГО ГОНАДОТРОПИНА (ВАРИАНТЫ), МОЛЕКУЛА НУКЛЕИНОВОЙ КИСЛОТЫ (ВАРИАНТЫ), ВЕКТОР (ВАРИАНТЫ), ЛИНИЯ ТРАНСФОРМИРОВАННЫХ КЛЕТОК СНО (ВАРИАНТЫ), СПОСОБ ПРОДУЦИРОВАНИЯ ОДНОЦЕПОЧЕЧНОГО БЕЛКА (ВАРИАНТЫ) | 1995 |
|
RU2213099C2 |

