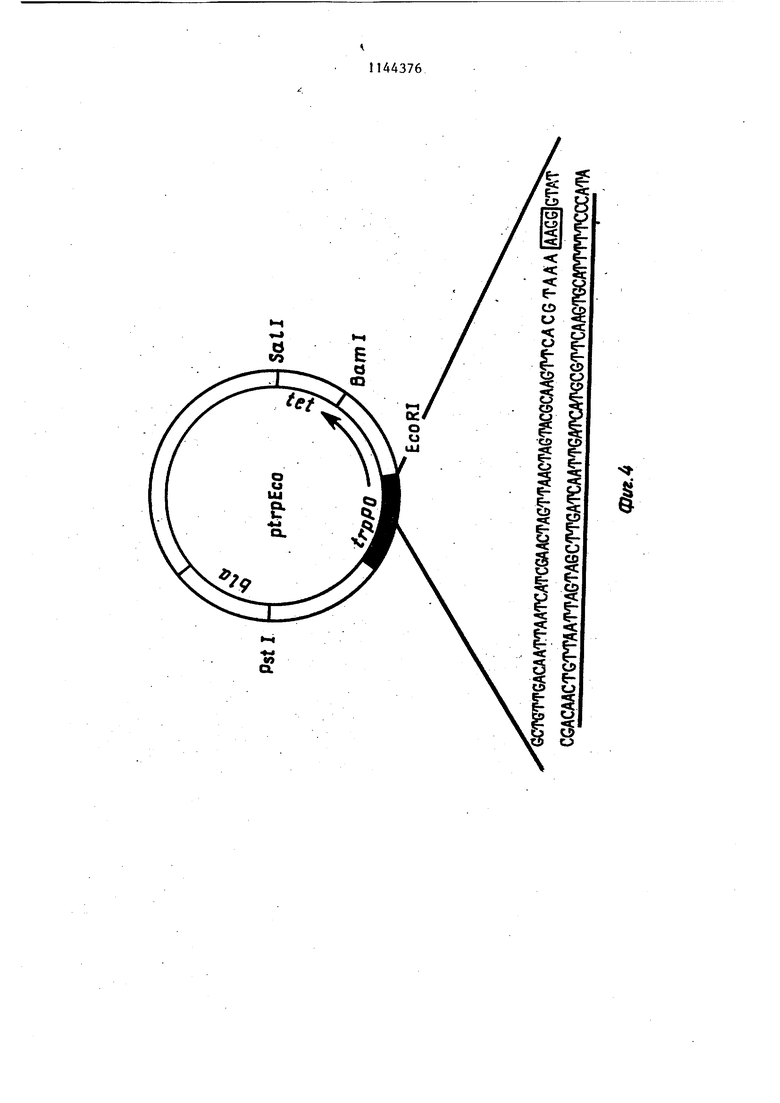
регуляторную область триптофано (вого оперона,содержащую промотор опе ратор и участок, кодирующий начало лидерной РНК без инициирующего кодо- на, локализованную между 4838 и 5636 нуклеотидами и присоединенную к EcoRI сайту pBR 322,
участок ДНК, соединяющий регуляторную область триптофанового оперона с геном ci-F и имеющий последовательность:. AAAAAGGGTATCGC GGAATTC ATG,
участки расщепления эндонуклеаза,ми рестрикции со следующими коорди:натами: Pvii II - 276 нуклеотид и 2543 нуклеот1ед Ват I- 851 нуклеотид. Hind III - 504 нуклеотид, Sa2l 1125 нуклеотид и PstI - 4088 нуклеотид.
TG-T G-AT СТО- ест CAG- АСС САС AGC CTG- G6T ААТ AGG AG& GCC TTGАТА СТС CTG GCA САА ATG G&A А&А АТС ТСТ ССТ ТТС TGC TCCCTG АА& GAC AGA CAT GAC ТТТ G(JA ТТС ССС CAG- GAG GAG ТТТ GAT GGC AA& САС ТТС САС ААС ССТ САА ССС АТС ТСТ СТС СТС CAT САС АТС АТС СА& CAG АСС ТТС ААТ СТС ТТС AGC АСА АА& GAG ТСА ТСТ GCT ACT TGG GAA CAG AGC СТС СТА SAA ААА ТТТ ТСС ACT GAA, СТТ ААС CAG. CAG- CTG- ААТ 6АС СТ& GAA GCC TGC СТ& АТА CAS GAG GTT GGGGTG- GAA GAG- ACT ССС CTG- ATG- AAT GTG- GAC TCC АТС CTG GCT GTG AAO AAA TAC TTC CAA АСА АТС ACT ОТ TAT CTG- АСА GAG-AAG- AAA TAC AGC CCT TGT GCC TGG GAG GTT GTC AGA GCA GAA ATG- ATG AGA TCC TTC TCT TTA TCA AAA ATT TTT CAA GAA A&A TTA AGG AGG AAG
GAATAAACTC,
регуляторную область триптофанового оперона, содержащзто промотор, оператор и участок, кодирующий начало лидерной РНК без инициирующего кодо на, локализованную между 4838 и 5640 нуклеотидами и присоединенную к EcoRI сайту pBR 322,
участок ДНК, соединяющий регуля- торную область триптофанового оперог на с геном pl-F и имеющий последова-; тельность: .
AAAAAGGGTATCGC GGAATTAATTC ATG, участки расщепления эндонукле- азами рестрикции со следующими когенетические маркеры: tet-ген, локализованный между 506 и 1000 нуклеотидайи, ген j -лактамазы, локализованный между 3500 и 4600 нуклеотидами.
2. Рекомбинантная плазмида pIFN-.Ы-F trp2, кодирующая синтез лейкоцитарного интерферона типа G(-F человека, характеризуется следующими признаками:
имеет длину 5664 п.о.;
содержит EcoRl.- Hind III большой фрагмент плазмиды pBR 322,
ген лейкоцитарного интерферона типаоС.-Р человека, локализованный с 1 по 505 нукл еотид и имеющий последовательность:
ординатами: PVU II - 276 нуклеотид и 2543 нуклеотид Ват 1-851 нукле- отид. Hind III - 504 нуклеотид, ,Sal 1 - 1125 нуклеотид, PSt I 4088 нуклеотид,
генетические маркеры: tet-ген, локализованный между 506 и 1000 нуклеотидами, ген р-лактамазы, локализованный между 3500 и 4600 нуклеотидами.
i
3. Рекомбинантная плазмида pIFN-d.-F trp3, кодирующая синтез лейкоцитарного интерферона типа
-человека, характеризуется еле- дзгющими признаками:
имеет длину 5356 п.о.; содержит EcoRl-Hind .IIIбольшой , фрагмент векторнойплазмиды pBR 322
Т&Т GAT CTG. ест GAG- AGO GAG AGO GTG- GGT AAT A6G AGO GGG TTGATA GTG GTG GCA GAA AT6 GGA AGA ATG TGT GGT TTG TGG TGC CTGAAG- GAG AGA GAT GAG TTT GGA TTG GGG GAG GAG- GAG TTT GAT GGG AAG GAG TTG GAG AAG GGT GAA GGG ATG TGT iTG GTG GAT GAG ATGATG GAG GAG-AGG TTG AAT GTG TTG AGG AGA AAG GAG TGA TGT GGT AGT TGG GAA GAG-AGG GTG GTA GAA AAA TTT TGG AGT GAA GTT AAG GAG- GAG GTG AAT GAG GTG GAA GGG TGG GTG ATA GAG GAG GTT GGG G-TG GAA GAG AGT GGO GTG- ATG- AAT GTG GAG TGG ATG GTG GGT GTG AAG AAA TAG TTG GAA AGA ATG AGT GTT TAT GTG AGA GAG AAG- AAA TAG AGK) GGT TGT GGG TGG GAG GTT GTG AGA GGA GAA ATG ATG- AGA TGG TTG TGT TTA TGA AAA ATT TTT GAA GAA AGA TTA AGG AGG AA& GAA TAA ACTG, ..
регуляторную область триптофа- нового оперона, содержащую промотор, оператор и участок, кодирующий начало лидерной РНК без инициирующего : кодона, локализованную между 4838 и 5636 нуклеотидами и присоединенную к EcoRl сайту рВЕ 322,
участок ДНК, соединяющий регуляторную область триптофанового one- рона с геномЫ-Р и имеющий последо вательность:
- . AAAAAGCiCrTATCGCCAC ATG, участки расщепления эндонукле- азами рестрикции со следующими координатами: PVU II - 276 нуклеотид и 2543 нуклеотид. Вага 1-851 нуклеотид. Hind III - 504 нуклеотид, Sail - 1125 нуклеотид, PSti - 4088 нуклеотид.
TGTSAT GTG GGT GAG AGG GAG AGG GT& GGT AAT AGG AGG GGG TTGATA GTG GTG GGA GAA ATG GGA AGA ATG TGT GGT TTG TGG TGG GTCAAG- GAG AGA GAT GAG TTT GGA TTG GGG GAfr GAG GAG TTT CAT GGC
ген лейкоцитарного интерферона типа o(-F человека, локализованный с 1 по 505 нуклеотид и имеющий последовательность:
; генетические маркеры: tet-ген, локализованный мекду 506 и 1000 нук- леотидами, ген р -лактамазы, локализованный между 3500 и 4600 нуклеоти дами.
4. Рекомбинантная плазнида pIFN-5(-F trp4, кодирзтощая синтез лейкоцитарного интерферона типа человека, характеризуется следу- Ю1ЩМИ признаками:
имеет длину 5655 п.о.;
содержит EcoBI - Hind III большой фрагмент векторной плазмиды рВН 322,
ген лейкоцитарного интерферона типао(-F человека, -локализованный с 1 по 505 нуклеотид и имеющий последовательность:1144376 .AAC CA& ТТС CAG- AAG- GCT CAA GCC АТС TCT GTC СТО CAT GAG- ATGАТС CAG- CA& ACC TTC AAT CTC TTC AGC АСА AAG- GAC TCA TCT GCT ACT TGG G.AA CAG- AGC CTC СТА G-AA AAA TTT TCC ACT GAA CTT AAC CAG- CAG. CTG-AAT GAG CTG- GAA GCC TGC GTG- ATA CAG-GAG- GTT GGGG-TCrGAA GAG-ACT CCC CTG-ATG-AAT CTG-frAC TCC АТС СТО/ GCT GTG- AAG. AAA TAC TTC. CAA АСА АТС ACT CTT TAT CTG- АСА OAfr AAG- AAA TAC AGC CCT TGT GCC T&G GAG GTT GTC AGA GCA GAA АТС AT& AGA TCC TTC. TCT TTA TCA AAA ATT TTT CAA GAA AGA .TTA AGG ASG- AAG-GAA TAA ACTC, регуляторную область триптофа.-генетические маркеры: tet-ген, нового оперона, содержащую промотор,локализованный между 506 и 1000оператор и участок, кодирующий иа-нуклеотидами, ген р -лактамазы, лочало лидерной РНК без инициирующегокализованный между 3500 и 4600 нуккодона, локализованную между 4838леотидамя. и 5636 нуклеотидами и присоединенную к EcoHI сайту pBR 322, 5. Рекомбинантная плазмида участок ДНК, соединяющий регуля-pIFN-o(-F trp5, кодирующая синтез лейторную область триптофанового оперо-коцитарного интерферона типао(-Г на с геномов-F и имеющий последова-человека, характеризуется следуютельность:Щими признаками: AAAAAGGGTATCGCCA ATG,имеет длину 5652 п.о.; участки расщепления эндонуклеаза-содержит ЕооМ -. Hind III больми рестрикции со следующими коорди-той фрагмент векторной плазмиды натами: Pvn II . 276 нуклеотид иpBR-322, 2543нуклеотид, Baml - 851 нуклео.-. ген лейкоцитарного интерферона тид. Hind III .- 504 нуклеотид,типао -F человека, локализованный Sati - 1125 нуклеотид, PSti - 4088с 1 по 505 нуклеотид и имеющий понуклеотип - -сл довательность: Г6Т GAT СТ& СОТ GAG- AGO GAG AGC GT& «Я AAT A( AGC GGC TTC , ATA CTC GT& GCA GAA ATG- G6A A&A ATG TGT COT TTG TCC TGC GT&. AAG- GAG AGA GAT GAG TTT GGA.TTG GCC GAG- (A(r GA& TTT GGC AAG GAS- TTG GAG- AAG- GCT GAA GCG ATG TGT G-TG GTC GAT &AG-ATGATG GAG- GAG- AGG TTG AAT GTG TTG AGC AGA AAG- GAC TCA TCT QCT AGT TGG-GAA GAG-АЭС GTG GTA fiAA AAA TTT TCC AGT C4AA ,GTT AAG GAG. GAG- GTG- AAT &AG GTG- GAA GGG TGG GT& ATA GAGr OAG- G-TT GGGG-TG- &AA GAG- AGT GGG GTG- ATG- AAT GTG GAG TCC АТС CTG GCT fe-TGAAG. AAA TAG TTG GAA AGA ATG AGT GTT TAT CT& АСА GAG-AA(r-AAA TAG AGG GGT T6T GGC TGO GAG-GTT G-TG A&A GCA. GAA ATG-ATG-AGA TGG TTG TGT TTA TCA AAA ATT lit CAA (MA AWA TTA AG{r A(:Kr-AAfr GAA TAA AGTC.
регуляторную область триптофане- вого оперона, содержащую промотор, оператор и участок, кодирующий начало лидерной РНК без инициирующего кодона, локализованную между 4838 и 5636 нуклеотидами и присоединенную к ЕсоНХ.сайту pBR 322,.участок ДНК, соединяющий регуляторную область триптофанового оперо- на с геном и имеющий последовательность :
AAAAAGGCiAATTC ATG,
участки расщепления эндонуклеазами рестрикции со следуюпщми координатами: PVU II - 276 нуклеотид, 2543 нуклеотид, Bani - 851 нуклеотид,. Hind III - 504 нуклеотид. Sail 1425 нуклеотид, PStl - 4088 нуклео- тид, .
генетические маркеры: tet-ген, локализованный между 506 и 1000 нук- леотидами, гено1-лактамазы, локал№ зованньгй между 3500 и 4600 нукле- отид.
6. Итамм Escherichia cell/ pIFN-o(-F trpl ЦМПМ B-2885 (коллекция Всесоюзного научно-исследователь- : ского института генетики и селекции
ВНИИ промьшшенных микроорганизмов
Генетика) - продуцент лейкоцитарного интерферона типаоС-F человека.
7.Штамм Escherichia coli/pIFN-d-F trp2 ЦМПМ В-2886 (коллекция Всесоюзного научно-исследовательского института генетики и селекции про мьшшенньк микроорганизмов ВНИИ Генетика) - продуцент лейкоцитарного интерферона типа . человека..
8.Штамм Escherichia coli/pIFN-c -F trp3 ЩШМ В-2887 (коллекция Всесоюзного научно-исследовательского -даститута генетики и селекции промышленных микроорганизмов ВНИИ Генетика) - продуцент лейкоцитарного интерферона типао1-F человека.
9.Штамм Escherichia coli/pIFN-o(-F trp4 ЦМПМ В-2888 (коллекция Всесоюзного научно-исследовательского института генетики и селекции промьшленных микроорганизмов ВНИИ Генетика) - продуцент лейкоцитарного интерферона, тштас{-F человека.
10.Штамм Escherichia coli/pIFNTot-F trp5 ЦМ1Ш.В-2889 (коллекция Всесоюзного научно-исследовательского института генетики и селекции промышленных микроорганизмов ВНИИ Генетика) - продуцент лейкоцитарного интерферона ( -F человека.











Изобретение относится к микробиологической промышленности, молекулярной биологии и генетической инженерии и представляет собой серию сконструированных in vitro рекомби нантных плазмид,обуславливающих син тез одного из интерферонов человека - лейкоцитарного интерферона типа isL-F в клетках кишечной .палочки E.coli, и штаммы Е.coli - продуценты данного интерферона.
Известна рёкомбинантная плаз,мида, кодирующая синтез лейкоцитарного интерферона типао -А человека и штаммы E.coli, содержащие зту плазмиду - продуценты интерферона типаоС-А человека.
Г
Указанная рёкомбинантная плазмигда состоит из гена интерферонао -А и триптофанового промотора, которые встроены в векторную плазмиду pBR - 322.
Однако уровень синтеза интерфе- рона в штамме E.coli, содержащем эту рекомбинантную плазмиду, недостаточно высок - 2,5-10 единиц активности с 1 л бактериальной суспензии. .
Наиболее близким к предлагаемому изобретению по технической сущности и достигаемому результату является рёкомбинантная плазмида pTMKS - .IFN - F - 71, кодирующая синтез интерферона типас(-F человека, и штамм E.coli, содержаш;ий эту плазмиду - продуцент интерферона THnao.-F человека..
Рёкомбинантная плазмида pTMKS- -IFH-F -- состоит из следующих элементов : -EcoRl-Hind III - большого фрагмента векторной плазмиды pBR-32
3
регуляторного участка триптофаново- го оперона Е.coli,содержащего промотор, оператор и участок, кодирующий начало лидерной РНК без инициирующего кодона, и присоединенного к EcoRl.сайту pBR-322; - гена лейкоцитарного интерферона типао -F человека, реконструированного для прямой экспрессии в клетках E.coli, - участка ДНК, соединяющего регуляторный з асток триптофанового оперона с геном cf.-F и имеющего следующую последовательность:
AAAMGGGTATCGCCAGGAATTC ATG .
Уровень синтеза интерферона в штамме E.coli, содержащем рекомбинантную плазмиду pTMKS-JFN-F-71, составляет 10 - 10® ед. активности интерферона с 1 л бактериальной суспензии при ее плотности Agg 1.
Недостатком рекомбинантной плаз- МИДЫ pTMKS-lFN-F-71 является недостаточно высокий выход интерферона d-F.
Т6-Т &АТ СТ6- ест CAG- АСС CAG АСС СТС ССТ ААТ AG& AG& GCC TTG АТА СТС 6TGССТ ТТС ТСС Т6С CTG- АА& GAC CAG- GAG- GAG- ТТТ GAT GGC AAC АТС ТСТ G-TC СТС CAT CAG-ATGTTC. AGC АСА AA6- GAC TCA TCT СТА GAA AAA TTT TCC ACT GAA CTG- GAA GCC TGC GTG ATA CAG CCC CTG ATG- AAT GTG GAC TCC TTC CAA АСА АТС ACT GTT TAT CCT TGT GCC TGiJ GAG- G-TT &TC TTC TCT TTA TCA AAA ATT TTT GAA TAA ACTC, регуляторную область гриптофано -, вого оперона, содержащую п1 омотор, оператор и участок, кодирующий начало лидерной РНК без инициирую-щего кодона, локализованную между 4838 и 5636 нуклеотидамя и присоединенную к EcoRl сайту pBR 322,
ЛАЗ 76
Целью описываемой группы изобретений являются новые рекомбинантные плазмиды, кодирующие синтез лейкоцитарного интерферона типа d -F чело- 5 века, и штаммы E.coli, содержащие эти плазмиды - продуценты лейкоцитарного интерферона типа 01-F человека, обеспечивающие более высокий интерферона. 10 JПоставленная цель достигается новой рекомбинантной плазмидой p;lFN-0/-Ftrp 1, кодирующей синтез лейкоцитарного интерферона типа 15 человека, которая характеризуется следующими признаками: имеет длину 5660 п.о.;
содержит: EcoRl - Hind III большой фрагмент векторной плазмиды 20 рВН 322,
ген лейкоцитарного интерферона THnaot-F человека, локализованный с 1 по 505 нуклеотид и имеющий последовательность : GCA CAA ATG- GGA AGA АТС TCT AGA CAT &AC TTT GGrA TTC CCC CAG- AAG- GCT CAGrTTC CAA GCC АТС CAGCAG ACC TTC AAI CTC GCT ACT TGG- GAA CAGr AGC CTC CTT AAC CAG CAG CTG AAT GAC GAG GTT GG& GT& GAA GAG ACT АТС CTG GCT GTG AAG AAA TAC CT& АСА GAG AAC AAA TAC AGC AGA GCA GAA АТС ATGAGA TCC CAA GAA AGA TTA AGG AOaMGучасток ДНК, соединяющий регуляторную область триптофанового one- рона с геноме/-F и имеющий последова55 тельность: AAAAAGGGTATCGCGGAATTC ATG, участки расщепления эндонуклеазами рестрикции со следующими коор- динатами: Pvu II - 276 нуклеотид и 2543 нуклеотид. Ват I - 851 нукл отид, Hind III - 504 нуклеотид, Sail 1125 нуклеотид и Psti .- 408 нуклеотид. Генетические маркеры: tet-reH Локализован между 506 и 1000 нукле отидами, ген р-лактамазы локализован между 3500 и 4600 нуклеотидами, Поставленная цель достигается т же новой рекомбинантной гшазмидой pIFN-o(-Ftrp2, кодирующей синтез ле коцитарного интерферона типао.- ч ловека, которая характеризуется те ми же признаками, что и рекомбинан ная плазмида pIFN- -Ftrpl, но имее длину 5660 п.о. и иную последовательность участка ДНК, соединяющую регуляторную область с геном o(-F AAMAGGGTATCGCGGAATTMTTC ATG. Поставленная цель достигается т же новой реко1 бинантиой плазмидой plFN-oi-Ftrp 3, кодирующей синтез лейкоцитарного интерферона типа oL- человека, которая характеризуется теми же признаками, что и рекомбин тная плазмида pIFN-0(.-Ftrp 1, но им ет длину 5656 п.о. и следующую последовательность участка ДНК, соединяющего регуляторную область с reHOHO -F: AAAAAGGGTATCGCCAC ATG. Поставленная цель достигается т же новой рекомбинантной плазмидой pIFN-o(-F-trp4, кодирующей синтез лейкоцитарного интерферона типaotчеловека, которая характеризуется теми же признаками, что и рекомби- нантная плазмида pIFN-oi-Ftrpl, но имеет длину 5655 п.о. и следующую последовательность участка ДНК, со единяющего регуляторну область с геном oC-F: AAAAAGGGTATCGCCA ATG , Поставленная цель достигается также новой рекомбинантной плазми- дои pIFN-o(-F-trp, кодирующей л ейкоцйтарного интерферона типа, человека, которая характе ризуется теми же признаками, что и рекомбинантная плазмида pIFN-(-F-trp но имеет длину 5652 п.о. и следу Вщую последовательность участка ДНК, соединяющего регуляторную область с геном AAAAAGGGAATTC ATG . Новые рекомбинантные плазмиДы plFN-cL-Ftrp отличаются от извест- 1,ной плазмиды pTMKS-IFN-F-71 .последовательностью участка ДНК, соединяющего регуляторную область трип- тофанового оперона с геномot-F. Поставленная цель достигается так же штаммами Escherichia coli/pIFN- p{.-FtrT)l ЦМПМ В 2885, Fscherichia coli/pIFN-o(.-Ftrp2 ЦМПМ В 2886, Escherichia coli/plFN-rt-F trp 3 ЦМПМ В 2887, Escherichia coli/pIFN-oC-F tfip 4 ЦМПМ В 2888, Escherichia coli/pIFN-oC-F trp 5 ЦМПМ В 2889.. (коллекция Всесоюзного научно-ис- следовательского института генетики к селекции промьшшенных микроорганизмов ВНИИ Генетика), которые являются продуцентами лейкоцитарного интерферона типа ot-F человека. Уровень синтеза интерферона при этом достигает 5--10 - 2-10 ед. активности с 1 л бактериальной суспензии, что в 5-200 раз вьше, чем при использовании известного штам- ма, содержащего плазмиду pTMKS-IFN-F-71. Способ конструирования рекомби- нантных плазмид для экспрессии состоит в следующем. В качестве источника гена интерферона oi,-F служит плазмида pIFN-c(-F pre, представляющая собой плазмиду pBR-322 в Hind III сайт которой вставлен ген интерферона o(.-F (IFN-F), после- довательность которого приведена на фиг. 1. Этот ген вырезают из плазмиды с помощью рестрикционной эндонуклеазы Hind III и реконструируют для прямой экспрессии в соответствии со схемой, приведенной на фиг. 2. Для этого ген неполностью расщепляют рестрикционной эн- донуклеазой SauSAl и путем электрофореза в полиакриламидном геле вьщеляют его С-концевую часть А, Эта часть лишена триплета TGT, кодирующего первую аминокислоту зрелого интерферона - цистеин. Для того, чтобы сконструировать полный ген, синтезируют химическим путем фрагент ДНК (как изображено на фиг. 2), структуре которого содержится ко- он TGT, предшествующий ему иници- аторный кодон ATG, а также два вытупающих липких конца, соответстующ11х концам, образз щимся при рас- щеплении ДНК рестрикционными эндо- нуклеазами EcoRl (слева) и SauSAl (справа). Присоединение этого фрагмента к части А гена интерферо- на, осуществляемое с помощью ДНК- лигаэы фага Т4, дает полный ген с от сеченным фрагментом, кодирующим сиг- напьный пептид. Ген снабжен иници- аторным триплетом ATG и липкими концами, соответствующими рестрикци онным эндонзтлеазам EcoRl (слева) и Hind III (справа). Для стабильного хранения и воспроизводства гена в клетках E.coli конструируют плазМИДУ plfFNpOt-F-0 (фиг, 3), в которой ген не экспрессируется. С этой целью плазмиду pBR-322 расщепляют реет рикционными эндонуклеазами EcoRl и Hind III и лишенный промотора вьг деленный большой фрагмент соединяют с помощью ДНК-лигазы с полным геном Штаммы Е. coli, содержащие эту ШIaзМИДУ, стабильно воспроизводят ее. Они устойчивы к антибиотику ампициллину и чувствительны к антибиотику тетрациклину. Ген Л FN-о(-F. может быть вырезан из плазмиды ptFN-T)( с помощью расщепления рестрикционными эндонуклеазами EcoRl и Hind III. В качестве первичного вектора для встраивания гена iFN-ot-F под контроль триптофанового промотора исполь эуют плазмиду ptrp Есо, генетическая и рестрикционная карта которой, а также первичная структура участка, окружакщего место расщепления реет- рикционной эндонуклеазой EcoRl и включающего часть регуляторной облас ти триптофанового оперона, приведена на фиг, 4. Способ конструирования рекомбинан ной плазмиды p fFN-rf-Ftrp1 заключается в том, что плазмиду plFNTtC, содержащую реконструированный ген ин . терферона rut-F, и плазмиду ptrp Есо расщепляют рестрикционными зндо- нуклеазами EcoRl и ,-образующие фрагменты лшгируют с помощью ДНК-лигазы, полз енной смесью рекомбинант- ных молекул трансформируют клетки штамма Есо11К802 или Ecoli HBjOl, или Ecoli 294, и из клонов, устойчивых к тетрациклину, отбирают целевую плазмиду .(, см. фиг. 5 Дпя получения рекомбинантной плазмиды pJ.FN-o -Ftrp2 плазмиду nJFN- -F trpi расщепляют рестрик- ционной. эндонуклеазой EcoRl, липкие концы заполняют ДНК-полимеразой и лигируют с помощью ДНК-лигазы фага Т4.. Для получения рекомбинантных плазмид.рЛРН- - Ftrp3 -и plFN-TSL-F trp4 плазмиду pJFN-ic) i;rpl. расщепляют эндонуклеазой рестрикции EcoRl, обрабатывают эндонуклеазой S, а затем лигируют с помощью ДНК- пигазы фага Т4. Рекомбинантную плазмиду pJFN й/-F trp5 конструируют из большого фрагмента, полученного расщеплением плазмиды ptrp Есо рестрикционными эндонуклеазами EcoRl и Satl с последующим удалением EcoRl-липкого конца обработкой S нуклеазой, и малого фрагмента, полученного при расщеплении плазмиды plFN- -F trpl, рестрикционными эндонуклеазами EcoRf и SaFl с последующим заполнением EcoRl-липкого конца ДНК-полимеразой фага Т4 в присутствии четырех дезоксинуклеозидтрифосфатов, путем лигирования этих фрагментов с помощью ДНК-лигазы фага Т4. Анализ полученных рекомбинантныя ДНК проводят с помощью рестрикцион- нык эндонуклеаз EcoRl, PvuII, Hind III, BamHi, Sail. Для получения штаммов-продуцен- ТО9 интерферона человека бинантные плазмиды plFN- -F trp трансформируют в штамм E.coli К802 (gal К, lac Y, met, hsr, hsm). Для получения штамма-продуцента E.coli/plFN-ot F trpl ЦМПМ В 2885 рекомбинантную плазмиду pilFN-ot-Ftrpl трансформируют в штамм E.coli К802, Для получения штамма продуцента E,coli/pa:FN- X-F trp 2 ЦМШВ 2886 рекомбинантную плазмиду p;tM-o(-F trp2трансформируют в штамм E.coli К802, Для получения штамма-продуцента E.coli/pjFN-A-F trp 3 ЦМПМ В 2887 рекомбинантную плазмиду piFN-cCr-F trp3 трансформируют в штамм E.coli , . - .. Для получения штамма-продуцента E.coli/pjfFN- -F trp4 ЦМПМ В 2888 рекомбинантную плазмиду pJfFN- rF trp 4 трансформируют в штамм Е.coli К802. Для получения штамма-продуцента E.coli/plFN-ot-F trp5 ЦМПМ В 2889 рекомбинантную плазмиду piFN-OC-F
trp5 трансформирзпот в штамм E.coli К802. ..
В качестве штаммов реципиентов могут быть использованы также штам мы E.coli НВ и E.coli 294.
При функционировании промотора триптофанового оперона (trp) в соста ве рекомбинантных плазмид p2FN-(i-F t штаммы, содержащие эти рекомбинантные плазмиды, имеют фенотип и синтезирздат интерферон. Используе- мый промотор триптофанового оперо- на содержит триптофановый оператор и регулируется путем взаимодействия триптофанового репрессора с .этим оператором, которое меняется в зависимости от присутствия в среде триптофана и индуктора -р-индолилакриловой кислоты. В присутствии триптофаиа репрёссор связан с оператором и экспрессия гена интерферона незначительна, что позволяет исключить возможную роль интерферона как ингибитора роста и деления клеток. При пониженной концентрации триптофана шш добавлении индуктора репрёссор теряет сродство к оператору и промотор начинает функционировать вследствие чего инициируется синтез интерферона. В качестве метода индукции использзют снижение концентрации триптофана в среде. Для этой цели клетки растят вначале в богатой триптофансодержащей среде LB, а затем разбавляют в 25 раз синтетической средой М9 с добавлеьшем казами- новых кислот.
Штаммы, содержащие рекомбинантные плазмиды plFN-al-F trpi, plFH- -F trp2, plFN-o(.-F trp3, piFN-oL-F trp4 и plFN-dL-F trp5, отличаются от штамма-реципиента E.coli К802 только по признакам, придаваемым плазмидой, т.е. являются ампициллини тетрациклин устойчивыми и сиите- зируют интерферон. Штаммы E.coli/ :/pIFN-flL-F trp характеризуются слегдующими признаками:
с перитрихиальными жгутиками, rpai отрицательные, неспороносные.
кие, круглые, блестящие, край ровный, при одинаковых условиях более мелкие, по сравнению с колониями изогенных штаммов E.coli, содержащих плазмиду pBR-322. При росте в жидких средах - LB и М9 - с добавлением казаминовых кислот образуют равномерную муть.
к ампициллину (до 100 мкг/мл при росте на твердой среде) и в незначительной степени к тетрациклину (10 мкг/мл). В условиях индукции промотора устойчивость к ампициллину сохраняется на прежнем уровне, устойчивость к тетрациклину возрастает до 80 мкг/мл в среде М9.
При поддержании штаммов в течение одного года в присутствии ампициллина не наблищается перестройки или утери плазмид.
Изобретение иллюстрируется следующими фигурами графических изобра жеНИИ.
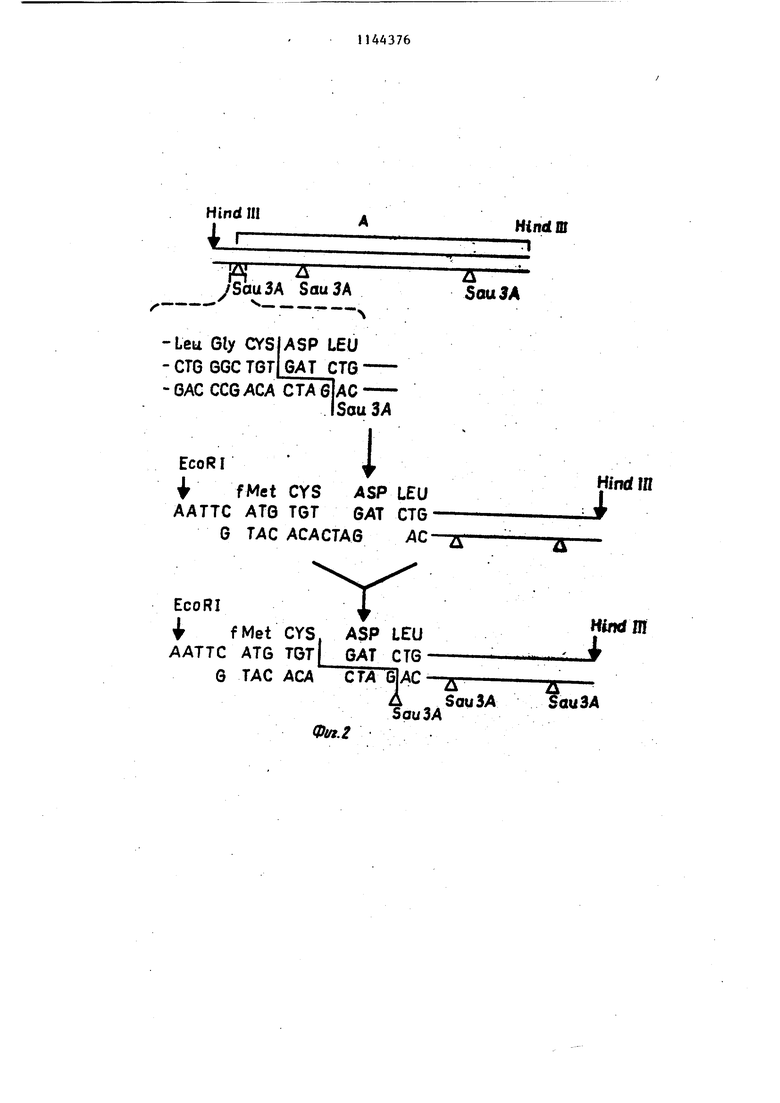
Фиг. 1. Нуклеотидная последовательность клонированного гена человеческого лейкоцитарного интерферона и его аминокислотная последовательность. Приведена последовательность комплементарной цепи ДНК, адекватная последовательность мРНК. Аминокислотная последовательность зрелого Интерферона дана прописными буквами, а части, соответствующей предшественнику, - строчными.
Фиг. 2. Схема реконструкции гена интерферонаоС-F для его прямой экспрессии в клетках E.coli,
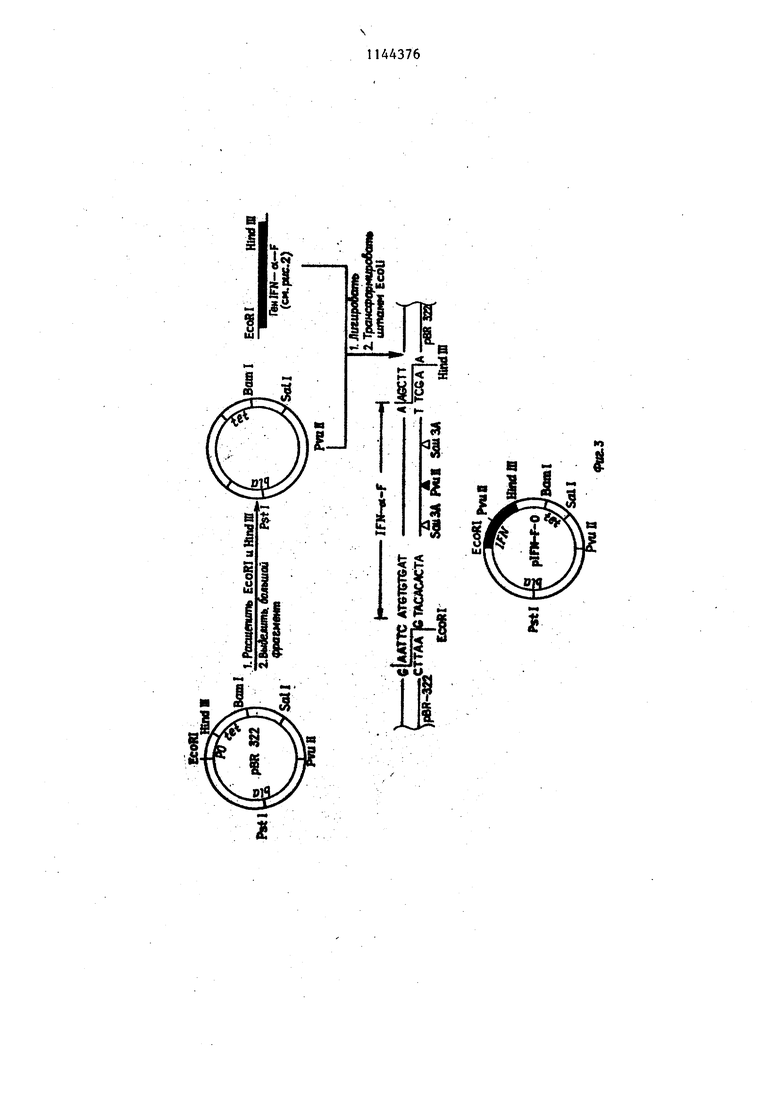
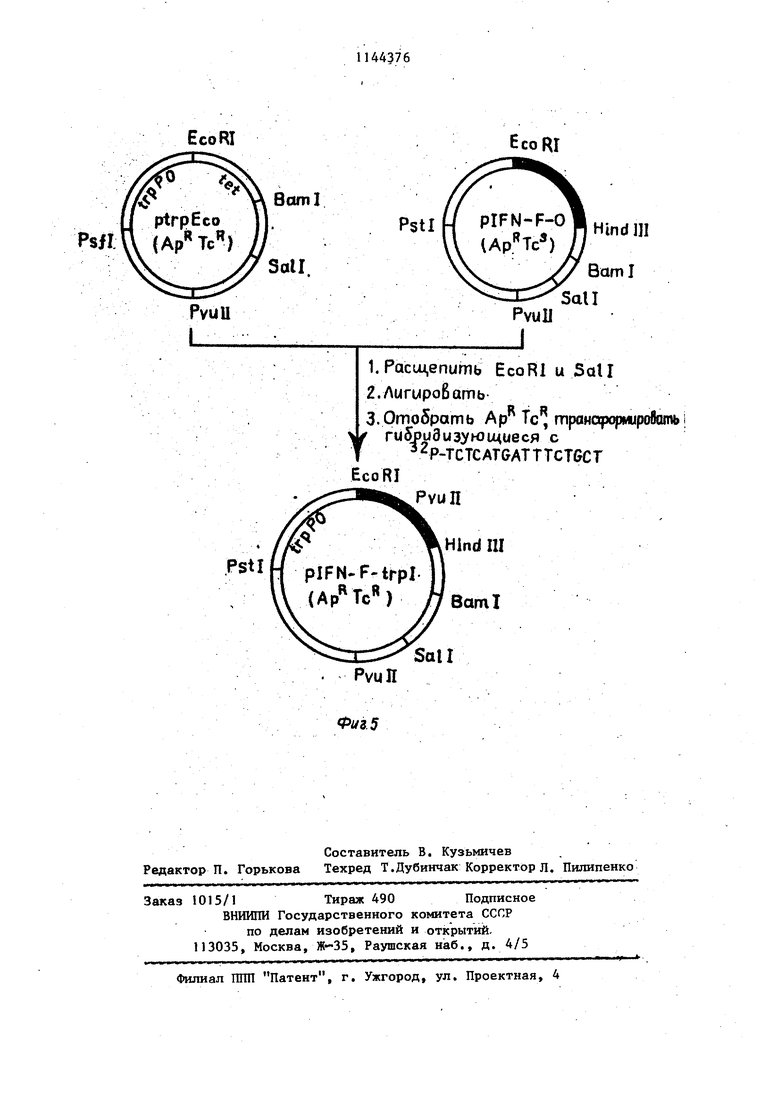
Фиг. 3. Схема получения и реет- риктная карта неэкспрессирующей плазмиды plFN-(, содержащей ген интерферона . Фиг. 4, Генетическая и рестрикт- ная карта исходной экспрессирую- щей плазмиды ptrpEco и последовательность промоторно-операторного участка trp оперона E.coli, исполь зуемого в качестве сигнального элемента. Стрелка указьшает начало и направление транскрипции. Подчеркну та последовательность названная в тексте собственно промоторной. Пред полагаемый участок Шайн-Дальгарно обведен. Фиг. 5. Схема конструкции экспре сирующей плазмиды piFN-tst-F trpl и ее физико-генетическая карта. Зачер нен участок, кодирующий интерферон oC-F. Пример 1. Получение реком- бинантной плазмиды р1№-о(-Ftrpl. Дпя получения плазмиды p,lFfvi-d-Fit-pi 1 мкг сконструированной плазмиды plFN-d-F-O, содержащей полный, ген . lFN-xs(.-F Г расщепляют 2 ед. рестрикци онной-эндонуклеазы EcoRI и 2-ед. Sall.l Мкг плазмиды ptrp Есо расщепляют 2 ед. EcbRl и 2 ед. Satl. Смесь расщепленных пла-змид прогревают при 74°С в течение 10 мин. Затем добавляют 10 мМ АТФ до концентрации 200 М, лигазный буфер (66 мМ трис- HCl, рН 7,5, 5 мМ MgCl,,,, 5 мМ ДТТ), 1 ед. лигазы и инкубируют при в течение 18ч. Полученной смесью ДНК трансформируют клетки E.coli. 1. Получение компетентных клеток 10 клеток со свежего ночного ко сяка суспендируют в 5 мл iВ среды. К 100 мл/В среды, содержащей.. 50 мкг/мл тимидина, добавляют 1 мл смыва с косяка и подращивают на-качалке при до плотности A.goO,6 Суспензию охлаждают 15-20 мин во льду и клетки собирают центрифугированием при (4000 об/мин, 5 ми центри-фуга К-23, Janets Ку). Затем, клетки суспендируют в 50 мл холодно го раствора, содержащего 60 мМ CaCl 10 мМ MOPS, рН 7,1, .0,5% глюкозы и 50 мкг/мл тимидина, инкубируют в этом растворе 5 мин при , центри фугируют в указанном выше режиме и суспендирукгт в 5 мп того же раствора. Вьщерживают 1 ч при . 2. Процедура трансформации. К 100 мкл раствора ДНК (0,010,1 мкг) добавляют 200 мкл полученных компетентных клеток и инкубируют вначале при в течение 5 мин, а затем при 42°С в течение 2 мин. Смесь разбавляют 1 мл LB среды и подращивают на качалке при в течение 45-90 мин. Затем высевают на чашки Петри с 1,5% агаром на LB среде и необходимым антибиотиком (ампициллин -. 50 мкг/мл, тетрациклин - 20 мкг/мп) и инкубируют при 37С в течение ночи. 3.Получение реплик. Клеточную суспензию после трансформации высевают непосредственно на фильтры HATF 29325 (Millipore, диаметром 10 см), которые помещают на на чашку Петри с агаром и соответствующим антибиотиком, инкубирует ночь при . Для получения реплики на стекло помещают стерильный лист Ватман ЭММ фильтр HATF с выращенными колониями, чистый фильтр HATf, лист Ватман и покрывают еще одним стеклом. Сверху помещают груз весом 1 кг. выдерживают в течение10 с. Нитро- целлюлозные фильтры разъединяют, и фильтр с реплицированными колониями переносят на чашку Петри с агаром. Чашку инкубируют при 37°С в течение 3-6 ч, после чего фильтр, переносят на чашку с агаром, содержащем 200 мкг/мл хлорамфеникола и инкубируют ночь при 37 С. 4.Обработка фильтров. Лист Ватман ЗММ пропитьтают 0,5М NaOH, помещают на него нитроцеллю- лозньш фильтры с реплицированными колониями и Вьщерживают 10 мин. Фильтры снимают с листа Ватмана, подсушивают на воздухе и повторяют утса- занную процедуру. После сушки фильтры помещают на лист Ватмана ЗММ, пропитанный 1М ацетатом аммония, выдерживают 10 мин, сушат и повторяют указанную процедуру. По окончании обработки фильтры сушат в вакуумном шкафу при температуре 80°С в течении 4ч. 5.Гибридизация. Высушенные фильтры замачивают в J мл раствора, содержащего 0,75М WaCl; 0,15М трис; рН 8,0; 0,5 мМ ЭДТА; 0,02% фикола; 0,02% поливинилш poлJшдoнa; 0,02% альбумина; 0,1% додещш сульфат а натрия; 250 мк тРНК; 0,5- 210® имп/мин синтетического
гексадекануклеотида, комплементарного фрагменту гена iFNi-xjt-F. Фильтры запаивают в полиэтиленовый пакет , инкубируют 30 мин при. темпера 5 туре , затем плавно опускают температуру до 37°С и инкубируют ночь при этой температуре. По окончании инкубации фильтры четырежды отмывают в растворе, содержащем 10 0,2М трис рН 8,0; 0,5М NaCl 0,5% додецилсульфат натрия при в течение 15 мин, используя по 10 мл раствора на каждый фильтр. Между отмывками фильтры сушат на воздухе. is Высушенные фильтры радиоавтографируют при комнатной температуре в течение ночи, используя рентгеновскую пленку РТ-1..
Гибридизующиеся клоны наращи- 20 вают в 3 мл LB среды, выделяют плазмидную ДНК щелочным способом и подвергают ее рестрикционному анализу. Выявленнзпо таким образом плаз МИДУ pIFN-nit-F trpl нарабатывают в 25 количествах, достаточных для анализа первичной структуры, которую определяют по модифицированному методу Максама-Гилберта.
Пример 2. Получение ре- ЗО комбинантной плазмиды р 1 FN-o(rFtrp2.
1 мкг плазмиды plFH-ot-F trpl расщепляют рестрикционной эндонуклеазой EcoR1, (1 ед.) липкие концы до.страивают ДНК-полимеразой фага Т4 35 в буфере, содержащем 20Мм трис, рН 7,6; ЮмМ 1 мМ ДТТ; по каждого из четырех дезокси- нуклеозидтрифосфатов и 0,5 ед. ДНК- полимер азы фага Т4. Обработку прово- 40 дят при ЗУС в течение 15 мин. Затем добавляют ДНК-лигазу (5 ед.), полученной смесью ДНК трансформируют клетки E.coli и отбирают клоны, как описано в примере 1. В результате 45 получают плазмиду plFN-ct-Fti.
Пример 3. Получение рекомбинантных плазмид plFN- XrFtrp3 и plFlHi-F trp4..
I мкг плазмиды plFH- -FtrpI рас-
щепляют рестрикционной эндонукле- азой Есов1 (1 ед.) и инкубируют в течение 30 мин при в присутствии иуклеазы (1 ед.) в буфере, со-
держащем О,ЗМ NaCl; 4,5i мгпСЦ; 55 ацетата натрия, рН 4,6, а затем сшивают с помощью ДНК-лигазы фага Т4 (5 ед.), трансформируют клетки E.coli и отбирйют клоны, как описано в примере 1. Девяносто полученных клонов анализируют на интерферо- новую активность. Из двух клонов, проявивших наиболее высокую противовирусную активность, выделяют плаз- МИДЫ, в которых определяют последовательность участка между собственно промоторной областью иначалом гена. Пример 4. Получение рекомбинантной плазмиды plFN-flt-Ftrp5.
а)I мкг плазмиды ptrpEco расщепляют рестрикционной эндонуклеазой Есоб1 (1 ед.) и инкубируют с ДНК-полимеразой (0,5 ед.), как описано в примере 2, но вместо смеси четырех дезоксинукзеозидтрифосфатов используют только один дезоксигуанозин- трифосфат. ДНК осаждают спиртом и инкубируют с S нуклеазой, как описано в примере 3. ДНК высаживают спиртом, растворяют в буфере для рестрикции SaEl и рестрицируют как в при мере 1.
б)Плазмиду plFN-t)( расщепляют рестрикционной эндонуклеазой- EcoRl, и липкие концы заполняют ДНКполимеразой фага Т4, как описано в примере 2. Полученную ДНК расщепляют рестрикционной эндонуклеазой Sal
в)Смесь расщепленных плазмид
(по 1 мкг каждой) лигируют и транс- формируют ею клетки E.coEi К802, высевая трансформанты на нитроцеллю лозные фильтры. Отбирают Ар Тс трансформанты, гибридизующиеся с олигонуклеотидным зондом, как описано в примере I. Плазмиду plFN- i-.Ftrp5 из одного полученного клона анализируют по последовательности участка между регуляторной промоторной областью и началом гена.
Пример 5. Получение штаммов продуцентов лейкоцитарного интерфе- poHaol -Р человека,
Рекомбйнантные плазмиды pIFN-ot- -Fthp вводят в штамм E.colj К802, как описано в примере 1, .и получают штaммь пpoдyцeнты лейкоцитарного ин- терферонао(- F человека с активностями, указанными в таблице.
Активность определяют методом ин- гибирования цитопатического действия вируса везикулярного стоматита на диплоидные фибробласты человека.
Таким образом, новые рекомбинант- ные плазмиды pjFN-A-Ftrp и итаммы
Е,со 1V, содержащие эти плазмидьрпродуцеиты лейкоцитарного интерфе- рона THnaOt-F человека, позволяют
Свойства штаммов-продуцентов лейкоцитарного интерферонасА-F человека, содержащих рекомбинантные плазмиды серии p1fN-OL-Ff р
Последовательность
Штамм между регуляторной областью и геном
E.coli/plFN-ct-F trpl ЦМПМ В 2885 AAAAAGGGTATCGCGGAATTC ATG5-10 ед/л
E.coli/plFN-ol-F trp2 ЦМПМ В 2886 AAAAAGGGTATCGCGGMTTMTTC ATG5-10 ед/л
E.coli/plFN-di.-F trp3 ЦМПМ В 2887 AAAAAGGGTATCGCCAC ATG5-10 ед/л
;E.coli/plFN- -F trp4 ЦМПМ В 2888 AAAAAGGGTATCGCCA ATG2-10 ед/л
E.coii/plFN-A-F trp5 ЦМПМ В 2889 AAAAAGGGAATTC ATG5-10 ед/л
АТС тег тст сте сес тег GAT сге
S5-IOS
-6ji-Ser- -j -Cyy-ASP-tEO- I06-I6S 166-226 4 227-286 287.-346 347-406 407-466 467-526 S27-S75
значительно увеличить выход интерферона - до 2-10 ед. активности с 1 л бактериальной суспензии.
Выход интерферона на 1 л бактериальной суспензии при
А 550 1
S23 I
10
CAG ACC CAC AGC CTG CGT AAT AC& - -THK-HI5-5ER-l.EO-6/.r-A5H-ARC. 20; 50 AG6 6CC TTG ATA CTC CTG GCA CM ATG GGA АСА АТС ТСТ ССТ ТТС ТСС ТСС CTG AAG GAC ARe-ALA-LEU-ILE-LEU-LEU-ALA-GLN-MET-GLY-ARG-ILE-SER-PRO-PHE-SER-CYS-LEU-LYS-ASP.- . . 40 .. . SO - AGA CAT GAC TTT GGA TTC CCC CAG GAG TTT GAT GGC AAC CAG TTC CAG AAG GCT CAA ARG-HYS-ASP-PHE-GLY-PHE-PRO-GLN 3LU-GLU-PHE-ASP SLY-ASN-GLN-PHE-GLN-LYS-ALA-GLNGCC АТС ТСТ GTC ТС CAT GAG ATG ,.TC CAG CAG ACC TTC AAT CTC TTC AGC АСА AAG GAC AlA-ILi SER-VAL-iEU-HIS-GLU-MET-ILE-GLN-GLH-THR-PHErASN-LEU-PHE-SER-THR-tYS-ASP 80 ..- . :90 TCA TCT COT ACT TOG GAA CAG AGC CTC СТА GAA AAA TTT TCC ACT GAA CTT AAC CAG CAG SER-SER-ALA-THR-TRP-GLU-GLN-SER-LEU-LEU-eLU-LYS-PHE-SER-THR-GLU-LEU-ASN-GLN-GLN.100110 CTG AAT GAC CTG GAA GCC TGC GTG ATA CAG GAG GTT GGG GTG GAA GAG ACT CCC CTG ATG LEU-ASN-ASP-LEU-GLU-ALA-CYS-VAL-ILE-GLN-GLU-VALrGLY-VAL-GLU-GLU-THR-PRO-LEU-METAAT GTG GAC .TCC АТС CTG GCT GTG AAG AAA TAC TTC CAA АСА АТС ACT CTT TAT CTG АСА .ASN-VAL-ASP-SER-ILE-LEU-ALA-VAL-LYS-LYS-TYR-PHE-GLN-ARG-ILE-THR-LEU-TYR-LEU-THRGAG AAG AAA TAfc AGC CCT TGT GCC TGG GAG GTT GTC AGA CCA GAA АТС ATG AGA TCC TTC GLU-LYS-LYS-TYR-SER-PRO-CYS-ALA-TKP-GLO-VAL-VAL-ARG-ALA-GLU-ILE-MET-ARG-SER-PHETCT TTA TCA AAA ATT TTT CAA GAA AGA TTA AGG AGG AAG GAA TAA ACTC SEH-LEU-SER-LYS-ILE-PHE-GLN-GLU-ARG-LEU-ARG-AHG-LYS-GLU ter 60.70 IZO 130 140ISO 160166 Фиг.1
-Leu Gly CYSJASP LEU - CTG GGC TGT GATCTG -GAG CCGACA CTASJAC lSou3A EcoRI I fMet CYS ASP LEU AATTC ATG TOT GAT CTG G TAG ACACTAG AC-TJ Т . - .
чг
EcoRI
Pstl
HcoRl
EcoRI
Pvun
| D | |||
| Goecldel et al | |||
| Nature, 1980, 287, 411-416 | |||
| TGT GAT GTG- CCT, CAG- ACC CAC АбС ATA CTC CT6 6UA CAA ATG 66A-A6A AA& GAG AGA CAT GAQ | |||
| TTT GGA TTC AAC GAG- TTG GAG AAG- GCT CAA OCC ATQ | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Овчинников Ю.А | |||
| и др | |||
| Доклады АН СССР, 1982, 265, 238-242 | |||
