Изобретение относится к области биотехнологии, в частности к генетической инженерии высших растений, и касается способа получения трансгенных растений, устойчивых к вирусной инфекции.
Известен способ получения трансгенных растений с антивирусной устойчивостью, которая обеспечивается с помощью экспрессии генов или некодирующих последовательностей антисмысловой РНК вирусного происхождения [Cooper В. Handbook of biological control. N.Y. Academic Press., 1999]. Основными недостатками способа являются специфическая устойчивость только к одному вирусу (или группе близкородственных вирусов), а также нестабильность вновь созданных механизмов устойчивости к вирусной инфекции.
Известны способы получения трансгенных растений, устойчивых к широкому спектру вирусов, и экспрессирующих гены рибосом-инактивирующих белков из фитолакки американской, ягод бузины и ириса [Lodge J.K., Kaniewski W.K, Tumer N.E. Proc. Natl. Acad. Sci. USA. 1993. V.90, p.7089-7093; Vandenbussche F. et al. Eur J Biochem. 2004. V.271, p.1508-15; Vandenbussche F. et al. Planta. 2004. V.220, p.211-221]. Основным недостатком данных способов является то, что данные белки являются цитотоксичными, что делает проблематичным их применение в сельском хозяйстве.
Наиболее близким к заявляемому способу - прототипом является способ получения трансгенных растений табака, экспрессирующих ген секреторной панкреатической рибонуклеазы, гидролизующей одноцепочечную РНК [Патент РФ №2300567, кл. C12N 15/82, оп. 10.06.2007], заключающийся в следующем. Конструируют рекомбинантную плазмиду PC27BN, состоящую из ДНК векторной плазмиды размером 12.5 т.п.н; фрагмента ДНК, содержащего ген панкреатической нуклеазы размером 455 п.н. Передают полученную рекомбинантную плазмиду pC27BN в штаммы Agrobacterium tumefaciens pGV2260 и AGL0 путем трехродительского скрещивания с использованием плазмиды-помощника pRK2013 или путем прямой трансформации и получения трансгенных растений табака путем трансформации листовых дисков Nicotiana tabacum cv. SR1 кокультивацией с Agrobacteria с последующим отбором на селективной среде.
Основными недостатками прототипа являются:
1. Использование гетерологичной рибонуклеазы животного происхождения, что делает этот способ недостаточно биобезопасным.
2. Использование только одной генетической конструкции с MAS2' промотором, локально индуцируемым при поранении, что сужает функциональные возможности способа и снижает его эффективность.
Технической задачей настоящего изобретения является создание новых рекомбинантных плазмид для трансформации растений и повышение устойчивости растений к вирусным инфекциям путем трансгенеза.
Поставленная техническая задача достигается созданием:
- новой рекомбинантной плазмиды pC27RNS, обеспечивающей экспрессию гена экстраклеточной рибонуклеазы Zinnia elegans ZRNaseII под управлением индуцибельного MAS2' - промотора в трансгенных растениях;
- новой рекомбинантной плазмиды pBiRNS, обеспечивающей экспрессию гена экстраклеточной рибонуклеазы Zinnia elegans ZRNaseII под управлением 35S промотора в трансгенных растениях;
- способа получения вирусоустойчивых форм растений.
Получают суммарную РНК из механически поврежденного листа Zinnia elegans. Затем с помощью обратной транскрипции с использованием праймеров oligo(dT) получают суммарную кДНК. Выделяют ген экстраклеточной рибонуклеазы Zinnia elegans ZRNaseII из суммарной кДНК с использованием метода ПЦР и клонируют полученный фрагмент в бинарных векторах:
а) рС27 под управлением индуцибельного MAS2'- промотора. Полученная рекомбинантная плазмида pC27RNS состоит из следующих элементов:
- ДНК векторной плазмиды размером 12.5 т.п.н.;
- фрагмента ДНК, содержащего кодирующую часть гена экстраклеточной рибонуклеазы Zinnia elegans ZRNaseII размером 720 п.н.
б) pBi101 под управлением конститутивного 35S промотора. Полученная рекомбинантная плазмида pBiRNS состоит из следующих элементов:
- ДНК векторной плазмиды размером 13.9 т.п.н.;
- фрагмента ДНК, содержащего кодирующую часть гена экстраклеточной рибонуклеазы Zinnia elegans ZRNaseII размером 720 п.н.;
- фрагмента ДНК, размером 442 п.н., содержащего промотор 35S РНК вируса мозаики цветной капусты.
Поставленная техническая задача достигается также предлагаемым способом получения вирусоустойчивых форм растений, заключающимся в следующем.
Описанные выше рекомбинантные плазмиды pC27RNS и pBiRNS, несущие кодирующую часть гена экстраклеточной рибонуклеазы Zinnia elegans ZRNaseII, путем прямой трансформации переносят в штамм Agrobacterium tumefaciens AGL0, затем полученными таким образом агробактериями проводят трансформацию растительного материала кокультивацией с Agrobacteria с последующим отбором трансгенных растений на селективной среде.
Определяющими отличиями предлагаемого изобретения по сравнению с прототипом являются:
- создание и использование для трансформации растений рекомбинантной плазмиды pBiRNS, обеспечивающей экспрессию гена экстраклеточной рибонуклеазы Zinnia elegans под сильным конститутивным промотором 35S РНК вируса мозаики цветной капусты;
- создание и использование для трансформации растений рекомбинантной плазмиды pC27RNS, обеспечивающей экспрессию гена экстраклеточной рибонуклеазы Zinnia elegans под индуцибельным поранением промотором MAS 2' Ti-плазмиды A.tumefaciens;
- использование в качестве целевого гена кодирующей части гена экстраклеточной рибонуклеазы растения Zinnia elegans, что позволяет эффективно защищать трансгенные растения от широкого спектра растительных вирусов, имеющих одноцепочечные РНК-геномы наиболее биобезопасным способом, в то время как секреторная панкреатическая рибонуклеаза быка (прототип) имеет гетерологичное (животное) происхождение. Экстраклеточная рибонуклеаза Zinnia elegans также локализуется в межклеточном пространстве и поэтому не цитотоксична для трансгенных растений;
- использование в способе двух векторных плазмид, что позволяет выбирать наиболее оптимальный для каждого объекта трансформации промотор. Выбор конструкции с конститутивным промотором 35S из вируса мозаики цветной капусты или индуцибельным MAS2' промотором может быть осуществлен для каждого конкретного объекта трансформации с учетом специфики инфицирующих объект вирусов.
Изобретение поясняется следующими примерами конкретного выполнения способа.
Пример 1.
Получение кодирующей части гена экстраклеточной рибонуклеазы Zinnia elegans ZRNaseII.
Суммарная РНК была выделена из механически поврежденного листа Zinnia elegans через 2,5 часа после поранения. С помощью обратной транскрипции с использованием праймеров oligo(dT) (НП АО «Силекс М») была получена суммарная кДНК с последующей амплификацией. Подбор последовательностей праймеров был осуществлен на основании известной нуклеотидной последовательности кДНК ZRNaseII. П1 5'-ACACTCGAGCACACAAACATGAAGA и П2 - 5' GAATCTAGAAATTTAGAATGAAGGA. Для проведения последующего клонирования в нуклеотидные последовательности синтезированных праймеров были включены сайты рестриктаз XhoI и XbaI. Фрагмент кДНК, полученный в результате амплификации, был клонирован в плазмиду pRT104 по сайтам рестриктаз XhoI и XbaI под контроль промотора 35S РНК вируса мозаики цветной капусты.
Пример 2.
Конструирование бинарного вектора (рекомбинантной плазмиды) pBiRNS, содержащего кодирующую часть гена экстраклеточной рибонуклеазы Zinnia elegans ZRNaseII.
кДНК с промотором, белок-кодирующей частью гена РНКазы, содержащей нуклеотидную последовательность лидерного пептида, а также сигналом полиаденилирования была амплифицирована с плазмиды pRT104 с помощью полимеразной цепной реакции [Carcana A., Confalone Е., Libonati М et al. Nucl. Acids Res. 1988, v.16, p.5491-5502], с праймерами (П3 5'-CAGGTCGACATGGTGGAGCA и П4 5'- GGAATTCGATTAAGTTGGGTA), содержащими сайты рестриктаз SalI и EcoRl. Полученный ПЦР-продукт был клонирован в вектор pBi101 по сайтам SalI и EcoRI с образованием плазмиды pBiRNS. Т-ДНК полученной плазмиды содержит ген неомицинфосфотрансферазы II под контролем нопалинсинтазного промотора Ti-плазмиды Agrobacterium tumefaciens (селективный маркер, определяющий устойчивость к канамицину) и белок-кодирующей частью целевого гена под управлением сильного конститутивного промотора 35S вируса мозаики цветной капусты. Схема т-ДНК вектора pBiRNS представлена на фиг.1.
Пример 3.
Конструирование бинарного вектора (рекомбинантной плазмиды) pC27RNS, содержащего кодирующую часть гена экстраклеточной рибонуклеазы Zinnia elegans ZRNaseII.
кДНК белок-кодирующей частью гена РНКазы, содержащей нуклеотидную последовательность лидерного пептида, была амплифицирована с плазмиды pRT104 с помощью полимеразной цепной реакции [Carcana A., Confalone Е., Libonati М et al. Nucl. Acids Res. 1988, v.16, p.5491-5502], с праймерами П1 5' - ACACTCGAGCACACAAACATGAAGA и П2 - 5' GAATCTAGAAATTTAGAATGAAGGA. Поскольку П2 содержит сайт рестриктазы XbaI, фрагмент кДНК, полученный в результате амплификации, был клонирован в плазмиду pC27 по сайту рестриктазы XbaI и тупому концу под контроль индуцибельного MAS2' Ti-плазмиды Agrobacterium tumefaciens. Схема т-ДНК вектора pC27RNS представлена на фиг.2.
Пример 4.
Получение трансгенных растений табака, несущих т-ДНК вектора pBiRNS.
Полученная по примеру 2 рекомбинантная плазмида pBiRNS, несущая кодирующую часть гена экстраклеточной рибонуклеазы Zinnia elegans ZRNaseII, была использована для трансформации клеток Agrobacterium tumefaciens штамма AGL0 путем прямой трансформации. Для агробактериальной трансформации в качестве исходных эксплантов были использованы листья трехнедельных растений табака Nicotiana tabacum SRI. Были получены 6 первичных трансформантов, устойчивых к канамицину (100 мг/л). Неомицинфосфотрансферазу (NPTII) в проростках тестировали по методу Рейса и соавторов [Reiss В., Sprendel R., Will Н. et al. Gene.1984, v.30, p.211-218]. Далее использовали только растения, устойчивые к канамицину и активно экспрессирующие неомицинфосфотрасферазу II (NPTII). На фиг.3 представлены результаты ПЦР анализа геномной ДНК проростков на наличие полноразмерной встройки целевого гена с праймерами П1-П2. Нанесения по дорожкам: 1 - трансгенный табак линии RNS1; 2 - трансгенный табак линии RNS5; 3 - трансгенный табак линии RNS10; 4 - трансгенный табак линии RNS16; 5 - нетрансгенный табак SR1; 3 - вектор pBiRNS; 4 - маркерная ДНК.
Далее провели селекцию трансформированных растений по признаку высокого уровня нуклеазной активности. Для измерения нуклеазной активности использовались растения одного возраста, выращенные стерильно в условиях климакамеры или в стандартных условиях теплицы. Листья отбирались приблизительно одного размера с одного уровня растений в каждом эксперименте. Нуклеазная активность листовых экстрактов оценивалась путем измерения изменения количества кислоторастворимого материала в суммарной РНК дрожжей [Blank А., McKeon Т.А. Proc. Natl Acad. Sci. USA 1989, v.86, р.3169-3173]. Результаты измерений нуклеазной активности приведены в таблице 1. В качестве контроля при измерении оптической плотности использовали реакционную смесь без добавления листового экстракта, время гидролиза в опыте составляло 60 минут [Galiana Е., Bonnet P., Conrod S. et al. Plant Physiol. 1997, v.115, p.1557-1567].
Из представленных в таблице 1 данных видно, что уровень нуклеазной активности грубых листовых экстрактов трансгенных растений табака, полученных с использованием конструкции pBiRNS, выше уровня активности контрольных растений. Так, нуклеазная активность экстрактов линий pBiRNS1 и pBiRNS2 приблизительно в 14 раз превышает активность экстрактов контрольных растений. Таким образом, заявляемый способ вкупе с созданной новой рекомбинантной плазмидой pBiRNS позволяет получать трансгенные растения с эффективно повышенным уровнем нуклеазной активности в листьях.
В таблице 2 представлены данные по тестированию устойчивости трансгенных растений табака линий pBiRNS1, pBiRNS5, pBiRNS10 и pC27RNS27 к вирусной инфекции.
Появление фенотипических симптомов и содержание антигена (мкг/мл) ВТМ в листьях контрольных (SR1) и трансгенных растений Nicotiana tabacum через 3 недели после инфекции демонстрирует повышенную устойчивость трансгенных растений к вирусу табачной мозаики вплоть до полной вирусоустойчивости при низких концентрациях вируса в инокулюме.
Пример 5.
Получение трансгенных растений табака, несущих Т-ДНК вектора pC27RNS.
Полученная по примеру 3 рекомбинантная плазмида pC27RNS, несущая кодирующую часть гена экстраклеточной рибонуклеазы Zinnia elegans ZRNaseII, была использована для трансформации клеток Agrobacterium tumefaciens штамма AGL0 путем прямой трансформации. В основном процедура проводилась также, как и в примере 4. Результаты измерений нуклеазной активности листовых экстрактов растений линий pC27RNS приведены в таблице 3.
Из представленных данных видно, что уровень нуклеазной активности в трансгенных растениях табака, полученных с использованием конструкции pC27RNS, достоверно выше уровня активности контрольных растений. При сравнении данных таблицы 1 и таблицы 3 можно заметить, что конструкция pC27RNS повышает уровень нуклеазной активности не так значительно, как конструкция pBiRNS. Авторы считают, что в некоторых случаях это может оказаться полезным для растения, так как не перегружает белоксинтезирующие и секретирующие системы растений. Кроме того, использованный в pC27RNS промотор индуцируется поранением и наши данные показывают, что в течение 24 часов после поранения нуклеазная активность в механически поврежденных растениях линий pC27RNS повышается в 2-2,5 раза. Представленные в таблице 2 данные по устойчивости линии pC27RNS27 к ВТМ показывают, что растения линии pC27RNS27 отличаются повышенной вирусоустойчивостью по сравнению с контрольными растениями.
Использование предлагаемого способа позволит повысить устойчивость растений к вирусным инфекциям путем трансгенеза.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННЫХ РАСТЕНИЙ, УСТОЙЧИВЫХ К ВИРУСНОЙ ИНФЕКЦИИ | 2005 |
|
RU2300567C1 |
| СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННЫХ РАСТЕНИЙ ТАБАКА С ПОВЫШЕННЫМ СОДЕРЖАНИЕМ ПРОЛИНА | 2006 |
|
RU2324736C1 |
| ХИМЕРНЫЙ РЕГУЛЯТОРНЫЙ УЧАСТОК ДЛЯ ЭКСПРЕССИИ ГЕНОВ В РАСТЕНИЯХ (ВАРИАНТЫ), КЛАСТЕР ДЛЯ ЭКСПРЕССИИ ГЕНА (ВАРИАНТЫ), КЛАСТЕР ДЛЯ ИНДУЦИБЕЛЬНОЙ ЭКСПРЕССИИ ЧУЖЕРОДНОГО ГЕНА (ВАРИАНТЫ), СПОСОБ ЭКСПРЕССИИ ГЕНА В РАСТЕНИИ (ВАРИАНТЫ), СПОСОБ ИНДУЦИБЕЛЬНОЙ ЭКСПРЕССИИ ЧУЖЕРОДНОГО ГЕНА В РАСТЕНИИ (ВАРИАНТЫ) И ПЛАЗМИДА (ВАРИАНТЫ) | 1994 |
|
RU2142998C1 |
| СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННЫХ РАСТЕНИЙ ТАБАКА С ПОВЫШЕННЫМ СОДЕРЖАНИЕМ ПРОЛИНА | 2006 |
|
RU2324737C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК PGII, ОПРЕДЕЛЯЮЩАЯ СИНТЕЗ БЕТА-ИНТЕРФЕРОНА ЧЕЛОВЕКА В ТРАНСГЕННЫХ РАСТЕНИЯХ, И СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННЫХ РАСТЕНИЙ, УСТОЙЧИВЫХ К ВИРУСНОЙ ИНФЕКЦИИ | 1994 |
|
RU2103361C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА pBM И СПОСОБ ПОЛУЧЕНИЯ С ЕЕ ИСПОЛЬЗОВАНИЕМ БЕЗМАРКЕРНЫХ ТРАНСГЕННЫХ РАСТЕНИЙ, СИНТЕЗИРУЮЩИХ ЦЕЛЕВЫЕ ПРОДУКТЫ | 2009 |
|
RU2410433C1 |
| СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННЫХ РАСТЕНИЙ МОРКОВИ, ПРОДУЦИРУЮЩИХ ИНТЕРЛЕЙКИН-10 ЧЕЛОВЕКА | 2007 |
|
RU2374321C2 |
| КОНСТРУКЦИЯ ДНК, КОНСТРУКЦИЯ РЕКОМБИНАНТНОЙ ДНК И СПОСОБ ПОЛУЧЕНИЯ РАСТЕНИЙ | 1994 |
|
RU2136757C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pBi101-IL18, КОДИРУЮЩАЯ СИНТЕЗ ИНТЕРЛЕЙКИНА-18 ЧЕЛОВЕКА В ТРАНСГЕННЫХ РАСТЕНИЯХ | 2005 |
|
RU2302460C1 |
| ПРОДУЦИРОВАНИЕ ТРЕГАЛОЗЫ В РАСТЕНИЯХ | 1994 |
|
RU2143496C1 |
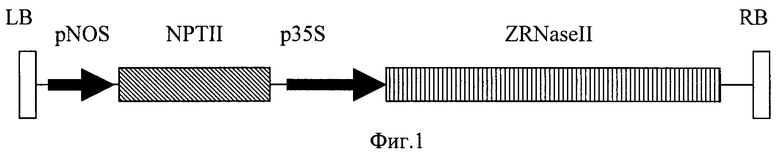
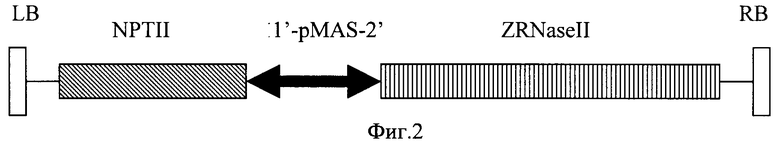

Плазмида pC27RNS содержит ДНК кодирующей части гена экстраклеточной рибонуклеазы Zinnia elegans ZRNaseII под управлением индуцибельного MAS2'-промотора. Плазмида pBiRNS включает ДНК кодирующей части гена экстраклеточной рибонуклеазы Zinnia elegans ZRNaseII под управлением 35S-промотора. Плазмиду pC27RNS или pBiRNS переносят в штамм Agrobacterium tumefaciens. Полученными агробактериями производят трансформацию растительного материала кокультивацией с Agrobacteria с последующим отбором трансгенных растений на селективной среде. Экспрессия гена экстраклеточной рибонуклеазы Zinnia elegans ZRNaseII в растениях придает им повышенную устойчивость к вирусным инфекциям. 3 н.п. ф-лы, 3 ил., 3 табл.
1. Рекомбинантная плазмида pC27RNS, обеспечивающая экспрессию гена экстраклеточной рибонуклеазы Zinnia elegans ZRNaseII под управлением индуцибельного MAS2'-промотора в трансгенных растениях, состоящая из следующих элементов:
ДНК векторной плазмиды размером 12,5 т.п.н.;
фрагмента ДНК, содержащего кодирующую часть гена экстраклеточной рибонуклеазы Zinnia elegans ZRNaseII размером 720 п.н.
2. Рекомбинантная плазмида pBiRNS, обеспечивающая экспрессию гена экстраклеточной рибонуклеазы Zinnia elegans ZRNaseII под управлением 35S промотора в трансгенных растениях, состоящая из следующих элементов:
ДНК векторной плазмиды размером 13,9 т.п.н.;
фрагмента ДНК, содержащего кодирующую часть гена экстраклеточной рибонуклеазы Zinnia elegans ZRNaseII размером 720 п.н.;
фрагмента ДНК размером 442 п.н., содержащего промотор 35S РНК вируса мозаики цветной капусты.
3. Способ получения вирусоустойчивых форм растений, включающий конструирование рекомбинантной плазмиды, содержащей целевой ген, экспрессия которого придает растению устойчивость к поражению вирусом, передачу полученной плазмиды в штамм Agrobacterium и инфицирование последним растительного материала с последующим отбором трансгенных растений на селективной среде, отличающийся тем, что в качестве рекомбинантной плазмиды используют плазмиду pC27RNS или плазмиду pBiRNS, при этом ген, экспрессия которого придает растению устойчивость к поражению вирусом, имеет растительное происхождение и является кодирующей частью гена экстраклеточной рибонуклеазы растения Zinnia elegans ZRNaseII.
| СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННЫХ РАСТЕНИЙ, УСТОЙЧИВЫХ К ВИРУСНОЙ ИНФЕКЦИИ | 2005 |
|
RU2300567C1 |
| ГЕН, ОБУСЛОВЛИВАЮЩИЙ УСТОЙЧИВОСТЬ РАСТЕНИЙ К БОЛЕЗНЯМ, И ЕГО ПРИМЕНЕНИЕ | 1997 |
|
RU2252960C2 |
| НОВЫЕ ГЕНЫ РАСТЕНИЙ И ИХ ПРИМЕНЕНИЕ | 2000 |
|
RU2241749C2 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК PGII, ОПРЕДЕЛЯЮЩАЯ СИНТЕЗ БЕТА-ИНТЕРФЕРОНА ЧЕЛОВЕКА В ТРАНСГЕННЫХ РАСТЕНИЯХ, И СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННЫХ РАСТЕНИЙ, УСТОЙЧИВЫХ К ВИРУСНОЙ ИНФЕКЦИИ | 1994 |
|
RU2103361C1 |
| JP 2000287569 A, 17.10.2000 | |||
| JP 6245660 A, 06.09.1994 | |||
| US 6320099 В1, 20.11.2001 | |||
| Прибор, замыкающий сигнальную цепь при повышении температуры | 1918 |
|
SU99A1 |

