Изобретение относится к области генной инженерии, биохимии и трансформации растений, в частности касается трансформации растительных клеток, приводящей к экспрессии химерного гена, кодирующего белок, токсичный для жесткокрылых насекомых.
Bacillus thuringiensis (B.t.) является спорообразующей почвенной бактерией, которая известна как бактерия, способная продуцировать параспоровый кристаллический белок, токсичный для широкого круга насекомых. Большинство штаммов активны по отношению к чешуекрылым насекомым (моли и мотылькам) и немногие штаммы активны по отношению к двукрылым насекомым (москитам и комарам, см. Aronson и др., 1986 г.) Токсиновые гены от различных штаммов были клонированы, и токсины были экспрессированы в гетерологических хозяйствах (Schnepf и др. 1981 г.; Klier и др., 1982 г.). За последние годы были идентифицированы штаммы В. t. вида tenebrionis (B.t.t., Krieg и др., 1983 г; Krieg и др. , 1984 г.) и В.t. вида san diego (B.t. sd., Herrnstadt и др., 1986) как штаммы, активные по отношению к жесткокрылым насекомым. Был клонирован токсиновый ген от В.t.sd., но токсин, продуцируемый в Е.сoli был больше по своему объему, чем токсин из кристаллов В.t.sd., но активность этого рекомбинантного токсина B.t.sd. была меньше.
Насекомые, чувствительные к действию белкового токсина бактерий Bacillus thuringiensis колеоптеранового типа (для жесткокрылых) включают (но не ограничиваются ими); жука колорадского (Leptinotarsa decemlineata), долгоносика хлопкового (Anthonomus grandis), желтого мучного червя (Tenebrio molitor), жука-листоеда (Pyrrhalta luteola) и южную блошку длинноусую (Diabrotica undecimpunctata howardi).
Таким образом, можно заранее предвидеть возможности растений, выращенных с использованием способов генной инженерии, которые обнаруживают токсичность или стойкость к жесткокрылым насекомым, если такие растения могут быть трансформированы таким образом, что экспрессируют токсин колеоптеранового типа в инсектицидно эффективном количестве. Сельскохозяйственные культуры, имеющие важное промышленное значение, на которые отрицательно воздействуют жесткокрылые насекомые, включают люцерну, хлопок, кукурузу, картофель, капусту (редьку), рис, табак, томаты, сахарную свеклу и подсолнух.
Хотя некоторые химерные гены были экспрессированы в трансформированных растительных клетках и растениях, эта экспрессия никак не является прогрессирующим. В частности, особенно проблематичная экспрессия белков токсина В.t. лепидоптеранового типа (для чешуекрылых). Имеющиеся в литературе данные, как было теперь установлено, касающиеся токсиновых белков лепидоптеранового типа (для чешуекрылых) В.t. в растениях, не распространяются на токсиновые белки В.t. колеоптеранового типа (для жесткокрылых). Этот вывод прямо противоположен существующим ранее утверждениям, согласно которым считалось, что можно осуществлять одни и те же генетические манипуляции для получения полезного выражения таких токсинов в трансформированных растениях.
Согласно одному из аспектов изобретения предусматривается способ получения генетически трансформированных растений, которые выражают токсичность по отношению к жесткокрылым насекомым, включающий:
(а) вставку в генном растительной клетки, чувствительной к жесткокрылым насекомым, химерного гена, включающего:
(i) промотор, который функционирует в растительных клетках таким образом, что вызывает продуцирование рибонуклеиновой кислоты (РНК);
(ii) последовательность дезоксирибонуклеиновой кислоты (ДНК), которая вызывает продуцирование последовательности РНК, кодирующей токсиновый белок Bacillus thuringiensis колеоптеранового типа; и
(iii) 3' нетранслированную последовательность ДНК, которая функционирует в растительных клетках таким образом, что вызывает присоединение аденилатнуклеотидов к 3' концу последовательности РНК;
(b) получение трансформированных растительных клеток и
(с) регенерацию из трансформированных растительных клеток генетически трансформированных растений, проявляющих стойкость к жесткокрылым насекомым.
Согласно следующему аспекту изобретения предусматривается химерный растительный ген, включающий в последовательном расположении;
(а) стимулятор, который функционирует в растительных клетках таким образом, что вызывает продуцирование РНК;
(b) последовательность ДНК, которая вызывает продуцирование последовательности РНК, кодирующей токсиновый белок Bacillus thuringiensis колеоптеранового типа; и
(с) 3' нетранслированную область, которая функционирует в растительных клетках таким образом, что вызывает присоединение полиаденилатнуклеотидов к 3' концу последовательности РНК.
Согласно следующему аспекту изобретения предусматриваются также бактериальные клетки, трансформированные растительные клетки и векторы трансформации растений, которые содержат соответственно ДНК, включающую указанные элементы (а), (b) и (с).
Согласно следующему аспекту данного изобретения предусматривается видоизмененное растение, которое включает трансформированные растительные клетки, как описано выше, которое проявляет токсичность по отношению к жесткокрылым насекомым. Кроме того, изобретение охватывает семена, которые продуцируют описанные трансформированные растения.
На фиг. 1 показаны ДНК пробы, используемые для выделения токсинового гена В. t. t.; на фиг. 2 - этапы, используемые для получения плазмиды pMON 5432; на фг. 3 - ориентация фрагмента 3,0 кб Hind III, кодирующего токсиновый ген в рМОN 5420 и рМON 5421, относительно мгновенной связующей молекулы PUC119; на фиг. 4 - операция, осуществляемая для построения последовательности токсинового гена В.t.t., содержащегося в рМОN 5420 и рМON 5421; на фиг. 5-14 - последовательность ДНК и расположение ограничительных точек для ORF 1932 bp. гена B.t.t., кодирующего 644 аминокислотный токсиновый белок; на фиг. 15 - показаны полосы, наблюдаемые для токсина В.t.t. путем анализа SDS-PAGE (электрофорез в полиакриламидном геле); на фиг. 16 - N-концевая группа белков, выраженных от токсинового гена В.t.t. или протеолитически продуцированных в условиях ин виво в В.t.t.; на фиг. 17-18 - видоизмененные гены В.t.t., используемые для анализа критичности 2-терминальной части токсина; на фиг. 19-20 - видоизмененные гены В.t.t., используемые для анализа критичности N-терминальной части токсина; на фиг. 21 - делеции, продуцированные при оценке мутантов токсиновых белков В.t.t.; на фиг. 22 - этапы, осуществляемые для получения плазмид рМОN 9758, рМОN 9754 и рМОN 9753; на фиг. 23 - этапы, осуществляемые для получения плазмиды рМОN 9791; на фиг. 24 - этапы, осуществляемые для получения плазмиды рМОN 9792; на фиг. 25 - плазмидная карта для кассетного вектора трансформации растения pМON 893; на фиг. 26 - этапы, осуществляемые для получения плазмиды рМОN 9741; на фиг. 27 - этапы, осуществляемые для получения плазмиды рМОN 5436; на фиг. 28 - элементы, включающие Т-ДНК область неусиленного Agrobacterium ACO; на фиг. 29 - последовательность ДНК для усиленного промотора СаМV 35S.
Изобретение охватывает способ трансформации растений с целью проявления токсичности по отношению к чувствительным жесткокрылым насекомым. В частности, изобретение охватывает трансгенные растения, которые выражают токсиновый белок колеоптеранового типа (для жесткокрылых) Вacillus thuringiensis в инсектицидной концентрации.
Согласно одному из аспектов, изобретение охватывает химерные гены, которые функционируют в растениях и продуцируют трансгенные растения, которые проявляют токсичность по отношению к чувствительным жесткокрылым насекомым. Выражение растительного гена, который существует в форме двухниточной ДНК, включает транскрипцию одной нити ДНК посредством РНК полимеразы, в результате чего продуцируется информационная РНК (мРНК), и обработку первичной транскрипты (мРНК) внутри ядра клетки. Эта обработка включает 3' нетранслированную область, которая соединяет полиаденилатнуклеотиды с 3' концом информационной РНК.
Транскрипция ДНК приводящая к продуцированию информационной РНК (мРНК), регулируются областью ДНК, которая обычно рассматривается как промотор. Эта промоторная область включает последовательность нуклеотидов, которая дает сигнал РНК полимеразе к ассоциации с ДНК, и инициирует продуцирование транскрипты мРНК с использованием нити молекулы ДНК, расположенной ниже от промотора, как модель для получения соответствующей нити РНК.
В литературе уже описаны различные промоторы, которые активны в растительных клетках. Эти промоторы включают нопалинсинтазу (NOS), октопинсинтазу (OCS) и маннопинсинтазу (МАS), которые несут индуцирующие опухоль плазмиды Agrobacterium tumefaciens, промоторы вирусамозаики цветной капусты (СаМV) 19S и 35S, и индуцируемый светом промотор от небольших субмолекулярных структур риболозо-бисфосфаткарбоксилазы (ss RUBISCO, очень обильный растительный полипептид). Эти типы промоторов были использованы для различных типов построений ДНК, которые были выражены в растениях; смотри, например, РСТ публикацию WO 84/02913 (Rogers и др., Монсанто).
Согласно изобретению могут использоваться промоторы, которые уже известны или которые, как обнаружено, вызывают продуцирование транскрипты мРНК в растительных клетках. Пригодные для данной цели промоторы могут включать как промоторы, которые получены от гена, который естественно выражен в растениях, так и синтетические последовательности промотора, которые могут включать избыточные или гетерологические усиливающие последовательности. Выбранный промотор должен быть способен вызывать выражение, достаточное для продуцирования эффективного количества токсинового белка для того чтобы придать растению токсичность к жесткокрылым насекомым. Для специалистов в данной области должно быть ясно, что количество токсинового белка, необходимого для индуцирования желаемой токсичности, может изменяться в зависимости от типа жесткокрылого насекомого, от которого должно быть защищено растение. Хотя промоторы СаMV 35S, ss RUBISCO и МАS являются предпочтительными, следует иметь в виду, что эти промоторы могут и не быть оптимальными для всех случаев, охватываемых изобретением.
мРНК, продуцируемая химерным геном, также содержит 5' нетранслированную основную последовательность. Эта последовательность может быть получена от специфического отобранного промотора, такого как промотор СаМV 35S, ss RUBISCO или МАS. 5' Нетранслированная область также может быть получена от другой подходящей последовательности эйкариотических генов или синтетических генов. Для специалистов в данной области должно быть известно, что требуемая функциональность 5' нетранслированной основной последовательности заключается в усилении связывания транскрипты мРНК с рибосомами растительной клетки для усиления трансляции мРНК при продуцировании кодированного белка.
Химерный ген также содержит структурную кодирующую последовательность, которая кодирует токсиновый белок Bacillus thuringiensis колеоптеранового типа или его инсектицидно активный фрагмент. Примерами источников таких структурных кодирующих последовательностей являются B.t. tenebronis и В.t. san diego. Согласно этому, изобретение охватывает структурную кодирующую последовательность от Bacillus thuringiensis вида tenebrionis и его инсектицидно активные фрагменты. Для специалистов в данной области должно быть известно, что может использоваться и другая структурная кодирующая последовательность, гомологичная токсину, кодирующему последовательность В.t.t., согласно описанию изобретения, что также охватывается объемом изобретения.
3' Нетранслированная область содержит сигнал полиаденилирования, который функционирует в растении, вызывая присоединение полиаденилатнуклеотидов к 3' концу молекулы РНК. Примерами таких 3' областей являются: (I) 3' транскрибированные нетранслированные области, содержащие полиаденилатный сигнал индуцирующих опухоль (Ti) плазмидных генов Agrobacterium, например нопалинсинтазный ген (NOS), и (2) растительные гены, типа накапливаемого белкового гена сои и ss RUBSICO. Примерами предпочтительных 3' областей являются области от генов NOS, ss RUBSICO и накапливаемых белковых генов, описанных более подробно в примерах.
Токсиновые белковые гены колеоптеранового типа, отвечающие изобретению, вставляются в геном растения любым подходящим способом. Пригодные для этой цели векторы трансформации растений включают векторы от Ti плазмиды Agrobacterium tumefaciens описанные, например, в публикации Европейского патента ЕРО 131 620 (Rogers и др.), Herrera-Esteella 1983 г., Bevan 1983 г. Кlee 1985 г., и публикация патента ЕРО 120 516 (Schilperoort и др.). Наряду с векторами трансформации растений, полученными от Ti или индуцирующих укоренение (Ri) плазмид Agrobacterium, могут использоваться альтернативные способы для вставки токсиновых белков генов колеоптеранового типа, отвечающих изобретению, в растительные клетки. Такие способы могут включать, например, липосомы, электропорацию, химические реагенты, которые увеличивают поглощение ДНК, и использование вирусов или пыльцы в качестве векторов. При желании в хромосомы растения может быть вставлен более чем один ген такими способами, как повторение трансформации, и более чем однократный цикл отбора.
Модифицированный таким путем растительный материал может быть проанализирован путем точечного покрытия Northern на присутствиe мРНК токсинового белка колеоптеранового типа. Если мРНК токсинового белка (или в слишком малом количестве для титра) обнаруживается, то промотор, используемый в построении химерного гена, заменяется другим, потенциально более сильным промотором, и повторно анализиpуется другое видоизмененное построение. Как вариант, концентрация токсинового белка может быть определена методом иммунопробы, таким как точечный метод Вестерна (Western). Во многих случаях наиболее чувствительным анализом токсинового белка является анализ методом биологической пробы на насекомых.
Контроль может осуществляться во всем объеме регенерированного растения. В любом случае, когда достигается требуемое продуцирование мРНК токсинового белка, и трансформированные клетки (или протопласты) регенерируются в весь объем растений, последние отбираются на стойкость к воздействию жесткокрылых насекомых. Выбор методики осуществления этапа регенерации не является критически важным, причем уже имеются протоколы осуществления операций для хозяев от Leguminosae (люцерна, соя, культурная, клевер, и т.д.), Umbelliferae (морковь, сельдерей, пастернак посевной), Cruciferae (капуста, редька, семя рапса, и т. д. ), Cucurbitaceae (дыня и огурцы), Gramineae (пшеница, рис, кукуруза и т.д.), Solanaceae (картофель, табак, томаты, перец), Malvaceae (хлопок, и т.д.), Chenopоdiaceae (сахарная свекла, и т.д.) и различные цветковые культуры. Смотри, например, публикацию Ammirato и др. , 1984 г. Все белковые структуры, приведенные в изобретении, представлены в общепринятом порядке; аминогруппа у N-терминального звена, находится слева, и карбоксильная группа у С-терминального звена находится справа. Аналогично, аминокислотная номенклатура для естественно присутствующих в белке аминокислот следующая: аланин (ala; A), аспаргин (Asn; А); аспаргиновая кислота (Asp; D), аргинин (Arg; R), цистеин (Сys; С); глутаминовая кислота (Glu; E), глутамин (Gln; Q); глицин (Gly; G), гистидин (His; H), изолейцин (Ile; I), лейцин (Leu; L), лизин (Lys; K), метионин (Меt; M), фенилаланин (Рhe; F), пролин (Pro; P), серин (Ser; S), треонин (Thr; T); триптофан (Тrp; W), тирозин (Tyr; V) и валин (Val; V).
Выделение токсинового гена В.t.t.
Ген В.t.t. кодирующий токсиновый белок колеоптеранового типа был выделен следующим образом.
Выделение белковых кристаллов
В. t. tenebrionis был выращен в сoевом питательном бульоне Триптиказы (TSB) с целью выделения белковых кристаллов. При попытке выделения неповрежденных кристаллов из В.t.t.. было отмечено значительное различие между этими кристаллами и кристаллами лепидоптеранового типа. Хотя кристаллы лепидоптеранового типa обычно выделяются на градиентах, образуемых из Ренографина, Хипак (Hypague) или NaBr, установлено, что кристаллы B.t.t. растворяются в этих градиентах среды. Установлено, что кристаллы В.t.t. стойки в градиентах сахарозы, и градиенты сахарозы использовались для выделения кристаллов В.t.t.;
Выделение токсина В.t.t. из кристаллов
Очищенные кристаллы анализировались на их белковый состав методом SDS электрофореза на полиакриламидном геле. Результаты этих экспериментов показывают, что кристаллы B.t.t. содержат, по меньшей мере, два белковых компонента с мол.м. соответственно примерно 68-70 кДа и примерно 60 кДа. Относительные количества компонентов были различными при различных приготовлениях препаратов. Кроме того, можно утверждать, что более высокомолекулярный компонент может состоять более чем из одного белка. Бернардом (1986 г) были описаны белки с мол.м. примерно 68 кДа и 50 кDa как компоненты кристаллов B. t.t. Herrnstadt и др., 1986 г., сообщал о том, что кристаллы B.t. sah diego состоят из белка мол.м. примерно 64 кDa. В противоположность этому, штаммы В. t. t. типа лепидоптерана, такие как В.t. kurstaki обычно содержат более высокомолекулярный белок с мол.м. от 130 до 140 кDa. Этот результат показывает значительное различие в структуре между лепидоптерановыми и колеоптерановыми токсиновыми белками.
Были сделаны некоторые попытки очистки отдельных белковых компонентов кристаллов. Изоэлектрическое фокусирование не было успешным для данной цели, поскольку все белки осаждались. Анионообменная жидкостная хроматография высокого давления (НPLC) на колонке Моно Q не позволила разделить компоненты. Катионообменная хроматография на колонке Моно S в присутствии мочевины 4М позволила выделить пять пиков. Анализ этих пиков методом SDS гель электрофореза показал, что пик А соответствует лишь полосе более высокомолекулярного компонента от всего кристалла. Пик В богат данной полосой, соответствующей более высокомолекулярному компоненту, и включает незначительное число полос, соответствующих более низкомолекулярному компоненту. Пик С богат полосой, соответствующей более низкомолекулярному компоненту, и включает незначительное число полос, соответствующих более высокомолекулярному компоненту. Пики D и Е представляют собой смеси обеих полос. В большинстве препаратов полоса, соответствующая более высокомолекулярному компоненту, отвечающая пикам А и В, представляла собой полосу преобладающего белка в кристаллах. Для материала, разделенного методом НPLC пики А и В представляли собой наибольшую часть выделенного белка.
Были определены N-терминальные аминокислотные последовательности, соответствующие пикам А, В и С. Пики А и В, как установлено, имели одинаковую N-терминальую последовательность, в то время как последовательность С пика была другой. Определяемые последовательности были следующими:
Пик А и В
М t Asn Pro Asn A
t Asn Pro Asn A Arg Ser Glu His Asp Thr Ile Lys Thr T
Arg Ser Glu His Asp Thr Ile Lys Thr T
Пик С
M t X Pro X T
t X Pro X T r Arg Ala Leu A
r Arg Ala Leu A Asp Thr Ile Lys A
Asp Thr Ile Lys A
V Ile Glyn Lys, где Х является неопределяемой аминокислотой.
Ile Glyn Lys, где Х является неопределяемой аминокислотой.
Токсичность белков B.t.t. к насекомым
Некоторые препараты В. t.t. и белков B.t.t. испытывали на токсичность действия по отношению к различным насекомым, включая чешуекрылых и жесткокрылых. Никакой активности действия не было обнаружено по отношению к чешуекрылым насекомым (совка хлопковая, совка ипсилон, бабочка-бражник и совка капустная). Из числа жесткокрылых наблюдалось активное действие против колорадского жука (Leptinotarsa decemlineata) и долгоносика хлопкового (Anthonomus grandis). Более низкая активность действия наблюдалась в отношении блошки П-точечной Говарда (Diabrotica undecimpunctata howardi). Было обнаружено инсектицидное действие у сырых бактериальных культур, очищенных кристаллов, солюбилизированных кристаллов и выделенных пиков С. D. Е (соединенных), А и В.
Анализ на токсичность действия по отношению к колорадскому жуку осуществлялся на томатных листьях так, что вредные насекомые могли находиться на обработанных листьях в течение четырех дней. Анализ на токсичность действия против долгоносика хлопкового и блошки П-точечной Говарда осуществлялся путем ввода испытываемого материала в соответствующую питательную смесь.
Идентификация и клонирование токсинового гена B.t.t. в Е. соli и Pseudomonas
Используя информацию от N-терминальной белковой последовательности, были приготовлены пробы синтетической ДНК (фиг. 1), которые использовались при выделении клонов, содержащих токсиновый ген В.t.t. Эти пробы по концам метились [γ-32 p] АТР согласно Maniatis (1982 г.) В. thuringiensis разновидности tenebrionis выращивался в течение 6 ч при 37оС в среде Spizizen (Spizizen, 1958), дополненной 0,1% дрожжевым экстрактом и 0,1% глюкозой (SPY) для выделения всей ДНК. Вся ДНК была выделена из В.t.t. способом Kronstad (1983 г.) Клетки выращивались на агаровых пластинах Luria для выделения кристаллов В.t.t., используемых в исследовании на токсичность. Культуры Е. coli и Pseudomonas выращивались обычным образом в питательном бульоне Luria Broth (LB) с ампициллином (Ар, 200 мкг/мл), канамицином (Кm, 50 мкг/мл), или гетамицином (Gm, 15 мкг/мл), вводимых для отбора и сохранения плазмиды.
Выделение и обработка ДНК
Плазмида ДНК была экстрагирована из Е.coli и Pseudomonas клеток способом Birnboim и Doly (1979 г.), и большие количества этой плазмиды были очищены с использованием смолы NACS-52 (Научно-исследовательская лаборатория Bethesda) cогласно инструкциям изготовителя. Согласно инструкциям изготовителя использовались ограничительные эндонуклеазы, щелочная фосфатаза теленка и лигаза Т4 ДНК (Биолаборатория New England). Ограничительные продукты вываривания анализировались на 0,8% агарозном геле, подвергнутом электрофорезу в трис-ацетатном буферном растворе. Фрагменты ДНК для клонирования очищались от агарозы с использованием метода замораживания - оттаивания. Построение молекул рекомбинантной ДНК осуществлялось согласно Maniatis и др. , 1982 г. Трансформация в E.coli осуществлялась согласно Maniatis (1982 г.).
Клонирование токсинового гена B.t.t.
Осуществлялся анализ Southern (Southern, 1975), модифицированным методом с использованием высушенного геля (Conner и др., 1983 г.). Для обнаружения токсиновых клонов B.t.t. осуществлялась фильтр гибридизация колонии с использованием метода тетраметиламмонийхлорида (Wood и др., 1985 г.).
Анализ методом Southern общей ДНК B.t.t.. вываренной с BamHI и Hind III, позволил идентифицировать фрагмент 5,8 кb BamHI и 3,0 кв Hind III, который гибридизировался с синтезированной АI пробой. BamHI фрагменты ДНК B. t. t. (5,4-6,5 кb) были очищены от агарозных гелей и сшиты с обработанной щелочной фосфатазой рU C119, вываренной с BamHI. PU C119 приготавливалась путем выделения фрагмента 476 bp HgiAI/DraI бактериофага М13 и затупления концов данного фрагмента полимеразой Т4 ДНК (Биолаборатории New England). Затем этот фрагмент вставлялся в pU C19, который был выварен c Nde I и был наполнен ДНК полимеразой Кленова (Биолаборатории New England). Сшитая В.t. t. и рUC119 ДНК затем использовалась для трансформации клеток Е. coli IM101. После нескольких попыток были получены лишь стойкие колонии 150 Ар. Фрагменты Hind III B.t.t. ДНК (2,8-3,5 кb) также были клонированы в точку Hind III плазмиды рUC119, и было получено 1100 колоний. Все колонии просеивались путем гибридизации колонии с пробой AI (фиг. I). Одиннадцать клонов Hind III показали сильную гибридизацию, но ни одна из колоний Bam HI не показала какой-либо гибридизации. Затем колонии, идентифицированные гибридизацией с АI, были отсеяны с использованием синтезированной пробы А2 (фиг. 1), и две колонии показали гибридизацию со второй пробой. Ограничительные вываренные структуры двух колоний показали, что один и тот же фрагмент 3,0 кb Hind III находится в обоих колониях, но имеет противоположные ориентации. Эти клоны были обозначены рMON 5420 и рМОN 5421 (фиг. 3). Для подтверждения того, что эти клоны не содержат ген для токсинового белка B.t.t. была построена последовательность однониточной ДНК из обоих клонов с использованием вырожденных проб А1 и А2 в качестве затравок для ди-деоксипоследовательности (Sanger, 1977 г.). Анализ последовательности пробой А1 в качестве затравки обнаруживает открытый для отсчета скелет (ORF), последовательность которого идентична аминокислотам с 9 по 15 аминокислотной последовательности, определяемой для очищенных пиков А и В токсинового белка В.t.t.
Проба А2 продуцировала последовательность ДНК, которая начиналась за пределами конца заданной аминокислотной последовательности, но эта последовательность ДНК была идентична последовательности, продуцируемой АI. Эти результаты подтверждают, что был клонирован желаемый токсиновый ген B.t.t.
Гибридизация Southern с общей ДНК В.t.t. с использованием вырожденных проб на основе N-терминала пика С не позволила обнаружить специфические полосы, подтверждающие, что данная аминокислотная последовательность, определяемая для пика С, была неправильной или наиболее вероятно была получена из смеси двух или более белков, заключающих в себе пик С.
Анализ белков, продуцируемых в Е.coli.
Осуществлялось исследование кристаллических белков B.t.t. и рекомбинантных белков B. t. t. методом SDS электрофореза на полиакриламидном геле (Laemmli, 1970 г. ). Один миллилитр (Е.coli) центрифугировали, гранулы повторно суспензировали в 100 мкг буферного раствора пробы SDS и 10 мкл пробы подвергались электрофорезу на 7,5%-ных полиакриламидных гелях. Эти гели либо окрашивались красителем Комассьи синим, либо анализировались на перекрестную активность к антителам, вызываемым против очищенных токсиновых кристаллов В. t.t. Осуществлялся точечный анализ Вестерна с использованием антитела, образующего сопряженную связь с пероксидазой хрена обыкновенного (Towbin и др., 1984 г.). Высокомолекулярные сигнальные гены были получены из Bio Rad.
Следующим подтверждением было то, что продуцированный клонами токоин В. t.t. был получен путем точечного анализа Вестерна белков, продуцированных в Е. coli. Клетки E.Coli IMIOI, содержащие либо pUC119, либо pMON 5420, либо рMON 5421, были выращены в течение ночи в присутствии IPTG (0,1 ммоль), индуцируя промотор lac. Осуществлялся анализ двойных проб методом электрофореза на полиакриламидном геле с использованием в качестве контрольных образцов очищенных кристаллических белков B.t.t. Точечный анализ Вестерна одного геля обнаружил продуцирование двух перекрестно активных белков посредством Е. coli содержащим рМОN 5420 или рМОN 5421. Эти белки были идентичными по размеру основному и второстепенному белкам кристалла B.t.t. Молекулярная масса этих белков определяли путем сравнения со стандартными молекулярными весами на втором геле, окрашенном красителем Комассьи синим. Определено, что основной токсиновый белок имеет мол.м. 74 кDa и второстепенный токсиновый белок имеет мол.м. 68 кDa. Количество токсиновых белков B. t.t. продуцируемых посредством рМОN 5420, увеличивалось путем ввода IPТG, в то время как продуцирование токсиновых белков посредством рМОN 5421 оставалось неизменным.
Продуцирование токсина (токсинов) B.t.t. в Pseudomonas fluorescens.
Вектор хозяина широкого диапазона, рМОN 5432, был построен путем клонирования pMON 5420, вываренной с Bam HI, в точку Bam HI pMON 5420, как показано на фиг. 2. Этот вектор был затем спарен в Р. fluorescens 701E1 с целью анализа продуцирования токсина. Осуществлялись трехродительские спаривания в Pseudomonas fluorescens как описывалось ранее (Ditta и др., 1980 г.). Приготавливались пробы ночных культур, выращенных с IPTG и без IPTG, с целью проведения точечного анализа Вестерна и исследования токсичности к насекомым. Белки, продуцируемые Pseudomonas, идентичны по своему размеру продуцируемым E.coli белкам, и выражение белка увеличилось с вводом IPTG.
Анализ на токсичность к насекомым.
Осуществляли анализ на токсичность действия токсина на жесткокрылых насекомых, используя свежевыведенных колорадских жуков (Leptinotarsa decemlineata) на томатных листьях. Культуры E.coli и Pseudomonas были выращены в течение ночи в присутствии IPTG, затем они подвергались центрифугированию и повторному суспензированию при различных концентрациях в 10 ммоль MgSO4. Клетки разрушались под действием прозвучивания (три 15-секундные пульсирующие обработки на льду). Вводили Твин-20 (0,1%), и пробу наносили на томатный лист, помещенный в чашку Петри (9 см) с влажной фильтровальной бумагой. На каждый лист помещали десять личинок колорадского жука. По прошествии четырех дней рассчитывали процент смертности (процент выживших насекомых и контрольных пробах минус процент выживших насекомых в обработанных пробах поделенный на процент выживших насекомых в контрольных пробах), используя формулу Аббота (Abbott, 1925 г.). Анализы осуществлялись по два раза и данные суммировались. В качестве позитивной контрольной пробы использовался препарат B.t.t. кристалл/cпора.
Культуры Е.соli клонов pMON 5420 и рМОN 5421 анализировались на токсичность к жесткокрылым с использованием различных концентраций культур, выращенных с добавлением IPТG. Сравнение активностей рекомбинантного и дикого типа токсина B.t.t. показано в табл. 1. Данные результаты показывают, что рекомбинантные белки B. t. t. токсичны по отношению к колорадскому жуку. Двукратно концентрированная индуцированная IPTG культура pMON 5420 умерщвляла 100% насекомых, как и контрольная культура В.t.t. спора/кристалл. Эти токсичности показывают, что такой клонированный В.t.t. ген является геном, который кодирует токсиновый белок B.t.t.
Анализ с подачей насекомых показал, что продуцированные Pseudomonas токсичны по отношению к колорадскому жуку. Относительная токсичность культур Pseudomonas зависит от количества продуцируемого токсинового белка, которое определяется точечным анализом Вестерна, по сравнению с культурами E.coli.
Последовательность токсинового гена B.t.t.
Расположение и ориентацию гена B.t.t. в клонированном фрагменте определяли основываясь на следующей информации:
а) Последовательность ДНК была получена от однониточной модели pMON 5421.
b) Точка Pst I, идентифицированная путем анализа последовательности ДНК, около начала трансляции, размещалась на карте в рМОN 5420 и pMON 5421.
с) Были нанесены на карту некоторые другие ограничительные точки.
d) была построена делеция от точки Bgl II до точки Bam HI, которая отбирает 130 основных пар (bp), и были продуцированы оба белка полной длины цепи. Эта информация использовалась для построения карт рМОN 5420 и pMON 5421. Как показано на фиг. 4 кодирующая область токсина начинается с 500 bp от точки 5' Hind III, и 150 bp вверх от точки Pst I. Эта кодирующая зона завершается примерно 450 bp от точки 3' Hind III. Точка Bgl II составляет примерно 350 bp ниже от стоп-кодона.
Плазмиды.
В табл. 2 приведены плазмиды, образуемые для построения последовательности инсектицидного токсинового гена B.t.t. Родительские плазмиды, pMON 5420 и рМОN 5421 являются независимыми изолятами от фрагмента Hind III, клонированного в pU С119 в противоположной ориентации.
Приготовление однониточной модели для построения последовательности.
Осуществление операции как указано обеспечивает воспроизводимо высокие выходы однониточной модели для построения последовательности. Однониточная колония, содержащая pU C119 с данным фрагментом, последовательность которого должна быть построена, подвергалась штриховой разводке на L-агаре (10 г триптона, 5 г дрожжевого экстракта, 5 г NaCl и 15 г агара на литр), содержащем ампициллин (200 мкг на мл). Одиночная колония от этой модели инокулировалась в 3 мл L-питательного бульона (200 мкг на мл ампициллина) и инкубировалась при температуре 37оС в течение ночи с взбалтыванием. От этой культуры 50 мкл инокулировалось в 10 мл 2 х УТ (20 г триптона и 10 г дрожжевого экстракта на литр) с 200 мкг ампициллина на мл в 150-миллилитровой колбе с боковой рукояткой и инкубировалось при температуре 37оС с взбалтыванием. После 2-3 ч (показатель Клетта 50) вводили 100 мкл М13К07 (вспомогательный фаг), выращенный в Е.соli IМ101 для индуцирования культуры. Колбу встряхивали в течение 1 ч, после чего вводили 20 мл 2 Х УТ, доводя конечную концентрацию канамицина до 70 мкг на мл ампициллина до 200 мкг на мл. Культуры взбалтывались в течение 16-18 ч при температуре 37оС. Как было определено, в общей сложности 3 м л индуцируемой в течение ночи культуры было достаточно для извлечения требуемого количества модели для четырех экспериментов построения последовательности. Эти 3 мл вытягивались в 1,5 мл трубках эппендорфа в течение 1 мин, декантировались и фильтровались через 0,2 мкм Gelman Sciences Acrodisc. Данный этап обеспечивал удаление клеточных остатков и неповрежденных Е.сoli. После осаждения в полиэтиленгликоле (20% полиэтиленгликоля, 2,5 мол. NaCl, 500 мкл на 2 мл лизата) при комнатной температуре в течение 10 мин осуществлялось центрифугирование в течение 10 мин. Поверхностный слой удаляли, после чего осуществлялось быстрое вытягивание (15 с) и удаление остаточного полиэтиленгликоля. Любой оставшийся полиэтиленгликоль проходил через выделенную модель и оказывал нежелательное влияние на реакции построения последовательности ДНК. Полученные гранулы суспензировались в 100 мкл ТЕ (10 ммоль трис- 1 ммоль этилендиамин тетрауксусной кислоты, рН 8,0) соединялись и тщательно перемешивались с 200 мкл буферного раствора фенола (буферный раствор приготавливался путем получения равновесной смеси с равным объемом 1 моль) трис-HCl, рН 8,0 затем 0,1 моль трис-HCl. рН 8,0, и затем с равным объемом ТЕ). После инкубации при температуре 55оС в течение 10 мин добавляли равный объем (200 мкл) смеси фенол-хлороформ (1:1), подвергали вихревому перемешиванию и центрифугировали в течение 2 мин. Верхний слой удаляли, экстрагировали 200 мкл хлороформа, центрифугировали, и водную фазу удаляли. Однониточная молекулярная модель осаждалась посредством 25 мкл 3 моль ацетата натрия (рН 5,2) и 600 мкл 95%-ного этанола, инкубировалась на сухом льду в течение 5 мин и центрифугировалась в течение 10 мин. Осадок снова суспензировали в 25 мкл Н2О и 2 мкл пробы анализировали на агарозном геле на точный размер, относительную концентрацию и загрязнение ДНК.
Образующие последовательность реагенты и условия
Процедуры построения последовательности ДНК описаны подробно в справочном пособии фирмы Amercham Corpiration. Реагенты (нуклеотиды, затравка, буферный раствор, вытягивающий раствор и полимераза Кленова) получены из системы Amercham М13 (каталог 4502). Используемые для построения последовательности смеси в системе Amersham подготавливались для эффективного получения последовательности обогащенного А - Т гена В.t.t. Вместо рекомендуемой смеси dNTH c dd NTP в соотношении 1:1, наиболее подходящими были следующие соотношения компонентов: 40 мкл dATP: 10 мкл ddATP, 35 мкл dТТР; 15 мкл ddТТР, 15 мкл dGTP: 35 мкл ddGTP, и 10 мкл dCTP: 40 мкл ddСТР. В реакциях построения последовательности использовалась радиоактивная сера [(α-35S] dATP) (Каталог Amercham # 1304). Образующие последовательность гели (приготовленные как описано в справочном пособии Amersham) обрабатывались в аппаратуре Hoeffer "Роker Face" мощностью 70 Вт (1200-1400 В), что обеспечивало очень высокое разрешение. Более высокое напряжение приводило к нечетким полосам.
Последовательность токсинового гена В.t.t.
Выделенные плазмиды, pMON 5420 и рМON 5421, содержали фрагмент 3,0 Hind III противоположной ориентации (см. фиг. 3). Основной белок кристалла B.t. t. , который использовался как основа для олигонуклеотидных проб, имел мол. м. примерно 73-76 кДа, соответствующий примерно 2 кb ДНК. Начальная последовательность от А1 и А2 затравок (синтезированные олигонуклеотиды, основанные на аминокислотной последовательности пика А; (см. табл. 3) подтверждает, что данная последовательность ДНК соответствует предполагаемой аминокислотной последовательности.
Точка Pst I располагалась в начальной последовательности, которая использовалась для идентификации положения и вероятностной ориентации генa B.t. t. в рМON 5420 и рМОN 5421 (см. фиг. 3 и 4). Построение карты ограничительных точек с числом ферментов (НраI, Xba I, Nde I, Eco RV и Bgl II) и многочисленными одиночными очистками, оставшимися в части рU C119 как плазмиды рМОN 5420, так и плазмиды pMON 5421, дает возможность получить последовательность с использованием универсальной образующей последовательность затравки. Были образованы делеции как в рМОN 5420, так и в рМОN 5421, несущие гомологичную область универсальной затравки в непосредственном соседстве с внутренними зонам гена. На участках, где нелегко образуются последовательность за счет генерируемых делеций, в качестве затравок использовались синтезированные олигонуклеотиды, соответствующие областям с построенной последовательностью в кодирующей последовательности (табл. 3) для получения продолжений областей с построенными последовательностями. Области с построенной последовательностью (координаты последовательности, табл. 4) и направление последовательности приведены на фиг. 4.
Компьютерный анализ инсектицидного токсинового гена B.t.t.
Из рМON 5420 и рМON 5421 получали суммарно 2615 основных пар последовательности. Компьютерный анализ последовательности показал один скелет открытой считки от основной пары 205 до 2136. Как показано на фиг. 5-14, инсектицидный токсиновый ген В.t.t. составляет 1932 основные пары, кодирующие белок, заключающий 644 аминокислот мол.м. 73,091 Дальтон. Этот белок имеет чистый заряд -17 и содержание в нем G-C составляет 34%.
Связь между токсиновыми генами и белками колеоптеранового жесткокрылых и лепидоптеранового (чешуекрылых) типов.
Несмотря на то, что токсины колеоптеранового типа и токсины лепидоптеранового типа получены из Bacillus thuringiensis, существуют значительные различия между токсиновыми генами и токсиновыми белками этих двух типов. Выделенные из Bacillus thuringiensis оба типа токсинов обнаруживаются в параспоровых кристаллах; однако, как указывалось, характеристики растворимости этих кристаллов явно различны. Кроме того, размеры токсиновых белков, находящихся в солюбилизированных кристаллах, совершенно отличны. Токсиновые белки лепидоптеранового типа обычно имеют мол.м. порядка 130 кDa, в то время как токсиновые белки колеоптеранового типа имеют размер примерно 70 кDa.
Выделение и анализ последовательности ДНК токсинового гена колеоптеранового типа, происходящего от B.t. tenebrionis, позволяет предсказать аминокислотную последовательность токсинового белка (фиг. 5-14). Как нуклеотидная последовательность, так и образованная аминокислотная последовательность токсинового гена колеоптеранового типа сопоставились с нуклеотидной и аминокислотной последовательностью типичного токсина лепидоптеранового типа.
Это сопоставление осуществлялось с использованием компьютерной программы BESTFIT, Devereux и др. (1984) в которой используется алгоритм Смита (Smith) и Ватермана (Waterman) 1981 г. Программа BESTFIT показывает максимальное выравнивание как нуклеотидных, так и аминокислотных последовательностей. BESTFIT позволяет рассчитать два параметра, качества и сопоставительного коэффициента, которые могут быть использованы как показатели выравнивания при проведении сопоставления различных выравниваний. Сопоставительный коэффициент изменяется в пределах от 0 до 1,0. Более высокое значение коэффициента показывает лучшее выравнивание (лучшее сходство) между двумя последовательностями.
Выравнивание BESTFIT показывает, что оба типа токсиновых генов связаны как с нуклеотидной последовательностью, так и с аминокислотной последовательностью. Однако, выравнивание также показывает, что обе последовательности явно различны и имеют много областей, не соответствующих друг другу как в отношении нуклеотидной, так и в отношении аминокислотной последовательности. Так, например, сопоставительный коэффициент, определяющий связь между двумя аминокислотными последовательностями, составляет лишь 0,22. В случае нуклеотидной последовательности максимальное выравнивание достигается лишь за счет ввода многих гэпов в обе последовательности, и сопоставительный коэффициент составляет лишь 0,072.
Существует много примеров последовательности токсиновых генов лепидоптеранового типа; аналогичное сопоставление этих генов показало, что гены от B. t. kurstaki HD-1, описанные Schnepf и др., 1985 г., и гены от B.t. kurstaki HD-73, описанные Adang и др., 1985 г., представляют собой два наиболее отличные друг от друга токсиновые гена лепидоптеранового типа (чешуекрылых). При сопоставлении с коэффициентами, рассчитанными, как указано, для выравниваня гена колеоптеранового типа и лепидоптеранового типа, сопоставительный коэффициент для аминокислотной последовательности двух наиболее отличных друг от друга белков лепидоптеранового типа составляет 0,811, и сопоставительный коэффициент для этих двух генов лепидоптеранового типа для нуклеотидиной последовательности составляет 0,755. Это показывает, что хотя гены колеоптеранового типа и лепидоптеранового типа могут быть сопоставимы друг с другом, они явно различны в отношении как нуклеотидной, так и аминокислотной последовательности.
Высокая степень продуцирования рекомбинантного B.t.t. токсина в Е.соli
Для облегчения очистки больших количеств рекомбинантного B.t.t. токсина, необходимо, чтобы ген B.t.t. клонировался в высоковыраженные векторы E.coli. Точечно направленный мутагенез был использован для вставки ограничительной точки Nco I и рМОN 5420 в кодоне ATG в начале скелета открытой очистки.
Точечно направленный мутагенез
Точечно направленный мутагенез для вставки новых ограничительных точек осуществлялся способом Kunkel, 1985 г.
Плазмида рМОN 5420 вставлялась путем трансформации в BW 313 штамм E.coli, который содержал мутации dut- и ung- с целью ввода дедоксиуридина в ДНК. Одиночная трансформированная колония выращивалась в течение ночи в среде 2Х УТ, содержащей 100 мкг/мл ампициллина и 0,25 мкг/мл уридина. 0,5-миллилитровая аликвота этой культуры вводилась в 10 мл той же среды и инкубировалась в течение 1 ч при температуре 37оС с тщательным взбалтыванием до достижения плотности 0,23 (А600). Для того, чтобы вызвать образование содержащих одиночную нить фаговых частиц, был введен вспомогательный фаг M13К07 с множественностью примерно 10, и продолжалась инкубация в течение 1 ч до достижения плотности 0,4 (А6 00). Культура разбавлялась путем ввода 30 мл указанной среды, и вводился канамицин до конечной концентрации 70 мкг/мл. Инкубация продолжалась в течение 15 ч, после чего клетки удалялись путем центрифугирования. Фаговые частицы осаждались из 25 мл всплывшего на поверхность слоя путем добавления 5 мл 20%-ного полиэтиленгликоля (2,5 моль NaCl)0, 50 мкг/мл рибонуклеазы А с последующей инкубацией на льду в течение 15 мин. Фаг извлекали путем центрифугирования и растворяли в 0,8 мл буферного раствора ТЕ. ДНК выделяли из данных частиц путем трех экстракций 0,8 мл раствора фенол - хлороформ - изоамиловый спирт в соотношении 25:24:1 с последующим осаждением этанолом. Гранула ДНК растворялась в 100 мкл воды до конечной концентрации примерно 1 мг/мл (установлено путем электрофореза на агарозном геле).
Синтезированные олигонуклеотидные затравки для мутации суспензировались в воде до концентрации примерно 10 пикомоль/мкл. Олигонуклеотиды фосфорилировались с использованием Т4 полинуклеотидокиназы в реакционной смеси, содержащей 50 микомоль олигонуклеотида, 1 ммоль, АТР, 25 ммоль Трис-HCl, рН 8, 10 ммоль MgCl2, 0,2 ммоль, спермидина-НСl, 1 ммоль ДТТ и 2 единицы фермента. Реакционная смесь инкубировалась при температуре 37оС в течение 30 мин, и затем нагревалась при 70о С в течение 5 мин. Фосфорилированная затравка аннилировалась содержащим деоксуридин фагом ДНК путем смешивания примерно 1 пикомоль фага ДНК (2 мкг) с 10 ммоль затравки в реакционной среде, содержащей 6,6 ммол. Трс-НСl 6,6 ммоль MgCl2, 6,6 ммоль NaCl и 5 ммоль ДТТ. Смесь нагревалась до 70оС в течение 7 мин и затем медленно охлаждалась до комнатной температуры. Анилированная затравка - модельная молекула использовалась в качестве субстрата для синтеза двухниточной замкнутой кольцевой молекулы ДНК путем ввода каждой из: dNTP до 0,5 ммоль, АТР до 0,5 ммоль, 5 единиц ДНК полимеразы фрагмента Кленова и 400 единиц Т4 ДНК лигазы (Биолаборатория New England). Реакция осуществлялась в том же буферном растворе соли, что и аннеляция, при температуре 15оС в течение примерно 15 ч. В этот момент вводили дополнительно 400 единиц лигазы, и инкубация продолжалась в течение 2 ч.
Половина данной реакционной смеси использовалась для трансформации 0,15 мл обработанных CaCl2 клеток IM101, клетки распределялись на пластинах LB, содержащих 100 мкг/мл ампициллина. На каждую реакцию мутагенеза извлекалось от 30 до нескольких сотен колоний. Одиночные колонии выращивались в течение ночи в LВ, содержащей ампициллин, и плазмидные минипрепараты приготовлялись щелочным SDS методом. Плазмиды анализировались на присутствие новой ограничительной точки, в присутствии этой точки подтверждалось анализом указанной последовательности.
Плазмида, содержащая точку Nco I (pMON 9759) в начале инсектицидного токсидного гена В.t.t., получалась путем точечно-специфического мутагенеза. Используемая затравка GATTGTTCGGATCCATGGTTCTTCCTCCCT, желаемая точка NcoI
Образование точки NcoI в N-терминале изменяло вторую аминокислоту от аспарагина до аспарагиновой кислоты. Это изменение не оказывало нежелательного действия на токсичность к насекомым. Точки Bam HI и Sty I также образовывались как следствие ввода данной точки NcoI. Плазмида, содержащая точку NcoI, обозначена для рМОN 9759. Фрагмент 2,5 kb Nco I-Hind III, содержащий кодирующий токсин сегмент от рМОN 9759, затем клонировался в рМОN 5634, вываренную с NcoI - Hind III, продуцируя рМОN 5436. Как показано на фиг. 27, рМОN 5634 представляет собой плазмиду на основе PBR 327, которая также содержит фаговый f1 источник репликации. Этот вектор содержит синтезированный rec A промотор, который индуцирован налидиксиновой кислотой. Присутствует также основной ген 10 от фага Т7. Патентная заявка США N 005821, 04.02.1987), который увеличивает выражение в Е.coli. Синтезированная связующая молекула с множеством клонирующих точек была введена для вставки генов ниже промотора и последовательности основного гена 10.
Для индуцирования промотора rec A выращенные в течение ночи культуры разбавлялись в соотношении 1:50 в минимальной среде М9 (Miller, 1972 г) с вводимыми 0,2% казаминовой кислоты и 0,25% глюкозы. Налидиксиновая кислота, 150 ед. Клетта, вводилась в 50 мкг/мл, и клетки после индуцирования собирались в течение 3 ч. Уровень токсина B.t.t., продуцируемого рМОN 5436, индуцированной налидиксиновой кислотой, сопоставлялся с рМОN 5420, индуцированной 1РТG, путем анализа SDS-РАGE. Гель, окрашенный Коммасье синим, не обнаружил B. t.t. продуцируемого рМОN 5420, в то время как уровень B.t.t.. продуцируемого посредством рМОN 5436, составлял примерно 5% от общего белка. Такой принцип использовался для извлечения больших количеств рекомбинантных токсиновых белков B.t.t. для изучения степеней токсичности, специфичности насекомых и принципа действия.
Характеризация токсина B.t.t.
Идентификация числа и происхождения белков B.t.t.
B. t. разновидности tenebrionis продуцирует ряд токсиновых белков колеоптеранового типа (жесткокрылых), присутствующих в белковых кристаллах, которые продуцируются сопутствующим образом со споруляцией (см. фиг. 15). Эти белковые кристаллы вводятся в данную среду по мере автолиза клеток в процессе или после споруляции. Для определения числа белковых токсинов, продуцируемых В. t. разновидности tenebrionis, 500 мл культур этого организма выращивалось в 2-литовых колбах в 15% TSB среде в 100 ммоль буферном растворе 2-(N-морфолино)этансульфокислоты (МЕS), рН 7,0, при температуре 30оС в течение 7 дней. В этот момент культуры спорулировались и клетки лизировались. Белковые кристаллы и споры собирались путем центрифугирования при 20000 х g (ускорение силы тяжести) в течение 20 мин при 4оС. Гранулы промывались три раза избыточным количеством воды, после чего промывались три раза 2 моль NaCl. Полученная гранула хранилась при температуре 4оС в воде плюс 0,02% азида натрия. Токсиновый белок B.t.t. солюбилизировался из кристаллов путем суспензирования гранулы в 100 ммоль буферного раствора карбоната натрия, рН 10, и перемешивания этой суспензии в течение 2 ч при комнатной температуре. После центрифугирования при 20 000 x g в течение 20 мин с целью удаления несолюбилизированого материала, всплывший на поверхность слой фильтровали через 0,2 мкм фильтр с целью удаления любых оставшихся спор. Токсиновый белок B.t.t., приготовленный указанным образом, как и кристаллы, солюбилизированные в 125 ммоль Трис-НСl, 4% SDS, 20% глицерина и 10% -2-меркаптоэтанола, рН - 6,8 (буферный раствор пробы SDS используется для приготовления образцов для SDS - РАGE анализа) включал четыре основные и различные белка, как определено SDS-РАGE анализом. Пять единственных в своем роде продуктов были идентифицированы путем анализа на N-терминальной аминокислоты. Для определения того, все ли пять этих белков были получены из одного и того же гена, а также для определения того, два или более чем два гена требуется для их синтеза, определяли N-терминальную аминокислотную последовательность каждого из этих белков, используя автоматизированный метод химического разложения Эдмана.
Был использован (Hunkapiller и др., 1983 г) газофазный секвенсор модели 470А.
(Foster City, Калифорния). Соответствующие РТН-аминокислотные производные были идентифицированы методом жидкостной хроматографии высокого давления RP - HPLC, в той же системе, с использованием анализатора Applied Biosystems, Inc. Модели 120А РТН, снабженного колонкой Brownlee РТН-С18 внутренним диаметром 2,1 мм. Определение N-терминальной аминокислотной последовательности каждого белка позволяет выявить, все ли эти белки получены от токсинового гена B. t.t., как описано выше. Построение последовательности этих белков заключалось в построении последовательности токсиновых белков B.t.t. соответствующих полосам 1 и 3 (смотри фиг. 15) из клона E.coli IM101 (pMON 5436), полос 2, 3 и 4, методом элктроэлюирования белков, продуцируемых B.t. разновидности tenebrionis из гелей SDS-PAGE. Последовательность 1 и 3 B.t. t. определяли с использованием очищенных от IM101 (pMON 5436). IM101 (pMON 5436), так же как и другие построения E.сoli (pMON 5450, 5456, и 5460, смотри ниже) продуцирует B.t.t. в форме нерастворенных преломляющих тел после того как культуры индуцируются для высокого уровня выражения. Построения Е. coli были выращены в модифицированной среде М9 при температуре 37оС. Культура, выращенная в течение ночи, использовалась для инокуляции 400 мл модифицированной среды М9 в 2,4-литровых колбах Фернбах до начальной плотности, составляющей 10 единиц Клетта.
Налидиксиновая кислота, в 1,1 н. NaOH, вводилась в данные культуры при 100 ед. Клетта до достижения конечной концентрации 50 мкг/мл, индуцируя выражение токсинового белка B.t.t. После дополнительных 4 ч инкубации культуры собирались путем центрифугирования при 20 000 x g в течение 20 мин при температуре 4оС. Клеточные гранулы суспензировались в воде до плотности эквивалентности 5000 ед. Клетта на мл, и прозвучивались в ледяной бане с использованием ультразвуковой системы с нагревом (Heat Systems Ultrasonics) с энергией 9, с рабочим циклом 50% в течение в общей сложности 5 мин. Подвергнутый прозвучиванию препарат подвергался центрифугированию в течение 20 мин при 20 000 x g при 4оС. Гранулы, содержащие преломляющие тела и клеточные осколки, двукратно промывались холодной водой и суспензировались при эквивалентах 10 000 е. Клетта на мл в воде +25% сульфолана. После перемешивания при комнатной температуре в течение 2 ч солюбилизированные препараты преломляющего тела снова центрифугировались при 20 000 х g при температуре 4оС для удаления несолюбилизированных материалов. В поверхностный слой центрифугирования вводили Трис-HCl до конечной концентрации 50, ммоль, рН 7,6. Полосы 1 и 3 В.t.t. совместно очищались в ионообменной колонке на HR5/5 Mono Q с использованием от 75 ммоль до 200 ммоль NaCl градиента в 50 ммоль трис-HCl, 25% сульфолана, рН 7,6. Фракции, содержащие полосы 1 и 3 B. t.t. идентифицировались путем 9% SDS-PAGE анализа, объединялись, диализировались в 100 ммоль буферного раствора карбоната натрия, рН 10, и концентрировались в концентратах Америкон Центрикон. Токсиновый белок B.t.t., соответствующий полосе 3, очищался от IM101 (pMON 5456) аналогичным образом.
Полосы, соответствующие лишь 2, и соединенные полосы 3, 3' и 4 (см. фиг. 15) подвергались электроэлюированию из 7% SDS-PAGE мыльных гелей, которые пропускались вместе с 48 мкл кристаллов B.t.t., солюбилизированных в буферном растворе 100 ммоль карбоната натрия, 20 ммоль дитиостреитола (ДТТ), рН 10. Гели окрашивались в течение 10 мин красителем Комассье синим R250 и обесцвечивались в 50% метаноле, 10% кислоте в течение 20 мин. Соответствующие полосы извлекались лезвием бритвы и осуществлялось электроэлюирование белка B.t.t. Зная аминокислотную последовательность, выведенную из последовательности ДНК токсинового гена B.t.t., клонированного в E.coli, идентифицировали все пять N-терминалов этих единственных в своем роде белков (фиг. 16).
Белки, соответствующие полосе 1 и 3, образованы в результате двух случаев независимого трансляционного инициирования, которые начинаются у метионина в положениях 1 и 48 (фиг. 15-16 соответственно. Белки, соответствующие полосам 2, 3 и 4 B.t.t., наблюдаемые лишь в B.t. разновидности tenebrionis, но не в построениях E.coli, по всей вероятности образуются за счет протеолитического расщепления любой из полос 1 или 3. Эти результаты говорят о том, что все пять белков происходят от одного и того же гена.
Очистка полос 1 и 3 B.t.t. для испытания на токсичность к насекомым.
Белки B. t.t., продуцированные в Е.соli, соответствующие полосам 3 и 1 плюс 3, которые солюбилизировались в 25% сульфолане и очищались хроматографическим методом на Mono Q, подвергались анализу на N-терминальную аминокислотную последовательность, который не обнаружил токсического действия против колорадского жука. В последующих экспериментах было показано, что сульфолан как таковой инактивирует B.t.t. В связи с этим был разработан другой способ очистки, и осуществлялось сопоставление относительных инсектицидных токсичностей полос 1 и 3 B.t.t., продуцированных в E.coli, c инсектицидными токсичностями B. t. t. солюбилизированными от естественных кристаллов B.t. разновидности tenebrionis. Культуры выращивались, индуцировались, извлекались и выделялись преломляющие тела, как описано выше. Различные B. t.t. белки солюбилизировались из преломляющих тел с использованием 100 ммоль карбоната натрия, рН 10. Солюбилизированный токсин B.t.t., концентрированный с использованием перемешанных клеток Амикон с мембранами УМ-10, очищался в колонке гель фильтрации FPLC, Pharmacia Superose-12, в которой происходило отделение полос 1 и 3 B.t.t. от других загрязняющих белков. Соответствующие фракции, определяемые исходя из SDS-PAGE анализа, соединялись, концентрировались и использовались для экспериментов на токсичность действия против насекомых с использованием колорадского жука. Белки, соответствующие полосe 1 (pMON 5436), полоса 1 (рМОN 5460) и полосе 3 (рМON 5456) составляли по чистоте более 90% на основе анализа SDS-РАGE.
Полоса 1, продуцированная pMON 5460, заключала изолейцин при аминокислоте 48, вместо метионина (указано далее).
Для получения естественного белкового токсина из B.t. разновидности tenebrionis для сопоставления токсичности, естественные кристаллы извлекались и очищались путем центрифугирования с использованием сахарозного градиента, как описано выше. Кристаллы солюбилизировались в 100 ммоль карбоната натрия, 20 ммоль ДТТ, рН 10, и использовались для испытаний на токсичность к насекомым.
Все препараты токсинового белка B.t.t. и контрольные препараты для испытания на насекомых содержали 0,3% Твина 20, поверхностно-активного вещества, которое повышает способность данных растворов связываться с томатными листьями. Эксперименты на токсичность к насекомым осуществлялись путем нанесения на верхнюю и нижнюю поверхности одиночных томатных листьев 3-4 недельного возраста буферных растворов, содержащих белки B.t.t. с указанными концентрациями белка. После того как растворы были высушены воздухом на поверхности томатных листьев, отдельный лист и 10 колорадских жуков помещали в чашку Петри и инкубировали при 22оС в течение 4 дней. Определяли число умерших насекомых и получали данные токсичности, выраженные как процент смертности (% СМ), согласно формуле Абботта. Все эксперименты осуществлялись по два раза и все, кроме случая полосы 1 B.t.t. от рМОN 5460, повторялись в разные дни. Результаты данных испытаний показаны в табл. 5.
Относительная токсичность очищенных белков от различных типов E.coli сопоставлялась с токсичностью естественных солюбилизированных B.t.t. кристаллов. Полоса I (pMON 5436) и полоса 3 (pMON 5456) были очищены как описано. Полоса I (pMON 5460) была очищена посредством хроматографии с использованием гель-фильтрации. Естественные B.t.t. кристаллы были солюбилизированы в 100 ммоль Na2CO3, рН 10.
Количество токсина B.t.t., необходимое для умерщвления 50% колорадских жуков, были практически идентичны полосе I B.t.t., выделенной из рМON 5436 и рМОN 5460, и полосе 3 B.t.t., выделенной из рМОN 5456 (табл. 5). Аналогичным образом, все эти очищенные препараты B.t.t. от E.coli продемонстрировали токсичность в основном идентичную токсичности, наблюдаемой для солюбилизированного карбонатом натрия естественного токсина из B.t. разновидности tenebrionis.
Определение токсических фрагментов токсиновых белков B.t.t.
Несколько групп ученых (Schnepf и др., 1985 г., Hofte и др., 1986, Wabiko и др., 1986 г.) уже сообщали о том, что С-терминальные обрывы цепи токсинов лепидоптеранового типа не снижают токсичность (обрыв цепи от аминокислоты 1155 до аминокислоты 607 не приводит в результате к потере токсичности). Ввиду этого, для токсичности не требуется С-терминальная половина белка. Другие группы ученых сообщали о том, что токсиновые гены лепидоптеранового типа, которые включают С-терминальные делеции, в более высокой степени выражены в трансформированных растениях. Имеются также сообщения о том, что для сохранения токсичности могут быть сделаны лишь небольшие обрывы цепи в N-терминальной части (Schnepf и др., 1985 г, и Hofte и др., 1986 г.). В противоположность этим сведениям, сейчас было установлено, что токсин B. t. t. колеоптрептанового типа (жесткокрылых насекомых) значительно отличен по своим свойствам. Так, С-терминальная часть является критически важной для токсичности, и следовательно практически допускается исключение обрыва цепи. Однако, могут быть осуществлены N-терминальные делеции, и токсичность сохраняется. Эти различия были выявлены с использованием описанных ниже построений:
Построение рМОN 5426 (Делеция Bgl II/Bam H1.
рМОN 5420 вываривался с Bgl II и Bam HI, сшитыми и трансформированными в IM101 для создания рМОN 5426. Эта делеция была построена для подтверждения того, что точка Bgl II не находился в пределах кодирующей зоны токсинового гена B.t.t.
Построение рМОN 5438 (НраI, С-терминальная делеция 463 bp).
рМОN 5420 вываривался с НраI и сшивался с синтезированной терминаторной связывающей молекулой. Эта связывающая молекула содержит несущественные кодоны в каждом каркасе счистки и выступающей части Bgl II 5'.
5' -TAGTAGGTAGCTAGCCA-3
3 '-ATCATCCATGATCGGTCAG-5'
Эта сшивка вываривалась с Bgl II для удаления многозвенных связывающих вставок и затем повторно сшивалась. Данная сшивка трансформировалась в IM101 и выделялся рМОN 5430. Для генерации точки Nco I в начале усеченного гена, фрагмент 2,35 кb Pst I от рМОN 9759 заменялся фрагментом 1,47 кb Pst I от рМОN 5430, это новое построение было обозначено pMON 5434. Фрагмент 1,57 kb Nco I/Hind III от рМОN 5434 клонирован в высоковыраженный вектор рМОN 5634 Е.coli, и в результате создается pMON 5438.
Построение рМОN 5441 (Eco RV, C-терминальная делеция 327 bp).
рМОN 5420 был выварен с Есо RV и сшит с синтезированной терминаторной связующей молекулой. Эта сшивка вываривалась с Bgl II для удаления многозвенных связывающих вставок и затем снова сшивалась. Сшивка трансформировалась в IM101 и был выделен рМОN 5431. Для генерации точки Nco I в начале усеченного гена, фрагмент 2,32 kb Pst I от рМОN 9759 был заменен фрагментом 1,61 lb Pst от рМОN 5431, это новое построение было обозначено рМОN 5435. Фрагмент 1,71 кb Nco I/Hind III от рМОN 5435 был клонирован в высоковыраженный вектор рМОN 5433 E.coli и был образован pMON 5441.
Построение рМОN 5449 (Ва131, С-терминальная делеция 190 bp)
Вываренный с Blg II, pMON 9759 обрабатывался нуклеазой Ва131 в течение 5 мин согласно инструкциям изготовителя. ДНК подвергалась электрофорезу в 0,8% агарозном геле и очищалась от агарозы способом замораживания - оттаивания. Синтезированная терминаторная связующая молекула затем сшивалась с очищенной ДНК, выделялся рМОN 5442. Фрагмент Nco I/Bgl II от pMON 9759 был заменен усеченным генным фрагментом от рМОN 5442, в результате был создан рМОN 5445. Фрагмент Nco I/Hind III от рМОN 5445 был клонирован в высоковыраженный вектор рMO 5634 E.coli и был образован pMON 5449. Путем анализа ДНК последовательности была определена концевая точка в созданной Ва131 делеции.
Построение рМОN 5448 (Хmn I, С-терминальная делеция 16 bp).
рМОN 5436 был выварен с Хmn I и сшит с синтезированной терминаторной связывающей молекулой. Эта сшивка затем вываривалась с Nco I и Bgl II, фрагмент 1,92 kb Nco I/Bgl II, содержащий усеченный ген, клонировался в рМОN 9759, вываренный с Nco I и Bgl II, вытесняя полной длины ген и создавая рМОN 5446. Фрагмент Nco I/Hind III от рМОN 5446 был клонирован в высоковыраженный вектор рМОN 5634 E.coli, в результате чего был образован рМОN 5448.
Построение рМОN 5450 (дополненные Nco I концы, удаление первого ATG из токсина ORF)
рМОN 5436 вываривался с Nco I, концы дополнялись с использованием ДНК полимеразы фрагмент Кленова, сшивались и трансформировались в IM101, образуя рМОN 5450. Эта плазмида выражает лишь белок полосы 3.
Построение рМОN 5452 (N-терминал, делеция 224 bp)
Ген B.t.t. содержал две точки Sty I (227 и 1587), третья точка вводилась путем мутагенеза, образуя точку Nco I в рМОN 9759. Последующие эксперименты осуществлялись с целью делеции 5' B.t.t. ДНК до основной пары 227, рМОN 5434 (производное делеции НраI, указанное выше) вываривался с Sty I, концы дополнились ДНК полимеразой Кленова, сшивались и трансформировались в IM101, с выделением рМОN 5444. Такая процедура приводила к разрушению точек расщепления как NcoI, так и StyI. Такая процедура создавала слияние с первым метионином (аминокислотой I) и лейцином (аминокислота 77). С-терминал данного гена вводили путем клонирования фрагмента 1,9 kb Nde I/Kpn I от pMON 9759 в рМОN 5444, и в результате был образован рМОN 5452. Построение рМОN 5456 (Мутант с полосой 3, N-терминальная делеция 140 bp)
Точка NcoI вставлена в рМОN 5420 и АТG для полосы 3 путем точечно направленного мутагенеза, как описано выше, с использованием затравки.
Затравка мутагенеза - BT L OOP
CGTATTATATCTGCATCCATGGTTCTTCCTCCCT
с образованием рМON 5455. Данный мутагенез приводил также к делеции находящейся выше последовательности, которая кодирует N-терминал 48 аминокислот полосы I. Фрагмент Nco I/Hind III от рМОN 5455 был клонирован в высоковыраженный вектор рМОN 5634 E.соli, образуя pMON 5456. Эта плазмида выражает лишь полосу 3. Генерация точки Nco I изменяет вторую аминокислоту с тионина на аспаргиновую кислоту.
Построение рМОN 5460 (Ген с полосой I мутанта с МЕТ48, измененный на ILE.
Кодон для метионина в положении 48 в рМОN 9759 был изменен как кодон для изолейцина путем точечного направленного мутагенеза, как описано выше, с использованием затравки.
Затравка мутагенеза - ВТTМЕТ
АТТАТТАТСТGCAGTTATTCTTAAAAACTCTTAT с образованием рМОN 54578. Фрагмент Nco I/Hind III от рМОN 5458 был клонирован в высоковыраженный вектор рМОN 5634 Е. соli с образованием рМОN 5460. В результате удаления АТG кодона, который инициирует трансляцию белка полосы 3, рМОN 5460 продуцирует только белок полосы 1 с изолейциновым остатком в положении 48.
Построение рМОN 5467 (Мутант полосы 5. N-терминальная делеция 293 bp)
Точка Nco I вставлялась в рМОN 5420, создавая N-терминальную делецию 28 аминокислот путем точечно направленного мутагенеза с использованием затравки.
Затравка мутагенеза
ТСАСТТGGCCAAATTGCCATGGTATTTAAAAAGTTTGT
с образованием рМОN 5466. Метионин и аланин также вставлялись путем мутагенеза. Фрагмент Nco I/Hind III от рМОN 5466 был клонирован в высоковыраженный вектор рМОN 5634 Е.соli c образованием рМОN 5467.
Результаты инсектицидной токсичности
С-терминальные усечения
Колеоптерановое токсиновое действие определяли посредством анализа с подачей свежевыведенных колорадских жуков на томатные листья, как описывалось вышее. Мутантные гены B.t.t. используемые для анализа С-терминалов, показаны на фиг. 17-18 и 21. рМОN 5438 содержал 490 аминокислот токсинового белка В. t.t. плюс 3 аминокислоты, кодированные связывающей молекулой, используемой в построении вектора. Усеченный белок продуцировался в высокой степени в E.coli, но не обладал активностью против колорадского жука. pMON 5441 продуцировал белок, который содержал 536 аминокислот токсина B.t.t. Усеченный белок был продуцирован в высокой степени в E.coli, но не обладал активностью против колорадского жука. рМОN 5449 содержал 582 аминокислоты белка B. t.t. плюс две аминокислоты, кодированные связующей молекулой, используемой в данном построении вектора. Усеченный белок был продуцирован в высокой степени в Е.coli, но не обладал активностью против колорадского жука. рМОN 5448 содержал 640 аминокислот белка В.t.t., плюс 2 аминокислоты, кодированные связующей молекулой, используемой в данном построении вектора. Этот усеченный белок продуцировался в высокой степени в Е.coli, но данный белок не обладал активностью против колорадского жука. Эти результаты подтверждают, что для токсичности действия против колорадского жука требуется С-терминал токсинового белка B.t.t. Делеция лишь 4 амино (рМОN 5448) кислот приводила в результате к полной потере активности. Эти результаты прямо противоположны сообщающимся в литературе данным о токсинах B.t. лепидоптеранового типа (для чешуекрылых).
Результаты N-терминальной мутации и делеций
Другие мутантные гены B.t.t., используемые для анализа N-терминалов, показаны на фиг. 19-21. Анализ белка, продуцируемого рМОN 5450, показал, что продуцированные полосы 3 в Е.coli обусловлено инициированием трансляции в МЕТ48, а не продуктом расщепления протеазы. Изучение токсичности показало также, что полоса 3 была токсичной, рМОN 5456 продуцировал белок, который начинался у аминокислоты 48 с аминокислотой 49, изменяемой от треонина до аспаргиновой кислоты. Этот белок был продуцирован в высокой степени в Е.соli, был токсичен к колорадскому жуку, pMON 5472 продуцировал белок, который начинался у аминокислоты 77. Этот белок был выражен в Е.coli и обладал активностью действия против колорадского жука. рМОN 5467 продуцировал белок, который начинался у аминокислоты 99 и имел также две аминокислоты, введенные в N-терминал (метионин и аланин). Данный белок продуцировался в Е.соli не обладал активностью действия против колорадского жука, однако степень выражения этой делеции была значительно ниже, чем других типов делеций.
Данные результаты подтверждают, что N-терминал токсинового белка B.t.t. может быть устойчивым к делециям. Делеция 76 аминокислот обнаруживала токсичность. Делеция 99 аминокислот, однако приводила в результате к потере токсичности, рМОN 5460 содержал мутацию, которая изменяла метионин в положении 48 на изолейцин, что предотвращало продуцирование полосы 3. Токсичность полосы I, продуцируемой рМОN 5460, была эквивалентна токсичности полосы 3, продуцируемой рМОN 5456.
Построение векторов растительной трансформации.
Токсиновый ген В.t. разновидности tenebrionis, содержащий в рМОN 5420, был модифицирован для вставки в векторы выражения растений. Точка Bgl II была вставлена непосредственно выше ATG кодона, который определяет инициирование трансляции токсинового белка B.t.t. полной длины (полоса I) с использованием точечно специфического мутагенеза согласно Kunkel, 1985 г. как описывалось выше. Последовательность данного токсинового гена В.t.t. в области инициатора АТG следующая:
ATGATAAGAAAGGGAGGAAGAAAAATGAATCCGAACAATCGAAGTGAACATGATACAATA
Met Asn Pro Asn Arg Ser Glu His Asp Thr Ile
Затравка для данного мутагенеза (bttbgl) составляла 27 нуклеотидов (по длине) и имела следующую последовательность:
СGGATTCATT TTAGATCTTC CTCCCTT
После мутагенеза плазмида, содержащая новую точку Bgl II, была идентифицирована путем вываривания с Bgl II, данное изменение было проверено путем анализа последовательности ДНК. Полученная в результате плазмида, содержащая токсиновый ген B.t.t. с новой точкой Bgl II, была обозначена рМОN 9758 (фиг. 22).
Токсиновый ген В.t.t. в рМОN 9758 был вставлен в кассетный вектор выражения рМОN 316 (Sanders и др., 1987 г.) рМОN 316 содержал промотор СаМV 35S и 3' конец от гена нопалинсинтазы (NOS) с точкой Bgl II для генной вставки между этими двумя элементами. Плазмида рМОN 9758 вываривалась с Вgl II, был выделен фрагмент приблизительно 2,3 kb. Этот фрагмент простирался от точки Bgl II непосредственно над кодоном АТG до точки Bgl II примерно на 350 bp ниже терминального кодона для токсинового гена B.t.t. Таким образом, данный фрагмент содержал полную кодирующую последовательность гена B.t.t., а также примерно 350 bp некодирующей последовательности 3' до терминального кодона. Этот фрагмент Bgl II был сшит с рМОN 316, вываренным с Bgl II. После трансляции в Е.соli, идентифицировалась колония, в которой токсиновый ген B.t.t. был вставлен в рМОN 316 таким образом, что 5' конец токсинового гена находился в непосредственном соседстве с промотором СаМV 35S. Такая плазмида была обозначена как рМОN 9753. Плазмида, содержащая токсиновый ген B. t.t. в противоположной ориентации в pMON 316, была выделена и обозначена как рМОN 9754 (фиг. 22).
Как рМОN 9753, так и рМОN 9754 были вставлены путем трехродительского спаривания в АSE штамм Agrobacterium tumefaciens, который содержал неусиленную Ti плазмиду. Соинтеграты между pMON 9753 или рМОN 9754 и неусиленной Ti плазмидой были идентифицированы как описано Fraley и др. (1985 г.) и их структуры были подтверждены анализом всей ДНК Agrobacterium.
Были построены также дополнительные векторы выражения растений, содержащие токсиновый ген B.t.t. (см. фиг. 23 и 24). В этих векторах токсиновый ген B.t.t. был вставлен в вектор выражения растений рМОN 893 (фиг. 25). Как показано на фиг. 25, кассетный вектор выражения рМОN 893 состоит из усиленного промотора СаМV 35S и 3' конца, включающего сигналы полиаденилирования от гена сои культурной, кодирующей альфа-затравочную субъединицу бета-конглицина (смотри как 7S ген). Между этими двумя элементами имеется многозвенная связывающая молекула, содержащая множество ограничительных точек для генной вставки.
Усиленный промотор СаМV 35S был построен следующим образом. Был предварительно построен фрагмент СаМV 35S промотора, простирающегося между положением - 343 и + 9 в рUC13, Odell и др., 1985 г. Этот сегмент содержал область, идентифицированную Odell и др., 1985 г., как область, необходимую для максимального выражения промотора СаМV 35S. Он был вырезан как фрагмент Cla I - Hind III затуплен по концам ДНК полимеразой I (фрагментом Кленова) и вставлен в точку Hinc II плазмиды pUC18. Область выше промотора 35S была вырезана из данной плазмиды как фрагмент Hind III - Eco RV (простирающийся от -343 до д-90) и вставлена в ту же плазмиду между точками Hind III и Pst I. Таким образом, усиленный промотор СаМV 35S содержал дубликат последовательностей между -343 и -90 (см. фиг. 29).
3' конец 7S гена образован от гена 7S содержащегося на клоне, обозначенном 17.1 (Schuler и др. , 1982 г.). Этот 3' концевой фрагмент, который включает сигналы полиаденилирования, простирается от точки Ava II, расположенной примерно на 30 bp выше концевого кодона бета-конглицининового гена в клоне 17,1 до точки Есо RI, расположенной примерно на 450 bp ниже этого концевого кодона.
Остальная часть рМОN 893 содержит сегмент от рВ R322, который обеспечивает источник репликации в Е.coli и область гомологичной рекомбинации с неусиленной Т-ДНК в АСО штамме Agrobacterium (описано ниже); область ori V от плазмиды RK2 хозяина широкого диапазона; стойкий к спректиномицину ген от Tn 7; и химерный ген NPT11, содержащий промотор СаМV 35S и 3' конец нопалинсинтазы (NOS), который обеспечивает стойкость к канамицину в трансформированных растительных клетках.
рМОN 9753 содержит примерно 400 bp 3' некодирующей последовательности за пределами концевого кодона. Поскольку эта область не является необходимой для продуцирования токсина, то она удаляется из сегментов токсинового гена В.t.t., вставленных в рМОN 893. Для того, чтобы создать токсиновый ген B. t. t., не содержащий никакой 3'-боковой последовательности, точка Bgl II была вставлена непосредственно после концевого кодона методом Kunkel, 1985 г. Последовательность данного токсинового гена B.t.t. около концевого кодона следующая:
GTTTATATAGACAAAATTGAATTTATTCCAGTGAATTAAATTAACTAGAAAGTAAAGAAG
Val Tyr Ile Asp Lys Ile Glu Phe Ile Pro Val Asn End
Мутагенез осуществлялся с затравкой (bttcterm), имеющий последовательность:
CTTTCTAGTTAAAGATCTTT AATTCACTG
Мутагенез токсинового гена B.t.t. осуществлялся в рМОN 9758. Плазмида, которая содержала новую точку Bgl II, была обозначена рМОN 9787 (фиг. 23). Поскольку рМОN 9787 содержал точку Bgl II непосредственно выше инициирующего кодона АТG, то полная кодирующая последовательность токсинового гена B.t.t., не содержащая ни 5' ни 3' боковой последовательности, находилась на фрагменте Bgl II, составляющем примерно 1940 bp.
Этот фрагмент на 1940 bp был выделен из рМON 9787 и сшит с рМОN 893, вываренным с Blg II. Плазмида, в которой 5' конец токсинового гена B.t.t. был смежен с усиленным промотором СаMV35S, была идентифицирована и обозначена как рМОN 9791 (фиг. 23).
Другой вид токсина B. t.t. полной длины был продуцирован в Е.coli из второго метионинового инициирующего кодона. Этот белок, обозначенный как "полоса 3", был в такой же степени токсичен к колорадскому жуку, как и токсин полной длины цепи ("полоса 1"). Возможно, что, как и в случае гена B.t.t., усеченные формы гена B.t.t. могут быть более легко выражены в растительных клетках. Таким образом, был построен модифицированный токсиновый ген B. t.t., в котором область выше АТG кодона полосы 3, была удалена. Для того, чтобы удалить эту последовательность, точка Bgl II была вставлена непосредственно выше АТG полосы 3 методом Kunkel, 1985 г. Последовательность около АТG полосы 3 следующая:
CCAAATCCAACACTAGAAGATTTAAATTATAAAGAGTTTTTAAGAATGACTGCATAAT
ProAsnProThrLeuGluAspLeuAsnTyrLysGluPheLeuArgMetThrAlaAspAsn
Мутагенез осуществлялся с затравкой (bttnterm), имеющий указанную последовательность:
ATCTGCAGTC ATTGTAGATC TCTCTTTATA ATTT
Мутагенез с этой затравкой осуществлялся на токсиновом гене B.t.t., содержащемся в рМОN 5420. Плазмида, содержащая новую точку Bgl II, была обозначена как рМОN 9788. Усеченный B.t.t. токсиновый ген, начинающийся в точке Bgl II данной полосы 3 и простирающийся до точки Bgl II, расположенной по периферии концевого кодона в рМОN 9787 был построен в рМОN 893 следующим образом. pMON 9788 (фиг. 24) был выварен с Bgl II и Xba I, и был выделен фрагмент, составляющий примерно 1250 bp. Этот фрагмент простирался от ATG полосы 3 до уникальной точки Xba I в середине токсинового гена B.t. t. рМОN 9787 вываривался также с Bgl II и Xba I, был выделен фрагмент, составляющий примерно 550 bp. Этот фрагмент простирался от уникальной точки Xba I в середине токсинового гена до точки Bgl II, расположенной по периферии концевого кодона. Эти два фрагмента смешивались и сшивались с рМОN 893, вываренным с Bgl II. Была идентифицирована плазмида, в которой 5'-конец токсинового гена был смещен с усиленным промотором СаМV 35S и которая была обозначена как рМОN 9792. Эта плазмида рМОN 9792 включает n-терминальное усеченное производное токсинового гена B.t.t. (фиг. 24), которое кодирует лишь полосу 3.
Как рМОN 9791, так и рМОN 9792 были вставлены в АСО штамм A. tumefaciens, который содержит неусиленную плазмиду Ti.
Были отобраны соинтеграты, которые использовались для трансформации томатов и картофеля.
АСО представляет собой неусиленный штамм, аналогичный рTi B6SE, описанному Fraley и др., 1985 г. Для построения АСО исходный штамм Agrobacterium представлял собой штамм А208, который содержал Ti плазмиду нопалинового типа. Ti плазмида была усилена аналогично описанию Fraley и др., 1985 г., таким образом, чтобы удалялась практически вся естественная Т-ДНК, за исключением левого края и нескольких сотен основных ар Т-ДНК внутрилевого края. Остальная часть Т-ДНК, простирающаяся до точки непосредственно за правым краем, была заменена новой частью ДНК, включающей (слева направо) сегмент рВ R322, область ori V от плазмиды RK2 b cтойкий к канамицину ген от Tn 601. Сегменты рВR 322 и oriV аналогичны сегментам в рМОN 893, и обеспечивают область, гомологичную для соинтеграта. Структура АСО Ti плазмиды показана на фиг. 28.
Химерный токсиновый ген В.t.t., использующий промотор МАS
Промотор MAS был выделен из pTi A6 как фрагмент 1,5 kb Eco RI - Cla I. Этот фрагмент ДНК простирался от точки Cla I с нуклеотидом 20 138 до точки Есо RI с нуклеотидом 21 631 в последовательности Barker и др., 1983 г. Как показано на фиг. 26, фрагмент Есо RI - Cla I был сшит с бинарным вектором рМОN 505 (Horsch и др., 1986 г.), который был предварительно выварен с Есо RI и Cla I. Образующаяся в результате плазмида была обозначена как рМОN 706. Фрагмент, содержащий конец 3' NOS, был вставлен ниже промотора МАS, в результате чего был получен кассетный вектор выражения MAS - NOS 3'. Фрагмент NOS 3' был вырезан из (рМОN 530 как фрагмент 300 bp Bgl II - Bam HI, и был вставлен с рМОN 706, вываренный с Bgl II. Полученная плазмида обозначена как рМОN 707.
Была построена плазмида рМОN 530 путем расщепления рМОN 200 с Nde I с целью удаления фрагмента 900 bp Nd I, что приводило к образованию рМОN 503. Плазмида рМОN 503 расщеплялась Hind III и Sma I и смешивалась с плазмидой рT IS75 (Schmidhauser и Helinski, 1985), которая также расщеплялась Hind III и Sma I. Плазмида, которая содержала фрагмент 3,8 kb Hind III - Sma I от рTIS75, соединенный с фрагментом 8 kb Hind III - Sma I от рМОN 503, была выделена и обозначена как рМОN 505. Следующая кассета CaMV35S - NOS 3' была перенесена в рМОN 505 путем расщепления рМОN 316 с Stu I и Hind II и выделения фрагмента 2,5 kb, Stu I - Hind III, содержащего сигнальный ген NOS - NPTII' - NOS и кассету Cа МV35S - NOS 3'. Она была введена в рМON 505 ДНК и расщеплена Stu I и Hind III. После сшивки и трансформации была выделена плазмида, несущая кассету CaMV35S - NOS 3' в pMON 505, и обозначена как рМОN 530. Поскольку некоторые бинарные векторы имеют значительно пониженные частоты трансформации в томатах, по сравнению с соинтегрирующими векторами (Mc Cormick и др., 1986 г.), то кассета МАS - NOS 3' была удалена из рМОN 707 в соинтегрирующий вектор рМОN 200 (Fraley и др., 1985 г.). Плазмида рМОN 200 была выварена с Stu I и Hind III, и фрагмент 7,7 kb был выделен путем гель-электрофореза на агарозе. Плазмида рМОN 707 вываривалась аналогичным образом с Stu I и Hind III, и фрагмент 3 kb Stu I - Hind III, содержащий кассету MAS - NOS 3' был выделен путем гель-электрофореза на агарозе и извлечения на DEAE мембранах с последующим элюированием в 1 моль NaCl. Эти два фрагмента ДНК сшивались, и образованная в результате плазмида была обозначена как рМОN 9741 (фиг. 26). Эта плазмида содержала кассету МАS - NOS 3' в соинтегрирующем рМОN 200 фоне. Химерные токсиновые гены B.t.t., стимулированные промотором МаS, приготавливались путем вываривания либо рМОN 9791, либо рМОN 9792 c Bgl II, извлечения кодирующего токсин фрагмента и удаления этого фрагмента в рМОN 9741 согласно данному описанию.
Эти промежуточные векторы могут быть использованы для трансформации растений, проявляющих токсичность к жесткокрылым насекомым, чувствительным к токсиновому белку B.t.t.
Выражение в растениях токсинового гена колеоптеранового типа (жесткокрылых насекомых)
Трансформация томатных растений
Штаммы А. tumefaciens pMON 9753-ASE и рМON 9754-АSE использовались для трансформации дисков томатных листьев методом Mc Cormick и др., 1986 г. Трансформированные томатные растения восстанавливались, как описано в заявке, и анализировались на стойкость к канамицину.
Токсичность к насекомым трансгенных томатных растений.
Томатные растения, трансформированные токсновым геном B.t.t. содержащимся в рМОN 9753, анализировались на выражение токсинового гена путем биоанализа с использованием колорадского жука (Leptinotarsa decemlineata). Подвергаемые анализу срезы листьев помещались в чашки Петри, содержащие насыщенную водой фильтровальную бумагу. На каждый срез листа вводили десять или двадцать свежевыведенных колорадских жуков. По прошествии четырех дней рассчитывали смертность насекомых. Кроме того, насекомых исследовали на возможность замедленного темпа роста (остановка в росте), и оставшуюся листовую ткань исследовали на выявление относительного повреждения.
В каждом эксперименте в качестве контрольных использовались много нетрансформированных растений. В качестве контрольных анализировали от 50 до 100 нетрансформированных растений. Из числа этих контрольных растений более чем 80% не показали смертности колорадского жука, примерно 15% растений показали 10% смертности и 5% или менее растений показали 20% смертности. Контрольные растения не показали смертность насекомых более чем 20%.
В табл. 6 суммированы данные токсичности, полученные с некоторыми томатными растениями, содержащими рМОN 9753,
Токсичность трансгеных томатных растений, содержащих рМОN 9753 против колорадского жука
Как показано в табл. 6, некоторые растения были восстановлены, показывая одновременно повышенную степень смертности колорадского жука по сравнению с нетрансформированными контрольными растениями. Эти результаты показывают, токсиновый ген B.t.t. выражается в такой степени, которая достаточна для умерщвления значительного количества насекомых, подаваемых на данные растения.
Выражение токсина жесткокрылых насекомых в картофеле
Кончики побегов картофельного растения сорта Кеппевес пересевались в среде, содержащей основную и минимальную MS соль, 0,17 г/л первичного кислого фосфата натрия, 0,4 мг/л тиамина НСl, 0,1 г/л инозитола, 3% сахарозы, 2,0 г/л Герлита (Kelco Co.) c pH 5,6. Культуры выращивались в течение 4 недель при 24оС в 16-часовом фотопериоде. Междоузлия стебля разрезали на отрезки длиной примерно 8 мм и разрезанные поверхности промазывались штаммом Agrobacterium pMON 9753 - ASЕ, который подвергался штриховой разводке на пластине с LB агаром и выращивался в течение 2-3 дней. Указанный рМОN 9753-ASE содержал химерный токсиновый ген B.t.t., стимулированный промотором CaMV 35S. Как возможный вариант использовались штаммы Agrobacterium pMON 9791 - ACO или рМОN 9792-АСО, содержащие химерные токсиновые гены B.t. t. Секции стебля помещались в отвержденную 0,8% агаром среду, содержащую соли и органические продукты присоединения.
Jarret и др., 1980 г., 3% сахарозы, 3 мг/л ВА и 0,1 мг/л NAA при величине рН 5,6. Спустя четыре дня экспланты переносились в среду того же состава, но с карбонициллином концентрацией 500 мг/л и канамицином в качестве реагента избирательного действия для трансформированных клеточных растений концентрацией 100 мг/л. Спустя четыре недели экспланты снова переносились в среду того же состава, но с GA3 равной 0,3 мг/л, как единственного гормона. Каллус, образованный в присутствии 100 мг/л канамицина, содержал фермент NPTII, как установлено в ходе испытания методом точечного анализа, и это указывало на то, что клетки картофельного растения трансформированы. Неинокулированная контрольная ткань ингибировалась при данной концентрации канамицина. Трансформированная картофельная ткань выражает токсиновый ген B. t. t. мРНК токсина B.t.t. может быть обнаружена методом анализа Northern и токсиновый белок B.t.t. может быть обнаружен методом иммуноанализа, например методом точечного анализа Вестерна. Однако во многих случаях наиболее чувствительным анализом на присутствие токсина B.t.t. является биоанализ. Личинки колорадского жука, подаваемые на трансформированную ткань, страдали от действий токсина.
Эта операция продуцирования стойких к канамицину трансформированных клеток картофельного растения была в последующем использована для регенерации побегов. Побеги длиной 1-2 см удалялись от эксплантов и помещались в физиологический раствор кончиков побегов, как описано выше, в котором эти побеги легко укоренялись.
Растения, выращиваемые таким образом, подвергались испытанию на трансформацию путем анализа с целью определения выражения фермента NPTII и по способности сегментов стебля образовывать каллус в содержащей канамицин среде. Трансформированные растения выражают токсиновый ген B.t.t. мРНК токсина B.t.t. могла быть обнаружена методом анализа Northern и токсиновый белок B. t.t. мог быть обнаружен методом иммуноанализа, таким как точечный анализ Вестерна. Личинки колорадского жука, подаваемые на трансформированную ткань, отпадали от действия токсина.
Выражение токсина жесткокрылых насекомых в хлопке.
Семена хлопка поверхностно стерилизовались путем первоначального погружения их на 10 мин в моющий раствор воды, в который добавляли мыло Sparkleen, и последующего перемешивания этого раствора в течение 20 мин в 30% растворе Хлорокс, содержащем 2 капли Твина 20 на 40 мл, после чего осуществлялась их двукратная промывка стерилизованной дистиллированной водой. Данные семена затем погружали в 0,4% бенолат на 10 мин. Бенолат сливали до асептического ввода семян на отвержденные агаром соли МS половинной крепости. Семена проращивались в течение 3-10 дней в темноте при температуре 32оС. Затем семядоли и подсемядольные колена асептически удаляли и разделяли на сегменты. Эти сегменты помещали: (I) в отвержденную агаром среду МS, содержащую 3% глюкозы, 2 мг/л нафталинуксусной кислоты (NAA) и 1 мг/л кинетина (Среда МSS), или (2) в отвержденную Герлитом среду MS, cодержащую 3% глюкозы, витамины В5, 100 мг/л инозитола, 0,75 мг/л MgCl2, 0,1 мг/дихлорфеноксиуксусной кислоты (2,4-D) и 0,1 или 0,5 мг/л кинетина (среда MST). Каллус сохранялся в течение 16/8 фотопериода при 28оС на любой из этих сред до тех пор, пока не инициировался эмбриогенез. Субкультура эмбриогенного каллуса приготавливалась в той же среде, что и среда инициирования, но содержащей 3% сахарозы вместо глюкозы. Соматические эмбрионы проращивались путем переноса их в отвержденую Герлитом среду Stewart без регуляторов роста растений, но содержащую 0,75 г/л MgCl2. Пророщенные эмбрионы переносятся в почву в камере роста, в которой они продолжают расти. Затем растения переносятся в теплицу для посадки семян и цветения.
Трансформация тканей хлопкового растения и продуцирование трансформированных каллусов и растений осуществлялись следующим образом. Приготавливались асептические сеянцы, как и для регенерации растения. Сегменты подсемядольных колен и семядолей инокулировались жидкими ночными культурами Agrobacterium или Agrobacterium, выращенными на питательных чашках. Эти экспланты совместно культивировались в течение 2-3 дней в среде MSS или MST, содержащей соли MS с разведением 1/10. Экспланты наносились на фильтровальную бумагу с целью удаления избытка бактерий и высевались на чашках со средой MSS или MSN, содержащей 500 мг/л карбонициллина и 30-100 мг/л канамицина. Каллус, который был трансформирован, должен быть выращен на данной среде и будет продуцировать эмбрионы. Эти эмбрионы растут, превращаясь в растения, как указано для регенерации. Растения испытываются на трансформацию путем анализа выражения NPT11.
Когда используемый для трансформации штамм Agrobacterium содержит токсиновый ген B. t. t., такой как pMON 9753, pMON 9791 или рМОN 9792, то токсиновый ген B. t. t. выражается в трансформированный каллус, эмбрионы стимулируются от этих каллус и трансформированные растения стимулируются от этих эмбрионов. Для всех этих случаев выражение мРНК токсина B.t.t. может быть обнаружено путем анализа Northern и выражение токсинового белка B.t.t. может быть определено методом иммуноанализа, такого как точечный анализ Вестерна. Биоанализ насекомых может быть наиболее чувствительным методом анализа на присутствие токсинового белка.
Инсектицидная токсичность каллус, эмбрионов или растений определяется методом биоанализа на личинках долгоносика хлопкового (Anthonomous grandis). Личинки долгоносика хлопкового, подаваемые на трансформированные клетки хлопкового растения, или на растения, выражающие токсиновый ген B.t. t. страдают от действия токсина.
Выражение токсинового гена жесткокрылых в кукурузе
Описание приготовления протопластов из кукурузы, введение химерных токсиновых генов B. t. t. в протопласты путем электропорации и выделение устойчиво трансформированных стойких к канамицину клеток кукурузного растения, выражающих химерные токсиновые гены B.t.t.
Приготовление протопластов кукурузного растения.
Приготавливались протопласты из суспензии Мексиканской сладкой кукурузы (ВМS), ВМS1 (ATCC 54022), как описано Fromm и др., (1985 и 1986 г.). Клетки суспензии BMSI выращивались в среде BMS, содержащей соли MS, 20 г/л сахарозы, 2 мг/л (2,4-дихлорфенокси)уксусной кислоты, 200 мг/л инозитола, 130 мг/л аспарагеина, 1,3 мг/л ниацина, 0,25 мг/л тиамина, 0,25 мг/л пирипоксина, 0,25 мг/л пантотената кальция, рН-5,8, Сорок миллилитров культуры в 125-миллилитровых колбах Эрленмейера взбалтывали со скоростью 150 об/мин при температуре 26оС. Культура разбавлялась равным объемом свежеприготовленной среды каждые 3 дня. Протопласты извлекались из клеток активированного роста через 1-2 дня после ввода свежеприготовленной среды. Для извлечения протопластов клетки гранулировались в центрифуге с подвешивающимся качающимся столиком, со скоростью 200 х g. Всплывший на поверхность слой хранился как кондиционированная среда для выращивания протопластов. Шесть миллилитров уплотненных клеток повторно суспензировались в 40 мл 0,2 моль маннита (5 ммоль CaCl2) 10 ммоль ацетата натрия, содержащего 1% целлюлозы, 0,5% гемицеллюлазы и 0,02% пектиназы. После инкубации в течение 2 ч при температуре 26оС протопласты отделялись путем фильтрации через найлоновую сетку размером отверстий 60 мкм, центрифугировались при скорости центрифуги 200 х g и однократно промывались в том же растворе без ферментов.
Трансформация протопластов кукурузы ДНК векторам токсинового гена B.t. t. с использованием методa электропорации.
Протопласты для электропорации приготавливались путем промывки в растворе, содержащем 2 ммоль фосфата калия, рН 7,1, 4 ммоль хлорида кальция, 140 ммоль хлорида натрия и 0,2 моль маннита. После промывки протопласты снова суспензировали в том же растворе при концентрации 4 х 106 протопластов/мл. Полмиллилитра содержащего протопласты раствора смешивал с 0,5 мл того же раствора, содержащего 50 мкг ДНК вектора чрезмерно скрюченной плазмиды и помещали в 1-миллилитровую кювету электроплатинирования. Электроплатинирование осуществлялось как описано (Fromm и др. (1986 г.). Как описывалось, электрический импульс подавался от конденсатора на 122 или 245 микрофарад, заряженного до 200 Вт. По прошествии 10 мин выдержки при 4оС и 10 мин выдержки при комнатной температуре протопласты разбавлялись 8 миллилитрами среды, содержащей MS соли 0,3 моль маннита, 2% сахарозы, 2 мг/л 2,4-D, 20% кондиционированной среды ВМS (смотри выше) и 0,1% низкоплавкой агарозы. По прошествии двух недель выдержки в темноте при температуре 26оС вводили среду, не содержащую маннит и содержащую канамицин, так что конечная концентрация канамицина составляла 100 мг/л жидкого раствора. Спустя еще две недели микрокаллусы удаляли из жидкости и помещали на мембранный фильтровальный диск над отвержденной агарозой средой, содержащей 100 мг/л канамицина. Стойкие к канамицину каллусы, состоящие из трансформированных клеток кукурузного растения, обнаруживались спустя 1-2 недели.
Выражение токсиновых генов B.t.t. в клетках кукурузного растения
Как описывалось Fromm и др. (1986 г.), трансформированные клетки кукурузного растения могут быть отобраны путем роста в содержащей канамицин среде с последующей электропорацией векторами ДНК, содержащими химерные стойкие к канамицину гены, состоящие из промотора CaMV35S, кодирующей зоны NPT11 и 3' конца NOS, pMON 9791 и рМО 9792 содержат такие химерные гены NPT11 и содержат также химерные токсиновые гены B.t.t. Как описывалось выше, кукурузные протопласты трансформируются путем электропорации с векторами ДНК, где векторы ДНК представляют собой рМОN 9791 и рМОN 9792. После отбора на стойкость к канамицину трансформированные клетки кукурузного растения анализируются на выражения токсинового гена B.t.t.
Проводили испытания на определение мРНК B.t.t. методом точечного анализа Northern, и на определение токсинового белка B.t.t. методом иммуноанализа, например точечного анализа Вестерна.
Анализы на инсектицидную токсичность осуществлялись путем подачи каллусов трансформированного кукурузного растения на личинки жука-блошки длинноусой (Diabrotica undecimpunctata howardi). Как возможный вариант, из клеток трансформированного кукурузного растения приготавливался белковый экстракт, содержащий токсиновый белок B.t.t., и этот экстракт вводился в рацион для питания личинок жука-блошки длинноусой. Личинки жука-блошки длинноусой, подаваемые на трансформированные каллусы или белковые экстракты таких каллусов, страдали от данного токсина.
Изложенные примеры разъясняют прак- тические аспекты изобретения, но не ограничивают объем изобретения.
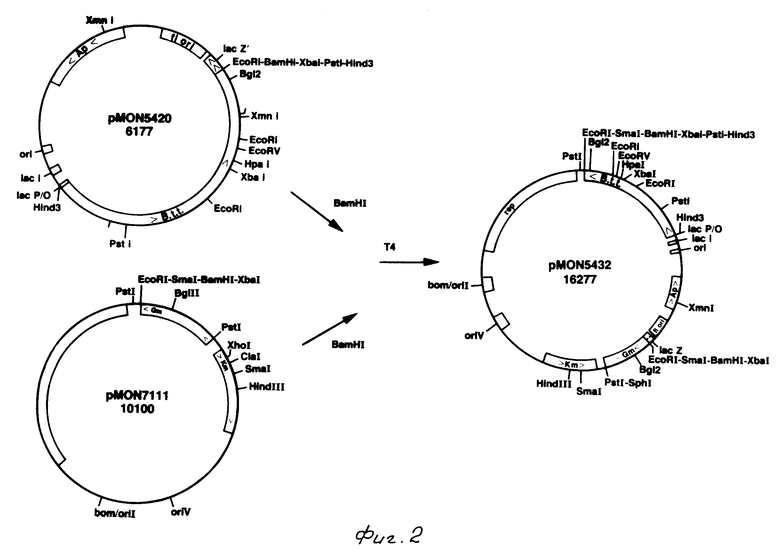
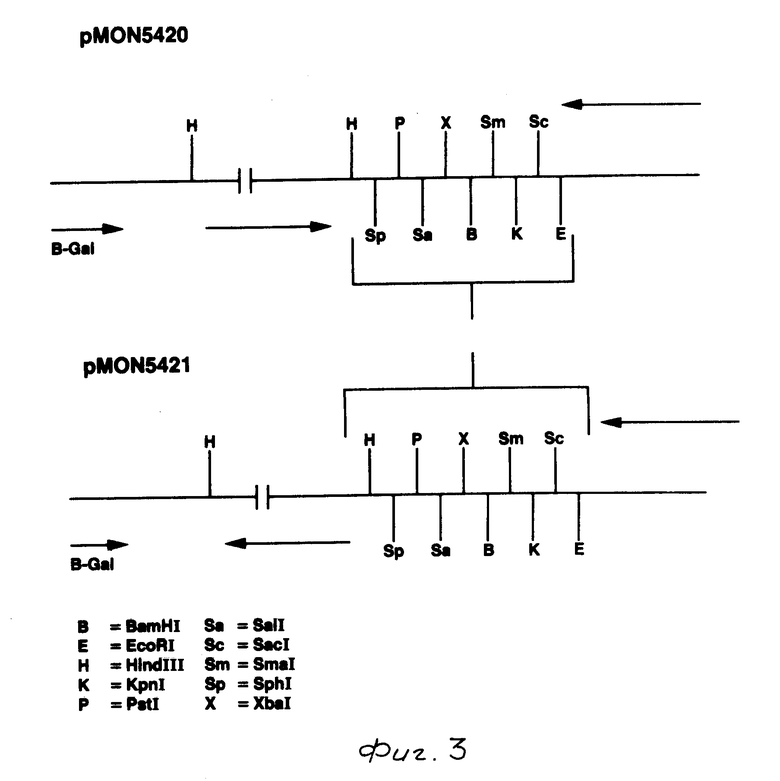
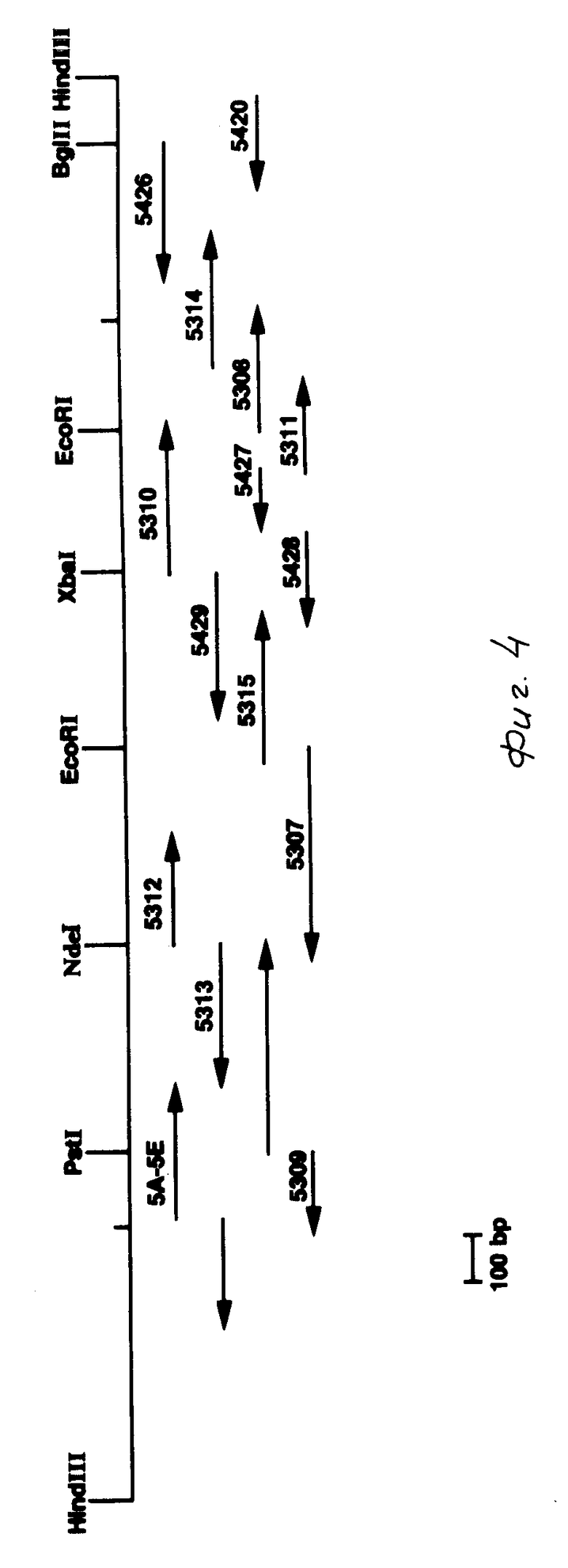

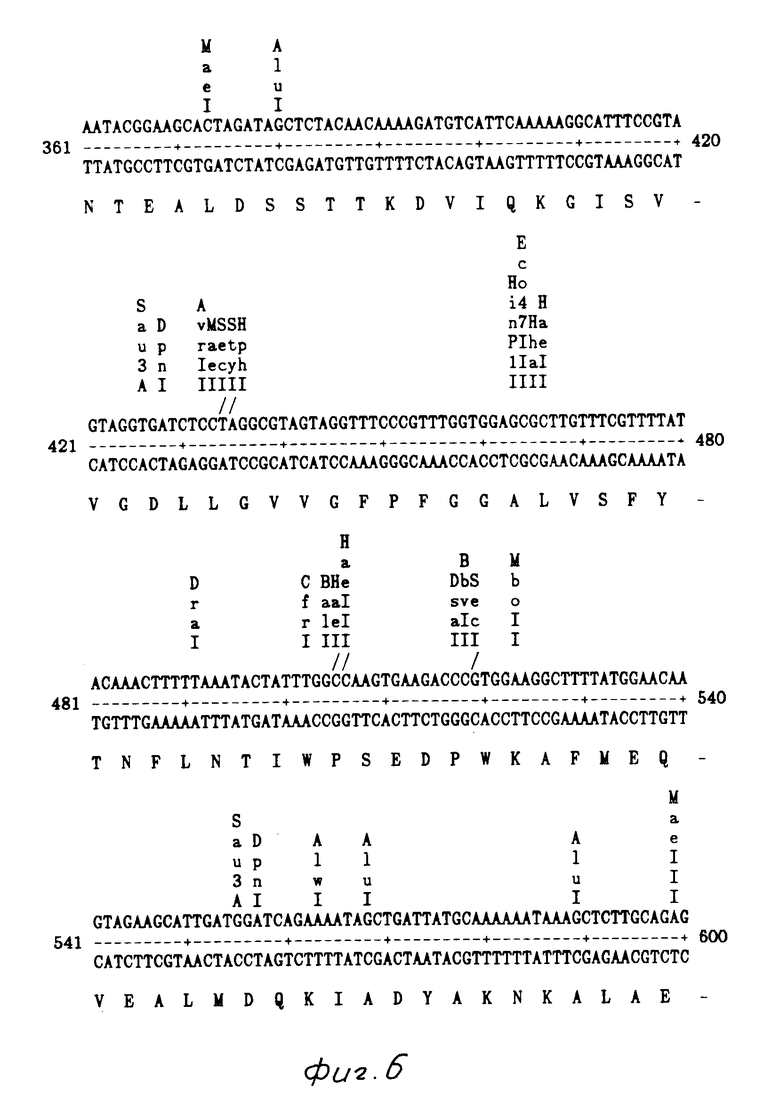
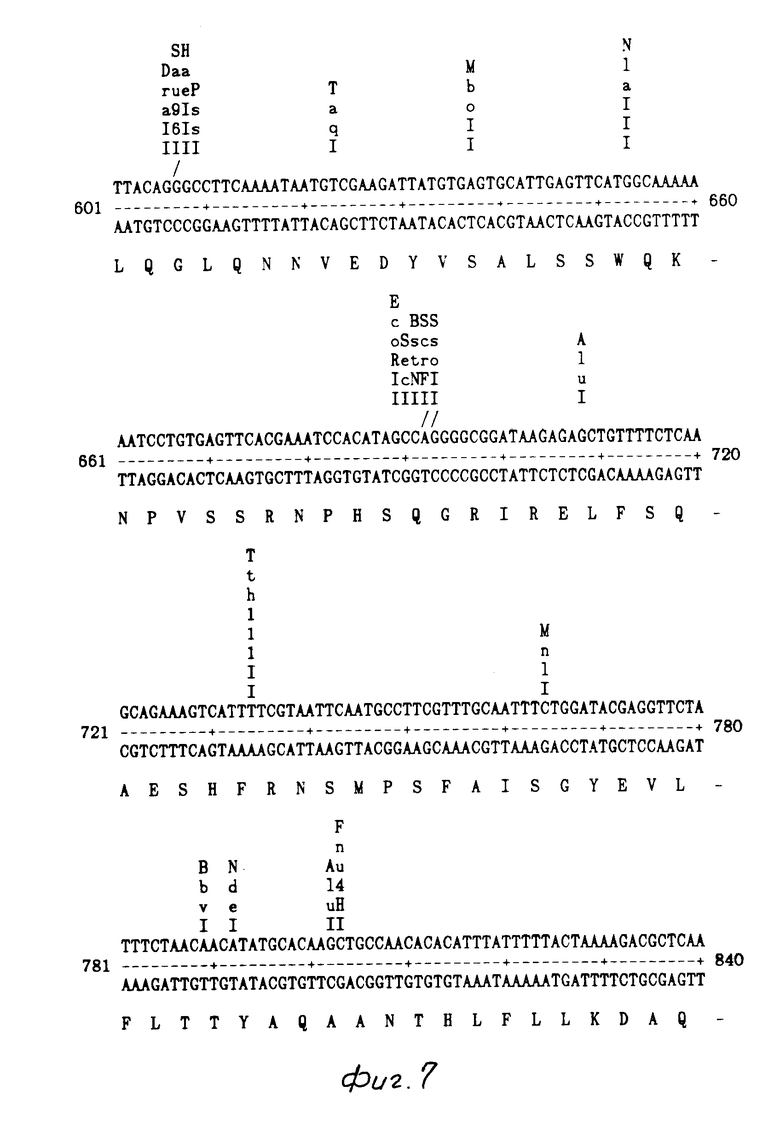
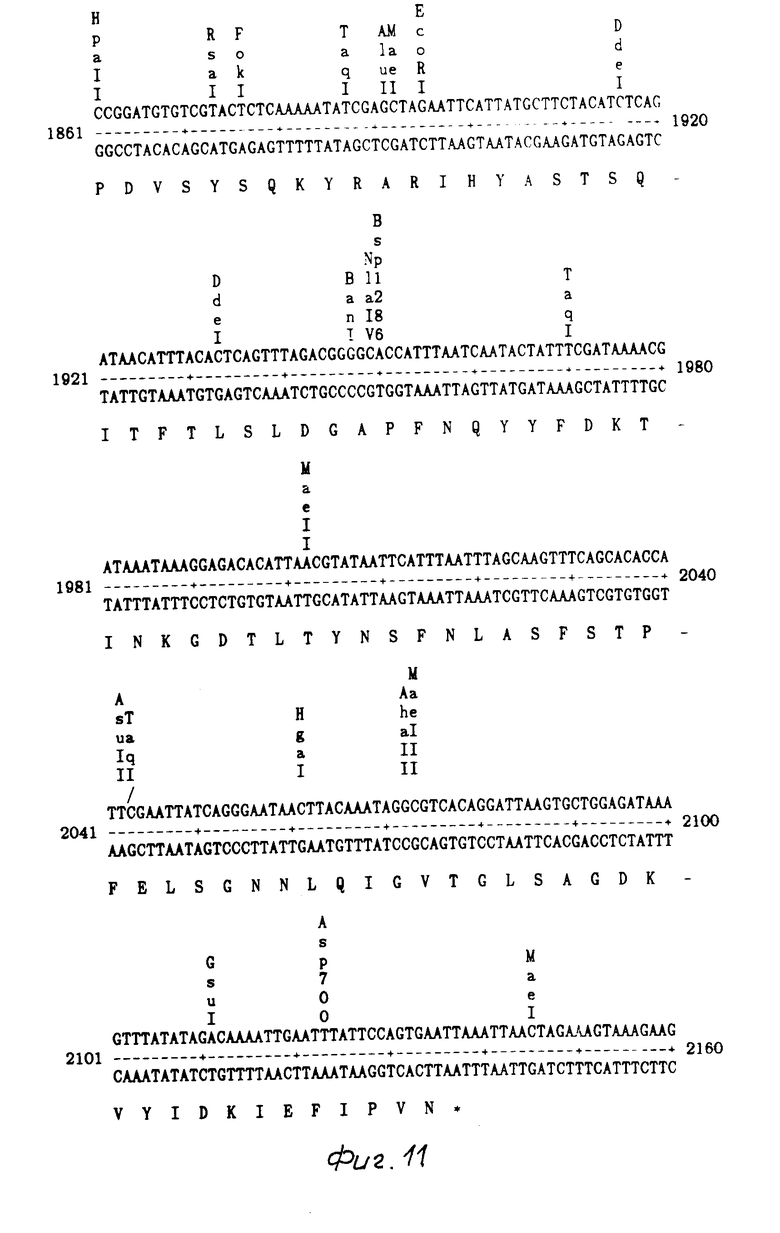
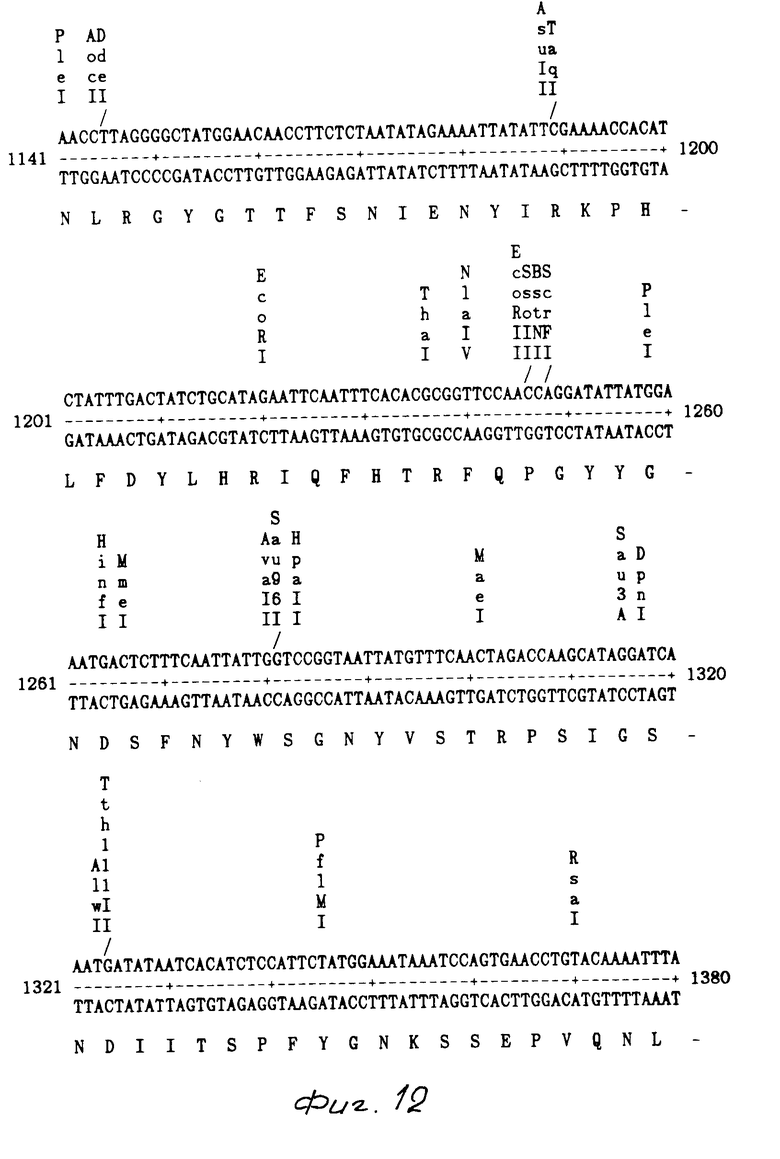
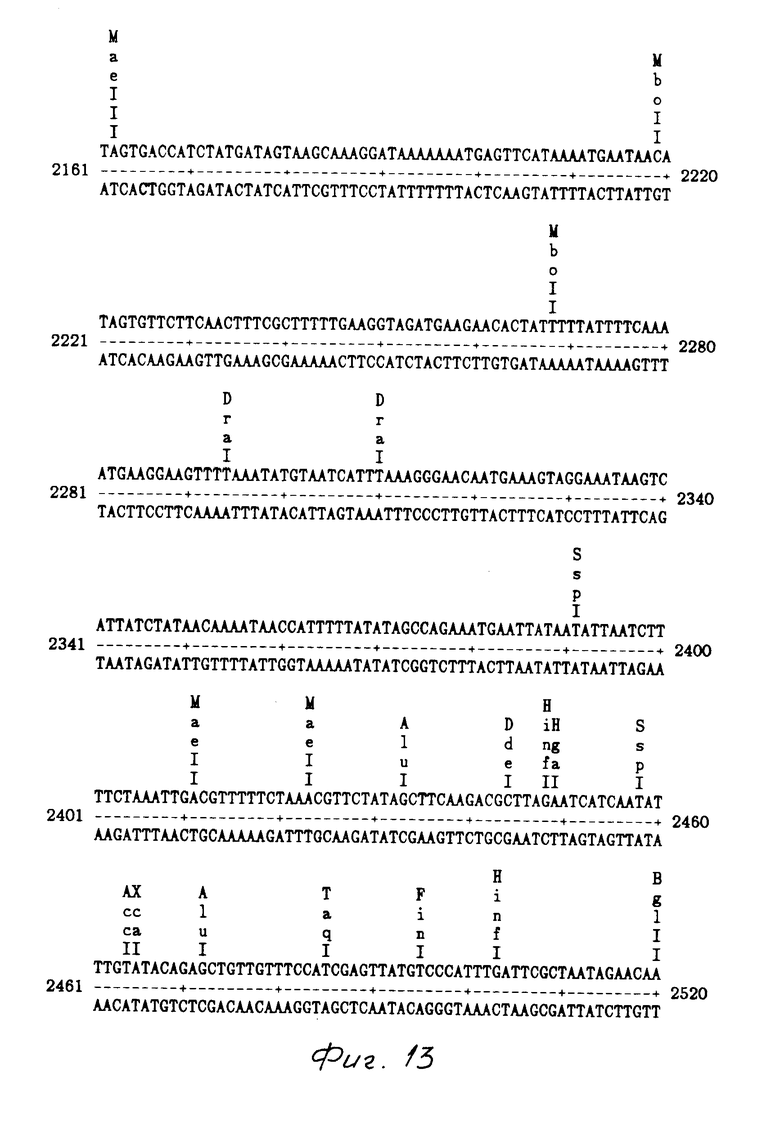
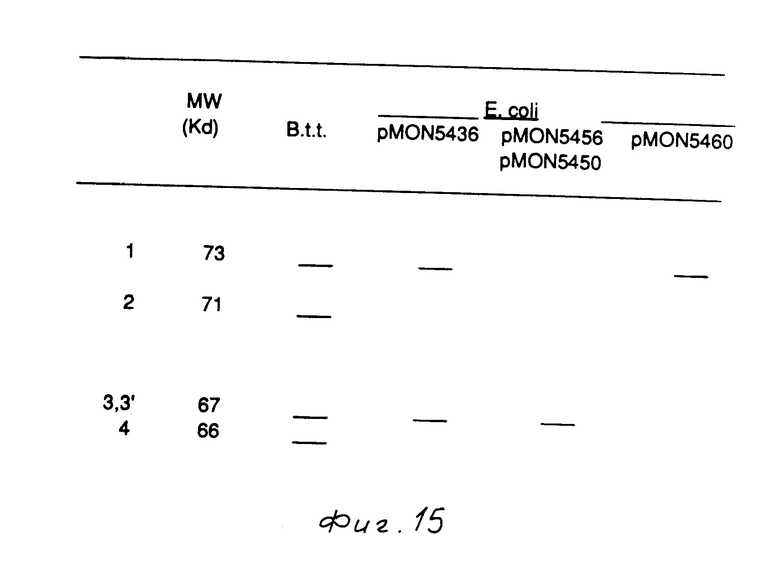

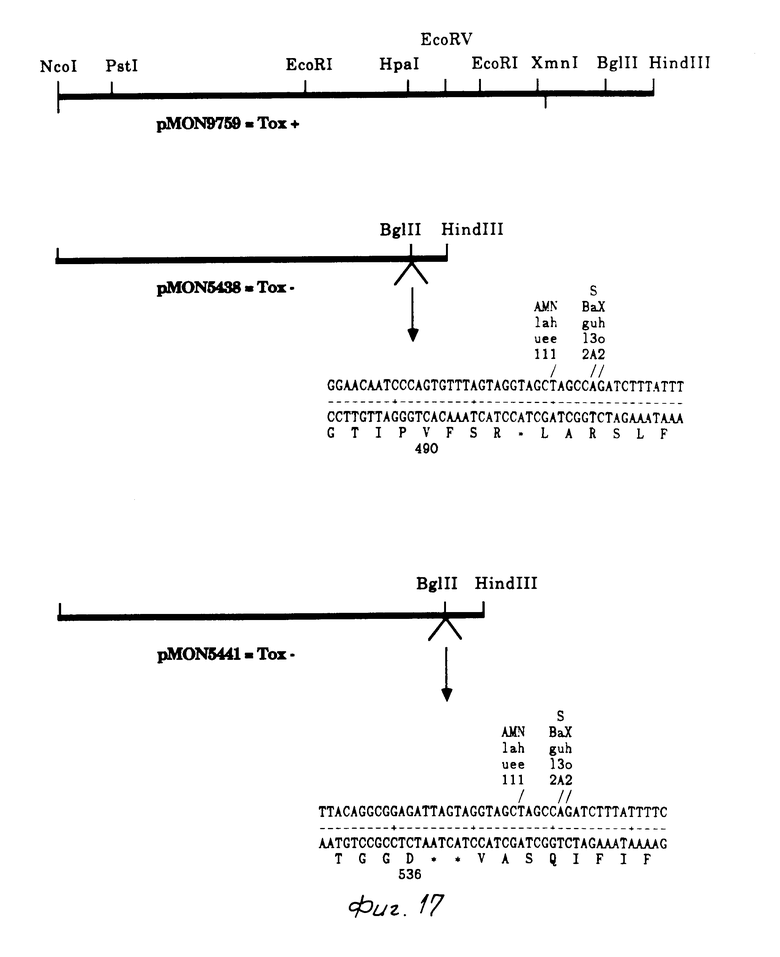
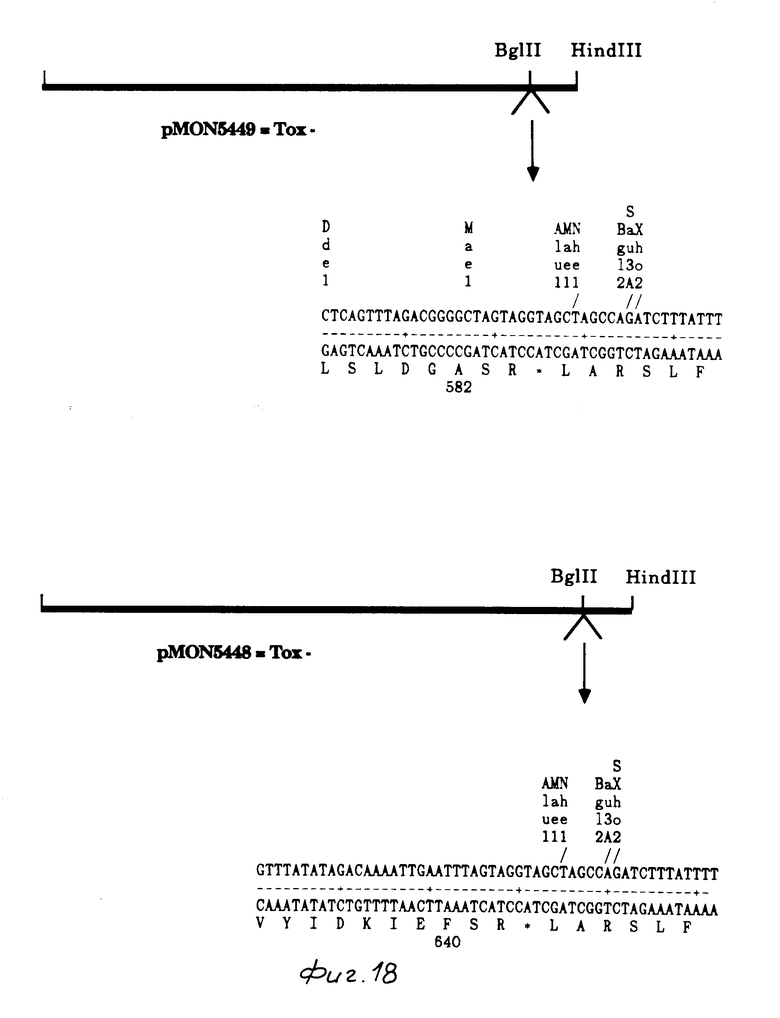
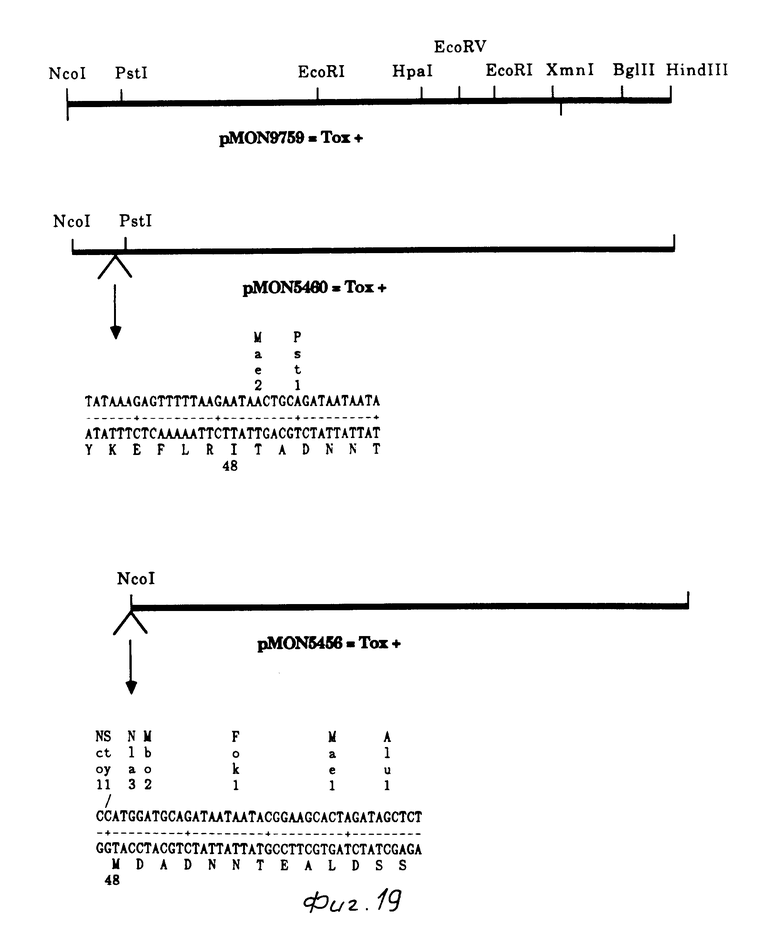
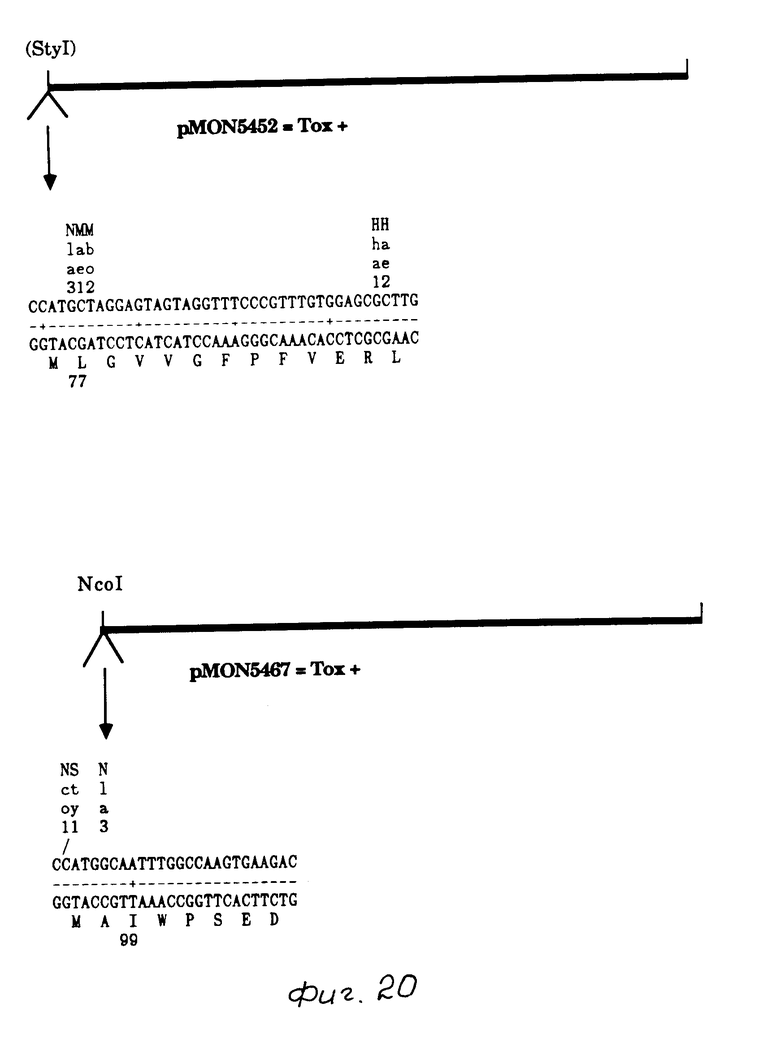

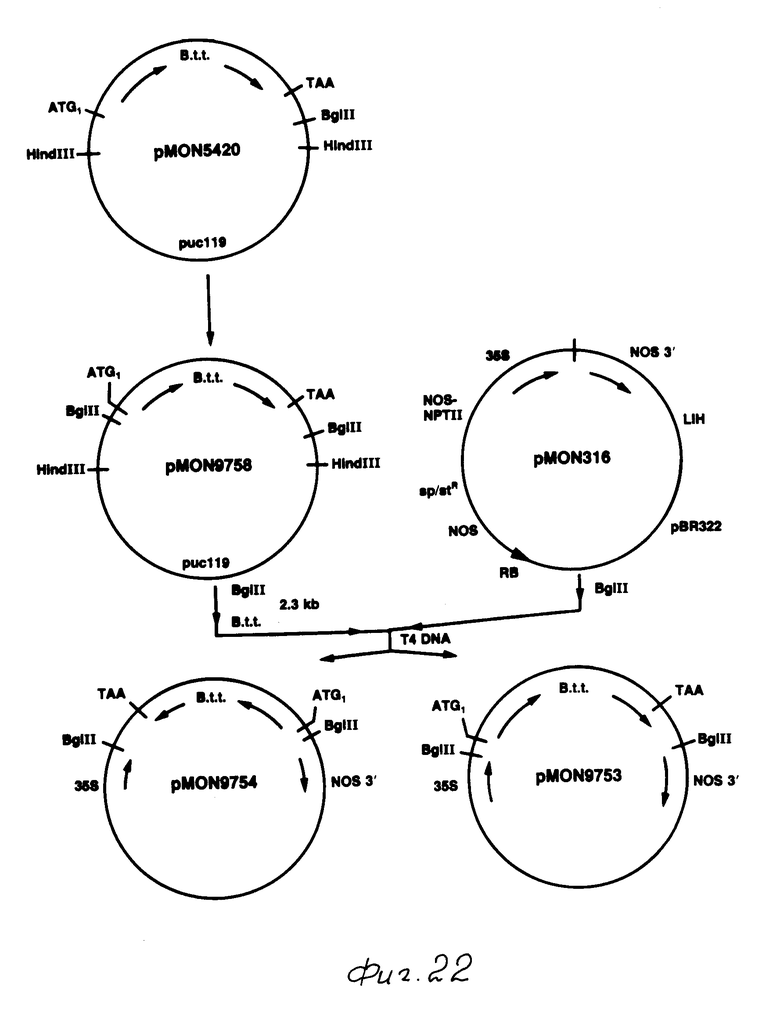
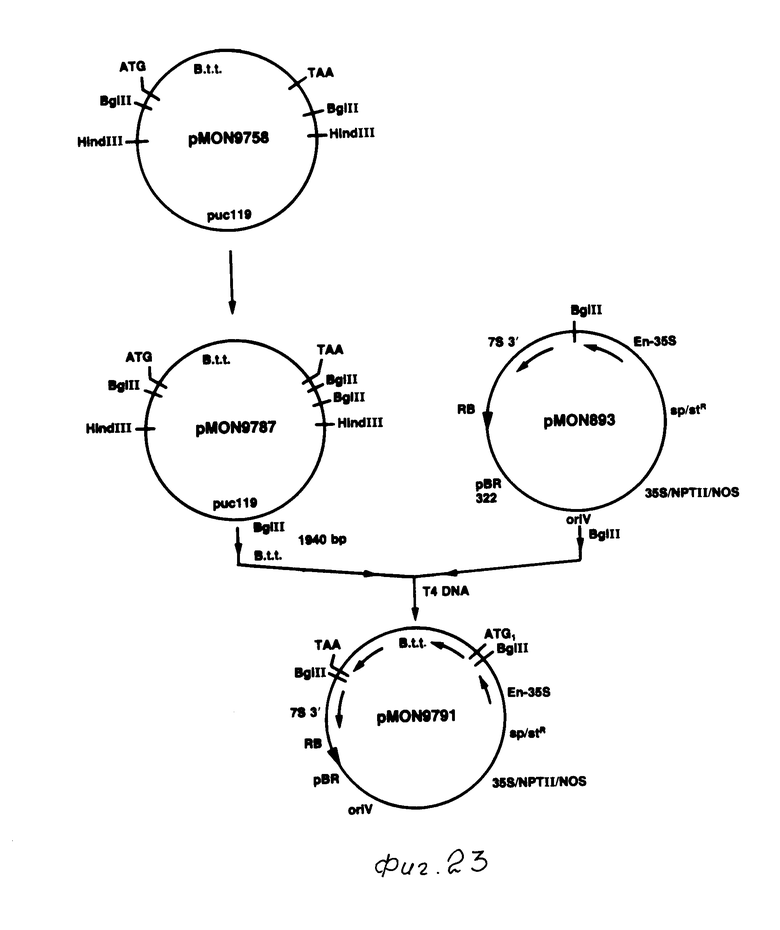
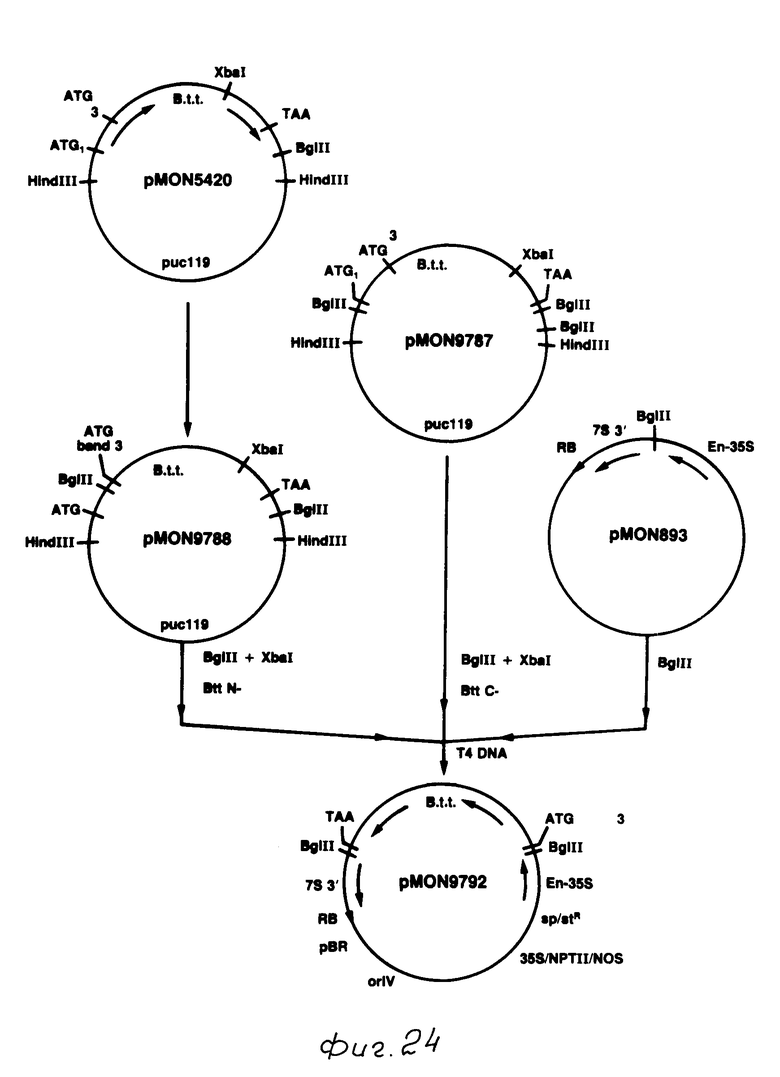
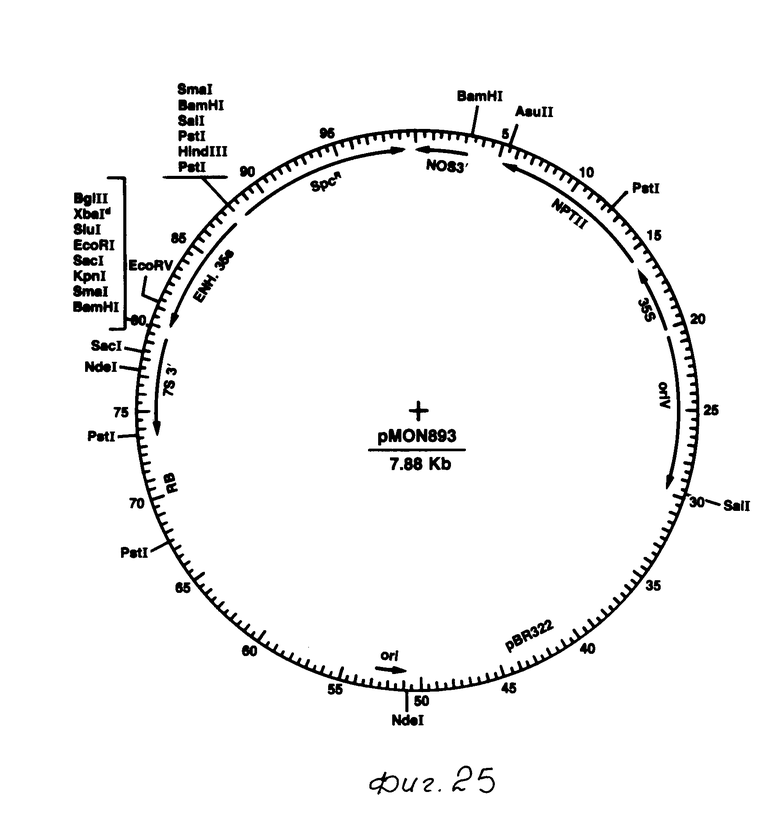
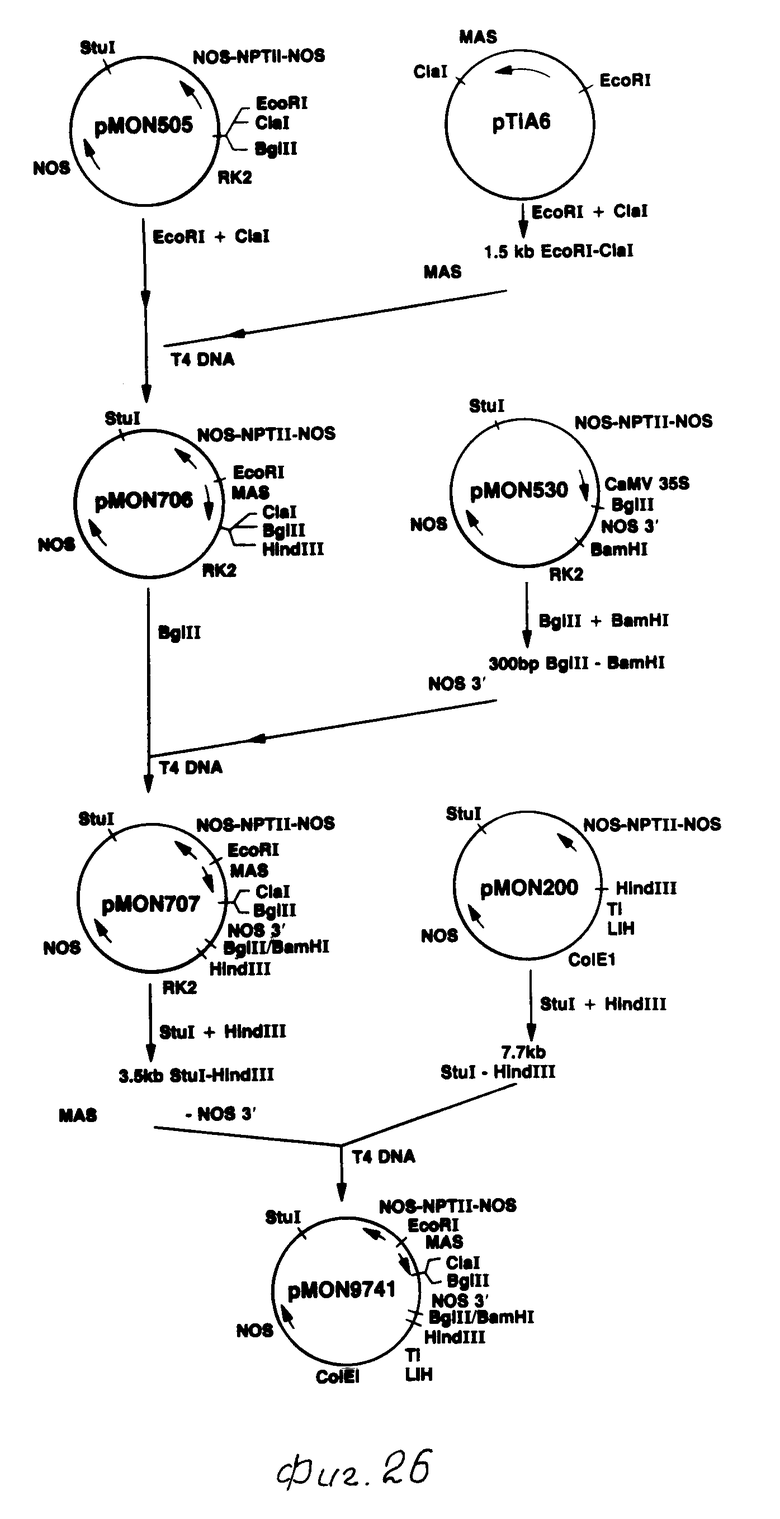
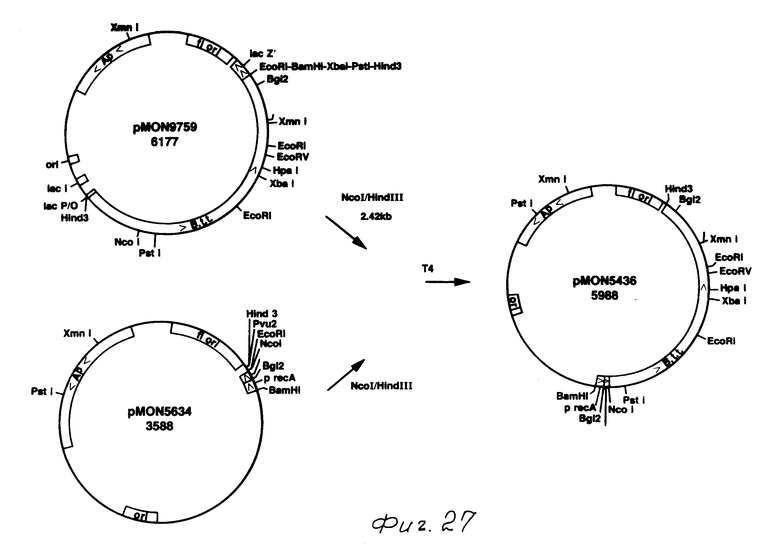

Использование: генетическая инженерия растений, получение устойчивых к насекомым растений. Сущность изобретения: конструируют рекомбинантную плазмидную ДНК, содержащую химерный ген, содержащий промотор CaMV35S или MAS, или SSRUBISCO, последовательность ДНК, кодирующую кристаллический белок B.thuringiensis, токсичный для жесткокрылых и 31 -нетранслируемую последовательность гена накоплении белка сои культурной. 29 ил., 6 табл.
СПОСОБ ПОЛУЧЕНИЯ РАСТЕНИЙ, УСТОЙЧИВЫХ К НАСЕКОМЫМ, предусматривающий конструирование рекомбинантной плазмидной ДНК, содержащей химерный ген, кодирующий токсин Bacillus thuringiensis, введение химерного гена в геном растения, получение трансформированных растительных клеток, культивирование клеток и получение регенерантов, отличающийся тем, что химерный ген содержит промотор CaMV35S, или MAS, или ssRUBISCO, последовательность ДНК, кодирующую кристаллический белок B. thuringiensis, токсичный для жесткокрылых, нуклеотидной последовательностью, приведенной в описании изобретения.
| СПОСОБ БЕСКАМЕРНОЙ ОЦЕНКИ ИСКРОБЕЗОПЛСНОСТИ ЭЛЕКТРИЧЕСКИХ ЦЕПЕЙ | 0 |
|
SU199259A1 |
| Способ гальванического снятия позолоты с серебряных изделий без заметного изменения их формы | 1923 |
|
SU12A1 |

