Изобретение относится к биотехнологии, в частности к средствам защиты клеток растений от вредного воздействия гербицидов.
Создание микроорганизмов и клеток высших организмов с новыми генетически обусловленными признаками открывает множество самых неожиданных направлений в науке и технике. Одним из таких направлений является создание различных средств или агентов, которые обладают цитотоксическим эффектом. Например, многие соединения, получившие распространение в сельском хозяйстве, предназначены для уничтожения вредителей, сорняков и т.д. Для этих соединений характерно сравнительно длительное время пребывания в окружающей среде или организме, или же продолжительный остаточный эффект.
Вместе с тем, во многих ситуациях задача заключается в том, чтобы дифференцировать виды, которые необходимо сохранить от видов, подлежащих уничтожению. Например, весьма часто желательно обеспечить избирательное уничтожение сорняков, нанося при этом минимально допустимое повреждение культурным растениям. Многие гербициды широкого спектра действия оказывают значительный неблагоприятный эффект на культурные растения и их употребление ограничено главным образом предвсходовым периодом или осторожным послевсходовым применением.
Поэтому огромный интерес представляет появившаяся возможность модификации жизнеспособных клеток для придания и резистентности к стрессам, например в результате воздействия цитотоксических сpедств или агентов.
Известно применение бактериального aro-А-гена для придания чувствительным к глифосату клеткам резистентности к этому гербициду. Описано микробиологическое разложение гербицидов семейства 3,5-дигалогено-4-гидроксибензонитрилов. Известно разложение бромоксинила в тяжелых глинах района Регина.
Техническая задача изобретения заключается в том, чтобы разработать средства, предназначенные для детоксикации местообитаний, содержащих бромоксинил и родственные ему гербициды, и для защиты клеток от цитотоксического воздействия таких гербицидов.
Предложен способ получения устойчивых к 3,5-дигалоген-4-гидроксибензонитрилу клеток табака.
Используемый в способе структурный ген может быть получен из бактерии, способной использовать бензонитрил в качестве источника азота. В дальнейшем при указании понятия "бензонитрил" имеется в виду, что бензонитрилом является галоидированный n-гидроксибензонитрил, в частности, 3,5-дииодо- или 3,5-дибромо-4-гидроксибензонитрил, под нитрилазой имеется в виду нитрилаза, способная использовать такой галоидированный бензонитрил в качестве источника азота.
Фермент нитрилаза может быть получен из бактерии, которая существует в природе в условиях окружающей среды, содеpжащей бромоксинил или оксинил. В частности, представляют интерес энтеробактерии, более конкретно вида Klebsiella. Может использоваться Klebsiella pneumoniae, конкретнее, ее разновидность ozaenae. Другие микроорганизмы могут быть выращены в почве или другой среде при возрастающих концентрациях бензонитрила и уменьшающихся количествах альтернативных источников азота до тех пор, пока не будут получены организмы, которые выживают, используя бензонитрил в качестве единственного источника азота.
Полученный фермент должен быть подвергнут скринингу, чтобы убедиться в эффективности нитрилазы в отношении детоксификации бензонитрила. Кроме того, нитрилаза должна быть специфичной в отношении бензонитрила, а не других аналогов, в которых нет атомов галогена или имеются другие заместители и т. п. Таким образом нитрилаза должна быть специфичной в отношении бензонитрилов в том виде, как они определены, и быть сравнительно неактивной в отношении аналогов или значительно менее активной в отношении аналогов. При этом не должно быть значительного снижения скорости размножения, т.е. должно наблюдаться менее, чем 10%-е снижение размножения бактерии в присутствии нормального источника азота, например аммиака, по сравнению с бензонитрилом в качестве источника азота при сопоставимых концентрациях. Подобный результат не должен наблюдаться для неспецифических бензонитрилов.
После выявления штаммов организма хозяина используют методики идентификации последовательности, кодирующей нитрилазу. Ген может присутствовать на хромосоме или плазмиде. Геном фрагментируют с помощью эндонуклеазы рестрикции, для чего используют одну или множество эндонуклеаз, чтобы получить фрагменты 5-50 тыс. пар оснований. Эти фрагменты клонируют в подходящих векторах, полученными рекомбинантными ДНК трансформируют бактерии, например E. coli, и осуществляют скрининг трансформантов по нитрилазной активности, причем организм-реципиента обеспечивает отрицательный фон.
После идентификации одного или большего числа клонов, проявляющих нитрилазную активность, вне хромосомные элементы, содержащие требуемый ДНК-фрагмент выделяют традиционными методами. Лизируют реципиент, осаждают ДНК и отделяют векторную плазмидную или вирусную ДНК от хромосомной ДНК. Внехромосомные элементы расщепляют с помощью рестриктазы и целевые фрагменты выделяют различными методами разделения и идентификации фрагментов разных размеров, например с помощью электрофореза, центрифугирования в градиенте плотности и т.п.
В зависимости от размера фрагмента с ним обычно могут проводиться дальнейшие манипуляции для уменьшения его размеров с целью более близкого приближения к размеру гена и его фланговым регуляторным областям. Существуют различные методы манипуляции с фрагментом, содержащим последовательность, кодирующую фермент и его регуляторные фланговые последовательности. Может использоваться частичное расщепление разными рестриктазами в разных реакционных смесях с последующим клонированием фрагментов для определения того, какие из них еще сохраняют способность обеспечивать экспрессию нитрилазы.
В альтернативном варианте фермент может быть выделен и подвергнут частично анализу последовательности. На основе аминокислотной последовательности могут быть подготовлены зонды, которые затем используют для идентификации тех фрагментов, в которых имеется ген. При сочетании этого подхода с рестрикционно-ферментным расщеплением фрагменты могут быть клонированы и подвергнуты скринингу на присутствие требуемого гена. Кроме того, можно использовать экзонуклеазы, например ВаL31, для удаления нуклеотидов с одного или обоих концов фрагмента для дальнейшего уменьшения числа ненужных нуклеотидов.
В альтернативе ген может быть клонирован в подходящем хозяине и информационная РНК выделена скринингом с помощью зонда. Выделенная информационная РНК может быть затем использована для получения к ДНК с применением традиционных методов, включающих обратную транскриптазу и образование комплементарной цепи с помощью ДНК-полимеразы. В этом случае результирующий структурный ген лишается регуляторных областей, сопряженных с транскрипцией.
ДНК-последовательность, содержащая структурный ген, кодирующий нитролазу, может быть соединен с целым рядом ДНК-последовательностей.
При использовании в качестве реципиентов прокариотических клеток с помощью трансформации, конъюгации, трансдукции или трансфекции в них могут быть введены ДНК-последовательности с помощью целого ряда векторов. Например, можно использовать в качестве векторов рBR 322, рАСУС184, рМВ9, рR2К290, космиды, такие как рVK100, или вирусы, такие как р22 и т.п.
В случае эукаристических хозяев может использоваться целый ряд методов введения ДНК в хозяйский организм, например, трансформация с помощью СА++-осажденной ДНК, включение нереплицирующейся ДНК-последовательности, плазмиды, или минихромосомы, трансформация с помощью Т-ДНК-содержащей последовательности в Agrobacterium, микроинъекция микропипеткой, или электропорация. В зависимости от присутствия в ДНК-конструкции репликационной системы, определяют, может ли ДНК реплицироваться в качестве эписомного элемента, или же ДНК может быть интегрирована в хозяйский геном, и структурный ген экспрессирован в хозяйском организме. Могут быть использованы такие эписомные элементы, как опухоле-индуцирующие плазмиды, например Ti или Ri, или их фрагменты, или вирусы, например CaMV, TMV или их фрагменты, которые не являются летальными для хозяина, и где структурный ген присутствует в эписомных элементах.
Полученные фрагменты, кодирующие нитрилазу, могут быть клонированы с помощью подходящего клонирующего вектора. Клонирование может быть проведено в прокариатическом микроорганизме, например бактерии, такой как E.coli.
При необходимости, можно использовать космиду, когда частичное или полное переваривание дает фрагменты, имеющие требуемый размер. Например, космида рVK100 может быть частично переварена подходящим рестрикционным ферментом и связана с фрагментами, получаемыми в результате либо частичного, либо полного переваривания плазмиды, хромосомы, или их фрагмента. Упаковка обеспечивает то, что упаковываться и трансдуцироваться в реципиенте будут только фрагменты требуемого размера.
Реципиент может быть выбран по резистентности к бензонитрилу. Штаммы-реципиенты могут быть модифицированы, т.е. содержать подходящие генетические маркеры, которые позволяют провести отбор трансдуктантов. В микроорганизмах трансдуктанты могут использоваться для конъюгации с другими микроорганизмами, при использовании в случае необходимости мобилизирующих плазмид. Для уменьшения размера фрагмента, содержащего ген нитрилазы, могут использоваться различные методы. Например, космидный вектор может быть выделен, расщеплен рестрикционными эндонуклеазами, например EcoRI, BglII, SmaI и т.п. и результирующие фрагменты клонированы в подходящем векторе, для удобства в ранее использованном космидном векторе. Вместо космидного вектора существует множество клонирующих векторов малого размера, таких как рАСУС177 и рАСУС184. Фрагменты, кодирующие фермент, должны иметь размер менее 5 тыс. пар оснований, обычно менее 4 тыс. пар оснований, более предпочтительно менее 2 тыс. пар оснований.
Желательно, чтобы фрагмент имел около 1 тыс. т.п. и менее 5 тыс. п.о. предпочтительно менее 4 тыс. п.о. в частности не менее 1047 пар оснований, более конкретно с включением фланговых областей размером не менее 1100 пар оснований, предпочтительно менее 1,5 тыс.п.о. Особый интерес представляет BglII-SmaI фрагмент из Klebsiella ozaene, конкретнее PstI-Hinc II фрагмент размером около 1210 пар оснований.
Нитрилазный фермент может быть экспрессирован с помощью любого источника как прокариотного, так и эукариотного, включая бактерии, дрожжи, нитевидные грибы, растительные клетки и т.п. В тех случаях, когда не обеспечивается секреция, осуществляют лизис клеток и выделение нитрилазы известным путем. Например, проводят хроматографию, электрофорез, афинную хроматографию и т.п. Для удобства бромоксинил может быть конъюгирован с помощью карбоксильной группы с нерастворимой подложкой, и использоваться для выделения нитрилазы.
Нитрилазо-специфическая активность составляет не менее 0,1 мкмоль аммиака/мин/мг белка, как правило не менее 0,5 или выше.
Очищенный фермент может использоваться самыми разными способами. Он может использоваться при проведении анализов на бромоксинил, иоксинил и другие родственные бензонитрилы. В альтернативе рассматриваемый фермент будучи конъюгированным с представляющим интерес аналитом,например гептеном, или с антителом может применяться в качестве метки для диагностических анализов.
Кодирующая нитрилазу ДНК-последовательность может использоваться множеством способов. ДНК-последовательность может применяться в качестве зонда для выделения нитрилаз дикого типа или мутированных. В альтенративе, ДНК-последовательность может использоваться для интеграции путем рекомбинации в организме-реципиена для придания хозяину резистентности к бензонитрилу.
В случае растительных клеток, структурный ген, в качестве части конструкции, может быть введен в ядро растительной клетки с помощью инъекции микропипеткой для интеграции путем рекомбинации в хозяйский геном. В альтернативе может использоваться электропорация. В случае, когда структурный ген получен из источника, имеющего регуляторные сигналы, которые не распознаются растением-хозяином, необходимо ввести подходящие регуляторные сигналы для экспрессии. Когда используется вирус или плазмида, например опухоле-индуцирующая плазмида, и для нее построена карта, рестрикционный сайт, в который встраивается структурный ген, должен располагаться ниже промотера. Если ДНК-последовательность не имеет подходящего рестрикционного сайта, можно провести переваривание с экзонуклеазой в течение различных периодов времени, например с Bal 31 и сшить с синтетическим линкером.
Особый интерес представляет использование опухолеиндуцирующей плазмиды, например Ti или Ri, где нитрилазный ген может быть интегрирован в хромосомы растительных клеток.
При использовании правой границы Т-ДНК или обеих границ, в случае, когда границы фланкируют структурный ген, под контролем регуляторных элементов, распознаваемых растением-хозяином, экспрессионная кассета может быть интегрирована в геном растения, что обеспечивает экспрессию фермента в растительной клетке на различных стадиях дифференцирования.
Могут быть получены различные конструкции, обеспечивающие экспрессию в растительных клетках. Для обеспечения транскрипции может использоваться множество промоторных областей, являющихся составной частью конститутивных и ин дуцируемых областей.
Область, запускающая транскрипцию, соединяется со структурным геном, кодирующим нитрилазу. Выше кодона запуска, обычно в пределах 200 пар оснований должна содержаться 5'-область нетранслируемая, которая не имеет АТG кодона.
3' конец структурного гена должен содеpжать один или более стоп-кодонов, которые соединяются с областью прекращения транскрипции, функционирующей в растении-хозяине. Область прекращения транскрипции может быть ассоциирована с тем же или иным структурным геном, что и область, начала транскрипции.
В качестве регуляторных областей могут использоваться спиновые промотерные и терминаторные области, которые обеспечивают конструктивную экспрессию нитрилазного гена. Могут использоваться промотеры и/или терминаторы, которые обеспечивают индуцируемую экспрессию или регулируемую экспрессию в растении-хозяине. Промотерные области, такие, как октопино-синтазный промотер, нопалино-синтазный промотер, агропино-синтазный промотер, маннопиносинтазный промотер и т.п. Другие промотеры включают вирусные промотеры, такие как промотер области VI CaMV или промотер полной длины 35S, промотеры, ассоциированные с рибулозо-1,5-бисфосфат-карбоксилатными генами, например, небольшая субъединица, гены, ассоциированные с фазеолином, накоплением белка, Б конглицинином, образованием целлюлозы и тому подобным.
Различные последовательности могут быть соединены традиционными методами. Промотерная область может быть идентифицирована как область, являющейся 5' концом от структурного гена, например, опинового гена, и может быть выбрана с помощью рестрикционной карты. Аналогична терминаторная область может быть выделена как область 3' от структурного гена. Последовательности могут быть клонированы в нужной ориентации для обеспечения конститутивной экспрессии нитрилазного гена в растении-хозяине.
Клетки культурного растения модифицируют введением функционального гена, экспрессирующего нитрилазный фермент, можно использовать бромоксинил, иоксинил или аналогичный гербицид, который обеспечивает полное уничтожение или уничтожение сорняков, оставляя культурное растение относительно непораженным. Тем самым можно добиться существенной экономии благодаря тому, что удобрения и вода могут использоваться более эффективно и устраняются неблагоприятные эффекты, обусловленные присутствием сорняков.
Рекомбинантная плазмидная ДНК, экспрессирующая нитрилазный фермент, может быть введена в целый ряд растений, как однодольных, так и двудольных, включая кукурузу, пшеницу, сою, табак, хлопчатник, томаты, картофель, крестоцветные Brassica sp. рис, арахис, петунию, подсолнечник, сахарную свеклу, газонная трава и т.п. Ген может присутствовать в клетках или частях растений, включая каллюс, ткань, корни, клубни, побеги, проростки, семена, листья рассаду, пыльцу и т.п.
Бромоксинил может использоваться как таковой для послевсходовой обработки сорняков в сочетании с культурными растениями, которым обеспечена безопасность.
Традиционно пестициды вносятся на поля в виде препаратов 0,1-4 фунт/акр, (0,1-4,5 кг/га) предпочтительно 0,2-2 фунт/акр (0,2-2,25 кг/га) бромоксинила, тогда как другой гербицид вносится в количестве, дающем 0,1-4 фунт/арк (0,1-4,5 кг/га) активного ингредиента. Препараты включают и другие добавки, такие как поверхностно-активные вещества, вспомогательные вещества, агенты для улучшения распределения препарата, агенты для улучшения прилипания препарата, стабилизаторы и тому подобное. Препараты могут быть как мокрыми, так и сухими, включая растекающиеся порошки, эмульгируемые концентраты, и жидкие концентраты, как это общеизвестно в данной области техники.
Гербицидные растворы могут наноситься традиционными методами, например опрыскиванием, поливом, опылением и т.п.
Материалы и методики.
Ферменты рестрикции и Т4-лигаза для связывания использовались в соответствии с рекомендациями изготовителя.
Для всех экспериментов по клонированию использовали штамм E.coli ММ294.
При использовании антибиотиков обеспечивались следующие уровни: (хлорамфеникол) 25 кмг/мл, Тс (тетрациклон) 10 мкг/мл, Ар(пенициллин) 300 мкг/мл.
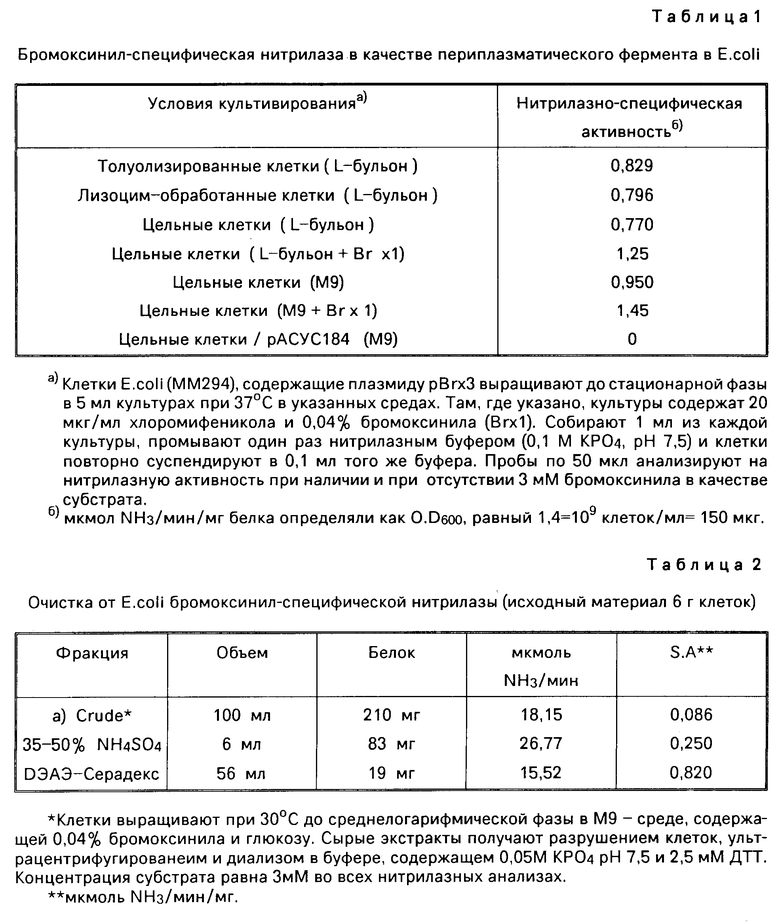
П р и м е р 1. Бактериальные культуры из загрязненного бромоксинилом образца почвы выделяют и проводят скрининг. Один такой организм идентифицируют как Klebsiella pneumoniae, подвид ozaenae. Частичная очистка бромоксинилспецифической нитрилазы из указанного организма дает активный фермент с молекулярной массой 34 кДаль.
После повторного субкультивирования K.ozaenae на твердом L-агаре выделяют мутант, который не в состоянии использовать бромоксинил в качестве единственного источника азота при выращивании в жидкой среде, содержащей на 1 литр: КН2РО4 (1,5 г), К2НРО4 (3,5 г), MgSO4˙7Н2О (0,1 г), дрожжевого экстракта (50 мг), цитрат глицерин и сукцинат на уровне 0,1% и микроэлементы.
В дальнейшем эта среда именуется как мультиуглеродная среда УЕТЕ. Мультиуглердоная среда УЕТЕ содержит 0,05% бромоксинила. Хотя этот организм не использовал бромоксинил в качестве единственного источника азота, он вырастал до полной плотности в L-бульоне, содержащем 0,05% бромоксинила. Выбирают колонию варианта К.ozaenae и выращивают в 10 мл L-бульона. Три независимые колонии К.ozaenae снимают с LB-чашки, содержащей бромоксинил и выращивают в тех же условиях. Те же четыре колонии К.ozaenae одновременно выращивают в 10 мл L-бульона с добавлением 0,05% бромоксинила. Культуры выращивают до полной плотности при 30оС и мини-препаративную плазмидную ДНК выделяют из каждой культуры. Непереваренные плазмидные ДНК подвергают электрофорезу на 0,5%-агарозном геле и плазмидные полосы проявляют окрашиванием этидий-бромидом.
Мутант К.ozaenae давал одиночные плазмидные агрегаты (размером 68 тыс.п. о. ) при выращивании как в присутствии, так и в отсутствии бромоксинила. Три колонии К. ozaenae дают более крупные плазмидные агрегаты (90 тыс.п.о.) при выращивании в присутствии 0,05% бромоксинила. В отсутствие бромоксинила в двух из трех колоний К.ozaenae присутствуют обе плазмидные формы. Эти данные свидетельствуют о конверсии более крупных плазмидных агрегатов до меньшей формы, с потерей приблизительно 22 тыс.п.о. плазмидной ДНК при устранении отбора по бромоксинилу.
Все четыре колонии выращивают в 200 мл L-бульона, содержащего 0,055% бромоксинила. Клетки разрушают на прессе Френча, высокоскоростные надосадочные жидкости диализуют против буфера, содержащего 0,05 КРО, рН 7,5, 2,5 мМ дитиотреитола (DTT) и индивидуальные сырые экстракты анализируют на бромоксинил-специфическую нитрилазную активность. Сырой экстракт, полученный из варианта. К. ozaenae, не проявляет нитрилазной активности, тогда как сырые экстракты других К. ozaenae проявляют нитрилазо-специфическую активность, равную 0,124, 0,105 и 0,143 мкмоль NH3/мин/мг белка, соответственно. Клетки (200 мл) выращивают при 30оС до среднелогарифмической фазы в среде М9, содержащей 0,1% глюкозы, и 0,04% бромоксинила. Сырые экстракты получают в результате разрушения клеток ультрацентрифу- гированием и диализом надосадочной жидкости в буфере, содержащем 0,05 М КРО4, рН 7,5 и 2,5 мМ ДТТ. Во всех анализах концентрация субстрата составляет 3 мМ бромоксинила. Контролируют выделение NH3. Способность утанта К.ozaenae расти в L-бульоне, содержащем бромоксинила приводит к устойчивости организма к данному соединению. Однако, данный организм не может расти в указанных средах, используя бромоксинил в качестве единственного источника азота.
Ген, кодирующий нитрилазы К.ozaenae, находится на плазмиде. Кодирующий фермент ген(ы) представляет собой сегмент плазмидной ДНК длиной 22 тыс. п.о. который утрачивается плазмидой К.ozaenae в отсутствии отбора по бромоксинилу.
П р и м е р 2. Экспрессия бромоксинил-специфичной нитрилазы К.ozaenae в E.coli.
Плазмидную ДНК из K.ozaenae, выращенной в условиях отбора по 0,05% бромоксинила, готовят и ДНК трансформируют в штамм E.coli ММ294 (thi, gyr А96, end I-, nsd R17). Трансформанты отбирают на дефицитной по азоту твердой, агарозной, минимальной среде (содержащей на 1 литр: КН2РО4 (1,5 г), К2НРО4 (3,5 г), MgSO4˙7H2O (0,1 г) и 0,1% глюкозы) с добавлением 0,05% бромоксинила в качестве единственного источника азота. После инкубирования в течение 5 суток на селекционных чашках появлялось 10 колоний. Эти колонии переносят на L-агаровые чашки, содержащие 0,05% бромоксинила, и проверяют на присутствие тиамин-акуситрофного маркера в ММ294. Ни одна из колоний, выращенных в минимальной среде в отсутствии тиамина, не являлась штаммом E.coli ММ294. Все колонии растут в среде М9 с добавлением тиамина и 0,05% бромоксинила в качестве единственного источника азота. В отсутствии бромоксинила рост в этой среде не наблюдался. Две из колоний отбирают для дальнейшего анализа. Когда препараты сырого экстракта E. coli ММ294, содержащие плазмиду размером 90 тыс. п. о. анализируют на бромоксинил-специфическую нитрилазную активность. При этом специфическая активность нитролазы составляет 0,216 мкмоль выделяемого NH3 мин/мг Е.coli ММ294. Колонии, содержащие меньшие плазмидные агрегаты, не давали обнаруживаемой нитрилазной активности. Большую (90 тыс.п. о. ) плазмиду в Е.coli обозначают рВrx1, тогда как меньшую плазмиду (68 тыс. п.о.) обозначают рВrx1А.
Чтобы убедиться, что штамм Е. сoli ММ294, содержащий плазмиду рBrx1, продуцирует нужный метаболит в результате бромоксинил-специфической нитрилазной реакции, 2 мл культуры ММ294 (рВrx1) выращивают в течение 24 часов при 30оС в среде М9 с добавлением 0,05% бромоксинила. Образец культурального фильтрата хроматографируют на колонке С18-ВЭЖХ. Весь поступивший бромоксинил в культуральном фильтрате преобразуют в пик нового метаболита. Метаболитовый пик по данным спектрального анализа относят на счет 3', 5'-дибром-4-гидроксибензойной кислоты (ДБГБ). Таким образом, продукт экспрессии в Е.coli плазмиды, кодирующей бромоксинил, оказывается тем же, что и в случае K.ozaenae.
П р и м е р 3. Клонирование гена кодирующего нитрилазу в Е.coli.
Плазмиду рВrx1 переваривают с помощью BamHI, что дает две полосы по 53 тыс. п. о. и 37 тыс.п.о. соответственно, BamHI фрагменты встраивают в BamHI сайт плазмидного вектора рАСУС184 Е.coli и трансформируют в штамм Е.соli ММ294. Клонирование в BamHI сайте вектора рАСУС184 приводит к инактивированию гена устойчивости к тетрациклину. Выбирают 10-хлорамфеникол-резистентных, тетрациклин-чувствительных колоний ММ294, выделяют ДНК, переваривают ее с помощью BamHI. Четыре клона содержат фрагмент размером 37 тыс.п.о. BamHI, тогда как один клон имел больший (53 тыс.п.о.) Bam HI ДНК-фрагмент плазмиды рВrx1. Пять клонов, содержащих клонированный BamHI фрагмент, обнаруживают также в плазмиде рВrx1Δ которая соответствует ДНК-последовательности, остающейся после отщепления 22 тыс.п.о. у плазмидной ДНК от рВrx1. Все 10 клонов выращивают в 200 мл L-бульона в присутствии 20 мкг/мл хлорамфеникола (для отбора плазмид), получают препараты сырого экстракта и анализируют на бромксинил-спецфическую нитрилазную активность. Четыре клона, содержащие BamHI фрагмент размером 37 тыс.п.о.проявляют нитрилазо-специфическую активность в диапазоне 0,140 мкмоль выделенного NH3/мин/мг белка, тогда как в других шести клонах нитрилазной активности обнаружено не было. Эти данные показывают, что ген, кодирующий бромоксинилспецифическую нитрилазную активность, располагается на BamHI-фрагменте размером 37 тыс.п.о. клонированном из плазмиды рВrx1. ДНК-фрагмент длиной 22 тыс.п.о. самопроизвольно теряемый в отсутствии отбора по бромоксинилу, характерен для BamHI-фрагмента размером 37 тыс.п.о.
Чтобы убедиться в ориентации BamHI фрагментов относительно вектора рАСУС184, ДНК из указанных четырех клонов переваривают с помощью ЕсоRI и подвергают электрофорезу на 0,07%-агарозном геле. Скомбинированные продукты ЕсоRI-переваривания плазмид рВrx1 и рВrxIA также анализируют.
Обе ориентации BamHI-фрагмента 37 тыс. п.о. относительно вектора рАС УС184 определяют и обозначают. соответственно, как плазмиды рВrx2 и pBrx3. Отмечают также, что три ЕсоRI фрагмента являются характерными для ДНК-фрагмента размером 22 тыс. п.о. который самопроизвольно отщепляется от плазмид рВrx2 и pBrx3. Размеры этих EcoRI-фрагментов соответственно 18 тыс.п.о. 3 тыс.п.о. и 1,9 тыс.п.о. Ген, кодирующий бромоксинил-специфическую нитрилазу, должен располагаться в одном из этих трех EcoRI фрагментов, если нитрилазный структурный ген не рассекается EcoR I рестрикционным сайтом.
Исследована локализация бромоксинил-специфической нитрилазы E. coli (pBrx3). Ниже приведены следующие результаты.
Эти данные показывают, что клеточным местоположением нитрилазного фермента является периплазматическое пространство. Второе наблюдение заключается в том, что фермент экспрессируется в отсутствии бромоксинила в среде, свидетельствуя, что индукция бромоксинилом не требуется для экспрессии фермента.
Дальнейшая очистка бромоксинил-специфической нитрилазы.
Дальнейшая очистка нитрилазы К.ozaenae проводилась с получением следующих результатов.
Колонку 2,5 см2 х 10 см уравновешивают в буфере, содержащем 0,05% КРО4, рН 7,5 2,5 мМ ДТТ и 1 мМ ЭДТУК. Пробу наносят и колонку элюируют 300 мл линейного градиента от 0,02 М до 0,40 М NaCl указанного буфера. Буфер, содержащий 1 м NaCl вводят в конце градиента. Собирают фракции по 5 мл и аликвотные пробы по 0,075 мл чередующихся фракций анализируют на нитрилазную активность. Одиночный пик ферментной активности элюируют при 0,22 М соли. Приблизительно 75% нитрилазной активности выделяют в активных фракциях.
Фракции, дающие нитрилазный пик из колонки ДЭАЗ, диализируют относительно 0,02 М КРО4, рН 7,5 и 50 мкл (6 мкг белка), каждой фракции наносят на 11,25% денатурирующий гель Лэммли. Обогащенная белком полоса, которая соответствует пику активности из ДЭАЭ-колонки, представляет собой полипептид с молекулярной массой 34000. Эти данные подтверждают, что бромоксинилспецифическая нитрилаза представляет собой полипептид молекулярной массой приблизительно 34000 и является продуктом единственного гена.
Клон рBrx2 полностью переваривают с помощью EcoRI и выделяют фрагмент приблизительно 19 тыс.п.о. Фрагмент вставляют в EcoRI-сайт вектора рАСУС184 (3,9 тыс.п.о.) для получения плазмиды рВrx5, которую трансформируют в Е.соli по описанной выше методике. Плазмиду выделяют обычным путем и переваривают с помощью Bgl II для получения фрагмента приблизительно 6,7 тыс.п.о. который остается встроенным в вектор рАСУС184. Выделенную плазмиду рВrx7 затем переваривают с помощью SmaI и BglII для получения фрагмента размером приблизительно 3,9 тыс.п.о. который встроен в SmaI-BamHI переваренную рАСУС177 (3,7 тыс. п.о.). Результирующую плазмиду с геном устойчивости к пенициллину трансформируют в Е.соli, как описано выше, и трансформанты отбирают на пенициллин-селективной среде для по лучения плазмиды рВrx8, которая несет нитрилазный ген на фрагменте 3,9 тыс.п.о. рВrx8 частично переваривают с помощью PstI и фрагменты вставляют в PstI переваренную рVC18. Результирующие плазмиды клонируют в Е.соli и проводят скрининг на нитрилазную активность. Один клон имел плазмиду рВrx9 длиной 5,3 тыс.п.о. которую выделяют и дополнительно переваривают с помощью PstI и Hinc II, что давало фрагмент 1210 п.о. имеющий в направлении от Pst I к Hinc II рестрикционные сайты ClaI, SalI, Sca I и Sph I, сравнительно равномерно распределенные. Pst I-Hinc II фрагмент используют для определения последовательности. Pst I Hinc II фрагмент, 5'- и 3'-фланговых областей лигируют с EcoR I линкерами, переваривают с помощью EcoRI и вводят в экспресионную плазмиду, предназначенную для растений, путем вставки в EcoRI-сайт рСGN451.
рСGN 451 включает октопиновую последовательность, которая содержит около 1566 п. о. 5'-некодирующей области, слитой через EcoR I-линкер с 3'-концом гена, и около 1349 п. о. 3'некодирующей ДНК. рТi-координаты от 11.207 до 12.823 для 3'-области и 13.643-15.208 для 5'-области, 5'-фрагмент получают следующим образом. Небольшой субклонированный фрагмент, содержащий 5'-конец кодирующей области, в качестве BamHI-ЕсоRI фрагмента клонируют в плазмиду pCGN 407. BamHI-EcoRI-фрагмент имеет Хmn I сайт в кодирующей области, тогда как рBR 322 имеет два ХmnI-сайта. рCGN 407 переваривают с помощью Хmn I, повторно рассекают нуклеазой Bal 31 и к ферментам добавляют EcoRI-линкеры. После EcoRI и BamHI переваривания фрагменты фракционируют по размеру, фракции клонируют и определяют последовательность. В одном случае была удалена вся кодирующая область и 10 п.о. 5'-нетранслируемой последовательности, что оставляет 5'-нетранскрибируемую область, "сар" сайт мРНК и 16 п.о. 5'-нетраслируемой области (до сайта Bam HI). Этот небольшой фрагмент получают фракционированием на 7% -м акриламидном геле и элюируют фрагменты длиной приблизительно 130 п.о. Фракционированную по ДНК связывают с М13mp9 и несколько клонов подвергают определению последовательности. Последовательность сравнивают с неизвестной последовательностью октопино-синтазного гена. Конструкция М13 обозначают р14, и эту плазмиду переваривают с помощью Bam HI и EcoR I, что дает небольшой фрагмент, который связывают с фрагментом от XhoI-BamHI, содержащим 5' последовательности из рТiА6 и с фрагментом от EcoR-XhoI, содержащим 3'-последовательности. Результирующий XhoI-фрагмент клонируют в XhoI сайт производного рU38, обозначенного как рСGN 426. Эта плазмида отличается от рUC8 наличием единственного EcoRI-сайта, заполненного ДНК полимеразой I, и утратой сайта Pst I и Hind III из-за нуклеазного загрязнения Hinc II рестрикционной эндонуклеазой, когда Xho I линкер был встроен в единственный HInc II сайт плазмиды рU38. Результирующая плазмида рСGN451 имеет единственный EcoR I сайт для вставки кодирующих белок последовательностей между 5'-некодирующей областью (которая содержит 1550 п.о. 5'-нетранскрибируемой последовательности, включающей правую границу Т-ДНК, "сар", сайт и РНК- и 16 п.о. 5'-нетранслируемой последовательности) и 3'-область (которая содержит 267 пар оснований кодирующей области: стоп кодон, 196 п.о. 3'-нетранслируемой ДНК, полиА-сайт и 1153 п.о. 3'-нетранскрибируемой последовательности).
XhoI фрагмент, содержащий октопино-синтетазную (ос) кассету, вставляют в плазмиду рСGN 517, которая имеет гены резистентности к тетрациклину и канамицину. рСGN517 получают из рНС79 введением в единственный Pst I сайт Kanr-гена из рVC4r pCGN 517 переваривают с помощью Sal I в единственный Sal I сайт вставляют Xho I-фрагмент. Xho I фрагмент. Затем этот фрагмент вставляют также во вторую плазмиду рСGN 529 pCGN 529 получают из рАСУС184 вставкой Kanr -гена из Tn5.
Bgl II фрагмента размером 2,4 тыс.п.о. из Т-ЛДНК рRiA4, вставляют в Bam HI-сайт, остающийся после замещения Hind III Bam HI фрагмента плазмиды рАСУС184 Kanr-геном из Tn5. XhoI-фрагмент, содержащий осs-кассету, в которую EcoRI-нитрилазный ген встроен в уникальной EcoR I-сайт ocs-кассеты, вставляют в рСGN 517 и рСGN 529 с получением двух плазмид рN1 и pN2, соответственно, которые используют для введения в А.tumefaciens или А.rhizogenes, соответственно, для интеграции с Т-ДНК Т; или Ri плазмид. Интеграцию в плазмиды достигают трехродительским скрещиванием. Культуры Е.coli реципиента, содержащие плазмиды рL К2073, pN1 или рN2, А.tumefaciens А722 или A.rhizogenes А4Т культивируют в течение ночи. Культуры смешивают и распределяют по АВ-чашкам, содержащим 150 мкг/мл канамицина. Одиночные колонии подвергают повторной штриховой разводке дважды. Правильность интеграции проверяют анализом по Саузерну общей ДНК-Agrobacterium. Переваренную с эндонуклеазой ДНК зондируют с помощью рВrх8 с транслированным сочетанием генов.
П р и м е р 4. Экспрессия бромоксинил-специфического нитрилазного гена в галловой ткани.
Плазмиду рВrx9, которая несет нитрилазный ген на фрагменте длиной 2,6 тыс. п. о. переваривают с помощью BamHI, и обрабатывают с помощью Bal 31 для удаления некоторой 5'-фланкирующей области. Добавляют Bam HI-линкеры и осуществляют повторное замыкание. Результирующие плазмиды, которые обеспечивали ампициллиновую резистентность, трансформируют в E.coli по описанной выше методике и трансформанты отбирают на ампициллин-селективной среде с получением плазмид рВrх16 и рВrх17 размером 5,2 тыс. пар оснований, которые несут нитрилазный ген на фрагменте 2,6 тыс. пар оснований. рВrx16 переваривают с Bam HI и затем частично переваривают с Hinc II, что дает фрагмент 1,2 тыс. п.о. с нитрилазным геном.
Bam HI-Hinc II фрагмент вставляют в переваренную Bam HI Sma I плазмиду рСGN 46 с получением плазмиды рВr х 22 длиной 6,6 тыс.п.о. содержащей фрагмент нитрилазного гена. Плазмида рСGN 46 содержит MAS промотер и осs 3'-область. Плазмиду рСGN 46 конструируют следующим образом. Приблизительно 5,5 тыс. п. о. EcoR I фрагмент размером 55 тыс.п.о. (Есо 13 или ЕсоС), несущий часть Т-R ДНК, включающий область маннопино-синтазного промотера (Рmas), клонируют в векторе, обозначенном рVK322. После переваривания рVK232 с помощью ЕсоRI, EcoR13, полученный фрагмент вставляют в ЕсоRI-сайт рАСУС184 с получением плазмиды рСGN14, pCGN 14 переваривают с помощью Sph I и Cla I (соответственно в положении 21562 и 20128 последовательности) для удаления Pmas области, которую встраивают в рVС19. рVC19 переваривают с помощью Sph I и AccI с получением РСGN 40. Pmas область включает литилированный сайт распознавания Cla I внутри, что защищает его от переваривания.
рСGN40 переваривают с помощью EcoRV и EcoRI, причем ЕсоRV сайт находится в Т-ДНК, тогда как ЕсоRI-сайт находится в полилинкере плазмиды рVC19. Это обеспечивает получение фрагмента, и меющего Рmas область, рСGN 451, содержащую октопино-синтазную кассету, переваривают с помощью Sma I и EcoRI и выделяют больший фрагмент, из которого удаляют октопино-синтазную 5'-область.
ЕсоRV-EcoRI-область Pmas встраивают в рСGN 451 вместо октопиносинтазной 5-области, где область начала транскрипции и стоп кодон разделены полилинкером, с получением рСGN46.
Плазмиду рВrx22, содержащую ген нитрилазы длиной 1,2 тыс.п.о. трансформируют в Е. соli. Плазмиду выделяют обычным путем и переваривают с помощью XhoI с получением фрагмента 4,1 тыс.п.о. содержащего МАS промотер, бромоксинильный ген, содержащий 25 пар оснований бактериальной 5'-нейтранслируемой последовательности и осs-3'-область. Фрагмент 4,1 тыс.п.о. вставляют в SalI- переваренную плазмиду рСGN 783 с получением плазмиды рВrx28 размером приблизительной 31 тыс.п.о.
П р и м е р 5. Построение рСGN 783.
Построение рСGN 167.
Для построения рСGN 167, Alu I-фрагмент СаМV (пары оснований 7144-7735) получают перевариванием с помощью Alu I- и клонируют в Hinc II-Сайт М13mp7 c образованием С614. ЕсоRI-переваривание С614 дало ЕсоRI- фрагмент из С614, содержащий 35S-промотер, который клонируют в ЕсоRI- сайт pUC8 c получением рСGN 146.
Bgl II-сайт (7660 пара оснований) обрабатывают с помощью BglII и Bal 31, а затем Bgl II-линкер присоединяют к Bal 31 обработанный ДНК для получения рСGN 147. рСGN 148 а, содержащую промотерную область, выбираемый маркер (КАN с двумя АТG) и 3'-область, получают перевариванием рСGN 528 (см. ниже) с помощью Bgl II и вставкой Bam HI-Bgl II промотерного фрагмента из рCGN 147. Этот фрагмент клонируют в Bgl II сайт плазмиды рСGN 528 так, чтобы Bgl II сайт располагался вблизи канамицинового гена рСGN 528.
Челночный вектор, использованный для этого построения, рСGN 528, получают следующим образом. рСGN 525 получают перевариванием плазмиды, содержащей Tn5, в которой располагается канамициновый ген и встраиваются Hind III-BamHI фрагмента, содержащего канамициновый ген в место Hind III Bam HI фрагмента с тетрациклиновым геном рАСУС184. рСGN 526 получают вставкой Bam HI- фрагмента 19 из рTi А6 в Bam HI4сайт рСGN 525. рСGN 527 получают удалением небольшого Хho I-фрагмента из рСGN 526 в результате переваривания с помощью Хho I и повторным связыванием.
рСGN 149 а получают клонированием BamHI фрагмента гена канамицина из рМВ9Каn XXI в BamHI-сайт рcGN148а.
рМВ9Каn XXI представляет собой вариант рVС4К, который не имеет Xho I-сайта, но содержит функциональный канамициновый ген из Tn 903, чтобы обеспечить эффективный отбор в Agrobacterium.
рСGN149a переваривают с помощью Bgl II и Sph I. Этот небольшой Bgl II Sph I фрагмент рСGN149а замещают BamHI-SphI фрагментом из МI (см. ниже), выделеннного перевариванием с помощью BamHI и Sph I. Это дает рCGN 167 конструкцию, содержащую СаМV промотер на полную длину, 1ATG канамициновый ген, 3'- конец и бактериальный канамициновый ген типа Tn 903 М1, расположенные на ЕсоRI фрагменте из рСGN550 (см. построение рСGN 587), которые клонированы в ЕсоRI-клонирующем сайте М13mp9 таким образом, чтобы Pst I-сайт в 1АТG-канамициновом гене был вблизи полилинкерной области М13mp9.
Построение 709 (1АТG канамицин-3'-область)
рСGN566 содержит ЕсоRI-Hind III-линкер из рUС18, вставленный в ЕсоRI и Hind III-сайты pUC13-Cm, Hind III Bgl II фрагмет из pNW 31с-8,29-1, содержащий ORF 1 и 2 (см. выше).
Субклонируют в Hind III-Bam HI-фрагмент из рСGN 566, получая рСGN 703.
Sau 3A фрагмент из рСGN 703, содержащий 3'-область транскрипта 7 из рTi A6 (соответствует основаниям 2396-2920 в рТ115955) субклонируют в BamHI-сайт pUC18 (см. выше), получая рСGN 709.
Построение рСGN766с (35S промотер-3'-область)
Hind III-Bam HI-фрагмент рСGN167 (построение см. здесь же), содержащий СаМV-35S-промотер, 1АТG-канамициновый ген и Bam HI-фрагмет 19 из рТiA6, клонируют в BamHI-Hind III-сайты рUC19, получая рСGN 976.
35S-промотер и 3'-область из транскрипта 7 сшивают с Hind III-EcoR I-фрагментом размером 0,7 тыс.п.о. из рСGN 976 (35S-промотер) и ЕсоRI-Sal I-фрагментом 0,5 тыс. п.о. из рСGN 709 (транскрипт 7:3') и встраивают в Hind III и Sal I-сайты рСGN 566, получая рСGN766 c.
Окончательное построение рСGN783 Hind III-Eco RI-фрагмент 0,7 тыс.п.о. из рСGN 66 с (СаМV-35S-промотер) связывают с ЕсоRI-Sal-фрагментом 1,5 тыс.п. о. из рСGN 726 с (1-АТG-KAN-3' область) и встраивают в Hind III-Sal I сайтах puC119 с получением рСGN778.
Hind III-Sal I-фрагмент, содержащий CaMV 35S промотер (1-АТG-KAN-3'-область из плазмиды рСGN 778 размером 2,2 тыс.п.о. замещают HInd III-Sal I- полилинкерной областью из рСGN739 с получением рСGN 783.
рВrx17 переваривают с помощью Bam HI и затем частично переваривают с помощью Hinc HI, что дает фрагмент с нитрилазным геном длиной 1,2 тыс.п.о. BamHI-Hinc II фрагмент вставляют в BamHI и Sma I переваренную рСGN 566 с получением плазмиды рВrx25,3,7 тыс.п.о. содержащей ген нитрилазы.
рСGN 566 строят следующим образом. PUC13 (CmR) переваривают с помощью ЕсоR I и HInd III, и полилинкеры из рUC18 и рUC19 вставляют, соответственно, в линеаризованную рUC13 с получением рСGN566, которая несет маркер резистентности к хлорамфениколу.
Плазмиду рBrx25, содержащую ген нитрилазы, трансформируют в Е.coli. Плазмиду выделяют обычным путем и переваривают с Bam HI и EcoRI для получения фрагмента с нитрилазным геном размером 1,2 тыс.п.о. BamHI- EcoRI-фрагмет вставляют и в Bam HI и EcoR I переваренную рСGN46 с получением плазмиды рВrx27 длиной 6,6 тыс.п.о. содержащей нитрилазогенный фрагмент.
рВrx27 трансформируют в Е.coli. Плазмиду выделяют обычным путем и переваривают с Xho I с получением фрагмента 4,1 тыс.п.о. содержащего MAS-промотер, бромоксинильный ген, содержащий II пар оснований бактериальной 5'-транслируемой последовательности и осs-3-области. Фрагмент 4,1 тыс.п.о. вставляют в Sal I-переваренную рCGN 783 с получением плазмиды рВrx29 приблизительно 31 тыс.п.о.
Выявление экспрессируемой нитрилазы. Плазмиды рВrx28 и рВrx29 трансформируют в Agrobacterium tumefacium, штамм К12 (рВrx28) и К12 (рBrx29) используют для образования гель на каланхоэ.
Приблизительно 1 г (свежая масса) галовой ткани растирают в жидком азоте в буфере, содержащей 0,1 М Трис рН 7,5, 10 мМ ЭДТУК, 0,15 М NaCl, 0,05% NP-40% 25 мг/мл, ВSA, 1 мМ ДТТ И 0,13 мкг/мл лейпептина. Образцы гомогенизируют после добавления 0,05 г поливинилпирролидона, затем центрифугируют при 15000g в течение 15 мин при 4оС, 25 мкл антисыворотки, приготовленной инъекцией очищенной нитрилазы кроликам, и 250 мкл 10% (мас./об) суспензии S. aureus добавляют к каждой надосадочной жидкости и инкубируют в течение 16 ч при 4оС. Затем образцы центрифугируют и осадок дважды промывают 20 мМ Трис рН 7,5 1 мМ ЭДТУК, 150 мМ NaCl и 0,05% NP-40. Осадки повторно суспендируют в 100 мкл, 0,125 М Трис рН 6,8, 4% СДН, 20% глицерина и 10% BMе, и нагревают в течение 2 мин при 90оС. Образец в целом подвергают электрофорезу на 10% акриламидных гелях. Разделенные полипептиды переносят на нитроцеллюлозные фильтры, как это описано. Нитроцеллюлозные фильтры, затем инкубируют в "БЛОТТО" в течение 1-3 ч при 42оС с последующим инкубированием в течение ночи при комнатной температуре в "БЛОТТО", содержащей раствор 1:50 антинитрилазной сыворотки. Фильтры промывают в течение 10 мин в 20 мМ Трис рН 7,5, 150 мМ NaCl, в течение 20 мин в том же буфере, содержащем 0,05% "Твин-20" и в течение еще 10 мин в буфере без "Твин-20" "БЛОТТО", содержащую 106 счетов в мин/мл 125 1-меченного белка А (9 мккюри/сг NEN), затем добавляют к фильтрам и инкубируют при комнатной температуре в течение 2 ч. Фильтры промывают в течение ночи в 50 мМ Трис рН 7,5, 1 М NaCl и 0,4% саркозила. После прополаскивания и сушки фильтры экспонируют на рентгеновской пленке "Кодак" AR" при -70оС, используя интенсифицирующую сыворотку "Дюпон-Кронекс".
П р и м е р 6. Трансформация и регенерация табака.
Табачные растения культивируют следующим образом (25оС) белый свет (16 ч), МS (1 мг в л 1АА, 0,15 мг/л кинетина). Трехнедельные растения, сформировавшиеся по основному пробегу, используют в качестве тканевых доноров. Молодые листья отбирают, на перфораторе вырезают листовые диски диаметром 2 мм и их помещают в чашки Петри (диаметром 3 см) в 1 мл среды MS с 1 мг/л 1АА. После выдерживания дисков в течение ночи в полной темноте к этим культурам добавляют Agrobacterium (А772 х рN1 или рN2) клетки (108-109)/мл в растительной культуральной среде). Совместное культивирование проводят в течение 18-24 ч в темноте. Листовые срезы освобождают от Agrobacterium промывкой трижды М-средой без гормонов и содержащей 250 мг/л цефотоксина. Листовые срезы переносят в чашки Петри диаметром 9 см в 10 мл М-среды без гормонов. Чашки Петри типа "Фитагар" ("Джибко" 0,6% цефотоксин, 530 мг/л) герметизируют парафильмом и выдерживают при тех же условиях, что и растения-доноры ткани. Корни появлялись спустя 2-4 недели, их вырезают и помещают в те же условия в той же среде плюс 2 мг/л 1АА и 2 мг/л кинетина. Регенерирующие корни становятся видимыми в последующие 2-5 недель.
Растения опрыскивают на стадии 6 листа направлением струи раствора бромоксинила на растение в горшке. Каждый горшок размером 4 дюйма содержит растение и получает 2,5 мл раствора для опрыскивания. Растения выращивают в камере при 25оС, относительной влажности 70% на протяжении 60-часового светового периода. Состояние роста оценивают спустя 9 сут после опрыскивания подсчетом новых листьев длиной более 0,5 см.
Следуя описанным выше процедурам, получают растения, которые являются резистентным к бромоксинилу и могут использоваться на поле в присутствии бромоксинила, не испытывая значительного неблагоприятного воздействия на свой рост.
Предлагаемое изобретение обеспечивает улучшение растений благодаря приданию им резистентности к гербицидам, в частности к конкретным бензонитрильным гербицидам. Так, ген кодирующий нитрилазу, может быть введен в растение, в результате чего ген экспрессируется и придает растению резистентность к бензонитрилу. Кроме того, фермент может быть получен при клонировании гена в подходящем бактериальном хозяине, что обеспечивает экспрессию фермента.
Использование: биотехнология, в частности средства защиты клеток растений от вредного воздействия гербицидов. Сущность изобретения: конструируют рекомбинантную плазмидную ДНК pBr 28 или pBr 29, или p N 1 или p N 2, срезы табачного листа трансформируют полученной рекомбинантной ДНК в результате культивирования со штаммом Agrobacterium tumefaciens, культивируют трансформированные клетки и получают регенеранты табака. 2 табл.
СПОСОБ ПОЛУЧЕНИЯ РАСТЕНИЙ ТАБАКА, УСТОЙЧИВЫХ К 3,5-ДИГАЛОГЕН-4-ГИДРОКСИБЕНЗОНИТРИЛАМ, заключающийся в том, что конструируют рекомбинантную плазмидную ДНК pBrX28 или pBrX29, или pN1, или pN2, экспрессирующую нитрилазу, трансформируют срезы табачного листа данной плазмидой в результате культивирования со штаммом Agrobacterium tumefaciens, культивируют трансформированные клетки с последующим получением регенерантов табака.
| Аналогов в научно-технической и патентной литуратуре не обнаружено. |
