Изобретение относится в основном к достижениям в молекулярной биологии и технологии рекомбинантных ДНК, изобретение касается способов и средств включая анализы и фармацевтические наборы, содержащие требуемые реагенты и средства, для обнаружения в установке in vitro и ex vivo присутствия в биологической пробе целевой нуклеиновокислотной последовательности или экстраполированной КРН-последовательности из соответствующей целевой ДНК-последовательности, и путем дедукции соответствующего полипептида, который кодирует последовательность РНК (ДНК).
Изобретение отличается проведением амплификации такой конкретной целевой последовательности нуклеиновых кислот в самоподдерживающейся in vitro системе, причем амплификацию целевой нуклеиновокислотной последовательности осуществляют путем получения множества транскрипт-продуктов, которые имеют необязательную способность саморепликации. Эта самоподдерживающаяся система амплификации устраняет необходимость повторной денатурации нуклеиновокислотных дуплексов, которые нуждаются в температурной циклизации. Изобретение по-новому объединяет все реагенты, необходимые для образования в одной реакционной установке амплифицированного продукта или продукта, пригодного для обнаружения в амплифицированной форме, представляющей наличие целевой нуклеиновокислотной последовательности.
Среди применений, в которых находит использование изобретение, следует отметить анализы РНК-последовательностей или путем экстраполирования ДНК-последовательностей, которые являются характерными для конкретного или общего патогенного заболевания или состояния путем зондовой гибридизации общих вод организма и тканей, содержащих требуемую целевую нуклеиновокислотную последовательность (и).
В данной области техники одной из целей является обнаружение различных последовательностей нуклеиновых кислот в биологической пробе, в которой данная последовательность, так называемая целевая нуклеиновая кислота, присутствует в малых количествах по отношению к ее присутствию среди широкого круга других нуклеиновокислотных видов, включая РНК, ДНК или и то и другое. Таким образом желательно обнаружить нуклеиновую кислоту, кодирующую полипептиды, которые могут ассоциироваться с патологическими заболеваниями или состояниями, как например, нуклеиновая кислота, взаимосвязанная с нуклеиновой кислотой вируса иммунодефицита человека (ВИЧ-1).
Кроме определения нуклеиновых кислот, кодирующих полипептиды, желательно определить другие нуклеиновые кислоты, характерные для патологического заболевания или состояния, как например, дефективный ген, как в случае обнаружения дефективного гена человеческого бетаглобина, имеющего место при гемофилии.
Нуклеиновые кислоты, ассоциируемые с ними, присутствуют в малых количествах относительно всей нуклеиновой кислоты в данной биологической пробе, такой как проба крови или другой общей воды организма, или проба ткани данного исследуемого пациента. Обнаружение таких нуклеиновокислотных видов предполагает такую специфичность, чтобы данная разновидность нуклеиновых кислот могла быть обнаруживаемой и измеряемой среди широкого круга других нуклеиновокислотных видов, с которыми она ассоциируется. Некоторые из этих видов могут нести тесную гомологию по крайней мере в выделенных сегментах с целевой нуклеиновой кислотой. Кроме того, эти целевые нуклеиновокислотные виды очень часто находятся только в очень малых количествах в биологической пробе, подлежащей исследованию. Для проведения правильного диагноза состояния болезни важно, чтобы даже маленькие количества такой целевой нуклеиновой кислоты можно было обнаружить недвусмысленно для правильности системы анализа.
Для достижения данной цели известного уровня техники разработаны различные подходы, один из которых заключается в том, что количество нуклеиновой кислоты в пробе не изменяется или не подвергается взаимодействию. Кроме того, разработана система, при которой большое число обнаруживаемых молекул, соответствующих нуклеиновокислотной мишени, продуцируется для целей легкого обнаружения и измерения. Такая система представляет собой сигнал-генерирующую систему, ассоциированную с целевой нуклеиновой кислотой, продуцирующей обнаруживаемый сигнал, который является репрезентативным ряда молекул целевой последовательности.
Разработан другой подход, который включает повышение числа копий самой целевой нуклеиновокислотной последовательности. Это можно сделать путем селективной амплификации целевой последовательности нуклеиновых кислот. Можно рафинировать культуральные методики пробы таким образом, чтобы целевая нуклеиновокислотная последовательность амплифицировалась предпочтительно к другим нуклеиновокислотным последовательностям. Данные методики затруднительны и расходуют много времени, подвергаясь пробам и ошибкам.
Другим примером данного подхода является амплификация целевой нуклеиновокислотной последовательности в так называемой "цепной полимеразной реакции" ЦПР (Саики и др. Science 230,1350,(1985); Муллис и др., заявка ЕР N 200362 и 201184, патент США N 4683195 и 4683202), и в частности предполагает гибридизацию к сегменту целевой нуклеиновокислотной последовательности праймера, удлинение указанного праймера полимеразой и придание одноцепочности дуплексам, полученным в результате реакции праймерного удлинения. Данную методику можно повторять на протяжении нескольких циклов с тем, чтобы амплифицировать целевую нуклеиновокислотную последовательность.
Известно использование стадии получения нового РНК-трансерипта в сочетании с синтезированной копией двунитевой кДНК целевой последовательности, оперативно связанной с промотором (заявка на патент США N 07/064141 и N 07/202978; РСТ N WO 88/10315). Благодаря стадии транскрипции, являющейся доминантной в смысле новизны, можно говорить о системе амплификации на транскрипционной основе (ТАС).
Изобретение предусматривает обнаружение in vitro или ex - vivo по крайней мере одной специфической нуклеиновокислотной последовательности (целевой последовательности или сегмента) в пробе, содержащей нуклеиновую кислоту, при этом предлагается способ получения двунитевой нуклеиновой кислоты, содержащей последовательность, соответствующую целевой последовательности, оперативно связанной с РНК-полимеразным промотором, использование двунитевой нуклеиновой кислоты в качестве матрицы двунитевой нуклеиновой кислоты для получения из нее множества РНК-транскриптов, каждый из которых несет РНК-последовательность, соответствующую целевой последовательности, и обнаружение присутствия указанной РНК-последовательности, а также по аналогии присутствия целевой последовательности.
Двунитевый нуклеиновокислотный темплат (матрицу) в свою очередь получают путем создания первого нуклеиновокислотного праймера или зонда, содержащего промоторную последовательность, оперативно связанную с последовательностью, соответствующей сегменту целевой последовательности, гибридизации в пригодных условиях первого нуклеиновокислотного праймера с целевой последовательностью в пробе, содержащей нуклеиновую кислоту, удлинения гибридизированного первого нуклеиновокислотного праймера в реакции полимеразного удлинения комплементарно целевой последовательности с образованием соответствующей дуплексной нуклеиновой кислот, отделения нитей дуплекса, гибридизации в пригодных условиях к отделенной нити промоторсодержащей последовательности второго нуклеиновокислотного праймера в конце, который противоположен промоторной последовательности, и удлинение второго нуклеиновокислотного праймера в цепной полимеразной реакции комплементарно, промоторсодержащей последовательности.
Таким образом изобретение позволяет получить однонитевый РНК-транскрипт или РНК-ДНК-дуплекс, образованный из него, когда не предпринимаются меры для предотвращения его образования, причем он имеет последовательность, соответствующую целевой нуклеиновой кислоте. Продукт однонитевого РНК-транскрипта позволяет непосредственно обнаружить целевой сегмент без использования затруднительных, склонных к ошибкам повторных ЦПР-циклов и разделения нитей. Такие преимущества не дает методика ЦПР, которая приводит к получению двунитевой ДНК (одна нить которой содержит целевой сегмент, а другая нить содержит комплемент целевого сегмента), которую не нужно разделять перед обнаружением, а только после того, как большое число повторных циклов достигли приемлемых уровней амплификации.
Целью изобретения является использование основного репликативного процесса для амплификации с тем, чтобы облегчить обнаружение целевых нуклеиновокислотных последовательностей, достигая тем самым экспоненциального копирования без необходимости в температурной циклизации, и иным образом регистрируя течение способа амплификации в отношении прибавления реагентов и т.д. Другой целью изобретения является объединение новым образом достижений методик транскрипции и удлинения продуктов в качестве средства обнаружения и измерения соответствующей целевой нуклеиновой кислоты.
Главной целью изобретения является ферментативное применение селективного переваривания РНК-нити РНК-/ДНК-дуплекса, образованного гибризацией праймера с целевой последовательностью нуклеиновых кислот, с последующим праймерным удлинением, как средство получения ДНК-нити в качестве матрицы с целью дальнейшей гибридизации с ней и с последующим праймерным удлинением. Двунитевый ДНК-дуплекс продукта содержит по крайней мере одну промоторную последовательность, которая распознается ДНК-зависимой РНК-полимеразой и таким образом служит в качестве матрицы для получения множества транскриптов, которые в конце концов обнаруживают и измеряют в качестве средств обнаружения и измерения целевой нуклеиновокислотной последовательности. Данная цель обеспечивает получение преимуществ амплификации целевой последовательности, т.е. амплификация, является самоподдерживающейся без необходимости температурной циклизации в единой реакционной смеси, содержащей три подходящие ферментативные активности и по крайней мере один праймер, содержащий промотор, оперативно узнаваемый ДНК-зависимой РНК-полимеразой.
Таким образом, полной целью изобретения является объединение и решение задач известного уровня техники и создание селективных средств для дальнейшей благоприятной амплификации целевых нуклеиновокислотных последовательностей.
Изобретение также позволяет создать простую методику, которую можно использовать репродуктивно за приемлемо короткий период времени в системе изотермической реакции, используя известные реагенты и имея точность, необходимую для достижения последовательных научных результатов, причем данную методику можно использовать в репродуктивном анализе и приспосабливать для применения в наборах для лабораторных/клинических анализов.
Изобретение направлено на использование средств и способов "нитеразделения" дуплекса РНК/ДНК с тем, чтобы освободить нить ДНК для гибридизации с олигонуклеотидами, содержащими РНК-полимераза-связующем последовательности (РСП), с последующим удлинением праймера с тем, чтобы образовать ДНК-дуплекс (двунитевую ДНК), который может служить в качестве матрицы для получения множества соответствующих РНК-транскриптов, которые восприимчивы к обнаружению и измерению, в качестве выведенного анализа на присутствие целевой нуклеиновокислотной последовательности. Дуплекс РНК/ДНК в свою очередь получают гибридизацией с мишенью РНК или праймером ДНК, оперативно содержащими промоторную последовательность, после чего осуществляют праймерное удлинение.
Средства изобретения, вызывающие "нитеразделение", включают использование фермента, имеющего РНКаза Н-подобную активность, такую как РНКаза Н, который бы селективно и предпочтительно переваривал РНК-нить дуплекса с тем, чтобы ДНК-нить можно было освободить для дальнейшего процессинга. Использование такого фермента устраняет использование температурного цикла для денатурации дуплекса.
Целевой последовательностью нуклеиновых кислот может быть та, которая присутствует от природы в пробе нуклеиновой кислоты, или ею может быть продукт экстраполирования соответствующей ДНК целевой последовательности. Продукт экстраполирования получают денатурацией целевой последовательности двунитевой ДНК, гибризацией с ней последовательности праймера, имеющей оперативно ассоциированную промоторную последовательность, с последующей реакцией праймерного удлинения, образуя дуплекс ДНК. Данный ДНК/РНК-дуплекс в свою очередь денатурируют, и нить, содержащую промоторную последовательность, гибридизируют к противоположному от промотора концу с вторым праймером, с последующим праймерным удлинением, образуя ДНК-дуплекс, который при взаимодействии с ДНК-зависимой РНК-полимеразой продуцирует соответствующий транскрипт, продукт экстраполяции. РНК затем служит в качестве целевой нуклеиновокислотной последовательности для целей изобретения.
После получения целевой последовательности нуклеиновых кислот (каким бы ни был ее источник) в соответствии с изобретением ее гибридизируют с праймерной последовательностью, оперативно ассоциированной с промоторной последовательностью, с последующим праймерным удлинением, получая РНК/ДНК-дуплекс, содержащий промоторную последовательность у 5'-конца нити ДНК. Реакцию праймерного удлинения можно осуществлять с любой пригодной полимеразой, например, обратной транскриптазой. Основной вариант изобретения тогда служит для освобождения ДНК-нити РНК/ДНК-дуплекса путем обработки РНК/ДНК-дуплекса ферментом, который селективно переваривает РНК-нить, таким как РНКаза Н. Освобожденную таким образом ДНК-нить либо подвергают самогенерированному праймерному удлинению посредством РНК-праймера, полученного в результате предыдущего селективного переваривания, либо гибридизируют с имеющим внешнее происхождение олигонуклеотидным праймером, по выбору несущим промоторную последовательность. Удлинение праймера создает двунитевую ДНК, содержащую одну или две промоторные последовательности , которая служит в качестве матрицы (темплата) для ДНК-зависимой РНК-полимеразой индукции транскрипции с получением множества транскриптов. Последовательность реакций можно осуществлять изотермически и в одной смеси с тремя соответствующими ферментными активностями, такими, которые дают обратная транкриптаза, РНКаза и и ДНК-зависимая РНК-полимераза, при этом различные стадии в вышеприведенном процессе осуществляют непрерывно, одновременно в течение определенного периода времени. Таким образом в конечной точке процесса продуцируемые транскрипты можно обнаружить и измерить на выведенное присутствие исходной целевой нуклеиновокислотной последовательности. Однако, хотя непрерывный и одновременный характер вышеописанной последовательности реакций является независимым от температурной циклизации и предполагает только единственное, первоначальное прибавление ферментных активностей, необходимых для осуществления данных реакций, сами продукты транксриптов подвергаются гибридизации с праймером, необязательно несущим дополнительную промоторную последовательность. Данный гибридизационный комплекс подвергают реакции праймерного удлинения с получением второго РНК-ДНК-дуплекса, который, в свою очередь, подвергают воздействию фермента селективного переваривания РНК для освобождения из него ДНК-нити, которую в свою очередь гибридизируют с самогенерирующимся РНК-праймером или имеющим внешнее происхождение олигонуклеотидным праймером, присутствующим в реакционной смеси, который может по выбору нести оперативно ассоциированную промоторную последовательность с тем, чтобы получить второй ДНК-дуплекc, восприимчивый к распознаванию ДНК-зависимой РНК-полимеразой, получая множество транcкриптов, имеющих смысл противоположный транскриптам, продуцированным первоначально.
Хотя еще не выявлен полностью механизм вышеприведенной последовательности (ей) реакций, полагают, что поскольку реакционную смесь используют в сочетании с тремя приемлемыми ферментными активностями, такими, которые дают обратная транскриптаза, РНКаза Н и ДНК-зависимая РНК-полимераза, и по крайней мере одним праймером, содержащим промотор, распознаваемый полимеразой, и поскольку реакции не зависят от температурной циклизации, используются два промоторсодержащих олигонуклеотидных праймера, и реакции могут проходить через несколько циклов одновременно и непрерывно, продуцируя как смысловые, так и антисмысловые транскрипты, которые могут быть обнаружены и измерены с получением амплификационного анализа количества целевой нуклеиновокислотной последовательности, присутствующего в исследуемой пробе.
Сущность изобретения способствует созданию реакционной смеси, находящейся в состоянии покоя в течение периода времени при пригодной температуре без необходимости в проведении циклизации между более высокими и более низкими температурами и без необходимости в периодическом прибавлении фермента или других реагентов, посредством чего целевая нуклеиновокислотная последовательность амплифицируется непрерывно и одновременно самогенерирующимся образом в присутствии по крайней мере одного праймера, оперативно несущего промоторную последовательность и ферментную активность такую, которую дает РНК-полимераза, которая распознает полимераза-связанный сайт промотора, обратная транскриптаза, фермент, такой как РНКаза Н, который селективно переваривает РНК, когда РНК гибридизируют к ДНК в дуплексной форме, и необходимые субстраты нуклеозид-трифосфата для РНК-полимеразы и обратной транскриптазы. В такой системе воспроизводимые уровни амплификации до 107 могут быть достигнуты приблизительно за 2 ч при температуре около 37oC, используя, например, РНК-полимеразу Т7, обратную транскриптазу АМV и РНКазу Н Е. соli. Операбельной является температура приблизительно 4 - 50°С, предпочтительно около 40°С. В предпочтительных вариантах размер целевой последовательности нуклеиновых кислот составляет менее, чем приблизительно 250 оснований. Другие переменные величины могут воздействовать на оптимизацию амплификации, такие как использование РНК-полимеразы, использование обратной транскриптазы, рН концентрации солей, концентрации нуклеозид-трифосфата.
Таким образом, изобретение включает обнаружение in vitro или ex vivo по крайней мере одной специфической нуклеиновокислотной целевой последовательности в пробе, содержащей гетерогенную коллекцию РНК. Изобретение предусматривает способ, включающий получение двунитевой ДНК, кодирующей последовательность, соответствующую целевой последовательности и имеющую оперативный РНК- полимеразный промотор, причем двунитевую ДНК получают, в свою очередь, гибридизацией, с последующим праймерным удлинением к ДНК-нити, которая освобождена от ее РНК-комплемента в дуплексе РНК/ДНК, под действием фермента селективного переваривания РНК, при этом дуплекс РНК/ДНК образован гибридизацией с праймером, несущим оперативно промоторную последовательность, к целевой нуклеиновокислотной последовательности с последующим праймерным удлинением. Двунитевая ДНК служит в качестве темплата для получения множества РНК-транскриптов, каждый из которых переносит РНК-последовательность, соответствующую целевой нуклеиновокислотной последовательности. Присутствие РНК-последовательности, а путем дедукции, присутствие целевой последовательности можно определить и измерить в соответствии с изобретением.
Изобретение направлено на все ассоциированные способы и средства получения и использования таких РНК-транскриптов. В одном варианте изобретения предлагается необязательно периодически повторяющийся способ получения двунитевого нуклеиновокислотного темплата, определенного выше, с получением первого нуклеиновокислотного праймера, содержащего промоторную последовательность, оперативно связанную с последовательностью, соответствующей сегменту целевой нуклеиновокислотной последовательности, гибридизацией в пригодных условиях первого нуклеиновокислотного праймера с целевой нуклеиновокислотной последовательностью в пробе, содержащей нуклеиновую кислоту, удлинением гибризированного первого нуклеиновокислотного праймера в реакции полимеразного удлинения комплементарно целевой последовательности, с образованием соответствующей нуклеиновой кислоты РНК/ДНК-дуплекса, ферментативным расщеплением РНК РНК/ДНК-дуплекса, гибридизацией к освобожденной нити промоторсодержащее, ДНК последовательности в пригодных условиях второго нуклеиновокислотного праймера в конце, который противоположен промоторной последовательности, либо посредством продукта, имеющего происхождение от РНК-праймера, либо посредством олигонуклеотидного праймера, необязательно содержащего промоторную последовательность, оперативно связанную с ним, удлинением гибридинированного второго нуклеиновокислотного праймера в реакции полимеразного удлинения комплементарно промотор содержащей последовательности.
Изобретение в другом варианте направлено на способы и средства использования двунитевой нуклеиновой кислоты в качестве темплата для получения множества РНК-трансриптов в реакции, катализируемой ДНК-зависимой РНК-полимеразой, которая распознает его промотор, и после осуществления дополнительной необязательной циклизации обнаруживают и измеряют наличие РНК-трансриптов.
В другом варианте изобретение направлено на усовершенствование в способе амплификации целевой нуклеиновокислотной последовательности, генерированной как таковой или в виде экстраполяционного продукта из целевой ДНК-последовательности, причем осуществляют гибридизацию с целевой последовательностью ДНК-праймера, имеющего промоторную последовательность, оперативно связанную с ней, с последующим праймерным удлинением, получая соответствующий РНК/ДНК-дуплекс, гибридизацию освобожденного продукта ДНК-удлинения в виде нити, несущей промоторную последовательность дуплекса, с вторым праймером в конце, противоположном поромоторной последовательности, с последующим праймерным удлинением, образующим двунитевый ДНК-темплат, пригодный для получения по выбору реплицируемых РНК-транскриптов для обнаружения как таковых или для рециркуляции, как определено выше. Усовершенствование включает высвобождение ДНК-нити продукта удлинения, несущей промоторную последовательность праймера, дуплекса РНК/ДНК, путем селективного ферментативного расщепления РНК-нити РНК/ДНК-дуплекса.
В одном варианте изобретение направлено на продукт процесса обработки РНК дуплекса РНК/ДНК, имеющий примыкающую к 5'-концу ДНК-последовательности промоторную последовательность, с помощью фермента селективного РНК-переваривания.
В другом варианте изобретение направлено на создание наборов, включающих требуемые реагенты и ассоциируемые средства, пригодные при обнаружении in vitro или ex - vivo по крайней мере одной специфической нуклеиновокислотной целевой последовательности в пробе, содержащей РНК, в использованием способов и средств, определенных выше.
На чертеже показано схематическое изображение варианта изобретения. Схематическое изображение создает представление о 12 стадиях, которые в предпочтительных вариантах являются непрерывными, самогенерирующимися стадиями в присутствии требуемых трех ферментов в реакционной среде вместе с целевой последовательностью и данной операбельной температурой. Тремя ферментами являются перечисленные выше обратная транскриптаза (RT), используемая для реакции прафмерного удлинения, РНКаза Н, которая является ферментом селективного РНК-переваривания, представляя основной аспект изобретения, и РНК-полимераза Т7 в качестве примера фермента, пригодного для получения транскриптов из темплата ДНК-дуплекса. Сущность изобретения представлена стадией 3 схематического изображения, причем продукт РНК-транскрипта стадии 6 может быть обнаружен и измерен как таковой или может быть подвергнут непрерывной реакции, как указано в стадиях 7 - 12, с образованием антисмысловой нити РНК-транскрипта. Генерированные РНК-транкрипты обнаруживают и измеряют как результат присутствия целевой нуклеиновокислотной последовательности. РВS обозначает сайт полимеразного связывания промоторной последовательности, ТСS обозначает целевую комплементарную последовательность.
В изобретении используются ссылки на стандартные руководства по молекулярной биологии, которые содержат определения, а также способы и средства для осуществления основных методик изобретения, например:
получение ДНК-зонда или праймера, включая синтез ДНК или выделение последовательностей из натуральных источников посредством расщепления рестрикционными ферментами и их наращивание с тем, чтобы они были пригодны как таковые или, когда присоединения к другой ДНК, для использования в качестве праймера или зонда;
получение олигонуклеотидов с различными функциональными последовательностями для использования при гибридизации;
методология гибридизации, включающая варианты в строгих условиях для получения большей или меньшей гибридизации, зависящей от степени гомологии праймера к целевой ДНК-последовательности;
идентификация, выделение или получение промоторов или более конкретно промоторов или сайтов, распознаваемых бактериофаг ДНК-зависимой РНК-полимеразой и бактериофаг РНК-зависимой РНК-полимеразой, или при использовании эукариотных систем, вирусной ДНК- и РНК-зависимой РНК-полимеразы, например аденовирус, кодируемой РНК-полимеразы и РНК-полимеразы вируса мозаики костра;
идентификация, выделение или получение РНК-полимеразы, способной распознавать промоторы или способной к реакции праймерного удлинения;
условия, способствующие производству РНК-транскриптов, включая так называемые транскрипционно-энхансерные последовательности;
условия, способствующие инициации и сохранению реакций праймерного удлинения, включая использование ДНК-зависимой полимеразы и дНТФ;
механизм и методология для индуцированной репликации, и т.д. и т.п. (Maniatis at al., Molecular Cloning: A laboratory Manual, Cold Spring Harbor Laboratory, New York, 1982; and Colowick et al., Methods in Enzymology Vol. 152, Academic Press, Inc, 1987; Hohg, Biocsience Reports 1, 243, 1981; Cooke et al. J.Biol. Chem. 255, 6502, 1980; and Zoller et al., Methods in Enzymology 100, 468-500, 1983; and Crea et al., Nucleic Acids Res. 8, 2331, 1980; Narang et al. , Meth. Enzyme. 68, 90, 1979; Beaucage et al., Tetrahedron Letters 22, 1859, 1981; Brown et al., Meth. Enzym. 68, 109, 1979; Caruthers et al., Meth. Enzum. 154, 287, 1985; Hitzeman et al., J. Biolog. Chem. 255, 2073, 1980, Lee et al., Science 239, 1288, 1988; Milligan et al. Nucleic Acils Res. 15, 8783, 1987; Miller et fl, Virology 125, 236, 1983; Aahlguist et al. , J. Mol.Biol. 153, 23, 1981; Miller et al., Nature 313, 681, 1985; Ahlguist et al., J.Mol.Biolog. 172, 369, 1984; Ahlguist et al., Plant. Mol. Biol. 3, 37, 1984; Ou et al., Pnas 79, 5235, 1982; Chu et al., Nucl. Acids Res. 14, 5591, 1986; заявка на Европатент N (ЕРА) 194809, Marsh et al., Positive STrahg RNA Viruses, p. 327-33, Aalan R.Liss (опублик, Нью-Йорк) 1987, материалы Симпозиума Ucla, 1986, Miller et al., J.Vol.Biol, 187,537, 1986; Staflet et al., Science 239, 491, 1988; Kramer et al., J.Mol.Biol. 89, 719, 1974Saris et al., Nucl. Acids Res. 10, 4831, 1982; Bresser et al., Pnas 80, 6523, 1983; Chu et al., Nucl. Acids Res. 16, 3671, 1989; Gubler et al., Gene 25, 263, 1983; and D"Alessio et al., Nucleic Acids Res., 16, 1999, 1989), а также отсылки, приводимые в изобретении, причем каждая из этих вышеприведенных публикаций введена в качестве отсылки.
Под термином "промотор" подразумевается последовательность нуклеиновых кислот (встречающаяся в природе или полученная синтетически, или как продукт рестрикционного переваривания), которая специфически распознается РНК-полимеразой, которая связывается с распознанной последовательностью и инициирует процесс транскрипции, посредством чего продуцируется РНК-транскрипт. Промотор может необязательно содержать нуклеотидные основания, простирающиеся за пределы фактического сайта распознавания, придавая дополнительную устойчивость относительно процессов деградации, и также может включать дополнительные плюс (+) нуклеотиды, примыкающие к сайту транскрипционной инициации. В принципе можно использовать любую промоторную последовательность, для которой существует известная и доступная полимераза, способная распознавать последовательность инициации. Типичными известными и полезными промоторами являются тем, которые распознаются некоторыми бактериофаг-полимеразами, такими как бактериофаг Т3, Т7 или SР6 (Siebenlist et al., Cell, 20, 269, 1980). Они являются примерами тех полимеров, которые можно использовать в практике изобретения в сочетании с их ассоциируемыми промоторными последовательностями.
"РНК-транскрипт" представляет собой последовательность рибонуклеиновой кислоты, продуцируемую после инициации транскрипции вслед за распознаванием РНК-полимеразой промоторной последовательности. Получение таких транскриптов является более или менее непрерывным, частично в зависимости от количества присутствующей полимеразы.
Под термином "зонд" или "праймер" подразумевается однонитевая последовательность нуклеиновых кислот (встречающаяся в природе или полученная синтетически, или же являющаяся продуктом рестрикционного переваривания), которая имеет достаточную комплементарность с целевой последовательностью, такой, которая при пригодных условиях гибридизации способна гибридизировать, т.е. связывается с соответствующей (целевой) последовательностью. Типичный зонд или праймер имеет по крайней мере около 10 нуклеотидов в длину и, наиболее предпочтительно, около 20 или более нуклеотидных оснований в длину, и в наиболее предпочтительном своем варианте разделяет идентичность или очень высокую комплементарность с соответствующей (целевой) последовательностью (ЕР, заявка N 128042, опублик. 1984). Такой зонд или праймер таков, что он гибридизирует к комплементарной последовательности для целевой реакции праймерного удлинения в присутствии соответствующих реагентов и условий.
Термин "оперативно связанный" или "ассоциируемый", или его грамматические варианты, в частности, в смысле связи промоторной последовательности в пределах РНК-кодирующей ДНК-последовательности, относится к функциональности при получении соответствующих РНК-транскриптов, когда промотор распознается пригодной полимеразой.
Известны методики формирования сигнала обнаружения, например, путем радиоактивного мечения или хромогенных средств с использованием хромогенного восприимчивого фермента.
Проба, на которой осуществляют метод испытания в соответствии с изобретением, может быть неочищенным образцом биологического материала, например сывороткой или другой общей водой организма, средой тканевой культуры или пищевым материалом. Более типично, когда метод осуществляют на пробе, которая является переработанным образцом, имеющим происхождение от сырого образца, с использованием различных обработок с тем, чтобы удалить материалы, которые могли бы воспрепятствовать обнаружению мишени, например, вызывая неспецифическое связывание родственных молекул. Известны методы переработки сырых образцов с получением пробы, которая является более пригодной для метода испытания изобретения.
Транскрипты (РНК) могут быть обнаружены различными путями. Обнаружение можно проводить ультрафиолетовым поглощением РНК, например, методом контактной фотопечи (Kutateladze et al. Anal. Biochem. 100,129, 1979).
Используя радиоактивно меченый рибонуклеозид - 5'-трифосфат в реакции, например, 3H-меченый или альфа-32РО4-меченый, так, что РНК является радиоактивной, РНК можно обнаружить любым из целого ряда известных способов посредством ее радиоактивности.
Биотин или иминобиотин можно вводить в РНК, которую затем можно обнаружить известными способами с аддуктом ферментавидина или фермен стрептавидина, который связывается с РНК-связанным биотином и катализирует получение удобно обнаруживаемого хромогена. Введение биотина или иминобиотина можно осуществлять путем использования УТФ (уридин-5'-трифосфат), который является биотинилированным посредством спейсера к углероду-5-урацильной части субстрата для репликазы в реакции репликации. Такие УТФ являются известными соединениями. Кроме того, известно, что такие УТФ представляют собой субстраты для QВ-репликазы, и что РНК, которые включают урацилы, биотинилированные посредством спайсерных групп, присоединенных к положению углерода-5'- вследствие использования таких УТФ в синтезе, являются темплатами для репликации, катализируемой QB-репликазами.
РНК можно сделать флуоресцентными путем использования реакции, катализируемой РНК-лигазой Т4, с прикреплением нуклеотидов, модифицированных к условиям флуоресценции, к 3'- концу репликативной РНК (Cosstick et al., Nucl. Acids Res, 12,1791, 1984. Флуоресценцию полученной РНК можно использовать для обнаружения РНК любыми стандартными методами. Среди других методов, которые могут быть использованы для обнаружения РНК, такие, в которых вещество, связываемое специфически с нуклеиновой кислотой, прибавляют в систему, в которой имела место репликация, или в среду, такую как положительно заряженный носитель, например бумага ECTEOL A, на которой выделяют реплицированную РНК и измеряют сигнал от данного вещества. Такие вещества включают хромогенные красители, такие как "универсальный краситель" (Lahlhberg et al.,J.Molec. Biol. 41, 139, 1969), метиленовый голубой (Dingman et al., Biochemistry, 7, 659, 1968) и серебрянный краситель (Sammons et al., Electrophoresis 2, 135, 1981; Igloi, Anal.Biochem. 184, 1983), флуорогенные соединители, которые связываются с РНК, например бромид этидия (Sharp et al., Biochemistry 12, 3055, 1973; Bailey et al., Anal.Biochem. 70, 75, 1976), а также флуорогенные соединения, которые связываются специфически с РНК, являющимися темплатами для репликации путем ОВ-репликаз, например фикобилипротеин (Oi et al., J.Cell. Biol. 93, 981, 1982; Stryer et al., патент США N 4520110), сопряженный с вирусной субединицей ОВ-репликазы.
В анализах в соответствии с изобретением исследования осуществляют одновременно в условиях, которые как можно более схожи относительно исследуемых и контрольных проб. Известно, что контрольные пробы аналогичны исследуемым пробам, однако не содержат ни мишени, ни известного количества мишени. Контрольная проба без мишени устанавливает "фон", ниже которого нет возможности отличать пробы, которые содержат мишень, от тех, которые не содержат. Путем сравнения количества или концентрации РНК, продуцируемой в анализе исследуемой пробы, с количеством или концентрацией, продуцируемой контрольными пробами, исследуемыми одновременно, можно определить наличие мишени в исследуемой пробе на уровне выше фонового. Если контрольные пробы в пределах известных концентраций мишени используются в анализе, можно оценить концентрацию мишени в исследуемой пробе.
Известно использование "репликазы" для автокаталитической индукции репликации необязательно реплицирующихся РНК-транскриптов изобретения. Пригодные примеры таких репликаз, которые пригодны в изобретении, включают так называемые репликазы вируса QВ, которые распознают определенные сайты последовательности нуклеиновых кислот с концов данного РНК-транскрипта, а также репликазы вируса мозаики костра (ВМV) и репликазы альфа-вируса, которые распознают сайты последовательности нуклеиновых кислот с 3'-конца данного РНК-транскрипта. Эти репликазы служат для репликации РНК-транскриптов и комплементов с тем, чтобы умножить их копии. Когда такой фермент присутствует в реакции в течение процесса транскрипции, можно предполагать, что множество транскриптов, продуцируемых во время транскрипции, могут сами подвергаться репликации с тем, чтобы эскпоненциально повышать количество продукта РНК-транскрипта.
Сущностью изобретения является способность совершать множество циклов амплификации in vitro без необходимости в термической циклизации или прибавлении дополнительных ферментов.
На чертеже изображена схема предпочтительного варианта изобретения. Принципиальным аспектом изобретения является включение фермента РНКазы Н. Стадии 1 и 2 на схеме идентичны тем, которые используются в так называемом протоколе ТАS, однако в стадии 3 вместо этапа термической денатурации гибридный дуплекс РНК/ДНК "нитераздуляют" путем селективного переваривания мишени РНК с использованием РНКазы Н. Активность РНКазы Н имеет специфичность для РНК только тогда, когда она присутствует в гибридном дуплексе РНК/ДНК. Продукты этого переваривания могут представлять собой или уникальные РНК-олигомеры, или множественные РНК-олигомеры (стадии 4), и в свою очередь эти олигомеры могут действовать как праймеры для ДНК-синтеза с использованием обратной транскриптазы (RT) в качестве катализатора этой кДНК-реакции (стадия 5). Двунитевая ДНК, показанная в стадии 5, может действовать в качестве темплата для Т7 РНК полимераза-направленной транскрипции (стадия 6). Этот амплифицированный РНК-продукт теперь служит в качестве молекул обнаружения для целевой последовательности и/или служит в качестве дополнительных целевых молекул для продолжения самоциклирующихся реакций (стадии 7 - 12).
Наиболее успешные реакции, приводящие к получению приблизительно 106-кратной целевой амплификации, функционируют с тремя ферментами и двумя олигонуклеотидными праймерами, содержащими последовательность связывания РНК-полимеразы Т7 (РВS). Необходимыми ферментами являются обратная транскриптаза АМV, РНК-полимераза Т7 и РНКаза Н E. соli. Прибавление РНКазы Н Е. соli в реакцию пополняет активность РНKазы в обратной трансксиптазе АМV и является необходимым для получения высоких уровней амплификации. Отбор оптимальных олигонуклеотидных праймеров концентрируется на областях длины последовательности, включения полимераза-связующей последовательности на одном или обоих праймерах и эффективности праймеров, содержащих полимераза-связующие последовательности в качестве транскрипционного промотора. Все три области влияют на уровень амплификации. Включение двух олигонуклеотидных праймеров, каждый из которых содержит РВS, приводит к большему уровню амплификации нежели включение единственного РВS-содержащего праймера и не-РВS-содержащего праймера.
Целевую молекулу мРНК отжигают целевым специфическим олигодезоксирибонуклеотидом, вводя последовательность связывания промотора РНК-полимеразы Т7.
Данный праймер удлиняют ДНК-полимеразной активностью обратной транскриптазы АМV, синтезируя первую кДНК-нить.
Активность РНКазы Н обратной транскриптазы АМV и РНК-азы Н Е. соli разлагает РНК гибридного дуплекса РНК/ДНК, делая ДНК пригодной в качестве теплата для синтеза кДНК второй нити.
Самогенерирующиеся олигорибонуклеотиды, полученные в результате переваривания РНКазы Н, или синтетический олигодезоксирибонуклеотид, предпочтительно инкорпорирующий последовательность связывания промотора РНК-полимеразы Т7 (не показана), примируют синтез кДНК вторых нитей. Обратная транскриптаза АМV затем удлиняет праймер с образованием двунитевой ДНК, которая инкорпорирует оперативную последовательность связывания промотора РНК-полимеразы Т7.
РНК-полимераза Т7 связывается с последовательностью связывания двунитевого промотора и транскрибирует копии РНК, комплементарные целевой нуклеиновой кислоте.
Второй олигодезоксирибонуклеотидный праймер с РВS прижигают к РНК-транскрипту.
Обратная транскриптаза АМV катализирует синтез нити кДНК.
РНКаза Н разлагает РНК гибридного дуплекса РНК/ДНК и делает ДНК пригодной в качестве темплата для двунитевого синтеза.
Олигодезоксирибонуклеотидный праймер гибридизирует к двунитевой кДНК, и обработанная транскриптаза АМV синтезирует ДНК. Происходит транскрипция, и циклизация продолжается.
Пример 1. Амплификация первой области env ВИЧ-1.
Область РНК ВИЧ-1 амплифицирует в изотермической ферментативной реакции, которая генерирует в 106 больше копий данной области к концу реакции.
а) Реакция амплификации.
Исходным нуклеиновокислотным веществом является цезийосажденная РНК, которую экстрагируют из ВИЧ-1-зараженных клеток СЕМ (линия клеток человеческой лимфобластомы) (Folks et al., Proc. Natl.Sсi: USA 4531, 1985) методикой градиента в гуанидий-изоцианате цезий-хлориде (Маниатис и др.). Ожидают, что ВИЧ-1 специфические последовательности составляют 1 - 10% от общего количества присутствующих нуклеиновых кислот. 1/10 аттомоль целевой нуклеиновой кислоты помещают в реакционную смесь (общий объем 100 мкл), содержащую (конечные концентрации) : 40 мМ Трис. HCl, pH 8,1 12 мМ MgCl2, 25 мМ NaCl, 2 мМ Спермидина- (HCl)3, 5 мМ Дитиот рейтола, 80 мкг/мл БСА, 1 мМ каждого издАТФ, дЦТФ, дГТФ, дТТФ, 1 мМ каждого из АТФ, ЦТФ, ГТФ, УТФ, 0,25 мкг каждого примирующего олигонуклеотида (88-211 и 88-347).
Реакционные компоненты объединяют в 1,5 мл пробирке Эппендорфа и затем быстро и слегка перемешивают. Целевую нуклеиновую кислоту денатурируют путем нагревания пробирки до 65oC в течение 1 мин в водяной бане. После охлаждения до 37oC в течение 1 мин добавляют следующие ферменты: 25 ед. обратной транскриптазы AMV (15 - 25 ед./мкл), 100 ед. РНК-полимеразы Т7 (100 ед./мкл), 4 ед. РНКазы H E.coli (2 ед./мкл). Реакцию продолжают в течение 3 ч при 37oC без дальнейших манипуляций.
b) Обнаружение продуктов амплификации.
Приблизительно через 1 ч продукты обнаруживают с использованием 32P-меченного олигонуклеотида, комплементарного области в 30 пар оснований в пределах амплифицированного фермента (88-298). Аликвоту реакционной смеси, образующую объем 1/40 от общего объема, денатурируют в 95,0 мкл 7,4% формальдегида и 10 x SSC (Маниатис и др.) при 55oC в ванне с водой в течение 20 мин. Сразу же аликвоту охлаждают на льду и затем наносят на нитроцеллюлозную мембрану с использованием устройства для слот-блоттинга. Нуклеиновые кислоты иммобилизируют к нитроцеллюлозе посредством ультрафиолетового облучения (254 м)
После прикрепления фильтры предварительно гибридизируют при 55oC в течение 15 мин в 50 - 100 мкл буфера/см2 фильтра, содержащего 0,5% БСА, 0,5% поливинилпирролидон, 5 x SSPE (20 = 3,6 M NaCl, 200 мМ NaH2PO4, 20 мМ ЭДТК, pH 7 - 8) и 1% додецилсульфат натрия (ДДС-Na). Гибридизацию осуществляют при 55oC в течение 2 ч в том же буфере, содержащем 2 - 5 x 106 циклов в минуту/мл фосфорилированного олигонуклеотидного зонда. Зонд прибавляют к предварительно гибридизационному буферу. Фильтры промывают три раза при комнатной температуре в течение 3 мин, используя каждый раз 1 мл буфера/см2 фильтра: 1 x SSPE, 1% ДДС-Na, затем в течение 1 мин в том же буфере при 55oC. Фильтры подвергают авторадиографии при -70oC с одним усиливающим экраном.
Уровни амплификации оценивают путем сравнения интенсивности сигнала, продуцируемого амплифицированным продуктом, и сигнала, продуцируемого известными количествами РНК ВИЧ-1 или pAPV7A/2 (Luciw et al., Nature 312, 760, 1984), плазмиды, содержащей кДНК-копию генома ВИЧ-1, инсенцированного в сайт EcoRI плазмиды pVC19.
В примере 1 уровень амплификации составляет 1 x 106. Назерн-блот и Саузерен-блот-анализ с использованием обнаружительных зондов 88-297 и 88-298 показывает, что он представляет собой смесь ДНК и РНК, причем преобладающим видом является РНК. Продукт имеет дискретный размер (приблизительно 210 пар оснований) в пределах узкого интервала (20 - 40 пар оснований).
Пример 2. Амплификация второй области envВИЧ-1.
Амплификацию второй области envВИЧ-1 осуществляют в соответствии с методикой, описанной в примере 1, за исключением того, что для амплификации используют примирующие олигонуклеотиды 87-284 и 88-347, а для обнаружения используют обнаружительные олигонуклеотиды 86-272 и 86-273. Достигают 103-кратную амплификацию.
Пример 3. Амплификация области sor ВИЧ-1.
Амплификацию области sor ВИЧ-1 осуществляют в соответствии с методикой, описанной в примере 1, за исключением того, что для амплификации используют примирующие олигонуклеотиды 88-77 и 87-292, а для обнаружения используют обнаружительные олигонуклеотиды 86-31 и 86-32. Достигают 103-кратную амплификацию.
Пример 4. Амплификация первой области envВИЧ-1 из клинических проб крови больных СПИДом.
Амплифицируют РНК из трех ВИЧ-1-инфицированных клинических проб. РНК экстрагируют протоколом органической экстракции.
Амплификацию проводят как в примере 1, используя примирующие олигонуклеотиды 88-211 и 88-347, а также обнаружительные олигонуклеотиды 88-297 и 88-298.
Две пробы дают положительные результаты. Общая амплификация для реакций в этих экспериментах составляет 105-кратную. Поскольку сигнал, обнаруженный после амплификации, прямо пропорционален количеству исходного вещества, присутствующего в начале реакции (см. пример 5), возможно, что третий клинический образец не идентифицирован в качестве ВИЧ-1-инфецированного, так как он содержит очень маленькую исходную целевую последовательность ВИЧ-1.
Пример 5. Амплификация первой области envВИЧ-1 в незаряженных клетках СЕМ, смешанных с различными количествами ВИЧ-1 зараженных клеток СЕМ.
Амплификацию осуществляют как в примере 4, используя примирующие олигонуклеотиды 88-211 и 88-347, а также обнаружительные олигонуклеотиды 88-297 и 88-298. Целевая амплификация с использованием всех нуклеиновых кислот, экстрагированных (см. пример 7) из 103 - 1,0 клеток СЕМ, зараженных ВИЧ-1, в популяции 106 незараженных клеток СЕМ показывает сигнал, пропорциональный количеству зараженных клеток, присутствующих в пробе. Негативный контроль, т.е. 106 незараженных клеток СЕМ, проявляет малый фон. Данный фоновый сигнал значительно менее интенсивен, чем сигнал, полученный из пробы 10 зараженных клеток СЕМ.
Пример 6. Реагенты и олигонуклеотиды.
а) Реагенты: Нуклеозидтрифосфаты получают из Сигма, транскриптазу AMV(обратную) получают из Лайф Сайенсез, РНК-полимеразу Т7 получают из Стратагена, рибонуклеазу H E. coli и БСА получают из Бефесда Рисерч Лэбз.
b) Олигонуклеотиды синтезируют фосфорамидитной химией с использованием Эпплайд Байосистемз 380А, затем очищают ВЭЖХ. Олигонуклеотиды, используемые в качестве праймеров и зондов, специфичны для ВИЧ-1 и соответствуют последовательностям (Ратнер и др, Нейчур 313, 277, 1985) для областей env и sor:
88-211: (примирующий олигонуклеотид, nt 6450-6479, env)
5' AATTTAATACGACTCACTATAGGGATCTATTGTGCCCCGGCTGTTTTGCGATTCTA - 3'
88-297: (обнаружительный олигонуклеотид, nt 6560-6581, env)
5' - TGGCCTAATTCCATGTGTACATTGTACTGT - 3'
88-298: (онаружительный олигонуклеотид, nt 6581-6560, env)
5' - ACAGTACAATGTACACATGGAATTAGGCCA - 3'
88-347: (примирующий олигонуклеотид, nf 6661-6632, env)
5' AATTTAATACGACTCACTATAGGGATGTACTATTATGGTTTTAGCATTGTCTGTGA - 3'
88-77: (nt 5018-4988, env) примирующий
5' - AATTTAATACGACTCACTATAGGGACACCTAGGGCTAACTATGTGTCCTAATAAGG - 3'
87-292: (nt 4766-4796, env) примирующий
5' - TAATACGACTCACTATAGGGAAAGAATAAGTTCAGAAGTACACATCCCACT - 3'
86-31: (nt 4901-4932, env) обнаружительный
5' - GCACAEAAGTAGACCCTGAACTAGCAGACCA - 3'
86-32: (nt 4932-4901, env) обнаружительный
5' - TGGTTCTGCTAGTTCAGGGTCTACTTGTGTGC - 3'
87-284: (nt 6551-6579, env) примирующий
5' - TAATACGACTCACTATAGGGAAATTAGGCCAGTAGTATCAACTCAACT - 3'
86-272: (nt 6591-6620, env) обнаружительный
5' - TCTAATTACTACCTCTTCTTCTGCTAGACT - 3'
86-273: (nt 6620-6591, env) обнаружительный
5' - AGTCTAGCAGAAGAGGTAGTAATTAGA - 3'
Каждый примирующий олигонуклеотид содержит последовательность у 5'-конца, которая является последовательностью связывания РНК-полимеразы Т7 и предпочтительным сайтом транскрипционной инициации. Остальная часть последовательности целевой последовательности.
В отношении целевой последовательности ВИЧ-1 олигонуклеотиды 88-211, 88-298, 87-292, 86-31, 87-284 и 86-273 комплементарны негативной нити, а олигонуклеотиды 88-347, 88-297, 88-77, 86-32, и 86-272 комплементарны позитивной нити.
Пример 7. Органическая экстракция РНК.
Протокол для получения нуклеотидных кислот на пробах зараженных клеток.
Осаждают клетки: 5 к-об/мин в течение 10 мин из 1 мл Трис-забуференного солевого раствора. Вытягивают и сливают супернантат, оставляя 50 мкл с осадком. Ресуспендируют осадок после центрифугирования в 600 мкл Лизисбуфера. Перемешивают энергично и инкубируют при 50oC в течение 45 мин, перемешивая в течение 10 - 15 с каждые 10 мин. Прибавляют 600 мкл фенол : хлороформ : изоамиловый спирт 50 : 48 : 2. Встряхивают и перемешивают с тем, чтобы эмульгировать смесь. Центрифугируют при 14 к-об/мин в течение 2 мин для разделения фаз. Вытягивают 575 мкл из водной (верхней) фазы. Прибавляют 600 мкл фенол : хлороформ : изоамиловый спирт 50 : 48 : 2. Встряхивают и перемешивают с тем, чтобы эмульгировать смесь. Центрифугируют при 14 к-об/мин в течение 2 мин для разделения фаз. Вытягивают 525 мкл из водной фазы. Прибавляют 600 мкл хлороформ : изоамиловый спирт 24 : 1. Встряхивают и перемешивают для эмульгирования смеси. Центрифугируют при 14 к-об/мин в течение 2 мин для разделения фаз. Вытягивают 400 мкл из водной фазы. (Не следует переносить никакой клеточный дебрис, который может быть на поверхности раздела). Прибавляют 1/10 объем (40 мкл) 8М Cl. На данной стадии пробы могут быть расщеплены для процессинга. Прибавляют 3 объема 100% этанола, охлажденного на льду, к пробам, которые были расщеплены. Прибавляют 2,5 объема 100% этанола, охлажденного на льду, к пробам, которые не были расщеплены. Хорошо перемешивают, преципитируют при - 20oC в течение ночи или в ванне сухой лед/этанол в течение 15 мин.
Реагенты:
Лизис-буфер : 20 мМ Трис, pH 7,5 150 NaCl, 10 мМ ЭДТК, 0,2% ДДС-Na, 200 мкг/мл протеиназы К;
трис-забуференный солевой раствор: 137 мМ NaCl, 5,1 мМ KCl, 24,8 мМ Трис- основание, pH доводят до 7,4 путем прибавления 1 н. раствора HCl, стерилизуют автоклавированием.
Вышеприведенное описание касается специфических методов, которые можно использовать для осуществления изобретения. Имея в наличии подробно описанные специфические методы, используемые первоначально для описания системы изометрической амплификации, специалист в данной области техники будет хорошо знать, как создать альтернативные эквивалентные методы в результате аналогичной амплификации в изометрической однореакторной установке и с целью распространения данной информации на целевые нуклеиновые кислотные, нежели те, которые раскрываются специфически, не выходя за пределы объема описания и формулы изобретения.
Изобретение относится к молекулярной биологии и технологии рекомбинантных ДНК. Сущность изобретения: проводят взаимодействие сегмента-мишени РКН с реакционной смесью, содержащей первый и второй праймеры и ферменты, обладающие активностью к РНК - и ДНК полимеразе с образованием матрицы, содержащей функциональный двунитевой промотор и последовательность сегмента-мишени РНК с последующим образованием РНК путем транскрипции с матрицей. 5 з. п. ф-лы, 1 ил.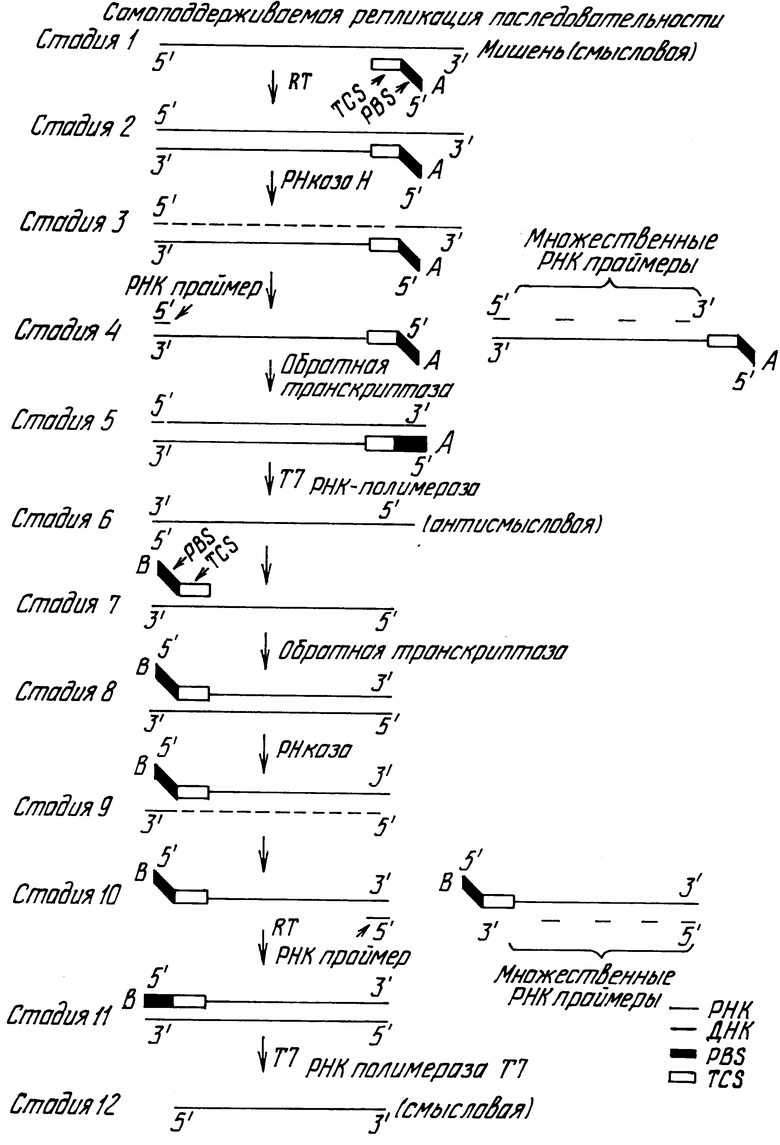
| РСТ, заявка, 88/10315, кл | |||
| Способ гальванического снятия позолоты с серебряных изделий без заметного изменения их формы | 1923 |
|
SU12A1 |
