Это изобретение относится к терапевтическим агентам, предназначенным для использования в лечении аутоиммунных заболеваний млекопитающих, в особенности человека. Изобретение также относится к терапевтическим агентам, применяемым в лечении лейкемий человека Т-клеточного происхождения, как так называемые "вакцинные носители", и агентам, использующимся для предупреждения у человека отторжения трансплантата и при заболевании трансплантат против хозяина (GVHD).
В статье под названием "Morfologic and Functional Alterations of Mucosal Т-сells by Cholera Toxin and its В-subunit" by Charles O. Elson et al. The Journal of Immunology, 1995, 154; 1032-1040 обнаруживается, что холерный токсин (Ctx) и CtxB-субъединица ингибируют CD8+ и CD4+ Т-клетки.
Также делались ссылки на статью под названием "Prevention of Acute Graf-Versus-Host Disease by Treatment with a Novel Immunosuppressant" by B. Yankelvich et al. The Journal of Immunology, 1995, 154; 3611-3617. В ней CtxB определяется как агент, предназначенный для использования при трансформации костного мозга для предупреждения острого заболевания трансплантат против хозяина (GVHD).
WO 95/10301 раскрывает иммунологические агенты, индуцирующие толерантность и содержащие молекулы, связанные со слизистой оболочкой и сцеплением со специфическим толерогеном.
При использовании здесь термин "Ctx" обозначает холерный токсин, и "CtxB" обозначает В-субъединицу холерного токсина. В других текстах, они могут иногда определяться соответственно как "СТ" или "Ct" и "СТВ" и "CtB". Термин "Etx" здесь обозначает теплолабильный энтеротоксин Е. coli, и "EtxB" - это В-субъединица Etx. В других текстах, они могут иногда называться как "LT" или "Lt", и "LTB" или "LtB" соответственно.
Обоснованием всех сторон настоящего изобретения является тот факт, что EtxB (чистая В-субъединица теплолабильного энтеротоксина Е.coli) связывается с GM-1-ганглизидными рецепторами, которые обнаруживаются на поверхности клеток млекопитающих, и что это связывание вызывает возникновение эффектов в популяции лимфоцитов, включающих специфическое истощение CD8+ Т-клеток и ассоциированную активацию В-клеток. Эти эффекты отсутствуют при применении мутантного EtxB белка, не имеющего GM-1-связывающей активности.
Аутоиммунные заболевания
Аутоиммунитет - это термин, использующийся для описания механизма, посредством которого организм дает иммунный ответ на собственные антигены.
В соответствии с первым аспектом настоящего изобретения, предлагается:
(i) агент, обладающий GM-1-связывающей активностью, отличный от Ctx или Etx, или В-субъединица Ctx или Etx; или
(ii) агент, обладающий воздействием на GM-1 опосредованные внутриклеточные сигнальные события, но не имеющий GM-1-связывающей активности;
для использования в качестве средства для лечения или предупреждения аутоиммунных заболеваний.
Обнаружено, что агент, в соответствии с настоящим изобретением, модулирует популяции лимфоцитов, приводя к индукции апоптоза в CD8+ Т-клетках, повышенной активности CD4+-клеток и поликлональной активации В-клеток. Эти явления вероятно приводят к сдвигу иммунного ответа в сторону индукции Тh2 ассоциированных цитокинов. Считается, что подобные ответы на собственные или перекрестно-реагирующие агенты опосредуют защиту для определения аутоиммунных заболеваний.
В первом воплощении этого первого аспекта настоящего изобретения, агент используется в методе лечения аутоиммунного заболевания в момент его прогрессирования. В этом воплощении агент назначается пациенту с или без одновременного применения собственных или перекрестно-реагирующих антигенов. Применение агента в соответствии с этим воплощением первого аспекта изобретения модулирует природу иммунного ответа на собственный антиген, исключая активацию вызывающего болезнь воспаления, и, следовательно, предохраняет от аутоиммунного заболевания.
Во втором воплощении этого первого аспекта настоящего изобретения агент используется для "вакцинации" представителей млекопитающих против аутоиммунных заболеваний, при которой агент применяется одновременно с собственным или перекрестно-реагирующими антигенными детерминантами (или с комбинацией различных собственных или перекрестно-реагирующих антигенных детерминант), связанными с названным заболеванием. Подобным образом иммунный ответ субъекта на собственный антиген или на перекрестно-реагирующий антиген исключается из активации патогенеза, что предохраняет в будущем от аутоиммунного ответа на собственный антиген.
В этом первом аспекте изобретения, терапевтический агент и собственная или перекрестно-реагирующая антигенная детерминанта должны или могут быть совместно назначены субъекту. Таким образом мы имеем в виду, что место и время введения каждого терапевтического агента и антигенной детерминанты таковы, чтобы достигалась необходимая модуляция иммунной системы. Таким образом, хотя терапевтический агент и антигенная детерминанта могут быть назначены в один и тот же момент времени и в одном и том же месте, могут существовать преимущества в применении терапевтического агента и антигенной детерминанты в разное время и в разном месте.
Хотя единичная доза терапевтического агента и антигенной детерминанты могут быть достаточными, множественные дозы рассматриваются, как входящие в область этого аспекта изобретения.
В этом втором воплощении первого аспекта изобретения, терапевтический агент и антигенная детерминанта могут связываться, например, ковалентной связью, с образованием единого активного агента, хотя раздельное применение, при котором терапевтический агент и антигенная детерминанта не связаны, предпочтительней, так как это делает возможным раздельное применение различных фрагментов.
Специфические аутоиммунные заболевания, которые можно лечить в соответствии с этим аспектом настоящего изобретения, это такие аутоиммунные заболевания, как ревматоидный артрит, рассеянный склероз и диабет, в которых патология связана с клеточно-опосредованным иммунитетом.
Дополнительно, под этим первым аспектом настоящего изобретения предлагается использование Ctx, Etx или В-субъединиц Ctx или Etx в производстве лекарств для использования, в качестве агентов для предупреждения аутоиммунных заболеваний.
Также предполагается фармацевтическая композиция для лечения аутоиммунного заболевания человека, содержащая
(i) агент, обладающий GМ-1-связывающей активность; или
(ii) агент, обладающий воздействием на GM-1 опосредованные внутриклеточные сигнальные события, но не имеющие GM-1-связывающей активности;
и их фармацевтически приемлемый носитель или растворитель.
Фармацевтическая композиция этого аспекта изобретения может быть представлена в таком виде, чтобы его можно было ввести в организм через слизистую оболочку, например с помощью спрея через нос, или парентерально, причем соединение представлено в форме, пригодной для инъекций, например, для применения внутривенно, внутримышечно или подкожно.
Фармацевтическая композиция может быть оформлена в готовую форму вместе с соответствующим собственным или перекрестно-реагирующим антигеном. И наоборот, комплект может содержать отдельные композиции для каждого терапевтического агента и антигенной детерминанты.
Специфические терапевтические агенты, которые могут быть использованы в этом аспекте изобретения, - это EtxB и CtxB или их мутантные разновидности, сохраняющие GM-1-связывающую активность.
Агенты для использования в первом аспекте настоящего изобретения должны предпочтительно быть в основном нетоксичными, хотя некоторый уровень токсичности может быть допустим при тяжелом лечении подобного рода.
Этот первый аспект изобретения распространяется на использование всех агентов, обладающих GM-1-связывающей активностью, для использования в лечении аутоиммунных заболеваний человека, так же как и тех агентов, которые обладают воздействием на GM-1 опосредованные внутриклеточные сигнальные события, и которые, следовательно, действуют подобно GM-1-связывающим агентам.
Таким образом, этот первый аспект настоящего изобретения не ограничивается использованием EtxB белка в качестве терапевтического агента в лечении аутоиммунных заболеваний человека. Однако использование EtxB белка (который является пентамером из пяти идентичных субъединиц) для подобного лечения представляет собой предпочтительное воплощение настоящего изобретения. В дополнение к дикому типу EtxB, этот предпочтительный аспект изобретения также распространяется на мутантные EtxB, которые имеют GM-1-связывающую активность, так же как и на другие эквивалентные белки, такие как В-субъединица холерного токсина (CtxB), и их мутантные формы, которые обладают GM-1-связывающей активностью.
Другими терапевтическими агентами для лечения аутоиммунных заболеваний в соответствии с первым аспектом этого изобретения являются человеческие моноклональные антитела, которые связываются с GM-1. Способы известного уровня техники для определения и получения таких агентов хорошо известны.
Т-лимфоцитарные лейкемии
В соответствии со вторым аспектом данного изобретения предлагается:
(i) агент, имеющий GM-1-связывающую активность, отличный от Ctx и Etx, или В-субъединицы Ctx или Etx; или
(ii) агент, обладающий воздействием на GM-1 опосредованные внутриклеточные сигнальные события, но не обладающий GM-1-связывающей активностью;
для использования в лечении лейкемий человека Т-клеточного происхождения, таких как лейкемии человека CD8 Т-клеточного происхождения.
Агенты для использования во втором аспекте настоящего изобретения должны предпочтительно быть в основном нетоксичными, хотя некоторый уровень токсичности может быть допустим при тяжелом лечении такого рода.
Дополнительно под вторым аспектом настоящего изобретения может предусматриваться использование Ctx или Etx, или В-субъединицы Ctx и Etx для получения медикаментов для лечения лейкемий человека Т-клеточного происхождения, таких как лейкемии человека CD8 Т-клеточного происхождения.
Также предлагается фармацевтическая композиция для лечения лейкемий человека Т-клеточного происхождения, содержащая
(i) агент, обладающий GM-1-связывающей активностью; или
(ii) агент, обладающий воздействием на GM-1 опосредованные внутриклеточные сигнальные события, но не обладающий GM-1-связывающей активностью; и их фармацевтически приемлемые носители и растворители.
Фармацевтическая композиция этого аспекта изобретения может быть представлена в такой форме, чтобы его можно было ввести в организм через слизистую оболочку, например с помощью спрея через нос, или парентерально, причем соединение представлено в форме, пригодной для инъекций, например, для применения внутривенно, внутримышечно или подкожно.
Этот второй аспект изобретения распространяется на все вещества, обладающие GM-1-связывающей активностью, для использования в лечении лейкемий человека Т-клеточного происхождения, так же как те агенты, которые имеют воздействие на GM-1 опосредованные внутриклеточные сигнальные события, и таким образом действуют подобно GM-1-связывающим агентам.
Таким образом, второй аспект настоящего изобретения не ограничивается использованием EtxB белка в качестве терапевтического агента для лечения лейкемий человека Т-клеточного происхождения. Однако использование EtxB белка для такого лечения представляет предпочтительное воплощение настоящего изобретения. В дополнение к дикому типу EtxB, этот предпочтительный аспект изобретения также распространяется на мутантные EtxB, которые имеют GM-1-связывающую активность, так же как и на другие эквивалентные белки, такие как В-субъединица холерного токсина (CtxB), и их мутантные формы, которые обладают GM-1-связывающей активностью.
Другими терапевтическими агентами для лечения аутоиммунных заболеваний в соответствии с первым аспектом этого изобретения являются человеческие моноклональные антитела, которые связываются с GM-1. Способы известного уровня техники для определения и получения таких агентов хорошо известны.
Отторжение трансплантата и GVHD
В соответствии с третьим аспектом данного изобретения предлагается:
(i) агент, обладающий GM-1-связывающей активностью, отличный от Ctx и Etx, или В-субъединиц Ctx или Etx; или
(ii) агент, обладающий воздействием на GM-1 опосредованные внутриклеточные сигнальные события, но не обладающий GM-1-связывающей активностью;
для использования в качестве терапевтического агента для предупреждения/лечения отторжения трансплантата GVHD.
Дополнительно под этим третьим аспектом настоящего изобретения предлагается использование Ctx или Etx или В-субъединиц Etx или Ctx для получения лекарств для предупреждения отторжения трансплантата или GVHD.
В предпочтительном воплощении этого изобретения описанный терапевтический агент может использоваться для предупреждения реакции отторжения трансплантатов твердых органов, как аллогенетических, так и ксеногенетических. Они также могут быть использованы для предупреждения острого заболевания трансплантат против хозяина (GVHD), например при проведении трансплантации костного мозга.
В воплощении этого аспекта изобретения, где пациент получает препарат до трансплантации, терапевтическое вещество должно быть применено одновременно с аллоантигеном и ксеноантигеном. В воплощениях, в которых пациент получает лечение после трансплантации, терапевтический агент применяется без одновременного применения антигена.
В воплощении этого аспекта изобретения, где терапевтическое вещество и алло- и ксено- антигенные детерминанты применяются одновременно, мы имеем в виду, что место и время применения каждого терапевтического вещества и антигенной детерминанты таковы, что достигается необходимая модуляция иммунной системы. Таким образом, хотя терапевтический агент и антигенная детерминанта могут применяться в один и тот же момент и в одно и то же место, есть преимущества в применении терапевтического вещества в другое время и в другом месте, чем антигенная детерминанта. Кроме того, терапевтическое вещество и антигенная детерминанта могут быть ковалентно связаны, образуя единый активный агент, хотя раздельное применение, при котором терапевтический агент и антигенная детерминанта на связаны таким образом, предпочтительно, так как это делает возможным раздельное применение различных фрагментов.
Хотя единичная доза терапевтического агента и антигенной детерминанты могут быть достаточными, множественные дозы рассматриваются, как входящие в область этого аспекта изобретения.
В этом аспекте изобретения, когда агент используется для предупреждения GVHD, он как правило может применяться для воздействия непосредственно на клетки, например клетки костного мозга, предназначенные для трансплантации.
Агент должен быть предпочтительно в основном нетоксичным, хотя некоторый уровень токсичности может быть допустим при тяжелом лечении такого рода.
Также предлагается фармацевтическая композиция для лечения отторжения трансплантата, содержащая
(i) агент, обладающий GM-1-связывающей активностью; или
(ii) агент, обладающий воздействием на GM-1 опосредованные внутриклеточные сигнальные события, но не имеющий GM-1-связывающей активности; и их фармацевтически приемлемый носитель или растворитель.
Фармацевтическая композиция этого аспекта изобретения может быть представлена в таком виде, чтобы его можно было ввести в организм через слизистую оболочку, например с помощью спрея через нос, или парентерально, причем соединение представлено в форме, пригодной для инъекций, например для применения внутривенно, внутримышечно или подкожно.
Фармацевтическая композиция может быть оформлена в готовую форму вместе с соответствующими алло- или ксено- антигенными детерминантами. И наоборот, комплект может содержать отдельные композиции для каждого терапевтического агента и антигенной детерминанты.
Этот третий аспект изобретения распространяется на использование всех агентов, обладающих GM-1-связывающей активностью для использования в предупреждении/лечении отторжения трансплантата или GVHD, так же как и тех агентов, которые обладают воздействием на GM-1 опосредованные внутриклеточные сигнальные события, и которые, следовательно, действуют подобно GM-1-связывающим агентам.
Таким образом, этот третий аспект настоящего изобретения не ограничивается использованием EtxB белка в качестве терапевтического агента в лечении отторжения трансплантата. Однако использование EtxB белка (который является пентамером из пяти идентичных субъединиц) для подобного лечения представляет собой предпочтительное воплощение настоящего изобретения. В дополнение к дикому типу EtxB, этот предпочтительный аспект изобретения также распространяется на мутантные EtxB, которые имеют GM-1-связывающую активность, так же как и на другие эквивалентные белки, такие как В-субъединица холерного токсина (CtxB), и их мутантные формы, которые обладают GM-1-связывающей активностью.
Другими терапевтическими агентами для лечения аутоиммунных заболеваний в соответствии с первым аспектом этого изобретения являются человеческие моноклональные антитела, которые связываются с GM-1. Способы известного уровня техники для определения и получения таких агентов хорошо известны.
Вакцинация
CtxB и EtxB уже были предложены в качестве так называемых "вакцинных носителей". В настоящее время обнаружено, что основой этого эффекта, частично, является способность EtxB модулировать популяции лимфоцитов (как обсуждалось выше) путем связывания GM-1 рецептора.
Таким образом, в соответствии с четвертым аспектом настоящего изобретения, предлагается:
(i) агент, обладающий GM-1-связывающей активностью, отличной от Ctx или Etx, или В-субъединиц Ctx или Etx; или
(ii) агент, обладающий воздействием на GM-1 опосредованные внутриклеточные сигнальные события, но не обладающий GM-1-связывающей активностью; для использования и вакцинации млекопитающего.
Агент способен к модуляции иммунного ответа, когда представлен вместе с неродственной чужеродной антигенной детерминантой. Когда агент вводится парентерально, такая иммуномодуляция с точки зрения иммунного ответа будет "прямой" в особенно ожидаемом направлении. Когда агент вводится через слизистую оболочку вместе с неродственным антигеном, таким как так называемый "слизистый адъювант", агент способен к облегчению слизистого иммунного ответа на неродственный антиген. Антиген и агент могут быть введены вместе как отдельные фрагменты, или могут быть связаны вместе, например, ковалентной связью.
Антиген должен быть предпочтительно нетоксичным. Кроме того, когда агент предназначен для введения через гастроэнтеральную слизистую, он должен быть способен сохранять стабильность во время передвижения по гастроэнтеральному тракту; например, он должен быть устойчив к детергентному влиянию желчи.
Также предлагается фармацевтическая композиция для использования при вакцинации млекопитающих, содержащая
(i) агент, обладающий GM-1-связывающей активностью; или
(ii) агент, обладающий воздействием на GM-1 опосредованные внутриклеточные сигнальные события, но не имеющий GM-1-связывающей активности;
и их фармацевтически приемлемый носитель или растворитель.
Фармацевтическая композиция этого аспекта изобретения может быть представлена в таком виде, чтобы его можно было ввести в организм через слизистую оболочку, например с помощью спрея через нос, или парентерально, причем соединение представлено в форме, пригодной для инъекций, например для применения внутривенно, внутримышечно или подкожно.
Фармацевтическая композиция может быть оформлена в готовую форму вместе с соответствующей антигенной детерминантой. И наоборот, комплект может содержать отдельные композиции для каждого терапевтического агента и антигенной детерминанты.
Этот четвертый аспект изобретения распространяется на использование всех агентов, обладающих GM-1-связывающей активностью, таких как иммуномодуляторы, так же как и тех агентов, которые обладают воздействием на GM-1 опосредованные внутриклеточные сигнальные события, и которые, следовательно, действуют подобно GM-1-связывающим агентам.
Таким образом, этот четвертый аспект настоящего изобретения не ограничивается использованием EtxB белка в качестве иммуномодулятора. Однако использование EtxB белка (который является пентамером из пяти идентичных субъединиц) подобным образом представляет собой одно из воплощений настоящего изобретения. В дополнение к дикому типу EtxB, этот предпочтительный аспект изобретения также распространяется на мутантные EtxB, которые имеют GM-1-связывающую активность, так же как и на другие эквивалентные белки, такие как В-субъединица холерного токсина (CtxB), и их мутантные формы, которые обладают GM-1-связывающей активностью.
Другими терапевтическими агентами для использования в качестве иммуномодуляторов, в соответствии с этим аспектом изобретения, являются человеческие моноклональные антитела, которые связываются с GM-1. Способы известного уровня техники для определения и получения таких агентов хорошо известны.
Когда терапевтический агент настоящего изобретения является белком, таким как EtxB-субъединица или CtxB- субъединица, он может быть получен, для использования во всех аспектах данного изобретения, способом, в котором ген или гены, клонирующие специфическую полипептидную цепь (или цепи), из которых образован белок, встраиваются в подходящий вектор и затем используются для трансфекции подходящего хозяина. Например, ген, кодирующий полипептидную цепь, составляющую EtxB, может быть встроен в, например, плазмид рММВ68, который затем используется для трансфекции клеток хозяина, такого как Vibrio sp. 60. Белок очищается и выделяется способом, известным per se. Мутантные гены, экспрессирующие мутантный EtxB белок, могут затем быть получены известными методами из гена дикого типа.
Как ранее сообщалось, агенты, обладающие GM-1-связывающей способностью, такие как специфически предназначенные человеческие моноклональные антитела, могут быть задуманы и получены, как подчеркивалось выше, известными в технологии способами.
Во всех аспектах изобретения, агент, обладающий GM-1-связывающей активностью, может также быть способен к перекрестному связыванию GM-1-рецепторов. EtxB является одним из таких агентов, которые способны к перекрестному связыванию GM-1-рецепторов благодаря своей пентамерной форме.
Теперь изобретение будет проиллюстрировано ссылками к сопровождающим чертежам и следующими примерами:
фиг. 1 представляет анализы физико-химических свойств EtxB и мутантных форм EtxB (EtxB (G33D));
фиг. 2 иллюстрирует, что рецепторное связывание EtxB необходимо для их сильной иммуногенности in vivo;
фиг. 3 иллюстрирует кинетику пролиферации лимфоцитов, следующую за инъекцией мыши EtxB;
фиг. 4 иллюстрирует, что EtxB вызывает повышение активации В-клеток;
фиг. 5 иллюстрирует, что EtxB вызывает повышение активности CD4+ Т-клеток и истощение CD8+ Т-клеток;
фиг. 6 показывает выборочное истощение OVA-ответственных CD8+ Т-клеток под действием EtxB;
фиг. 7 показывает, что рецепторное связывание EtxB вызывает изменения морфологических характеристик лимфоцитарного ядра клеток, подвергнутых апоптозу;
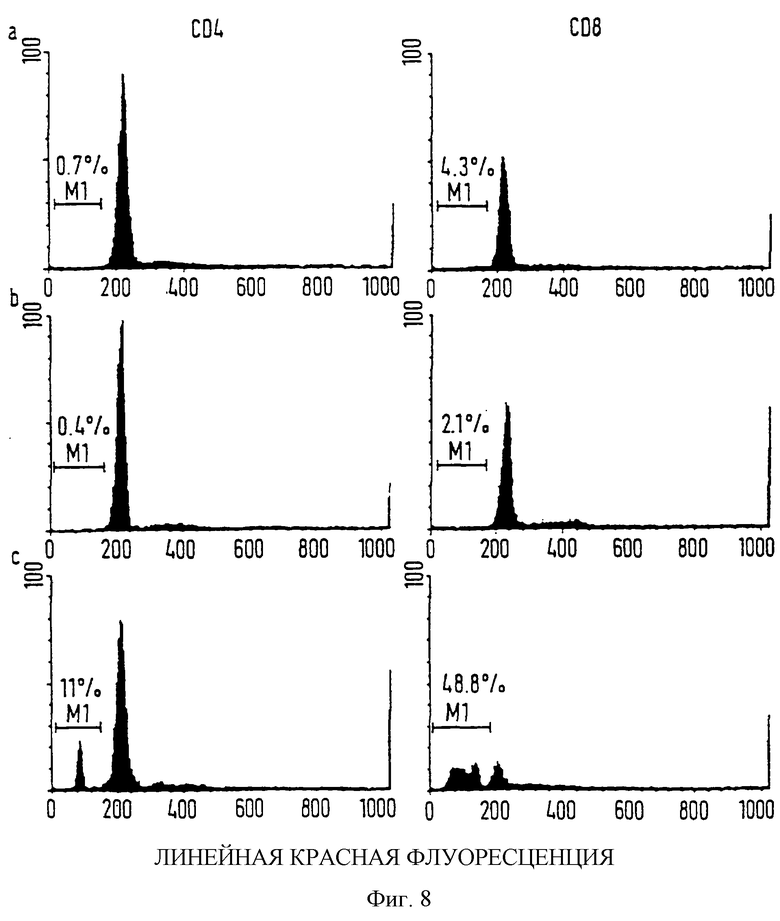
фиг. 8 показывает EtxB рецепторно опосредованный апоптоз CD8+ Т-клеток, как определено при исследовании клеточного цикла;
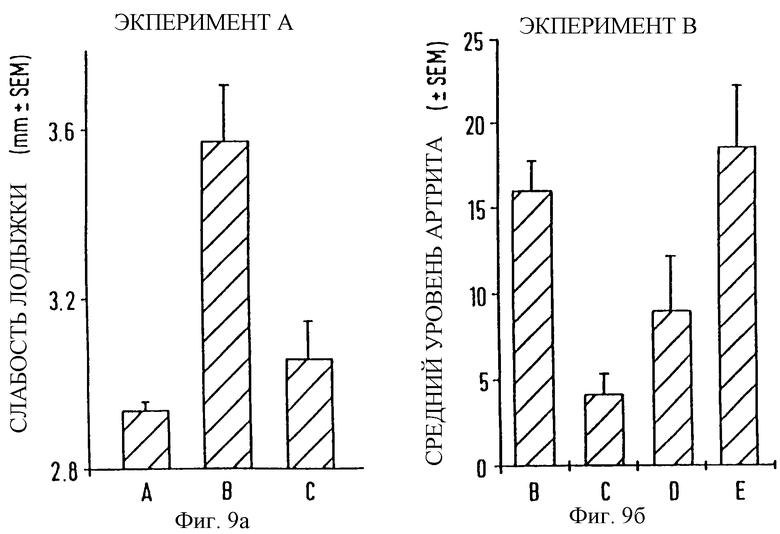
фиг. 9а и 9б показывает результаты экспериментов, проводимых с целью показать, что GM-1-связывание EtxB вызывает развитие коллаген индуцированного артрита на модели животного;
фиг. 10 показывает результаты эксперимента, проводимого для того, чтобы проиллюстрировать, что EtxB, но не EtxB (G33D), вызывает апоптоз в популяции нормальных человеческих мононуклеарных клеток периферической крови;
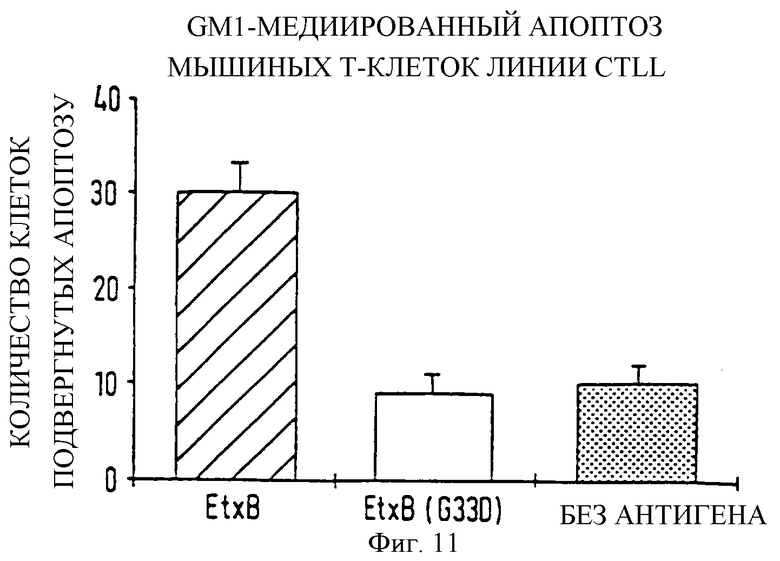
фиг. 11 показывает результаты эксперимента, который показывает, что перекрестное связывание GM1 ведет к апоптозу в части мышиных CTLL клеток.
Детальное описание чертежей
Фиг. 1
Анализы физико-химических свойств EtxB и EtxB (G33D).
(а) SDS-PAGE анализы EtxB или EtxB (G33D): 5 мкг каждого белка анализировались в восстановительных условиях в присутствии β-меркаптоэтанола с или без предварительного нагревания. Линия 1, дикий тип EtxB, ненагретый. Линия 2, EtxB (G33D), ненагретый. Линия 3, дикий тип EtxB, нагретый до 95oС. Линия 4, EtxB (G33D), нагретый до 95oС. Молекулярный вес стандартов (Bio-Rad.) показан слева от панели.
(б) Определение действительной молекулярной массы EtxB и EtxB (G33D) методом гель-фильтрационной хроматографии: стандартная кривая (кружки) получена с использованием, с верху до основания: бычьего сывороточного альбумина (66 кД), куриного яичного альбумина (45кД), карбоангидразы бычьих эритроцитов (29 кД) и цитохрома сердца лошади (12,4 кД); EtxB и EtxB (G33D) элюировали с соответствующими молекулярными массами 36 и 38 кД соответственно; Ve - элюирующий объем белка, Vo - свободный объем гель-фильтрационной колонки.
(в) ELISA для сравнительного связывания EtxB и EtxB (G33D) с ганглиозидом GM-1; платы были покрыты GM-1, блокированы и инкубировались с 1 мкг/мл или EtxB или EtxB (G33D) разведенными серийно (3 кратно) от 1 мкг/мл.
Фиг. 2
Рецепторное связывание EtxB необходимо для их сильной им-муногенности in vivo.
Мышам BALB/c (по 4 в каждой группе) проводились подкожные инъекции или EtxB или EtxB (G33D) в PBS, или белки вводились через рот в бикарбонатном буфере. Сыворотка исследовалась в течение 10 дней, следующих за двумя подкожными инъекциями (А) или в течение 1 недели, следующей за 3 оральными дозами (В), и кишечные выделения исследовались в течение 1 недели, следующей за 3 оральными дозами (С). В образцах, полученных от контрольных мышей, реакция не была определена. Результаты выражены в титрах антител IgG в сыворотке, в то время как IgA в кишечных выделениях выражены в "специфической активности", как описано ниже.
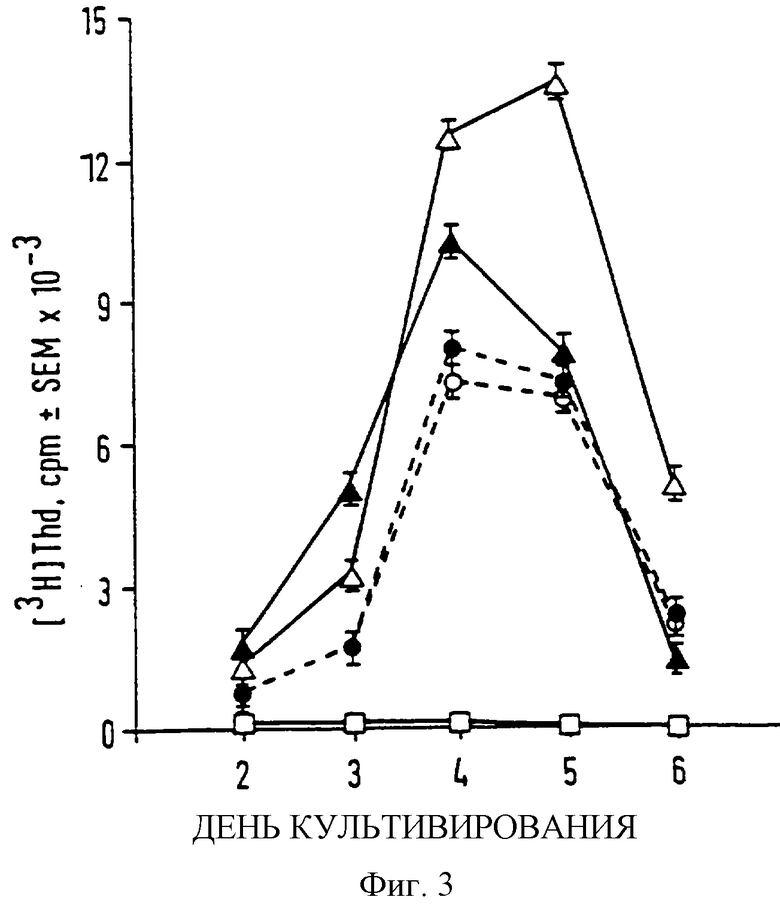
Фиг. 3
Кинетика пролиферации лимфоцитов
Мышам проводились интраперитонеальные инъекции 30 мкг EtxB (G33D) в полном адъюванте Фрейда. MLNs были изолированы 10 днями позже и клетки инкубировали в отсутствие антигена (незакрашенный квадрат) или в присутствии 80 мкг/мл EtxB (закрашенный треугольник), EtxB (G33D) (незакрашенный треугольник) или несобранных мономерных форм EtxB (закрашенный треугольник) и EtxB (G33D) (незакрашенный круг), полученных при нагревании до 95oС. После 6 часов каждого дня исследования клетки импульсно метили с 1 мкКи (3Н) Thd. Данные представляют среднее срm (число импульсов в минуту) и SEM из трех лунок.
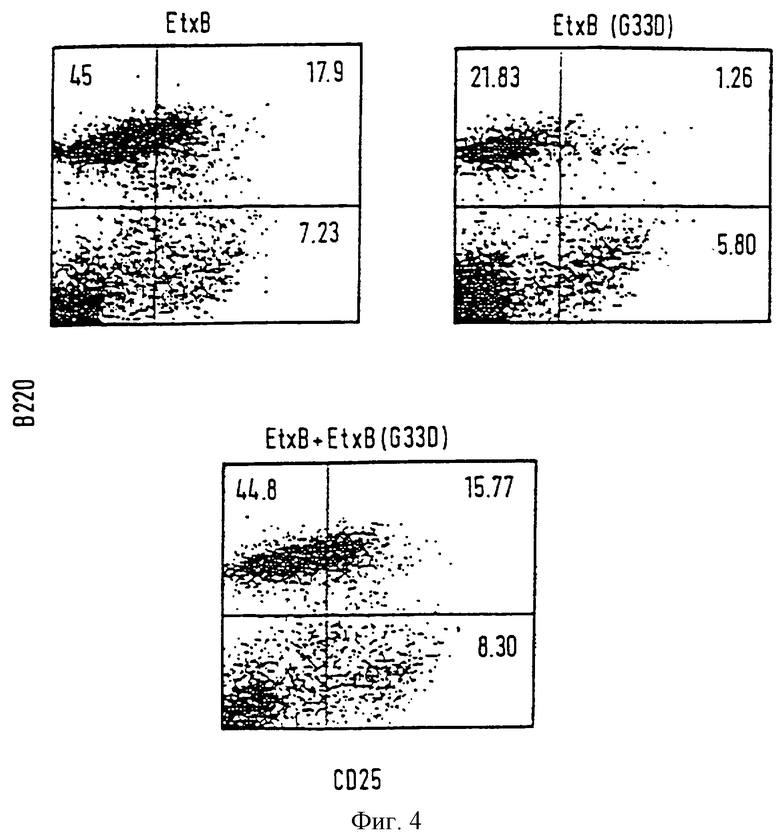
Фиг. 4
EtxB вызывают повышенную активность В-клеток.
Мышей иммунизировали EtxB (G33D) в CFA. Клетки изолировали из MLN 10 дней спустя и инкубировали в присутствии 80 мкг/мл или EtxB или EtxB (G33D), или смеси из 40 мкг/мл каждого белка. Клетки метили биотинилированным анти-СD25 (7D4) и Фикоэритрином (РЕ) анти-В220 (Ra3-6D2). Стрептавидин FITC использовался в качестве вторичного конъюгата антител. Включались также контроли для антител (не показаны). Двойной проточный цитометрический анализ проводился на 4 день пролиферации.
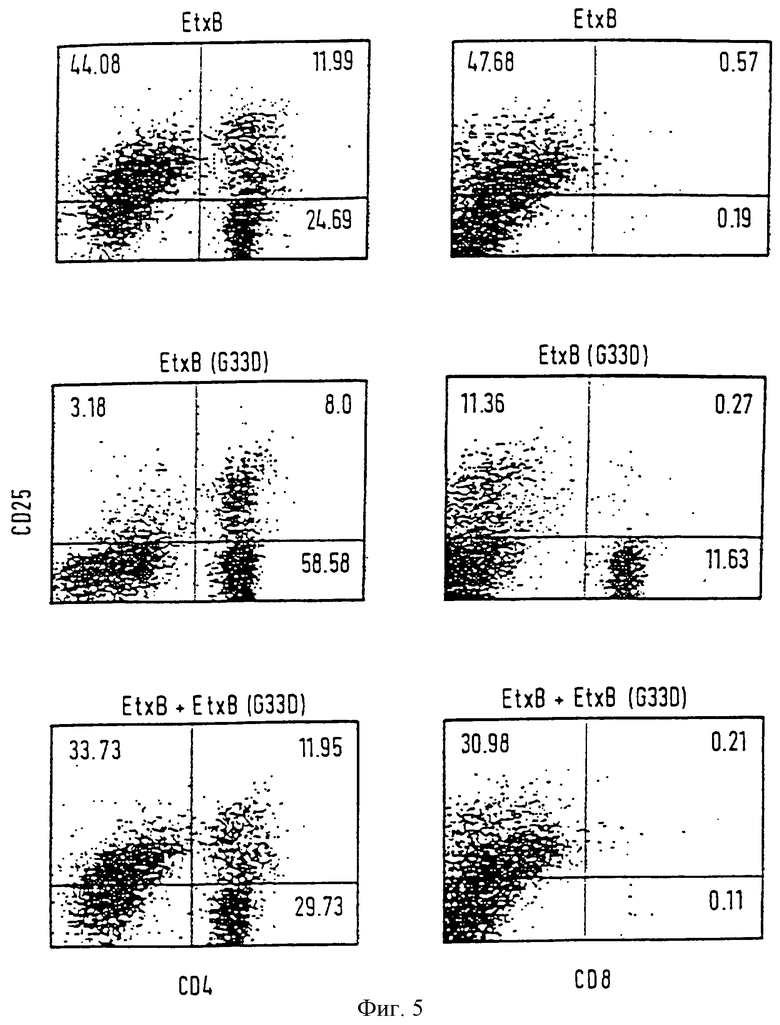
Фиг. 5
EtxB вызывает повышенную активность CD4+ Т-клеток и истощение CD8* Т-клеток.
Процесс иммунизации, выделение клеток и контрольное заражение in vitro такие же, как в описании фиг. 4. Для определения CD25 использовались биотинилированный анти-СD25 (7D4) и Стрептавидин FITC. Для определения CD4 и CD8 использовались FITC, меченный aнти-CD4 (RNRM4-5), и FITC, меченный aнти-CD8α (53-6.7). Соответствующие контроли для антител включались (не показаны).
Фиг. 6
Выборочное истощение ОVА-чувствительных CD8+ Т-клеток с помощью EtxB.
Культуры клеток из MLN, взятых у OVA-праймированных мышей, культивировались в течение 5 дней в отсутствие антигена или в присутствии OVA+EtxB, OVA+EtxB (G33D) или одного OVA при 100 мкг OVA и по 40 мкг/мл EtxB или EtxB (G33D) или 100 мкг только одного OVA. Клетки метили следующими крысиными антителами: FITC-aнти-CD4 или FIТС-анти-СD8α, и они оба с биотин-анти-CD25 (IL-2Rα) с последующим Стрептавидин-фикоэритрином. Непомеченные клетки или клетки, помеченные только вторым антителом, также включались в качестве контроля. Клетки исследовались с помощью FACS (Becton Dickinson). Большее увеличение количества клеток, являющихся CD25+ в культурах, содержащих EtxB по сравнению с другими способами обработки, происходит благодаря присутствию большего количества В-клеток, экспрессирующих этот маркер (не показано). Шкала интенсивности флюоресценции дана в виде логарифма.
Фиг. 7
Рецепторное связывание EtxB вызывает изменение в морфологической характеристике ядра лимфоцитов в клетках, подвергнутых апоптозу.
MLNC, содержащие >90% CD3+ Т-клеток и лишенные макрофагов, инкубировали в течение 18 часов с 80 мкг/мл EtxB или с 80 мкг/мл EtxB (G33D) и метили акридоном оранжевым. Клетки исследовались под обычным или конфокальным флуоресцентным микроскопом (Leica TCS 4D). Показана характерная микроскопическая картина (• 540) для каждого опыта [EtxB, снимок слева; EtxB (G33D), снимок справа] . Клетки, которые инкубировали в присутствии антигена, дают такой же результата, как клетки, обработанные EtxB (G33D) (не показано).
Фиг. 8
EtxB рецепторно-опосредованный апоптоз CD8+ Т-клеток, как определено при анализе клеточного цикла.
Количество CD4+ и CD8+ SPLTC в суб-G0/G1 стадии клеточного цикла определялось проточным цитометрическим анализом содержания DNA, после окрашивания с пропидиум йодидом. SPLTC выделяли из селезенки отрицательным отбором, как описано выше. Клетки обрабатывали в течение 18 часов: (а) без антигена, (b) с 80 мкг/мл EtxB (G33D) или (с) с 80 мкг/мл EtxB и затем метили FITC-крысиными aнти-CD4 или FITC-крысиными анти-CD8α. Клетки последовательно метили пропидиум йодидом. Количество клеток, помеченных пропидиум йодидом, определялось путем установки дискриминационного окна на клетки, меченные или анти-СD4, или анти-CD8 антителами.
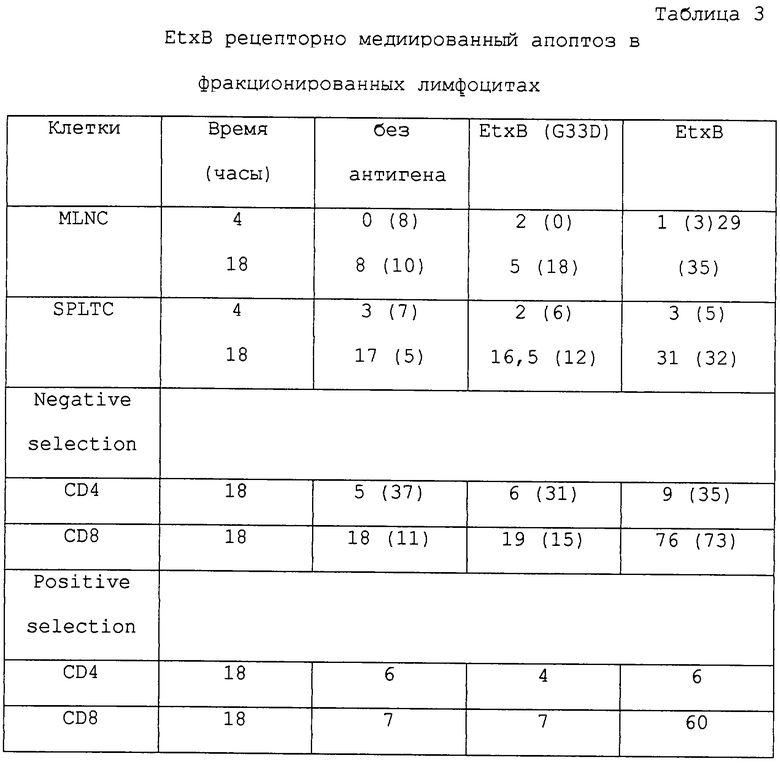
Этот эксперимент проведен на клетках, результаты также представлены на фиг. 7 и в таблице 3.
Примеры
Пример 1
Этот пример иллюстрирует условия GM-1-связывания для достижения различных эффектов на популяции лимфоцитов.
Материалы и методы
Получение рецептор-связанного мутанта EtxB.
Замещение Gly-33 на Asp происходит на участке, ответственном за рецепторное связывание человеческого EtxB, с использованием плазмида pTRH29, производного фагемид вектора pBluescript IIKS+, который содержит гены для А- и В- субъединиц (частиц) Etx (Yu J. , Webb H. & Hirst T.R. (1992). Molec. Microbiol. 6, 1949-1958). Мутагенез осуществляют с оленуклеотид-направленным комплектом для проведения мутагенеза in vitro (Amersham International), используя одноцепочечный pTRH29 в качестве матрицы и синтетический олигонуклеотид (5'-TCTCTTTTATCTGCCATCG-3')(из Microanalytical Facility, IAPGR, Cambridge Research Station, UK) в качестве мутагенного праймера. Правильность замещения Gly на Asp подтверждают дидеокси последовательностью, используя Sequenase II (United States Biochemical Corp.), и полученный в результате плазмид называют pTRH56. Мутантный etxB ген из pTRH56 возбуждается при использовании EcoRI и Spel рестрикционных ферментов, и встраивается в рММВ68 (Sandkvist М. , Hirst T. R. & Bagdasarian, M.,(1987) J. Bacteriol. 169, 4570-4576) для получения широкого радиуса действия экспрессирующего вектора, pTRH64 экспрессирующего EtxB (G33D).
Антигены
Дикий тип EtxB и EtxB (G33D) очищают от культуральных супернатантов Vibrio sp. 60 (рММВ68) и Vibrio sp. 60 (pTRH64) соответственно, используя модификацию метода, предложенного Amin и Hirst (Amin, Т., & Hirst T.R. (1994) Prot. Express, and Purif. 5, 198-204). Кратко, белки очищают дифракцией и гидрофобной связывающей хроматографией и концентрируют анион-обменной хроматографией. Растворы белка обессоливают на колонке PD10 (Pharmacia, UK), уравновешенной фосфатным буферным солевым раствором (PBS; 10 мМ фосфата натрия, 150 мМ NaCl, pH 7,4) и хранят при -30oС.
Чистота EtxB и EtxB (G33D) подтверждается электрофорезом на SDS полиакриламидном геле. Молекулярная масса отдельных мономеров подтверждается лазерной дезорпционной масс-спектрометрией (Protein Science Facility, University of Kent).
Приблизительную молекулярную массу EtxB и EtxB (G33D) определяют гель-фильтрационной хроматографией с использованием SMART системы (Pharmacia). Белки элюируют из колонки Superdex 75 PC 3.2/30 в PBS, pH 7,5.
Необратимая денатурация В-субъединиц пентамера для использования в исследовании лимфотарной пролиферации (см. ниже) достигается нагреванием белков до 95oС в течение 5 минут.
Животные, набор образцов и схемы иммунизации
Мыши BALB/c (H-2d; с высоким ответом на EtxB) 7-12 недельного возраста были приобретены в Charles River Laboratories и содержались в питомнике Университета Кента. Ответы антител на EtxB или EtxB (G33D) измеряли после подкожной инъекции мышам 30 мкг белка в PBS, с последующей поддерживающей инъекцией через 10 дней. Другой группе мышей тот же белок вводили через рот в бикарбонате натрия (50 мкг/мл) за три приема с интервалом в одну неделю. Контрольным мышам давали PBS. Забор крови производился в течение 10 дней, следующих за последней подкожной инъекцией, или в течение 1 недели, следующей за последним оральным применением. Кишечные выделения живых мышей выделяли в раствор ингибитора протеаз, как ранее описывалось (Elson С.О., Еаlding, W. & Lefkowitz, J. (1984) J. Immunol. Meth. 67, 101-108), в течение одной недели, следующей за последним кормлением. Образцы затем разрушали ультразвуком и осветляли цетрифугацией (13,226 • g, 10 минут, при 4oС).
Для исследования пролиферации мышам проводили интраперитонеальную инъекцию с 30 мкг EtxB или EtxB (G33D) в полном адъюванте Фрейда (CFA), и мезентериальные лимфоузлы выделялись 10 днями позже. В исследование также включались контрольные иммунизированные мыши и их лимфоузлы, выделенные подобным же образом.
Энзим-связанный иммуносорбентный анализ (ELISA's) (Твердо-фазный иммуносорбентный метод)
Связывание EtxB или EtxB (G33D) с GM-1 исследовались методом GM-1-ELISA (Amin, Т., & Hirst. T.R. (1994) Prot. Express. and Purif. 5, 198-204).
Сыворотку и кишечные выделения исследуют в присутствии анти-В-субъединиц IgG и IgA антител методом ELISA's, в котором образцы помещают в микротитровальные платы (Immulon I, Dyna-teck, USA), покрытые 5 мкг/мл или EtxB или EtxB (G33D) в PBS. Анти-В-субъединицы антител IgA в супернатантах кишечных выделений экстраполируют из стандартной кривой, полученной при покрытии двух рядов лунок на каждой плате 1 мкг/мл кроличьего антимышиного IgA (α цепь специфичная; Zymed, Lab, USA) в PBS, с последующим добавлением 1мкг/мл IgA мышиной миеломы (МОРС 315, Sigma, USA). Для измерения общего количества IgA лунки покрывают кроличьим анти-мышиным IgA с последующим добавлением супернатантов кишечных выделений. Все образцы серийно разводят. Пероксидазный конъюгат козьего анти-мышиного IgG (Fc специфический фрагмент; Jackson Lab., USA) или козьего анти-мышиного IgA (специфическая цепь; Sigma) разводят и добавляют во все лунки. Определяют титр анти-В-субъединиц IgG, дающий A450nm≥0,2. Ответ анти-В-субъединиц IgA для EtxB и EtxB (G33D) в кишечных выделениях высчитывают как "IgA специфическую активность" [среднее отношение IgA анти-В-субъединиц (мкг/мл)/общее количество IgA (мкг/мл)].
Метод ELISA для измерения уровня цитокинов IL-2, IL-4, IL-5, IL-10 и IFN-γ применялся, как ранее описано (Harper H.M., PhD thesis, University of Bristol (1995)). Коротко, микротитровальные платы покрывают крысиными антителами к мышиным IL-2, IL-4, IL-5, IL-10 и IFN-γ. Платы блокируют с 2% (вес/объем) бычьего сывороточного альбумина. Супернатант из культуральной среды добавляют в лунки и разводят. Один ряд на каждой пластинке для каждого цитокина содержит стандартное количество рекомбинантных цитокинов. Затем пластинки инкубируют с 0,5 мкг/мл биотинилированных антицитокин моноклональных антител, с последующим добавлением авидин-перксидазы и 3,3', 5,5'-Тетраметил-бензиденового (ТМВ) субстрата и читают при A450nm.
Исследование пролиферации лимфоцитов
Мышей убивают шейным вывихом, мезентеральные лимфоузлы асептически выделяют и измельчают в сите из нержавеющей стали в сбалансированном солевом растворе Хенкса (HBSS). (Flow Laboratories Irvine, Renfrewshire, UK.). Клетки промывают центрифугацией (500•g, 10 минут, 4oС) в HBSS и ресуспендируют в модифицированной Eagle's среде (Flow), к которой добавлено 20 мМ Hepes (Flow), 100 IU Пенициллина, 100 мкг/мл Стрептомицина, 4 мМ L-глютамина (Flow) и 2-меркаптоэтанол (полная среда). Свежую аутологичную нормальную мышиную сыворотку иммунизированных мышей добавляют до конечной концентрации 0,5% (объемных). Культуры содержат 2•106 жизнеспособных клеток/мл в каждых 2 мл объема в платах с 24 лунками или в 8 мл объема в 25 см3 флаконах (Nunc A/S, Roskide, Denmark) и культивируют в присутствии или в отсутствие антигенов, как показано в описании чертежей. Культуры инкубируют при 37oС во влажной атмосфере из 5% CO2 и 95% воздуха в течение шести дней. В ожидаемый момент времени 0,1 мл образцы извлекают из среды и переносят на платы с 96 лунками с U-образным дном (Nunc) и импульсно метят 1 мкКи/лунку [3H]-Thd (Amersham, UK) за 6 часов пред тем, как собрать (Mash III harvesting 96 Tomtec, Orange, Conn. USA) и посчитать стандартной жидкой сцинцилляцией 1450 Micro β plus, LKB-Wallas, Turku, Finland). Подобным образом 0,5 мл супернатанта выделяют из культуры для анализа цитокинов. Клетки осаждают и супернатанты до исследования хранят при 68oС.
Фенотипический анализ культивируемых клеток
Культивируемые клетки, собранные на 4 день культивирования, промывают, и жизнеспособные клетки извлекают на поверхность раздела градиента HBSS/18% метризамида (Nyegaard и Со.,Oslo, Norway) с последующей центрифугацией при 500•g в течение 15 минут при 20oС. Клетки дважды промывают и ресуспендируют в HBSS, содержащем 0,2% азида натрия (Sigma) и 10% нормальной крысиной сыворотки. Использовались следующие крысиные антитела (Pharmingen, San Diego, USA): флуоресцентный изотиоцианат (FINC), меченный анти-СD4 (RNRMA-5), FITC, меченный анти-СD8 (53-6.7), биотин, меченный анти-СD25 (7D4) и фикоэритрин (РЕ), меченный анти-В220 (RA3-6D2). Дополнительно для биотинмеченых антител использовался Стрептавидин-РЕ или Стрептавидин FITC (Serotech, UK). Все антитела разводят в HBSS, содержащей азид, и используют в предварительно определенных концентрациях. 200 мкл 2•106 клеток и 200 мкл каждого из антител смешивают и инкубируют на льду в течение 30 минут. Когда необходимы Стрептавидин-РЕ или FITC вторичные антитела, клетки инкубируют с этими антителами еще дополнительно 30 минут. Соответствующие контроли для FITC и РЕ антител также включают в исследование. Клетки промывают HBSS и затем анализируют 2 проточной цитометрией (Becton Dickinson).
Результаты
Получение и характеристика рецепторно связанного мутанта EtxB.
Замещение Gly на Asp встраивались в В-субъединицу тепло-лабильного энтеротоксина Е. coli путем олигонуклеотидно-направленного мутагенеза EtxB с целью производства мутантной формы В-субъединицы, имеющую дефект в рецепторном распознавании. Мутантный белок, названный EtxB (G33D), и дикий тип EtxB очищали до гомогенного состояния (смотри материалы и Методы). Молекулярную массу очищенного EtxB и EtxB (G33D) определяли лазерной дезорпционной масс-спектрометрией. Массы входили в 20 Д теоретических масс в 11702 и 11760 Д для мономеров EtxB и EtxB (G33D) соответственно. При анализе с SDS-PAGE без предварительного нагревания как EtxB, так и EtxB (G33D) мигрировали как дискретные стабильные олигомеры с соответствующим молекулярным весом в 42 кД и 56 кД (фиг.1а, линия 1 и линия 2 соответственно). Наблюдающаяся электрофоретическая подвижность и SDS-стабильность EtxB является характерным свойством В-субъединицы пентамера ( см. Sandkvist М., Hirst T.R. & Bagdasarian, M. , (1987) J. Bacteriol. 169, 4570-4576). Медленная электрофоретическая подвижность олигомерного EtxB (G33D) не является следствием различия в количестве составляющих В-субъединиц мономеров, так как оба пентамерных EtxB и EtxB (G33D) проявляют одинаковое время задержки при гель-фильтрационной хроматографии с высокой разрешающей способностью. Таким образом, отличие в электрофоретической подвижности EtxB (G33D) олигомера от дикого типа EtxB возможно существует благодаря встроенному отрицательно заряженному Asp остатку, вызывающему снижение в SDS-связывании и последующее замедление миграции.
EtxB и EtxB (G33D) также сравнивались по их стабильности в буферах с низким рН, устойчивости к 1,0 мг/мл трипсина или протеиназы К, и по относительной реактивности с рядом анти-В-субъединиц моноклональных и поликлональных антител. В каждом из
этих опытов EtxB (G33D) проявлял такие же свойства, как и дикий тип EtxB. Из этого делается вывод, что замещение Gly на Asp в остатке 33 в EtxB не меняет олигомерную конфигурацию, SDS, рН и протеазную стабильность или реактивность антител по сравнению с диким типом EtxB.
Способность EtxB (G33D) связываться со своим рецептором GM-1, оценивалась с использованием метода GM-1-ELISA (фиг.1в). Он показывает очень значительное снижение способности мутанта к связыванию GM-1 по сравнению с диким типом белка (>99% снижения при чтении А450nm). Более того, в отличие от дикого типа EtxB, EtxB (G33D) имеет недостаточную способность в связывании с СНО-клетками при проверке методом иммунофлуоресценции. Делается вывод, что EtxB (G33D) имеет дефект способности к связыванию GM-1 ганглиозида in vitro и in situ.
Сильная иммуногенность EtxB in vivo зависит от рецепторного связывания.
Важность рецепторного связывания для иммуногенности EtxB оценивалась у мышей после орального введения, или подкожной инъекции EtxB или EtxB (G33D) в PBS. В результате орального введения EtxB определяется высокий титр IgG антител в сыворотке и IgA антител в кишечных выделениях (фиг.2). И наоборот, при таком же режиме оральной иммунизации EtxB (G33D) не обнаружено никакой определимой активности антител. EtxB вызывает ответ сывороточных антител после введения путем подкожной инъекции, хотя этот ответ значительно ниже по сравнению с ответом антител на дикий тип EtxB, с 160-кратным снижением титра антител, 1050 против 171000 соответственно. Делается вывод, что рецепторное связывание EtxB необходимо для их сильной иммуногенности in vivo.
Рецепторное связывание не влияет на пролиферацию лимфоцитов в присутствии EtxB или EtxB (G33D).
Проверялось влияние EtxB и EtxB (G33D) на пролиферацию лимфоцитов in vitro. Лимфоциты выделяли из подколенных и мезентериальных лимфоузлов (MLN) мышей, иммунизированных или EtxB, или EtxB (G33D), и стимулировали in vitro одним или другим белком или препаратом денатурированного нагреванием EtxB или EtxB (G33D). Пролиферативный ответ лимфоцитов, полученных из подколенных или MLN лимфоузлов, был одинаковым. В каждом случае пролиферация в ответ на каждый из белковых препаратов повышалась с повышением концентрации В-субъединиц. Представительное число данных эксперимента с использованием MLN показано в таблице 1. Величина ответа на дикий и мутантный тип пентамеров сравнивалась с ответом, полученным в присутствии денатурированного нагреванием дикого и мутантного типов мономеров. Фиг. 3 показывает кинетику пролиферативного ответа, полученного в присутствии 80 мкг/мл каждого из белковых препаратов. Реактивность зависела от присутствия антигена и носила одинаковый характер в присутствии любого белка. Реактивность становилась очевидной на 3 день культивирования с инкорпорированным [3H]-Thd, достигая пика на 4 день и колеблясь после этого. Минимальные различия во времени наступления пика ответов, очевидные на фиг. 3, не наблюдались при повторных проверках, показывая что анамнестические характеристики ответов на EtxB и EtxB (G33D) являются сравнимыми. Из этого делается вывод, что уровень стимуляции в присутствии нативного белка скорее всего не зависел от рецепторного связывания или встроенной мутации.
Рецепторное связывание токсина вызывает иммуномодуляцию В-клеточной и Т-клеточной популяций.
Чтобы проверить, оказывает ли рецепторное связывание какое-либо влияние на популяцию лимфоидных клеток in vitro, лимфоциты выделялись у MLN мышей, праймированных интраперитониально EtxB (G33D), и затем стимулировались или EtxB, или EtxB (G33D), или их смесью. Кроме того, параллельно проводился эксперимент с использованием лимфоцитов MLN-происхождения от мышей, инъецированных EtxB, причем были получены результаты, идентичные полученным при опыте с EtxB (G33D) праймированными мышами.
(i) EtxB вызывает повышение активации В-клеток.
Воздействие EtxB и В-клетки проверялись путем экспрессии активирующего маркера CD25 (IL-2Rα) в ассоциации с В-клеточным маркером В220 (CD4 5R). Как показано на фиг.4, количество В-клеток в культуре, стимулированной EtxB, составляло 62,9% от общего количества клеток, из которых большое количество (28,4%) экспрессировало маркер клеточной активации CD25. В противоположность этому, количество В-клеток после стимуляции в присутствии EtxB (G33D) было вполовину меньше по сравнению с диким типом (22,26%) и меньшее количество было активировано (5,6%). Для подтверждения, является ли воздействие, оказанное EtxB, преобладающим, клетки инкубировали в присутствии эквимолярных концентраций EtxB и EtxB (G33D). Данные проточной цитометрии были одинаковыми с данными, полученными при стимуляции в присутствии только одного дикого типа EtxB (60,6% В-клеток, из которых 26% было активировано). Делается вывод, что способность EtxB к рецепторному связыванию опосредует повышение активации В-клеток in vitro.
(ii) EtxB вызывает повышенную активацию CD4+ Т-клеток и полное истощение CD8+ Т-клеток.
Для проверки влияния рецепторного связывания В-субъединиц на Т-клетки, лимфоциты метили антителами к CD4 или CD8 в ассоциации с антителами CD25 (фиг. 5). Кроме того, некоторые клетки были избирательно помечены антителами к CD3 маркеру (не показано). Количество Т-клеток, экспрессирующих CD4 маркер, при стимуляции в присутствии EtxB составляло 36,7%, большое число которых (32,7%) было активировано. В противоположность этому, в культуре, содержащей EtxB, не присутствовало определимого количества CD8+ Т-клеток.
Для сравнения, как CD4+, так и CD8+ Т-клетки присутствовали в культуре, стимулированной в присутствии EtxB (G33D). Такие культуры содержали большее количество CD4+ Т-клеток (66,6%), но только 12% из них было активировано. Количество CD8+ Т-клеток, определенных в присутствии EtxB (G33D), составляло 11,7% от общего числа клеток, но очень немногие из них были активированы, что определено по отсутствию CD25 маркера. Кроме того, в присутствии смеси, состоящей из эквимолярных концентраций EtxB и EtxB (G33D), характер клеточного ответа был таким же, как и в присутствии одного дикого типа EtxB; с 41,68% CD4+ Т-клеток (из которых 28,6% были CD25+) и без определимых CD8+ Т-клеток (фиг. 5). Во всех этих исследованиях количество клеток, меченных CD3, было приблизительно равно сумме клеток, экспрессирующих CD4 и CD8 маркеры. Эти данные показывают, что повышение активации В и CD4+ Т-клеток и избирательное истощение CD8+ Т-клеток опосредовано рецепторным связыванием токсина.
Продукция цитокинов
Чтобы оценить, может ли воздействие EtxB на популяции лимфоцитов зависеть от изменения в продукции цитокинов, культуру клеток инкубировали или с EtxB, или с EtxB (G33D) и супернатанты удаляли на 2, 3, 4, 5 и 6 дни исследования. Результаты, полученные от образцов, собранных на 5 день, когда определялась максимальная концентрация цитокинов, показаны в таблице 2. Как IFN-γ, так и IL-2 определялись в супернатантах культур, стимулированных в присутствии EtxB или EtxB (G33D), хотя относительные уровни этих цитокинов варьировались. Среду от клеток, инкубированных с диким EtxB, содержащую 3-кратную повышенную концентрацию IL-2 и 1,5-кратно сниженный уровень IFN-γ, сравнивали с супернатантами из культур, стимулированных в присутствии EtxB (G33D). Несмотря на то, что было обнаружено, что другие культуры пролиферирующих Т-клеток, отвечающих на другие антигены, вырабатывают высокий уровень IL-4, IL-5 и IL-10, ни один из этих цитокинов не был определен в культуре, стимулированной с EtxB или EtxB (G33D). Повышенные уровни IL-2 и понижение уровня IFN-γ вслед за стимуляцией EtxB, по сравнению с EtxB (G33D), наиболее вероятно отражает состояние активации В и CD4+ Т-клеток. Тем не менее, результаты показывают, что абсолютное воздействие дикого типа EtxB на CD8+ Т-клетки вряд ли опосредовано большим сдвигом в пролиферации цитокинов, вследствие блокады рецепторов.
Обсуждение
Это исследование показывает, что введение единичной точечной мутации (G33D) в область рецепторного связывания EtxB вызывает значительную потерю способности к связыванию с GM-1. Важно отметить, что мутантный EtxB (G33D) проявляет физико-химические свойства, идентичные свойствам дикого типа в том, что касается структуры, что обнаруживается при гель- хроматографии, стабильности в SDS, кислотах и протеазах. При определении специфического антительного ответа после иммунизации EtxB или EtxB (G33D), были отмечены серьезные различия. В результате подкожного инъецирования мышам EtxB (G33D) титр антител значительно падал по сравнению с диким типом (са >160 раз), в то время как после орального применения не определялось никакого антительного ответа. Возможно, что эти различия явились результатом разрушения доминантного эпитопа, включенного или в распознавание молекулы антителом, или в стимуляцию эффективной Т-клеточной помощи для продукции антител. Однако заслуживает внимания, что замещение Gly на Asp не влияет на распознавание В-субъединицы рядом специфических поликлональных и моноклональных антител. В дальнейшем, пролиферативные ответы, полученные при добавлении к культурам EtxB или EtxB (G33D), были сравнимы вне зависимости от того, какой из белков использовался для праймирования in vivo; это показывает, что Т-клеточная реактивность не специфична для этих двух молекул. Из этого делается вывод, что рецепторное связывание EtxB необходимо для проявления сильной иммуногенности in vivo.
Важность рецепторного связывания для сильной иммуногенности EtxB может объясняться различным образом. Прежде всего, связывание В-субъединицы с Etx и Ctx с GM-1 может повышать эффективность поглощения этих белков, повышая локальную концентрацию белка, подходящего для иммунной системы. Обнаружено, что другие классы белков, которые способны связываться со слизистыми оболочками, являются эффективными иммуногенами (De Aizpura H.J. & Russell-Jones G. J. (1988) J. Exp. Med. 167, 440-451). Наблюдаемые различия в иммуногенности EtxB и его мутанта после орального применения могут, конечно, существовать благодаря эффективному всасыванию EtxB из просвета кишечника. Однако серьезные отличия, отмеченные после парентеральной иммунизации (когда антиген поступал локально с высокой концентрацией), заставляют думать о других влияниях. Например, связывание EtxB с GM-1 может воздействовать на эффективность активности антиген-представляющих клеток. Такое связывание может вызывать активацию II-несущего класса клеток, особенно это касается экспрессии необходимых ко-стимулированных молекул, таких как В7, которые связаны со своей приобретенно усиленной антиген-представляющей активностью (Jenkins М.К. & Johnson J. G. (1993) Curr. Opin. Immunol. 5, 361-367). С другой стороны, рецепторное связывание может непосредственно воздействовать на субпопуляции лимфоцитов. Количество наблюдений в этом исследовании обеспечивает большую степень достоверности данного наблюдения.
Исследования in vitro показывают, что EtxB способен вызывать пролиферацию праймированных клеток из лимфатических узлов. Это свойство на зависит от рецепторного связывания, так как ответы с одинаковыми анамнестическими характеристиками были получены при использовании как дикого типа EtxB и EtxB (G33D), так и денатурированных нагреванием мономерных форм этих белков, которые не связываются с GM-1. Эти наблюдения интересны сами по себе, так как широко сообщалось, что коммерческие препараты Ctx и CtxB или очищенного рекомбинантного CtxB являются сильными ингибиторами лимфоцитарной пролиферации in vitro. Видимое противоречие может явиться результатом того факта, что предыдущие эксперименты были проведены с очищенными лимфоцитами, и что широко использовались лимфоцитарные культуры, стимулированные мутогеном (которые не являются клональными рестрикционными ответами), когда могли быть включены другие различные механизмы. С этим согласуется наше наблюдение, что пролиферация Con A-стимулированных лимфоцитов без сомнения ингибируется EtxB. Однако анализ клеточных популяций в культурах праймерованных клеток из лимфоузлов, стимулированных или EtxB, или EtxB (G33D), выявил важные различия, касающиеся как В-клеток, так и CD4 и CD8 -несущих Т-клеток.
В-клетки определялись через 4 дня после культивирования в присутствии или EtxB, или EtxB (G33D). Однако, по сравнению с EtxB (G33D), относительное количество В-клеток, присутствующих в среде с EtxB, было выше приблизительно на 100%. Это повышение сочеталось с экспрессией CD25 на очень большом количестве В-клеток. В показанном эксперименте отвечающие лимфоциты были праймированы EtxB (G33D) in vivo. Подобные эксперименты с клетками мышей, иммунизированных EtxB, показали сравнимые результаты. Таким образом, независимо от любых эффектов in vivo, связанных с рецепторным связыванием, среды в присутствии EtxB содержали большее количество В-клеток по сравнению со средами, стимулированными с EtxB (G33D). Подобные эффекты на В-клетках также проявлялись независимо, или частично завися, от регуляции Т-клетками, in vitro, так как результат не предполагает большого сдвига в профиле обнаруженных цитокинов. Таким образом, in vitro рецепторное связывание EtxB оказывается связанным с прямым воздействием на В-клетки, давая в результате как пропорциональное распространение этой популяции, так и ее активацию. Также заслуживает внимания, что CtxB показал повышение экспрессии МНС класса II на чистые В-клетки, свойство, которое не проявлялось GM-1- связывающим мутантом CtxB (G33D) (Francis M.L., Ryan J., Jobling M.G., Holmes R.K., Moss J. & Mond J. J. (1992) J. Immunol. 148, 1999-2005). Результат этих экспериментов предполагает наличие прямого мутантного эффект EtxB на антиген-праймированные В-клетки и показывает, что подобный эффект опосредован рецепторным связыванием.
Кроме воздействия EtxB на В-клетки в культуре, проточный цитометирческий анализ выявляет, что этот токсоид вызывает полное истощение любых определимых CD8+ клеток. И снова показано, что этот эффект зависит от рецепторного связывания, так как эта же популяция Т-клеток не истощалась в среде, содержащей EtxB (G33D). Далее, наблюдалось полное истощение в культуре, содержащей EtxB, CD8+ клеток от мышей, иммунизированных диким типом EtxB. Существует три возможных механизма, которые могут опосредовать подобный эффект. 1) Известно, что связывание Ctx или CtxB с GM-1 на крысиных MLN- клетках вызывает образование бляшек и шапочки (Craig S.W. and Cuatrecasas P., (1975) Proc. Natl. Acad. Sci. USA, v. 72, p. 3844-3848). Возможно, что в этом процессе EtxB-GM-1 комплекс и другие молекулы, включая CD8, интернализированы. Подобный процесс может препятствовать определению этих клеток при проточной цитометрии при использовании CD8 в качестве маркера и может привести к их гибели благодаря ассоциативной потере пространственного TCR комплекса. Хотя последнее можно рассматривать как отсутствие CD8+ Т-клеток в культуре, другие не обнаруживают отсутствия TCR комплекса на поверхности Т-клеток человека линии Jarkat при использовании CtxB (Imboden J.B., Shoback D.M., Pattison G. & Stobo J.D., (1986) Proc. Natl. Acad. Sci. USA 83, 5673-5677). Отсутствие эффекта в результате кэппинга поддерживается тем, что обнаружено отсутствие влияния маркеров CD3 и CD4. 2) Альтернативный механизм включает эффект, осуществляемый цитокинами в культуре.В этом исследовании определялись как IL-2, так и IFN-γ. Результат, однако, не предполагает настолько большой сдвиг в профиле цитокина, который бы объяснил такое сильное воздействие на CD8+ Т-клетки. 3) Отсутствие CD8+ Т-клеток может быть следствием активной индукции апоптоза. Гибель лимфоцитов в результате апоптоза может включать кэппинг-феномен, как описывалось выше, или при отсутствии кэппинга может быть опосредовано воздействием на сигнальные события в клетке. Индуцированная активацией запрограммированная гибель зависит от Са2+ и включает фосфатазы и киназы. Показано, что связывание CtxB с лимфоцитами ингибирует протеинкиназную С-зависимую пролиферацию и индуцирует явное повышение внутриклеточного Са2+ события, которые не связаны с повышением уровня CAMP. Способность EtxB истощать CD8+, но не CD4+ Т-клетки может быть следствием воздействия различных сигналов, связанных с CD4/CD8-TCR комплексом, являясь результатом перекрестного связывания с GM-1 на поверхности клеток этих популяций лимфоцитов. Это может происходить, как результат различного связывания токсоида на мембране, как сообщалось для CtxB или наоборот, различных сигнальных механизмов в CD4+ и CD8+ Т-клетках.
При полном отсутствии определимых CD8+ Т-клеток, EtxB повышал количество CD4+ Т-клеток, которые были активированы, по сравнению с рецепторно связанным мутантом. Условия, необходимые для ответа CD4+ Т-клеток на Ctx продемонстрированы in vivo. Причины повышения активации этой Т-клеточной популяции, однако, остаются неясными. Заслуживает внимания, что показано, что CtxB стимулирует DNA синтез и клеточное деление в находящихся в покое нетрансформированных мышиных 3Т3 клетках. Избирательное мутагенное воздействие на CD4+ Т-клетки также было обнаружено в присутствии растительных лектинов, которые связываются с Galβ-1-3-3GalNAc, тем же компонентом, с которым EtxB связывается с GM-1. Нельзя исключить возможность, что EtxB опосредует зависимое от GM-1-связывания прямое воздействие на CD+ Т-клетки, вызывая их активацию. Однако, также возможно, что повышение активации CD4+ Т-клеток в культурах, содержащих EtxB, является результатом описанных изменений в В-клеточных и CD8+ Т-клеточных популяциях. Известно, что активация В-клеток связана с повышением их компетентности, как клеток, представляющих антиген для CD4+ Т-клеток. Кроме того, CD8+ Т-клетки широко связаны с регуляторной ролью в иммунной реактивности как in vivo, так и in vitro. Их удаление из Т-клеточных пролиферативных культур связано с пролонгированными и повышенными уровнями деления CD4+ Т-клеток.
Обобщив, можно предположить, что сильная иммуногенность EtxB in vivo, как показало исследование, оказывается результатом их способности повышать активацию В-клеток под влиянием эффектов регуляции роста после связывания с GM-1. Активность CD4+ Т-клеток и способность EtxB увеличивать IL-2 в культуре in vitro могут служить необходимым сигналом для дальнейшего распространения клонов В-клеток. Истощение CD8+ Т-клеток EtxB in vitro в этом исследовании может также предполагать другой механизм иммунопотенцирование in vivo после системного или орального введения, особенно в свете включения этой популяции клеток в суппрессию иммунного ответа и в оральную толерантность. С этой точки зрения показано, что как Ctx, так и Etx отменяют оральную толерантность к одновременно орально введенным растворимым белкам, другие исследования для объяснения этого механизма включают эффект Ctx и CtxB истощения интраэпителиальных лимфоцитов в кишечнике или в своде Пейровой бляшки. Также показано, что CtxB-ингибирующее воздействие на CD8+ Т-клетки in vitro предупреждает реакцию трансплантат против хозяина.
В заключение продемонстрировано наличие сильного иммуномодуляторного воздействия EtxB на антительный ответ in vivo, и на популяции лимфоцитов in vitro. Кроме того, показано, что это воздействие опосредовано рецепторным связыванием. Наши выводы также относятся к пониманию способности Etx и Ctx действовать подобно сильному адъюванту и потенциальному белковому носителю для других антигенов, и предполагается, что такие свойства основаны на способности этих токсоидов связывать ганглиозидные рецепторы на поверхности лимфоидных клеток.
Пример 2
Этот пример иллюстрирует, что воздействие на CD8 клетки не зависит от антигенного распознавания и медиируется апоптозом.
Рекомбинантные препараты EtxB и EtxB (G33D) получают, как в Примере 1. Оба белка хорошо охарактеризованы в отношении связывания GM-1, связывания с рядом моноклональных и поликлональных антител и различных других физико-химических свойств. Яичный альбумин (OVA) приобретен у Sigma (Poole, UK). Мезентеральные лимфоузлы (MNL) выделяют у мышей BALB/c [линия с высоким ответом к EtxB (Nashar Т.О. and Hirst T.R. 1995. Immunoregulatory role of H-2 and intra-H-2 alleles on antibody responses to recombinant preparations of B-subunits of Escherichia coli heat-labile enterotoxin (rEtxB) and cholera toxin (rCtxB). Vaccine 13:803.)] 8-10 недель жизни. Мышам делают интраперитонеальную инъекцию с 200 мкг OVA (Sigma) эмульгированным в неполном адъюванте Фрейда (Sigma). MNL удаляют через 10 дней после инъекции, измельчают в сите из нержавеющей стали в HBSS (Flow, Irvine, UK). Удаленные клетки промывают в HBSS центрифугацией (500•g, 10 минут, 4oС) и ресуспендируют в модифицированной Eagle's среде (Flow), содержащей 20 мМ HEPES (Flow), 100 IU пенициллина, 100 мкг/мл стрептомицина, 4мМ L-глютамина и 5•10-5 М 2-меркаптоэтанола (полная среда), к которой добавляют 0,5% (вес/вес) свежей аутологичной мышиной сыворотки. Культуры содержат 2•106 жизнеспособных клеток/мл в каждых 2 мл объема в платах с 24 лунками (Nunc, Roskide, Denmark) и культивируют в присутствии 100 мкг/мл OVA (экстенсивно диализованного в полной среде), или самого по себе, или с 40 мкг/мл EtxB или EtxB (G33D). Культуры инкубируют при 37oС в 5% CO2 и 95% воздуха в течение 5 дней. В ожидаемый момент времени 0,1 мл образцы извлекают из среды и переносят на платы с 96 лунками с U-образным дном (Nunc) и импульсно метят 1 мкКи/лунку [3H]-тимидина (Amersham, UK) за 6 часов перед тем, как собрать (Mash III harvesting 96; Tomtec, Orange, CT) и посчитать стандартной жидкой сцинцилляцией (1450 Micro b plus, LKB-Wallac, Turku, Finland). Для проточного цитометрического анализа (Becton Dickinson, Erenbodegem-Aalst, Belgium) Т-клетки метят следующими крысиными антителами: (PharMin-gen, Cambridge, UK): FITC, меченный анти-СD4 (RNRM4-5) или FIТС-анти-СD8α (53-6.7) и биотин, меченный анти-СD25 (IL-2Rα) (7D4) с последующим Стрептавидин-фикоэритрином. Кроме того, для биотинмеченых антител используют FITC-меченый Стрептавидин. FACS анализы выделенных клеток проводят на пике пролиферации (4 день), как определено путем [3Н]-тимидин инкорпорации.
Для оценки апоптоза, свежие MNL-клетки (MLNC) и селезеночные Т-клетки (SPLTC) выделяют у BALB/c мышей 8-10 недель жизни. MLNC, содержащие >90% CD3+ Т-клеток, как определено при проточном цитометрическом анализе, инкубируют в течение 2 часов в чашках Петри (Costar, Cambridge, MA) в полной среде, содержащей 10% FCS, при 37oС в 5% СO2 и 95% воздуха для удаления прилипших клеток. Неприлипшие клетки последовательно отсасывают пипеткой, осаждают и дважды промывают в HBSS перед использованием. SPLTC очищают отрицательным отбором, используя стеклянные гранулы, покрытые нормальной мышиной сывороткой с последующими кроличьими анти-мышиными глобулинами. Как было описано (Wigzell Н. (1976). Specific affinity fractionation of lymphocytes using glass or plastic bead columns. Scand. J. Immunol. 5: (suppl. 5) 23). Выделенная популяция Т-клеток содержала 90% CD3+, как определено проточным цитометрическим анализом.
CD4+ и CD8+ Т-клетки разделяют следующим образом: прилипшие MLNC метят крысиными фикоэритрин-анти-мышиными CD4 (4708-02) или FITC-анти-мышиными CD8α (53-6.7) (PharMingen) и затем инкибируют с MACS коллоидными супер-парамагнитными микрогранулами, конъюгированными с козьими анти-крысиными IgG (Н + L) F(ab')2 (PharMingen), в соответствии с инструкцией по получению. Их наносят на мини-МАСS колонку (Miltenyi Biotec, Bergisch Gladbach, Germany) для разделения положительных (>99% чистоты) и отрицательных (>90% чистоты) выбранных популяций CD4 и CD8+ Т-клеток, как определено проточным цитометрическим анализом.
Два метода используются для количественного анализа апоптоза: i) DNA метят акридин оранжевым для проверки морфологии ядра и ii) анализ клеточного цикла после того, как DNA был помечен пропидиум йодидом и анти-СD4 или анти-CD8 антителами.
Культуры 2•105/мл MLNC, SPLTC и фракционированных MLNC культивируют в полной среде, содержащей 10% FCS, в отсутствие или в присутствии 80 мкг/мл или EtxB или EtxB (G33D) и проверяют через 4-18 часов. После инкубации, клетки осаждают, промывают с HBSS и метят 5 мкг/мл акридина оранжевого (Sigma).
Тимоциты выделяют и обрабатывают в отсутствие или в присутствии 10-7 М дексаметазона и используют в качестве положительного контроля для клеток, подвергнутых апоптозу. Морфологические изменения ядра в лимфоцитах исследовали нормальной или конфокальной флуорисцентной микроскопией (Leica TCS 4D). Количество CD4+ и CD8+ SPLTC в суб-G0/G1 стадии клеточного цикла определяют при проточном цитометрическом анализе содержания DNA после окрашивания пропидиум йодидом, как было описано (О'Connor P.M., Jackman J., Jondle D., Bhatia K., Magrath I. and Kohn K.W. 1993. Role of p53 tumor suppressor gene in cell cycle arrest and radiosensitivity of Burkitt's lymphoma cell lines. Cancer. Res. 53:4776). Клетки выделяют из 18-часовой культуры SPLTC, инкубируют сами по себе или с 40 мкг/мл EtxB или EtxB (G33D) меченными FITC крысиными анти-СD4 или FITC-анти-CD8α. Меченые клетки доводят до 1•106/мл холодным HBSS, содержащим 20 мМ HEPES и 0,5 мМ EDTA и фиксируют холодным этанолом, добавляемым капельно. Затем добавляют 50 мкг/мл проподиум йодида и 40 мкг/мл рибонуклеазы A (DNasa свободная) и клетки инкубируют в течение 1 часа при комнатной температуре. Относительную интенсивность DNA окрашивания проподиум йодидом в CD4 и CD8+ Т-клетках определяют путем установки дискриминационного окна на клетки, одновременно помеченные с каждым mАВ.
В Примере 1, наблюдение, что CD8+ Т-клетки полностью истощаются из культуры клеток лимфоузлов, пролифилируя в ответ на EtxB, предполагает, что EtxB вызывает поликлональное воздействие на эту Т-клеточную популяцию. Для выяснения, зависит ли подобный эффект от активации EtxB чувствительных клеток, культуры создавали из клеток OVA-праймированных мышей и стимулировали только одним OVA или OVA с EtxB или с мутантным EtxB (G33D). Одинаковые пиковые уровни пролиферации (4 день культивирования в каждом случае) достигались в присутствии одного OVA, OVA плюс EtxB или OVA плюс EtxB (G33D) (9734±347, 12,031±135 и 9305±290 c.p.m. соответственно). Однако существуют серьезные различия в распределении популяций Т-клеток в этих культурах через 4 дня (фиг. 6). Все культуры содержат CD4+ Т-клетки, из которых одинаковое количество одновременно экспрессирует активирующий маркер CD25. Однако CD8+ Т-клетки не определяются в культурах, инкубированных с OVA плюс EtxB, но очевидно присутствуют (хотя не активированные, как определено по CD25 экспрессии) в культурах с OVA плюс EtxB (G33D) или с одним OVA. Это доказывает, что EtxB вызывает истощение CD8+ Т-клеток, реагируя на антиген, отличный от EtxB. Более того, отсутствие такого ответа на EtxB (G33D) указывает на то, что истощение запускается вслед за рецепторным воздействием токсоида. Также отмечалось, что присутствие дикого типа EtxB вызывает значительное повышение количества В-клеток, среди которых большое число составляют CD25+ (не показано), как раннее было обнаружено для EtxB чувствительных культур (Пример 1). Из этого делается вывод, что рецепторное связывание EtxB оказывает абсолютное иммуномодуляторное воздействие на лимфоциты независимо от их антигенной специфичности.
Исследовалась возможность того, что CD8+ Т-клетки подвергаются апоптозу, когда культивируются в присутствии EtxB. MLNC или очищенные SPLTC от непраймированных мышей инкубируют с EtxB или EtxB (G33D) и изменения в морфологии клеточного ядра после окрашивания акридином оранжевым обнаруживают через 4-18 часов (таблица 3 и фиг. 7). Клеточные морфологические изменения характеризуются присутствием скоплений хроматина, что проявляется в появлении дольчатости ядра (фиг. 7). Также наблюдались другие клеточные структуры, такие как пузырьки плазматической мембраны и присутствие апоптозных тел. Эти морфологические изменения появлялись в каждом третьем клеточном препарате, обработанном EtxB, тогда как значительно меньший охват наблюдался в клетках, культивированных с EtxB (G33D) или без экзогенного антигена (таблица 3). Так как CD8+ Т-клетки составляют 35-40% MLNC и SPLTC препаратов, истощение этих клеток не может быть ответственным за наблюдаемый апоптоз. Для выяснения, тот ли это случай, популяция очищенных CD8 и CD4+ Т-клеток культивировали в течение 18 часов в присутствии антигенов (таблица 3). Одинаковый процент морфологических изменений был вызван в отрицательно отобранных популяциях CD4+ Т-клеток (содержащих >90% CD4-несущих клеток) при обработке с EtxB, EtxB (G33D) или без антигена, указывая, что связывание EtxB со своим рецептором не является пусковым механизмом апоптоза в этой Т-клеточной популяции. В противоположность этому, в >70% отрицательно отобранных CD8+ Т-клеток (>90% очищения) выявлены морфологические изменения при культивировании с диким типом EtxB; в то время как инкубация или без антигена, или с EtxB (G33D) вызывала изменения только в 11-19% этих Т-клеточных популяций соответственно. Кроме того, присутствие небольшого числа загрязняющих клеток в использованной очищенной популяции (~10% в каждом случае) не могли считать за наблюдаемый эффект, так как более хорошо очищенные популяции, содержащие >99% CD8 или CD4+ Т-клеток (выделенных положительным отбором) отвечали на EtxB таким же образом (60% были апоптичными в присутствии EtxB по сравнению с 7% как для отсутствия антигена, так и для EtxB (G33D) обработки) (таблица 3). Апоптоз определялся в 40% и 98% тимоцитов после 18 часов инкубации в отсутствие или в присутствии дексаметазона соответственно.
Чтобы продемонстрировать, что морфологические изменения, наблюдавшиеся в наших культурах, согласуются с индукцией апоптоза, оценивалось появление субдиплоидного DNA в культурах SPLTC, обработанных EtxB в течение 18 часов. Клетки подвергали проточному цитометрическому анализу после того, как их одновременно метили пропидиум йодидом и или анти-CD8 или анти-СD4 антителами (фиг. 8). Приблизительно 48% CD8+ Т-клеток из культур, инкубированных с EtxB, падает ниже диплоидного G0/G1 пика пропидиум йодидного окрашивания, показывая, что они подвержены апоптозу (О'Connor P.M. et al. supra). Небольшое количество клеток, экспрессирующих CD4, в культурах EtxB, также показывает суб-G0/G1 уровни DNA (~11%; которые могут быть результатом гибели такого большого количества CD8+ Т-клеток). В противоположность этому, большинство CD4 или CD8+ Т-клеток, культивированных без антигена или в присутствии EtxB (G33D), находилось в G0/G1 фазе клеточного цикла, с <0,5% проявленного апоптоза. Мы делаем вывод, что наблюдаемые ядерные морфологические изменения и присутствие суб-G0/G1 уровней DNA в значительном количестве CD8+ Т-клеток, обработанных EtxB, демонстрируют избирательный апоптоз, запущенный холероподобным энтеротоксоидом. Недостаток в рецепторном связывании мутанта, EtxB (G33D), чтобы вызвать такое воздействие, демонстрирует, что индукция CD8+ Т-клеточного апоптоза связана с их способностью связывать GM-1 ганглиозид.
Пример 3
Группы из 8 самцов DBA/l мышей не подвергаются контрольному заражению (группа А) или инъецируются 100 мкг бычьего коллагена в CFA в 0 день интрадермальной инъекцией в бок. Мыши, инъецированные коллагеном, или не защищены (группа В; положительный контроль), или сделана попытка предупредить развитие болезни интрадермальным введением в область, смежную с областью заражения коллагеном, 100 мкг EtxB в IFA в 0 день (группа С), 100 мкг EtxB в IFA на 14 день (группа D), или 100 мкг EtxB (G33D) в IFA в 0 день (группа Е). Все животные, за исключением входящих в группу А, получали возрастающие дозы коллагена в IFA интрадермально на 21 день, и тяжесть заболевания оценивалась на 45 день измерением слабости лодыжки задней конечности (эксперимент А) или учитывая каждый палец задней конечности по распуханию (значения 0-3, где 0=норма, 3=максимум распухания; эксперимент В).
Полученные результаты проиллюстрированы на фиг. 9а и 9б. Они показывают, что EtxB, но не EtxB (G33D), существенно предохраняют мышей от развития коллаген-вызванного артрита.
Пример 4а
Два раздельных человеческих образца светлого слоя кровяного сгустка (полученного из нормальной человеческой крови доноров) используют в качестве источника мононуклеарных клеток. Клетки выделяют под Ficollpaque и экстенсивно промывают перед культивированием в отсутствие антигена или с 80 мкг/мл или EtxB, или EtxB (G33D), как указывалось. До культивирования клеточные популяции содержали 24% CD8+, 27% CD4+ и 27% CD8+, 22,9% CD4+ для каждого образца соответственно. После культивирования в течение 18 часов появление апоптических клеток определяют в образцах клеток, окрашенных акридином оранжевым (как детализировано в Примере 2). Полученные результаты показаны на фиг. 10; они иллюстритуют, что EtxB, но не EtxB (G33D) вызывает апоптоз в популяции нормальных человеческих мононуклеарных клеток периферической крови.
Пример 4б
Т-клетки мышиной линии, CTLL-2, культивируют до слияния, и затем клетки промывают перед повторным посевом при 1•106 клеток/мл в отсутствие антигена или с 80 мкг/мл или EtxB, или EtxB (G33D), как было указано. Через 18 часов образцы удаляют и определяют процентное содержание клеток, показывающих признаки апоптоза, используя акридин оранжевый (детали в Примере 2). Полученные результаты проиллюстрированы на фиг. 11. Они показывают, что перекрестное связывание GM-1 ведет к апоптозу в части мышиных CTLL клеток.
Мышей инъецируют интраперитонеально 30 мкг EtxB (G33D) в полном адъюванте Фрейда (CFA). Мезентериальные лимфоузлы выделяют через 10 дней. Клетки выделяют и инкубируют в течение 4 дней в присутствии EtxB, EtxB (G33D) или разобранных мономеров этих белков (*), полученных при нагревании до 95oС. Пролиферацию определяют при добавлении 1 мкКи (3H) dThd в последние 6 часов 4 дня. Данные представляют среднюю срm и SEM из трех лунок. Клетки, выделенные из иммунизированных мышей, дают <1500 срm (доза 160 мкг/мл).
Мышей инъецируют EtxB (G33D) в CFA и клетки мезентериальных лимфоузлов изолируют 10 днями позже. Затем клетки инкубируют in vitro или с EtxB, или EtxB (G33D) и образцы супернатантов исследуют на содержание цитокинов на 5 день клеточной пролиферации.
Нуклеарные морфологические изменения во фракционированных CD4 и CD8+ Т-клетках после 4 или после 18 часов инкубации в отсутствие антигена, или с 80 мкг/мл EtxB, или EtxB (G33D) исследуют флуоресцентной микроскопией с акридином оранжевым. Все MLN из приросших были истощены. SPLTC выделяют отрицательным выделением со стеклянными гранулами, покрытыми мышиными γ-глобулинами и кроличьими анти-мышиными, как вторичными антителами. Фракционированные SPLTC получают после того, как их метят крысиными фикоэритрин-анти-мышиными CD4 или FITC-анти-мышиными CD8α, которые затем инкубируют с MACS коллоидными суперпарамагнитными микрогранулами, конъюгированными с козьими анти-крысиными IgG (Н+L) F(ab')2. Их разделяли, используя мини-MACS колонку для получения как положительных (>99% чистоты), так и отрицательных (>90% чистоты) избирательных фракций CD4 и CD8+ Т-клеток. Нуклеарные морфологические изменения выявлялись с 4 по 18 часы в случайно выбранных образцах из 200 клеток на опыт, как описывалось в описании к фиг. 7. Максимальное число клеток с апоптозом появилось после 18 часов. Данные в скобках, когда указывается результат из другого отдельного эксперимента. Данные для MLN и SPLTC представлены как общие из четырех экспериментов.
| название | год | авторы | номер документа |
|---|---|---|---|
| ВЕКТОРЫ ДЛЯ ДОСТАВКИ МОЛЕКУЛ К ЭКСПРЕССИРУЮЩИМ CD11b КЛЕТКАМ | 2001 |
|
RU2312143C2 |
| БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА К CD25 И Fc ГАММА-РЕЦЕПТОРУ ДЛЯ ЭЛИМИНАЦИИ ОПУХОЛЕСПЕЦИФИЧЕСКИХ КЛЕТОК | 2017 |
|
RU2759970C2 |
| МИКРООРГАНИЗМ-НОСИТЕЛЬ НУКЛЕОТИДНЫХ ПОСЛЕДОВАТЕЛЬНОСТЕЙ, КОДИРУЮЩИХ АНТИГЕНЫ И БЕЛКОВЫЕ ТОКСИНЫ | 2007 |
|
RU2447145C2 |
| СПОСОБ ЛЕЧЕНИЯ АУТОИММУННОГО ЗАБОЛЕВАНИЯ С ИСПОЛЬЗОВАНИЕМ БИОСОВМЕСТИМЫХ БИОАБСОРБИРУЕМЫХ НАНОСФЕР | 2011 |
|
RU2635537C2 |
| СПОСОБ ЛЕЧЕНИЯ РАКА | 2016 |
|
RU2770447C2 |
| ВАКЦИНА НА ОСНОВЕ В-КЛЕТОК, НАГРУЖЕННЫХ ЛИГАНДОМ Т-КЛЕТОК-ПРИРОДНЫХ КИЛЛЕРОВ И АНТИГЕНОМ | 2006 |
|
RU2420311C2 |
| КОМБИНИРОВАННАЯ ТЕРАПИЯ ДЛЯ ЛЕЧЕНИЯ РАКА ПУТЕМ ВНУТРИВЕННОГО ВВЕДЕНИЯ РЕКОМБИНАНТНОГО MVA И АНТИТЕЛА | 2018 |
|
RU2795103C2 |
| КОМБИНИРОВАННЫЙ ПРЕПАРАТ ДЛЯ УВЕЛИЧЕНИЯ ИММУНОГЕННОСТИ ВАКЦИНЫ (ВАРИАНТЫ) | 2008 |
|
RU2491090C2 |
| АГЕНТЫ И СПОСОБЫ, ОСНОВАННЫЕ НА ПРИМЕНЕНИИ ДОМЕНА EDA ФИБРОНЕКТИНА | 2006 |
|
RU2430738C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ПРОФИЛАКТИКИ И ЛЕЧЕНИЯ ЗЛОКАЧЕСТВЕННЫХ НОВООБРАЗОВАНИЙ | 2011 |
|
RU2623038C2 |


Изобретение относится к медицине и касается терапевтического агента для лечения аутоиммунного заболевания. Сущность изобретения включает применение термолабильного энтеротоксина [Etk], В-субъединицы термолабильного энтеротоксина [EtxB] и их смесей, обладающих ганглиозид GМ-1 (GM-1)-связывающей активностью для лечения или предупреждения аутоиммунных заболеваний. Преимущество изобретения заключается в том, что указанные субъединицы, связываясь с GM-1 ганглиозидными рецепторами, вызывают активацию В-клеток. 2 з.п.ф-лы, 11 ил., 3 табл.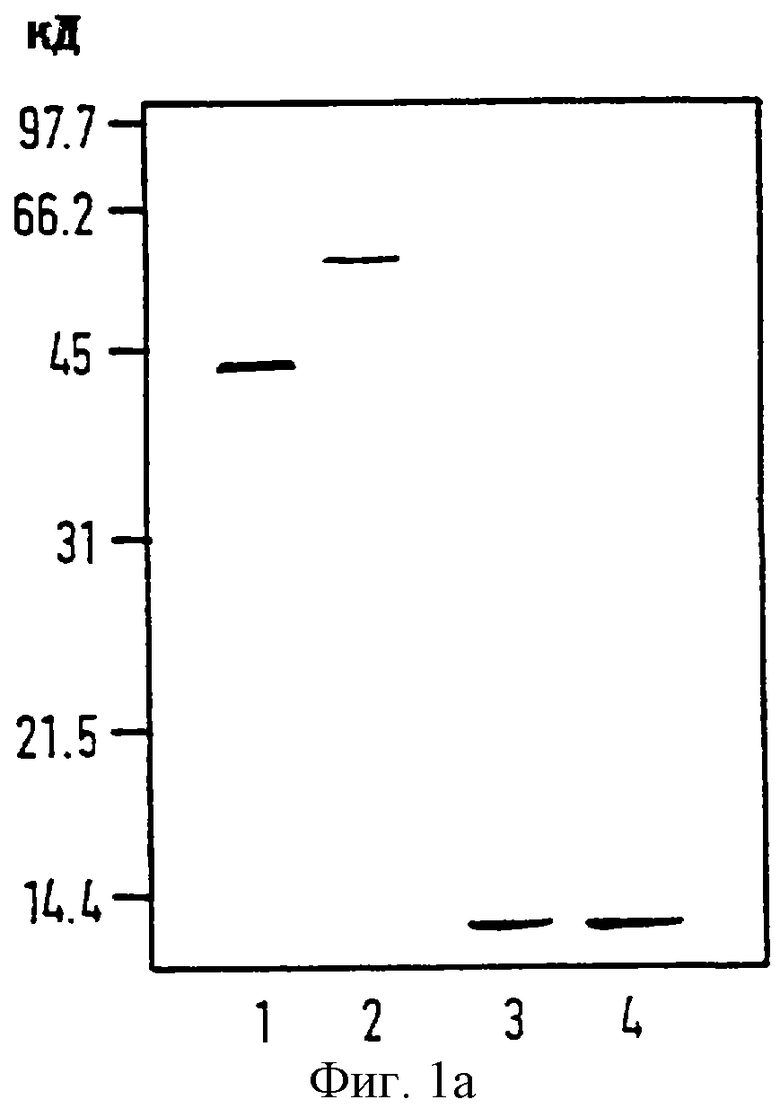
| US 4361549 А, 30.11.1982 | |||
| ELSON et al | |||
| Morfologic and functional alterations of Mucosal Т-cells by cholera toxin and its B-subunit | |||
| J | |||
| of Immunology, 1995, v.154, pp.3611-3617. |

