Область техники.
Изобретение относится к биотехнологии, конкретно к способу получения рекомбинантного хорионического гонадотропина человека (ХГЧ), который может быть использован в медицине, методом рекомбинантных ДНК.
Предшествующий уровень техники.
Хорионический гонадотропин человека (ХГЧ) широко применяется в медицине совместно с фолликулостимулирующим гормоном (ФСГ) при применении вспомогательных репродуктивных технологий (ВРТ) у человека, в качестве финального триггера созревания ооцита в протоколах овариальной стимуляции, проводящихся в клиниках экстракорпорального оплодотворения (ЭКО). У мужчин препараты, содержащие ХГЧ, применяются при гипогонадизме и задержке полового развития, связанных с пониженным синтезом гонадотропинов гипофизом, при крипторхизме без анатомической обструкции и идиопатической диспермии.
На сегодняшний день существует два класса фармацевтических препаратов, содержащих ХГЧ: полученные из мочи беременных женщин и полученные с использованием рекомбинантных ДНК. Препараты ХГЧ мочевого происхождения (представляющие собой ХГЧ, образовавшийся в плаценте и затем выделенный в мочу) появились на рынке в середине ХХ века и на сегодняшний день представлены препаратами с низкой биохимической чистотой ХГЧ, содержащими существенные количества белков, не относящихся к гонадотропинам (Bassett R, et al. Curr Med Res Opin 2005;21:1969-1976). Также на сегодняшний день во всем мире наблюдается дефицит мочевых препаратов как содержащих ХГЧ, так и ФСГ, ввиду возникшей сложности сбора, обработки и процессинга биоматериала, в том числе в условиях пандемии из-за распространения коронавируса SARS-CoV-2. В то же время, применение технологии рекомбинантной ДНК позволяет получать многие биофармацевтические препараты в биореакторах без использования биологических жидкостей человека, что повышает безопасность данного класса препаратов при применении у человека, а также улучшает их молекулярно-биологические характеристики как по показателям чистоты (>99%), так и постоянству биологической активности от партии к партии.
Разработка новых методов и подходов, позволяющих получать бóльшие количества рекомбинантного биологически активного ХГЧ в культивируемых клетках млекопитающих, представляет большой интерес для фармацевтической промышленности вследствие постоянного роста потребления лекарственных препаратов ХГЧ и низкой продуктивности разработанных в конце 80-х годов прошлого века систем экспрессии ХГ, использованных для производства оригинальных препаратов рекомбинантного ХГ. Оптимизация эукариотической системы экспрессии рекомбинантного ХГ, проведённая путём многократного увеличения удельной продуктивности получаемой моноклональной клеточной линии, лежит в основе настоящего изобретения.
ХГЧ представляет собой гетеродимерный гликопротеиновый гормон (34 кДа), состоящий из двух нековалентно связанных друг с другом субъединиц, кодирующихся разными генами. Альфа-субъединица ХГЧ содержит 92 а.о. и также представлена в фолликулостимулирующем, лютеинезирующем и тиреотропном гормонах гипофиза. Альфа-субъединицы этих гормонов подвергаются N-гликозилированию по остаткам аспарагина 52 и 78. Бета-субъединица ХГЧ состоит из 145 а.о. и на 80 % гомологична по структуре бета-субъединице лютеинизирующего гормона. Основное различие между бета-субъединицами ХГЧ и ЛГ заключается в том, что у ХГЧ присутствует 24-аминокислотная последовательность карбоксиконцевого пептида (CTP), О-гликозилированная по остаткам серина 121, 127, 132, 138, в то время как у полипептидного предшественника бета-субъединицы ЛГ CTP отщепляется, и зрелая бета-цепь она имеет длину 121 а.о. Кроме того, бета-субъединицы ХГЧ и ЛГ отличаются и по N-гликозилированию: бета-субъединица ХГЧ подвергается N-гликозилированию по остаткам аспарагина 13 и 30, а в случае ЛГ имеет только один сайт гликозилирования - аспарагин 30.
Как ХГ, так и ЛГ обладают микрогетерогенностью по углеводному компоненту гликопротеинов, то есть существуют в виде смеси изоформ с разным уровнем ветвления олигосахаридных групп и разным содержанием концевых сиаловых кислот, что и предопределяет физико-химические свойства изогормонального профиля ХГЧ. Гетерогенность состава ХГЧ обуславливает фармакокинетические параметры молекулы ХГЧ как на уровне связывания с рецептором ХГЧ, так и на уровне времени полувыведения ХГЧ из кровяного русла. В то же время известно, что уровень сиалирования О-связанных олигосахаридов не влияет на биологические свойства ХГЧ, а уровень сиалирования N-связанных олигосахаридов влияет в очень значительной степени. Изоформы ЛГ имеют изоэлектрическую точку (pI) от 4,0 до 7,2, тогда как pI ХГЧ колеблется от 3 до 7 из-за более высокой кислотности некоторых изоформ ХГЧ. Типичный Z-индекс рекомбинантного ХГЧ, отражающего степень сиалирования, используемого в клинической практике, составляет около 150 (Gervais A, Hammel YA, Pelloux S et al. Glycosylation of human recombinant gonadotrophins: characterization and batch-to-batch consistency. Glycobiology. 2003;13(3):179-89). Согласно исследованию Bousfield et al. (Rev. Endocr. Metab. Disord. (2011) 12:289-302), гипофизарные гонадотропины имеют как α2,3-, так и α2,6-связанную сиаловую кислоту, как и ХГЧ мочевого происхождения. Рекомбинантный ХГЧ, продуцирующийся в клетках яичника китайского хомячка (линия клеток CHO), имеет только α2,3-связанную сиаловую кислоту. Таким образом, рекомбинантный ХГЧ, содержащийся в качестве активного фармацевтического компонента в присутствующих на рынке лекарственных препаратах, содержит молекулы, обладающий только α2,3-связанными сиаловыми кислотами.
На сегодняшний день зарегистрированы в РФ и имеют действующее регистрационное удостоверение следующие лекарственные препараты, содержащие ХГЧ, со следующими международными непатентованными наименованиями (МНН): МНН: хориогонадотропин альфа, препарат Овитрель® (Мерк Сероно С.п.А., Италия); МНН: гонадотропин хорионический, мочевой ХГЧ (ФГУП "Московский эндокринный завод", Россия) и препарат Хорагон® (Ферринг ГмбХ, Германия).
Общим недостатком мочевых препаратов является сниженная удельная биологическая активность из-за наличия неконтролируемых родственных и неродственных примесей в лекарственном препарате, а также неудобная форма выпуска - лиофилизат, требующая восстановления лиофилизированного осадка перед планируемой инъекцией пациентом самостоятельно, что влечет ошибки в постановке дозе и потери действующего вещества.
Лекарственный препарат, содержащий рекомбинантный ХГЧ (хориогонадотропин альфа, Овитрель®) изготавливается в виде раствора в предварительно заполненных шприцах объемом 0,5 мл, содержащих однократную дозу равную 250 мкг (эквивалентно 6500 МЕ) ХГЧ и вспомогательные вещества: маннитол, метионин, полоксамер 188, фосфорная кислота, натрия гидроксид. Данный рекомбинантный ХГЧ продуцируется клетками яичника китайского хомячка (СНО). Овитрель имеет биологическую активность 26000 МЕ/мг (Bassett R, De Bellis C, Chiacchiarini L et. al. Comparative characterization of a commercial human chorionic gonadotrophin extracted from human urine with a commercial recombinant human chorionic gonadotrophin. Curr Med Res Opin. 2005;21(12):1969-76.). Его получение раскрыто в истекшем патенте США US5767251, а фармацевтическая композиция - в другом истекшем патенте США US6706681.
Известен другой способ создания эукариотических продуцентов рекомбинантных гетеродимерных гормонов человека, в том числе ХГЧ, в клетках DUKX CHO (DHFR-minus) (патент США US5240832) c использованием пары плазмид, одна из которых кодирует альфа-субъединицу и открытую рамку считывания DHFR для селективного отбора и амплификации в эукариотических клетках, а вторая - бету-субъединицу и ген орнитин декарбоксилазы (ODC) мыши для селективного отбора. Однако в примерах раскрыто только получение указанным образом продуцентов ФСГ и ТТГ, уровень экспрессии которых был невысок и снижался вследствие использования несплайсированного гена альфа-цепи, интроны которого предположительно отрицательно влияют на экспрессию. В тесте данного патента также отсутствовали количественные данные по уровню биосинтеза биологически активного ХГЧ в такой системе экспрессии генов.
Известны способы повышения биологической активности и продления времени полувыведения рекомбинантного ХГЧ за счёт увеличения его гликозилирования. Описываемая модификация рекомбинантного ХГЧ, продуцируемого в человеческой клеточной линии, имеет более кислый профиль, чем существующий на рынке лекарственный препарат ХГЧ Овитрель® из клеточной линии СНО, благодаря более высокому содержанию сиаловой кислоты вследствие наличия как α-2,3, так и α 2,6-связанной сиаловой кислоты (патенты РФ RU2724528, EA034022). Такой ХГЧ содержит 19,1 моль сиаловой кислоты/моль белка. Среди недостатков данной линии относительно высокий уровень секреции свободной β-субъединицы, что требует введения дополнительных процедур очистки гетеродимера ХГЧ, также не указана продуктивность полученной клеточной линии.
Известен также высокогликозирированный ХГЧ с пониженным содержанием свободной бета-цепи, заявленный в патенте EA034022B1. Для его получения векторами, кодирующими альфа- и бета-субъединицы ХГЧ, модифицируют культивируемые клетки линии PER.C6 или их же с интегрированной в геном кДНК, кодирующей α-2,3-сиалилтрансферазу. В тексте данного патента также отсутствовали количественные данные по уровню промышленного биосинтеза биологически активного ХГЧ в такой системе экспрессии генов.
Известны способы продления времени полувыведения рекомбинантного ХГЧ за счёт его конъюгации с полимерами (патенты CN103619358B, EA009122) или использования фьюжн-партнеров, как слитые в открытой рамке считывания кодирующие последовательности ХГЧ и варианта альфа-1-антитрипсина (патент RU2503688C2). В данных изобретениях не указана продуктивность полученной клеточной линии, не определены физико-химические свойства необходимые для терапевтических белков, а также не определены иммуногенные свойства получаемых химерных белков.
Краткое описание настоящего изобретения
Технической задачей, решаемой авторами, являлось создание высокопродуктивной технологии получения рекомбинантного хорионического гонадотропина человека для биофармацевтического производства.
Технический результат достигался путём создания пар новых взаимно совместимых экспрессионных плазмидных ДНК p1.1-Tr2-HCG-AIB и p1.2-GS-bhCG, p1.1-Tr2-HCG-BIA и p1.2-GS-ahCG, кодирующих открытые рамки считывания альфа- и бета-цепей гетеродимерного белка ХГЧ, и создание на их основе клональных клеточных линий-продуцентов и технологии получения ХГЧ, пригодного для биофармацевтического производства.
В основе данной технологии лежит разработанная пара совместимых плазмидных ДНК p1.1-Tr2-HCG-AIB (длина 8137 пар оснований, SEQ ID NO: 1, Фиг.1) и p1.2-GS-bhCG (длина 13291 пар оснований, SEQ ID NO: 4, Фиг.2), а также пара совместимых плазмидных ДНК p1.1-Tr2-HCG-BIA (длина 8137 пар оснований, SEQ ID NO:5, Фиг.3) и p1.2-GS-ahCG (длина 13144 пар оснований, SEQ ID NO: 6, Фиг.4).
Плазмиды p1.1-Tr2-HCG-AIB и p1.1-Tr2-HCG-BIA различаются только взаиморасположением ОРС альфа- и бета-субъединиц, и состоят из:
фрагмента ДНК длиной 6699 п.о. плазмиды p1.1 (SEQ ID NO: 1 1 - 6700) и (SEQ ID NO: 5 1 - 6700), в следующем порядке содержащего регуляторные элементы, обеспечивающие экспрессию целевого белка:
последовательность (SEQ ID NO: 1 21 - 608) аттенюированного IRES вируса энцефаломиокардита (EMCV), обеспечивающая бицистронную экспрессию в животных клетках и последовательность (SEQ ID NO: 1 621 - 1184) ОРС дигидрофолатредуктазы (DHFR), обеспечивающая устойчивость трансфицированных клеток к воздействию метотрексата,
3’ нетранслируемую область (3’НТО) гена фактора элонгации 1 альфа китайского хомячка (EEF1A1), включающую терминатор и сигнал полиаденилирования этого гена, а также фланкирующие нетранскрибируемые области этого гена, обеспечивающие эухроматинизацию сайтов инсерции и транскрипционную активность генетической кассеты в геноме CHO (SEQ ID NO: 1 1186 - 1800);
область начала репликации плазмиды pUC, открытую рамку считывания бета-лактамазы (bla) и прокариотический промотер гена bla, обеспечивающей устойчивость бактерий к антибиотику ампициллину, позволяющие проводить препаративную наработку плазмиды в E. coli (SEQ ID NO: 1 1965-2638 и 2780-3739), и участок терминального повтора вируса Эпштейн-Барр человека (EBVTR) (SEQ ID NO: 1 3775 - 4176);
5’ нетранслируемую область (5’НТО) гена фактора элонгации 1 альфа китайского хомячка, включающие фланкирующие нетранскрибируемые области этого гена, обеспечивающие эухроматинизацию сайтов инсерции и транскрипционную активность генетической кассеты в геноме CHO, и его функциональный промотор (SEQ ID NO: 1 4186 - 6697).
фрагмента ДНК длиной 1438 п.о., в следующем порядке содержащего:
синтетическую последовательность Козак (SEQ ID NO: 1 6705 - 6713) и (SEQ ID NO: 5 6705 - 6713), обеспечивающую кэп-зависимую инициацию трансляции мРНК,
ОРС субъединиц ХГЧ, разделенных последовательностью внутреннего сайта связывания рибосом вируса энцефаломиокардита EMCV дикого типа; в случае p1.1-Tr2-HCG-AIB - последовательность альфа-субъединицы ХГЧ (SEQ ID NO: 1 6714 - 7061) с блоком стоп-кодонов (SEQ ID NO: 1 7062 - 7067), IRES вируса энцефаломиокардита (EMCV) дикого типа (SEQ ID NO: 1 7074 - 7638), ОРС бета-субъединицы гонадотропного гормона человека (SEQ ID NO: 1 7636 - 8130) с блоком стоп-кодонов (SEQ ID NO: 1 8131 - 8136); в случае и p1.1-Tr2-HCG-BIA: последовательность бета-субъединицы ХГЧ (SEQ ID NO: 5 6714 - 7208) с блоком стоп-кодонов (SEQ ID NO: 5 7209 - 7214), IRES вируса энцефаломиокардита (EMCV) дикого типа (SEQ ID NO: 5 7221 - 7785), ОРС альфа-субъединицы гонадотропного гормона человека (SEQ ID NO: 5 7783 - 8130) с блоком стоп-кодонов (SEQ ID NO: 5 8131 - 8136).
Плазмиды p1.2-GS-ahCG и p1.2-GS-bhCG состоят из:
фрагмента 12775 п.о. плазмиды p1.2-GS (SEQ ID NO: 4 1 - 122775) и (SEQ ID NO: 6 1 - 122775), в следующей последовательности содержащего регуляторные элементы, обеспечивающие экспрессию целевого белка:
3’ нетранслируемую область (3’НТО) гена фактора элонгации 1 альфа китайского хомячка, включающую терминатор и сигнал полиаденилирования этого гена, а также фланкирующие нетранскрибируемые области этого гена, обеспечивающие эухроматинизацию сайтов инсерции и транскрипционную активность генетической кассеты в геноме CHO (SEQ ID NO: 4 1 - 4257);
промотор вируса SV40, последовательность гена для метаболической селекции глутаминсинтазы крысы, сигнал полиаденилирования и терминатор вируса SV40 (SEQ ID NO: 4 4327 - 6331),
область начала репликации плазмиды pUC, открытую рамку считывания бета-лактамазы (bla) и прокариотический промотер гена bla, обеспечивающей устойчивость бактерий к антибиотику ампициллину, позволяющие проводить препаративную наработку плазмиды в E. coli (SEQ ID NO: 4 6480 - 8150),
участок терминального повтора вируса Эпштейн-Барр человека (EBVTR) (SEQ ID NO: 4 8290 - 8691),
5’ нетранслируемую область (5’НТО) гена фактора элонгации 1 альфа китайского хомячка, включающую фланкирующие нетранскрибируемые области этого гена, обеспечивающие эухроматинизацию сайтов инсерции и транскрипционную активность генетической кассеты в геноме CHO функциональный промотор этого гена, а также его функциональный промотор (SEQ ID NO: 4 8703 - 12773).
фрагмента ДНК, длиной 516 п.о., в следующем порядке содержащего синтетическую последовательность Козак (SEQ ID NO: 1 12781-12789), ОРС цепи бета-субъединицы гонадотропного гормона человека (SEQ ID NO: 1 12790-13284) и блок стоп-кодонов (SEQ ID NO: 1 13285-13290) - в случае p1.2-GS-bhCG, и фрагмента ДНК, длиной 363 п.о., в следующем порядке содержащего синтетическую последовательность Козак (SEQ ID NO: 6 12781-12789), ОРС цепи альфа-субъединицы гонадотропного гормона человека (SEQ ID NO: 6 12790-13137) и блок стоп-кодонов (SEQ ID NO: 6 13138-13143) - в случае p1.2-GS-ahCG.
Плазмиды были последовательно введены методом электротрансфекции (Straus S.E. et al, J. Virol., 1981, Jul; 39(1):290-4) в клетки CHO (Invitrogen cGMP banked, США; Mol Cell Biol 5, 1750-1759 Kaufman RJ et al. 1985), адаптированные к суспензионному культивированию в бессывороточной ростовой среде. В случае плазмиды p1.1-Tr2-HCG-AIB использовали клетки линии CHO DG44, в случае плазмиды p1.1-Tr2-HCG-BIA использовали клетки CHO S. После трансфекции клеток указанными плазмидами вели культивацию в безбелковой среде, не содержащей гипоксантин и тимидин, и дополнительно содержащей 0,2 мкМ ингибитора DHFR метотрексата (MTX) до восстановления жизнеспособности клеток более 90%. После этого была проведена амплификация целевых генов цепей ХГЧ в геноме путём последовательных культивирований в присутствии возрастающих концентраций MTX, максимальная концентрация MTX 8 мкМ. Была получена поликлональная популяция клеток, устойчивых к высокой концентрации МТХ и продуцирующих увеличенные количества ХГЧ и значительные количества свободной цепи ХГ, ОРС которой расположен в первом цистроне экспрессионной кассеты (альфа-цепи в случае p1.1-Tr2-HCG-AIB, бета-цепи в случае p1.1-Tr2-HCG-BIA). Клетки полученных популяций соответственно трансфицировали плазмидами p1.2-GS-bhCG и p1.2-GS-ahCG, после чего культивировали в присутствии 25 мкМ метионилсульфоксимина (MSX) до восстановления жизнеспособности клеток более 90%. Были получены популяции клеток, секретирующих бóльшие количества гетеродимера ХГЧ. Клетки данных популяций подвергли клонированию методом предельных разведений. Полученные клоны клеток-продуцентов были проанализированы методом иммуноферментного анализа и были отобраны клоны, дающие максимальный уровень экспрессии гетеродимера ХГЧ. Среди них были выбраны клоны D-pTr2-hCG-55 и S-pTr2-hCG-8, при культивировании которых в суспензионной культуре в бессывороточной среде конечная концентрация гонадотропного гормона человека составляла: 5,4 мкг/мл для D-pTr2-hCG-55 и 6,3 мкг/мл для S-pTr2-hCG-8. Полученные клеточные линии депонированы во Всероссийской Коллекции Промышленных Микроорганизмов (ВКПМ): D-pTr2-hCG-55 - под регистрационным номером ВКПМ Н-194, S-pTr2-hCG-8 - под регистрационным номером ВКПМ Н-213.
Целью настоящего изобретения является предоставление плазмид для экспрессии рекомбинантного гонадотропного гормона человека, в частности, плазмид p1.1-Tr2-HCG-AIB, p1.1-Tr2-HCG-BIA, p1.2-GS-ahCG и p1.2-GS-bhCG. Также целью настоящего изобретения является предоставление моноклональных линий клеток млекопитающего, в частности, клеток яичника китайского хомячка, - продуцентов рекомбинантного гонадотропного гормона человека, содержащих в геноме множественные копии экспрессионных кассет, соответствующих линеаризованным экспрессионным плазмидам, в частности, плазмидам p1.1-Tr2-HCG-AIB, p1.1-Tr2-HCG-BIA, p1.2-GS-ahCG и p1.2-GS-bhCG, в частности, их комбинациями p1.1-Tr2-HCG-AIB и p1.2-GS-bhCG, а также p1.1-Tr2-HCG-BIA и p1.2-GS-ahCG. Иллюстративными примерами указанных моноклональных линий клеток являются клетки яичника китайского хомячка клонов D-pTr2-hCG-55 и S-pTr2-hCG-8.
Также целью настоящего изобретения является предоставление способа получения рекомбинантного ХГЧ, включающий культивирование описанных выше линий клеток в питательной среде и выделение полученного ХГЧ из культуральной жидкости.
Также целью настоящего изобретения является предоставление способа получения рекомбинантного ХГЧ, в котором выделение полученного ХГЧ из культуральной жидкости осуществляют центрифугированием культуральной жидкости для отделения клеток и дебриса с последующим выделением целевого ХГЧ с помощью хроматографии. Указанный способ позволяет производить очистку целевого ХГЧ от родственных примесей, белков линии-продуцента и ДНК линии-продуцента до фармакопейных требований.
Также целью настоящего изобретения является предоставление описанного выше способа, в котором культивируют линии-продуценты ХГЧ D-pTr2-hCG-55 или S-pTr2-hCG-8.
Подробное описание настоящего изобретения
Для реализации настоящего изобретения главной технической задачей явилось создание технологии высокопродуктивного получения рекомбинантного хорионического гонадотропина человека для биофармацевтического производства с использованием культивируемых клеток яичника китайского хомячка, адаптированных к суспензионному культивированию в безбелковой среде, содержащих в геноме множественные копии генетических кассет, представляющих собой линеаризованные экспрессионные плазмиды, содержащих фрагмент ДНК, кодирующих альфа и бета цепи гонадотропного гормона человека под контролем промотора и регуляторных элементов, функционирующих в эукариотической клетке.
Термин «экспрессионная плазмида» («экспрессионная плазмидная ДНК») означает плазмидную ДНК, содержащую все необходимые генетические элементы для экспрессии внедрённого в него гена, такие как промотор, терминатор, сигнал полиаденилирования. Конкретным примером генетических элементов, необходимых для экспрессии рекомбинантного хорионического гонадотропина человека в составе экспрессионной кассеты, согласно настоящему изобретению, является, но не ограничивается им, промотор гена фактора элонгации 1 альфа китайского хомячка.
Фрагментами ДНК, кодирующими рекомбинантный хорионический гонадотропин человека, согласно настоящему изобретению, являются гены, кодирующие ОРС полипептидов альфа- и бета-цепей ХГ, которые могут быть получены, например, как указано в Примере 1. Также указанные фрагменты ДНК могут быть получены, например, с использованием технологии клонирования фирмы SloningBioTechnology, описанной в заявке РСТ WO2005071077.
Последовательность искусственного гена, кодирующего ОРС обеих субъединиц рекомбинантного хорионического гонадотропина человека, согласно настоящему изобретению, представлена в Перечне последовательностей под номером SEQ ID NO: 1 (цепь альфа-субъединицы - нуклеотиды 6714 - 7061, цепь бета-субъединицы - нуклеотиды 7636 - 8130). Аминокислотная последовательность секретируемого гонадотропного гормона человека, согласно настоящему изобретению представлена, в Перечне последовательностей под номерами SEQ ID NO: 2 и 3, и представляет собой продукт трансляции нуклеотидов 6714 - 7061 и 7636 - 8130 в последовательности ОРС SEQ ID NO: 1, включая 24 аминокислоты N-концевого сигнального пептида альфа-цепи (Mdyyrkyaaiflvtlsvflhvlhs) и 20 аминокислот (MEMFQGLLLLLLLSMGGTWA) N-концевого сигнального пептида бета-цепи, отделяющихся при пост-трансляционном процессинге цепей ХГЧ и отсутствующих в зрелом секретированном гетеродимерном белке. Часть молекул цепей ХГЧ может быть секретирована в свободной форме, для целей настоящего изобретения важно накопление только двуцепочечной формы гетеродимера цепей ХГЧ в культуральной среде.
Чтобы обеспечить эффективную трансляцию гена в клетках китайского хомячка, предпочтительно, чтобы ОРС предварялась последовательностью для кэп-зависимой инициации трансляции (последовательность Козак), например, синтетической. Для кэп-независимой инициации трансляции предпочтительно использование вирусных регуляторных элементов класса внутренних сайтов связывания рибосом (IRES).
Фрагменты ДНК, которые кодируют по существу тот же белок, могут быть получены, например, путём модификации нуклеотидной последовательности фрагмента ДНК SEQ ID NO: 1 (цепь альфа - нуклеотиды 6714 - 7061, цепь бета - нуклеотиды 7636 - 8130), например, посредством метода сайт-направленного мутагенеза таким образом, что один или несколько аминокислотных остатков в определенном сайте будут делетированы, заменены, вставлены или добавлены. Фрагменты ДНК, модифицированные, как описано выше, могут быть получены с помощью традиционных методов обработки с целью получения мутации. Фрагменты ДНК, которые кодируют по существу такой же функциональный белок, могут быть выявлены путём экспрессии фрагментов ДНК, имеющих мутацию, описанную выше, в соответствующей клетке и установления активности экспрессируемого продукта.
ХГЧ - гетеродимерный гликопротеиновый гормон (массой около 46 кДа), продуцируемый трофобластом плаценты, состоящий из двух нековалентно связанных друг с другом субъединиц, кодирующихся разными генами (PierceJ.G., ParsonsT.F. (1981), "Glycoproteinhormones: structureandfunction". Annu.Rev. Biochem.,50: 465-495).
Показатели функциональной активности, при которой считается, что полученный белок обладает свойствами рекомбинантного гонадотропного гормона человека, определяются по его способности увеличивать массу комплекса добавочных половых желёз у крыс-самцов (семенных пузырьков и предстательной железы) или увеличивать массу матки у крыс-самок, состоящей из тела и двух рогов, после трех последовательных инъекций по 1 разу в стуки. Исследования проводят в сравнении со стандартом ХГЧ (ГФ РФ XIII). Концентрацию гетеродимерного ХГЧ определяют при помощи иммуноферментного анализа в сравнении со стандартом, в качестве которого можно использовать образцы плазмы крови женщин, калиброванные против первичного международного стандарта ХГ, изготовляемого Всемирной Организацией Здравоохранения, например, стандарта hCG 99/688 (https://doi.org/10.1373/49.1.144). Удельную активность ХГЧ определяют как отношение активности и концентрации ХГЧ и выражают в МЕ/мг. Считается, что вариант белка обладает свойствами рекомбинантного хорионического гонадотропина человека при условии, что удельная активность указанного варианта составляет не ниже 10 % от удельной активности очищенного природного хорионического гонадотропина человека, выделяемого из мочи с типичной удельной активностью 2500 МЕ/мг, то есть не менее 250 МЕ/мг.
Экспрессионная плазмида, согласно настоящему изобретению, содержит фрагмент ДНК, кодирующий цепи альфа- и бета-субъединиц ХГЧ или индивидуальную субъединицу ХГЧ под контролем промотора и регуляторных элементов, функционирующих в эукариотической клетке. В качестве рекомбинантной плазмиды, согласно настоящему изобретению, могут использоваться различные плазмиды, обладающие способностью к экспрессии в клетке-реципиенте, такие как плазмиды pcDNA3.1, pCMV-Myc, pDEF38, pCHO и подобные им, но список плазмид не ограничивается ими.
Пара совместимых плазмид, согласно настоящему изобретению, означает две такие экспрессионные плазмиды, которые могут быть использованы для одновременной или последовательной трансфекции культивируемых клеток млекопитающих и последующей селекции стабильно трансфицированных клеток. Необходимым условием совместимости векторов является наличие в их составе различных генов устойчивости к действию селекционных агентов, сообщающих трансфицированным клеткам устойчивость к действию только одного из нескольких используемых селекционных агентов. Экспрессионные плазмиды из пары совместимых плазмид могут кодировать ген, содержащий ОРС одной или обеих цепей ХГ.
Конкретным вариантом реализации настоящего изобретения являются пары совместимых плазмид:
пара I: p1.1-Tr2-HCG-AIB (длина 8137 пар оснований, SEQ ID NO: 1, Фиг. 1) и p1.2-GS-bhCG (длина 13291 пар оснований, SEQ ID NO: 4, Фиг. 2),
пара II: p1.1-Tr2-HCG-BIA (длина 8137 пар оснований, SEQ ID NO: 5, Фиг. 3) и p1.2-GS-ahCG (длина 13144 пар оснований, SEQ ID NO: 6, Фиг. 4).
Данные плазмиды содержат следующие функциональные элементы, перечисленные в порядке их расположения:
Для плазмиды p1.1-Tr2-HCG-AIB (SEQ ID NO: 1, Фиг. 1):
область начала репликации плазмиды pUC (1965-2638), открытую рамку считывания бета-лактамазы (bla) (2780-3640) и прокариотический промотер гена bla (3635-3739), позволяющие проводить препаративную наработку плазмиды в E. coli;
участок терминального повтора вируса Эпштейн-Барр человека (EBVTR), представляющий собой фрагмент конкатемера терминальных повторов вируса Эпштейна-Барр (3775-4176), обеспечивающей увеличение частоты интеграции генетической кассеты в геном клеток CHO и увеличение скорости амплификации кассеты в геноме;
фрагмент последовательности, фланкирующей ген фактора элонгации 1 альфа китайского хомячка «выше по течению» (4186-6697), включающий нетранскрибируемые области этого гена, промотор этого гена, первый интрон (5742-6685) этого гена, обеспечивающий конститутивную экспрессию гена гонадотропного гормона человека в геноме клеток CHO;
синтетическую последовательность Козак (сайт связывания рибосом) (6705-6713), обеспечивающую кэп-зависимую инициацию трансляции мРНК в животных клетках;
последовательность, кодирующую открытую рамку считывания альфа-субъединицы ХГЧ (6714-7061) с блоком стоп-кодонов (7062-7067);
последовательность внутреннего сайта связывания рибосом вируса энцефаломиокардита EMCV дикого типа (7074-7638), обеспечивающая кэп-независимую инициацию трансляции полицистронных РНК в животных клетках;
последовательность, кодирующую открытую рамку считывания бета-субъединицы ХГЧ (7636-8130) с блоком стоп-кодонов (8131-8136);
последовательность аттенюированного внутреннего сайта связывания рибосом вируса энцефаломиокардита EMCV (21-608), обеспечивающую кэп-независимую инициацию трансляции полицистронных РНК в животных клетках;
последовательность, кодирующую открытую рамку считывания дигидрофолатредуктазы (621-1184) - фактора устойчивости трансфицированных клеток к воздействию метотрексата;
фрагмент последовательности, фланкирующей ген фактора элонгации 1 альфа китайского хомячка «ниже по течению» (1186-1800), включающий терминатор и сигнал полиаденилирования, а также нетранскрибируемые области этого гена, обеспечивающий конститутивную экспрессию гена ХГЧ в геноме клеток CHO.
Для плазмиды p1.1-Tr2-HCG-BIA (SEQ ID NO: 5, Фиг. 3):
область начала репликации плазмиды pUC (1965-2638), открытую рамку считывания бета-лактамазы (bla) (2780-3640) и прокариотический промотер гена bla (3635-3739), позволяющие проводить препаративную наработку плазмиды в E. coli;
участок терминального повтора вируса Эпштейн-Барр человека (EBVTR), представляющий собой фрагмент конкатемера терминальных повторов вируса Эпштейна-Барр (3775-4176), обеспечивающей увеличение частоты интеграции генетической кассеты в геном клеток CHO и увеличение скорости амплификации кассеты в геноме;
фрагмент последовательности, фланкирующей ген фактора элонгации 1 альфа китайского хомячка «выше по течению» (4186-6697), включающий нетранскрибируемые области этого гена, промотор этого гена, первый интрон (5742-6685) этого гена, обеспечивающий конститутивную экспрессию гена гонадотропного гормона человека в геноме клеток CHO;
синтетическую последовательность Козак (сайт связывания рибосом) (6705-6713), обеспечивающую кэп-зависимую инициацию трансляции мРНК в животных клетках;
последовательность, кодирующую открытую рамку считывания бета-субъединицы ХГЧ (6714 - 7208) с блоком стоп-кодонов (7209 - 7214);
последовательность внутреннего сайта связывания рибосом вируса энцефаломиокардита EMCV дикого типа (7221-7785), обеспечивающая кэп-независимую инициацию трансляции полицистронных РНК в животных клетках;
последовательность, кодирующую открытую рамку считывания альфа-субъединицы ХГЧ (7783-8130) с блоком стоп-кодонов (8131-8136);
последовательность аттенюированного внутреннего сайта связывания рибосом вируса энцефаломиокардита EMCV (21-608), обеспечивающую кэп-независимую инициацию трансляции полицистронных РНК в животных клетках;
последовательность, кодирующую открытую рамку считывания дигидрофолатредуктазы (621-1184) - фактора устойчивости трансфицированных клеток к воздействию метотрексата;
фрагмент последовательности, фланкирующей ген фактора элонгации 1 альфа китайского хомячка «ниже по течению» (1186-1800), включающий терминатор и сигнал полиаденилирования, а также нетранскрибируемые области этого гена, обеспечивающий конститутивную экспрессию гена ХГЧ в геноме клеток CHO.
Для плазмиды p1.2-GS-ahCG (SEQ ID NO: 4, Фиг. 2):
область начала репликации плазмиды pUC (6480-7153), открытую рамку считывания бета-лактамазы (bla) (7295-8155) и прокариотический промотер гена bla (8150 - 8254), позволяющие проводить препаративную наработку плазмиды в E. coli;
участок терминального повтора вируса Эпштейн-Барр человека (EBVTR), представляющий собой фрагмент конкатемера терминальных повторов вируса Эпштейна-Барр (8290-8691), обеспечивающей увеличение частоты интеграции генетической кассеты в геном клеток CHO и увеличение скорости амплификации кассеты в геноме;
фрагмент последовательности, фланкирующей ген фактора элонгации 1 альфа китайского хомячка «выше по течению» (8703-12773), включающий нетранскрибируемые области этого гена, промотор этого гена, первый интрон этого гена, обеспечивающий конститутивную экспрессию гена бета-цепи ХГЧ в геноме клеток CHO;
синтетическую последовательность Козак (сайт связывания рибосом) (12781-12789), обеспечивающую кэп-зависимую инициацию трансляции мРНК в животных клетках;
последовательность, кодирующую открытую рамку считывания альфа-цепи гонадотропного гормона человека (12790-13284) с блоком стоп-кодонов (13285-13290);
фрагмент последовательности, фланкирующей ген фактора элонгации 1 альфа китайского хомячка «ниже по течению» (1-4257), включающий терминатор и сигнал полиаденилирования, а также нетранскрибируемые области этого гена, обеспечивающий конститутивную экспрессию гена бета-субъединицы гонадотропного гормона человека в геноме клеток CHO.
последовательность для метаболической селекции трансфицированных клеток - ген глутаминсинтазы крысы (4668-5789) и регуляторные элементы для его экспрессии - промотор ранних генов (4327-4651), сигнал полиаденилирования и терминатор вируса SV40 (6196-6331).
Для плазмиды p1.2-GS-bhCG (SEQ ID NO: 6, Фиг. 4):
область начала репликации плазмиды pUC (6480-7153), открытую рамку считывания бета-лактамазы (bla) (7295-8155) и прокариотический промотер гена bla (8150 - 8254), позволяющие проводить препаративную наработку плазмиды в E. coli;
участок терминального повтора вируса Эпштейн-Барр человека (EBVTR), представляющий собой фрагмент конкатемера терминальных повторов вируса Эпштейна-Барр (8290-8691), обеспечивающей увеличение частоты интеграции генетической кассеты в геном клеток CHO и увеличение скорости амплификации кассеты в геноме;
фрагмент последовательности, фланкирующей ген фактора элонгации 1 альфа китайского хомячка «выше по течению» (8703-12773), включающий нетранскрибируемые области этого гена, промотор этого гена, первый интрон этого гена, обеспечивающий конститутивную экспрессию гена бета-цепи ХГЧ в геноме клеток CHO;
синтетическую последовательность Козак (сайт связывания рибосом) (12781-12789), обеспечивающую кэп-зависимую инициацию трансляции мРНК в животных клетках;
Последовательность, кодирующую открытую рамку считывания альфа-цепи гонадотропного гормона человека (12790-13137) с блоком стоп-кодонов (13138-13143);
фрагмент последовательности, фланкирующей ген фактора элонгации 1 альфа китайского хомячка «ниже по течению» (1-4257), включающий терминатор и сигнал полиаденилирования, а также нетранскрибируемые области этого гена, обеспечивающий конститутивную экспрессию гена бета-субъединицы гонадотропного гормона человека в геноме клеток CHO.
последовательность для метаболической селекции трансфицированных клеток - ген глутаминсинтазы крысы (4668-5789) и регуляторные элементы для его экспрессии - промотор ранних генов (4327-4651), сигнал полиаденилирования и терминатор вируса SV40 (6196-6331).
Плазмида p1.1-Tr2-HCG-AIB имеет размер 8137 п.о. и содержит уникальные сайты узнавания эндонуклеазами рестрикции NheI (1), AscI (1831), PvuI (3224), EcoRI (4183), AbsI (6700), XbaI (6886).
Плазмида p1.1-Tr2-HCG-BIA имеет размер 8137 п.о. и содержит уникальные сайты узнавания эндонуклеазами рестрикции NheI (1), AscI (1831), PvuI (3224), EcoRI (4183), AbsI (6700), XbaI (7955).
Плазмида p1.2-GS-bhCG имеет размер 13291 п.о. и содержит уникальные сайты узнавания эндонуклеазами рестрикции PvuI (7739), AbsI (12776).
Плазмида p1.2-GS-ahCG имеет размер 13144 п.о. и содержит уникальные сайты узнавания эндонуклеазами рестрикции PvuI (7739), AbsI (12776), XbaI (12962).
Структура плазмиды p1.1-Tr2-HCG-AIB приведена на Фигуре 1.
Структура плазмиды p1.2-GS-bhCG приведена на Фигуре 2.
Структура плазмиды p1.1-Tr2-HCG-BIA приведена на Фигуре 3.
Структура плазмиды p1.2-GS-ahCG приведена на Фигуре 4.
Плазмиды p1.1-Tr2-HCG-AIB и p1.1-Tr2-HCG-BIA сконструированы на базе экспрессионной плазмиды р1.1, подробно описанной в патенте РФ 2488633 (Фиг. 2 в патенте РФ 2488633). Плазмида р1.1 содержит функциональные промотор и терминатор гена фактора элонгации 1 альфа китайского хомячка, обеспечивающие конститутивную экспрессию целевого гена в клетках CHO, фланкированные 5’ и 3’ НТО этого гена, обеспечивающими эухроматинизацию сайтов интеграции экспрессионной кассеты в геном CHO; участок для клонирования открытых рамок считывания целевых белков; внутренний сайт связывания рибосом вируса энцефаломиокардита (EMCV IRES), обеспечивающий реинициацию трансляции на бицистронной мРНК в клетках млекопитающих и полное генетическое сцепление уровней продукции целевого белка и DHFR; ОРС DHFR мыши, экспрессирующуюся в составе бицистронной мРНК вместе с целевым геном (IRES DHFR) и обеспечивающую устойчивость стабильно трансфицированных клеток к отсутствию в среде тимидина и дозозависимую устойчивость к воздействию метотрексата (MTX), что позволяет вести направленную селекцию высокопродуктивных клонов с множественными копиями экспрессионной кассеты в геноме CHO; участок терминального повтора вируса Эпштейн-Барр человека (EBVTR), обеспечивающий повышенный уровень интеграции кассет в геном клеток CHO и ускорение процесса амплификации амплификации целевого гена в геноме под действием MTX. Использование плазмиды р1.1 позволяет осуществлять высокочастотную интеграцию и ускоренную амплификацию экспрессионной кассеты в клетках млекопитающих, а также получать линии-продуценты с высоким уровнем продукции целевого белка и высокой стабильностью.
Плазмиды p1.2-GS-ahCG и p1.2-GS-bhCG сконструированы на базе экспрессионной плазмиды p1.2 для моноцистронной экспрессии, подробно описанной в патенте РФ 2552170 (в патенте РФ 2552170 плазмида представлена в SEQ ID: 1 и на Фиг. 2). Плазмида р1.2 содержит функциональные промотор и терминатор гена фактора элонгации 1 альфа китайского хомячка, обеспечивающие конститутивную экспрессию целевого гена в клетках CHO, фланкированные 5’ и 3’ НТО этого гена, обеспечивающими эухроматинизацию сайтов интеграции экспрессионной кассеты в геном CHO; участок для клонирования открытых рамок считывания целевых белков; участок терминального повтора вируса Эпштейн-Барр человека (EBVTR), обеспечивающий повышенный уровень интеграции кассет в геном клеток CHO и кассету, обеспечивающую экспрессию селекционного маркера гена устойчивости к антибиотику, под контролем генетических элементов для экспрессии в клетках млекопитающего. Использование плазмиды р1.2 позволяет осуществлять высокочастотную интеграцию экспрессионной кассеты в клетках млекопитающих, а также получать линии-продуценты с высоким уровнем продукции целевого белка при одновременной или последовательной трансфекции культивируемых клеток млекопитающих плазмидами на основе p1.1 и p1.2 и последующей селекции стабильно трансфицированных клеток.
При помощи созданных плазмид, согласно настоящему изобретению, в частности, пары плазмид p1.1-Tr2-HCG-AIB и p1.2-GS-bhCG или пары плазмид p1.1-Tr2-HCG-BIA и p1.2-GS-ahCG, можно трансфицировать эукариотическую клетку, предпочтительно иммортализованную клетку яичника китайского хомячка. Выбор конкретной линии клеток не является критическим, поскольку методология и приёмы трансфекции хорошо известны специалисту в данной области техники. После проведения одного или нескольких раундов селекции и клонирования популяции трансфицированных клеток могут быть получены клональные линии-продуценты рекомбинантного ХГЧ человека. Методология и приёмы селекции и клонирования клеток известны специалисту в данной области техники. И хотя в зависимости от вида клетки и условий культивирования и селекции трансфектантов уровень экспрессии ХГЧ может варьироваться, факт транзиентной экспрессии целевого белка будет иметь место при условии успешной трансфекции клетки-реципиента, факт стабильной экспрессии целевого белка будет иметь место при успешном проведении раунда селекции популяции клеток-трансформантов.
«Трансфекция клетки плазмидой» означает введение плазмиды в клетку с помощью методов, хорошо известных специалисту в данной области техники. Методы трансфекции включают любые стандартные методы, известные специалисту в данной области техники, например электропорацию, использование трансфекционных агентов, например, метод, описанный в техническом документе компании Invitrogen, Inc. “Lipofectamine® 2000 Reagent” Pub. No. MAN0000995 Rev. Date 20 July 2012.
Согласно настоящему изобретению, «клеточная линия-продуцент хорионического гонадотропина человека» означает клональную линию клеток млекопитающих, обладающую способностью к продукции и секреции ХГЧ, когда она согласно настоящему изобретению выращивается в указанной питательной среде. Используемый здесь термин «клеточная линия-продуцент хорионического гонадотропина человека» также означает линию клеток, которые способна секретировать хорионический гонадотропин человека в количестве не менее, чем 1 мкг/мл при концентрации клеток 1-2 млн/мл, более предпочтительно, не менее, чем 5 мкг/мл при концентрации клеток 1-4 млн/мл в течение трех дней культивирования в суспензионной культуре в бессывороточной среде. Указанный хорионический гонадотропин человека секретируется указанными клетками в культуральную жидкость.
Среди клеток млекопитающих предпочтительно использование клеток китайского хомячка (Cricetulus griseus), предпочтительно клеток яичника (CHO). В качестве примера предпочтительных клеток яичника китайского хомячка могут быть приведены линия CHO DG44 и линия CHO S (Invitrogen cGMP banked, США; Mol Cell Biol 5, 1750-1759 Kaufman RJ et al. 1985). Круг сублиний CHO не ограничен каким-либо образом, например, могут быть использованы сублинии CHO CHOZN DHFR, CHO DUKX B11 и подобные им.
Конкретным примером линии для получения продуцента гонадотропного гормона человека согласно настоящему изобретению являются линия CHO DG44 и CHO S, но спектр линий клеток не ограничиваются только этими линиями.
Линия CHO DG44 характеризуется следующими культурально-морфологическими, физиолого-биохимическими и генетическими признаками:
Данные по видовой принадлежности: китайский хомячок Cricetulus griseus, яичник.
Контроль видовой идентичности: кариологический, изоферментный (ЛДГ и Г6ФДГ).
Маркерные признаки и методы их оценки: иммунологические, цитогенетические, биохимические, физиологические.
Морфология: эпителиоподобная.
Кариология: 2n=22, пределы изменчивости по числу хромосом 10-28, модальное число хромосом 22, псевдодиплоид, имеются микрохромосомы, количество полиплоидов 9,0%.
Признаки опухолевого роста in vivo, in vitro: отсутствуют.
Онкогены: отсутствуют.
Морфология клеточной линии: при анализе в световой микроскоп, культура представлена суспензией круглых клеток с овальным ядром, содержащих небольшие эндосомы, время удвоения 28 часов.
Культуральные свойства, маркерные признаки: способ культивирования - суспензионный, посевная доза - 300 000 на мл., кратность рассева - 1:2 - 1:6, процедура пересева - центрифугирование суспензии на скорости 1200 об/мин, 5 мин., ресуспендирование в новой ростовой среде. Оптимальная плотность 1,0-1,2х106 клеток/мл. Клетки адаптированы к росту в культуральной среде без добавления сыворотки.
Рекомендуемые условия для замораживания: при плотности ≥ 2,0х106 живых клеток/мл; ростовая среда, 10% ДМСО
Линия CHO S характеризуется следующими культурально-морфологическими, физиолого-биохимическими и генетическими признаками:
Данные по видовой принадлежности: китайский хомячок Cricetulus griseus, яичник.
Контроль видовой идентичности: кариологический, изоферментный (ЛДГ и Г6ФДГ).
Маркерные признаки и методы их оценки: иммунологические, цитогенетические, биохимические, физиологические.
Морфология: эпителиоподобная.
Кариология: 2n=22, пределы изменчивости по числу хромосом 16-25, модальное число хромосом 21, псевдодиплоид, имеются микрохромосомы, количество полиплоидов 3,0%.
Признаки опухолевого роста in vivo, in vitro: отсутствуют.
Онкогены: отсутствуют.
Морфология клеточной линии: при анализе в световой микроскоп, культура представлена суспензией круглых клеток с овальным ядром, содержащих небольшие эндосомы, время удвоения 28 часов.
Культуральные свойства, маркерные признаки: способ культивирования - суспензионный, посевная доза - 300 000 на мл., кратность рассева - 1:2 - 1:6, процедура пересева - центрифугирование суспензии на скорости 1200 об/мин, 5 мин., ресуспендирование в новой ростовой среде. Оптимальная плотность 1,0-1,2х106 клеток/мл. Клетки адаптированы к росту в культуральной среде без добавления сыворотки.
Рекомендуемые условия для замораживания: при плотности ≥ 2,0х106 живых клеток/мл; ростовая среда, 10% ДМСО
Трансфекция клеток линии CHO DG44 парой плазмид p1.1-Tr2-HCG-AIB (длина 8137 пар оснований, SEQ ID NO: 1, Фиг.1) и p1.2-GS-bhCG (длина 13291 пар оснований, SEQ ID NO: 4, Фиг.2) с последующей селекцией, амплификацией трансгена, кодируемого плазмидой p1.1-Tr2-HCG-AIB, и клонированием приводит к получению клеточных линий-продуцентов рекомбинантного ХГЧ.
Линия-продуцент D-pTr2-hCG-55 обеспечивает синтез и секрецию рекомбинантного гонадотропного гормона человека в количестве 5,4 мкг гонадотропного гормона на 1 мл среды при концентрации клеток 1,7 млн/мл при культивировании в суспензионной культуре в бессывороточной среде. Линия-продуцент D-pTr2-hCG-55 депонирована во Всероссийской Коллекции Промышленных Микроорганизмов (ВКПМ) под регистрационным номером ВКПМ Н-194.
Трансфекция клеток линии CHO S парой плазмид p1.1-Tr2-HCG-BIA (длина 8137 пар оснований, SEQ ID NO: 5, Фиг.3) и p1.2-GS-ahCG (длина 13144 пар оснований, SEQ ID NO: 6, Фиг.4) с последующей селекцией, амплификацией трансгена, кодируемого плазмидой p1.1-Tr2-HCG-BIA, и клонированием приводит к получению клеточных линий-продуцентов рекомбинантного ХГЧ.
Линия-продуцент S-pTr2-hCG-8 обеспечивает синтез и секрецию рекомбинантного гонадотропного гормона человека в количестве 6,3 мкг гонадотропного гормона на 1 мл среды при концентрации клеток 3,2 млн/мл при культивировании в суспензионной культуре в бессывороточной среде. Линия-продуцент S-pTr2-hCG-8 депонирована во Всероссийской Коллекции Промышленных Микроорганизмов (ВКПМ) под регистрационным номером ВКПМ Н-213.
Выращивание клеток, выделение и очистка целевого белка из культуральной или подобной ей жидкости может быть осуществлена способом, подобным традиционным способам культивирования, в которых рекомбинантный белок продуцируется с использованием клеток млекопитающих.
Питательная среда, используемая для культивирования, может быть как синтетической, так и натуральной, при условии, что указанная среда содержит источники углерода, азота, незаменимые аминокислоты, нуклеозиды, незаменимые липиды, минеральные добавки, инсулин и, если необходимо, соответствующее количество питательных добавок, требующихся для роста клеток. В качестве источника углерода могут использоваться различные углеводы, такие как глюкоза или сахароза и другие органические соединения. В качестве источника азота может использоваться аминокислота глутамин, или ее дипептиды, или природные источники азота, такие как соевый гидролизат. В качестве минеральных добавок могут использоваться фосфат калия, сульфат магния, хлорид натрия, сульфат железа, сульфат марганца, хлорид кальция и подобные им соединения. В качестве витаминов могут использоваться тиамин, дрожжевой экстракт и т.п.
Для получения фармацевтически пригодного гонадотропного гормона человека предпочтительно использование бессывороточной питательной среды для культивирования.
Выращивание может осуществляться в аэробных условиях, предпочтительно с повышенным содержанием СО2 (5%), таких как перемешивание культуральной жидкости в колбах, при температуре в пределах от 20 до 40°С, предпочтительно в пределах от 34 до 38°С. Обычно выращивание в течение от 24 часов до 20 дней приводит к накоплению целевого рекомбинантного белка в культуральной среде. После выращивания твердые остатки, такие как клетки и их фрагменты, могут быть удалены из культуральной жидкости методом центрифугирования, а затем целевой белок может быть выделен и очищен методами хроматографии и/или концентрирования.
Особенности созданных экспрессионных плазмид, линии клеток и результаты практического применения изобретения приведены на следующих фигурах.
Краткое описание Фигур.
На Фигуре 1 показана карта экспрессионной плазмиды p1.1-Tr2-HCG-AIB (длина 8137 пар оснований). Используются следующие обозначения: pUC origin - область начала репликации плазмиды pUC; bla - открытая рамка считывания бета-лактамазы, обеспечивающей устойчивость к ампициллину; bla promoter - прокариотический промотер гена bla; EBV TR - участок терминального повтора вируса Эпштейна-Барр человека; 5CHEF функциональный промотор гена фактора элонгации 1 альфа китайского хомячка, фланкированный 5’ НТО этого гена; transcription start - точка начала транскрипции; TATA - TATA бокс, 3CHEF функциональный терминатор и сигнал полиаденилирования гена фактора элонгации 1 альфа китайского хомячка, фланкированный 3’ НТО этого гена; pA - сигнал полиаденилирования; EMCV IRES - внутренний сайт связывания рибосом вируса энцефаломиокардита (EMCV) дикого типа; att EMCV IRES - аттенюированный внутренний сайт связывания рибосом вируса энцефаломиокардита (EMCV); DHFR - открытая рамка считывания дигидрофолатредуктазы мыши для селективного отбора и амплификации в эукариотических клетках; Kozak - область, кодирующая последовательность Козак для кэп-зависимой инициации трансляции, hCG-A-chain - ОРС альфа-субъединицы ХГ; hCG-B-chain - ОРС бета-субъединицы ХГ; stop - блок стоп-кодонов. Стрелками указаны направления транскрипции генов, в скобках указаны номера первого и последнего нуклеотидов фрагментов. Курсивом выделены сайты узнавания эндонуклеаз рестрикции, в скобках указаны номера нуклеотидов в точках разрезания.
На Фигуре 2 показана карта экспрессионной плазмиды p1.2-GS-bhCG (длина 13291 пар оснований). Используются обозначения аналогично фигуре 1, а также SV40 promoter - область промотора вируса SV40; SV40pA, terminator - терминатор и сигнал полиаденилирования вируса SV40; GS ORF - ОРС гена глутаминсинтазы крысы.
На Фигуре 3 показана карта экспрессионной плазмиды p1.1-Tr2-HCG-BIA (длина 8137 пар оснований). Используются обозначения аналогичные указанным для Фиг. 1.
На Фигуре 4 показана карта экспрессионной плазмиды p1.2-GS-ahCG (длина 13144 пар оснований). Используются обозначения аналогичные указанным для Фиг. 2.
На Фигуре 5 показаны результаты иммуноблоттинга культуральной среды для оценки доли секреции гетеродимера и свободных цепей ХГЧ в культуральную среду. Используются следующие обозначения: anti-alpha - антитела против альфа-цепи ХГЧ. anti-beta - антитела против бета-цепи ХГЧ. OVI - коммерческий препарат «Овитрель» (рекомбинантный ХГЧ); HA8 - поликлональный продуцент ХГЧ CHO DG-44/p1.1-Tr2-HCG-AIB, 8 мкМ МТХ; HA8+BGS - поликлональный продуцент ХГЧ, дополнительно трансфицированный конструкцией, кодирующей бета-цепь ХГЧ; HB8 - поликлональный продуцент ХГЧ CHO S/p1.1-Tr2-HCG-BIA, 8 мкМ МТХ; HB8+AGS - поликлональный продуцент ХГЧ, дополнительно трансфицированный конструкцией, кодирующей альфа-цепь ХГЧ. Цифрами обозначены отдельные клоны. Стартовая нагрузка калибратора и образцов - по 250 нг гетеродимера/дорожка, далее серия двукратных разведений. Серии разведений обозначены черными треугольниками, молекулярные массы полос маркера обозначены в кДа. OVI 95°C - обработка препарата сравнения нагревом до 95°C перед электрофорезом для разделения цепей без восстановления дисульфидных связей.
На Фигуре 6 показана электрофореграмма очищенного ХГЧ - продукта, полученного из линий D-pTR2-hCG-55 и S-pTR2-hCG-8 - в сравнении с лекарственным препаратом «Овитрель» (OVI) при различных способах пробоподготовки: без нагрева и восстановления, с нагревом (100°С), с нагревом и восстановлением 10 мМ дититрейтола (DTT). 2 мкг белка в каждой дорожке, окраска коллоидным Кумасси синим. Стрелками обозначены позиции полос или серий полос гетеродимера ХГЧ, бета-цепи ХГЧ, альфа-цепи ХГЧ.
На Фигуре 7 приведена оценка стабильности свойств моноклональных линий-продуцентов ХГЧ при продолжительном культивировании в течение 60 дней в бессывороточной культуральной среде. Показаны динамика клеточной плотности и накопления целевого белка.
На Фигуре 8 показаны данные ПЦР-РВ по копийности областей ОРС альфа-цепи и бета-цепи ХГЧ в геноме клеток для поликлональных и моноклональных популяций. Нормализация по ДНК из гена Rab1. Планки погрешностей обозначают стандартные отклонения, n=3.
На Фигуре 9 представлены результаты определения биологической активности ХГЧ в сравнении с 6-м международным стандартов ВОЗ. Расчет произведен с использованием функции линейной регрессии, в которой масса семенников использовалась в качестве величины ответной реакции вводимой дозе ХГЧ. Определенная биологическая активность ХГЧ составила 24,7 МЕ/мкг, 95% доверительный интервал составил 16,7 - 35,7 МЕ/мкг (P=0,95).
На Фигуре 10 представлен профиль гликанов образца ХГЧ. Рассчитанный Z-индекс составил 164. На хроматограмме обозначены группы пиков гликанов: 0S - нейтральные гликаны; 1S, 2S, 3S, 4S - моно-, ди-, три, тетрасиалированные гликаны, соответственно.
На Фигуре 11 представлена типичная хроматограмма профиля образца ХГЧ для определения его концентрации после очистки из культуральной жидкости.
Практическая применимость настоящего изобретения иллюстрируется следующими примерами.
Пример 1. Дизайн, получение и верификация последовательности ДНК конструкций, кодирующих альфа- и бета-субъединицы гонадотропного гормона человека pUC57-Kan-HCG-AIB и pUC57-Kan-HCG-BIA.
Для расчета синтетической ДНК, кодирующей ОРС цепей ХГЧ были использованы природные последовательности гена, в силу того что по имеющимся литературным данным оптимизация кодонов и другие манипуляции с кодирующей последовательностью могут приводить к снижению уровней экспрессии целевых генов http://www.freepatentsonline.com/y2012/0034655.html. В качестве источника последовательности природного гена использована публичная базы данных GeneBank. Для ОРС альфа-субъединицы - NM_000735.3 (http://www.ncbi.nlm.nih.gov/nuccore/NM_000735.3; использовали транскрипционный вариант 2, имеющий длину, совпадающую с указанной для хориогонадотропина альфа, что соответствует последовательности транслируемого белка SEQ ID NO: 2. Для ОРС бета-субъединицы ХГч использовали эталонную последовательность - NM_000737.3 (https://www.ncbi.nlm.nih.gov/nuccore/NM_000737.3), что соответствует последовательности транслируемого белка SEQ ID NO: 3.
Фрагменты ДНК, кодирующие цепи альфа- и бета-субъединицы гонадотропного гормона человека, предваряемые консенсусной последовательностью Козак, содержащие между ОРС субъединиц внутренний сайт связывания рибосом вируса EMCV (IRES), и фланкированные адапторными сайтами узнавания эндонуклеаз рестрикции AbsI и AsuNHI были синтезирован по заказу ООО «Клонинг Фасилити» (Москва, Россия), синтетические фрагменты были клонированы в плазмидный вектор pUC57-Kan, рестрицированный эндонуклеазой EcoRV, с образованием плазмид pUC57-Kan-HCG-AIB и pUC57-Kan-HCG-BIA. Приведенные плазмиды различаются положением ОРС цепей гормона в экспрессионной кассете (альфа-цепь - IRES - бета-цепь в случае AIB и бета-цепь - IRES - альфа-цепь в случае BIA).
Плазмиды трансформировали в клетки E.coli Top10 (F-mcrAΔ(mrr-hsdRMS-mcrBC) ϕ80lacZΔM15 ΔlacZX15 ΔlacX74 recA1 araD139 Δ(ara-leu)7697 galUgalKrpsL (StrR) endA1 nupG) (Invitrogen, Великобритания). Для этого к 200 мкл замороженной суспензии клеток E. coli добавляли 1 мкл раствора ДНК, инкубировали на льду 10 минут для сорбции плазмидной ДНК, нагревали до 42°С на 45 секунд и инкубировали на льду 1-5 минут. После чего добавляли 800 мкл питательного бульона SOC (20 г/л триптона, 5 г/л дрожжевого экстракта, 0,5 г/л NaCl, 250 мМ KCl, 10 мМ MgCl2) и инкубировали при 37°С 60 минут. После инкубации переносили суспензию на чашку Петри с твёрдой агаризованной средой 2xYT-агар (16 г/л триптона, 10 г/л дрожжевого экстракта, 10 г/л NaCl, 18 г/л агара), содержащей канамицин в концентрации 30 мкг/мл и помещали в термостат на 37°С на 18 часов. Отдельные клоны трансформантов инокулировали в 5 мл питательного бульона 2xYT (16 г/л триптона, 10 г/л дрожжевого экстракта, 10 г/л NaCl) с добавлением канамицина в концентрации 30 мкг/мл, наращивали 18 часов при 37°С, и выделяли плазмидную ДНК набором реактивов Plasmid Miniprep (ЗАО «Евроген», Россия) в соответствии с рекомендациями фирмы производителя. Полностью секвенировали области вставки полученных плазмид с праймеров M13dir и M13rev (SEQ ID NO: 7 и 8) с использованием набора BigDye® Terminator v. 3.1 Cycle Sequencing Kit (AppliedBiosystems, США) и капиллярного секвенатора ABI PRISM 3730 Genetic Analyzer (AppliedBiosystems, США), анализ данных вели при помощи программы Chromas 1.45 (Technelysium Pty Ltd, Australia). Плазмиды с верифицированной последовательностью обоих цепей использовали для получения экспрессионных конструкций.
Пример 2. Создание экспрессионных конструкций p1.1-Tr2-HCG-AIB и p1.1-Tr2-HCG-BIA, кодирующих альфа- и бета-субъединицы гонадотропного гормона человека.
На базе плазмидного вектора p1.1, описанного в патенте РФ 2488633 (Фиг. 2 в патенте 2488633), созданы экспрессионные конструкции для трицистронной экспрессии генов альфа- и бета-субъединиц гонадотропного гормона человека и ОРС дигидрофолатредуктазы мыши.
Вектор p1.1 был укорочен в областях 5’CHEF и 3’CHEF, укороченный вариант назвали p1.1-Tr2. Для этого эндонуклеазами EcoRI и PciI («СибЭнзим», Россия) вырезали дистальный фрагмент из области 5’CHEF вектора р1.1, проводили достройку выступающих концов Большим фрагментом Кленова ДНК-полимеразы I из E. coli («СибЭнзим», Россия) с последующим внутримолекулярным лигированием тупых концов ДНК-лигазой фага Т4 (Fermentas, Литва). Фрагмент 3’CHEF заменяли на укороченный фрагмент 3’CHEF-F1, который вырезали из плазмиды pAL-3CHEF-F1 по сайтам AsuNI-XbaI («СибЭнзим», Россия), и лигировали в плазмиду pBL-ID-EBV-5CHEF, представляющую собой р1.1 без 3’CHEF, рестрицированную эндонуклеазой XbaI.
Плазмиды pUC57-Kan-HCG-AIB и pUC57-Kan-HCG-BIA расщепляли эндонуклеазами AbsI и AsuNI («СибЭнзим», Россия) и переносили фрагменты, содержащие ОРС обеих цепей ХГЧ, интактный IRES EMCV и участок консенсусной последовательности Козак, в вектор p1.1-Tr2, рестрицированный теми же эндонуклеазами, c образованием плазмиды p1.1-Tr2-HCG-AIB и плазмиды p1.1-Tr2-HCG-BIA соответственно. Приведенные плазмиды различаются положением ОРС цепей гормона в экспрессионной кассете (альфа-цепь - IRES - бета-цепь в случае «AIB» и бета-цепь - IRES - альфа-цепь в случае «BIA»).
Рестрикции проводили по инструкции производителя ферментов. Продукты рестрикции смешивали в соотношении 5:1 с буфером для нанесения «6x Loading dye» («Fermentas», Литва) и проводили разделение в 1% агарозном геле с добавлением бромистого этидия (0,4 мкг/мл). Для выделения ДНК, полосу, визуализированную при помощи УФ-лампы, вырезали из агарозного геля, помещали в пробирку объемом 1,5 мл, после чего проводили выделение ДНК, набором Cleanup Standard («Евроген», Россия) согласно рекомендациям фирмы производителя. Лигирование очищенных фрагментов проводили ДНК-лигазой фага Т4 (Fermentas, Литва) по методике производителя фермента. Полученной лигазной смесью трансформировали как описано в Примере 1 клетки E. coli штамма Top10 (Invitrogen, Великобритания) и высевали на чашку Петри с твёрдой агаризованной средой 2xYT-агар (16г/л триптона, 10г/л дрожжевого экстракта, 10г/л NaCl, 18 г/л агара), содержащей ампициллин в концентрации 100 мкг/мл и помещали в термостат на 37°С на 18 часов.
Отбор клонов E. coli проводился методом ПЦР с клонов с использованием праймеров SQ-5CH6-F и SQ-FA-R (SEQ ID NO: 9 и 10). ПЦР с колоний E. coli проводили с использованием набора реактивов «ScreenMix-HS» (ЗАО Евроген, Россия), по методике производителя, помещая образцы бактериальных клонов в пробирки с 10 мкл готовой смеси для ПЦР. Продукты ПЦР анализировали электрофорезом в 1% агарозном геле, позитивные клоны наращивали в 5 мл среды 2xYT с добавлением ампициллина в концентрации 50 мкг/мл 18 часов при 37°С, и выделяли плазмидную ДНК набором реактивов Plasmid Miniprep (ЗАО «Евроген», Россия).
Области открытых рамок цепей ХГЧ в полученных плазмидах p1.1-Tr2-HCG-AIB и p1.1-Tr2-HCG-BIA были полностью отсеквенированы, как описано выше, с использованием праймеров SQ-5CH6-F (SEQ ID NO: 9) и SQ-FA-R (SEQ ID NO: 10), SQ-FA-F (SEQ ID NO: 11), AD-HCGb-SpeR (SEQ ID NO: 12) и AD-IHCGb-NheF (SEQ ID NO: 13), мутации в кодирующих областях не выявлены.
Полученные экспрессионные конструкции для трицистронной экспрессии цепей альфа и бета гонадотропного гормона человека и ОРС дигидрофолатредуктазы мыши p1.1-Tr2-HCG-AIB и p1.1-Tr2-HCG-BIA имеют размер 8137 п.о. и содержат уникальные сайты узнавания эндонуклеазами рестрикции NheI (1), AscI (1831), PvuI (3224), EcoRI (4183), AbsI (6700), XbaI (6886) - в случае p1.1-Tr2-HCG-AIB, и NheI (1), AscI (1831), PvuI (3224), EcoRI (4183), AbsI (6700), XbaI (7955) - в случае p1.1-Tr2-HCG-BIA.
Данные конструкции предназначены для трансфекции эукариотических клеток генами альфа- и бета-субъединиц гонадотропного гормона человека с последующим отбором и амплификацией по сцепленному признаку DHFR+. Они сообщают клеткам-реципиентам следующие фенотипические признаки: при трансформации прокариот - устойчивость к ампицилину, карбенициллину; при трансфекции эукариот - устойчивость к отсутствию гипоксантина и тимидина, ограниченная устойчивость к метотрексату, увеличивающаяся при селективном отборе, экспрессия рекомбинантного гонадотропного гормона человека. Созданные генетические конструкции использовали для получения линий клеток-продуцентов ХГ.
Пример 3. Создание экспрессионных конструкций p1.2-GS-ahCG и p1.2-GS-bhCG, кодирующих альфа- и бета-субъединицы хорионического гонадотропина человека соответственно.
На базе плазмидного вектора p1.2 для гетерологичной экспрессии рекомбинантных белков в клетках млекопитающих подробно описанного в патенте РФ 2552170 (в патенте РФ 2552170 представлен в SEQ ID: 1, Фиг. 2) созданы экспрессионные конструкции для моноцистронной экспрессии индивидуальных альфа или бета субъединиц ХГЧ в векторе, содержащем метаболический маркер - фермент глутаминсинтазу крысы.
Получение p12-GS содержащего глутаминсинтазу крысы описано в статье Kovnir, S. V., N. A. Orlova, C. Khodak Ycapital A, M. P. Kondrashova, A. G. Gabibov, K. G. Skryabin, A. I. Vorobiev and Vorobiev, II (2017). "Approaches to Controlled Co-Amplification of Genes for Production of Biopharmaceuticals: Study of the Insertion and Amplification Dynamics of Genetic Cassettes in the Genome of Chinese Hamster Ovary Cells during Co-Expression of Compatible Pair of Plasmids." Bull Exp Biol Med 163(2): 245-249). ОРС глутаминсинтетазы амплифицировали c адапторными праймерами AD-GS-BglNcoF (SEQ ID NO: 14) и AD-GS-XbaR (SEQ ID NO: 15), субклонировали по сайтам рестрикции NcoI-XbaI в плазмиду pAL-Zeo с образованием плазмиды pAL-GS. Рестриктный фрагмент AscI-AscI, включающий промотор и терминатор вируса SV40 и ОРС глутаминсинтазы переносили в плазмиду p1.2.
Плазмиды pUC57-Kan-HCG-AIB и pUC57-Kan-HCG-BIA расщепляли эндонуклеазами AbsI и SpeI («СибЭнзим», Россия) и переносили фрагменты, содержащие соответственно ОРС альфа- или бета-субъединицы ХГЧ, предваряемые участком консенсусной последовательности Козак, в вектор p1.2-GS, рестрицированный эндонуклеазами AbsI и AsuNHI («СибЭнзим», Россия), c образованием плазмид p1.2-GS-ahCG и p1.2-GS-bhCG соответственно.
Рестрикции проводили по инструкции производителя ферментов. Продукты рестрикции выделяли из 1% агарозного геля как описано в Примере 1 набором Cleanup Standard («Евроген», Россия) согласно рекомендациям фирмы производителя. Лигирование очищенных фрагментов проводили ДНК-лигазой фага Т4 (Fermentas, Литва) по методике производителя фермента. Полученной лигазной смесью трансформировали как описано в Примере 1 клетки E. coli штамма Top10 (Invitrogen, Великобритания) и высевали на чашку Петри с твёрдой агаризованной средой 2xYT-агар (16г/л триптона, 10г/л дрожжевого экстракта, 10г/л NaCl, 18 г/л агара), содержащей ампициллин в концентрации 100 мкг/мл и помещали в термостат на 37°С на 18 часов. Отбор клонов E. coli вели методом ПЦР с клонов с использованием праймеров SQ-5CH6-F (SEQ ID NO: 9) и SQ-3CH1-R (SEQ ID NO: 16) и набора «ScreenMix-HS» (ЗАО Евроген, Россия), по методике производителя, помещая образцы бактериальных клонов в пробирки с 10 мкл готовой смеси для ПЦР, с последующим электрофорезом в 1% агарозном геле. Позитивные клоны наращивали в 5 мл среды 2xYT с добавлением ампициллина в концентрации 50 мкг/мл 18 часов при 37°С, и выделяли плазмидную ДНК набором Plasmid Miniprep (ЗАО «Евроген», Россия). Область открытой рамки бета-цепи ХГЧ полученной плазмиды была полностью отсеквенирована как описано выше с использованием праймеров SQ-5CH6-F (SEQ ID NO: 9) и AD-HCGb-SpeR (SEQ ID NO: 12), SQ-3CH1-R (SEQ ID NO: 16), мутаций не выявлено.
Полученные экспрессионные конструкция для моноцистронной экспрессии индивидуальных цепей ХГЧ в векторе, содержащем кассету устойчивости к метионилсульфоксимину имеют следующие размеры p1.2-GS-ahCG - 13144 п.о. и содержит уникальные сайты узнавания эндонуклеазами рестрикции PvuI (7739), AbsI (12776), XbaI (12962), а p1.2-GS-bhCG, имеет размер 13291 п.о. и содержит уникальные сайты узнавания эндонуклеазами рестрикции PvuI (7739), AbsI (12776).
Данные конструкции предназначена для трансфекции эукариотических клеток геном кодирующим альфа (в случае p1.2-GS-ahCG) или бета (в случае p1.2-GS-bhCG) субъединицу гонадотропного гормона человека с последующим отбором по способности синтезировать глутамин и признаку устойчивости к метионилсульфоксимину (MSX). Они сообщают клеткам-реципиентам следующие фенотипические признаки: при трансформации прокариот - устойчивость к ампицилину, карбенициллину; при трансфекции эукариот - экспрессия фермента глутаминсинтазы, устойчивость к MSX, экспрессия бета-субъединицы рекомбинантного гонадотропного гормона человека (в случае p1.2-GS-bhCG) или экспрессия альфа-субъединицы рекомбинантного гонадотропного гормона человека (в случае p1.2-GS-ahCG).
Схематическая карта p1.2-GS-bhCG приведена на Фиг. 2, p1.2-GS-ahCG - на Фиг.4.
Созданные генетические конструкции использовали для получения линий клеток-продуцентов ХГ в следующих комбинациях: p1.1-Tr2-HCG-AIB и p1.2-GS-bhCG или p1.1-Tr2-HCG-BIA и p1.2-GS-ahCG.
Пример 4. Препаративное выделение плазмидной ДНК для трансфекций и получение транзиентно трансфицированных популяции клеток CHO DG-44/p1.1-Tr2-HCG-AIB и CHO S/p1.1-Tr2-HCG-BIA.
Плазмидные ДНК для трансфекции нарабатывали в клетках Escherichia coli штамма Top10, трансформированных экспрессионными конструкциями. Клетки культивировали в 50-150 мл среды 2xYT в течение 16 ч, и выделяли плазмиды с использованием набора GeneJet™ Midi по протоколу фирмы-производителя (Fermentas, Литва). Переосаждали этанолом, осадки ДНК растворяли в стерильной воде.
Хозяйские клетки CHO DG-44 и CHO S культивировали в безбелковой среде ProCHO 5 (Lonza, Швейцария) с добавлением 8 мМ L-глутамина (Invitrogen, Ltd., США) и гипоксантина-тимидина (HT, Панэко, Россия). Клетки выращивали в стерильных одноразовых колбах Эрленмейера (30 мл суспензии в колбах вместимостью 125 мл) на орбитальной качалке при 130 об/мин в атмосфере, содержащей 5% углекислого газа при температуре 37°C. Подсчет плотности клеточной суспензии и доли живых клеток проводили в счётной камере Горяева при окрашивании трипановым синим. Пассирование культуры проводили по достижении плотности 2,0 млн клт/мл, посевная концентрация 0,3 млн клт/мл. За 24 ч до трансфекции клетки дополнительно пассировали с посевной концентрацией 0,5 млн клт/мл.
Полученными препаратами очищенных плазмид p1.1-Tr2-HCG-AIB и p1.1-Tr2-HCG-BIA были трансфицированы клетки CHO DG-44 и CHO S, соответственно. К препаратам плазмид p1.1-Tr2-HCG-AIB или p1.1-Tr2-HCG-BIA перед трансфекцией добавляли 5% (по массе) плазмиды pEGFP-N2 (Clontech, США), кодирующей зелёный флуоресцентный белок под контролем эукариотического промотора.
Трансфекцию клеток проводили методом нуклеофекции при помощи аппарата Invitrogen Neon и набора Neon 100 mkl transfection kit (Invitrogen, США), 1700 В, 1 импульс, раствор R. Трансфекцию клеток проводили для 10 млн живых клеток и 50 мкг плазмидной ДНК, растворенной в буфере R в концентрации 1-3 мг/мл.
После трансфекции культивировали клетки в течение 48 ч в неселективной среде, измеряли эффективность трансфекции как долю живых клеток, флюоресцирующих в зеленом свете. Эффективность трансфекции, измеренная как доля флуоресцирующих клеток, составила 6%. Титр ХГЧ в среде определяли при помощи иммуноферментного анализа (ИФА) по схеме «сэндвич», используя в качестве нижних антител моноклональные антитела против бета-цепи ХГЧ (ХН51, ХЕМА-Медика), в качестве верхних антител для детекции - конъюгат пероксидазы хрена с моноклональными антителами против альфа-цепи гонадотропных гормонов (XF1*, ХЕМА-Медика). В качестве калибратора использовали коммерческий препарат рекомбинантного ХГЧ «Овитрель» (Мерк, Италия).
Концентрация секретированного ХГЧ для транзиентно трансфицированных культур составила: 1 нг/мл для p1.1-Tr2-HCG-AIB, 4 нг/мл для p1.1-Tr2-HCG-BIA.
Пример 5. Получение стабильно трансфицированных популяций клеток CHO DG-44/p1.1-Tr2-HCG-AIB и CHO S/p1.1-Tr2-HCG-BIA и амплификация целевого гена под действием возрастающих концентраций метотрексата.
После выращивания в неселективной среде в течение 48 ч культуру пассировали с добавлением последовательно возрастающих концентраций селекционного агента МТХ (0,2 мкМ, 2 мкМ, 8 мкМ). На каждом шаге вели культивирование в течение 20-30 дней в селекционной среде с периодическим пассированием культур через 3-4 дня. Во всех случаях были получены делящиеся популяции клеток с жизнеспособностью более 90%, при этом при повышении концентраций MTX наблюдали увеличение титра целевого белка. Концентрация секретируемого гетеродимера ХГЧ приведена в Таблице 1 для CHO DG-44/p1.1-Tr2-HCG-AIB (код HA) и Таблице 2 для CHO S/p1.1-Tr2-HCG-BIA (код HA).
Таблица 1. Титр ХГЧ для поликлональной культуры CHO DG-44/p1.1-Tr2-HCG-AIB, при селекции и амплификации. Последовательная культивация в бессывороточной среде в присутствии возрастающих концентраций МТХ, оценка методом ИФА.
Таблица 2. Титр ХГЧ для поликлональной культуры CHO S/p1.1-Tr2-HCG-BIA при селекции и амплификации. Последовательная культивация в бессывороточной среде в присутствии возрастающих концентраций МТХ, оценка методом ИФА.
Таким образом, в результате продолжительного культивирования в присутствии возрастающих концентраций МТХ продуктивность клеток была увеличена в несколько раз. Для поликлональной популяции HA8 на третьи сутки выращивания удельная продуктивность составила 3,4 пг/клетку, для поликлональной популяции HB8 - 3,7 пг/клетку.
Пример 6. Получение популяции клеток-продуцентов ХГч, стабильно трансфицированных вспомогательными плазмидами p1.2-GS-ahCG и p1.2-GS-bhCG.
Для кондиционированной культуральной среды, полученной от популяций клеток HA8 и HB8, определяли относительный уровень секреции цепей ХГЧ при помощи иммуноблоттинга с моноклональными антителами к обеим цепям (Фиг. 5). Установили, что в обоих случаях клеточными популяциями секретируется преимущественно ХГЧ в форме гетеродимера, но также наблюдается присутствие в культуральной среде мономерных цепей гормона, при этом уровень свободной цепи, расположенной в первом цистроне, значительно превосходит уровень свободной цепи, расположенной во втором цистроне.
Предположили, что эффективность трансляции области открытой рамки считывания субъединицы в первом цистроне снижена относительно области открытой рамки считывания субъединицы во втором цистроне, то есть эффективность реинициации трансляции EMCV IRES дикого типа ниже 100%. Для компенсации уровня экспрессии недостающей цепи в клетки ввели дополнительные копии гена данной цепи: популяцию CHO DG-44/p1.1-Tr2-HCG-AIB (HA8) повторно трансфицировали плазмидой p1.2-GS-bhCG с получением популяции HA8+BGS, популяцию CHO S/p1.1-Tr2-HCG-BIA (HB8) повторно трансфицировали плазмидой p1.2-GS-ahCG с получением популяции HB8+AGS. Трансфекцию проводили, как указано в Примере 4.
Через 48 часов к культурам добавили MSX в концентрации 25 мкМ. Пассирования культуры проводили по достижении плотности 2,0 тыс.кл/мл, но не позднее чем через пять суток после предыдущего пересева. Через 20-30 дней после повторной трансфекции были получены стабильно трансфицированные популяция клеток HA8+BGS и HB8+AGS с долей живых клеток более 90%. После восстановления жизнеспособности провели измерение концентрации ХГЧ методом ИФА. Установили, что продуктивность клеток при введении дополнительных копий недостающей цепи в геном продуцентов была увеличена в 2-3 раза: 5,2 пг/клетка для HA8+BGS против 2,3 пг/клетка для HA8, 3,9 пг/клетка для HB8+AGS против 1,0 пг/клетка для HB8.
Полученные поликлональные популяции HA8+BGS и HB8+AGS были использованы для получения клональных линий-продуцентов ХГ.
Пример 7. Получение клональных линий-продуцентов ХГЧ.
Для поликлональных популяций HA8+BGS HA8+BGS и HB8+AGS было проведено клонирование методом предельных разведений в 96-луночных планшетах (200 мкл среды на лунку в культуральной среде EXCELL-CHO (Sigma), с добавлением 8 мМ глутамина, 2 мМ гипоксантина и 2 мМ тимидина (Панэко) Клетки разводили до концентрации 10 жизнеспособных клеток/мл, помещали в лунки 96-луночных планшетов (Corning, США), конечная плотность посева составила 1 клетка на лунку, инкубировали в течение 14 дней при 37°C, 5% CO2 в инкубаторе. Эффективность клонирования составила около 15%. Для лунок, содержащих единичные колонии, проводили последовательное масштабирование в планшеты следующего калибра (24-луночные, затем 6-луночные), отбирая такие колонии, для которых титр ХГ, измеренный по ИФА, был наибольшим. На финальном этапе масштабирования после адаптации культур к суспензионному культивированию (перемешиваемые колбы Эрленмейера объемом 125 мл, объем культивации 30 мл) были отобраны три наиболее перспективные клональные линии D-pTr2-hCG-1, D-pTr2-hCG-55, D-pTr2-hCG-69, обладающие удовлетворительными ростовыми показателями при выращивании в суспензионной культуре и максимальным титром ХГЧ (Таблица 3).
Таблица 3. Титр ХГЧ для моноклональных культур при культивации в бессывороточной среде. Оценка методом ИФА.
Таблица 4. Титр ХГЧ для моноклональных культур при культивации в бессывороточной среде. Оценка методом ИФА.
Пример 8. Измерение копийности вставок трансгена методом количественного ПЦР в реальном времени (ПЦР-РВ).
Методом количественного ПЦР в реальном времени (ПЦР-РВ) оценили копийность вставок трансгена в геномной ДНК штамма-продуцента, выделенной при помощи набора ДНК Wizard SV (Promega, США) по инструкции производителя. Для проведения ПЦР использовали реакционную смесь qPCRmix-HS-SYBR (Евроген) и праймеры RT-LTHa-F (SEQ ID NO: 17) и RT-LTHa-R (SEQ ID NO: 18) для альфа цепи, RT-bhCG-F (SEQ ID NO: 19) RT-bhCG-R (SEQ ID NO: 20) - для бета-цепи, RT-Rab-F (SEQ ID NO: 21) и RT-Rab-R (SEQ ID NO: 22) - для контрольного ампликона Rab, встречающегося в геноме CHO один раз. Проводили предварительную денатурацию 10 мин при 95°С, 40 циклов амплификации (денатурация 10 с при 95°С, отжиг 15 с при 55°С, элонгация 15 с при 72°С) при помощи амплификатора iCycler iQ (Bio-Rad, США), детекцию осуществляли на стадии элонгации. Для калибровки использовали высокоочищенную плазмиду p1.1-Tr2-HCG-AIB с известной концентрацией.
Копийность областей ОРС альфа- и бета-цепей ХГч многократно возрастала при геномной амплификации поликлональных популяции клеток от десятков до сотен копий на гаплоидный геном (Фигура 6). После трансфекции клеток второй экспрессионной плазмидой и отбора дважды стабильно трансфицированной популяции клеток наблюдали в обоих случаях увеличение копийности области ОРС дополнительной цепи ХГч. В трех полученных клональных линиях клеток копийность ОРС альфа- и бета-цепи многократно различалась между линиями, во всех линиях копийность области ОРС альфа-цепи превышала копийность области ОРС бета-цепи. Для кандидатных клональных линий-продуцентов копийность цепей составила: для D-pTr2-hCG-55 - 324±86 копий для альфа-цепи, 344±66 копий для бета-цепи; для S-pTr2-hCG-8 - 1557±424 копий для альфа-цепи, 153±50 копий для бета-цепи.
Пример 9. Непрерывное культивирование кандидатных моноклональных линий-продуцентов ХГЧ в течение 60 дней.
Для оценки стабильности уровней экспрессии целевых генов три моноклональные линии клеток-продуцентов (1, 55, 69), полученные на основе клеток линии CHO-DG44, и три моноклональные линии клеток-продуцентов (8, 18, 21), полученные на основе клеток линии CHO-S, непрерывно культивировали в течение 60 дней в бессывороточной среде определенного химического состава. Пассирование культуры проводили с трех-четырехдневными интервалами методом разбавления свежей средой 1:3-1:5, посевная концентрация составляла 0,2 млн клт./мл, на момент пересева плотность клеток достигала 0,6-1,4 млн клеток/мл (Фигура 7А, В). Титр продукта (Фигура 7Б, Г) при этом оставался стабильным в течение всего времени культивации для всех исследованных линий, кроме 18. Для клона 18 наблюдали трехкратное падение титра по результатам 60 дней культивирования, из чего следует, что данный клон не пригоден для промышленного применения.
Пример 10. Выделение целевого белка из супернатанта культуральной жидкости, полученной при культивации продуцентов ХГЧ на основе CHO DG44 и CHO S.
Кондиционированную культуральную среду отделяли от клеток и дебриса центрифугированием. Далее рН культуральной жидкости доводили до 5,0 при помощи концентрированной соляной кислоты. Проводили фильтрацию через капсулу Sartoguard NF MidiCaps (Sartorius, Германия) площадью 2200 см2. Далее получаемый материал объемом до 15 л наносили на сорбент Capto Blue (Cytiva, США), предварительно уравновешенный буфером, содержащим 0,02 М цитрата натрия, 0,005 М гистидина, 0,001 М метионина, 0,15 М хлорида натрия, pH 5,0±0,2, элюцию проводили буфером содержащим 0,02 М Трис (трис(гидроксиметил)аминометана), 2 М хлорида натрия, 10% изопропанола, pH 8,0±0,2. В полученный материал добавляли сульфат аммония до конечной концентрации 1,25 М и наносили на сорбент Capto Butyl ImpRes (Cytiva, США), элюцию целевой молекулы с сорбента проводили буфером, содержащим 0,02 М цитрата натрия, 0,005 М гистидина, 0,001 М метионина, 0,31 М сульфата аммония, pH 6,0±0,2. Перевод в буфер, содержащий 0,01 М дигидрофосфата натрия, 0,005 М гистидина, 0,001 М метионина, pH 7,0±0,2, осуществляли через сорбент Sephadex G25 (Cytiva, США). Далее полученный элюат наносили на сорбент Ceramic Hydroxyapatite Type I. Отсечку нецелевых изоформ ХГЧ осуществляли на сорбенте 40Q (WorkBeads, Швеция), предварительно уравновешенном буфером, содержащим 0,01 М Бис-Трис, 0,005 М гистидина, 0,001 М метионина, pH 7,0±0,2, элюции проводили с помощью буфера, содержащего 0,01 М Бис-Трис, 0,005 М гистидина, 0,001 М метионина, 0,5 М хлорида натрия, pH 7,0±0,2. Очистку от свободных субъединиц проводили на сорбенте Superdex 75 (Cytiva, США), промытым буфером, содержащим 0,01 М фосфорной кислоты, 0,299 М манитола, 0,00134 М метионина, 0,0009791 М полаксомера 188, pH 7,0±0,2.
Пример 11. Измерение биологической активности очищенного рекомбинантного ХГЧ.
42 неполовозрелых самца крыс WISTAR возрастом в диапазоне 21-23 дня, различающихся по возрасту не более чем на 3 дня, и с различиями по массе, не превышающими 10 гр между самой крупной и самой легкой особью, были сгруппированы в 6 групп по 7 крыс. В пределах каждой из 6 групп особям были проведены подкожные инъекции растворов, содержащих ХГЧ в дозе 1, 2, 4 МЕ на животное как в группах крыс с использованием стандартного образца (6-й международный стандарт ВОЗ хориогонадотропина альфа, кат. № 18/244, NIBSC, Великобритания), так и в группе с тестируемым образцом (очищенный ХГЧ серии № 010223). Инъекции повторяли через 24 и 48 часов после первой инъекции. Через 24 ч после последней инъекции животных подвергали эвтаназии, после чего у животных незамедлительно извлекали по одному семенные пузырьки, удаляли постороннюю жидкость и ткани, не относящиеся к изучаемому органу, и взвешивали.
Для определения биологической активности использовали метод сравнения логарифмических параллельных кривых, построенных для вводимых доз раствора стандартного объекта с известной концентрацией ХГЧ и раствора тестируемого объекта с предполагаемой концентрацией ХГЧ, где масса двух семенных пузырьков рассматривалась в качестве величины ответной реакции на вводимые дозы. Согласно Европейской Фармакопее (статья 01/2011:0498), для определенной величины удельной биологической активности ХГЧ должны выполняться следующие условия:
- удельная биологическая активность ХГЧ должна быть не менее 80% и не более 125% биологической активности стандартного образца (или от 20,8 до 32,5 МЕ/мкг);
- 95% доверительный интервал относительной активности (Relative Potency) должен быть не менее 64% и не более 156% (или от 16,64 до 40,56 МЕ/мкг).
Пример 12. Определение профиля гликозилирования ХГЧ и расчет Z-индекса.
Определение профиля гликозилирования рХГЧ проводили в следующих хроматографических условиях:
Thermo Scientific, кат. № 082470
Z-индекс рассчитывался по следующему уравнению:
Z=A0 x 0 + A1 x 1 + A2 x 2 + A3 x 3 + A4 x 4
где: A0, A1, A2, A3, A4 - процентные площади пиков нейтральных, моно-, ди-, три- и тетрасиалированных гликанов молекулы ХГЧ, соответственно. 0, 1, 2, 3, 4 - содержание сиаловых кислот для каждой группы гликанов. Z-индекс должен составлять от 120 до 170.
Пример 13. Количественное определение ХГЧ методом ВЭЖХ.
Определение концентрации ХГЧ проводили в следующих хроматографических условиях:
Концентрацию гетеродимера ХГЧ (С, г/л) определяли по формуле:
С = (Vис*Сст)/Vст г/л, где:
V ис - объем пика гетеродимера ХГЧ на хроматограмме испытуемого раствора;
С ст - концентрация гетеродимера ХГЧ в использованном стандартном образце, г/л;
V ст - объем пика гетеродимера ХГЧ на хроматограмме стандартного образца.
Хотя указанное изобретение описано в деталях со ссылкой на Примеры, для специалиста в указанной области техники очевидно, что могут быть совершены различные изменения и произведены эквивалентные замены, и такие изменения и замены не выходят за рамки настоящего изобретения.
--->
<?xml version="1.0" encoding="UTF-8"?>
<!DOCTYPE ST26SequenceListing PUBLIC "-//WIPO//DTD Sequence Listing
1.3//EN" "ST26SequenceListing_V1_3.dtd">
<ST26SequenceListing originalFreeTextLanguageCode="ru"
dtdVersion="V1_3" fileName="HCG-sequence listing.xml"
softwareName="WIPO Sequence" softwareVersion="2.3.0"
productionDate="2023-12-28">
<ApplicantFileReference>HCG-patent-2023</ApplicantFileReference>
<ApplicantName languageCode="ru">Общество с ограниченной
ответственностью "АйВиФарма"</ApplicantName>
<ApplicantNameLatin>IVPharma ISC</ApplicantNameLatin>
<InventionTitle languageCode="ru">Плазмиды для экспрессии
рекомбинантного хорионического гонадотропина человека (ХГЧ), плазмиды
для экспрессии рекомбинантных альфа- и бета-субъединиц ХГЧ,
моноклональные линии клеток млекопитающих - продуценты ХГч и способ
получения ХГЧ</InventionTitle>
<SequenceTotalQuantity>22</SequenceTotalQuantity>
<SequenceData sequenceIDNumber="1">
<INSDSeq>
<INSDSeq_length>8137</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..8137</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q2">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
<INSDFeature>
<INSDFeature_key>CDS</INSDFeature_key>
<INSDFeature_location>6714..7061</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>protein_id</INSDQualifier_name>
<INSDQualifier_value>2</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier>
<INSDQualifier_name>translation</INSDQualifier_name>
<INSDQualifier_value>MDYYRKYAAIFLVTLSVFLHVLHSAPDVQDCPECTLQENPFF
SQPGAPILQCMGCCFSRAYPTPLRSKKTMLVQKNVTSESTCCVAKSYNRVTVMGGFKVENHTACHCSTCY
YHKS</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
<INSDFeature>
<INSDFeature_key>CDS</INSDFeature_key>
<INSDFeature_location>7636..8130</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>protein_id</INSDQualifier_name>
<INSDQualifier_value>3</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier>
<INSDQualifier_name>translation</INSDQualifier_name>
<INSDQualifier_value>MEMFQGLLLLLLLSMGGTWASKEPLRPRCRPINATLAVEKEG
CPVCITVNTTICAGYCPTMTRVLQGVLPALPQVVCNYRDVRFESIRLPGCPRGVNPVVSYAVALSCQCAL
CRRSTTDCGGPKDHPLTCDDPRFQDSSSSKAPPPSLPSPSRLPGPSDTPILPQ</INSDQualifier_v
alue>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>ctagcggccgcgcagttaacgccgcccctctccctcccccccccctaac
gttactggccgaagccgcttggaataaggccggtgtgcgtttgtctatatgttattttccaccatattgc
cgtcttttggcaatgtgagggcccggaaacctggccctgtcttcttgacgagcattcctaggggtctttc
ccctctcgccaaaggaatgcaaggtctgttgaatgtcgtgaaggaagcagttcctctggaagcttcttga
agacaaacaacgtctgtagcgaccctttgcaggcagcggaaccccccacctggcgacaggtgcctctgcg
gccaaaagccacgtgtataagatacacctgcaaaggcggcacaaccccagtgccacgttgtgagttggat
agttgtggaaagagtcaaatggctctcctcaagcgtattcaacaaggggctgaaggatgcccagaaggta
ccccattgtatgggatctgatctggggcctcggtacacatgctttacatgtgtttagtcgaggttaaaaa
aacgtctaggccccccgaaccacggggacgtggttttcctttgaaaaacacgatgataatatggccacaa
gatctgccaccatggttcgaccattgaactgcatcgtcgccgtgtcccaaaatatggggattggcaagaa
cggagacctaccctggcctccgctcaggaacgagttcaagtacttccaaagaatgaccacaacctcttca
gtggaaggtaaacagaatctggtgattatgggtaggaaaacctggttctccattcctgagaagaatcgac
ctctaaaggacagaattaatatagttctcagtagagaactcaaagaaccaccacgaggagctcattttct
tgccaaaagtttggatgatgccttaagacttattgaacaaccggaattggcaagtaaagtagacatggtt
tggatagtcggaggcagttctgtttaccaggaagccatgaatcaaccaggccacctcagactctttgtga
caaggatcatgcaggaatttgaaagtgacacgtttttcccagaaattgatttggggaaatataaacttct
cccagaatacccaggcgtcctctctgaggtccaggaggaaaaaggcatcaagtataagtttgaagtctac
gagaagaaagactaatcatattacccctaacacctgccaccccagtcttaatcagtggtggaagaacggt
ctcagaactgtttgtctcaattggccatttaagtttaatagtgaaagactggttaatgataacaatgcat
cggaaaaccttcaggaggaaaggagaatgttttgtggaacatttttgtgtgtgtggcagttttaagttat
tagttttcaaaatcagtactttttaatggaaacaacttgaccaaaaatctgtcacagaattttgagaccc
attaaaatacaagtttaatgagaagtctgtctctgttaatgctgaagtcattactaagtgcttagcttag
caaggtatgtggatgcccatttgtgttccaagggattggactgttcatcaggacccagagctgagtttca
agggctcaagagatggcttattacctgtgggtgtcttgaaggttctggttgggacaaattaggaatgttt
ttggcagacatggtgactacccttcatctgggtgagttcagttgatttgtcttgagcctttggggtttac
acaagtaaatgacatcatacagttagtgtattgttagtgaatattaatatatgaggcaggctttgctcta
gctagacatatgtcgacgtccggagctcaggcgcgccatggctcttccgcttcctcgctcactgactcgc
tgcgctcggtcgttcggctgcggcgagcggtatcagctcactcaaaggcggtaatacggttatccacaga
atcaggggataacgcaggaaagaacatgtgagcaaaaggccagcaaaaggccaggaaccgtaaaaaggcc
gcgttgctggcgtttttccataggctccgcccccctgacgagcatcacaaaaatcgacgctcaagtcaga
ggtggcgaaacccgacaggactataaagataccaggcgtttccccctggaagctccctcgtgcgctctcc
tgttccgaccctgccgcttaccggatacctgtccgcctttctcccttcgggaagcgtggcgctttctcat
agctcacgctgtaggtatctcagttcggtgtaggtcgttcgctccaagctgggctgtgtgcacgaacccc
ccgttcagcccgaccgctgcgccttatccggtaactatcgtcttgagtccaacccggtaagacacgactt
atcgccactggcagcagccactggtaacaggattagcagagcgaggtatgtaggcggtgctacagagttc
ttgaagtggtggcctaactacggctacactagaagaacagtatttggtatctgcgctctgctgaagccag
ttaccttcggaaaaagagttggtagctcttgatccggcaaacaaaccaccgctggtagcggtttttttgt
ttgcaagcagcagattacgcgcagaaaaaaaggatctcaagaagatcctttgatcttttctacggggtct
gacgctcagtggaacgaaaactcacgttaagggattttggtcatgagattatcaaaaaggatcttcacct
agatccttttaaattaaaaatgaagttttaaatcaatctaaagtatatatgagtaaacttggtctgacag
ttaccaatgcttaatcagtgaggcacctatctcagcgatctgtctatttcgttcatccatagttgcctga
ctccccgtcgtgtagataactacgatacgggagggcttaccatctggccccagtgctgcaatgataccgc
gagacccacgctcaccggctccagatttatcagcaataaaccagccagccggaagggccgagcgcagaag
tggtcctgcaactttatccgcctccatccagtctattaattgttgccgggaagctagagtaagtagttcg
ccagttaatagtttgcgcaacgttgttgccattgctacaggcatcgtggtgtcacgctcgtcgtttggta
tggcttcattcagctccggttcccaacgatcaaggcgagttacatgatcccccatgttgtgcaaaaaagc
ggttagctccttcggtcctccgatcgttgtcagaagtaagttggccgcagtgttatcactcatggttatg
gcagcactgcataattctcttactgtcatgccatccgtaagatgcttttctgtgactggtgagtactcaa
ccaagtcattctgagaatagtgtatgcggcgaccgagttgctcttgcccggcgtcaatacgggataatac
cgcgccacatagcagaactttaaaagtgctcatcattggaaaacgttcttcggggcgaaaactctcaagg
atcttaccgctgttgagatccagttcgatgtaacccactcgtgcacccaactgatcttcagcatctttta
ctttcaccagcgtttctgggtgagcaaaaacaggaaggcaaaatgccgcaaaaaagggaataagggcgac
acggaaatgttgaatactcatactcttcctttttcaatattattgaagcatttatcagggttattgtctc
atgagcggatacatatttgaatgtatttagaaaaataaacaaataggggttccgcgcacatttccccgaa
aagtgccacctgcatctcatcacggtcacgcatggctgcgcccgcgcagcgccccccgacccccccccga
tccccggaacgccagccgccatccaccgcccgcagcccccggagcccgcggaccccgaccccccgccgct
cgcccgccgttgcgccgctctctgcgggggggctagggggccgcggggggaaggccacgccccctccact
ttttccaggaatgcgcggcatgacccgccccccccatgacccgccaacacccccccgatgccccccgggg
gtctttcctggggggcctttgtcagggttgcctgtgtcacccacgggccccttggccacgcttgcctggg
tcaccccggagccccgggtcacgtttgccggggtcagccctggggcgcttcttcacgctccattgccatg
cagaattcccacatgtggacaccggacagtaggtcctcaaatgctccttattaggttggttcaataatat
caattgtttgttactaggcagtgatgttgtacatctggaggagatctcttgagcccataatcaggttatt
aggaataaatactctaaggctaaaaatgtagcttagtgataagagtgcttgcctggtgtgctgagaccct
cggttccatctccacaaccccatattccattacaaaataccttttcaccgtccctagcattaagaaacaa
aacaacaaagaagtttttctttcttctgagatcctgcccggagaggcatttaaaactggccagggccaaa
aaaaaaaaaaaaaaaagaaaaaaaagaaaagaaaacaggctagggccggcatggtggcgcacgcctttaa
tcccagcacgcaggaggcagaggcagggcggatctctgtgagtttgaggtcagcctggtctacctagtga
gtttcagggcacccagggctaaagagactgtctcaaaaacaaaacagccacacaatcagaaccacagcaa
aacgcagttatgatccttggaactgtaggaatgacaagcatttaaataataggacgagccatttttgaga
agctctgatttcacaagtgtcagggatgggctctgggcgagtaagattgctaatgctggcctctaaatga
gaccacgtggagttgattagattcttttcatgttcctcgtgctctatcaaataactgtacccaaatacac
acacacacacacacacacaatgcgcgcacacacaaaatccttttttagcttaagaagcccagaatcagaa
gtaaagctaactgtgggacttaagtattattctgaacggaactcccagggcgtgaagcgcgcttcaggct
tccagagaagcagctggcgctggatggaatgaaccaagaggccagcacaggggcagatccgtcgagctct
cggccaccgagctgagcccttaggttctggggctgggaagggtccctaggattgtgcacctctcccgcgg
gggacaagcagggggatggcggggctgacgtcgggaggtggcctccacgggaagggacacccggatctcg
acacagccttggcagtggagtcaggaagggtagggacagattctggacgccctcttggccagctcctcac
cgccccacccccgactggagccgagagtaattcatacaaaaggagggatcgccttcgcccctgggaatcc
cagggaccgtcgctaaattctggccggcctcccagcccggaaccgctgtgcccgcccagcgcggcgggag
gagcctgcgcctagggcggatcgcgggtcggcgggagagcacaagcccacagtccccggcggtgggggag
gggcgcgctgagcgggggcccgggagccagcgcggggcaaactgggaaagtggtgtcgtgtgctggctcc
gccctcttcccgagggtgggggagaacggtataaaagtgcggtagtcgcgttggacgttctttttcgcaa
cgggtttgccgtcagaacgcaggtgagtggcgggtgtggcctccgcgggcccgggctccctcctttgagc
ggggtcggaccgccgtgcgggtgtcgtcggccgggcttctctgcgagcgttcccgccctggatggcgggc
tgtgcgggagggcgagggggggaggcctggcggcggccccggagcctcgcctcgtgtcgggcgtgaggcc
tagcgtggcttccgccccgccgcgtgccaccgcggccgcgctttgctgtctgcccggctgccctcgattg
cctgcccgcggcccgggccaacaaagggagggcgtggagctggctggtagggagccccgtagtccgcatg
tcgggcagggagagcggcagcagtcgggggggggaccgggcccgcccgtcccgcagcacatgtccgacgc
cgcctggacgggtagcggcctgtgtcctgataaggcggccgggcggtgggttttagatgccgggttcagg
tggccccgggtcccggcccggtctggccagtaccccgtagtggcttagctccgaggagggcgaggcccgc
ccgcccggcaccagttgcgtgcgcggaaagatggccgctcccgggccctgtagcaaggagctcaaaatgg
aggacgcggcagcccggcggagcggggcgggtgagtcacccacacaaaggaagagggccttgcccctcgc
cggccgctgcttcctgtgaccccgtggtgtaccggccgcacttcagtcaccccgggcgctctttcggagc
accgctggcctccgctgggggaggggatctgtctaatggcgttggagtttgctcacatttggtgggtgga
gactgtagccaggccagcctggccatgaaagtaattcttggaatttgcccattttgagtttggagcgaag
ctgattgacaaagctgcttagccgttcaaaggtattcttcgaactttttttttaaggtgttgtgaaaacc
tcgaggccgccaccatggattactacagaaaatatgcagctatctttctggtcacattgtcggtgtttct
gcatgttctccattccgctcctgatgtgcaggattgcccagaatgcacgctacaggaaaacccattcttc
tcccagccgggtgccccaatacttcagtgcatgggctgctgcttctctagagcatatcccactccactaa
ggtccaagaagacgatgttggtccaaaagaacgtcacctcagagtccacttgctgtgtagctaaatcata
taacagggtcacagtaatggggggtttcaaagtggagaaccacacggcgtgccactgcagtacttgttat
tatcacaaatcttaatagactagtccccccccctaacgttactggccgaagccgcttggaataaggccgg
tgtgcgtttgtctatatgttattttccaccatattgccgtcttttggcaatgtgagggcccggaaacctg
gccctgtcttcttgacgagcattcctaggggtctttcccctctcgccaaaggaatgcaaggtctgttgaa
tgtcgtgaaggaagcagttcctctggaagcttcttgaagacaaacaacgtctgtagcgaccctttgcagg
cagcggaaccccccacctggcgacaggtgcctctgcggccaaaagccacgtgtataagatacacctgcaa
aggcggcacaaccccagtgccacgttgtgagttggatagttgtggaaagagtcaaatggctctcctcaag
cgtattcaacaaggggctgaaggatgcccagaaggtaccccattgtatgggatctgatctggggcctcgg
tgcacatgctttacatgtgtttagtcgaggttaaaaaacgtctaggccccccgaaccacggggacgtggt
tttcctttgaaaaacacgatgataatatggagatgttccaggggctgctgctgttgctgctgctgagcat
gggcgggacatgggcatccaaggagccgcttcggccacggtgccgccccatcaatgccaccctggctgtg
gagaaggagggctgccccgtgtgcatcaccgtcaacaccaccatctgtgccggctactgccccaccatga
cccgcgtgctgcagggggtcctgccggccctgcctcaggtggtgtgcaactaccgcgatgtgcgcttcga
gtccatccggctccctggctgcccgcgcggcgtgaaccccgtggtctcctacgccgtggctctcagctgt
caatgtgcactctgccgccgcagcaccactgactgcgggggtcccaaggaccaccccttgacctgtgatg
acccccgcttccaggactcctcttcctcaaaggcccctccccccagccttccaagcccatcccgactccc
ggggccctcggacaccccgatcctcccacaataatagg</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="2">
<INSDSeq>
<INSDSeq_length>116</INSDSeq_length>
<INSDSeq_moltype>AA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..116</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>protein</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q36">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>MDYYRKYAAIFLVTLSVFLHVLHSAPDVQDCPECTLQENPFFSQPGAPI
LQCMGCCFSRAYPTPLRSKKTMLVQKNVTSESTCCVAKSYNRVTVMGGFKVENHTACHCSTCYYHKS</I
NSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="3">
<INSDSeq>
<INSDSeq_length>165</INSDSeq_length>
<INSDSeq_moltype>AA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..165</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>protein</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q20">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>MEMFQGLLLLLLLSMGGTWASKEPLRPRCRPINATLAVEKEGCPVCITV
NTTICAGYCPTMTRVLQGVLPALPQVVCNYRDVRFESIRLPGCPRGVNPVVSYAVALSCQCALCRRSTTD
CGGPKDHPLTCDDPRFQDSSSSKAPPPSLPSPSRLPGPSDTPILPQ</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="4">
<INSDSeq>
<INSDSeq_length>13291</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..13291</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q5">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
<INSDFeature>
<INSDFeature_key>CDS</INSDFeature_key>
<INSDFeature_location>12790..13284</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>protein_id</INSDQualifier_name>
<INSDQualifier_value>3</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier>
<INSDQualifier_name>translation</INSDQualifier_name>
<INSDQualifier_value>MEMFQGLLLLLLLSMGGTWASKEPLRPRCRPINATLAVEKEG
CPVCITVNTTICAGYCPTMTRVLQGVLPALPQVVCNYRDVRFESIRLPGCPRGVNPVVSYAVALSCQCAL
CRRSTTDCGGPKDHPLTCDDPRFQDSSSSKAPPPSLPSPSRLPGPSDTPILPQ</INSDQualifier_v
alue>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>ctagcatattacccctaacacctgccaccccagtcttaatcagtggtgg
aagaacggtctcagaactgtttgtctcaattggccatttaagtttaatagtgaaagactggttaatgata
acaatgcatcggaaaaccttcaggaggaaaggagaatgttttgtggaacatttttgtgtgtgtggcagtt
ttaagttattagttttcaaaatcagtactttttaatggaaacaacttgaccaaaaatctgtcacagaatt
ttgagacccattaaaatacaagtttaatgagaagtctgtctctgttaatgctgaagtcattactaagtgc
ttagcttagcaaggtatgtggatgcccatttgtgttccaagggattggactgttcatcaggacccagagc
tgagtttcaagggctcaagagatggcttattacctgtgggtgtcttgaaggttctggttgggacaaatta
ggaatgtttttggcagacatggtgactaccttcatctgggtgagttcagttgatttgtcttgagcctttg
gggtttacacaagtaaatgacatcatacagttagtgtattgttagtgaatattaatatatgaggcaggct
ttgctctagcaattttagaactagttttcaggaaagggttcatcttgtgcattggatgtttgattctatc
acttagagtttaaactgaaagtgctcaagaggttttatttaggctgggatataaataagcctttctgtag
cttgtaatggtatcaggaatttaaaaggccatctggggcacaaagattaagcagaaaaggtagaaggtga
gattgggggactttgagtacttcacacactttaatgtgtgagtgctttagtgcatatagtacaactgcca
gataagggcatccacatctgattgtttggaaggcaccttgtggtttctgggaattcagaattgggagaaa
aatgctcccaaccgctgaagcccttggtaatctgcagggtgtttatttagcaggagataaggacaaaaag
ttatagtgtggagttggttgagttggtagatgtcattacaacaggtggtcttaaattgggttaggagtca
ctttgaaatacctgggccataagcaaagtggcattttcacctttcaggagaaactggtacacttatccat
tctatagtgcatgcttgttcaattgggctgatgactaaaccggtgactaaaggtttgtcagtataaatgg
atgggttgtaggcagacggtgaggaattactatacctgcaaggagtcattgcctgatctgcctggaaagg
ggcaggattgagtctcagaaggtgtacaccataggatatggaaaaatttgtcacgcctagcattcaactt
agtggtgtagcgccacctactggcactttaaaagcttagcatagaggagcatgtgtgttaggagctcgga
tgggatccagggcctcaaggtttgcatgtaaataaaagccctttaccaaattaactacataccagcatac
atcagtcctttagtgttgaaaaacagaagtgaaagctaatatatatagtgcttgctttatttaagtctag
ctgattacgtgtttggttgccagtgtgactagtctggagttgaatttgtcctcagacacgtaaaatggaa
tttgggattcacaacactctagtatgagggacctaatggcctgtaccaggcacaaacgtgtctataaact
acacaaaacgaaggaatttacaggaattaggaaggtattcttaacattaaaacattatgggcattttaaa
aaaagctttgacaggatttctttgtcatggctgtcctggagctagttgtgtagaccaggctgggctgaaa
tcttgtctgcctgcctggcttggacacttttttattatgtatacaacattctgcttccatgtatatctgc
acattagaagacggcaccagatctcctaatggatggttgtgagccaccatgtggttgctgggaattgaac
tcaggacctctggaagagcagtgctcttaacctctgagccatctccagccccagcttgggcacattttta
atggctgggaaatcaaaccccctaggccttctgtcagtaatgaagggcttttggctaccgagagtaggat
ttaaggttattcggagctgcaggtctgcctcagtgcaggtttgggagtccagcatcttagaaaatgcagt
gaagccaagctgagctatattttgtttaaaaaaaaaataagtgggtaaagtgctgctgagcctgatgacc
aagctgggacacaagtagaagaacataggccaatgctctatattaaaagcatgggtcatttttaatgctc
ttgagaaggctatgcctacactactctcagccaccgcagcgtgtttaaattaaactagtttggaaatttt
ctttgggggtaagctatttaacctagtgccttggcaggtatactactgaactctcctcctcattcctttt
tgttttttaagaatttcagtcaggctcaggcagcccttaaacttgtgattaagcctgagaacagttacga
ttatgagcctattagtataccgatcaatatgtgaatttttttgggatgggggtcaggcctccctgcctcc
caaatactgggactaaaggctgcaccaccacaacctggctcttgaaatacttttctacattttttggggg
gcatgggtgggagagcagggtttctctgtattagccctggctctcctggaactctgtagaccaggctatt
cttgagctcagattagcctgtctctgcctcctaaattctgggattaaaggtgtgtgctactgctgcctgg
ctacaaagacatttttttttttcttaaatttaaaaacaaaagtggttcttttagaagggtggttggtgtt
ggcacatactccaagcactcaggttttgagtttgtcccaggaatgaagactgcattactgccgcccctcc
ctggtaagggctacacagagaaatcctatttggagcctgtcctggtaactcgctctgtagaccaggctgg
cctcgaactcaagagaaccacctgcctctgaatgctggtattaagggcaggcaccaccaacacccagcct
aaaaaatgtcttttttttaaagatttttttttttttttttacagaataaacattctgtttacaatattct
gcttctatgtatatctgcacactagaagagggcacccgatctcataatggatggttgtgagccaccaagt
ggttgctgggaattgaactcagaacctctggaagagcagtcagtgctcttaacctctgagccatctctcc
agcccctaaaaatggctcttgagatagggtctcaagtagtttgagactgagttggctatataaacaaggc
tggcacatagcaccatgtacagctgggtttagtttacatggggtgtttttgtctctggaggcaggaggat
catttgagcatagggagttaatagtgaggtcatgttttatctactcttctgaattgagaactaagctgat
gcaaagcaagtttgactgaagaagtccagtttatgagaacaagggtggaaactaatgtgtcaaagatggc
cttgcatgtgttttagatgatgacccagtcacttgggaattactggatgtgtaagacctatatcttgaca
ggagtgaacagtgtcttataggtcctatatgaaagaaatgagacatacccattttgtttcccctaagaat
tcacttttcctaacctggttcatgctatttaggttattttacttgcaaatcctagggtgctcccttaccc
agtattgcttatgtggcaccaaagtcactcactcccatgatttgcaagtctctgggaacttccatgacaa
cctagaatagcaactcaaatacattttctcagtaccaattttgaagaaaaaatattttgcaaaatagctg
tatggatgggtactaaatagtgaggttatctccagaaggcctatgaagaattaaggttgagttcagttga
gttcagcagcaagtttaaggttcatccatttttgtacagtgttttcctattacggtaagtgttttgcctg
caggaatatctgtaccacatgcttgcctggtacctatatcggccagaagagggctttggatcctctggac
ttgaattacagatgggtattagccaccatttaggtgctgggaattgaaaccaagtcctctggaagaacag
caagtgactcgacgtccggagctcaggcgcgcctgtggaatgtgtgtcagttagggtgtggaaagtcccc
aggctccccagcaggcagaagtatgcaaagcatgcatctcaattagtcagcaaccaggtgtggaaagtcc
ccaggctccccagcaggcagaagtatgcaaagcatgcatctcaattagtcagcaaccatagtcccgcccc
taactccgcccatcccgcccctaactccgcccagttccgcccattctccgccccatggctgactaatttt
ttttatttatgcagaggccgaggccgcctcggcctctgagctattccagaagtagtgaggaggctttttt
ggaggcctaggcttttgcaaaaagctcccgggagcttggatatccattttcgcatcgatgcttccaccat
ggccacctcagcaagttcccacttgaacaaaggcatcaagcagatgtacatgaacctgccccagggcgag
aagatccaactcatgtatatctgggttgatggtaccggggaagggctacgctgcaagacccgtactctgg
actgtgaccccaagtgtgtagaagagttacccgagtggaactttgatggttctagtacgtttcagtctga
aggctccaacagcgacatgtacctccatcctgtggccatgtttcgagaccccttccgcagagaccccaac
aagctggtgttctgcgaagtattcaagtataaccggaagcccgcagagaccaacctgaggcacagctgta
agcgtataatggacatggtgagcagccagcacccctggtttggaatggaacaggagtatactctcatggg
aacagacggccaccctttcggctggccttctaatggcttccctggaccccaaggaccctattactgcggt
gtgggagctgacaaggcttatggccgagatatcgtggaggctcactaccgggcctgcttgtatgctggaa
tcaagatcacagggacaaatgccgaggttatgcctgcccagtgggaattccagataggaccctgcgaagg
gatccgcatgggagatcatctctgggtagcccgttttatcttgcatcgggtatgcgaagactttggggtg
atagcaacctttgaccccaagcccattccagggaactggaatggggcaggctgccacaccaactttagca
ccaaggccatgcgggaggagaatggtctgaggtgcattgaggaggccattgataaactgagcaagaggca
ccagtaccacatccgtgcctacgaccccaaggggggcctggacaacgcccgccgtctgactggattccac
gaaacctccaacatcaacgacttttccgctggcgttgccaaccgcagcgccagtatccgcattccccgga
ttgtcggccaggagaagaagggttactttgaagaccgtcggccttctgccaattgcgacccctatgcggt
gacggaagccatcgtccgcacgtgtctcctcaacgaaactggcgacgagcccttccaatacaagaactaa
gcggactcgacttccagtgatcttgagcccttcctagttcaccccactcccaactgttccctctcccact
ggtccccactgtaactcaaaaggatggaataccaaggtctttttattccttgcgcccagttaatttttgc
ctttattggtcagaatagaggggtcaggttcttaatctctacacacccaaccccttctttcctagctatc
gccttcttgacgagttcttctgagcgggactctggggttcgaaatgaccgaccaagcgacgcccaacctg
ccatcacgagatttcgattccaccgccgccttctatgaaaggttgggcttcggaatcgttttccgggacg
ccggctggatgatcctccagcgcggggatctcatgctggagttcttcgcccaccccaacttgtttattgc
agcttataatggttacaaataaagcaatagcatcacaaatttcacaaataaagcatttttttcactgcat
tctagttgtggtttgtccaaactcatcaatgtatcttatcatgtctgtataccgtcgacctctaggcgcg
ccatggctcttccgcttcctcgctcactgactcgctgcgctcggtcgttcggctgcggcgagcggtatca
gctcactcaaaggcggtaatacggttatccacagaatcaggggataacgcaggaaagaacatgtgagcaa
aaggccagcaaaaggccaggaaccgtaaaaaggccgcgttgctggcgtttttccataggctccgcccccc
tgacgagcatcacaaaaatcgacgctcaagtcagaggtggcgaaacccgacaggactataaagataccag
gcgtttccccctggaagctccctcgtgcgctctcctgttccgaccctgccgcttaccggatacctgtccg
cctttctcccttcgggaagcgtggcgctttctcatagctcacgctgtaggtatctcagttcggtgtaggt
cgttcgctccaagctgggctgtgtgcacgaaccccccgttcagcccgaccgctgcgccttatccggtaac
tatcgtcttgagtccaacccggtaagacacgacttatcgccactggcagcagccactggtaacaggatta
gcagagcgaggtatgtaggcggtgctacagagttcttgaagtggtggcctaactacggctacactagaag
aacagtatttggtatctgcgctctgctgaagccagttaccttcggaaaaagagttggtagctcttgatcc
ggcaaacaaaccaccgctggtagcggtttttttgtttgcaagcagcagattacgcgcagaaaaaaaggat
ctcaagaagatcctttgatcttttctacggggtctgacgctcagtggaacgaaaactcacgttaagggat
tttggtcatgagattatcaaaaaggatcttcacctagatccttttaaattaaaaatgaagttttaaatca
atctaaagtatatatgagtaaacttggtctgacagttaccaatgcttaatcagtgaggcacctatctcag
cgatctgtctatttcgttcatccatagttgcctgactccccgtcgtgtagataactacgatacgggaggg
cttaccatctggccccagtgctgcaatgataccgcgagacccacgctcaccggctccagatttatcagca
ataaaccagccagccggaagggccgagcgcagaagtggtcctgcaactttatccgcctccatccagtcta
ttaattgttgccgggaagctagagtaagtagttcgccagttaatagtttgcgcaacgttgttgccattgc
tacaggcatcgtggtgtcacgctcgtcgtttggtatggcttcattcagctccggttcccaacgatcaagg
cgagttacatgatcccccatgttgtgcaaaaaagcggttagctccttcggtcctccgatcgttgtcagaa
gtaagttggccgcagtgttatcactcatggttatggcagcactgcataattctcttactgtcatgccatc
cgtaagatgcttttctgtgactggtgagtactcaaccaagtcattctgagaatagtgtatgcggcgaccg
agttgctcttgcccggcgtcaatacgggataataccgcgccacatagcagaactttaaaagtgctcatca
ttggaaaacgttcttcggggcgaaaactctcaaggatcttaccgctgttgagatccagttcgatgtaacc
cactcgtgcacccaactgatcttcagcatcttttactttcaccagcgtttctgggtgagcaaaaacagga
aggcaaaatgccgcaaaaaagggaataagggcgacacggaaatgttgaatactcatactcttcctttttc
aatattattgaagcatttatcagggttattgtctcatgagcggatacatatttgaatgtatttagaaaaa
taaacaaataggggttccgcgcacatttccccgaaaagtgccacctgcatctcatcacggtcacgcatgg
ctgcgcccgcgcagcgccccccgacccccccccgatccccggaacgccagccgccatccaccgcccgcag
cccccggagcccgcggaccccgaccccccgccgctcgcccgccgttgcgccgctctctgcgggggggcta
gggggccgcggggggaaggccacgccccctccactttttccaggaatgcgcggcatgacccgcccccccc
atgacccgccaacacccccccgatgccccccgggggtctttcctggggggcctttgtcagggttgcctgt
gtcacccacgggccccttggccacgcttgcctgggtcaccccggagccccgggtcacgtttgccggggtc
agccctggggcgcttcttcacgctccattgccatgcagaattccacgttgtgcatagaaacagatgcagg
caaaacatccacacatataaaacaaaaaattaaaaccaataaaactcctaaacttttggtctttcttgaa
tcttcaatccctcaggttatgaaataatcatttatgcagtcaaaaatttgccattcttgttgccaggtgt
ggtgatgattcggggaagcagaagcaggcagatctctgtgaatgaggccagcctggtctacaaagtgagt
cccaggacagtcaggcctgttacacagagaaaccttgaaaaaaaaaagataatatgtactgttgtattac
cccaatatataaggctaaaccattagaagcacaacactgttaagtacggaaaataatatctagtgtggta
cagttactactactataatacactaatatagctgtgggaaactagttccaaagatgaattactaaccagt
gtttccaaggaaataaatgaaagcagagagattagttctattgctagtgtttcattttcgtatatttctt
acaatttctcttgttacaaataggcactagggtatcaagataattttaacgactggctgagaaccctaga
aaatctctgtgaaaaagggatttgtgaaatgagagagggtaatgtggccattatagaaaaggcttttgtg
tgccttgcatgcatagaccctgtgtttgatctcttaacaccctccttgaccagaaaaagcttctgtggat
agaaaatgattagttatatatacttttagggaaacgtagttctggattctttggttacaattaacagaat
taagtgcaaacaaagccagaaacctcctgataaatgagaaaacctgcttgtagaaggttgtaaggctctg
taatataggaattaggagaaaagaaacctgtgtggtggggcacgtctgtaatcccagcattgggaagtag
aggtagaagattagaaatcaaaggccagcctcagcaacacagtgagtttgaggccaccctgaactacatc
aggttctgtctcctttcttttttttttttttcttttctttttttggtttctctgtgtagttttggagcct
atcctggcactagctctgaagagcaggctggcctcgaactcagagatcagccagcctctgctgggattaa
aggtatgcaccaccaacgccccaggttttgtctcaaacaaacaaaaataacatcaggaggtggtgagagg
gctcagtggtcacaggcattctctgcaaagcctgactctgagttggatcctttagagctacatggttgag
ggaagagaactgactcctggaaggtgtcctctggtccccacacatagctatacacagcatgtgcattcac
acacactaaataatgctatttttaaaaaaattaaaaacaacaacagtttgggttgtgaaaactagaacta
gataataggtaagaatcaagtatcatgtaaatttgctttcaactcatcccaaaatttgttttatatttca
gtttttttccttcctagcttgactgtggagtcttgtccggaagcaaatagttcctttgcagatcccacat
gtggacaccggacagtaggtcctcaaatgctccttattaggttggttcaataatatcaattgtttgttac
taggcagtgatgttgtacatctggaggagatctcttgagcccataatcaggttattaggaataaatactc
taaggctaaaaatgtagcttagtgataagagtgcttgcctggtgtgctgagaccctcggttccatctcca
caaccccatattccattacaaaataccttttcaccgtccctagcattaagaaacaaaacaacaaagaagt
ttttctttcttctgagatcctgcccggagaggcatttaaaactggccagggccaaaaaaaaaaaaaaaaa
aagaaaaaaaagaaaagaaaacaggctagggccggcatggtggcgcacgcctttaatcccagcacgcagg
aggcagaggcagggcggatctctgtgagtttgaggtcagcctggtctacctagtgagtttcagggcaccc
agggctaaagagactgtctcaaaaacaaaacagccacacaatcagaaccacagcaaaacgcagttatgat
ccttggaactgtaggaatgacaagcatttaaataataggacgagccatttttgagaagctctgatttcac
aagtgtcagggatgggctctgggcgagtaagattgctaatgctggcctctaaatgagaccacgtggagtt
gattagattcttttcatgttcctcgtgctctatcaaataactgtacccaaatacacacacacacacacac
acacaatgcgcgcacacacaaaatccttttttagcttaagaagcccagaatcagaagtaaagctaactgt
gggacttaagtattattctgaacggaactcccagggcgtgaagcgcgcttcaggcttccagagaagcagc
tggcgctggatggaatgaaccaagaggccagcacaggggcagatccgtcgagctctcggccaccgagctg
agcccttaggttctggggctgggaagggtccctaggattgtgcacctctcccgcgggggacaagcagggg
gatggcggggctgacgtcgggaggtggcctccacgggaagggacacccggatctcgacacagccttggca
gtggagtcaggaagggtagggacagattctggacgccctcttggccagctcctcaccgccccacccccga
ctggagccgagagtaattcatacaaaaggagggatcgccttcgcccctgggaatcccagggaccgtcgct
aaattctggccggcctcccagcccggaaccgctgtgcccgcccagcgcggcgggaggagcctgcgcctag
ggcggatcgcgggtcggcgggagagcacaagcccacagtccccggcggtgggggaggggcgcgctgagcg
ggggcccgggagccagcgcggggcaaactgggaaagtggtgtcgtgtgctggctccgccctcttcccgag
ggtgggggagaacggtataaaagtgcggtagtcgcgttggacgttctttttcgcaacgggtttgccgtca
gaacgcaggtgagtggcgggtgtggcctccgcgggcccgggctccctcctttgagcggggtcggaccgcc
gtgcgggtgtcgtcggccgggcttctctgcgagcgttcccgccctggatggcgggctgtgcgggagggcg
agggggggaggcctggcggcggccccggagcctcgcctcgtgtcgggcgtgaggcctagcgtggcttccg
ccccgccgcgtgccaccgcggccgcgctttgctgtctgcccggctgccctcgattgcctgcccgcggccc
gggccaacaaagggagggcgtggagctggctggtagggagccccgtagtccgcatgtcgggcagggagag
cggcagcagtcgggggggggaccgggcccgcccgtcccgcagcacatgtccgacgccgcctggacgggta
gcggcctgtgtcctgataaggcggccgggcggtgggttttagatgccgggttcaggtggccccgggtccc
ggcccggtctggccagtaccccgtagtggcttagctccgaggagggcgaggcccgcccgcccggcaccag
ttgcgtgcgcggaaagatggccgctcccgggccctgtagcaaggagctcaaaatggaggacgcggcagcc
cggcggagcggggcgggtgagtcacccacacaaaggaagagggccttgcccctcgccggccgctgcttcc
tgtgaccccgtggtgtaccggccgcacttcagtcaccccgggcgctctttcggagcaccgctggcctccg
ctgggggaggggatctgtctaatggcgttggagtttgctcacatttggtgggtggagactgtagccaggc
cagcctggccatgaaagtaattcttggaatttgcccattttgagtttggagcgaagctgattgacaaagc
tgcttagccgttcaaaggtattcttcgaactttttttttaaggtgttgtgaaaacctcgaggccgccacc
atggagatgttccaggggctgctgctgttgctgctgctgagcatgggcgggacatgggcatccaaggagc
cgcttcggccacggtgccgccccatcaatgccaccctggctgtggagaaggagggctgccccgtgtgcat
caccgtcaacaccaccatctgtgccggctactgccccaccatgacccgcgtgctgcagggggtcctgccg
gccctgcctcaggtggtgtgcaactaccgcgatgtgcgcttcgagtccatccggctccctggctgcccgc
gcggcgtgaaccccgtggtctcctacgccgtggctctcagctgtcaatgtgcactctgccgccgcagcac
cactgactgcgggggtcccaaggaccaccccttgacctgtgatgacccccgcttccaggactcctcttcc
tcaaaggcccctccccccagccttccaagcccatcccgactcccggggccctcggacaccccgatcctcc
cacaataataga</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="5">
<INSDSeq>
<INSDSeq_length>8137</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..8137</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q29">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>ctagcggccgcgcagttaacgccgcccctctccctcccccccccctaac
gttactggccgaagccgcttggaataaggccggtgtgcgtttgtctatatgttattttccaccatattgc
cgtcttttggcaatgtgagggcccggaaacctggccctgtcttcttgacgagcattcctaggggtctttc
ccctctcgccaaaggaatgcaaggtctgttgaatgtcgtgaaggaagcagttcctctggaagcttcttga
agacaaacaacgtctgtagcgaccctttgcaggcagcggaaccccccacctggcgacaggtgcctctgcg
gccaaaagccacgtgtataagatacacctgcaaaggcggcacaaccccagtgccacgttgtgagttggat
agttgtggaaagagtcaaatggctctcctcaagcgtattcaacaaggggctgaaggatgcccagaaggta
ccccattgtatgggatctgatctggggcctcggtacacatgctttacatgtgtttagtcgaggttaaaaa
aacgtctaggccccccgaaccacggggacgtggttttcctttgaaaaacacgatgataatatggccacaa
gatctgccaccatggttcgaccattgaactgcatcgtcgccgtgtcccaaaatatggggattggcaagaa
cggagacctaccctggcctccgctcaggaacgagttcaagtacttccaaagaatgaccacaacctcttca
gtggaaggtaaacagaatctggtgattatgggtaggaaaacctggttctccattcctgagaagaatcgac
ctctaaaggacagaattaatatagttctcagtagagaactcaaagaaccaccacgaggagctcattttct
tgccaaaagtttggatgatgccttaagacttattgaacaaccggaattggcaagtaaagtagacatggtt
tggatagtcggaggcagttctgtttaccaggaagccatgaatcaaccaggccacctcagactctttgtga
caaggatcatgcaggaatttgaaagtgacacgtttttcccagaaattgatttggggaaatataaacttct
cccagaatacccaggcgtcctctctgaggtccaggaggaaaaaggcatcaagtataagtttgaagtctac
gagaagaaagactaatcatattacccctaacacctgccaccccagtcttaatcagtggtggaagaacggt
ctcagaactgtttgtctcaattggccatttaagtttaatagtgaaagactggttaatgataacaatgcat
cggaaaaccttcaggaggaaaggagaatgttttgtggaacatttttgtgtgtgtggcagttttaagttat
tagttttcaaaatcagtactttttaatggaaacaacttgaccaaaaatctgtcacagaattttgagaccc
attaaaatacaagtttaatgagaagtctgtctctgttaatgctgaagtcattactaagtgcttagcttag
caaggtatgtggatgcccatttgtgttccaagggattggactgttcatcaggacccagagctgagtttca
agggctcaagagatggcttattacctgtgggtgtcttgaaggttctggttgggacaaattaggaatgttt
ttggcagacatggtgactacccttcatctgggtgagttcagttgatttgtcttgagcctttggggtttac
acaagtaaatgacatcatacagttagtgtattgttagtgaatattaatatatgaggcaggctttgctcta
gctagacatatgtcgacgtccggagctcaggcgcgccatggctcttccgcttcctcgctcactgactcgc
tgcgctcggtcgttcggctgcggcgagcggtatcagctcactcaaaggcggtaatacggttatccacaga
atcaggggataacgcaggaaagaacatgtgagcaaaaggccagcaaaaggccaggaaccgtaaaaaggcc
gcgttgctggcgtttttccataggctccgcccccctgacgagcatcacaaaaatcgacgctcaagtcaga
ggtggcgaaacccgacaggactataaagataccaggcgtttccccctggaagctccctcgtgcgctctcc
tgttccgaccctgccgcttaccggatacctgtccgcctttctcccttcgggaagcgtggcgctttctcat
agctcacgctgtaggtatctcagttcggtgtaggtcgttcgctccaagctgggctgtgtgcacgaacccc
ccgttcagcccgaccgctgcgccttatccggtaactatcgtcttgagtccaacccggtaagacacgactt
atcgccactggcagcagccactggtaacaggattagcagagcgaggtatgtaggcggtgctacagagttc
ttgaagtggtggcctaactacggctacactagaagaacagtatttggtatctgcgctctgctgaagccag
ttaccttcggaaaaagagttggtagctcttgatccggcaaacaaaccaccgctggtagcggtttttttgt
ttgcaagcagcagattacgcgcagaaaaaaaggatctcaagaagatcctttgatcttttctacggggtct
gacgctcagtggaacgaaaactcacgttaagggattttggtcatgagattatcaaaaaggatcttcacct
agatccttttaaattaaaaatgaagttttaaatcaatctaaagtatatatgagtaaacttggtctgacag
ttaccaatgcttaatcagtgaggcacctatctcagcgatctgtctatttcgttcatccatagttgcctga
ctccccgtcgtgtagataactacgatacgggagggcttaccatctggccccagtgctgcaatgataccgc
gagacccacgctcaccggctccagatttatcagcaataaaccagccagccggaagggccgagcgcagaag
tggtcctgcaactttatccgcctccatccagtctattaattgttgccgggaagctagagtaagtagttcg
ccagttaatagtttgcgcaacgttgttgccattgctacaggcatcgtggtgtcacgctcgtcgtttggta
tggcttcattcagctccggttcccaacgatcaaggcgagttacatgatcccccatgttgtgcaaaaaagc
ggttagctccttcggtcctccgatcgttgtcagaagtaagttggccgcagtgttatcactcatggttatg
gcagcactgcataattctcttactgtcatgccatccgtaagatgcttttctgtgactggtgagtactcaa
ccaagtcattctgagaatagtgtatgcggcgaccgagttgctcttgcccggcgtcaatacgggataatac
cgcgccacatagcagaactttaaaagtgctcatcattggaaaacgttcttcggggcgaaaactctcaagg
atcttaccgctgttgagatccagttcgatgtaacccactcgtgcacccaactgatcttcagcatctttta
ctttcaccagcgtttctgggtgagcaaaaacaggaaggcaaaatgccgcaaaaaagggaataagggcgac
acggaaatgttgaatactcatactcttcctttttcaatattattgaagcatttatcagggttattgtctc
atgagcggatacatatttgaatgtatttagaaaaataaacaaataggggttccgcgcacatttccccgaa
aagtgccacctgcatctcatcacggtcacgcatggctgcgcccgcgcagcgccccccgacccccccccga
tccccggaacgccagccgccatccaccgcccgcagcccccggagcccgcggaccccgaccccccgccgct
cgcccgccgttgcgccgctctctgcgggggggctagggggccgcggggggaaggccacgccccctccact
ttttccaggaatgcgcggcatgacccgccccccccatgacccgccaacacccccccgatgccccccgggg
gtctttcctggggggcctttgtcagggttgcctgtgtcacccacgggccccttggccacgcttgcctggg
tcaccccggagccccgggtcacgtttgccggggtcagccctggggcgcttcttcacgctccattgccatg
cagaattcccacatgtggacaccggacagtaggtcctcaaatgctccttattaggttggttcaataatat
caattgtttgttactaggcagtgatgttgtacatctggaggagatctcttgagcccataatcaggttatt
aggaataaatactctaaggctaaaaatgtagcttagtgataagagtgcttgcctggtgtgctgagaccct
cggttccatctccacaaccccatattccattacaaaataccttttcaccgtccctagcattaagaaacaa
aacaacaaagaagtttttctttcttctgagatcctgcccggagaggcatttaaaactggccagggccaaa
aaaaaaaaaaaaaaaagaaaaaaaagaaaagaaaacaggctagggccggcatggtggcgcacgcctttaa
tcccagcacgcaggaggcagaggcagggcggatctctgtgagtttgaggtcagcctggtctacctagtga
gtttcagggcacccagggctaaagagactgtctcaaaaacaaaacagccacacaatcagaaccacagcaa
aacgcagttatgatccttggaactgtaggaatgacaagcatttaaataataggacgagccatttttgaga
agctctgatttcacaagtgtcagggatgggctctgggcgagtaagattgctaatgctggcctctaaatga
gaccacgtggagttgattagattcttttcatgttcctcgtgctctatcaaataactgtacccaaatacac
acacacacacacacacacaatgcgcgcacacacaaaatccttttttagcttaagaagcccagaatcagaa
gtaaagctaactgtgggacttaagtattattctgaacggaactcccagggcgtgaagcgcgcttcaggct
tccagagaagcagctggcgctggatggaatgaaccaagaggccagcacaggggcagatccgtcgagctct
cggccaccgagctgagcccttaggttctggggctgggaagggtccctaggattgtgcacctctcccgcgg
gggacaagcagggggatggcggggctgacgtcgggaggtggcctccacgggaagggacacccggatctcg
acacagccttggcagtggagtcaggaagggtagggacagattctggacgccctcttggccagctcctcac
cgccccacccccgactggagccgagagtaattcatacaaaaggagggatcgccttcgcccctgggaatcc
cagggaccgtcgctaaattctggccggcctcccagcccggaaccgctgtgcccgcccagcgcggcgggag
gagcctgcgcctagggcggatcgcgggtcggcgggagagcacaagcccacagtccccggcggtgggggag
gggcgcgctgagcgggggcccgggagccagcgcggggcaaactgggaaagtggtgtcgtgtgctggctcc
gccctcttcccgagggtgggggagaacggtataaaagtgcggtagtcgcgttggacgttctttttcgcaa
cgggtttgccgtcagaacgcaggtgagtggcgggtgtggcctccgcgggcccgggctccctcctttgagc
ggggtcggaccgccgtgcgggtgtcgtcggccgggcttctctgcgagcgttcccgccctggatggcgggc
tgtgcgggagggcgagggggggaggcctggcggcggccccggagcctcgcctcgtgtcgggcgtgaggcc
tagcgtggcttccgccccgccgcgtgccaccgcggccgcgctttgctgtctgcccggctgccctcgattg
cctgcccgcggcccgggccaacaaagggagggcgtggagctggctggtagggagccccgtagtccgcatg
tcgggcagggagagcggcagcagtcgggggggggaccgggcccgcccgtcccgcagcacatgtccgacgc
cgcctggacgggtagcggcctgtgtcctgataaggcggccgggcggtgggttttagatgccgggttcagg
tggccccgggtcccggcccggtctggccagtaccccgtagtggcttagctccgaggagggcgaggcccgc
ccgcccggcaccagttgcgtgcgcggaaagatggccgctcccgggccctgtagcaaggagctcaaaatgg
aggacgcggcagcccggcggagcggggcgggtgagtcacccacacaaaggaagagggccttgcccctcgc
cggccgctgcttcctgtgaccccgtggtgtaccggccgcacttcagtcaccccgggcgctctttcggagc
accgctggcctccgctgggggaggggatctgtctaatggcgttggagtttgctcacatttggtgggtgga
gactgtagccaggccagcctggccatgaaagtaattcttggaatttgcccattttgagtttggagcgaag
ctgattgacaaagctgcttagccgttcaaaggtattcttcgaactttttttttaaggtgttgtgaaaacc
tcgaggccgccaccatggagatgttccaggggctgctgctgttgctgctgctgagcatgggcgggacatg
ggcatccaaggagccgcttcggccacggtgccgccccatcaatgccaccctggctgtggagaaggagggc
tgccccgtgtgcatcaccgtcaacaccaccatctgtgccggctactgccccaccatgacccgcgtgctgc
agggggtcctgccggccctgcctcaggtggtgtgcaactaccgcgatgtgcgcttcgagtccatccggct
ccctggctgcccgcgcggcgtgaaccccgtggtctcctacgccgtggctctcagctgtcaatgtgcactc
tgccgccgcagcaccactgactgcgggggtcccaaggaccaccccttgacctgtgatgacccccgcttcc
aggactcctcttcctcaaaggcccctccccccagccttccaagcccatcccgactcccggggccctcgga
caccccgatcctcccacaataatagactagtccccccccctaacgttactggccgaagccgcttggaata
aggccggtgtgcgtttgtctatatgttattttccaccatattgccgtcttttggcaatgtgagggcccgg
aaacctggccctgtcttcttgacgagcattcctaggggtctttcccctctcgccaaaggaatgcaaggtc
tgttgaatgtcgtgaaggaagcagttcctctggaagcttcttgaagacaaacaacgtctgtagcgaccct
ttgcaggcagcggaaccccccacctggcgacaggtgcctctgcggccaaaagccacgtgtataagataca
cctgcaaaggcggcacaaccccagtgccacgttgtgagttggatagttgtggaaagagtcaaatggctct
cctcaagcgtattcaacaaggggctgaaggatgcccagaaggtaccccattgtatgggatctgatctggg
gcctcggtgcacatgctttacatgtgtttagtcgaggttaaaaaacgtctaggccccccgaaccacgggg
acgtggttttcctttgaaaaacacgatgataatatggattactacagaaaatatgcagctatctttctgg
tcacattgtcggtgtttctgcatgttctccattccgctcctgatgtgcaggattgcccagaatgcacgct
acaggaaaacccattcttctcccagccgggtgccccaatacttcagtgcatgggctgctgcttctctaga
gcatatcccactccactaaggtccaagaagacgatgttggtccaaaagaacgtcacctcagagtccactt
gctgtgtagctaaatcatataacagggtcacagtaatggggggtttcaaagtggagaaccacacggcgtg
ccactgcagtacttgttattatcacaaatcttaatagg</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="6">
<INSDSeq>
<INSDSeq_length>13144</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..13144</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q35">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>ctagcatattacccctaacacctgccaccccagtcttaatcagtggtgg
aagaacggtctcagaactgtttgtctcaattggccatttaagtttaatagtgaaagactggttaatgata
acaatgcatcggaaaaccttcaggaggaaaggagaatgttttgtggaacatttttgtgtgtgtggcagtt
ttaagttattagttttcaaaatcagtactttttaatggaaacaacttgaccaaaaatctgtcacagaatt
ttgagacccattaaaatacaagtttaatgagaagtctgtctctgttaatgctgaagtcattactaagtgc
ttagcttagcaaggtatgtggatgcccatttgtgttccaagggattggactgttcatcaggacccagagc
tgagtttcaagggctcaagagatggcttattacctgtgggtgtcttgaaggttctggttgggacaaatta
ggaatgtttttggcagacatggtgactaccttcatctgggtgagttcagttgatttgtcttgagcctttg
gggtttacacaagtaaatgacatcatacagttagtgtattgttagtgaatattaatatatgaggcaggct
ttgctctagcaattttagaactagttttcaggaaagggttcatcttgtgcattggatgtttgattctatc
acttagagtttaaactgaaagtgctcaagaggttttatttaggctgggatataaataagcctttctgtag
cttgtaatggtatcaggaatttaaaaggccatctggggcacaaagattaagcagaaaaggtagaaggtga
gattgggggactttgagtacttcacacactttaatgtgtgagtgctttagtgcatatagtacaactgcca
gataagggcatccacatctgattgtttggaaggcaccttgtggtttctgggaattcagaattgggagaaa
aatgctcccaaccgctgaagcccttggtaatctgcagggtgtttatttagcaggagataaggacaaaaag
ttatagtgtggagttggttgagttggtagatgtcattacaacaggtggtcttaaattgggttaggagtca
ctttgaaatacctgggccataagcaaagtggcattttcacctttcaggagaaactggtacacttatccat
tctatagtgcatgcttgttcaattgggctgatgactaaaccggtgactaaaggtttgtcagtataaatgg
atgggttgtaggcagacggtgaggaattactatacctgcaaggagtcattgcctgatctgcctggaaagg
ggcaggattgagtctcagaaggtgtacaccataggatatggaaaaatttgtcacgcctagcattcaactt
agtggtgtagcgccacctactggcactttaaaagcttagcatagaggagcatgtgtgttaggagctcgga
tgggatccagggcctcaaggtttgcatgtaaataaaagccctttaccaaattaactacataccagcatac
atcagtcctttagtgttgaaaaacagaagtgaaagctaatatatatagtgcttgctttatttaagtctag
ctgattacgtgtttggttgccagtgtgactagtctggagttgaatttgtcctcagacacgtaaaatggaa
tttgggattcacaacactctagtatgagggacctaatggcctgtaccaggcacaaacgtgtctataaact
acacaaaacgaaggaatttacaggaattaggaaggtattcttaacattaaaacattatgggcattttaaa
aaaagctttgacaggatttctttgtcatggctgtcctggagctagttgtgtagaccaggctgggctgaaa
tcttgtctgcctgcctggcttggacacttttttattatgtatacaacattctgcttccatgtatatctgc
acattagaagacggcaccagatctcctaatggatggttgtgagccaccatgtggttgctgggaattgaac
tcaggacctctggaagagcagtgctcttaacctctgagccatctccagccccagcttgggcacattttta
atggctgggaaatcaaaccccctaggccttctgtcagtaatgaagggcttttggctaccgagagtaggat
ttaaggttattcggagctgcaggtctgcctcagtgcaggtttgggagtccagcatcttagaaaatgcagt
gaagccaagctgagctatattttgtttaaaaaaaaaataagtgggtaaagtgctgctgagcctgatgacc
aagctgggacacaagtagaagaacataggccaatgctctatattaaaagcatgggtcatttttaatgctc
ttgagaaggctatgcctacactactctcagccaccgcagcgtgtttaaattaaactagtttggaaatttt
ctttgggggtaagctatttaacctagtgccttggcaggtatactactgaactctcctcctcattcctttt
tgttttttaagaatttcagtcaggctcaggcagcccttaaacttgtgattaagcctgagaacagttacga
ttatgagcctattagtataccgatcaatatgtgaatttttttgggatgggggtcaggcctccctgcctcc
caaatactgggactaaaggctgcaccaccacaacctggctcttgaaatacttttctacattttttggggg
gcatgggtgggagagcagggtttctctgtattagccctggctctcctggaactctgtagaccaggctatt
cttgagctcagattagcctgtctctgcctcctaaattctgggattaaaggtgtgtgctactgctgcctgg
ctacaaagacatttttttttttcttaaatttaaaaacaaaagtggttcttttagaagggtggttggtgtt
ggcacatactccaagcactcaggttttgagtttgtcccaggaatgaagactgcattactgccgcccctcc
ctggtaagggctacacagagaaatcctatttggagcctgtcctggtaactcgctctgtagaccaggctgg
cctcgaactcaagagaaccacctgcctctgaatgctggtattaagggcaggcaccaccaacacccagcct
aaaaaatgtcttttttttaaagatttttttttttttttttacagaataaacattctgtttacaatattct
gcttctatgtatatctgcacactagaagagggcacccgatctcataatggatggttgtgagccaccaagt
ggttgctgggaattgaactcagaacctctggaagagcagtcagtgctcttaacctctgagccatctctcc
agcccctaaaaatggctcttgagatagggtctcaagtagtttgagactgagttggctatataaacaaggc
tggcacatagcaccatgtacagctgggtttagtttacatggggtgtttttgtctctggaggcaggaggat
catttgagcatagggagttaatagtgaggtcatgttttatctactcttctgaattgagaactaagctgat
gcaaagcaagtttgactgaagaagtccagtttatgagaacaagggtggaaactaatgtgtcaaagatggc
cttgcatgtgttttagatgatgacccagtcacttgggaattactggatgtgtaagacctatatcttgaca
ggagtgaacagtgtcttataggtcctatatgaaagaaatgagacatacccattttgtttcccctaagaat
tcacttttcctaacctggttcatgctatttaggttattttacttgcaaatcctagggtgctcccttaccc
agtattgcttatgtggcaccaaagtcactcactcccatgatttgcaagtctctgggaacttccatgacaa
cctagaatagcaactcaaatacattttctcagtaccaattttgaagaaaaaatattttgcaaaatagctg
tatggatgggtactaaatagtgaggttatctccagaaggcctatgaagaattaaggttgagttcagttga
gttcagcagcaagtttaaggttcatccatttttgtacagtgttttcctattacggtaagtgttttgcctg
caggaatatctgtaccacatgcttgcctggtacctatatcggccagaagagggctttggatcctctggac
ttgaattacagatgggtattagccaccatttaggtgctgggaattgaaaccaagtcctctggaagaacag
caagtgactcgacgtccggagctcaggcgcgcctgtggaatgtgtgtcagttagggtgtggaaagtcccc
aggctccccagcaggcagaagtatgcaaagcatgcatctcaattagtcagcaaccaggtgtggaaagtcc
ccaggctccccagcaggcagaagtatgcaaagcatgcatctcaattagtcagcaaccatagtcccgcccc
taactccgcccatcccgcccctaactccgcccagttccgcccattctccgccccatggctgactaatttt
ttttatttatgcagaggccgaggccgcctcggcctctgagctattccagaagtagtgaggaggctttttt
ggaggcctaggcttttgcaaaaagctcccgggagcttggatatccattttcgcatcgatgcttccaccat
ggccacctcagcaagttcccacttgaacaaaggcatcaagcagatgtacatgaacctgccccagggcgag
aagatccaactcatgtatatctgggttgatggtaccggggaagggctacgctgcaagacccgtactctgg
actgtgaccccaagtgtgtagaagagttacccgagtggaactttgatggttctagtacgtttcagtctga
aggctccaacagcgacatgtacctccatcctgtggccatgtttcgagaccccttccgcagagaccccaac
aagctggtgttctgcgaagtattcaagtataaccggaagcccgcagagaccaacctgaggcacagctgta
agcgtataatggacatggtgagcagccagcacccctggtttggaatggaacaggagtatactctcatggg
aacagacggccaccctttcggctggccttctaatggcttccctggaccccaaggaccctattactgcggt
gtgggagctgacaaggcttatggccgagatatcgtggaggctcactaccgggcctgcttgtatgctggaa
tcaagatcacagggacaaatgccgaggttatgcctgcccagtgggaattccagataggaccctgcgaagg
gatccgcatgggagatcatctctgggtagcccgttttatcttgcatcgggtatgcgaagactttggggtg
atagcaacctttgaccccaagcccattccagggaactggaatggggcaggctgccacaccaactttagca
ccaaggccatgcgggaggagaatggtctgaggtgcattgaggaggccattgataaactgagcaagaggca
ccagtaccacatccgtgcctacgaccccaaggggggcctggacaacgcccgccgtctgactggattccac
gaaacctccaacatcaacgacttttccgctggcgttgccaaccgcagcgccagtatccgcattccccgga
ttgtcggccaggagaagaagggttactttgaagaccgtcggccttctgccaattgcgacccctatgcggt
gacggaagccatcgtccgcacgtgtctcctcaacgaaactggcgacgagcccttccaatacaagaactaa
gcggactcgacttccagtgatcttgagcccttcctagttcaccccactcccaactgttccctctcccact
ggtccccactgtaactcaaaaggatggaataccaaggtctttttattccttgcgcccagttaatttttgc
ctttattggtcagaatagaggggtcaggttcttaatctctacacacccaaccccttctttcctagctatc
gccttcttgacgagttcttctgagcgggactctggggttcgaaatgaccgaccaagcgacgcccaacctg
ccatcacgagatttcgattccaccgccgccttctatgaaaggttgggcttcggaatcgttttccgggacg
ccggctggatgatcctccagcgcggggatctcatgctggagttcttcgcccaccccaacttgtttattgc
agcttataatggttacaaataaagcaatagcatcacaaatttcacaaataaagcatttttttcactgcat
tctagttgtggtttgtccaaactcatcaatgtatcttatcatgtctgtataccgtcgacctctaggcgcg
ccatggctcttccgcttcctcgctcactgactcgctgcgctcggtcgttcggctgcggcgagcggtatca
gctcactcaaaggcggtaatacggttatccacagaatcaggggataacgcaggaaagaacatgtgagcaa
aaggccagcaaaaggccaggaaccgtaaaaaggccgcgttgctggcgtttttccataggctccgcccccc
tgacgagcatcacaaaaatcgacgctcaagtcagaggtggcgaaacccgacaggactataaagataccag
gcgtttccccctggaagctccctcgtgcgctctcctgttccgaccctgccgcttaccggatacctgtccg
cctttctcccttcgggaagcgtggcgctttctcatagctcacgctgtaggtatctcagttcggtgtaggt
cgttcgctccaagctgggctgtgtgcacgaaccccccgttcagcccgaccgctgcgccttatccggtaac
tatcgtcttgagtccaacccggtaagacacgacttatcgccactggcagcagccactggtaacaggatta
gcagagcgaggtatgtaggcggtgctacagagttcttgaagtggtggcctaactacggctacactagaag
aacagtatttggtatctgcgctctgctgaagccagttaccttcggaaaaagagttggtagctcttgatcc
ggcaaacaaaccaccgctggtagcggtttttttgtttgcaagcagcagattacgcgcagaaaaaaaggat
ctcaagaagatcctttgatcttttctacggggtctgacgctcagtggaacgaaaactcacgttaagggat
tttggtcatgagattatcaaaaaggatcttcacctagatccttttaaattaaaaatgaagttttaaatca
atctaaagtatatatgagtaaacttggtctgacagttaccaatgcttaatcagtgaggcacctatctcag
cgatctgtctatttcgttcatccatagttgcctgactccccgtcgtgtagataactacgatacgggaggg
cttaccatctggccccagtgctgcaatgataccgcgagacccacgctcaccggctccagatttatcagca
ataaaccagccagccggaagggccgagcgcagaagtggtcctgcaactttatccgcctccatccagtcta
ttaattgttgccgggaagctagagtaagtagttcgccagttaatagtttgcgcaacgttgttgccattgc
tacaggcatcgtggtgtcacgctcgtcgtttggtatggcttcattcagctccggttcccaacgatcaagg
cgagttacatgatcccccatgttgtgcaaaaaagcggttagctccttcggtcctccgatcgttgtcagaa
gtaagttggccgcagtgttatcactcatggttatggcagcactgcataattctcttactgtcatgccatc
cgtaagatgcttttctgtgactggtgagtactcaaccaagtcattctgagaatagtgtatgcggcgaccg
agttgctcttgcccggcgtcaatacgggataataccgcgccacatagcagaactttaaaagtgctcatca
ttggaaaacgttcttcggggcgaaaactctcaaggatcttaccgctgttgagatccagttcgatgtaacc
cactcgtgcacccaactgatcttcagcatcttttactttcaccagcgtttctgggtgagcaaaaacagga
aggcaaaatgccgcaaaaaagggaataagggcgacacggaaatgttgaatactcatactcttcctttttc
aatattattgaagcatttatcagggttattgtctcatgagcggatacatatttgaatgtatttagaaaaa
taaacaaataggggttccgcgcacatttccccgaaaagtgccacctgcatctcatcacggtcacgcatgg
ctgcgcccgcgcagcgccccccgacccccccccgatccccggaacgccagccgccatccaccgcccgcag
cccccggagcccgcggaccccgaccccccgccgctcgcccgccgttgcgccgctctctgcgggggggcta
gggggccgcggggggaaggccacgccccctccactttttccaggaatgcgcggcatgacccgcccccccc
atgacccgccaacacccccccgatgccccccgggggtctttcctggggggcctttgtcagggttgcctgt
gtcacccacgggccccttggccacgcttgcctgggtcaccccggagccccgggtcacgtttgccggggtc
agccctggggcgcttcttcacgctccattgccatgcagaattccacgttgtgcatagaaacagatgcagg
caaaacatccacacatataaaacaaaaaattaaaaccaataaaactcctaaacttttggtctttcttgaa
tcttcaatccctcaggttatgaaataatcatttatgcagtcaaaaatttgccattcttgttgccaggtgt
ggtgatgattcggggaagcagaagcaggcagatctctgtgaatgaggccagcctggtctacaaagtgagt
cccaggacagtcaggcctgttacacagagaaaccttgaaaaaaaaaagataatatgtactgttgtattac
cccaatatataaggctaaaccattagaagcacaacactgttaagtacggaaaataatatctagtgtggta
cagttactactactataatacactaatatagctgtgggaaactagttccaaagatgaattactaaccagt
gtttccaaggaaataaatgaaagcagagagattagttctattgctagtgtttcattttcgtatatttctt
acaatttctcttgttacaaataggcactagggtatcaagataattttaacgactggctgagaaccctaga
aaatctctgtgaaaaagggatttgtgaaatgagagagggtaatgtggccattatagaaaaggcttttgtg
tgccttgcatgcatagaccctgtgtttgatctcttaacaccctccttgaccagaaaaagcttctgtggat
agaaaatgattagttatatatacttttagggaaacgtagttctggattctttggttacaattaacagaat
taagtgcaaacaaagccagaaacctcctgataaatgagaaaacctgcttgtagaaggttgtaaggctctg
taatataggaattaggagaaaagaaacctgtgtggtggggcacgtctgtaatcccagcattgggaagtag
aggtagaagattagaaatcaaaggccagcctcagcaacacagtgagtttgaggccaccctgaactacatc
aggttctgtctcctttcttttttttttttttcttttctttttttggtttctctgtgtagttttggagcct
atcctggcactagctctgaagagcaggctggcctcgaactcagagatcagccagcctctgctgggattaa
aggtatgcaccaccaacgccccaggttttgtctcaaacaaacaaaaataacatcaggaggtggtgagagg
gctcagtggtcacaggcattctctgcaaagcctgactctgagttggatcctttagagctacatggttgag
ggaagagaactgactcctggaaggtgtcctctggtccccacacatagctatacacagcatgtgcattcac
acacactaaataatgctatttttaaaaaaattaaaaacaacaacagtttgggttgtgaaaactagaacta
gataataggtaagaatcaagtatcatgtaaatttgctttcaactcatcccaaaatttgttttatatttca
gtttttttccttcctagcttgactgtggagtcttgtccggaagcaaatagttcctttgcagatcccacat
gtggacaccggacagtaggtcctcaaatgctccttattaggttggttcaataatatcaattgtttgttac
taggcagtgatgttgtacatctggaggagatctcttgagcccataatcaggttattaggaataaatactc
taaggctaaaaatgtagcttagtgataagagtgcttgcctggtgtgctgagaccctcggttccatctcca
caaccccatattccattacaaaataccttttcaccgtccctagcattaagaaacaaaacaacaaagaagt
ttttctttcttctgagatcctgcccggagaggcatttaaaactggccagggccaaaaaaaaaaaaaaaaa
aagaaaaaaaagaaaagaaaacaggctagggccggcatggtggcgcacgcctttaatcccagcacgcagg
aggcagaggcagggcggatctctgtgagtttgaggtcagcctggtctacctagtgagtttcagggcaccc
agggctaaagagactgtctcaaaaacaaaacagccacacaatcagaaccacagcaaaacgcagttatgat
ccttggaactgtaggaatgacaagcatttaaataataggacgagccatttttgagaagctctgatttcac
aagtgtcagggatgggctctgggcgagtaagattgctaatgctggcctctaaatgagaccacgtggagtt
gattagattcttttcatgttcctcgtgctctatcaaataactgtacccaaatacacacacacacacacac
acacaatgcgcgcacacacaaaatccttttttagcttaagaagcccagaatcagaagtaaagctaactgt
gggacttaagtattattctgaacggaactcccagggcgtgaagcgcgcttcaggcttccagagaagcagc
tggcgctggatggaatgaaccaagaggccagcacaggggcagatccgtcgagctctcggccaccgagctg
agcccttaggttctggggctgggaagggtccctaggattgtgcacctctcccgcgggggacaagcagggg
gatggcggggctgacgtcgggaggtggcctccacgggaagggacacccggatctcgacacagccttggca
gtggagtcaggaagggtagggacagattctggacgccctcttggccagctcctcaccgccccacccccga
ctggagccgagagtaattcatacaaaaggagggatcgccttcgcccctgggaatcccagggaccgtcgct
aaattctggccggcctcccagcccggaaccgctgtgcccgcccagcgcggcgggaggagcctgcgcctag
ggcggatcgcgggtcggcgggagagcacaagcccacagtccccggcggtgggggaggggcgcgctgagcg
ggggcccgggagccagcgcggggcaaactgggaaagtggtgtcgtgtgctggctccgccctcttcccgag
ggtgggggagaacggtataaaagtgcggtagtcgcgttggacgttctttttcgcaacgggtttgccgtca
gaacgcaggtgagtggcgggtgtggcctccgcgggcccgggctccctcctttgagcggggtcggaccgcc
gtgcgggtgtcgtcggccgggcttctctgcgagcgttcccgccctggatggcgggctgtgcgggagggcg
agggggggaggcctggcggcggccccggagcctcgcctcgtgtcgggcgtgaggcctagcgtggcttccg
ccccgccgcgtgccaccgcggccgcgctttgctgtctgcccggctgccctcgattgcctgcccgcggccc
gggccaacaaagggagggcgtggagctggctggtagggagccccgtagtccgcatgtcgggcagggagag
cggcagcagtcgggggggggaccgggcccgcccgtcccgcagcacatgtccgacgccgcctggacgggta
gcggcctgtgtcctgataaggcggccgggcggtgggttttagatgccgggttcaggtggccccgggtccc
ggcccggtctggccagtaccccgtagtggcttagctccgaggagggcgaggcccgcccgcccggcaccag
ttgcgtgcgcggaaagatggccgctcccgggccctgtagcaaggagctcaaaatggaggacgcggcagcc
cggcggagcggggcgggtgagtcacccacacaaaggaagagggccttgcccctcgccggccgctgcttcc
tgtgaccccgtggtgtaccggccgcacttcagtcaccccgggcgctctttcggagcaccgctggcctccg
ctgggggaggggatctgtctaatggcgttggagtttgctcacatttggtgggtggagactgtagccaggc
cagcctggccatgaaagtaattcttggaatttgcccattttgagtttggagcgaagctgattgacaaagc
tgcttagccgttcaaaggtattcttcgaactttttttttaaggtgttgtgaaaacctcgaggccgccacc
atggattactacagaaaatatgcagctatctttctggtcacattgtcggtgtttctgcatgttctccatt
ccgctcctgatgtgcaggattgcccagaatgcacgctacaggaaaacccattcttctcccagccgggtgc
cccaatacttcagtgcatgggctgctgcttctctagagcatatcccactccactaaggtccaagaagacg
atgttggtccaaaagaacgtcacctcagagtccacttgctgtgtagctaaatcatataacagggtcacag
taatggggggtttcaaagtggagaaccacacggcgtgccactgcagtacttgttattatcacaaatctta
ataga</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="7">
<INSDSeq>
<INSDSeq_length>17</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..17</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q10">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>gttttcccagtcacgac</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="8">
<INSDSeq>
<INSDSeq_length>17</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..17</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q11">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>caggaaacagctatgac</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="9">
<INSDSeq>
<INSDSeq_length>18</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..18</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q12">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>gccgctgcttcctgtgac</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="10">
<INSDSeq>
<INSDSeq_length>19</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..19</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q14">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>cttggaccttagtggagtg</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="11">
<INSDSeq>
<INSDSeq_length>18</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..18</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q13">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>cacgctacaggaaaaccc</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="12">
<INSDSeq>
<INSDSeq_length>22</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..22</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q15">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>tactagtctattattgtgggag</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="13">
<INSDSeq>
<INSDSeq_length>20</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..20</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q16">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>ttgctagcctattattgtgg</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="14">
<INSDSeq>
<INSDSeq_length>30</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..30</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q17">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>agatctgccaccatggccacctcagcaagt</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="15">
<INSDSeq>
<INSDSeq_length>26</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..26</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q18">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>tctagattagttcttgtattggaagg</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="16">
<INSDSeq>
<INSDSeq_length>19</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..19</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q22">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>acaaacagttctgagaccg</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="17">
<INSDSeq>
<INSDSeq_length>22</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..22</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q23">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>ggtcacattgtcggtgtttctg</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="18">
<INSDSeq>
<INSDSeq_length>22</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..22</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q24">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>cagcccatgcactgaagtattg</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="19">
<INSDSeq>
<INSDSeq_length>19</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..19</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q25">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>atgtgcgcttcgagtccat</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="20">
<INSDSeq>
<INSDSeq_length>20</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..20</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q26">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>ggcagagtgcacattgacag</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="21">
<INSDSeq>
<INSDSeq_length>22</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..22</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q27">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>gagtcctacgctaatgtgaaac</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="22">
<INSDSeq>
<INSDSeq_length>19</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..19</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q28">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>ttccttggctgtggtgttg</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
</ST26SequenceListing>
<---
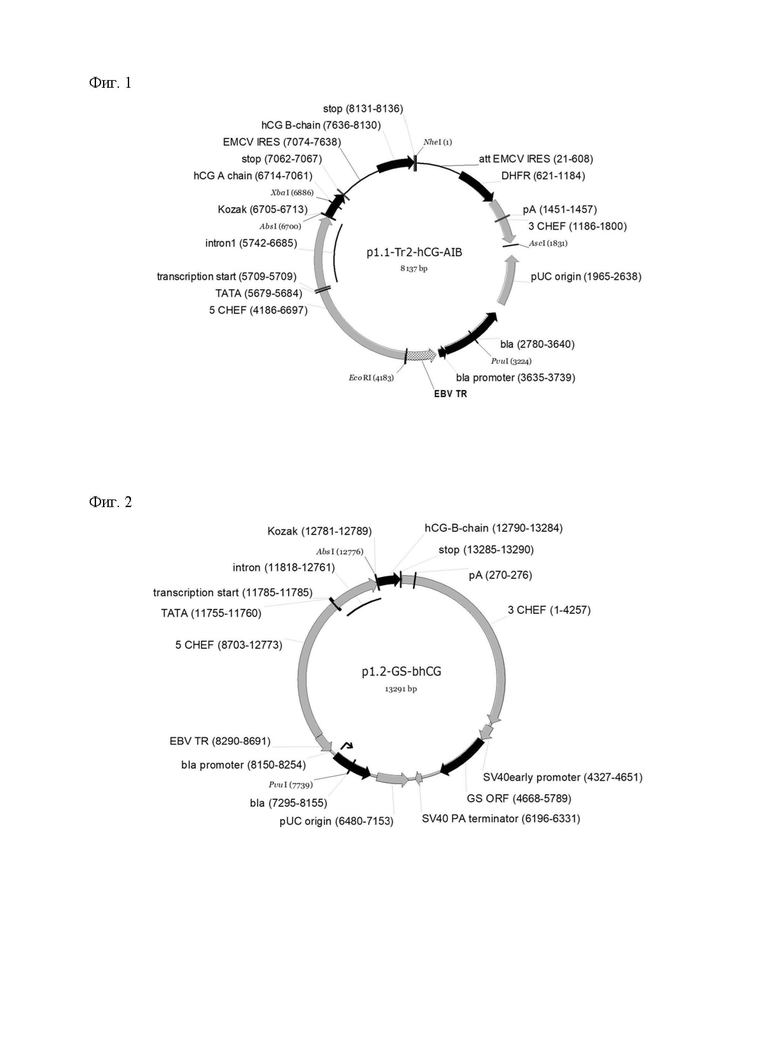
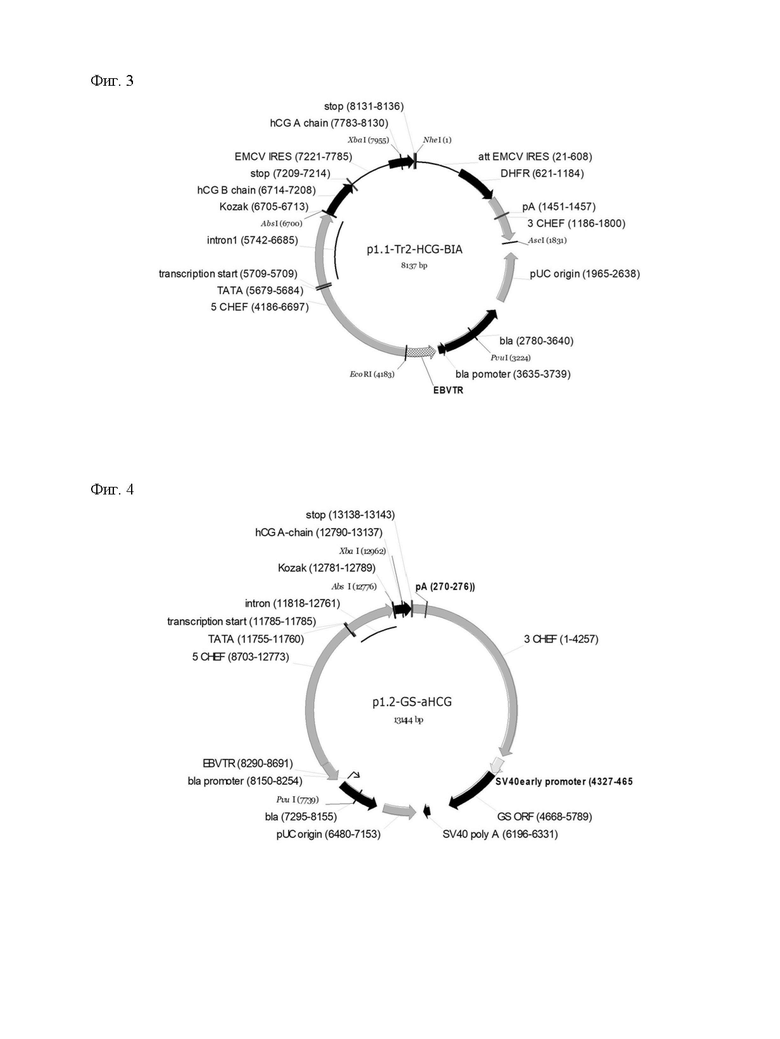
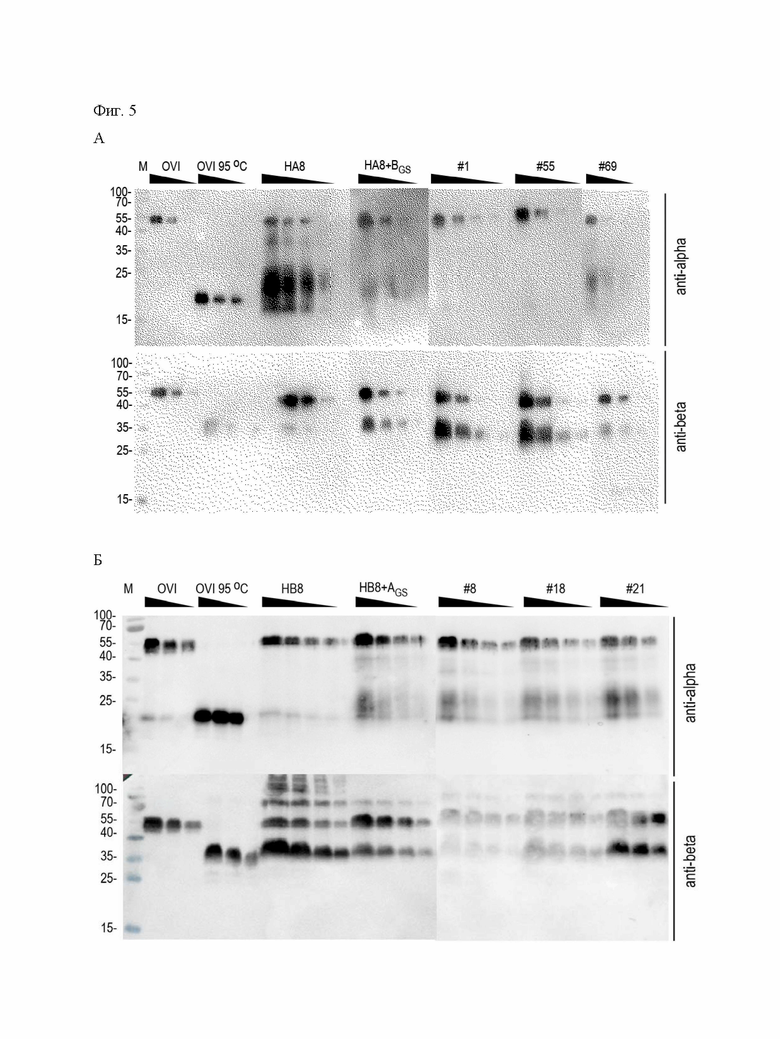
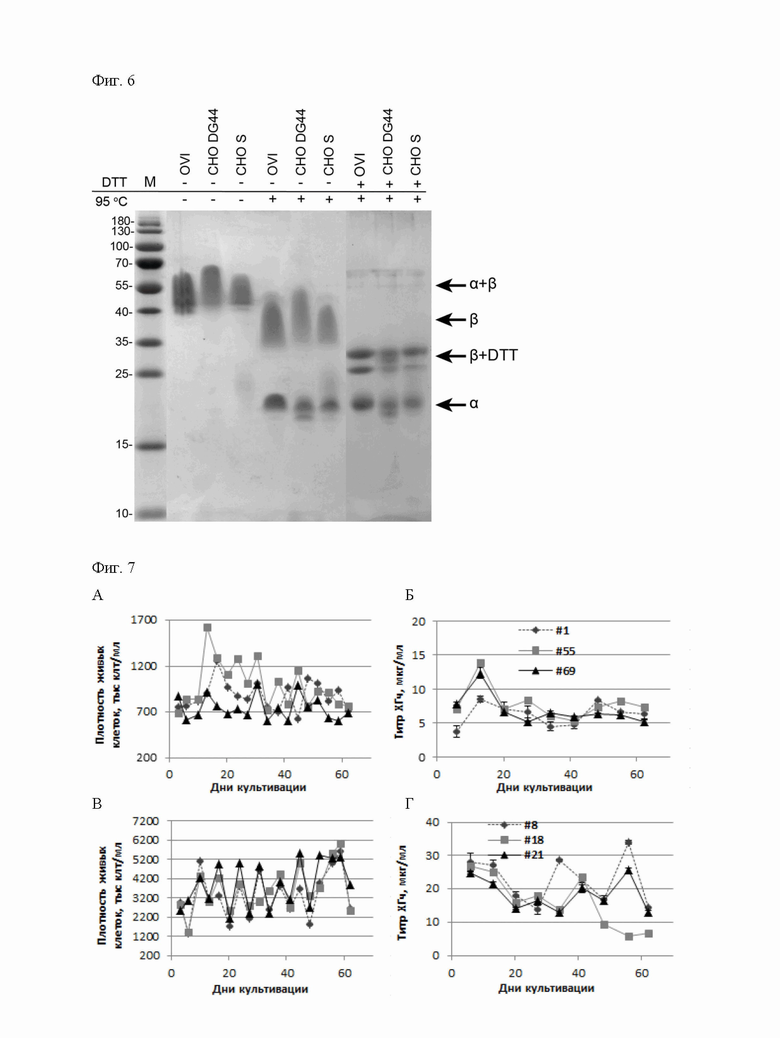
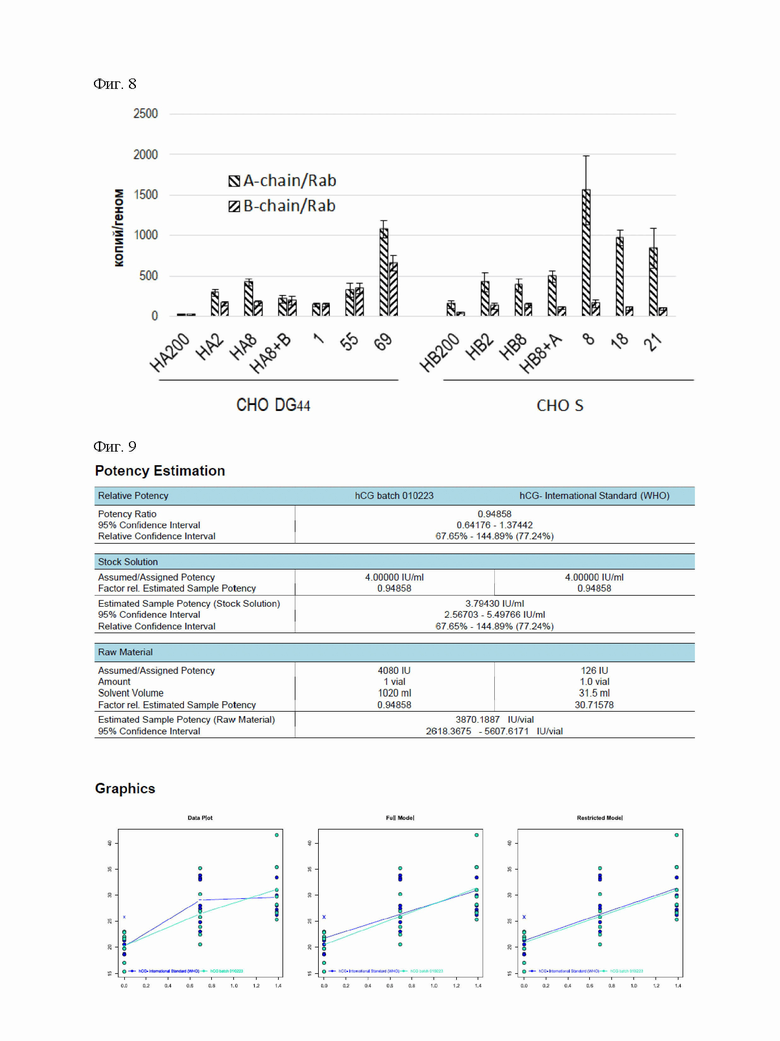
Изобретение относится к области биотехнологии, в частности плазмиде для экспрессии рекомбинантного хорионического гонадотропного гормона человека (ХГЧ). Настоящее изобретение также относится к плазмиде для экспрессии индивидуальных субъединиц альфа или бета рекомбинантного хорионического гонадотропного гормона человека (ХГЧ), моноклональной линии клеток млекопитающего, трансформированной обеими указанными плазмидами, и способу получения рекомбинантного ХГЧ. Настоящее изобретение обеспечивает высокопродуктивную технологию получения рекомбинантного хорионического гонадотропина человека для биофармацевтического производства. 4 н. и 13 з.п. ф-лы, 11 ил., 4 табл., 13 пр.
1. Плазмида для экспрессии рекомбинантного хорионического гонадотропного гормона человека (ХГЧ), в следующей последовательности по существу содержащая:
- область начала репликации плазмиды pUC;
- открытую рамку считывания бета-лактамазы (bla) и прокариотический промотор гена bla;
- участок терминального повтора вируса Эпштейн-Барр человека;
- функциональный промотор и первый интрон гена фактора элонгации 1 альфа китайского хомячка, фланкированный 5’ нетранслируемой областью этого гена;
- последовательность Козак;
- открытую рамку считывания, кодирующую альфа-субъединицу ХГЧ с блоком стоп-кодонов; внутренний сайт связывания рибосом (IRES); открытую рамку считывания, кодирующую бета-субъединицу ХГЧ с блоком стоп-кодонов, или
- открытую рамку считывания, кодирующую бета-субъединицу ХГЧ с блоком стоп-кодонов; внутренний сайт связывания рибосом (IRES); открытую рамку считывания, кодирующую альфа-субъединицу ХГЧ с блоком стоп-кодонов;
- аттенюированный внутренний сайт связывания рибосом;
- открытую рамку считывания дегидрофолатредуктазы (DHFR), экспрессирующуюся в составе трицистронной мРНК вместе с геном, кодирующим альфа- и бета-субъединицы ХГЧ; и
- функциональный терминатор гена фактора элонгации 1 альфа китайского хомячка, фланкированный 3’ нетранслируемой областью этого гена.
2. Плазмида по п. 1, отличающаяся тем, что открытые рамки считывания, кодирующие альфа- и бета-субъединицы ХГЧ, кодируют альфа- и бета-субъединицы ХГЧ, последовательности аминокислот которых приведены в SEQ ID NO: 2 и 3, соответственно.
3. Плазмида по п. 1, отличающаяся тем, что участок терминального повтора вируса Эпштейн-Барр человека (EBVTR) представляет собой фрагмент конкатемера терминального повтора EBVTR.
4. Плазмида по п. 1, отличающаяся тем, что открытая рамка считывания дегидрофолатредуктазы (DHFR) представляет собой открытую рамку считывания дегидрофолатредуктазы (DHFR) мыши.
5. Плазмида по п. 1, отличающаяся тем, что внутренний сайт связывания рибосом (IRES) представляет собой внутренний сайт связывания рибосом (IRES) вируса энцефаломиокардита (EMCV) лошадей.
6. Плазмида по п. 1, отличающаяся тем, что указанная плазмида выбрана из группы, включающей плазмиду p1.1-Tr2-hCG-AIB, представленную в Перечне последовательностей под номером SEQ ID NO: 1, и плазмиду p1.1-Tr2-hCG-BIA, представленную в Перечне последовательностей под номером SEQ ID NO: 5.
7. Плазмида для экспрессии индивидуальных субъединиц альфа- или бета-рекомбинантного хорионического гонадотропного гормона человека (ХГЧ), в следующей последовательности по существу содержащая область начала репликации плазмиды pUC, открытую рамку считывания бета-лактамазы (bla) и прокариотический промотор гена bla; участок терминального повтора вируса Эпштейн-Барр человека; функциональный промотор гена фактора элонгации 1 альфа китайского хомячка, фланкированный 5’ нетранслируемой областью этого гена; последовательность Козак; открытую рамку считывания гена, кодирующего альфа- или бета-субъединицу ХГЧ с блоком стоп-кодонов; функциональный терминатор гена фактора элонгации 1 альфа китайского хомячка, фланкированный 3’ нетранслируемой областью этого гена; промотор вируса SV40; ОРС глутаминсинтазы крысы; и функциональный терминатор вируса SV40.
8. Плазмида по п. 7, отличающаяся тем, что ген альфа-субъединицы ХГЧ кодирует полипептид, последовательность аминокислот которого приведена в SEQ ID NO: 2, а ген бета-субъединицы ХГЧ кодирует полипептид, последовательность аминокислот которого приведена в SEQ ID NO: 3.
9. Плазмида по п. 7, отличающаяся тем, что участок терминального повтора вируса Эпштейн-Барр человека представляет собой фрагмент конкатемера терминального повтора EBVTR.
10. Плазмида по п. 7, отличающаяся тем, что указанной плазмидой является p1.2-GS-bhCG, представленная в Перечне последовательностей под номером SEQ ID NO: 4, или p1.2-GS-аhCG, представленная в Перечне последовательностей под номером SEQ ID NO: 6.
11. Моноклональная линия клеток млекопитающего - продуцент рекомбинантного ХГЧ, трансформированная одновременно плазмидой по пп. 1-6 и плазмидой по пп. 7-10, при этом указанная плазмида по пп. 7-10 содержит открытую рамку считывания, кодирующую субъединицу ХГЧ, расположенную в плазмиде по пп. 1-6 второй.
12. Моноклональная линия клеток по п. 11, отличающаяся тем, что клетками млекопитающего являются клетки яичника китайского хомячка.
13. Моноклональная линия клеток по п. 11, отличающаяся тем, что указанной линией клеток является линия клеток яичника китайского хомячка D-pTr2-hCG-55, депонированная во Всероссийской Коллекции Промышленных Микроорганизмов (ВКПМ) под регистрационным номером ВКПМ Н-194, или линия клеток яичника китайского хомячка S-pTr2-hCG-8, депонированная в ВКПМ под регистрационным номером ВКПМ Н-213.
14. Способ получения рекомбинантного ХГЧ, включающий следующие стадии:
- культивирование в питательной среде линии клеток по пп. 11-13; и
- выделение полученного ХГЧ из культуральной жидкости.
15. Способ по п. 14, отличающийся тем, что выделение полученного ХГЧ из культуральной жидкости осуществляют центрифугированием культуральной жидкости для отделения клеток и дебриса с последующим выделением целевого ХГЧ с помощью хроматографии.
16. Способ по п. 14, отличающийся тем, что культивируют клетки яичника китайского хомячка D-pTr2-hCG-55 или S-pTr2-hCG-8, депонированные во Всероссийской коллекции промышленных микроорганизмов (ВКПМ) под регистрационным номером ВКПМ Н-194 или ВКПМ Н-213, соответственно.
17. Способ по п. 14, отличающийся тем, что питательная среда для культивирования является бессывороточной.

