Изобретение направлено на решение задачи генетической трансформации молочной железы животного. Изобретение относится к генетической инженерии животных, экспериментальной и молекулярной биологии, биотехнологии, может быть использовано в экспериментальной медицине, ветеринарии и племенном животноводчестве.
Известно множество различных способов генетической трансформации клеток. Большинство из них относятся к трансформации клеток в культуре, либо к получению трансгенных животных. Так, например, на трансгенных животных была показана возможность получения чужеродных белков в молоке. Однако только немногие из способов генетической трансформации могут быть применены для соматических клеток в организме. Подходящими для генетической трансформации путем введения генетического материала в соматические клетки организма in vitro могут быть способы: с использованием липосом, с использованием вирусов, в состав генома которых встроена необходимая генетическая информация, и с использованием специальных ДНК-переносящих конструкций. Наиболее перспективным для генетической трансформации соматических клеток in vivo представляется использование растворимых ДНК-переносящих комплексов.
Известны, например, способы генетической трансформации in vitro с использованием носителей - конъюгатов трансферрин- полилизин, антитела-полилизин и т.д. В этих способах, как и в предлагаемом, имеется носитель - конъюгат, который состоит из поликатиона и лиганда, обеспечивающего связывание с клетками; этот конъюгат образует с генетическим материалом - нуклеиновыми кислотами растворимый комплекс; поглощение комплекса клетками in vitro происходит путем рецептор-опосредованного эндоцитоза. Однако известные способы - аналоги практически непригодны для генетической трансформации молочной железы из-за очень низкой эффективности трансфекции (< 0,1%).
В другом известном способе генетической трансформации в организм животного генетический материал вводят в растворимом комплексе с носителями, которые направленно доставляют в клетку генетический материал путем специфического рецептор-опосредованного эндоцитоза. Причем ген, кодирующий рецепторы клеточной поверхности вводится в организм животного при помощи растворимого комплекса, включающего в себя генетический материал и носитель, имеющий ген-связывающий участок и участок, узнающий клетку данного типа, введение гена производится внутривенно. Последнее является существенным недостатком при решении задачи генетической трансформации клеток молочной железы, т. к. : при внутривенном введении происходит контакт вводимого растворимого комплекса со многими тканями и жидкостями организма, что может, с одной стороны, приводить к повышенной биодеградации водорастворимого комплекса и, с другой стороны, вызывать различные побочные эффекты в тканях, и для создания действующей концентрации водорастворимого комплекса необходимо большое количество материала.
Технический результат предлагаемого способа состоит в устранении указанных недостатков, т.е. в повышении надежности, безопасности и экономичности способа за счет доставки генетического материала именно в ядра клеток-мишеней выбранного типа и создания высокой локальной концентрации используемого для генетической трансформации растворимого комплекса, что исключает риск нежелательных побочных реакций в тканях и обеспечивает экономию генетического материала.
Предлагаемый способ направленной генетической трансформации молочной железы животного позволяет направленно доставлять генетический материал в ядра клеток выбранного типа (клеток-мишеней), а также дает возможность создавать высокие локальные концентрации используемых для генетической трансформации растворимых комплексов. Для достижения указанного технического результата в способе направленной генетической трансформации молочной железы животного, включающем введение генетического материала в организм животного в растворимом комплексе с носителями и направленную доставку генетического материала в клетку носителями путем специфического рецептор-опосредованного эндоцитоза, используют носители, направленно доставляющие генетический материал также и в ядро клетки путем цитоплазменно-ядерного транспорта, причем растворимый комплекс генетического материала с носителями вводят в организм животного через молочные протоки молочной железы.
Изобретение решает задачу направленной генетической трансформации клеток молочной железы in vivo. Клеточная мембрана является трудноопределимым барьером для крупных гидрофильных молекул, какими являются молекулы генетического материала - ДНК. Для преодоления этого барьера используется специальный носитель, который состоит из лиганда(ов) и поликатиона(ов). Интернализуемый клеткой рецептор-специфичный лиганд способен обеспечить направленный транспорт генетического материала в составе искусственного растворимого молекулярного комплекса через наружную клеточную мембрану. Другой барьер, который необходимо преодолеть для эффективной генетической трансформации неделящихся и интерфазных клеток, - ядерная мембрана. Добавление в состав конструкции лиганда, обеспечивающего проникновение в ядро, позволяет решить эту проблему. Использование носителя, способного к цитоплазменно-ядерному транспорту, позволяет направленно доставлять генетический материал в тот клеточный компартмент, в котором и происходит обработка генетической информации. Очевидно, что введение растворимого комплекса носитель - генетический материал в молочные протоки позволяет локально создать ту же концентрацию комплекса за счет меньшего количества использования материала по сравнению с введением его в кровь. Специфичность и эффективность способа достигается, таким образом, за счет сочетания нескольких приемов: локального введения раствора комплекса при помощи специально разработанного устройства, использования искусственного растворимого молекулярного комплекса - носителя лиганд(ы) - поликатион(ы) с ДНК, где в качестве лигандов используются как лиганд, обеспечивающий клеточную специфичность и транспорт искусственной молекулярной конструкции внутрь клеток при помощи рецептор-опосредованного эндоцитоза, так и лиганд, обеспечивающий транспорт в клеточное ядро при помощи цитоплазменно-ядерного транспорта. Причем использование цитоплазменно-ядерного транспорта для доставки конструкции в ядро может достигаться путем связывания лиганда, находящегося в составе конструкции, с каким-либо клеточным компонентом, обеспечивающим ее транспорт в ядро. Также можно использовать лиганд, который, наряду с транспортом конструкции внутрь клетки путем рецептор-опосредованного эндоцитоза, обеспечивает и транспорт ее в ядро путем цитоплазменно-ядерного транспорта.
В качестве таких лигандов могут быть использованы, например, инсулин, пролактин, фактор роста эпидермиса и другие. При использовании лигандов двух типов, а именно лиганда, обеспечивающего рецептор-опосредодванный эндоцитоз (лиганд первого типа) и лиганда, обеспечивающего цитоплазменно-ядерный транспорт (лиганд второго типа), в качестве лигандов первого типа могут быть использованы также соматотропин, трансферрин и другие интернализуемые лиганды. В этом случае необходим и лиганд второго типа, обеспечивающий цитоплазменно-ядерный транспорт, например, пептиды с сигналом ядерной локализации, генно-инженерные белки с этими сигналами, нуклеоплазми и другие кариофильные белки.
Подходящим лигандом для решения задачи доставки искусственной молекулярной растворимой конструкции в клеточное ядро для трансформации in vivo может служить инсулин. Известно, что для этого гормона имеются рецепторы на поверхности клеток различных типов; эти рецепторы связывают инсулин и в комплексе с гормоном поглощаются клеткой путем рецептор-опосредованого эндоцитоза. Одним из компартментов, в которых накапливается инсулин после рецептор-опосредованного эндоцитоза, является ядро. Использование инсулина для генетической трансформации клеток молочной железы представляется оправданным, поскольку клетки из этой ткани обладают инсулиновыми рецепторами. Для специфической генетической трансформации клеток молочной железы наиболее перспективным представляется местное введение искусственной водорастворимой молекулярной конструкции, поскольку при этом снижается ее расход и достигается ее высокая локальная концентрация. Существенно, что при таком способе первым типом клеток, с которым взаимодействуют вводимые вещества, являются секреторные клетки эпителия, представляющие наибольший интерес с биотехнологической точки зрения. В качестве поликатиона, обеспечивающего обратимое связывание носителя с нуклеиновыми кислотами, может быть использован полилизин, синтетические поликатионы, катионизированные белки и другие.
Исходя из вышеизложенного, в качестве носителя генетического материала в предлагаемом способе используется ковалентно связанный с полилизином инсулин (конъюгат инсулин - полилизин). Синтез иинсулинированного полилизина заключается в следующем: проводят предварительное мечение инсулина 125I, предварительное цитраконирование инсулина, предварительное мечение полилизина [14C] -динитрофторбензолом либо [35S]-бутоксикарбонилметионином, предварительное растворение сшиваемых компонентов в гуанидине, титрование пиридиндитионированного инсулина полилизином с 10%-ной степенью модификации аминогрупп, интенсивное перемешивание во время титрования, сшивание инсулина с полилизином при помощи сукцинимидилпиридиндитиопропионата, очистку полученного вещества, снятие цитраконильных групп с инсулин-полилизина.
Известно устройство для введения растворов, например, лекарств и/или смазки в молочные протоки молочной железы животного, содержащее расширитель соска, состоящий из эластичных элементов, которые при соединении образуют наконечник, и упор, ограничивающий введение наконечника в сосок животного. Упор может иметь дренажный канал, а эластичные элементы могут служить держателем для лекарства (растворов вещества). Эластичные элементы могут деформироваться и сжиматься при введении расширителя и расширяться после введения, приспосабливаясь к различным размерам сосков.
Недостаток указанного устройства состоит в том, что введение расширителя в выходное отверстие канала соска осуществляется вручную, причем данный расширитель может быть применен лишь для крупного рогатого скота. Отыскать и попасть в отверстие канала соска, например, мелких млекопитающих, имеющих соски размером 1-3 мм, при помощи такого расширителя практически невозможно. Кроме того при введении такого расширителя в молочный проток неизбежно происходит деформация не только ткани соска, но и профиля главного протока молочной железы животного, в который необходимо ввести расширитель, а упор ограничивает введение расширителя в сосок на постоянную глубину.
Наиболее близким к заявляемому устройству по совокупности существенных признаков является устройство по авторскому свидетельству СССР N 1123598, 1984, содержащее корпус, выполненный в виде доильного стакана, с полостью по формe соска животного, каналом для подачи отрицательного давления и плоским дном, вдоль оси которого выполнено сквозное сливное отверстие, а на плоском дне имеется расширитель сфинктера соска в виде кольцевого буртика, центр которого расположен на продольной оси стакана. Сосок животного вставляют в полость стакана, причем между вершиной соска и плоским дном гильзы имеется зазор. В канал подают постоянный вакуум и устройство фиксируют в области основания соска. Затем под действием переменного вакуума, подаваемого через сливное отверстие, под соском создается разрежение, сосок удлиняется, упираясь в кольцевой буртик, и сфинктер (выходное отверстие) раскрывается.
Несмотря на то, что устройство не содержит элементов для введения веществ в проток молочной железы, по своей технической сущности оно может быть использовано для введения растворов (например, веществ, применяемых для генетической трансформации) в проток молочной железы, например, путем впрыскивания через сливное отверстие в момент раскрытия сфинктера соска.
Однако это устройство обладает существенными недостатками, а именно, расширение сфинктера соска (отверстия протока молочной железы) животного происходит за счет увеличения объема ткани соска при подаче вакуума (при разрежении под соском), когда сосок удлиняется и увеличивается в объеме, заполняя весь объем стакана, полость которого (т.е. выемка, в которую помещается сосок), имеет большие размеры, чем сосок, и при разрежении ткань соска деформируется в сторону увеличения своего объема до тех пор, пока сосок не упрется в кольцевой буртик и не раскроется сфинктер соска.
Технический результат, достигаемый предлагаемым устройством состоит в устранении указанных недостатков, т.е. повышении надежности, безопасности и экономичности устройства за счет снижения степени деформирования соска животного, обеспечения возможности введения растворов веществ в проток молочной железы млекопитающих независимо от размеров соска, а также обеспечения точного дозирования вводимого раствора. Для достижения указанного технического результата устройство содержит корпус с выемкой по форме соска животного, канал для подачи отрицательного давления, сквозной осевой канал на дне выемки и расширитель сфинктера соска; оно содержит также канюлю с мерной полостью для дозированного объема растворимого комплекса генетического материала с носителями, причем выемка выполнена с обеспечением охвата ею соска без зазоров, диаметр сквозного осевого канала больше диаметра канюли, а расширитель сфинктера соска выполнен в виде кольцевой канавки, расположенной на внутренней поверхности дна выемки по периметру сквозного осевого канала, при этом кольцевая канавка соединена в ее углублении с каналом для подачи отрицательного давления. Канюля может быть выполнена в виде стеклянной пипетки, на острие которой сделан скос под углом 30-45 градусов с оплавленными кромками, а выемка с кольцевой канавкой, сквозным осевым каналом и каналом для подачи отрицательного давления могут быть заполнены изотоническим раствором с целью повышения травмобезопасности устройства.
Изобретение поясняется чертежами, где на фиг. 1 показана схема устройства для введения генетического материала в молочный проток молочной железы животного; на фиг. 2 - очистка носителя инсулин-полилизина при помощи гель-фильтрации на колонке с сефакрилом S-200; на фиг. 3 - зависимость растворимости комплекса носитель - генетический материал (инсулин-полилизин-плазмида) от молярного соотношения в них мономеров лизин/нуклеотид; на фиг. 4 - зависимость количества легкой и тяжелой фракций плазмидной ДНК в составе комплексов инсулин-полилизин-плазмида от молярного соотношения мономеров лизин/нуклеотид в комплексе при аналитическом ультрацентрифугировании; на фиг. 5 - зависимость констант седиментации легкой и тяжелой составляющих легкой фракции плазмидной ДНК в составе комплексов инсулин-полилизин-плазмида от молярного соотношения мономеров лизин/нуклеотид в растворимом комплексе при аналитическом ультрацентрифугировании; на фиг. 6 - вытеснение специфически связанного с клетками гепатомы линии PLC/PRF/5 [125I] инсулина растворимым комплексом носитель - генетический материал (инсулин-полилизин-плазмида рSVRP8), а также немеченым инсулином; на фиг. 7 - визуализация связывания растворимого комплекса носитель - генетический материал (флуоресцентно меченный конъюгат инсулин-полилизин - ДНК) с клетками линии PLC/PRF/5 при помощи конфокальной лазерной сканирующей микроскопии; на фиг. 8 - связывание растворимого комплекса носитель - генетический материал с клетками линии PLC/PRF/5; ФИТЦ-меченный комплекс инсулин-полилизин-ДНК на поверхности клеток, измерено при помощи конфокальной лазерной сканирующей микроскопии; на фиг. 9 - визуализация рецептор-опосредованного эндоцитоза растворимого комплекса носитель-генетический материал (флуоресцентно меченный конъюгат инсулин-полилизин-ДНК) с клетками линии PLC/PFR/5 при помощи видеоинтенсификационной микроскопии; на фиг. 10 - рецептор-опосредованный эндоцитоз растворимого комплекса носитель - генетический материал в клетках линии PLC/PRF/5; накопление ФИТЦ-меченного комплекса инсулин-полилизин-ДНК в цитоплазме, измеренное при помощи конфокальной лазерной сканирующей микроскопии; на фиг. 11 - кинетика транспорта носителя-конъюгата инсулин-полилизин в ядра клеток PLC/PRF/5; на фиг. 12 - цитоплазменно-ядерный транспорт растворимого комплекса носитель-генетический материал в клетках линии PLC/PRF/5; накопление флуоресцентно меченного комплекса инсулин-полилизин-ДНК, измеренное при помощи конфокальной лазерной сканирующей микроскопии; на фиг. 13 - направленная генетическая трансформация клеток в культуре, измеренная по эффективности экспрессии большого Т-антигена вируса SV40 в клетках гепатомы человека линии PLC/PRF/5, трансформированных комплексом инсулин-полилизин-плазмида pSVRP8; на фиг. 14 - генетическая трансформация секреторных клеток молочной железы в культуре геном светлячковой люциферазы при помощи направленной доставки растворимых комплексов инсулин-полилизин-плазмидная ДНК рRSVL; на фиг. 15 - генетическая трансформация молочной железы мышей in vivo.
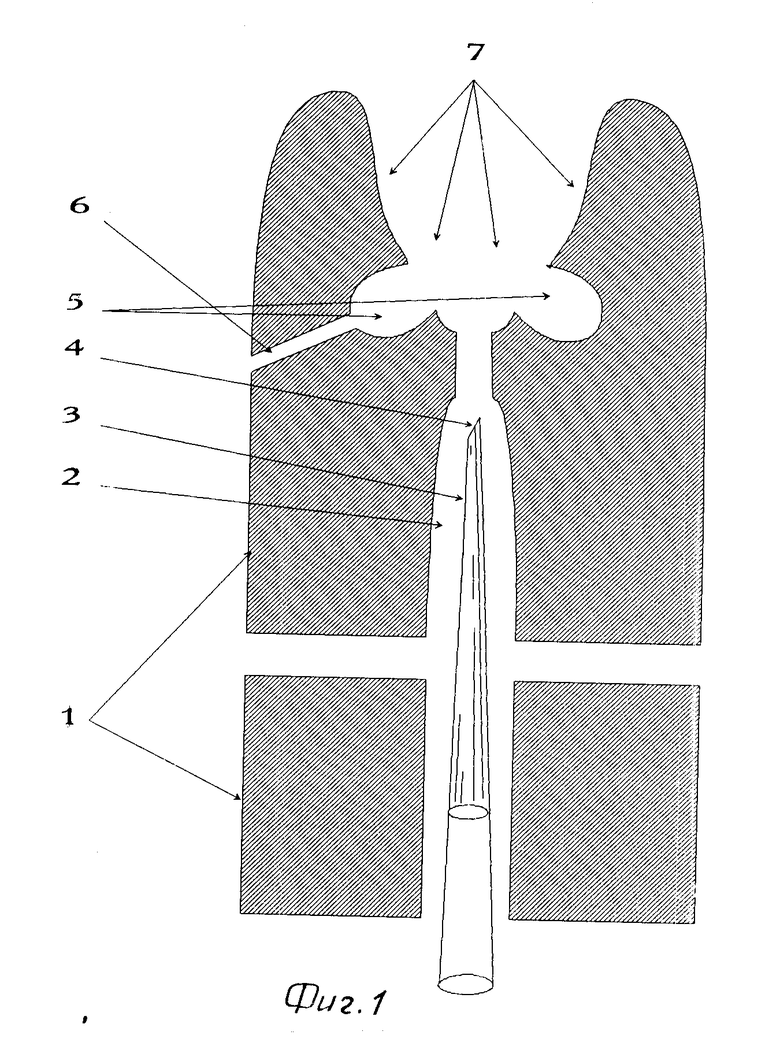
Устройство для введения генетического материала в молочный проток молочной железы животного содержит корпус 1, со сквозным осевым каналом 2 для канюли 3 с мерной полостью для дозированного объема растворимого комплекса генетического материала с носителем, причем рабочая часть 4 канюли 3 скошена на 30-45 градусов и оплавлена, по периметру сквозного осевого канала выполнена кольцевая канавка 5, соединенная с каналом 6 для присоса (подачи отрицательного давления). В корпусе выполнена выемка 7 по форме соска животного с обеспечением охвата соска без зазоров.
Устройство работает следующим образом. Выемку 7 корпуса 1 помещают на сосок молочной железы животного, прижимая торец корпуса к телу животного, затем через канал 6 подают отрицательное давление для обеспечения присоса соска. Поверхность ткани соска, присасываясь в кольцевой канавке 5, растягивается в области сфинктера соска раскрывая вход в молочный проток и фиксируется относительно сквозного осевого канала 2 корпуса 1. В канал 2 вводят канюлю 3, предварительно заполненную комплексом генетического материала с носителями. Затем рабочую часть кончика 4 канюли вводят в проход сфинктера соска и производят инъекцию дозированного объема комплекса в проток молочной железы. Выемка 7 в корпусе 1 в сочетании с кольцевой канавкой 5 обеспечивает установку соска молочной железы для облегченного введения канюли в выходное отверстие протока, так как при этом присос кольцевой канавкой дает возможность с одной стороны, растянуть центральную часть соска, свободную от занимаемой кольцевой канавкой площади, что обеспечивает расширение выходного отверстия протока, с другой, не дает возможности значительно деформироваться как ткани соска, так и главному протоку молочной железы во время введения в него канюли, так как сосок фиксируется вокруг сфинктера - места введения канюли. Кроме того, диаметр сквозного канала больше диаметра канюли для облегчения попадания канюлей в отверстие сфинкера, а оплавленная рабочая часть стеклянной канюли при введении ее в проток не повреждает эпителий протоков молочной железы.
Таким образом, данное устройство позволяет значительно упростить и ускорить процесс введения растворов веществ в проток молочной железы, обеспечивая при этом высокую сохранность биологического объекта, то есть снижая повреждение и деформацию соска и, что особенно важно, исключает необходимость применения дополнительных манипуляций практически для всех видов млекопитающих животных, начиная от мелких грызунов, и независимо от состояния животного, а именно - находится или не находится оно в состоянии лактации. Все это определяет положительный эффект от применения данного изобретения.
Способ направленной генетической трансформации молочной железы животного с использованием устройства для введения генетического материала в молочный проток молочной железы животного осуществляется следующим образом. Синтезируют носитель генетического материала - инсулинированный полилизин описанным выше образом. Синтезированный носитель может храниться в замороженном виде при -20оС в течение нескольких месяцев. Проводят образование растворимого комплекса с выбранным генетическим материалом. Полученный комплекс переводят в раствор Хэнкса с рН 7,4. Комплекс в растворе Хэнкса заправляют в устройство для введения генетического материала в молочный проток молочной железы. Комплекс в растворе при помощи этого устройства вводят в молочный проток животного.
Возможность осуществления способа направленной генетической трансформации молочной железы животного с использованием устройства для введения генетического материала в молочный проток молочной железы животного подтверждается следующими примерами.
П р и м е р 1. Синтез и очистка носителя генетического материала - инсулинированного полилизина.
Предварительно перед синтезом носителя бычий инсулин (Sigma) был иодирован 125I при помощи идогена в буфере 1 (50 мМ НЕРЕS (Sigma), рН 7,65 при 20оС, 150 мМ NaCl). Очистку [125I]-инсулина проводили гель-фильтрацией на колонке с сефадексом G-25 в буфере 1. Радиоактивность определяли на гамма-счетчике Ria Gamma 1271 (LKB, Швеция).
Поли-L-лизин (90 кД) (Sigma) предварительно перед синтезом был промечен [14C]-динитрофторбензолом (Amersham). Мечение проводили в 0,5 М натрий-фосфатном буфере при рН 9,5. Меченый полилизин очищали диализом против буфера 1. Радиоактивность полученных препаратов определяли на жидкостном сцинтилляционом счетчике Rack Beta 1217 (LKB, Швеция).
Бычий [125I]-инсулин был ковалентно присоединен к [14C]-полилизину при помощи бифункцонального сшивающего реагента сукцинимидил 3-(2-пиридилдитио)пропионата (SPDP) (Sigma) по схеме Юнга. Перед модификацией N-концевые аминогруппы инсулина были защищены цитракониевым ангидридом (Serva). К 20 мл (40 мг) бычьего радиоактивно меченного инсулина в 25 мМ ВES, рН 6,9 при 20оС, было добавлено при непрерывном перемешивании 6,6 мкл цитракониевого ангидрида. После инкубации при контролируемом рН 6,9 в течение 1 часа при 20оС инсулин диализовали против буфера 1. Далее к инсулину было добавлено 500 мкл 40 мМ SPDP в метаноле. После тридцатиминутной инкубации при 23оС образец также диализовали против буфера 1. Степень модификации инсулина контролировали спектрофотометрически по появлению продукта реакции восстановления - пиридин-2-тиона.
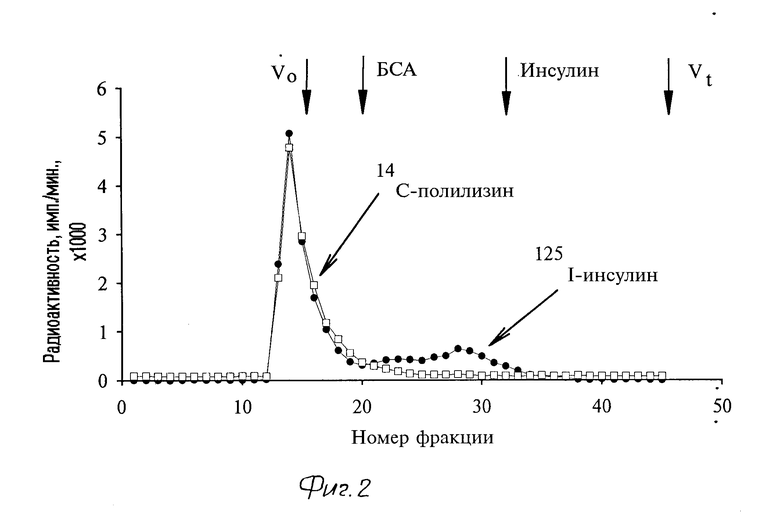
Полилизин был модифицирован сульфгидрильными группами при помощи SPDP. К 1050 мкл (28,3 мг) радиоактивно меченного полилизина в буфере 1 было добавлено 326 мкл 40 мМ SPDP в метаноле. После сорокапятиминутной инкубации при 25оС модифицированный полилизин диализовали против буфера 1. После диализа дитиопиридинильные остатки полилизина были восстановлены до SH-групп добавлением дитиотреита в конечной концентрации 50 мМ. После тридцатиминутной инкубации при 37оС материал был отдиализован против буфера 1. Степень модификации полилизина SPDP определяли перед диализом так же, как и в случае инсулина. При описанных условиях полилизин имел 10% сульфгидрильных групп. Перед реакцией сшивания инсулина и полилизина оба компонента переводили в буфер 2 (50 мМ HEPES рН 7,5 при 20оС, 150 мМ NaCl,2 M гуанидиин (Merck). Для получения конъюгата к 6280 мкл (9,7 мг) пиридиндитионированного инсулина было добавлено по каплям при интенсивном перемешивании 1356 мкл (9,6 мг) SH-полилизина. Конъюгацию проводили в буфере 2 в течение 18 ч при 4оС. Взаимодействие дитиопиридинильных остатков инсулина с полученными после восстановления дитиопиридильных остатков полилизина сульфгидрильными группами контролировали спектрофотометрически, как описано выше. Полученный конъюгат был очищен гель-фильтрацией на колонке с сефакрилом S-200 (Pharmacia) в буфере 2 (см. фиг. 2). Цитраконильные группы инсулина (в составе конъюгата) были удалены. К очищенному конъюгату в буфере 2 была добавлена 1 М НСl до рН 2,0. После четырехчасовой инкубации при 18оС (при контролируемом рН 2,0) добавлением 1 М NaOH pH раствора был резко доведен до 9,3. Далее конъюгат инсулин-полилизин был 3 раза отдиализован против 0,15 М NaCl и сконцентрирован при помощи ультрафильтрации. Молярное содержание инсулина и полилизина в конъюгате было рассчитано по соотношению в нем радиоактивных иода и углерода и составило от 8 до 10 остатков инсулина на 1 полилизин в различных партиях синтезированного препарата.
Примененный метод обеспечивает эффективное присоединение инсулина к полилизину о чем свидетельствуют результаты гель-фильтрации конъюгата (фиг. 2). Меченый инсулин и меченый полилизин выходят с колонки в одних и тех же фракциях. При реакции присоединяется до 80% участвующего в реакции инсулина и выход получаемого вещества составляет около 50%.
П р и м е р 2. Приготовление растворимого комплекса носитель-ДНК.
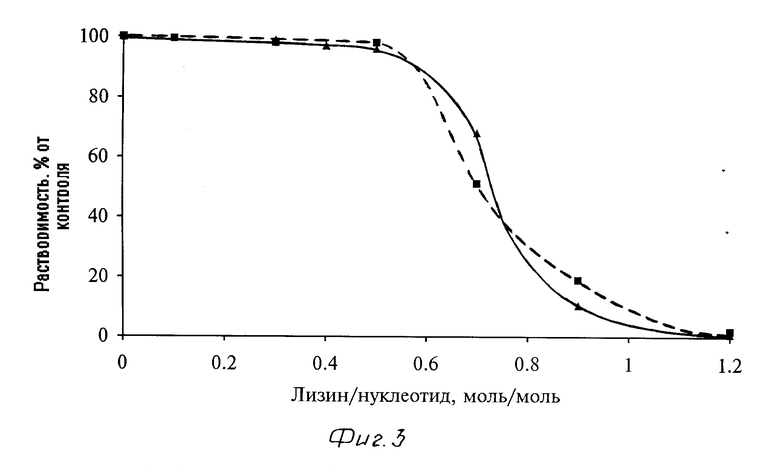
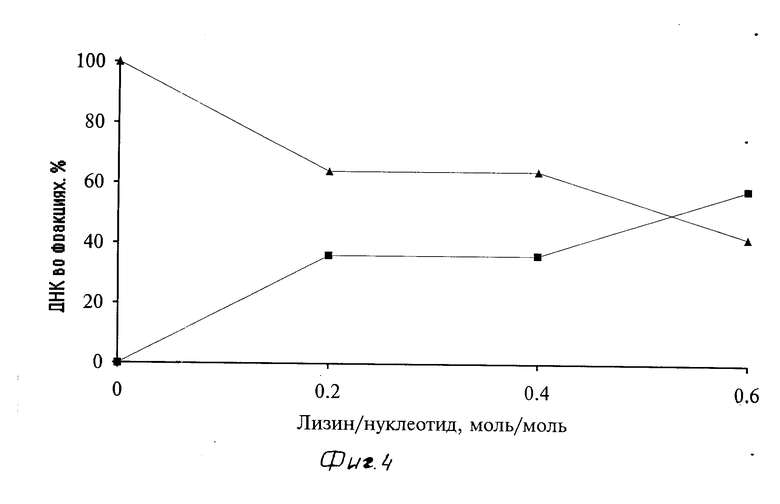
Кривые зависимости растворимости различных комплексов носитель - ДНК (инсулин-полилизин-плазмидная ДНК) от соотношения лизин/нуклеотид в комплексе представлены на фиг. 3. Растворимость комплексов инсулин-полилизин-плазмидная ДНК практически не зависит (см. фиг. 3) от типа и размера используемой в их составе плазмиды. Так, полученные комплексы с плазмидами рSVRP8 (5,9 тысяч пар оснований) и рSV3neo (8,6 тысяч пар оснований) были растворимы вплоть до молярного соотношения мономеров лизин/нуклеотид, равного 0,5. Формирование растворимых комплексов инсулин-полилизин-плазмида контролировали электрофорезом в агарозном геле и аналитическим ультрацентрифугированием. При электрофорезе (в 1%-ном агарозном геле) с увеличением соотношения лизин/нуклеотид (от 0,1 до 0,5) возрастала доля плазмидной ДНК в составе комплекса инсулин-полилизин-плазмида pSVRP8, остающейся на старте в виде неподвижной фракции ДНК. Аналогичные результаты были получены и при аналитическом ультрацентрифугировании растворимых комплексов инсулин-полилизин-плазмида pSV3neo с различным молярным соотношением мономеров лизин/нуклеотид (см. фиг. 4 и 5). Как следует из профиля седиментации, комплексы содержат легкую фракцию ДНК (состоящую из двух компонентов) с вариабельной константой седиментации 14-19 S (в зависимости от соотношения лизин/нуклеотид), а также тяжелую фракцию ДНК (растворимые микроорганизмы, которая осаждается при ультрацентрифугировании на дно пробирки. С увеличением соотношения лизин/нуклеотид (см. фиг. 4) снижается доля легкой фракции и, соответственно, увеличивается доля тяжелой фракции плазмидной ДНК в составе растворимого комплекса. В свою очередь (см. фиг. 5), с возрастанием соотношения лизин/нуклеотид (от 0,2 до 0,6) также увеличиваются и константы седиментации легкой и тяжелой составляющих легкой фракции плазмидной ДНК.
Приведенные данные позволяют утверждать, что синтезируемый носитель способен образовывать растворимые комплексы с генетическим материалом, а именно с плазмидой ДНК.
П р и м е р 3. Проверка рецептор-связывающих свойств растворимого комплекса носитель-ДНК.
Клетки РLC/PRF/5 имеют один тип рецепторов, связывающих инсулин, с константой диссоциации комплекса инсулин-рецептор - 3,7 нМ, и их количество в расчете на одну клетку - 160 000, что делает их удобным объектом для изучения рецептор-связывающих свойств растворимого комплекса носитель-генетический материал (инсулин-полилизин-плазмида).
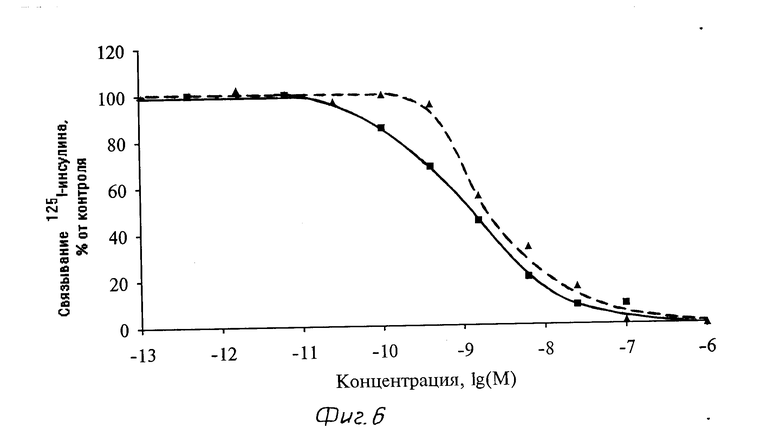
Средство комплексов инсулин-полилизин-плазмида pSVRP8 к инсулиновым рецепторам клеток линии PLC/PRF/5 определяли с помощью конкурентного радиолигандного анализа. Клетки культивировали в сред RPMI-1640 с 10%-ной фетальной телячьей сыворотки. Для определения связывания клетки инкубировали в течение 18 ч при 0оС с [125I]-инсулином (в концентрации 0,3-0,5 нМ) и с возрастающими концентрациями комплексов (здесь и далее концентрация дается в расчете на ДНК) или немеченого инсулина. Растворимые комплексы инсулин-полилизин-плазмида (соотношение мономеров лизин/нуклеотид - 0,4) эффективно конкурировали с радиоактивно меченным лигандом за инсулиновые рецепторы клеток гепатомы (см. фиг. 6) с таким же сродством, как и немеченый инсулин, так как кривые вытеснения в обоих случаях практически совпадали. Аналогичные результаты были получены и для комплексов носителя с другой плазмидой - pSV3neo. Полученные данные свидетельствуют о том, что инсулин в составе растворимого комплекса инсулин-полилизин-плазмидная ДНК сохраняет способность связываться с инсулиновыми рецепторами на поверхности клеток.
Визуализация связывания растворимых комплексов носитель - генетический материал была проведена при помощи конфокальной лазерной сканирующей микроскопии. Носитель - конъюгат предварительно был промечен флуоресцеинизотиоцианатом (ФИТЦ).
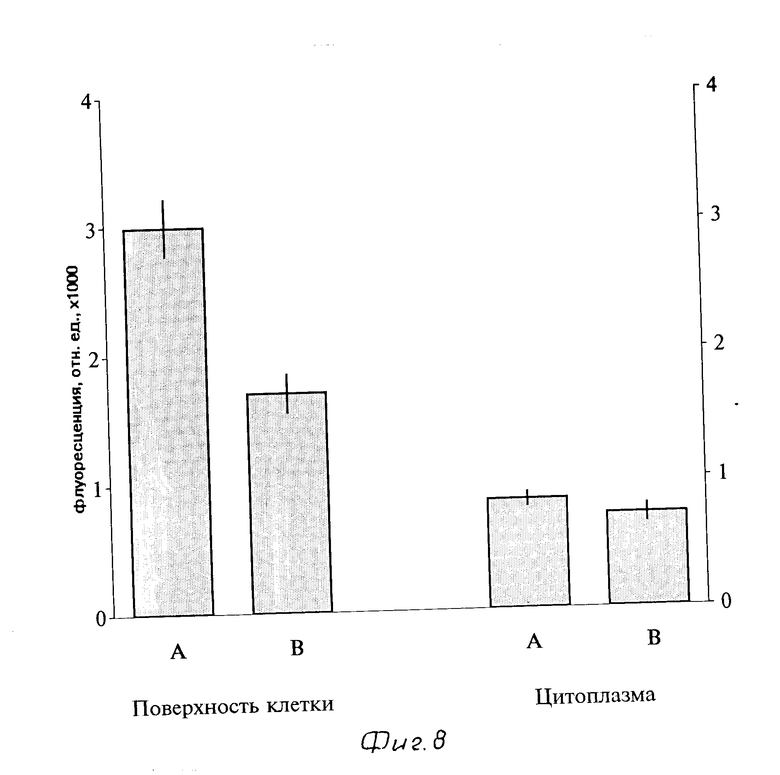
Спустя 7 мин после добавления комплексов в концентрации 3,7 нМ к клеткам РLC/PRF/5 (при температуре 21оС), были обнаружены ярко флуоресцирующие участки на поверхности клеток (фиг. 7). В присутствии 10 мкМ свободного инсулина средняя интенсивность флуоресценции этих ярких участков на поверхности клеток была значительно меньше по сравнению с клетками, инкубированными только с ФИТЦ-меченными комплексами (фиг. 8). В то же время не было выявлено достоверных различий во флуоресценции цитоплазмы клеток, инкубированных с растворимыми комплексами с или без избытка свободного инсулина (фиг. 8), т.е. при данных условиях еще не регистрируется рецептор-опосредованого эндоцитоза.
П р и м е р 4. Рецептор-опосредованный эндоцитоз растворимого комплекса носитель - генетический материал.
Для изучения рецептор-опосредованного эндоцитоза была проведена видеоинтенсификационная микроскопия клеток линии PLC/PRF/5 после поглощения флуоресцеин-меченной конструкции инсулин-полилизин-плазмида pSVRP8. Инсулин-полилизин был предварительно помечен ФИТЦ.
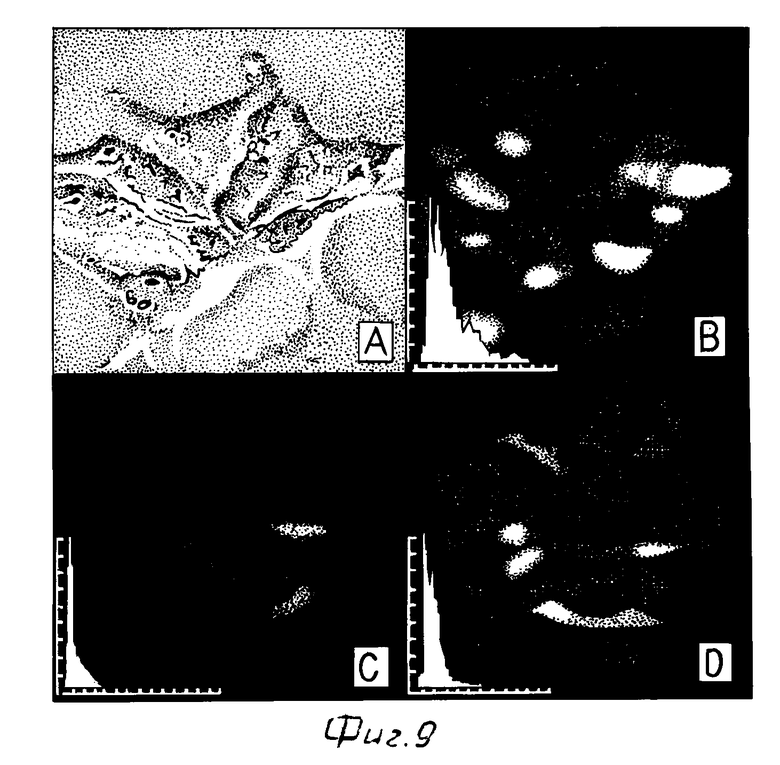
Клетки PLC/PRF/5 инкубировали с 7 нМ растворимым флуоресцентно меченным комплексом инсулин-полилизин-плазмида в течение 2 ч пр 37оС. Непосредственно перед регистрацией (за 10 мин) в среду был добавлен хлорид аммония (в конечной концентрации 10 мМ), и при помощи видеомикроскопии были получены флуоресцентные изображения клеток, накопленные и обработанные затем на анализаторе изображений (см. фиг. 9). Результаты экспериментов показывают, что ФИТЦ-меченный комплекс аккумулируется в клетках: так, флуоресценция цитоплазмы после инкубации с комплексом, меченным ФИТЦ (фиг. 9 В), была значительно ярче, чем в случае клеток, инкубированных с комплексами в присутствии избытка свободного инсулина (фиг. 9 Д), или же в клетках, инкубированных без такого комплекса (автофлуоресценция; фиг. 9 С). Частота распределения интенсивности флуоресценции цитоплазмы клеток, представленная на графиках (левый нижний угол на фиг. 9 В-Д), также подтверждает этот вывод. В клетках, инкубированных с ФИТЦ-меченными комплексами в присутствии избытка свободного инсулина (фиг. 9 Д), наблюдались также и более интенсивно флуоресцирующие, чем в случае автофлуоресценции (фиг. 9 С), отдельные участки цитоплазмы. Однако, интенсивность флуоресценции этих ярких участков цитоплазмы значительно ниже по сравнению с таковыми в клетках, инкубированных только с комплексами (фиг. 9 В).
Для получения более детальных и количественных данных о локализации в клетке комплексы инсулин-полилизин-плазмида клетки линии PLC/PRF/5 исследовали при помощи лазерной сканирующей конфокальной микроскопии.
В исследованиях по интернализации клеток инкубировали с 3,7 нМ флуоресцентно меченными комплексами при 37оС. После двухчасовой инкубации клеток в этих условиях наблюдалось специфическое (блокируемое свободным инсулином) накопление ФИТЦ-меченных комплексов инсулин-полилизин-плазмида внутри клеток (фиг. 10). Флуоресцентно меченные комплексы накапливались в околоядерных участках, интенсивность свечения которых почти втрое превышала уровень автофлуоресценции клеток, что свидетельствует об эндоцитозе комплексов носитель - генетический материал. Интенсивность флуоресценции околоядерных участков также была в 1,6 раза выше для клеток, инкубированных с комплексами в отсутствие (1703±165 относительных единиц), чем в присутствии избытка свободного инсулина (1062±46 отн. ед., достоверность различия р < 0,001). Таким образом, на основании вышеприведенных данных можно утверждать, что растворимые комплексы носитель - генетический материал могут поглощаться клетками путем специфического рецептор-опосредованного эндоцитоза.
Добавление хлорида аммония в среду, в конечной концентрации 10 мМ, за 10 мин до начала измерения методом лазерной сканирующей микроскопии заметно увеличивало флуоресценцию поглощенной комплексов в клетках. Это указывает на то, что после поглощения комплексы оказываются в кислых компартментах клеток. Добавление избытка свободного инсулина также существенно снижало интенсивность флуоресценции, измеренной в аналогичных условиях (фиг. 10, живые клетки). С этими результатами сходны данные, полученные на препаратах клеток, фиксированных р-формальдегидом (фиг. 10, фиксированные клетки). Полученные результаты позволяют сделать вывод, что ФИТЦ-меченные комплексы инсулин-полилизин-плазмидная ДНК поступают в клетки PLC/PRF/5 рецептор-опросредованным эндоцитозом через инсулиновые рецепторы и, как следствие, специфически транспортируются в кислые внутриклеточные компартменты.
П р и м е р 5. Цитоплазменно-ядерный транспорт носителя.
Проверка способности синтезированного нами носителя - конъюгата инсулин-полилизин к цитоплазменно-ядерному транспорту была проведена методом восстановления флуоресценции после фотоотбеливания. Метод позволяет количественно охарактеризовывать транспорт флуоуресцентно-меченных молекул в ядра живых клеток. Носитель - инсулин-полилизин был предварительно промечен ФИТЦ. Клетки линии PLC/PRF/5 были проинкубированы с 4 мМ флуоресцентно меченного конъюгата инсулин-полилизин 4 ч и затем подвергались исследованию на установке по измерению восстановления флуоресценции после фотоотбеливания. На фиг. 11 приведена кинетика транспорта конъюгата ФИТЦ-инсулин-полилизин в ядра клеток PLC/PRF/5. Проведенные измерения показали наличие транспорта носителя - конъюгата инсулин-полилизин в ядра клеток-мишеней. По результатам экспериментов по методу Петерса была рассчитана константа транспорта носителя в ядро, она оказалась равна 0,035±0,006 с-1. Таким образом, синтезированный носитель способен проникать в ядра клеток при помощи цитоплазменно-ядерного транспорта.
П р и м е р 6. Направленная доставка растворимых комплексов носитель - генетический материал (инсулин-полилизин-плазмида) в ядро.
Эксперименты, проведенные с растворимым комплексом носитель-генетический материал (флуоресцентно меченный конъюгат инсулин-полилизин-плазмидная ДНК), методом конфокальной лазерной сканирующей микроскопии на клетках линии PLC/PRF/5 показали наличие комплексов в ядре. Так, после двухчасовой инкубации клеток при 37оС с 3,7 нМ растворимого комплекса инсулин-полилизин-плазмида pSVRP8 наблюдается достоверное увеличение флуоресценции ФИТЦ-меченного комплекса в ядре (фиг. 12, живые клетки). Сходный результат был получен и на препаратах фиксированных клеток (фиг. 12). В клетках, которые были инкубированы с растворимым комплексом в присутствии избытка свободного инсулина, наблюдается меньшее накопление флуоресцентно-меченного комплекса в ядрах. После 4 ч флуоресценция комплексов, наблюдаемая в ядре, увеличивается.
П р и м е р 7. Проверка возможности генетической трансформации клеток при помощи растворимых комплексов носитель - генетический материал (инсулин-полилизин-плазмида).
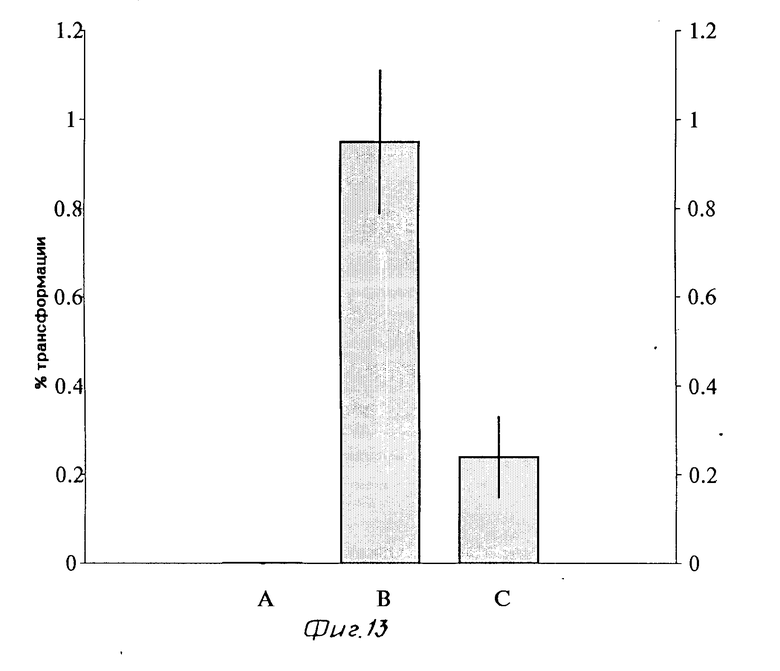
Клетки линии PLC/PRF/5, отмытые от сыворотки, инкубировали в течение 18 ч при 37оС с растворимыми комплексами инсулин-полилизин-плазмида pSVRP8. После фиксации и прокрашивания методом непрямой иммунофлуоресценции (первичные антитела против белка большого Т-антигена вируса SV-40 и флуоресцентно меченные вторичные антитела) были обнаружены клетки с ярко флуоресцирующими ядрами (т.к. продукт экспрессии является кариофильным белком), что указывает на высокий уровень экспрессии гена большого Т-антигена в этих клетках. Такие экспрессирующие Т-антиген клетки совершенно отсутствовали среди контрольных клеток, ядра которых не флуоресцировали. Эффективность экспрессии в случае инкубации клеток гепатомы с конструкцией составляла 0,95±0,16% (среднее±стандартная ошибка для трех репрезентативных серий экспериментов, в каждом из которых было просчитано около 15000 клеток - см. фиг. 13, В). Свободный инсулин, добавленный к клеткам вместе с растворимыми комплексами, достоверно снижал количество клеток, экспрессирующих Т-антиген, до 0,24±0,09% (р < 0,001 по сравнению с клетками, инкубированными только с конструкцией). Это снижение эффективности экспрессии при добавлении избытка свободного инсулина на 75% полностью согласуется с остаточным связыванием (10-30% ), наблюдаемым в экспериментах по связыванию [125I]-инсулина с клетками PLC/PRF/5 в аналогичных условиях. В клетках, инкубированных с плазмидой pSVRP8 без добавления носителя, не было обнаружено экспрессии большого Т-антигена (см. фиг. 13, А).
П р и м е р 8. Проверка возможности генетической трансформации секреторных клеток молочной железы при помощи растворимых комплексов носитель - генетический материал (инсулин-полилизиин-плазмида).
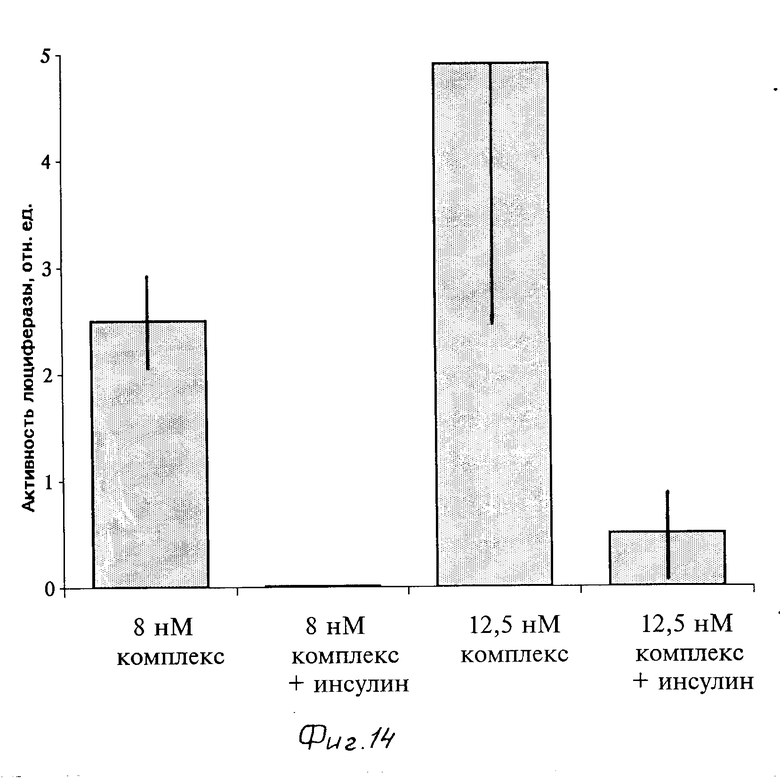
Эпителиальные секретирующие клетки молочной железы мышей линии НС-11 были подвергнуты генетической трансформации растворимыми комплексами носитель-ДНК (инсулин-полилизин-плазмида pRSVL). Данная плазмида имеет в своем составе ген светлячковой люциферазы, экспрессию которой легко определить в клетках. Клетки линии НС-11 имеют по нашим данным около 3000 инсулиновых рецепторов на клетку и константой диссоциации комплекса инсулин-рецептор около 4 нМ, что делает их пригодными для генетической трансформации растворимым комплексом носитель - генетический материал. В качестве носителя мы использовали конъюгат инсулин-полилизин, синтез которого описан в примере 1. Клетки были посеяны на 6-см чашки Петри (Costar) в среде RPMI-1640 с 10% фетальной телячьей сыворотки, их инкубировали в течение двух суток при 37оС и 5% СО2, после чего отмывали от среды и 3 ч инкубировали в среде RPMI-1640 без сыворотки. Растворимый комплекс носитель - генетический материал, приготовленный как описано в примере 2, был добавлен к клеткам в объеме 0,5 мл в среде RPMI-1640 с 25 мМ НЕРЕS, 2 мг/мл БСА рН 7,4. Клетки были проинкубированы с комплексами в течение 18 ч при 37оС, после чего к ним было добавлено 5 мл среды RPМI-1640 с 5% фетальной телячьей сыворотки. По прошествии 1 сут клетки были сняты с поверхности чашек и люциферазная активность измерена в клеточных экстрактах на люминометре фирмы "Рharmacia-LKB", модель 1250. Проведенные эксперименты показали, что секреторные клетки молочной железы могут быть генетически трансформированы при помощи растворимых комплексов носитель - генетический материал (фиг. 14). Избыток свободного инсулина (1,6 мкМ), добавленного к клеткам одновременно с растворимыми комплексами, снижал эффективность генетической трансформации примерно на порядок (фиг. 14). Таким образом можно утверждать, что клетки молочной железы в культуре могут быть специфически трасформированы растворимыми комплексами носитель - генетический материал.
П р и м е р 9. Направленная генетическая трансформация молочной железы in vivo.
С целью проверки доставки гена светлячковой люциферазы в клетки молочной железы мыши in vivo и возможности получения белка - продукта этого гена в клетках, в протоки молочной железы вводили при помощи устройства растворимый комплекс носитель - генетический материал, в состав которой входили конъюгат инсулин-полилизин в соотношении инсулин и полилизин 8:1 и плазмида pRVSL, несущая ген люциферазы светлячка. Носитель был синтезирован как описано в примере 1, растворимый комплекс приготовлен как описано в примере 2. Раствор комплекса (в концентрации 37,5 нМ) был переведен в сбалансированный солевой раствор Хэнкса следующего состава: NaCl (0,137 мМ), КCl (5,36 мМ), СаСl2 (1,26 мМ), MgSO4 x 7 H2O (0,41 мМ), MgCl2 x 6 H2O (0,49 мМ), Na2HPO2 x 7 H2O (0,34 мМ), КН2РО4 (0,44 мМ), рН 7,6, глюкоза (1,00 г/л) без красителя фенолового красного. Приготовленный раствор вводили по 25 мкл в каждый проток молочных желез самок лабораторных мышей BALB/c с помощью устройства. Устройство (фиг. 1) предварительно заполняли раствором Хэнкса так, чтобы заполнить выемку с кольцевой канавкой, сквозной осевой канал и канал для подачи отрицательного давления, и после присасывания соска животного вводили канюлю с растворимыми комплексами носитель - генетический материал сквозь заполняющий раствор в проток молочной железы.
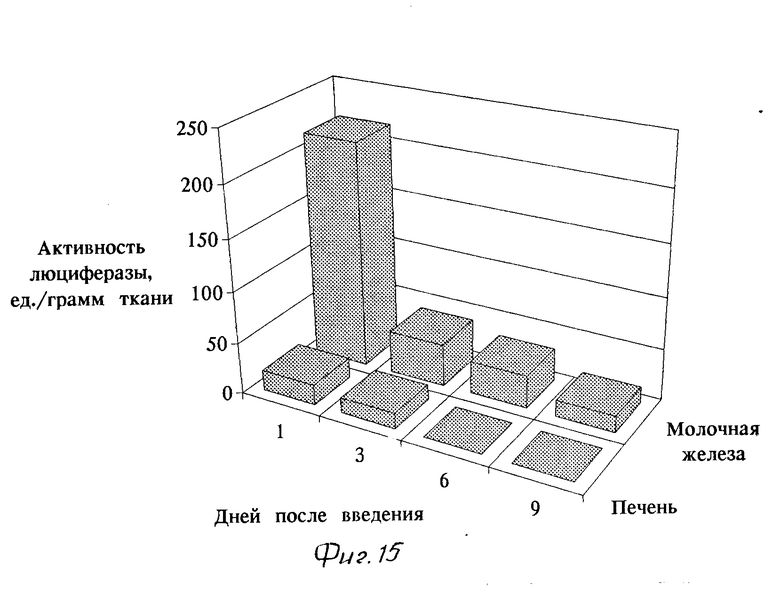
Результаты введения конструкции подтвердили, что ген люциферазы может быть доставлен при помощи выбранной конструкции по данному способу. Об этом свидетельствует наличие активности люциферазы в препаратах гомогената молочной железы мышей (фиг. 15), что было измерено при помощи люминометра. В тканях молочной железы контрольных мышей активности люциферазы обнаружено не было. Набольшая активность маркерного фермента - люциферазы наблюдалась через 1 сут после введения, но и через 9 сут в тканях еще оставался заметный уровень люциферазной активности (фиг. 15). При введении комплексов носитель - генетический материал только в часть молочных желез мыши, в других (не подвергшихся трансформации) молочных железах не было обнаружено активности люциферазы. В печени - основном органе-мишени для инсулина, обнаружены лишь следовые количества люциферазной активности. Тем самым показана возможность направленной доставки генетического материала в молочные железы животных.
П р и м е р 10. Подтверждение травмобезопасности и надежности устройства для введения генетического материала в молочный проток молочной железы животного.
Для определения возможных повреждений при введении канюли в проток молочной железы использовали лабораторных мышей BALB/c, которым вводили в молочные железы 0,4%-ный раствор красителя трипанового синего с молекулярным весом 960,8 в количестве по 100 мкл в каждый проток молочной железы при помощи стеклянных, специальным образом изготовленных на микрокузнице канюль с оплавленными (термически отшлифованными) рабочими кончиками.
Результаты введения анализировали под микроскопом типа "Биолам" с объективами 2,5х и 10х, а также окулярами 15х на препарированных молочных железах. Из 90 проверенных молочных желез ни одна не была повреждена.
Таким образом, предлагаемое устройство обеспечивает надежное и травмобезопасное введение генетического материала в молочный проток молочной железы животного.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННЫХ ЖИВОТНЫХ | 1996 |
|
RU2108714C1 |
| КОМПОЗИЦИЯ ДЛЯ ФОТОДИНАМИЧЕСКОГО ПОВРЕЖДЕНИЯ КЛЕТОК-МИШЕНЕЙ И СПОСОБ ФОТОДИНАМИЧЕСКОГО ПОВРЕЖДЕНИЯ КЛЕТОК-МИШЕНЕЙ | 1996 |
|
RU2066552C1 |
| СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНЫХ БЕЛКОВ В МОЛОЧНОЙ ЖЕЛЕЗЕ НЕТРАНСГЕННЫХ МЛЕКОПИТАЮЩИХ | 2003 |
|
RU2345088C2 |
| МОЛЕКУЛЯРНЫЕ КОНЪЮГАТЫ С ПОЛИКАТИОННЫМ УЧАСТКОМ И ЛИГАНДОМ ДЛЯ ДОСТАВКИ В КЛЕТКУ И ЯДРО КЛЕТКИ ДНК И РНК | 2009 |
|
RU2537262C2 |
| СПОСОБ ПЕРЕНОСА НУКЛЕИНОВОЙ КИСЛОТЫ В КЛЕТКИ ЖИВОТНЫХ | 1990 |
|
RU2098487C1 |
| СРЕДСТВО ДЛЯ ПЕРЕНОСА НУКЛЕИНОВЫХ КИСЛОТ ИЛИ ИХ ПРОИЗВОДНЫХ В КЛЕТКИ ЧЕЛОВЕКА ИЛИ ЖИВОТНЫХ | 1992 |
|
RU2113485C1 |
| СПОСОБ СТИМУЛИРОВАНИЯ РОСТА ПАРЕНХИМАТОЗНОЙ ТКАНИ МОЛОЧНОЙ ЖЕЛЕЗЫ ЖИВОТНОГО | 1987 |
|
RU2033178C1 |
| СПОСОБ РАННЕЙ ДИАГНОСТИКИ ЗЛОКАЧЕСТВЕННЫХ ОПУХОЛЕЙ ОРГАНОВ И ТКАНЕЙ (ЕГО ВАРИАНТЫ) - СПОСОБ ДААБУЛЯ И ВЕЩЕСТВО ДЛЯ ЕГО ОСУЩЕСТВЛЕНИЯ | 1997 |
|
RU2130775C1 |
| СПОСОБ ПОВЫШЕНИЯ ПРОДУКТИВНОСТИ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ И ПРЕПАРАТ ДЛЯ ЕГО ОСУЩЕСТВЛЕНИЯ | 1993 |
|
RU2034457C1 |
| СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННЫХ КРОЛИКОВ, ПРОДУЦИРУЮЩИХ БЕЛКИ В МОЛОЧНУЮ ЖЕЛЕЗУ | 2007 |
|
RU2402211C2 |





Использование: относится к генной инженерии в области животноводства, изобретение направлено на решение задачи генетической трансформации молочной железы животных. Сущность изобретения: способ включает введение в организм животного генетического материала в растворимом комплексе с носителями, которые направленно транспортируют генетический материал в клетку путем специфического рецептор-опосредованного эндоцитоза. Из ряда носителей используют носители направленно транспортирующие генетический материал также и в ядро клетки путем цитоплазменно-ядерного транспорта, при этом указанный растворимый комплекс вводят в организм животного через молочные протоки молочной железы. Устройство для введения генетического материала в молочный проток молочной железы животного содержит корпус с выемкой по форме соска животного, каналом для подачи отрицательного давления, сквозным осевым каналом на дне выемки и расширителем сфинктера сосца. Устройство содердит канюлю с мерной полостью для дозированного объема растворимого комплекса генетического материала с носителем, при этом выемка выполнена с возможностью охвата ею соска без зазора, диаметр сквозного осевого канала выполнен больше диаметра канюли, а расширитель сфинктера сосца выполнен в виде кольцевой канавки, расположенной на внутренней поверхности дна выемки по периметру сквозного осевого канала, причем кольцевая канавка соединена в ее углублении с каналом для подачи отрицательного давления. 2 н.п. ф-лы, 9 з.п. ф-лы, 15 ил.
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Однокамерный доильный стакан | 1983 |
|
SU1123598A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Колосниковая решетка с чередующимися неподвижными и движущимися возвратно-поступательно колосниками | 1917 |
|
SU1984A1 |
