Изобретение относится к микроорганизмам, трансформированным новой гибридной плазмидой, содержащей фрагмент ДНК, обеспечивающий использование бетаина, и к способам получения, применения этих микроорганизмов.
Из молекулярной микробиологии известно, что для получения некоторых соединений используют микроорганизмы, трансформированные рекомбинантной плазмидой, в которую встроены гены, кодирующие эти соединения.
В этом случае одна из проблем состоит в том, чтобы сохранить плазмиды в клетках микроорганизмов, поскольку при их делении может происходить бесконтрольное распределение плазмид в дочерние клетки.
Это обстоятельство приводит к тому, что во время ферментационных процессов образуется большое количество дочерних клеток, не содержащих плазмиды.
В лабораторном масштабе на этот недостаток можно воздействовать вводя в культуральную среду антибиотики, соответствующие гену резистентности, включенному в плазмиды. Однако при крупномасштабных ферментационных процессах добавление соответствующих антибиотиков вряд ли оправдано, поскольку некоторые антибиотики (например, тетрациклин) оказывают нежелательный эффект на способность микроорганизмов, содержащих плазмиды, к росту, делению и размножению (Bioscience Reports, 5, 1985, pp.29-37; Gene, 39, 1985, pp. 173-180). Другой недостаток стабилизирующего действия антибиотиков состоит в том, что их введение, особенно в случае крупномасштабных ферментационных процессов, слишком дорогостоящая операция. Кроме того, использование антибиотиков при производстве фармацевтических средств, а также пищевых и фуражных добавок нежелательно или недопустимо.
Другой метод, благодаря которому удается противодействовать потере плазмид, описан H. Sakoda и T. Imanaka (J. Ferment, and Bioeng. v.69, 1990, рp. 75-78). Он заключается в том, что вначале осуществляют делецию триптофанового оперона в хромосоме хозяина, вследствие чего клетка не усваивает триптофан. Затем клетку-хозяин трансформируют рекомбинантной плазмидой, содержащей этот оперон. Трансформированные клетки в дальнейшем выявляют по использованию триптофана.
Недостаток данного метода состоит в том, что из-за выделения триптофана вследствие диффузии в ходе ферментационного процесса могут возникать и размножаться клетки, не содержащие плазмид, что ведет к увеличению количества дочерних клеток, не содержащих плазмид.
Задача изобретения состоит в том, чтобы устранить указанные выше недостатки и получить микроорганизмы с плазмидами, построенными таким образом, чтобы они могли стабильно сохраняться благодаря дешевым и легко доступным веществам в ходе всего процесса ферментации, при этом способность к росту, делению и размножению микроорганизмов, содержащих плазмиду, должна быть гарантирована.
Эта задача решается с помощью описанных в изобретении микроорганизмов, содержащих рекомбинантную плазмиду с фрагментом ДНК, включающим генетическую последовательность, кодирующую использование бетаинов и характеризующуюся следующей рестрикционной картой (см.фиг.1)б и мутацию в хромосомном гене, кодирующем использование бетаина.
Далее под термином "бетаины" следует понимать такие соединения, как бетаин, холин, диметилглицин и сакрозин.
Получение микроорганизмов, стабильно сохраняющих плазмиды
I. В хромосомном гене микроорганизмов, утилизирующих бетаин, осуществляют мутацию с получением штамма, дефектного по использованию бетаина.
II. Выделяют фрагмент ДНК, содержащий генетическую последовательность, кодирующую использование бетаина.
III. Выделенный фрагмент ДНК клонируют в вектор экспрессии:
IV. с получением рекомбинантной плазмиды.
V. Трансформируют микроорганизм (клетку хозяина), полученный на стадии II, данной плазмидой и получают штамм, стабильно сохраняющий плазмиду.
V. Эти трансформированные микроорганизмы представляют устойчивый в отношении использования бетаина штамм, поскольку рекомбинантные плазмиды дополнительно содержат ген, кодирующий использование бетаина.
Мутация в хромосоме у микроорганизмов, использующих бетаин
В качестве микроорганизмов, использующих бетаин, могут служить микроорганизмы, способные расти и размножаться, при утилизации бетаина или бетаинов как единственного источника углерода, азота и энергии.
Примерами таких микроорганизмов могут служить Pseudomonas sp. Rhizobium/Agrobacterium sp. или Rhizobium sp.
Наиболее целесообразно использовать семейство Rhizobium/Agrobacterium. Предпочтительно использовать микроорганизмы Rhizobium/Agrobacterium sp. НК1349 DSM N 3944. Под этим номером указанный штамм был зарегистрирован 4.11.1991 в Германской коллекции микроорганизмов и клеточных культур ГмбХ, Машеродервег 1б, D-3300 Брауншвейг.
Следует отметить, что при идентификации изолята НК4 был использован комплекс фенотипических, хемотаксономических методов, а также технология нуклеиновых кислот.
Хроматографический анализ показал наличие у изолята НК4 убихинона с 10 изопреноидными единицами на поверхности цепи (Q-10), что подтверждает его принадлежность к альфа- субклассам Proteobacteria (Woese et а1. 1984, System. Appl. Microbiol. 5: 315; Stackerbrandt et al. 1988, Int. J. Syst. Bacteriol. 38: 321-325).
Количественные анализы полиаминов с помощью жидкостной хроматографии высокого разрешения у данного изолята (Busse and Auling, 1988, System. Appl. Microbiol. 11: 1-8) дали следующие результаты (данные представлены в микромолях на 1 г сухого веса): путрецин 0,7; спермидин 1,2; сим- спермидин 22,4. Структура этих полиаминов позволяет утверждать, что штамм НК4 относится к одному из субклассов Proteobacteria.
Множественная тестовая система AP120NE позволила идентифицировать изолят НК4 как Agrobacterium radiobacter. Однако присущая изоляту НК4 структура полиаминов, о которой сообщалось выше, отличается от структуры последних.
Исследования по индукции опухоли у Kalanchoe daigremonata изолятом НК4 были отрицательными.
Анализ фрагмента 16s рРНК 1220-1380 (Escherichia coli номенклатура) подтвердил, что изолят НК4 является членом альфа-2 субклассов Proteobacteria.
Частичное определение последовательности фрагмента изолята НК4 показало ее идентичность последовательности соответствующего фрагмента Rhizobium meliloti, "Pseudomonas" aminovorans (Green Gillis, 1989, J. Gen. Microbiol. 135: 2071-2076) изолята TE9, утилизирующего нитрилтриацетат (Egli et al. 1988, System Appl. Microbiol. 10: 297-505), и изолята FLB 300, способного разлагать фторбензонат (Engesser et al. 1990, Arch. Microbiol. 153: 193-199). Растворимые белки этих изолятов подвергались SDS-полиакриламидному гель электрофорезу с белками НК4 для обнаружения какого-либо сходства на субгенетическом уровне.
Фингерпритинг белков показал наличие большого сходства между изолятами НК4 и FLB 300.
Мутацию в хромосомном гене, кодирующем использование бетаина (в последующем именуется как beu), осуществляли методами, известными любому специалисту. Примерами таких методов могут служить
делеция вследствие гомологической рекомбинации;
мутагенез с помощью вещества, изменяющего структуру ДНК или встраивание транспозона.
При использовании микроорганизмов типа Rhizobium/Agrobacterium НК1349 делецию гена осуществляют методом гомологической рекомбинации.
Для этой цели сначала выделяют фрагмент ДНК, содержащий хромосомный ген beu, и клонируют его в так называемую вспомогательную плазмиду. Затем из этой "вспомогательной" плазмиды выделяют фрагмент ДНК, кодирующий beu.
При использовании так называемой "вспомогательной" плазмиды с делецией с помощью обычных для специалиста методов осуществляют делецию в хромосоме за счет обмена с помощью гомологичной рекомбинации (Mol. Gen. Genet. 210, 1987, pp.381-381; J. Bacteriol. 171. 1989, pp.4617-4622).
В результате этого микроорганизм Rhizobium/Agrobact. sp. НК1349 (DSM 3944) преобразуется в дефектный по гену beu микроорганизм НК 1349.4.
II. Выделение фрагмента ДНК, содержащего beu-ген
Источником фрагмента ДНК, содержащего beu-ген, могут служить описанные в разделе I виды микроорганизмов.
Предпочтительно, если в качестве такого источника использовать микроорганизм Rhizobium/Agrobacterium НК1349, зарегистрированный в коллекции микроорганизмов и клеточных культур под N 3944.
Вначале осуществляют локализацию фрагмента ДНК, содержащего генетическую последовательность, кодирующую использование бетаинов, на хромосоме Rhizobium/Agrobacterium НК1349.
Локализацию проводят с помощью тривиальных методов, например, путем инсерции транспозона в соответствующую хромосому микроорганизма. При этом искомый фрагмент ДНК маркируется с помощью транспозона.
Для выявления мутации с маркированным фрагментом ДНК применяют метод, подтверждающий отсутствие утилизации бетаинов как источника углерода, азота и энергии.
Фрагмент ДНК, содержащий инсерцию транспозона, выделяют с помощью рестриктазы EcoRI. Полученные при этом фрагменты клонируют с помощью обычных методов в плазмиду для Е.coli.
Образующиеся рекомбинантные плазмиды, отобранные по устойчивости к антибиотикам, содержат маркированный транспозоном фрагмент EcoRI-фрагмент ДНК размером 18,2 kb (12,5 kb и 5,7 kb для соответствующего транспозона).
Для выделения (не маркированного транспозоном) фрагмента ДНК, содержащего ген beu, выделяют ДНК Rhizobium/Agrobacterium НК1349. Затем выделенную ДНК полностью гидролизуют с помощью фермента EcoRI и отделяют. После этого EcoRI-фрагмент ДНК размером 12,0-13,0 kb клонируют в Е. coli с применением обычных методов. Путем конъюгации различных клонов с немаркированным транспозоном beu-геном в дальнейшем выделяют клоны, содержащие немаркированный ген. При этом рекомбинантная плазмида присутствует в Е. coli.
В результате дополнительных опытов с субклонами (клонами, содержащими делецию в различных участках EcoRI-фрагмента ДНК), был идентифицирован и выделен PstI-фрагмент ДНК, содержащий генетическую последовательность, кодирующую использование бетаинов размером 3 kb.
Этот фрагмент ДНК является предметом изобретения и характеризуется следующей картой рестрикции (см. фиг.1)
Этот фрагмент ДНК находится в составе рекомбинантной плазмиды pL032 и депонирован в коллекции микроорганизмов в штамме НК1349.4 под N 6712.
III. Встраивание фрагмента ДНК beu-гена в вектор экспрессии
Полученный при помощи традиционной методики фрагмент ДНК beu-гена лигируют с предварительно полученным вектором экспрессии с получением рекомбинантной плазмиды.
Обычно векторы экспрессии содержат подходящий промотор (активатор, контролирующий экспрессию). Оптимально, если за промотором в направлении транскрипции имеется один или несколько сайтов рестрикции для рестриктаз. В них впоследствии встраивается фрагмент ДНК, экспрессия которого представляет интерес.
Для получения описанных в изобретении рекомбинантных плазмид можно применять векторы экспрессии с широким спектром хозяев. Примеры таких векторов:
рКТ240 (Gene, 26, 1983, pp.273-282)
pME285 (Gene, 36, 1985, pp.27-36)
pVK100 (Gene, 8, 1982, pp.45-54)
Вектор экспресси рКТ240 обычно обрабатывают рестриктазой PstI и образующиеся при этом концы лигируют с заявленным фрагментом ДНК, например, с помощью Т4-ДНК лигазы.
IV. Рекомбинантные плазмиды
Изобретение относится к рекомбинантным плазмидам, содержащим фрагмент ДНК beu-гена.
В принципе могут быть использованы любые рекомбинантные плазмиды, способные экспрессировать бетаин.
Однако наиболее удачным для этой цели можно считать рекомбинантную плазмиду pL032, состоящую из фрагмента ДНК beu-гена и вектора экспрессии рКТ240, в котором фрагмент ДНК beu-гена расположен в направлении транскрипции к промотору Pbla, определяющему устойчивость к ампициллину.
Эта рекомбинантная плазмида хранится в Германской коллекции микроорганизмов и клеточных культур ГмбХ, Машеродервег 1б, D-3300 Брауншвейг в составе микроорганизма НК1349.4 (N 6712).
На фиг.2 представлена физическая карта рекомбинантной плазмиды pL032.
V. Трансформация
Полученными таким образом рекомбинантными плазмидами трансформируют микроорганизмы, полученные на стадии I.
Трансформированные микроорганизмы также являются частью данного изобретения.
Трансформацию микроорганизмов рекомбинантными плазмидами осуществляли с помощью известных в микробиологии методов. Выделение трансформированных микроорганизмов производили на питательной среде, в которую был добавлен бетаин как источник углерода или азота. При трансформации рекомбинантной плазмидой pL032 выделение или селекцию трансформированных микроорганизмов осуществляли на питательной среде, в которую добавлен бетаин как источник углерода или азота. После трансформации получали микроорганизмы НК1349.4, содержащие рекомбинантную плазмиду pL032 (N 6712).
Для стабильного сохранения плазмид в этих микроорганизмах в культуральные среды добавляли бетаины в концентрации 0,2 0,4% Культуральной средой при этом могут служить обычные в таких случаях среды, например среда минеральных солей по данным Кулла и соавт. (Arch. Microbiol. 135, 1983, pp.1-7).
VI. Получение штамма, стабильно сохраняющего плазмиду с фрагментом, обеспечивающим использование бетаина
Изобретение раскрывает применение фрагмента ДНК для получения плазмид, содержащих этот фрагмент, а также использование микроорганизмов для стабильного хранения этих плазмид. Таким образом, полученные штаммы также являются составной частью изобретения.
Эти штаммы могут быть получены путем транформации микроорганизмов, содержащих мутацию в хромосомном гене, кодирующем использование бетаина рекомбинантной плазмидой с фрагментом ДНК beu-гена.
Эти плазмиды получают с помощью тривиальных методов. При этом beu- ген может быть введен в рекомбинантные плазмиды, содержащие другой ген, путем встраивания фрагмента ДНК с геном beu в плазмиду, которая уже содержит какой-либо ген.
Оптимальным можно считать вариант, когда применяется рекомбинантная плазмида, которая содержит дополнительный ген, выделенный с помощью рестриктазы PstI, а затем соединенный с PstI-фрагментом ДНК beu-гена в составе этой плазмиды.
В случае, если используемая плазмида уже имеет фрагмент ДНК beu- гена, при встраивании другого фрагмента ДНК с beu-геном обработку осуществляют рестриктаз, в результате чего оттесняется в сторону содержащийся в плазмиде ген.
После такого преобразования микроорганизмы культивируют в среде с бетаинами при сохранении количества плазмид.
В качестве стабильно сохраняемой плазмиды предпочтительно использовать плазмиду pL032, содержащую дополнительный ген. Эта плазмида соответствует по своим характеристикам рекомбинантной плазмиде pL032. Этой плазмидой обычно трансформируют микроорганизм НК1349.9.
Примером штамма, стабильно сохраняющего плазмиду, может служить микроорганизм НК1349.4, содержащий плазмиду pLOL01. Для ее получения использовали плазмиду pL03, описанную в Европейском патенте 0477828. Плазмида pL03 состоит из вектора экспрессии рКТ240 и генов xyIMA (кодирующих фермент ксилолмонооксидазу). Эту плазмиду обработали рестриктазой PstI и лигировали с фрагментом ДНК beu-гена. При этом конечная плазмида соответствует плазмиде pL032 с той лишь разницей, что в отличие от нее имеет дополнительные xyIMA гены.
После встраивания pLOL01 в микроорганизмы НК1349.4 этот штамм становится способным трансформировать 2,5-диметилпиразин в 5-гидроксиметилпиразин в присутствии бетаина без потери плазмид.
Пример 1. Получение мутантов, способных вводить транспозон, и их фенотипическая идентификация
В результате селекции штамма Agrobacterium/Rhizobium sp. НК1349 (N по каталогу НК1349) в присутствии стрептомицина получают клетки, устойчивые к стрептомицину (1000 мкг/мл). Устойчивость сохранялась неизменной в течение более 50 генераций без селекции и могла рассматриваться как селекционный маркер.
0,2 мл культуры донора Tn5 Е. coli S 17-1/pSUP 2021, устойчивая к неомицину (R.Simon et al. Biotechnology, 1983, 1, pp.784-790), смешивают с 2 мл культуры реципиента НК1349 и центрифугируют.
Клеточную культуру промывают 0,9%-ным раствором соли (раствор NaCl) и повторно суспендируют в 100 мкл 0,9%-ного раствора соли. Конъюгация реципиента с донором происходила в течение ночи при 30oС на сухом агаре в качестве питательной среды. Вызревшие клетки в разбавленном состоянии высевают на пластину с селекционной средой для реципиента и транспозона.
Tn5-мутанты НК1349 получают путем селекции на агаре со стрептомицином (1000 мкг/мл) и неомицином (100 мкг/мл) в качестве питательной среды. Фенотипическую идентификацию производили по неиспользованию бетаинов как источника углерода или азота в среде с минеральными кислотами (Kulla et al. Arch. Microbiol. 1983, 135. pp.1-7).
Пример 2. Клонирование фрагмента ДНК, маркированного Tn-5 из НК1349-генома
Выделенный по известной методике фрагмент хромосомной ДНК (J. Моl. Biol, 1979, 130, рр.161-173) с Tn-5-мутацией НК1349 (5 мкг) полностью гидролизуют с помощью EcoRI (1 ед/мкг). После завершения процесса 2,5 мкг плазмиды обрабатывают раствором щелочной фосфатазы (0,1 ед/1-20 рмоль ДНК терминальной дефосфорилазы). Рекомбинантные плазмидные ДНК получают путем смешивания геномной ДНК и pBR325 с Т4-ДНК-лигазой (0,2 ед/мкг ДНК) в 400 мкл буфера (20 мМ трис-HCl, рН 7,2, 10 мМ дитиоэритритола ДДТ), 10 мМ MgCI2, 0,6 мМ аденозинтрифосфата (АТФ). Культуры выдерживают в течение ночи при 12oС. Аликвотные количества лигированной смеси используют в ходе эксперимента по трансформации (Cohen et al. Proc. National Acad. Sci. US, 1972, 96, pp.2110-2114) с применением Е. coli ЕД8654.
Отбор трансформантов производят по их устойчивости к ампициллину (100 мкг/мл, pBR325) и канамицину (25 мг/мл за счет введения Tn-5) на агаре в качестве питательной среды.
Все гибридные плазмиды содержат фрагменты (12,5 kb + 5,7 kb для Tn5), то есть маркированные Tn5.
При более точном картировании выявлено, что транспозон находится в том же самом геномном фрагменте на том же самом месте, что и фрагмент, отвечающий за фенотип Beu, т.е. неактивный beu ген, полученный при введении транспозона Tn5 (отобранные Tn5-мутанты). Далее эти мутанты именуются как НК4УII.
Пример 3. Клонирование фрагмента ДНК beu (немаркированного) из НК1349-генома
В соответствии с примером 2 выделяют и полностью гидролизуют с помощью рестриктазы EcoRI (4 ед/мкг) ДНК НК1349 с последующим отделением путем гель-электрофореза на агарозе. Большие фрагменты ДНК размером 12,0- 13,0 kb (маркированный фрагмент имел величину 12,6 kb) выделяют путем электрофореза на агарозе. Выделенную ДНК по аналогии с примером 2 обрабатывают EcoRI и также в соответствии с примером 2 лигируют с дефосфорилированным вектором рУК100 (Plasmid, 1982, 8, pp. 45-54). Аликвотные количества лигированной смеси используют по аналогии с примером 2 в эксперименте по трансформации в Е. coli S 17-1 (Biotechnol. 1983, 1, pp.1784-1791).
Производят отбор трансформантов на их устойчивость к тетрациклину (25 мкг/мл) и канамицину (25 мкг/мл).
Полученные трансформанты путем конъюгации с НК4УII с мутацией за счет транспозона (методом "patch mating") в качестве реципиентного штамма анализируют на введение заданного фрагмента ДНК beu-гена. Устойчивые к антибиотикам трансформанты отбирают с использованием образцов на питательной среде из агара с канамицином 2 мкг/мл. Параллельно пластины из агара засевают штаммом-реципиентом НК4У11, выделенным из газонной травы.
Для конъюгации трансформанты в виде отдельных клонов закрепляют на реципиентных штаммах, выделенных из клеток пророщенной травы, и выдерживают в течение ночи при 30oС.
И, наконец, для отбора полученных трансконъюгантов пластины маркируют в указанной в примере 1 среде, содержащей соли минеральных кислот с бетаином (0,2 мас.) в качестве субстрата, который не использовался донором и реципиентом. В более крупных по размеру трансконъюгантах реципиента (НК4УII) с геномным фрагментом ДНК, содержащим мутацию, комплементируют соответствующий неактивный фрагмент ДНК за счет введения гибридной плазмиды из штамма донора или подвергают гомологичной рекомбинации. Комплементарные гибридные плазмиды получили название pУK100s. Для предотвращения ревертантности штамма НК4УII в среду вводят неомицин (100 мкг/мл). С помощью гибридизации с клонированным, маркированным Tn5 фрагментом ДНК подтверждают успешное клонирование комплементарного фрагмента с неактивным геном beu из генома НК1349 в гибридной плазмиде рУК100s.
Пример 4. Идентификация субфрагмента ДНК, кодирующего бетаин
Путем клонирования фрагментов, полученных при использовании различныхных ферментов (Bg1II, XhoI, SphI, PstI), в гибридной плазмиде рУК100 с последующей комплементацией beu Tn5-мутанта НК4УII идентифицируют фрагмент ДНК размером 3 kb на плазмиде, кодирующей использование бетаина.
Этот фрагмент характеризуется следующей физической картой (см.фиг.1):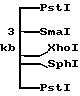
Пример 5. Стабильная мутация beu в геноме НК1349
Поскольку мутация с введением Tn при селекции с помощью антибиотиков или без них очень нестабильна, к штамму НК1349 добавляют стабильную делецию.
Введение делеции осуществляют с применением метода гомологичной рекомбинации.
Полученный в примере 3 EcoRI фрагмент размером 12,5 kb клонируют в суицидный вектор рАСУС184 (J. Bacteriol. 1978, 134, pp.1141-1156) и из него в результате обработки рестриктазой (1 ед/мг) получают фрагмент ДНК, кодирующий beu, размером 3 kb. Повторную лигацию проводят в течение ночи с применением 1 ед/мкг Т4-ДНК-лигазы.
Гибрид с делецией встраивают в Е. coli НВ101/рРК2013 и из него с помощью конъюгации переносят в НК1349. Полученные трансконъюганты подвергают селекции в отношении ауксотрофного роста донора (Pro пролинотрицательный) и устойчивости плазмиды к антибиотикам в среде, содержащей соли минеральных кислот, 0,4% глюкозы и 25 мкг/мл тетрациклина.
Только клетки с фрагментом, интегрированным в плазмиду в процессе гомологичной рекомбинации, обнаруживают устойчивость к тетрациклину и проявляют способность к росту и размножению в указанной среде.
Для выделения второго продукта рекомбинации вектора рАСУС184 и неактивного beu-гена, выделенного из хромосомы НК1349, трансконъюганты культивируют в той же среде без селекции в присутствии тетрациклина в течение более 100 генераций.
Чтобы увеличить количество чувствительных к тетрациклину beu- мутантов, проводят селекцию на интегрированный в вектор рАСУС184 неактивный beu-ген.
Для этой цели клетки вводят в 25 мл комплексной среды NYB (Oxoid, Wesel, BRD), содержащей тетрациклин (10 мкг/мл) и выдерживают в течение 6 ч при 30oС.
Затем к культуре добавляют 0,5 мг/мл D-циклосерина и 15 мг/мл пенициллина G. Выдерживают при 30oС в течение 84 ч, клетки центрифугируют, трижды промывают свежим NYB и высевают при соответствующем разбавлении на агар в качестве питательной среды.
18% полученных колоний обладают повышенной чувствительностью к тетрациклину, из них третья часть одновременно неактивна в отношении использования бетаинов. Правильность наличия делеции beu подтверждают гибридизацией с фрагментом 12,5 kb гибридной плазмиды pVK100s (из примера 3). Маркируют только один укороченный фрагмент EcoRI размером 3 kb.
Полученный таким образом НК1349.4 с мутацией комплементируют с помощью Pstl-фрагмента с геном beu размером 3 kb, клонированным в подходящем векторе. Описание векторов дано в примере 6.
Пример 6.
а) Клонирование beu-гена в различные векторы "с широким спектром клеток- хозяев"
В известные "векторы с широким спектром хозяев" рКТ240, рМЕ285, рУК100 клонируют PstI фрагмент размером 3 kb, кодирующий beu. (Current Protocols in Molecular Biology, John Wiley and Sons, New York, 1989, Abschnitt 3.16, Subcloning of DNA-Fragments). При этом существенную роль играет правильная ориентация фрагмента по отношению к промотору. В случае рКТ240 имеет значение его правильное расположение (расположение в направлении транскрипции) по отношению к промотору Pbla (промотор гена bla, определяющий устойчивость к ампициллину).
б) Введение beu-гена в рКТ240
рКТ240 линеаризуют с помощью PstI (1 ед/мкг). Линеаризованную ДНК лигируют с фрагментом размером 3 kb с помощью Т4-ДНК-лигазы (1 ед/мкг) в буфере (20 ммоль трис-HCI, рН 7,2, 10 ммоль ДТТ, 10 ммоль MgCI2, 0,6 ммоль АТФ).
Лигирование проводят в течение ночи при 12oС.
Затем полученную смесь конъюгируют согласно методу Lederberg und Cohen (J. Bacteriol. 1974, 119, pp.1072-1074) с Е. coli S.17-1. Селекцию проводят в среде NYB с канамицином (25 мкг/мл) по отношению к ампициллину (100 мкг/мл).
Гибридные плазмиды, содержащие фрагмент (чувствительность к ампициллину, устойчивость к канамицину), расположенный в направлении транскрипции от промотора, получили название pL032.
в) Конъюгация pL032 в НК1349.4
Конъюгированный перенос pL032 из Е. coli S.17-1 в НК1349.4 осуществляют указанными методами. Селекцию НК1349.4, содержащего гибридную плазмиду pL032, производят в присутствии бетаина (среда, содержащая соли минеральных кислот, в соответствии с примером 1, и 0,2 мас. бетаина).
Пример 7. Стабильность pL032 в НК1349.4
Продолжительность сохранения гибридной плазмиды в микроорганизмах НК1349.4 испытывают в различных средах. Оценку осуществляют по использованию бетаина (или холина и диметилглицина) в качестве единственного источника углерода. Стабильность плазмиды составила 100%
При снижении количества азота (например, в не содержащих азот средах рециклических культур) стабилизацию при использовании бетаина как единственного источника азота оценивали в 100%
Пример 8. Устойчивость гибридной плазмиды, обладающей способностью к биотрансформации
В качестве примера стабильно сохраняющейся плазмиды в beu- отрицательном штамме-хозяине НК49.4 следует назвать гибридную плазмиду pL03, описанную в Европейском патенте А-0477828.
При этом речь идет о гибридной плазмиде, состоящей из вектора рКТ240 и ClaI-HindIII-фрагмента (2,35 kb) TOL-плазмиды, которая содержит xyIMA- гены, клонированные под контролем промотора канамицинфосфотрансферазы.
а) Введение нового фрагмента ДНК, обеспечивающего устойчивость к канамицину (KmR)
Фрагмент ДНК, обеспечивающий устойчивость к канамицину (1,1 kb) (Harayama et al. J. Bacteriol. 1986, 167. pp.455-461), вырезают из pRMEI с помощью EcoRI (4 ед/мкг ДНК) и выделяют с помощью электрофореза на агарозном геле. 5'-выступающие концы фрагмента ДНК заполняют в ходе реакции Кленова (Current Protocols in Molecular Biology, John Wiley, New York, 1987, Abschnitt 3.5). рL03-ДНК обрабатывают HpaI рестриктазой (1 ед/мкл ДНК) и дефосфорилируют 4,8 ед. щелочной фосфатазой. После осаждения изопропанолом "тупой конец" вектора на срезе лигируют с "тупым концом" КmR-фрагмента в течение ночи пр 15oС (буфер: 20 ммоль трис, 10 ммоль MgCI2, 0,6 ммоль АТФ, рН 7,2, 10% полиэтиленгликоль 6000, 0,5 ед Т4-ДНК-лигазы).
Е. coli К12 трансформируют лигированной смесью по аналогии с примером 2 и полученные рL03-(KmR-трансформанты селектируют на агаре (в качестве питательной среды) с 50 мкг/мл Km.
б) Введение beu-гена в pL03 (KmR)
Гибридную плазмиду pL03 (KmR) "линеаризуют" (см. пример 6) с использованием PstI (1 ед/мкг ДНК) и лигируют с PstI-фрагментом beu-гена размером 3 kb. Трансформацию в Е. coli и конъюгацию в НК1349.4 производят в соответствии с примером 6. Гибридная плазмида получила название pLOL01 и соответствовала pL032 с дополнительной ксилолмонооксидазной активностью (xyIMA).
в) Биотрансформация устойчивой гибридной плазмидой
Agrobacterium/Rhizobium HK1349.4/pLOL01 помещают в среду, содержащую соли минеральных кислот (Arch. Microbiol. 1983, 135, pp.1-7), с 0,2% бетаина в качестве единственного источника углерода при 30oС. Выявляют биотрансформацию 0,1% (об/об) 2,5-диметилпиразина в 5- гидроксиметил-2-метилпиразин. Выход 5-гидроксиметил-2-метилпиразина через 2 дня составляет 20%
г) Стабильность pLOL01 в НК1349.4
По аналогии с примером 7 испытывают период стабильного сохранения гибридной плазмиды в микроорганизме НК1349.4. При использовании бетаина как единственного источника углерода и азота стабильность плазмиды в штамме, даже в условиях биотрансформации, была на уровне 100%



Использование: биотехнология. Сущность изобретения: разработан способ получения штамма бактерий, предназначенного для стабильного хранения плазмиды с фрагментом ДНК, обеспечивающим использование бетаина из субстрата путем осуществления мутации в хромосомном гене штамма бактерий Rhizobium /Agrobacterium с образованием дефектного по использованию бетаина штамма конструирования рекомбинантной плазмидой ДНК pL 032 с фрагментом ДНК, обеспечивающим использование бетаина из субстрата, имеющим указанную рестриктную карту. Последующий перенос рекомбинантной плазмидной ДНК в штамм, содержащий мутацию с последующим выращиванием трансформированного штамма бактерий Rhizobium /Agrobacterium DSM N 6712. Рекомбинантная плазмидная ДНК рL 032. Фрагмент ДНК размером 3 kb, имеющий указанную карту рестрикции. 4 c.и 2 з.п. ф-лы, 1 табл., 2 ил.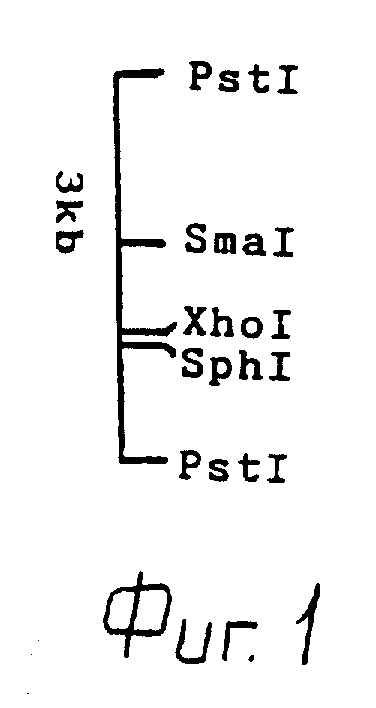

2. Рекомбинантная плазмидная ДНК pL 032, с фрагментом ДНК, обеспечивающим использование бетаина, содержащая Pst Pst фрагмент размером 3 kb с рестриктной картой, представленной в п. 1; ДНК вектора рКТ240; уникальные сайты рестрикции: SstI, HpaI, EcoRI, ClaI и HindIII; регуляторные элементы: Pb l a промотор гена устойчивости к ампициллину; генетические маркеры: Smr ген устойчивости к стрептомицину; Kmr ген устойчивости к канамицину.

