Настоящее изобретение относится к новому рекомбинантному фрагменту ДНК, используемому для придания растениям устойчивости к гербицидам на основе 3,5-дигалоид-4-оксибензонитрила, к способу трансформации растительных клеток с помощью этого фрагмента и к трансформированным растениям, регенерированным из этих клеток.
Из европейской заявки на патент 229042 известно придание растениям устойчивости к гербициду указанного выше типа, в частности к 3,5-дибром-4-оксибензонитрилу или бромоксинилу, путем введения в геном растения гена, кодирующего специфическую нитрилазу для деградации этих гербицидов. Хотя эта технология дает интересные результаты, она требует улучшения для увеличения шансов на успех и повышения его экономической ценности, в частности в том, что относится к уровню экспрессии в растениях и, следовательно, качеству устойчивости растений к этим гербицидам.
В настоящем описании под "растением" понимают любой многоклеточный дифференцированный организм, способный к фотосинтезу, а под "растительной клеткой" - любую клетку, происходящую из растения и могущую составлять недифференцированные ткани, такие как каллюс или зародыш, или дифференцированные ткани, такие как части растения, растения или семена.
Целью настоящего изобретения является удовлетворение этой потребности.
Оно относится к рекомбинантному фрагменту ДНК, используемому для придания растениям устойчивости к гербициду на основе 3,5-дигалоид-4-оксибензонитрила, состоящему из по крайней мере одного гена, кодирующего нитрилазу устойчивости к этому гербициду, чужеродного промотора и необязательно сигнальной зоны полиаденилирования, отличающемуся тем, что промотор происходит из гена, проявляющегося натурально в растительных клетках и выбранного в группе, состоящей из промотора dde РНК 35 S вируса мозаики цветной капусты /CamV 35S/ и промотора маленькой субъединицы /PSU/ 1,5-бис-рибулоза-фосфат карбоксилаза оксигеназы /Rubis CO/ подсолнечника /Heliauthus annuuas/.
Промотор фрагмента ДНК согласно изобретению происходит из гена, природно выражающегося в растениях, т.е. или нерастительного типа, например, вируса как РНК 35S вируса мозаики цветной капусты /CamV 35S/, или еще и предпочтительно растительного типа однодольных или двудольных растений, в частности маленькой субъединицы 1,5-бис-рибулоза-фосфат карбоксилаза окигеназы /Rubis CO/ подсолнечника /Heliauthus annuuas/. Можно использовать эти промоторы одни или в сочетании. Выбор зависит от природы трансформируемого растения /однодольное или двудольное/. Таким образом, предпочтительно использовать маленькую субъединицу Rubis CO подсолнечника для трансформации двудольного растения.
Каждый из этих промоторов может быть получен следующим образом.
1/ Промотор РНК 35S вируса мозаики цветной капусты /CamV 35S/.
Выделение промотора было описано Оделлом с сотр. /1985/. Выбирают клон /pJ05-2/, содержащий примерно 850 по выше сайта инициирования транскрипции, для описанных конструкций. Выделяют фрагмент EcoRI-Hind III, концы открыты полимеразой Кленова и клонируют в вектор pUC 19 /Янниш-Перрон с сотр. 1985/ в сайт Hind II. Этот клон переваривают под действием XbaI и Pst I и полученный фрагмент обрабатывают полимеразой фага T4, чтобы создать открытые концы. Этот фрагмент клонируют в pUC 19Cm /Баккли, 1985/, разрезают SmaI и XbaI и обрабатывают полимеразой Кленова. Полученный таким образом клон назван pRPA-BL 145. При обработке полимеразой Кленова сайта AccI 3'-конца и легировании с сайтом EcoRI, обработанным полимеразой Кленова, фрагментов, расположенных выше этого промотора, восстанавливают сайт EcoRI, и последовательность, полученная таким образом из сайта инициирования транскрипции, является следующей: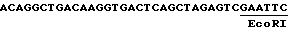
2/ Промотор маленький субъединицы 1,5-бис-рибулоза-фосфат карбоксилаза /Rubis CO/ подсолнечника /Heliauthus annuuas/.
Ген, из которого происходит этот промотор, был выделен Вакксманом с сотр. /1987/. Фрагмент EcoRI, содержащий промотор этого гена, был клонирован в mp 18, часть 3' промотора непосредственно перед полилинкером этого вектора. Затем этот клон линеаризуют BstXI и обрабатывают экзонуклеазой Bal31. Полученную таким образом смесь фрагментов обрабатывают SalI, потом полимеразой Кленова и наконец легируют при низкой концентрации ДНК. Клоны, полученные после такой манипуляции, секвенируют, и один из них имеет следующую последовательность за сайтом признанного начала транскрипции: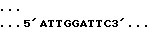
Линкер Cla I /ATCGAT/ был введен в сайт PstI этого клона. Итак, при обработке этого сайта Cla I полимеразой Кленова и легировании с сайтом EcoRI, обработанным полимеразой Кленова, фрагментов, которые помещены за этим промотором, воспроизводят сайт EcoRI, и последовательность, полученная таким образом из достоверного сайта начала транскрипции, является следующей: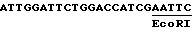
Согласно другому аспекту изобретения рекомбинантный фрагмент ДНК содержит нетранслируемую промежуточную зону /линкер/ между кодирующим геном и промотором, которая может быть выбрана в группе, состоящей из
с одной стороны, линкера pUC 19, модифицированного клонированием и имеющего следующую последовательность: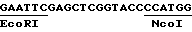 ,
,
с другой стороны, нетранслируемой зоны маленькой субъединицы Rubis CO кукурузы; эта зона происходит из кДНК, соответствующей гену, описанному Лебрюном с сотр. /1987/. Это фрагмент EcoRI-NcoI, который имеет следующую последовательность: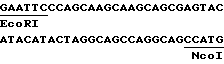 ,
,
с другой стороны, нетранслируемой зоны маленькой субъединицы Rubis CO подсолнечника; эта зона происходит из кДНК, выделенной Вакксманом и Фрейссинетом /1987/. Она не была выделена как таковая и всегда находится перед переходным пептида Rubis CO подсолнечника. Последовательность является следующей: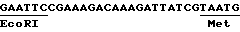
Рекомбинантный фрагмент ДНК
Химерический ген согласно изобретению включает необязательно зону или сайт полиаденилирования, который может быть, например
1/ сайтом полиаденилирования гена нопалин синтазы pTi 37/Беван с сотр., 1983/. Этот сайт содержится в фрагменте MboI 260 по /Фрели с сотр., 1983/, заявка на патент PCT 84 /02913/, который был обработан полимеразой Кленова и клонирован в сайт Smal M13 mp18 для введения сайтов BamHI и Eco RI соответственно на концы 5' и 3'. Сайт Bam HI был обработан нуклеазой Vigna vadiata и клонирован в сайт SalI, обработанный полимеразой Кленова pUC 19. Этот фрагмент содержит теперь на своем 5' конце сайт Hind III, который может быть легирован с фрагментом, расположенным на 3' конце гена нитрилазы.
2/ сайтом полиаденилирования гена маленькой субъединицы Rubis CO кукурузы: этот сайт был изолирован в виде фрагмента SmalI-BglII 540 по гена, описанного Лебрюном с сотр. /1987/. Линкер ClaI /ATCGAT/ был выведен на уровне сайта SmaI. После разреза Cla и заполнения полимеразой Кленова этот фрагмент был клонирован в pUC19, разрезанную PstI, потом обработан полимеразой фага T4 и разрезан BamHI. Эта манипуляция позволила ввести сайт Hind III B 5' сайта полиаденилирования. Полученная последовательность является следующей: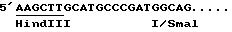 .
.
Согласно другому аспекту изобретения рекомбинантный фрагмент ДНК может необязательно и предпочтительно содержать между промежуточной зоной и геном нитрилазы зону, кодирующую переходный пептид, выбранный в группе, содержащей переходный пептид маленькой субъединицы Rubic CO кукурузы и переходный пептид маленькой субъединицы подсолнечника. Переходный пептид функционирует в природном гене, позволяя войти маленькой субъединицы Rubis CO в строму хлоропластов. Они должны направлять даже нитрилазу в этот участок в случае, когда их вводят между ранее описанной промежуточной зоной и структурным геном нитрилазы.
1/ Переходный пептид маленькой субъединицы Rubis CO кукурузы: этот фрагмент происходит из кДНК, соответствующей гену, описанному Лебрюном с сотр. /1987/. Этот фрагмент Hco I-5рпI 141 по, сайт HcoI, охватывающий кодон инициатора трансляции и сайт 5рпI, сайт разрыва переходного пептида. При обработке конца 5рпI этого фрагмента полимеразой фага 14 Т4 и легировании с HcoI концом обработанного полимеразой Кленова гена нитрилазы восстанавливают последовательность, позволяющую получить нитрилазу, не модифицированную в строме хлоропластов.
2/ Переходный пептид маленькой субъединицы Rubis CO подсолнечника: этот фрагмент происходит из кДНК, выделенной Вакксманом и Фрейссинетом /1987/. Эта последовательность не обладает источником сайти 5рпI на уровне разреза сайта переходного пептида. Последовательность на этом уровне является следующей:
5′CAATGCATGA*AG3
Заменяют направленным мутагенезом C на A, отмеченное звездочкой, создавая таким образом сайт Sph I. Для проведения этой манипуляции используют методику Золлера и Смита /1984/. Фрагмент EcoRI-SalI 27 по был клонирован в M13mp19am4. Этот вектор, происходящий из M13mp19, обладает амбр-мутацией в гене 4 по основанию 5327 и не может умножаться в штаммах, не обладающих суппрессором мутации этого типа. После очистки однониточной формы этого рекомбинантного фага три олигонуклеотида были гибридизованы в одну стадию. Последовательность этих фосфорилированных олигонуклеотидов является следующей: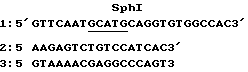
Они позволяют соответственно:
1: мутацию фрагмента на уровне сайта разреза переходного пептида,
2: коррекцию амбр-мутации,
3: начало последовательности выше мутированного фрагмента.
После одновременного действия полимеразы Кленова в присутствии четырех нуклеотидов и лигазы фага Т4 полученную смесь трансформируют в штамм HB2154, потом наслаивают монослоем на ковер HB2151 /Картер с сотр., 1985/. Среди полученных клонов те, что обладают дополнительным сайтом SphI, сиквенсировали, чтобы подтвердить структуру, и один из них был использован для создания фрагментов ДНК. На уровне сайта разреза переходного пептида теперь последовательность является следующей:
5′CAATGCATGC
Этот фрагмент используют по методике, идентичной методике, использованной для фрагмента, кодирующего переходной пептид маленькой субъединицы Rubis CO кукурузы.
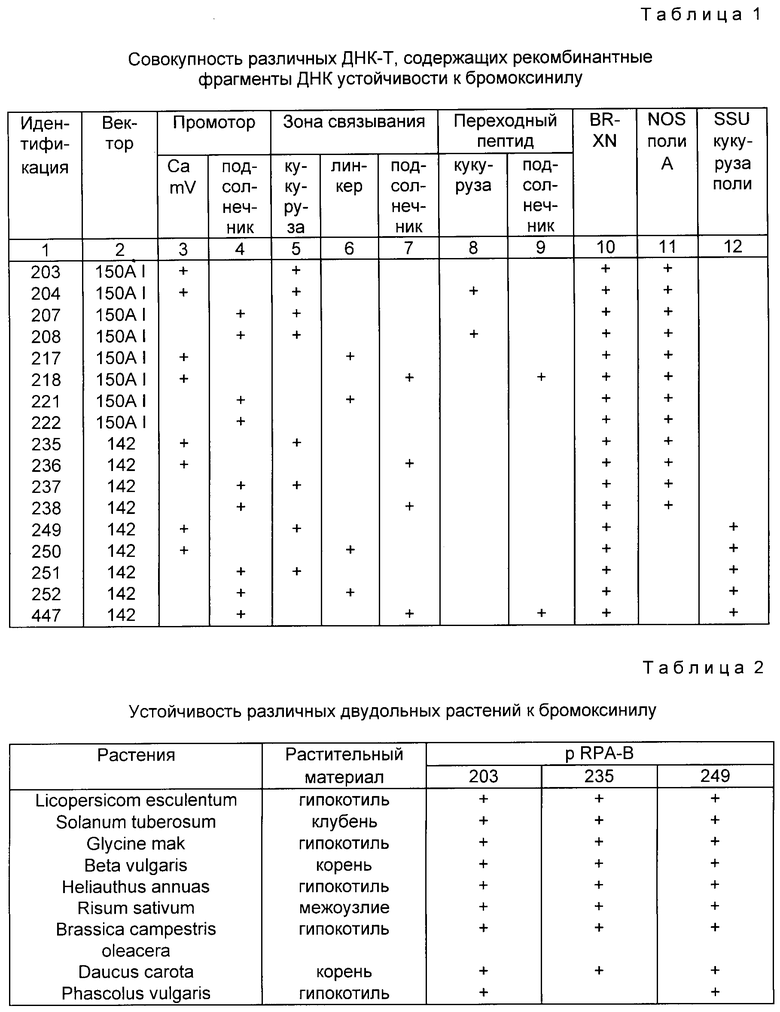
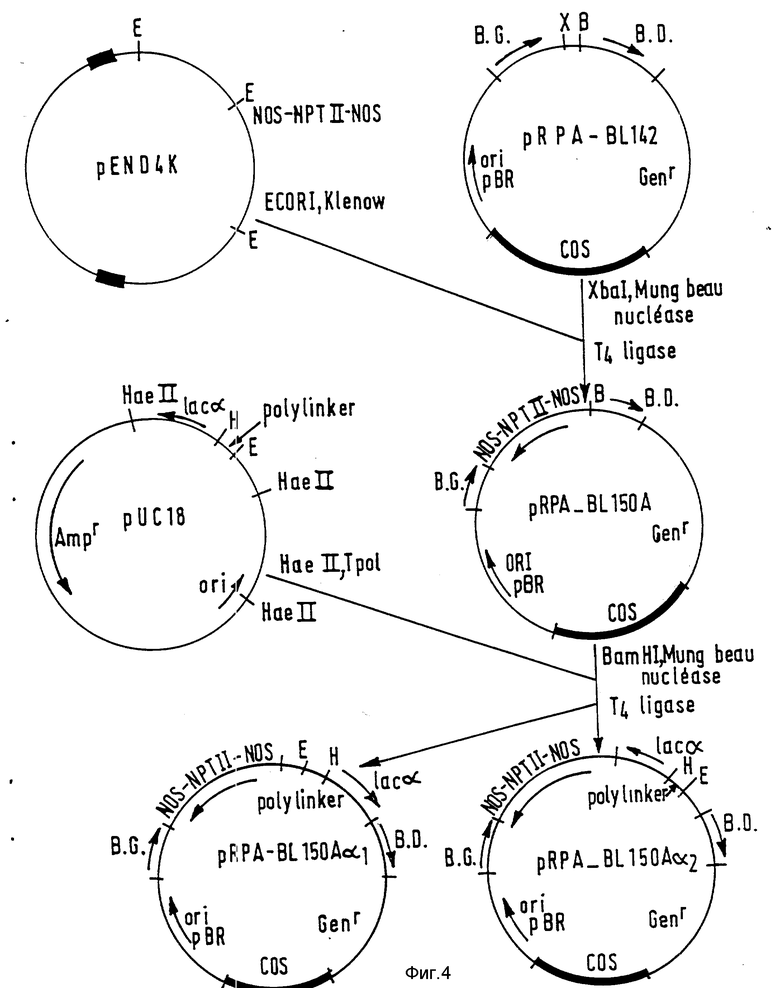
Объединение исходных фрагментов осуществляется согласно схеме на фиг. 1 - 4 с ранее описанными элементами. Созданные таким образом различные рекомбинантные фрагменты ДНК помещают в один или два типа векторов, и каждая совокупность была обозначена номером. Различные векторы, полученные таким образом, описаны в табл. 1.
Использованные векторы:
Различные химерические конструкции приводят к векторам pRPA-BL-142 и pRPA-BL-150Aa lphal, представленным на фиг. 1 - 4, они были введены в растения с помощью системы переноса Agrobacterium tumefacienc. Векторы переноса, сконструированные в данном случае, имеют следующие характеристики:
источник репликации и переноса, происходящий из pBR322,
ген бактериальной селекции, например устойчивости к гентамицину,
сайт COS, происходящий из фага лямбда,
два края, правый и левый, ДНК-Т pTiA6,
необязательно ген селекции эукариота, например устойчивости к канамицину,
необязательно фрагмент, содержащий ген комплементации lac альфа pUC18.
Конструкция pRPA-BL-142 /фиг. 1 - 4/
Правый и левый края /фиг. 1/ ДНК-Т левой pTiA6 вначале были целиком субклонированы.
Правый край:
фрагмент BamHI-EcoRI, лежащий от 13774 до 1602 по системе нумерации Баркера и др. /1983/, был клонирован в pGEM 1/Промега Биотеч/ в соответствующие сайты, данные pBL-17. Эта плазмида была переварена NruI /14276 и 14475/ и EcoRI /16202/ и обработана полимеразой Кленова. Лигация сайтов, заполненных NruI и EcoRI, регенерирует сайт EcoRI в 14276 и дает плазмиду pBL-19.
Левый край:
фрагмент Hind III, лежащий от 602 до 3390 в системе Баркера и др. /1983/, клонируют в соответствующий сайт pGEM I, получают pBL-21, в которой левый край находится напротив полилинкера. Эту плазмиду переваривает Acc I /1161 и 2687/ и обрабатывают полимеразой Кленова перед лигацией. Полученная в результате плазмида pBl-26 содержит фрагмент, лежащий от 602 до 1161, вставленный между сайтами Hind III и XbaI.
Создание ДНК-Т:
При введении фрагмента EcoRI-BamHI pBL-19 в соответствующие сайты pBL-26 реконструируют ДНК-Т, имеющую правый и левый края pTiA6 в их природной ориентации. Полученная плазмида обозначена pBl-70.
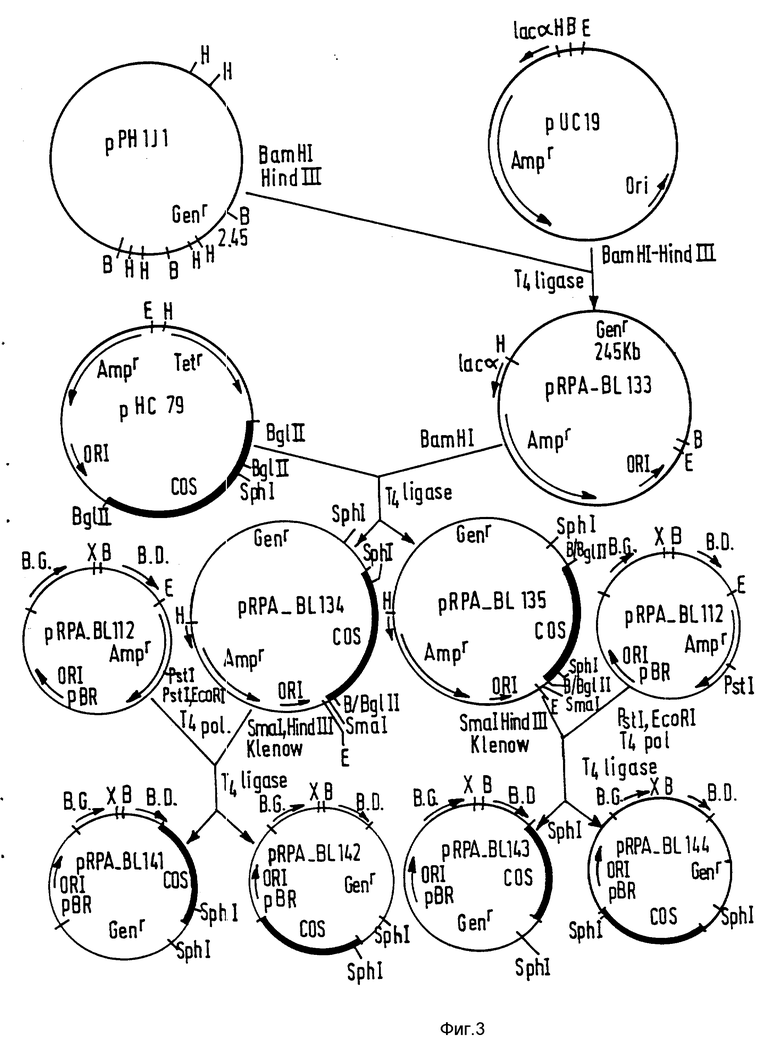
Введение ДНК-Т в pBR 322 /фиг. 2/:
После разреза pBL-70 c помощью Hind III этот сайт обрабатывают полимеразой Кленова и плазмиду снова разрезают EcoRI. Полученный фрагмент клонируют в pBR 322, разрезанный PmII-EcoRO. Полученный в результате клон обозначают pRPA-BL-112.
Клонирование гена устойчивости к гентамицину /фиг. 3/.
Ген устойчивости к гентамицину был получен из pPHI JII /Хирш и Брингер, 1984/. Эту плазмиду переваривают BamHI и Hind III и совокупность фрагментов клонируют в pUC19, разрезанную теми же ферментами. После селекции на ампициллин + гентамицин изолируют несколько клонов, содержащих фрагмент 2,45 кпо. Клон, полученный в результате манипуляций, назван pRPA-BL-133. Со стороны BamHI этого клона вводят фрагмент BglII, 1,6 ко, изолированный из pHC 79 /Хон и Коллинз, 1980/ и содержащий сайт COS фага лямбда. Этот фрагмент, вставленный в двух ориентациях, позволил получить два клона pRPA-Bl-134 и pRPA-Bl-135.
Получение интегрированного вектора /фиг. 3/.
Чтобы снова объединить в один и тот же вектор различные части, описанные ранее, плазмиды pRPA-Bl-134 и pRPA-Bl-135 переваривают Smal и Nind III и вставкой, содержащей ген устойчивости к гентамицину и сайт COS фага лямбда, обработанной полимеразой Кленова. Плазмиду pRPA-Bl-112 переваривают PstI и EcoRI и обрабатывают полимеразой фага Т4. Оба фрагмента легируют и отбирают клоны, содержащие одновременно устойчивость к гентамицину, сайт COS, ДНК-Т и источник репликации pBR 322. Плазмида pRPA-Bl-134 дает начало pRA-Bl-141 и pRPA-Bl-142, и плазмида pRPA-Bl-135 дает начало pRPA-Bl-143 и pRPA-Bl-144. Плазмида pRPA-Bl-142 была отобрана для введения рекомбинантных фрагментов ДНК, переносимых в растения. Исходя из этого вектора, была получена конструкция, содержащая маркерный ген NOS-NPTII-NOS /фиг. 4/.
Плазмиду pRPA-Bl-142 переваривают XbaI и восстанавливают концы действием нуклеазы Vigna radiata. Кроме того, pEND4 K /Кли и др., 1985/ переваривают EcoRI и обрабатывают полимеразой Кленова. Изолируют фрагмент 1,6 кпо, содержащий ген устойчивости к канамицину, и вводят в pRPA-Bl-142. Результат одного из этих слияний назван pRPA-Bl-15 A, и он был выбран для последующих манипуляций. Для облегчения клонирования в этот вектор вводят в сайт BamHI, обработанный нуклеазой Vigna radiata, фрагмент HacII, обработанный полимеразой фага Т4 и содержащий ген комплементации lac альфа, изолированный из pUC 18 /Янниш-Перрон и др., 1985/. Оба полученных вектора названы pRPA-Bl-150Aa lpha1 pRPA-Bl-150Aa lpha2. pRPA-Bl-150AalphaI служит основой для введения генов в растения.
Использование pRPA-Bl-142 и pRPA-Bl-150AalphaI
Эти векторы одни не сохраняются в Agrobacterium. Для сохранения их необходимо объединить простой рекомбинацией в плазмиду, находящуюся в этой бактерии. Это можно осуществить через фрагменты, такие, что сайт находится на космидах, таких как pVK 102 или других, или таких, как фрагмент pBR322 для плазмид, обладающих такими последовательностями. Это случай для плазмиды Ti штамма GV3850 /Замбриски и др., 1983/, который также является хозяином для pRPA-Bl-142 и pRPA-Bl-150AalphaI. Используя источник репликации pBR 322, эти плазмиды переносят в Agrobacterium с помощью трехчастной системы, описанной Диттом с сотр. /1980/.
Трансформация растительного материала
Для испытания эффективности этих химерических генов их переносят в материал в соответствии с методиками, описанными ниже.
А - Процедуры трансформации.
1. Табак
Вектор вводят в неонкогенный штамм Agrobacterium EHA 101 /Худ и др., 1987/, носитель космиды pTVK 291 /Комари и др., 1986/. Методика трансформации основана на процедуре Хорша и др. /1985/. Процедура регенерации промышленного табака РВД6 /провинция СЕЙТА, Франция/ описана ниже. Регенерация табака РВД6 из эксплантатов листьев осуществляют на среде на основе среды Мурасиге-Скуга /МС/, содержащий 30 г/л сахарозы. Листовые эксплантаты отделяют от растений, культивируемых в почве или in vitro, и трансформируют в соответствии с методикой листовых дисков /Science 1985, т. 227, с. 1229 - 1231/ в три последовательных этапа.
Первый этап заключается в индукции ростков на МС + 30 г сахарозы, содержащей 0,5 мг НУК и 2 мг/л БАП, в течение 15 дней. Второй этап позволяет развитие ростков, сформированных во время первого этапа; он осуществляется на МС + 30 г/л сахарозы, не содержащей гормона, в течение 10 дней. Третий этап позволяет укоренение ростков, отобранных индивидуально. Эта среда для укоренения содержит соли, витамины и сахар, разбавленные в половину. Она не содержит гормона. Через 10 - 15 дней укорененные ростки переносят в землю.
Определение гормонального баланса.
Оптимальная частота регенерации была получена при испытании 25 гормональных балансов БАП 0,2 - 0,5 - 1 - 1,5 - 2 и НУК 0 - 0,5 - 0 - 0,05, наблюдается для 1,5 и 2 мг/л БАП и 0,05 и 0,1 мг/л НУК. Была выбрана смесь 0,05 мг/л НУК и 2 мг/л БАП.
2. Другие двудольные.
Двудольные растения, представленные в табл. 2, были трансформированы с использованием онкогенного штамма Agrobacterium A281, носителя космиды pTVK291, ассоциированной с соответствующим вектором, используя растительный материал, такой как указано в табл. 2.
B - Измерение устойчивости к бромоксинилу
1. Табак
Устойчивость была измерена in vitro при выращивании каллюса в присутствии 20 мг/л бромоксинила в виде октоната и in vitro путем опрыскивания листьев бромоксинилом или оксинилом в дозах, предоставляющих 10-кратную дозу, рекомендованную для обработки в открытом поле. Полученные результаты приведены в табл. 3.
2. Другие двудольные растения
Устойчивость была измерена in vitro при выращивании каллюса в присутствии 10 мг бромоксинила /в виде октоната/ на 1 л среды. Во всех случаях результаты были положительными /табл. 2/.
Использование: изобретение относится к созданию рекомбинантного фрагмента ДНК для придания растениям устойчивости к гербицидам на основе 3,5-дигалоид-4 оксибензонитрила. Сущность: рекомбинантный фрагмент ДНК содержит кодирующий ген и гетерологический промотор, при этом необязательно фрагмент ДНК между кодирующим геном и промотором может содержать нетранслируемый линкер или сайт полиаденилирования, а гетерологический промотор представляет собой промотор 35S PHK вируса мозаики цветной капусты (Cam V 35S) и/или промотор маленькой субъединицы (PSU), 1,5-бис-рибулоза-фосфат карбоксилазы оксигеназы подсолнечника (Rubis CO). 2 с. и 3 з.п. ф-лы, 4 ил., 3 табл.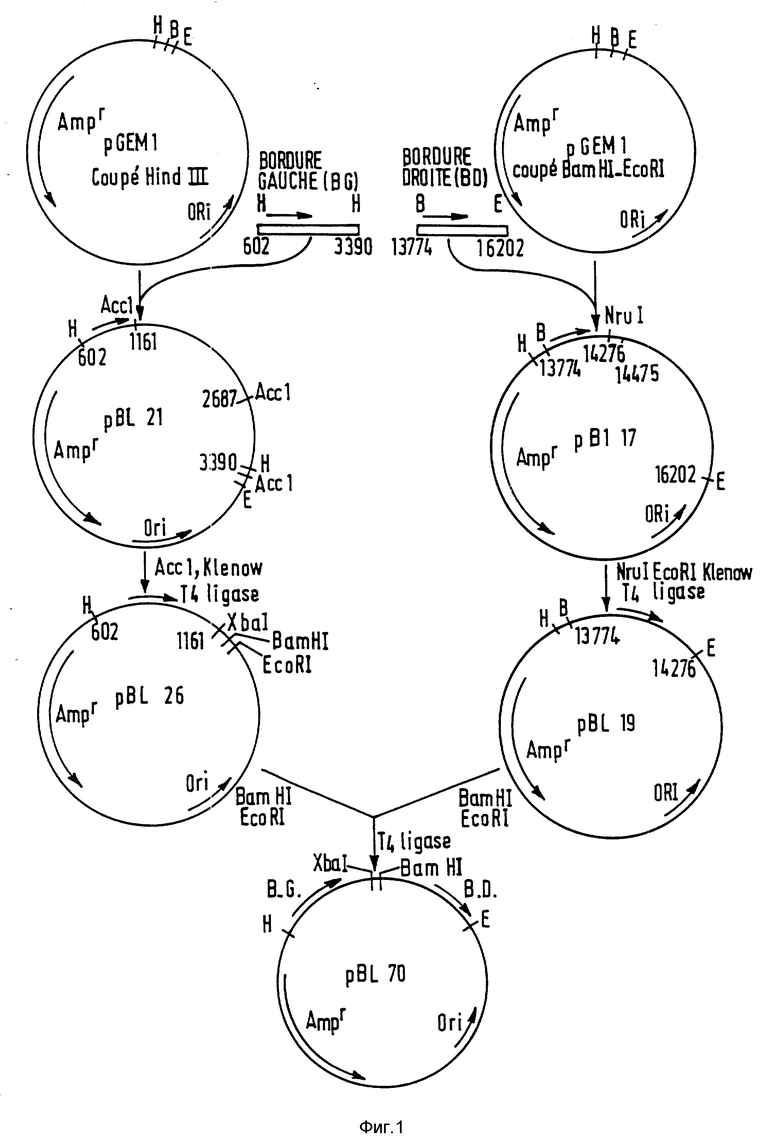
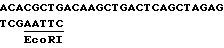
и/или промотор маленькой субъединицы (PSU) 1,5-бис-рибулоза фосфат карбоксилаза оксигеназы подсолнечника (Rubis CO), имеющий сайт инициации транскрипции с последовательностью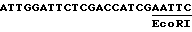
2. Рекомбинатнтный фрагмент ДНК по п.1, отличающийся тем, что между кодирующим геном и промотором содержит нетранслируемый линкер, имеющий последовательность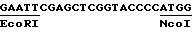
или последовательность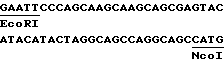
или последовательность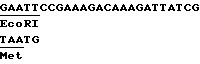
3. Рекомбинантный фрагмент ДНК по п.1, отличающийся тем, что содержит сайт полиаденилирования.
ori pBR 322;
ген устойчивости к гентамицину плазмиды pPHI J II;
cos - сайт фага λ ;
2Т ДНК рTiА6.
Ori pBR 322;
ген устойчивости к гентамицину плазмиды pPHI J II;
cos - сайт фага λ ;
Т ДНК рTiА6;
ген устойчивости к канамицину плазмиды рЕ ND4;
маркерный ген NOS- N PTII - NOS.

