Изобретение относится к биотехнологии, в частности к генетической инженерии высших растений, и может быть использовано для конструирования растений, устойчивых к заражению вирусами, а также для создания растительных продуцентов β -интерферона для фармакопеи.
Известно, что интерфероны человека всех трех подклассов α,β и γ ингибируют развитие вируса табачной мозаики на растениях табака [1, 2] Для интерферона α 2 ингибирующее действие продемонстрировано также на некоторых других парах вирус-растение [3] Интерфероны, по-видимому, имитируют какие-то эндогенные антивирусные факторы растений, так как показано, что α и β-интерфероны сходны с антивирусным фактором табака по физико-химическим свойствам и по механизму действия на клетку [1]
Известна рекомбинантная плазмидная ДНК, кодирующая синтез α -интерферона человека в растениях. Плазмида представляет собой вектор на основе вируса мозаики цветной капусты (СаМV) со встроенным вместо orfII и, таким образом, находящимся под управлением 35S промотора геном α -интерферона [4] Вектор обеспечивает довольно высокий уровень синтеза интерферона в зараженных клетках ≈2˙104 2˙105 е.а./г ткани.
Наиболее близкой к заявляемой плазмиде, прототипом является рекомбинантная плазмидная ДНК рST20, в которой ген рекомбинантного α -интерферона человека встроен в экспрессионный вектор pST6 под контроль сильного конститутивного промотора 35S вируса мозаики цветной капусты [5] Полученная рекомбинантная плазмидная ДНК рST20 перенесена в клетки Agrobacterium tumefaciens C58Cl RifR, несущие "разоруженную" Тi плазмиду pGV2260. Отобранные клоны трансконъюгантов, содержащие коинтеграт плазмид pGV2260 и pST20, были использованы для трансформации табака методом листовых дисков.
Основным недостатком данной плазмиды является то, что она в качестве переносимого гена содержит ген α -интерферона, тогда как известно, что β -интерферон как индуктор антивирусного ответа имеет ряд преимуществ перед α -интерфероном и проявляет антивирусную активность в более широком, чем α -интерферон, диапазоне концентраций [1] Кроме того, известная плазмидная ДНК кодирует белок только с внутриклеточной локализацией, что может быть неоптимально для проявления антивирусной активности.
Векторы для переноса гена β -интерферона в растения не описаны.
Целью изобретения является создание рекомбинантной плазмидной ДНК, кодирующей синтез β -интерферона человека в растениях.
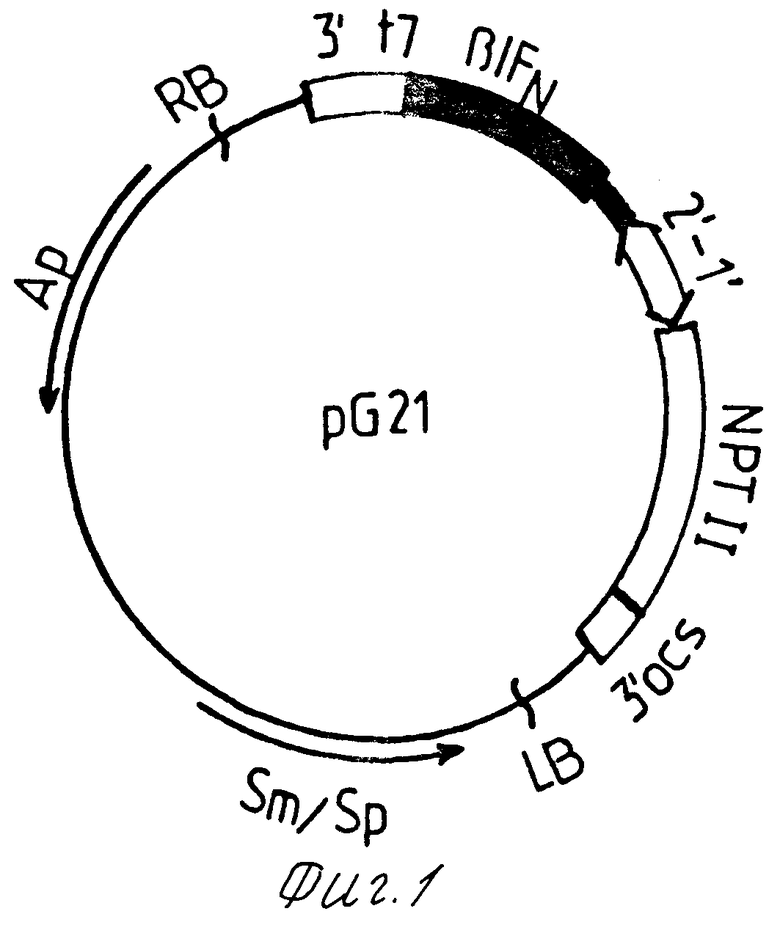
Это достигается тем, что сконструирована рекомбинантная плазмидная ДНК рG21 размером 9,1 т.п.н. предназначенная для создания трансгенных растений, продуцирующих β -интерферон человека и состоящая из следующих фрагментов:
ДНК векторной плазмиды рGSН160, размером 8,5 т.п.н.
HindIII-BamHI фрагмента ДНК, являющегося фрагментом хромосомного гена β -интерферона человека;
гена bla, кодирующего β -лактамазу;
гена Sm/Sp, кодирующего устойчивость к стрептомицину и спектиномицину;
гена NPTII, кодирующего неомицинфосфотрансферазу II, обеспечивающего устойчивость трансгенных растений к канамицину, промоторы РТ1 и РТ2.
Способ конструирования рекомбинантной плазмидной ДНК рG21 заключается в следующем: фрагмент структурной части гена β-интерферона, выщепленный из рекомбинантного фага λ HulF21 рестриктазами HindIII и BgIII, клонируют в полилинкере плазмиды πАN7, затем указанный ген, выщепленный из полученной плазмиды последовательной обработкой ферментами HindIII, polK и BgIII, переносят в плазмиду pGSH160, обработанную последовательно HindIII, polK и BamHI, содержащую функциональные элементы, необходимые для переноса и экспрессии гена в растениях.
Физическая карта плазмиды с указанием генетических маркеров приведена на фиг. 1. Плазмида имеет уникальные сайты рестрикции BamHI, Scal, KpnI и Ncol, несет бактериальные маркеры устойчивости к ампициллину и спектиномицину, фрагменты ДНК вектора pGSH160 RB, LB, 3'ocs, 3'Т7, РТ1' и РТ2', обеспечивающие перенос, интеграцию и экспрессию в геноме растений гена-репортера неомицинфосфотрансферазы II (NPTII) и гена β -интерферона человека.
Отличие полученной рекомбинантной плазмидной ДНК от прототипа состоит в использовании для конструирования целевой плазмидной ДНК структурной части хромосомного гена β -интерферона вместе с последовательностью, кодирующей транзиентный пептид, что позволяет получать растения, устойчивые к вирусной инфекции.
П р и м е р 1. Конструирование плазмиды pG21.
100 мкг ДНК рекомбинантного фага λHuIF21, содержащего фрагмент хромосомной ДНК человека с геном β -интерферона [6] обрабатывали 2 ч при 37оС рестриктазами HindIII (400 e.a.) и BglII (450 е.а.) в 500 мкл буфера, содержащего 20 мМ трис-HCl рН 7,6, 50 мМ NaCl, 10 мМ MgCl2 (буфер М). Полученный гидролизат использовали для выделения фрагмента ДНК, содержащего ген β -интерферона. С этой целью реакционную смесь разделяли электрофорезом в 1%-ном геле агарозы, гель окрашивали раствором бромистого этидия в воде (1 мкг/мл) и фрагмент ДНК размером 0,6 т.п.н. выделяли из геля электроэлюцией на DE-81.
2 мкг ДНК векторной плазмиды π AN7 обрабатывали 2 ч при 37оС рестриктазами HindIII (10 e.a.) и BglII (10 е.а.) в 30 мкл буфера М. Из полученного гидролизата выделяли фрагмент ДНК размером 0,7 т.п.н. С этой целью реакционную смесь разделяли электрофорезом в 1%-ном геле агарозы, гель окрашивали раствором бромистого этидия в воде (1 мкг/мл) и фрагмент ДНК размером 0,7 т. п.н. выделяли из геля электроэлюцией на DE-81.
Cмешивали 0,1 мкг HindIII-BglII фрагмента плазмиды π AN7 и 0,5 мкг HindIII-BglII фрагмента ДНК фага λHulF21, содержащего ген β -интерферона, в 20 мкл буфера, содержащего 20 мМ трис-HCl рН 7,6, 10 мМ MgCl2, 1 мМ дитиотреитола и 1 мМ АТФ (буфер Л), смесь охлаждали до 4оС, добавляли 70 е.а. ДНК-лигазы фага Т4 и инкубировали 16 ч. Лигазной смесью трансформировали компетентные клетки E. coli штамма МС1061(РЗ) и трансформантов, высевали на чашки с LB-агаром, содержащим 25 мкг/мл ампициллина, 12,5 мкг/мл тетрациклина и 50 мкг/мл канамицина. Клоны, несущие рекомбинантные плазмиды со встроенным геном, отбирали гибридизацией колоний с радиоактивно меченым олигонуклеотидом d(TTCTGAGACACTGAAAATT), комплементарным участку гена β-интерферона, кодирующему аминокислоты 14-19 в структуре зрелого белка (0). Плазмидные ДНК пяти клонов, дававших гибридизационный сигнал, анализировали с помощью гидролиза рестриктазами PstI, BglII и HindIII и с последующим разделением продуктов гидролиза гель-электрофорезом в 1%-ной агарозе. Во всех пяти плазмидах набор полученных в результате этого анализа фрагментов ДНК подтвердил встройку гена β-интерферона в плазмиду π AN7.
HindIII-BamHI фрагмент одной из таких плазмид ( πAN7-IFN) использован для конструирования плазмиды pG21. C этой целью 5 мкг ДНК плазмиды π AN7-IFN обрабатывали рестриктазой HindIII (20 e.a.), реакционную смесь охлаждали до 12оС, добавляли dATP, dTTP, dCTP и dGTP до концентрации 0,1 мМ и 3 е.а. фрагмента Кленова ДНК-полимеразы 1 (polK) и инкубировали 15 мин при 12оС. Ферменты удаляли экстракцией водным фенолом рН 8,0, ДНК трижды переосаждали этанолом из 2,5 М ацетата аммония, промывали 70%-ным этанолом, сушили, растворяли в 20 мкл буфера М и гидролизовали 30 е.а. рестриктазы BamHI в течение 2 ч при 37оС. Из полученного гидролизата гель-электрофорезом в 1%-ной агарозе выделяли фрагмент ДНК размером 0,6 т.п.н. как описано выше. 2 мкг ДНК плазмиды pGSH160 гидролизовали рестриктазой ClaI (5 e.a.) в 20 мкл буфера М в течение 2 ч при 37оС. Реакционную смесь охлаждали до 12оС, добавляли dCTP и dGTP до концентрации 0,1 мМ и 2 е.а. polK и инкубировали 15 мин при 12оС. Ферменты удаляли экстракцией водным фенолом рН 8,0, ДНК трижды переосаждали этанолом из 2,5 М ацетата аммония, промывали 70%-ным этанолом, сушили, растворяли в 20 мкл буфера рестрикции и гидролизовали 10 е.а. рестриктазы BamHI в течение 2 ч при 37оС. Ферменты удаляли фенолом и ДНК очищали от остатков фенола экстракцией хлороформом и спиртовым осаждением. 0,5 мкг полученной ДНК смешивали с 0,5 мкг ClaI-polK-BamHI фрагмента ДНК плазмиды π AN7-IFN в 20 мкл буфера Л и обрабатывали ДНК-лигазой (70 е.а.), как описано выше. Лигазной смесью трансформировали компетентные клетки E. coli штамма МС1061 и трансформантов, высевали на чашки с LB-агаром, содержащим по 100 мкг/мл ампициллина и спектиномицина. Клоны, несущие рекомбинантные плазмиды со встроенным геном β-интерферона, отбирали гибридизацией колоний с радиоактивно меченым олигонуклеотидом О. Плазмидную ДНК пяти клонов, дававших гибридизационный сигнал, анализировали с помощью гидролиза рестриктазами BamHI, PstI, ScaI, KpnI и NcoI и с последующим разделением продуктов гидролиза гель-электрофорезом в 1%-ной агарозе. Во всех пяти плазмидах набор полученных в результате этого анализа фрагментов ДНК подтвердил встройку гена β-интерферона в плазмиду pGSH160 и сохранность основных элементов структуры вектора.
П р и м е р 2. Получение трансгенных растений.
В качестве исходных эксплантов использовали листья трехнедельных растений табака Nicotiana tobaccum SR1, стерильно выращенных на среде MOW (культуральная среда MS с добавлением 8 г/л агара и 20 г/л сахарозы, рН 5,8). Листья разрезали на квадраты 5 х 5 мм и помещали (нижней стороной листа вверх) на чашку Петри с 15 мл жидкой МOW и 0,1 мл ночной культуры A. tumefaciens. Экспланты инкубировали двое суток в слегка затемненном месте, затем четырехкратно отмывали MOW c добавлением 500 мг/л клафорана и помещали на среду Т1 (культуральная среда MS с добавлением 1 мг/л бензиламинопурина (БАП), 0,1 мг/л нафтилуксусной кислоты (НУК), 500 мг/л клафорана, 50 мг/л канамицина, 8 г/л агара, 20 г/л сахарозы, рН 5,8).
Через 2-3 недели кусочки эксплантов с появившимися листочками переносили на среду Т2 (Т1, 0,1 мг/л БАП, без НУК), а еще через ≈2 недели на среду Т3 (Т2 без БАП). После появления корней растения переносили в почву во влажные камеры.
Выращенные в почве растения-регенеранты охарактеризованы стандартным тестом на активность гена-репортера, кодирующего бактериальную неомицинфосфотрансферазу (NPTII). Для всех полученных регенерантов наблюдали активность NPTII c характеристиками, идентичными бактериальному ферменту.
Интеграцию перенесенного фрагмента в геном растений доказывали двумя методами а) анализом расщепления по устойчивости к канамицину и по активности NPTII в F1 потомстве и б) блот-гибридизацией геномной ДНК трансгенных растений.
а) Семена F1 поколения стерилизовали 10%-ным гипохлоритом и спиртом по стандартной методике и высевали на чашки со средой MS c 500 мг/л канамицина. Чашки инкубировали две недели при 25оС со стандартным световым режимом (16 ч свет, 8 ч темнота) и подсчитывали соотношение белых и зеленых проростков. Показано, что это соотношение близко к 3:1 у большинства проанализированных линий. Выборочный анализ NPTII у канамицинустойчивых растений показал, что во всех проанализированных растениях присутствует активность NPTII c характеристиками, идентичными бактериальному ферменту.
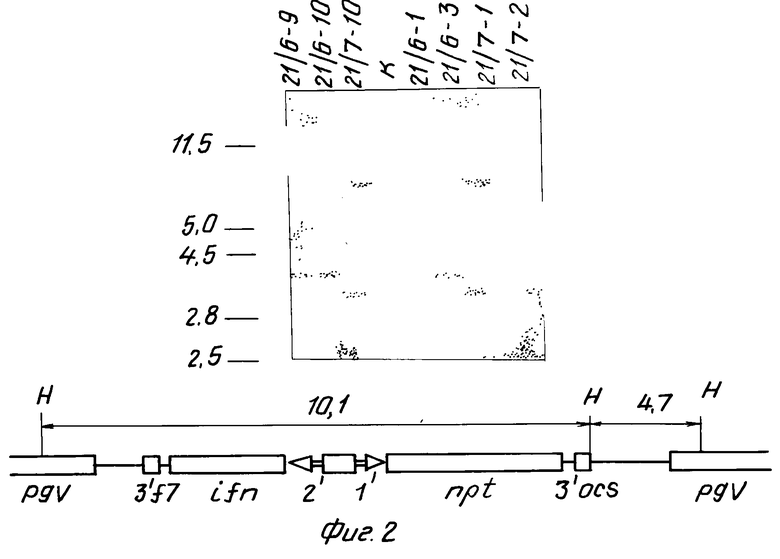
б) Геномную ДНК из листьев NPT+ растений поколения F1 и ДНК из клонов A. tumefaciens, содержащих коинтеграт pGV2260:pG21 (контроль), гидролизовали рестриктазой HindIII, фракционировали гель-электрофорезом, переносили на нитроцеллюлозный фильтр и гибридизовали с меченой 32Р ДНК плазмиды pG21. Результат представлен на фиг.2. Из фиг.2 видно, что, во-первых, в гидролизатах геномных ДНК трансгенных растений есть фрагменты, гибридизующиеся с зондом, и, во-вторых, размеры этих фрагментов различны в геномных ДНК трансгенных растений и в контроле.
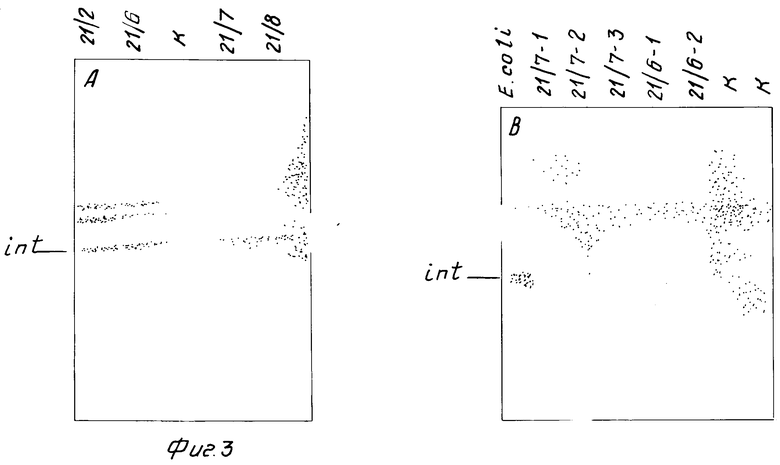
На фиг.3 представлены результаты тестирования β -интерферона в потомстве F1 NPT+ растений. Анализ проводился иммуноблоттингом по стандартной методике. Из фиг.3 видно, что в проанализированных экстрактах содержится белок, перекрестно реагирующий с моноклональными антителами к β -интерферону. Судя по электрофоретической подвижности, этот белок имеет молекулярную массу процессированного интерферона и, следовательно, проходит созревание при секреции.
П р и м е р 3. Тестирование устойчивости трансгенных растений к вирусу табачной мозаики (ВТМ).
Исходные молодые трансформированные растения после пересадки в почву и анализа NPT заражали ВТМ. Для этого суспензию очищенного ВТМ (0,05 мкг/мл) в 0,05 М натрий-фосфатном буфере с 10%-ным карборундом (200-400 меш) наносили на 2 молодых хорошо экспонированных листа (по 50 мкл на каждый лист) и слегка растирали стерильным стеклянным шпателем. За процессом инфекции следили, титруя через определенные промежутки времени вирус, развивающийся в листьях следующего яруса, на Nicotiana tobaccum cv Xanti nc. C этой целью кусочки листа, следующего за инокулированным, (≈100 мг) гомогенизировали при 4оС в 1 мл 0,05 М натрий-фосфатного буфера, центрифугировали и в супернатанте измеряли белок. Пробы выравнивали по содержанию белка, разбавляя буфером и по три аликвоты из каждой пробы наносили на три листа Nicotiana tobaccum cv Xanti nc, как описано выше. Через 4 дня подсчитывали число локальных некрозов.
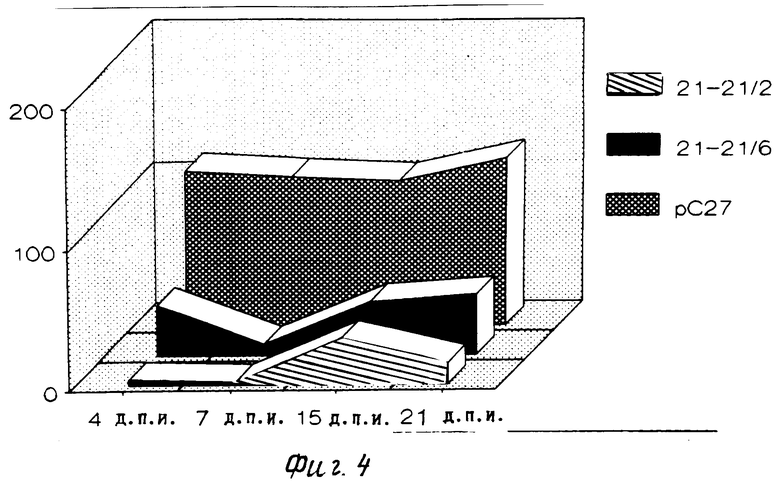
Результаты приведены на фиг.4. Из фиг.4 видно, что в растениях 21/2 и 21/6 титр вируса ниже, чем в контроле растениях, трансформированных "пустым" вектором рС27, в течение всего времени тестирования. (56) 1. Orchansky P. Rubinstein M. Sela, I. Proc.Natl.Acad.Sci. USA, 79 (1982), 2278-2280.
2. Vicente M. De-Fazio G. Menezes M.E. Golgher R.R. J.Phytopatol. 119 (1987) 25-31.
3. Kaplan, I.B. Taliansky, E. Malyshenko, S.I. Arch.Phytopatol. Pflanzenschutz (Berlin), 24 (1988) 3-8.
4. De Zoeten, G.A. Panswick, I.R. Horisberger, H.A. Virology, 172 (1989) 213-222.
5. Смирнов С.В. Теверовская Э.Х. Крашенинникова Л.В. Пухальский В.А. Генетика, 26 (1990), с. 2111-2121.
6. Ривкин М.И. Кузнеделов К.Д. Тезисы докладов V съезда ВОГИС им. Н.И. Вавилова, М. 1987, т.1, с. 236.
| название | год | авторы | номер документа |
|---|---|---|---|
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК PGII, ОПРЕДЕЛЯЮЩАЯ СИНТЕЗ БЕТА-ИНТЕРФЕРОНА ЧЕЛОВЕКА В ТРАНСГЕННЫХ РАСТЕНИЯХ, И СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННЫХ РАСТЕНИЙ, УСТОЙЧИВЫХ К ВИРУСНОЙ ИНФЕКЦИИ | 1994 |
|
RU2103361C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pBi101-IL18, КОДИРУЮЩАЯ СИНТЕЗ ИНТЕРЛЕЙКИНА-18 ЧЕЛОВЕКА В ТРАНСГЕННЫХ РАСТЕНИЯХ | 2005 |
|
RU2302460C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА pBi121-ESAT6-CFP10-gIFN, КОДИРУЮЩАЯ БЕЛКИ ESAT6 И CFP10 MYCOBACTERIUM TUBERCULOSIS И ГАММА-ИНТЕРФЕРОН ЧЕЛОВЕКА В ТРАНСГЕННЫХ РАСТЕНИЯХ | 2011 |
|
RU2468082C1 |
| СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННЫХ РАСТЕНИЙ ТАБАКА С ПОВЫШЕННЫМ СОДЕРЖАНИЕМ ПРОЛИНА | 2006 |
|
RU2324737C1 |
| СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННЫХ РАСТЕНИЙ ТАБАКА С ПОВЫШЕННЫМ СОДЕРЖАНИЕМ ПРОЛИНА | 2006 |
|
RU2324736C1 |
| СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННЫХ РАСТЕНИЙ МОРКОВИ, ПРОДУЦИРУЮЩИХ ИНТЕРЛЕЙКИН-10 ЧЕЛОВЕКА | 2007 |
|
RU2374321C2 |
| СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННЫХ РАСТЕНИЙ, УСТОЙЧИВЫХ К ВИРУСНОЙ ИНФЕКЦИИ | 2005 |
|
RU2300567C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА, ОБЕСПЕЧИВАЮЩАЯ ЭКСПРЕССИЮ ГЕНА ЭКСТРАКЛЕТОЧНОЙ РИБОНУКЛЕАЗЫ Zinnia elegans ZRNaseII В ТРАНСГЕННЫХ РАСТЕНИЯХ (ВАРИАНТЫ), И СПОСОБ ПОЛУЧЕНИЯ ВИРУСОУСТОЙЧИВЫХ ФОРМ РАСТЕНИЙ | 2009 |
|
RU2393226C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pBinPLUS-ARS-EPSPS, ОБЕСПЕЧИВАЮЩАЯ ЭКСПРЕССИЮ ГЕНА АГРОБАКТЕРИАЛЬНОЙ 5-ЕНОЛПИРУВИЛ-ШИКИМАТ-3-ФОСФАТ-СИНТЕТАЗЫ В ТРАНСГЕННЫХ МИКРОВОДОРОСЛЯХ РОДА ХЛОРЕЛЛА | 2009 |
|
RU2407794C1 |
| СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННЫХ РАСТЕНИЙ, РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК PTIVY4, ПРЕДНАЗНАЧЕННАЯ ДЛЯ ПОЛУЧЕНИЯ ТРАНСГЕННЫХ РАСТЕНИЙ И ШТАММ БАКТЕРИЙ AGROBACTERIUM TUMEFACIENS, ИСПОЛЬЗУЕМЫЙ ДЛЯ ТРАНСФОРМАЦИИ РАСТЕНИЙ | 1988 |
|
SU1547316A3 |
Использование: биотехнология, конкретно генетическая инженерия высших растений. Сущность изобретения: сконструирована рекомбинантная плазмидная ДНК pG 21, несущая копию хромосомного гена вместе с последовательностью, кодирующей транзиентный пептид, что позволяет получать растения, устойчивые к вирусной инфекции. Предлагаемая плазмида имеет размер 9,1 т.п.н. и сосоит из следующих фрагментов: ДНК векторной плазмиды PGS H 160, размером 8,5 т.п.н., Hind-III-Bam HI фрагмента ДНК, являющегося фрагментом хромосомного гена b - интерферона человека; гена bla , кодирующего b - лактамазу, гена Sm/Sp, кодирующего устойчивость к стрептомицину и спектиномицину, гена NP, TII, кодирующего неомицинфосфотрансферазу II, обеспечивающего устойчивость трансгенных растений к канамицину, промоторы PTI и PT2. Предлагаемая рекомбинантная плазмидная ДНК pG 21 кодирует синтез b - интерферона человека в растениях, что позволяет получать растения, устойчивые к вирусной инфекции. 4 ил.
РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК PG 21, КОДИРУЮЩАЯ β ИНТЕРФЕРОН ЧЕЛОВЕКА В РАСТЕНИЯХ размером 9.1 т.п.н. состоящая из:
ДНК векторной плазмиды pGSH 160 размером 8,5 т.п.н.
Hind III Bam HI фрагмента ДНК, являющегося фрагментом хромосомного гена b интерферона человека;
гена bla, кодирующего b лактамазу;
гена Sm/Sp, кодирующего устойчивость к стрептомицину и спектиномицину;
гена NPT II, кодирующего неомицинфосфотрансферазу II, обеспечивающего устойчивость трансгенных растений к канамицину;
промоторов PTI и PT 2.
| Ривкин М.И | |||
| и др | |||
| Тезисы докладов V съезда ВОГИС им.Н.И.Вавилова, М.: 1987, т.1, с.236. |
