Т и осмотического давления Пу). Способ возможно осуществить при помощи стандартного ЯМР спектрометра.
Способ осуществляют следующим образом.
Приготавливают два образца суспензии клеток в исходном состоянии (при исходных фиксированных значениях осмотического давления П и и температуры Тн). Измеряют в этом состоянии в основном образце амплитуду сигнала ЯМР от протонов внутриклеточной воды Маи, среднее время жизни молекул воды в клетках $ аи и в дополнительном образце - амплитуду сигнала ЯМР от протонов внутриклеточной воды МдИ. Переохлаждают клетки основного образца до температуры замораживания Т и измеряют параметры Моп, и -сщу а- тем замораживают клетки этого образца при температуре Т« и измеряют в этом состоянии амплитуду сигнала ЯМР от протонов внутриклеточной воды Мдо, среднее время жизни молекул воды в клетках Ј п « и осмотическое давлеч
ние внеклеточной среды По,. Затем размораживают клетки основного образца, повышая температуру до исходной Т, и получают исходное значение осмотического давления внеклеточной среды Пи, пропорционально изменяя концентрацию компонент во внеклеточной среде до исходной. При исходных значениях осмотического давления П и и темпе - ратуры Т и измеряют параметры
и г
зии
В дополнительном образце понижаю температуру до Т «у и повышают пропорционально концентрацию компонент во внеклеточной среде так, чтобы в переохлажденном состоянии при этой температуре давление внеклеточной среды в этом образце было равно осмотическому давлению П. внеклеточной среды в основном образце, замороженном при температуре Т,.
Измеряют в дополнительном образце в переохлажденном состоянии при Т амплитуду сигнала ЯМР от протонов внутриклеточной водыМдПг,и среднее время жизни молекул воды в клетках tgn. Затем, повышая температуру до исходной Ти и пропорционально изменяя концентрацию компонент во внеклеточной среде до исходного значения, переводят клетки .этого образца вновь в исходное состояние Тк ис- холным значениям П и Т и) и измеря 0
5
0
5
0
5
40
45
50
ют в этом состоянии параметры М°ий
и ° и °аии
По отношениям амплитуд в основно. и дополнительном образцах судят о проценте клеток, мембраны которых повреждены:в результате увеличения - осмотического давления внеклеточной среды при замораживании; из-за образования льда при замораживании;
вследствие рекристаллизации льда при отогреве.
По отношениям времен жизней в основном и дополнительном образцах судят об изменении диффузионной водной проницаемости (ДВП) вследствие: изменения осмотического давления в цикле замораживание - отогрев; механического воздействия льда при замораживании; действия льда в ходе цикла замораживание - отогрев.
Если в процессе замораживания клетки не повреждаются, то по отношениям амплитуд и времен жизней молекул воды в клетках основного и дополнительного образцов дополнительно судят об изменении ДВП в результате увеличения -осмотического давления внеклеточной среды в замороженном состоянии.
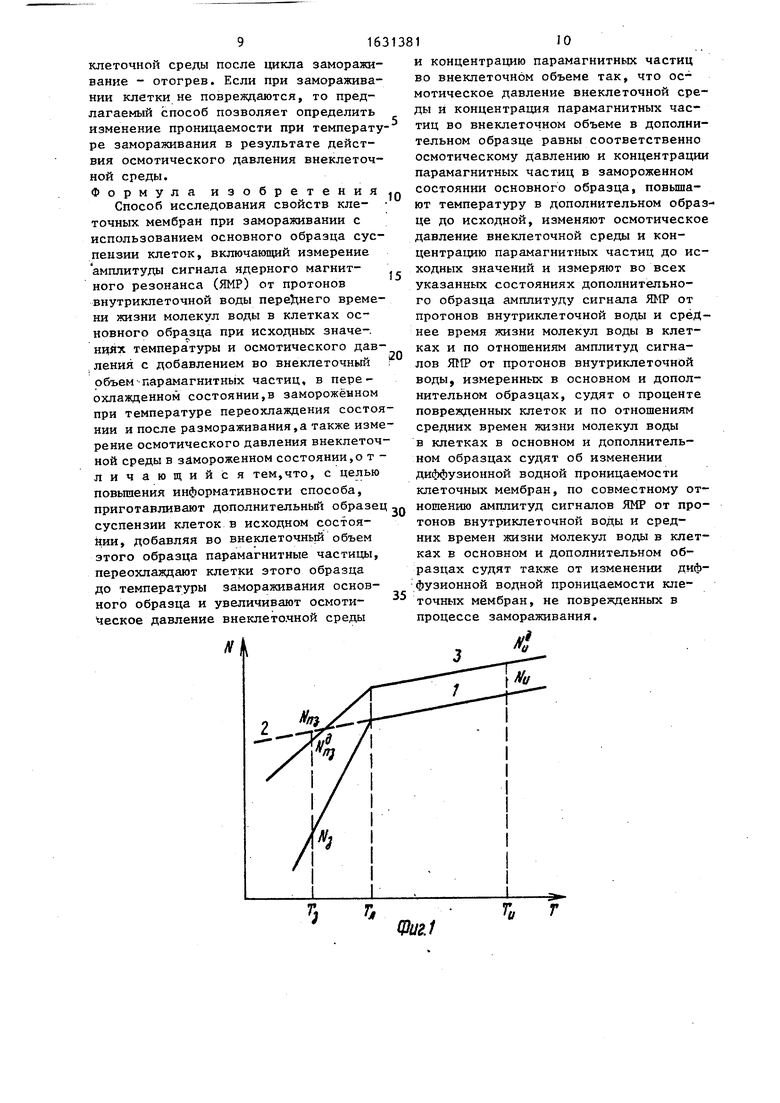
На фиг.1 схематически представлены упрощенные зависимости изменения количества неповрежденных клеток в основном N (кривая 2 - без замораживания, кривая 1 - с замораживанием) и дополнительном N (кривая 3) образцах при понижении температуры образцов от исходной Т до температуры замораживания (переохлаждения) TV, Тд - температура начала гибели клеток от образования льда в основном образце, N«, и N - количество клеток в основном и Дополнительном образцах при температуре Т..
Принимая во внимание фиг.2, оценивают процент погибших клеток в результате образования льда при замораживании
V(
1-OyNM)/().(1)
Очевидно, что
Van v™ и vav V«T
где Уаи и Уал, - объем воды в клетке в исходном и замороженном состояниях в основном образце;
Э 9 5t6
V и VQ - те же величины в до полнительном образце в исходном и переохлажденном при температуре замораживания Т состояниях соответственно.
Амплитуды сигнала ЯМР от протонов внутриклеточной воды в исходном сое- тоянии для обоих образцов, замороженном для основного и переохлажденном для дополнительного образца состоянии соответственно записываются
М«ИЗ k Vavi NH
ML k V« NH
V k v«yNv
Ma«V k V«VN r
где k - коэффициент пропорциональности. Тогда выражение (1) можно представить в виде
Ц- 1-Смв /мви)/() . (2)
Исходя из кривой 3 (фиг.1), оценивают процент гибели клеток от осмотического эффекта
где Мии - число клеток в исходном состоянии в дополнительном образце после перевода их из исследуемого переохлажденного состояния опять в исходное состояние. Здесь считается, что при обратном переводе число клеток в дополнительном образце сохраняется, т.е. Nn NjJM. Амплитуды сигналов ЯМР от внутриклеточной воды в дополнительном образце до и после перевода клеток этого образца в исследуемое (переохлажденное) состояние записываются соответственно
.9
м
аи
ьЛи Ни - м-ии
k vavw Nnn-9 9
где VQV| и Удии - объем воды в клетке в дополнительном образце при тех же условиях. Очевидно, что V . Поэтому
оии
U,
1 - MaV
аи
(4)
Определяют суммарный процент клеток, погибших в основном образце от осмотического эффекта и механического действия льда после всего цикла замораживание - отогрев
Лх NH-NHH /N« 1-м т/маи (5)
где Маи k-VQvi-Nw
и Mq(iM Ь ии НИи- амплитуды сигналов ЯМР от внутриклеточной воды в основном образце в исходном состоянии до и после цикла заморажива-: ние - отогрев.
Причем очевидно, что V. лии т.е. внутриклеточный объем до замораживания равен аналогичной величине после отогрева.
Процент погибших клеток от действия льда в процессе размораживания выражается соотношением
й2-Л«-АЛЈ или Я О
Ч МаЛ + ,Г )- 1 . (6)
Определяют также процент клеток, погибших ох совместного действия осмотического эффекта и льда при замораживании. Очевидно, что
+ 2 , (Ма3ии/ма9и) 9 з (мп„/мди)/(мли, /м°м).
«V
a«j
(7)
50 Оценивают изменение проницаемости мембраны. По определению ДВП Р равно
РҐ
(8)
где Уд - объем воды в клетке;
S - площадь поверхности клеточной мембраны;
va - среднее время жизни воды в клетке.
Проницаемость в исходном, замороженном и переохлажденном состояниях обоих образцов соответственно выражается
ри ws v р
-VJL/CS.,)(9)
Изменение проницаемости вследствие суммарного действия осмотического давления и льда после всего цикла замораживание - отогрев
UR.. (Ри- ,
ОЛ
(17)
Изменение проницаемости вследствие механического действия льда после цикла замораживание - отогрев



| название | год | авторы | номер документа |
|---|---|---|---|
| Способ определения осмотических свойств клеточных мембран | 1984 |
|
SU1224693A1 |
| Способ определения количества и консистенции сырой биомассы | 1988 |
|
SU1566274A1 |
| Способ определения морозостойкости растительных образцов | 1990 |
|
SU1738150A1 |
| УСИЛЕНИЕ ЯДЕРНОГО МАГНИТНОГО РЕЗОНАНСА (ЯМР) И МАГНИТОРЕЗОНАНСНОЙ ВИЗУАЛИЗАЦИИ (МРВ) В ПРИСУТСТВИИ ГИПЕРПОЛЯРИЗОВАННЫХ БЛАГОРОДНЫХ ГАЗОВ | 1997 |
|
RU2186405C2 |
| КРИОЗАЩИТНЫЙ АГЕНТ, СОДЕРЖАЩИЙ ЭКЗОПОЛИСАХАРИД ИЗ PSEUDOALTEROMONAS SP. СY01 | 2017 |
|
RU2779448C2 |
| СПОСОБ КРИОКОНСЕРВАЦИИ ЗАГОТОВЛЕННОЙ ТКАНИ МЛЕКОПИТАЮЩИХ ИЛИ КУЛЬТИВИРОВАННОГО ЭКВИВАЛЕНТА ТКАНИ И УСТРОЙСТВО ДЛЯ ЕГО ОСУЩЕСТВЛЕНИЯ | 1996 |
|
RU2178865C2 |
| КОМБИНИРОВАННЫЙ КРИОПРОТЕКТОР "ДИМЕТИЛСУЛЬФОКСИД/РЕОПОЛИГЛЮКИН" ДЛЯ КРИОКОНСЕРВАЦИИ СТВОЛОВЫХ КЛЕТОК И СПОСОБ ИХ КРИОКОНСЕРВАЦИИ ДЛЯ КЛИНИЧЕСКОГО ПРИМЕНЕНИЯ | 2014 |
|
RU2563117C1 |
| СПОСОБ ЗАМОРАЖИВАНИЯ ВОДНЫХ РАСТВОРОВ ГЕЛЕОБРАЗУЮЩИХ БИОЛОГИЧЕСКИХ ВЕЩЕСТВ ДО АМОРФНОГО СОСТОЯНИЯ И ПЛАВЛЕНИЯ АМОРФНЫХ ОБРАЗЦОВ ЭТИХ ВЕЩЕСТВ | 1995 |
|
RU2103870C1 |
| Способ обнаружения протонного обмена в гетерогенных системах | 1983 |
|
SU1112267A1 |
| СПОСОБ КРИОСОХРАНЕНИЯ МОРСКИХ МИКРОВОДОРОСЛЕЙ | 2012 |
|
RU2496318C1 |
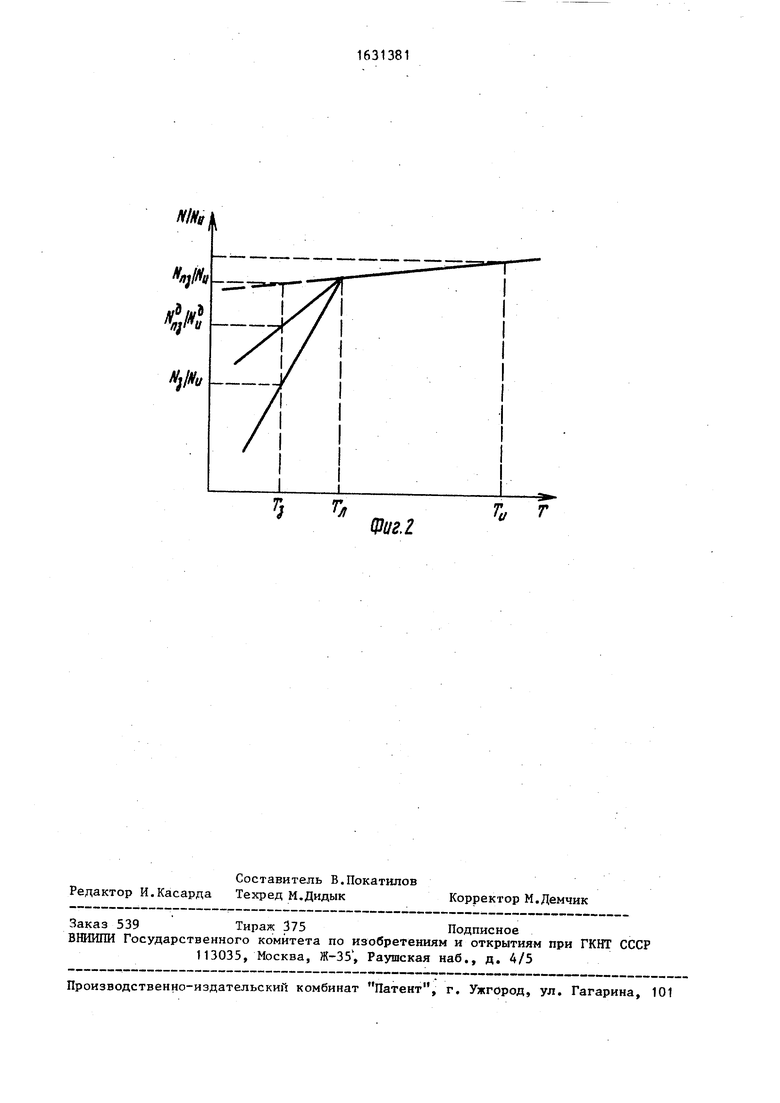
Изобретение может быть использовано в биотехнологии, пищевой промышленности, медицине и биологии. Целью изобретения является повышение информативности способа. В способе берут дополнительный образец суспензии клеток и измеряют в этом образце при исходных значениях темИзобретение относится к фи.ичес- ким методам исследования вещества на основе явления ЯМР и может быть использовано в биотехнологии, медицине, биологии и других областях естествен- ных наук, где в процессе замораживания или образования льдоподобных структур (типа клатратов газов) необходимо проводить быстрый и точный контроль свойств клеточных мембран или замкнутых полупроницаемых оболочек. Цель изобретения - повышение информативности способа. пературы и осмотического давления внеклеточной среды амплитуду сигнала ядерного магнитного резонанса (ЯМР) от протонов внутриклеточной воды и среднее время жизни молекул воды в клетках, переохлаждают этот образец до температуры замораживания и повышают в нем пропорционально концентрацию компонент внеклеточной среды таким образом, чтобы осмотическое давление внеклеточной среды в дополнительном образце при этой температуре в переохлажденном состоянии было равно осмотическому давлению внеклеточной среды в замороженном состоянии основного образца, измеряют те же параметры в переохлажденном состоянии, далее переводят образец в исходное состояние и измеряют в этом состоянии амплитуду сигнала ЯМР от протонов внутриклеточной воды и среднее время молекул воды в клетках. 2 ил. i (Л На Лиг.1 схематически представлены упрощенные зависимости изменения количества неповрежденных клеток в основном (сплошная кривая 1 и пунктирная кривая 2) и дополнительном (кривая 3) образцах при понижении температуры образцов от исходной температуры на фиг. 2 - те же кривые с нормированным значением ординаты N/N и , где N и N9 - количество клеток в основном и дополнительном образцах соответственно, NM и N.. - количество клеток в-основном и дополнительном образцах со-, ответственно в исходном состоянии (при исходных значениях температуры о со со 00
(12)
ДВП после всего цикла замораживание - отогрев
Рии W(S W W s4ami 25 /(И,
(13)
где VquH VqM.
ДВП в дополнительном образце 30 после перевода из переохлажденного состояния в исходное
vot,
Х,И/(), гл-е V4M« V™ Ґа9ии vaV
(14)
Здесь считается, что S const во всех состояниях и fyjj ajвследствие механического контакта со льдом.
Изменение ДВП в результате действия льда при замораживании выражается
AR - 1AV4
Изменение ДВП от осмотического
действия среды после переохлаждения при температуре IV , осмотическом давлении Па и возврата в исходное состояние
лкэ-ля -(Р8-РЭ Vr3-i-Ј3 /f9-
URo fiRo (РИ Рии)/Ги-1 WW - -(16)
ARA
(М$
йии °аии Мин
ЛиО/tf-u- 1ЛЛ-и.
(18)
Если при замораживании клетки не повреждаются, то изменение ДВП при температуре TV выражается соотношени ем у
AR«of pn -Pb)/pnje1- Mfl an /
,
- ,
П f)
(19)
где Мяп, и
an
1 и ьап
- амплитуда сигнала ЯМ от протонов внутриклеточной воды и среднее время жизни молекул воды в клетках в основном образце, в переохлажденном до температуры Т состоянии.
В этом случае изменение проницаемости в результате увеличения осмотического давления при замораживании с учетом (15) записывается
URo) АК -Јап}/Ц- ма$ Ц / МвпуЈп.(20)
Таким образом, введение указанным способом дополнительного образца клеток позволяет определить при любом строении клеточной стенки отдельно процент клеток, мембраны которых повреждены в результате увелич ия осмотического давления внеклеточной среды при замораживании, действия льда как при замораживанци, так и при размораживании, а также изменени ДВП вследствие механического воздействия льда при замораживании, суммарного действия льда при замораживании и оттаивании и интегрального действия осмотического давления внеклеточной среды после цикла замораживание - отогрев. Если при замораживании клетки не повреждаются, то предлагаемый способ позволяет определить изменение проницаемости при температуре замораживания в результате действия осмотического давления внеклеточной среды.
Формула изобретения
Способ исследования свойств клеточных мембран при замораживании с использованием основного образца суспензии клеток, включающий измерение амплитуды сигнала ядерного магнитного резонанса (ЯМР) от протонов внутриклеточной воды переднего времени жизни молекул воды в клетках основного образца при исходных значениях температуры и осмотического давления с добавлением во внеклеточный объем парамагнитных частиц, в переохлажденном состоянии,в замороженном при температуре переохлаждения состоянии и после размораживания,а также измерение осмотического давления внеклеточной среды в замороженном состоянии,о т - личающийся тем,что, с целью повышения информативности способа,
приготавливают дополнительный образец ™ ношению амплитуд сигналов ЯМР от просуспензии клеток в исходном состоянии, добавляя во внеклеточный объем этого образца парамагнитные частицы, переохлаждают клетки этого образца до температуры замораживания основного образца и увеличивают осмотическое давление внеклеточной среды
35
тонов внутриклеточной воды и средних времен жизни молекул воды в клетках в основном и дополнительном образцах судят также от изменении диффузионной водной проницаемости клеточных мембран, не поврежденных в процессе замораживания.
П
5
0
5
0
и концентрацию парамагнитных частиц во внеклеточном объеме так, что осмотическое давление внеклеточной среды и концентрация парамагнитных частиц во внеклеточном объеме в дополнительном образце равны соответственно осмотическому давлению и концентрации парамагнитных частиц в замороженном состоянии основного образца, повышают температуру в дополнительном образце до исходной, изменяют осмотическое давление внеклеточной среды и концентрацию парамагнитных частиц до исходных значений и измеряют во всех указанных состояниях дополнительного образца амплитуду сигнала ЯМР от протонов внутриклеточной воды и среднее время жизни молекул воды в клетках и по отношениям амплитуд сигналов ЯМР от протонов внутриклеточной воды, измеренных в основном и дополнительном образцах, судят о проценте поврежденных клеток и по отношениям средних времен жизни молекул воды в клетках в основном и дополнительном образцах судят об изменении диффузионной водной проницаемости клеточных мембран, по совместному от
тонов внутриклеточной воды и средних времен жизни молекул воды в клетках в основном и дополнительном образцах судят также от изменении диффузионной водной проницаемости клеточных мембран, не поврежденных в процессе замораживания.
PUit
Фиг. 2
| Способ определения осмотических свойств клеточных мембран | 1984 |
|
SU1224693A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Сахаров Б.В.и др | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Железнодорожный снегоочиститель | 1920 |
|
SU264A1 |
