Изобретение относится к биотехнологии, в частности к генетической инженерии, ветеринарии и медицине, и представляет собой: последовательность ДНК, несущественную для репликации вируса и содержащую ген тимидинкиназы (ТК-ген), из генома вируса оспы кур штамма "К", и имеющую первичную нуклеотидную последовательность, приведенную ниже; рекомбинантные плазмидные ДНК pTKlFPV и pTK2FPV, содержащие полученную последовательность ДНК; рекомбинантный плазмидный вектор pINTFPVl, обладающий способностью интегрироваться в геном вируса оспы кур.
Вирус оспы кур является представителем рода Avipoxvirus семейства Poxviridae. Оспа кур, заболевание вызываемое вирусом оспы кур, является высококонтагиозным заболеванием кур. Распространена болезнь повсеместно. Экономический ущерб заключается в резком и продолжительном снижении яйценоскости, вынужденном убое больных и переболевших птиц. Для специфической профилактики оспы кур используют живые вакцины двух типов, приготовленные из аттенуированных иммунногенных и ареактогенных штаммов гетерологичного вируса оспы голубей или вируса оспы кур [1].
Авипоксвирусы обладают многими свойствами, характерными для всех поксвирусов: геном состоит из линейной двухспиральной ДНК размером до 300 kb, репликация вируса происходит в цитоплазме инфицированных клеток, вирионы обладают неикосаэдрической морфологией [2]. В отличие от других поксвирусов авипоксвирусы обладают узким кругом хозяев: продуктивная инфекция происходит только в организме птиц или в культурах клеток, полученных из тканей птиц. Попытки получить продуктивную инфекцию авипоксвирусов в организме млекопитающих или в культурах клеток млекопитающих не увенчались успехом [3]. Из других свойств авипоксвирусов следует также отметить термоустойчивость и простоту наработки [1].
Стратегия получения рекомбинантных авипоксвирусов основана на подходе, хорошо зарекомендовавшего себя при получении рекомбинантных вирусов осповакцины. Суть данного подхода заключается в том, что на первом этапе конструируют гибридную плазмиду, содержащую гетерологичный ген, фланкированный последовательностями ДНК вируса осповакцины из несущественной области вирусного генома. Полученную плазмиду используют для введения в клетки, инфицированные вирусом осповакцины. За счет рекомбинации по областям гомологии происходит встройка чужеродной последовательности в геном вируса. Затем определенным образом в популяции вирусного потомства можно выявить гибридные вирусы [4].
Известно несколько районов в составе генома авипоксвирусов, пригодных для интеграции чужеродных генов: ген тимидинкиназы [5], район терминальных инвертированных повторов [6], район, гомологичный гену тимидинкиназы вируса осповакцины [7]. Самым распространенным способом отбора рекомбинантных авипоксвирусов является использование гена β-галактозидазы E.coli, что позволяет быстро идентифицировать рекомбинанты, используя хромогенный субстрат. Одной из наиболее распространенных конструкций для плазмиды интеграции является конструкция, предложенная в работе [7]. Особенностью этой конструкции является наличие двух противоположно направленных промоторных элементов внутри фланкирующих последовательностей. Причем под контролем одного из промоторов находится ген β-галактозидазы, что позволяет легко и быстро идентифицировать полученные вирусные рекомбинанты, а под контролем второго промотора находится исследуемый чужеродный ген.
В конце 80-х годов начались работы по получению рекомбинантных авипоксвирусов. В первую очередь были получены рекомбинантные вирусы оспы кур, содержащие гены вирусов, представляющих интерес для птицеводства: вируса болезни Марека [8, 9], вируса болезни Ньюкастла [6], вируса болезни Гамборо [10] и др. В опытах на птицах полученные рекомбинанты показали хорошие иммуннопротективные свойства, сравнимые с действием имеющихся коммерческих вакцин. В работе [3] было проведено сравнение использования рекомбинантных и имеющихся коммерческих вакцин и сделан вывод, что наилучший эффект дает комбинированное использование обоих типов вакцин. В этой работе также сделан вывод о необходимости получения рекомбинантных вирусов оспы кур, содержащих в своем геноме гены двух и более вирусов, то есть прототипов поливалентных вакцин.
Практически одновременно были начаты работы по получению рекомбинантных авипоксвирусов, содержащих в своем геноме гены вирусов, вызывающих заболевания у млекопитающих и человека. Были получены рекомбинантные вирусы оспы кур, содержащие в своем геноме гены вирусов бешенства и кори, показавшие хорошие протективные свойства в опытах на мышах и крысах [11 - 13]. В работе [14] было показано, что рекомбинанты, полученные на основе вируса оспы канареек, более эффективны в опытах на животных, чем аналогичные рекомбинанты, полученные на основе вируса оспы кур. Впоследствии все исследования по получению и изучению свойств рекомбинантных авипоксвирусов в экспериментах на млекопитающих проводятся с использованием вируса оспы канареек в качестве вирусного вектора. Из последних работ можно отметить работу, в которой показано успешное применение рекомбинантных авипоксвирусов при вакцинации против чумы плотоядных в опытах на собаках [15]. Представляют интерес работы, в которых сообщается об использовании рекомбинантных авипоксвирусов в иммуннотерапии бешенства [16], рака [17], СПИДа [18], а также в качестве продуцентов интерлейкина, интерферона, фактора некроза опухолей [19]. Этот краткий литературный обзор позволяет сделать вывод, что использование рекомбинантных авипоксвирусов может найти применение в птицеводстве, ветеринарии и медицине.
В России до настоящего времени не проводились какие-либо исследования, связанные с получением и изучением рекомбинантных авипоксвирусов отечественных штаммов.
Технической задачей изобретения является конструирование вирусного вектора интеграции на основе вируса оспы кур штамма "К", используемого в России для специфической профилактики оспы кур, с целью создания поливалентных вакцин для птицеводства, а также в ветеринарии и медицине в качестве эффективного эукариотеческого вектора.
Поставленная задача решается путем амплифицирования и клонирования фрагмента генома вируса оспы кур штамма "К", несущественного для репликации вируса и содержащего ТК-ген, и конструирования на его основе плазмидного вектора, обладающего способностью интегрироваться в геном вируса оспы кур.
Для этого: 1. Выделяют геномную ДНК из вируса оспы кур штамма "К". 2. Методом ПЦР получают последовательность ДНК, содержащую ТК-ген и несущественную для репликации вируса, используя специфические праймеры. 3. Используя полученную последовательность ДНК, с помощью генно-инженерных манипуляций получают рекомбинантные плазмиды pTKlFPV и pTK2FPV, содержащие полученную последовательность ДНК в виде двух примерно одинакового размера фрагментов, содержащих 5'- и 3'-концевые части ТК-гена с прилегающими участками генома вируса оспы кур. 4. Определяют первичную нуклеотидную последовательность полученной последовательности ДНК, используя плазмидные ДНК pTKlFPV и pTK2FPV. 5. Получают плазмидный вектор pINTFPVl при помощи генно-инженерных манипуляций, используя плазмиды pTKlFPV и pTK2FPV и другие вспомогательные плазмиды. 6. Проводят трансфекцию плазмидным вектором pINTFPVl культуры клеток фибробластов эмбрионов кур, зараженных вирусом оспы кур штамма "К". Используя хромогенный субстрат X-gal, из вирусной популяции выделяют рекомбинантные вирусы.
Сущность предлагаемых объектов изобретения состоит в следующем:
1. Последовательность ДНК, несущественная для репликации вируса и содержащая ТК-ген, из генома вируса оспы кур штамма "К" имеет первичную нуклеотидную структуру, приведенную на фиг.3. Она получена из генома вируса оспы кур штамма "К" посредством генной амплификации с помощью полимеразной цепной реакции, в которой использовались 4 синтетических одигонуклеотида, обозначенные как ТК11, ТК12, ТК21, ТК22 и имеющие следующие последовательности:
ТК11 - 5 AAGCTTCGCCACGTTATTCGAA - 3;
ТК12 - 5 GCGTGAAATGCTATGTCGACG - 3;
ТК21 - 5 ACTCGTTTAGAAATCGATGCGTC - 3;
ТК22 - 5 GCCAAGTACGCGCGATAAAG - 3.
Они представляют собой элемент новизны. Олигонуклеотиды ТК11, ТК12, ТК21 имеют в своем составе сайты рестрикции: HindIII, SalI, ClaI, которых нет в полученных фрагментах. Полученная последовательность ДНК для удобства получена в виде двух фрагментов, которые в дальнейшем будут использованы как фланкирующие последовательности при сборке плазмиды интеграции.
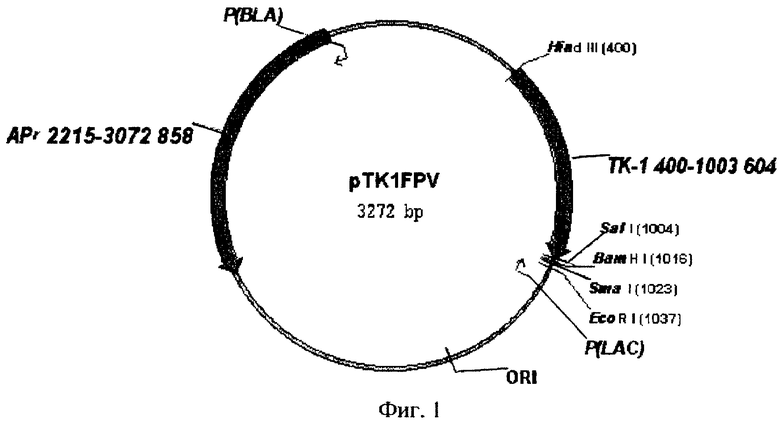
2. Рекомбинантная плазмидная ДНК pTKlFPV, содержащая 3'-конец ТК-гена вируса оспы кур и характеризующаяся следующими признаками:
имеет молекулярную массу 2,16 мегадальтон размером 3272 п.о.;
содержит амплифицированный фрагмент размером 604 п.о., обработанный эндонуклеазами рестрикции HindIII и SaiI, и кодирующий N-концевую часть тимидинкиназы вируса оспы кур;
содержит HindIII-SalI - фрагмент плазмидного вектора pUC-18, имеющего размер 2668 п.о.;
содержит участок начала репликации;
содержит ген устойчивости к ампициллину;
содержит уникальные сайты для эндонуклеаз рестрикции, имеющие следующие координаты: HindIII-400, SalI-1004, BamHI-1016, SmaI-1023, KpnI-1029, SacI-1037, EcoRI-1037.
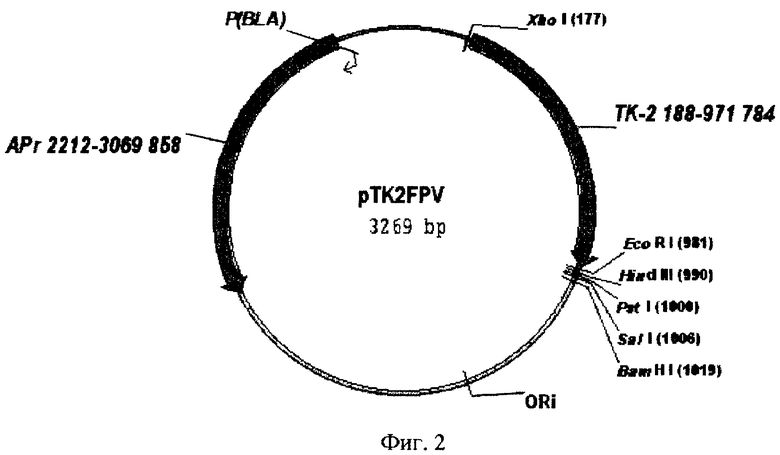
3. Рекомбинантная плазмидная ДНК pTK2FPV, содержащая 5'-конец ТК-гена вируса оспы кур и характеризующаяся следующими признаками:
имеет молекулярную массу 2,16 мегадальтон размером 3269 п.о.;
содержит амплифицированный фрагмент размером 780 п.о., обработанный эндонуклеазой рестрикции ClaI и кодирующий C-концевую часть тимидинкиназы вируса оспы кур;
содержит ClaI-SmaI фрагмент плазмиды pMTL-22 размером 2497 п.о.;
содержит участок начала репликации;
содержит ген устойчивости к ампициллину;
содержит уникальные сайты для эндонуклеаз рестрикции, имеющие следующие координаты: XhoI-177, Bg1II-181, ClaI-188, EcoRI-981, HindIII-990, PstI-1000, SalI-1006, BamHI-1019.
4. Рекомбинантая плазмидная ДНК pINTFPVl, обладающая способностью интегрироваться в геном вируса оспы кур и характеризующаяся следующими признаками:
имеет молекулярную массу 4,87 мегадальтон размером 7388 п.о.;
содержит HindIII-EcoRI фрагмент плазмиды pUC-18 размером 2635 п.о.;
содержит участок начала репликации;
содержит ген устойчивости к ампициллину;
содержит амплифицированный фрагмент размером 604 п.о., обработанный эндонуклеазами рестрикции HindIII и SalI и кодирующий N-концевую часть тимидинкиназы вируса оспы кур;
содержит амплифицированный фрагмент размером 780 п.о., обработанный эндонуклеазами рестрикции ClaI и кодирующий C-концевую часть тимидинкиназы вируса оспы кур;
содержит два SalI-BamHI фрагмента плазмиды pBRPZ размером 110 п.о., содержащих промотор 11К вируса осповакцины;
содержит BamHI-KpnI фрагмент плазмиды pUC-128 размером 66 п.о., включающий уникальные сайты для эндонуклеаз рестрикции, BamHI, SmaI, PstI, EcorI, HindIII, SalI, XhoI, ApaI, KpnI;
содержит BamHI-SalI фрагмент плазмиды pBRPZ размером 3000 п.о., включающий ген β-галактозидазы E.coli, обладающий способностью экспрессировать β-галактозидазу в составе генома вируса оспы кур;
содержит уникальные сайты для эндонуклеаз рестрикции, имеющие следующие координаты: BamHI-1113, SmaI-1121, Pst-1129, EcoRI-1131, HindIII-1143, SalI-1158, XhoI-1164, ApaI-1177, KpnI-2183.
Таким образом, при помощи генно-инженерных манипуляций из генома вируса оспы кур штамма "К" получена последовательность ДНК, несущественная для репликации вируса и содержащая ТК-ген; определена первичная нуклеотидная структура полученной последовательности ДНК; получены рекомбинантные плазмиды pTKlFPV и pTK2FPV, содержащие полученную последовательность ДНК; сконструирован плазмидный вектор pTNTFPVl, обладающий способностью интегрироваться в геном вируса оспы кур. Полученное техническое решение может найти применение при получении генно-инженерных вакцин на основе вируса оспы кур, которые могут найти применение в ветеринарии и медицине.
Фиг.1. Физическая карта рекомбинантной плазмиды pTKlFPV.
Фиг.2. Физическая карта рекомбинантной плазмиды pTK2FPV.
Фиг. 3. Первичная нуклеотидная структура полученной последовательности ДНК, содержащей ТК-ген и несущественной для репликации вируса, из генома вируса оспы кур штамма "К". Цветом выделен ТК-ген.
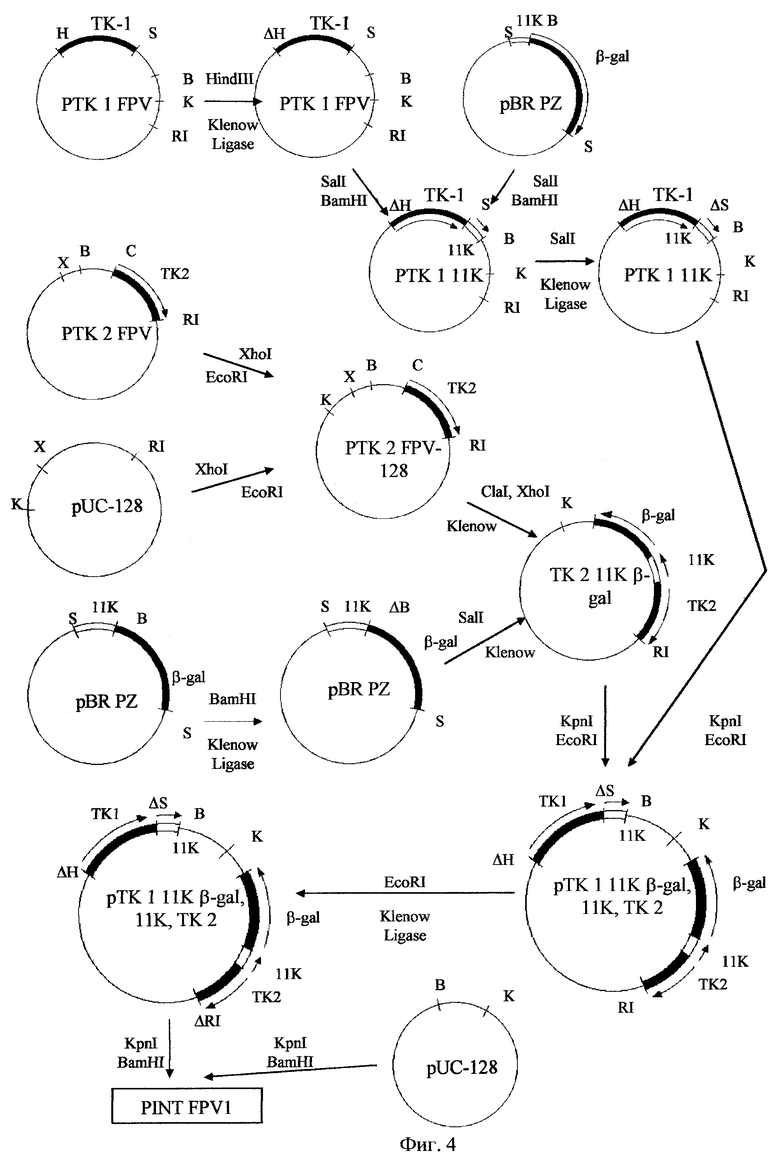
Фиг.4. Схема сборки плазмидного вектора pINTFPVI.
Фиг.5. Физическая карта рекомбинантного вектора pINTFPVI.
Пример 1. Получение последовательности ДНК, несущественной для репликации вируса и содержащей ТК-ген, из вируса оспы кур штамма "К".
В качестве вирусного вектора был выбран вирус оспы кур штамма "К", используемый в России для специфической вакцинации против вируса оспы кур. Штамм был получен из ВГНКИ Ветпрепаратов. Вирус нарабатывают, очищают по методике, описанной ранее в работе [21]. Вирусную ДНК выделяют по методике, описанной в работе [22]. Праймеры для проведения ПЦР рассчитывают по программе OLIGO [23] с учетом ранее опубликованных нуклеотидных последовательностей генов тимидинкиназы авипоксвирусов [2, 20]. Структуры праймеров приведены ниже:
ТК11 - 5 AAGCTTCGCCACGTTATTCGAA - 3;
TK12 - 5 GCGTGAAATGCTATGTCGACG - 3;
ТК21 - 5 ACTCGTTTAGAAATCGATGCGTC - 3;
ТК22 - 5 GCCAAGTACGCGCGATAAAG - 3.
ПЦР проводят по стандартной методике [24]. Амплифицированный фрагмент, полученный при проведении ПЦР с праймерами ТК11 и ТК12, обрабатывают рестриктазами HindIII и SalI. Из полученного гидролизата выделяют в 4% полиакриламидном геле фрагмент длиной примерно 600 п.о., что совпадает с рассчитанной длиной амплифицированного фрагмента 604 п.о. Амплифицированный фрагмент, полученный при проведении ПЦР с праймерами ТК11 и ТК12, обрабатывают рестриктазой ClaI. Из полученного гидролиза выделяют в 4% полиакриламидном геле фрагмент длиной примерно 780 п.о., что совпадает с рассчитанной длиной 780 п. о. Полученные таким образом амплифицированные фрагменты далее были использованы для получения рекомбинантных плазмидных ДНК pTK1FPV и pTK2FPV.
Пример 2. Получение рекомбинантных плазмидных ДНК pTKlFPV и pTK2FPV, содержащих полученную последовательность ДНК.
1. 10 мкг плазмидной ДНК pUC-18 обрабатывают рестриктазами HindIII и SalI по общепринятой методике [25]. Из полученного гидролизата выделяют векторный фрагмент размером 2680 п.о. в 4% полиакриламидном геле праймеров. Амплифицированный фрагмент длиной 604 п.о. и векторную часть плазмиды pUC-18 сшивают при помощи лигазной реакции в 20 мкл буфера для лигирования [25]. 10 мкл лигазной смеси используют для трансформации компетентных клеток E.coli JM-109 [25]. Трансформанты высевают на LВ-агар, содержащий 100 мкг/мл ампициллина, 5 мкг/мл Х-gal, 5 мкг/мл IPTG. Из белых клонов выделяют плазмидную ДНК и анализируют рестрикционным анализом [25]. Рекомбинантная плазмидная ДНК, содержащая фрагмент нужной длины, будет обозначена как pTK1FPV и в дальнейшем будет использована для определения нуклеотидной последовательности полученного амплифицированного фрагмента и для сборки плазмиды интеграции. Физическая карта плазмиды pTKlFPV приведена на фиг.1.
2. 10 мкг плазмидной ДНК pMTL-22 [26] обрабатывают рестриктазами ClaI и SmaI. Из полученного гидролизата выделяют в 4% полиакриламидном геле векторный фрагмент размером 2497 п.о. 780 п.о. Амплифицированный фрагмент длиной 780 п.о. и векторную часть плазмиды pMTL-22 сшивают при помощи лигазной реакции в 20 мкл буфера для лигирования. 10 мкл лигазной смеси используют для трансформации компетентных клеток E.coli JM-109. Трансформанты высевают на LB-агар, содержащий 100 мкг/мл ампициллина, 5 мкг/мл X-gal, 5 мкг/мл IPTG. Из белых колоний выделяют плазмидную ДНК и анализируют рестрикционным анализом. Рекомбинантные клоны, содержащие фрагмент нужной длины, будут обозначены как pTK2FPV и в дальнейшем будут использованы для определения первичной структуры полученного амплифицированного фрагмента и для сборки плазмиды интеграции. Физическая карта плазмиды pTK2FPV приведена на фиг.2.
Пример 3. Определение первичной нуклеотидной структуры полученной последовательности ДНК.
Первичную нуклеотидную структуру полученной последовательности ДНК определяют по методу Максама-Гилберта [25]. Для секвенирования используют по 3 независимых клона. Полученная последовательность приведена на фиг.3. Сравнение полученной первичной структуры с опубликованными ранее показало, что полученная последовательность ДНК содержит ген тимидинкиназы вируса оспы кур, причем 5'-конец гена тимидинкиназы расположен в амплифицированноv фрагменте длиной 780 п.о., а 3'-конец гена - в амплифицированном фрагменте длиной 604 п.о.
Пример 4. Получение плазмидного вектора интеграции pINTFPVl.
Для получения плазмидного вектора pINTFPVI используют плазмиды pTKlFPV и pTK2FPV, а также вспомогательные плазмиды pBRPZ [27] и pUC-128. Все генно-инженерные манипуляции проводят аналогично, как описано в примере 2, и согласно общепринятым методикам [25]. Схема сборки плазмиды pINTFPVI приведена на фиг.4. Физическая карта плазмиды pINTFPVI - на фиг.5.
Пример 5. Получение рекомбинантного вируса оспы кур, содержащего ген β-галактозидазы E.coli.
Плазмидной ДНК pINTFPVI трансфецируют культуру клеток фибробластов эмбрионов кур, зараженную вирусом оспы кур штамма "К". Трансфекцию и отбор клонов проводят по методике, описанной ранее [11]. Полученные рекомбинантные вирусные клоны обладают способностью окрашивать культуральную среду при добавлении хромогенных субстратов: X-gal, Y-gal и др. Наличие рекомбинантных клонов доказывает правильность сборки плазмиды интеграции, а также возможность использовать полученную плазмиду для конструирования рекомбинантов, содержащих целевые чужеродные гены.





Изобретение относится к биотехнологии, в частности к генетической инженерии, ветеринарии и медицине. Путем амплификации и клонирования фрагмента генома вируса оспы кур отечественного штамма "К" получают последовательность ДНК, несущественную для репликации вируса и содержащую TK-ген. Используя полученную последовательность ДНК, с помощью генно-инженерных манипуляций получают рекомбинантные плазмиды pTK1FPV и pTK2FPV, содержащие 5'- и 3'-концевые части ТК-гена. При помощи этих и других вспомогательных плазмид получают плазмидный вектор pINTFPV1, способный интегрироваться в геном вируса кур. Полученное техническое решение может найти применение при получении генно-инженерных вакцин на основе вируса оспы кур, которые могут быть использованы в ветеринарии и медицине. 4 с.п. ф-лы, 5 ил.
| Приспособление в пере для письма с целью увеличения на нем запаса чернил и уменьшения скорости их высыхания | 1917 |
|
SU96A1 |
| Устройство для контроля однородности физических параметров среды | 1972 |
|
SU443335A1 |
| Звукотеплоизолирующая панель | 1974 |
|
SU652287A1 |

