Настоящая заявка частично является продолжением американской заявки с серийным номером 487767, поданной 2 марта 1990 г., содержимое которой естественно вошло со ссылкой в настоящее описание.
На протяжении этой заявки в круглых скобках приводятся ссылки на различные публикации. Данные этих публикаций во всей полноте вошли тем самым со ссылкой в настоящую заявку, чем достигается большая полнота описания состояния техники, к которой относится это изобретение.
Изобретение касается клонирования и продуцирования человеческих аналогов фактора фон Виллебранда и способов использования таких аналогов.
Структурные признаки фактора фон Виллебранда.
Фактор фон Виллебранда (vWF) представляет собой большой белок плазмы, который синтезируется в эндотелиальных клетках, которые образуют внутреннюю поверхностную выстелку стенки кровеносных сосудов и через мегакариоциты предшественника тромбоцитов. Большие количества vWF-фактора находят в тромбоцитных α -гранулах, содержимое которых высвобождается в кровь при тромбоцитной активации. Вновь синтезированный в эндотелиальных клетках vWF-фактор может поступать в кровь двумя возможными способами. Часть его конститутивно секретируется в кровь в основном в виде связанных через дисульфидную группу димеров или небольших мультимеров массой в 250000 дальтоновых субъединиц. Или же честь его поступает в секреторные органеллы, названные телами Вейбеля-Палада (Weibel-Palade). Фактор vWF, находящийся в телах Вейбеля-Палада, является сильно мультимерным, изменяясь по размеру от димера до мультимеров в 50 или более субъединиц, и может быть высвобожден из клеток при воздействии секретатогов, таких как тромбин. Сильно мультимерный vWF-фактор является наиболее эффективным в отношении стимуляции слипания тромбоцитов.
Был выделен ген, кодирующий vWF-фактор, и было показано, что его длина превышает 150000 оснований. Он включает в себя более 20 эксонов. Информационная РНК (мРНК) vWF-фактора по длине примерно составляет 9000 оснований и кодирует пре-про vWF-фактор, состоящий из 2813 аминокислот. Остатки 1 - 22 образуют обработанную главную последовательность, которая предположительно расщепляется при вхождении белка в гранулярный эндоплазматический ретикулюм. N-Концевая часть про-vWF-фактора (741 аминокислота) представляет собой про-пептид, который не присутствует в зрелом vWF-факторе. Этот пептид присутствует в крови, и было показано, что он является идентичным белку крови, известному ранее как антиген 11 фон Виллебранда (vWF Agll). Про-пептид является существенным для протекания мультимеризации vWF-фактора. Клетки, в которые введена кДНК фактора vWF-, содержащие только зрелые vWF-последовательности, дают только димеры. Ничего не известно о функциональной роли образования про-пептид/vW Agll.
Из анализа ДНК-последовательности следует, что про-vWF-предшественник состоит из интервально повторяющихся субъединиц. Были идентифицированы четыре разные области. Зрелый vWF-фактор состоит из трех областей типа A, трех областей типа B и двух областей типа C. Имеются также две полные и одна частичная области типа D. Про-пептид состоит из двух областей типа D, из чего делается предположение, что он может обладать связанными с ними функциями.
Зрелый vWF-фактор представляет собой многовалентную молекулу, которая характеризуется наличием связующих сайтов, пригодных для нескольких белков. Один из связующих сайтов распознает тромбоцитный гликопротеид Ib/GPI/. Использованием протеолитических веществ было установлено, что у зрелого vWF-фактора этот сайт приходится на область, расположенную между аминокислотными остатками 449 и 728. Кроме того, vWF-фактор характеризуется наличием по крайней мере двух коллагеновых связующих сайтов, по крайней мере двух гепариновых связующих сайтов, сайта, связывающего фактор VIII, и RGD-сайта, посредством которого происходит присоединение к тромбоцитному GP IIb /IIIa-рецептору.
Участие vWF-фактора в тромбоцитном сцеплении с субъэндотелием.
Вывод о том, что vWF-фактор, в частности связывание vWF-фактора с тромбоцитным GPIB-рецептором, играет существенную роль в обычной тромбоцитной адгезии, основывается как на клинических наблюдениях, так и на данных исследований, проведенных in vitro. Пациенты с нарушением кровотечения в виде заболевания фон Виллебранда (болезнь Виллебранда-Юргенса/vWD) обладают пониженными уровнями фактора vWF или характеризуются полным отсутствием vWF-фактора. Или же они могут обладать дефектным vWF-фактором. Еще одно заболевание - синдром Бернарда-Соулира (Bernard-Soulier Syndrome) BSS//-характеризуется наличием тромбоцитов с отсутствующими GPIB-рецепторами.
В исследованиях in vitro система, которая наиболее близко аппроксимирует условия, существующие в поврежденном кровеносном сосуде, состоит из перфузионной камеры, в которой вывернутый участок кровеносного сосуда (кроличья аорта, человеческая посмертная почечная артерия или человеческая умбиликальная артерия) подвергается воздействию протекающей крови. После отделения слоя эндотелиальных клеток от сосуда крови позволяют протекать через камеру. Степень протекания тромбоцитной адгезии оценивают непосредственно методом морфометрии или косвенно, используя тромбоциты с радиоактивной меткой. Кровь, взятая у пациентов, страдающих болезнью Виллебранда-Юргенса или синдромом Бернарда-Соулира, не способствует тромбоцитной адгезии в этой системе, тогда как нормальная кровь способствует, указывая на необходимость наличия vWF-фактора или тромбоцитного гликопротеида Ib (GPIb). Более того, добавление моноклональных антител к GPIb также препятствует протеканию тромбоцитной адгезии. Зависимость тромбоцитной адгезии от vWF-фактора является более выраженной в условиях существования высоких касательных скоростей, типа существующих в артериальном потоке. В условиях существования низких касательных скоростей тромбоцитная адгезия может быть усилена воздействием других адгезионных белков, таких как фибронектин. Адгезионные силы, возникающие под воздействием этих других белков, не являются, видимо, достаточными для обеспечения адгезии при воздействии больших касательных сил; и становится очевидной зависимость от vWF-фактора. Кроме того, мультимерная природа vWF-фактора может оказаться способствующей обеспечению более прочной связи за счет связывания большего числа сайтов, имеющихся на тробмоците.
Примерно 20% пациентов, у которых был удален сгусток проведением пластической операции на сосудах или введением тканевого плазминогенного активатора (tPA), страдают от реокклюзии. Сказанное связано, видимо, с повреждением эндотелия, возникающим при проведении лечения, результатом чего является адгезия тромбоцитов на подвергнутой воздействию области внутренней поверхности сосуда. Сказанное сопровождается агрегацией многих слоев тромбоцитов и фибрина на ранее прилипших тромбоцитах, образующих тромб.
До настоящего времени отсутствуют указания на то, что какое-либо из препятствующих агрегации тромбоцитов веществ, описанных в литературе, могло бы противодействовать адгезии тромбоцитов на подвергнутом воздействию субъэндотелии, что тем самым исключало бы последующее образование сгустка.
Предметом изобретения являются негликозилированные биологически активные полипептиды, которые включают в себя связующую область GPIb фактора vWF (фактора фон Виллебранда). Эти полипептиды могут быть, между прочим, использованы для устранения адгезии и агрегации тромбоцитов при лечении субъектов с нарушениями, такими как цереброваскулярные расстройства и сердечно-сосудистые расстройства. Это изобретение также касается плазмид экспрессии, кодирующих эти полипептиды, и к тому же способов продуцирования посредством трансформации бактериальной клетки и выделения таких полипептидов. Кроме того, предметом изобретения являются способы лечения и предотвращения цереброваскулярных, сердечно-сосудистых и других расстройств посредством использования этих полипептидов для ингибирования агрегации тромбоцитов.
Изобретение касается негликозилированного биологически активного полипептида, обладающего следующей аминокислотной последовательностью:
X-A-[Cys Ser Arg Leu Leu Asp Leu Val Phe Leu Leu Asp Gly
Ser Ser Arg Leu Ser Glu Ala Glu Phe Glu Val Leu Lys Ala
Phe Val Val Asp Met Met Glu Arg Leu Arg Ile Ser Gln Lys
Trp Val Arg Val Ala Val Val Glu Tyr His Asp Gly Ser His
Ala Tyr Ile Gly Leu Lys Asp Arg Lys Arg Pro Ser Glu Leu
Arg Arg Ile Ala Ser Gln Val Lys Tyr Ala Gly Ser Gln Val
Ala Ser Thr Ser Glu Val Leu Lys Tyr Thr Leu Phe Gln Ile
Phe Ser Lys Ile Asp Arg Pro Glu Ala Ser Arg Ile Ala Leu
Leu Leu Met Ala Ser Gln Glu Pro Gln Arg Met Ser Arg Asn
Phe Val Arg Tyr Val Gln Gly Leu Lys Lys Lys Lys Val Ile
Val Ile Pro Val Gly Ile Gly Pro His Ala Asn Leu Lys Gln
Ile Arg Leu Ile Glu Lys Gln Ala Pro Glu Asn Lys Ala Phe
Val Leu Ser Ser Val Asp Glu Leu Glu Gln Gln Arg Asp Glu
Ile Val Ser Tyr Leu Cys]-B-COOH
где
X - группа NH2-метионин- или NH2-;
A - последовательность, состоящая из по крайней мере из одной аминокислоты, но при этом содержащая менее 35 аминокислот, которая присутствует в vWF-факторе естественного происхождения и у которой карбоксиконцевая аминокислота представляет собой тирозин с N 508;
B - последовательность, состоящая по крайней мере из одной аминокислоты, но при этом содержащая менее 211 аминокислот, которая присутствует в vWF-факторе естественного происхождения и у которой концевая аминогруппа аминокислоты представляет собой аспарагиновую кислоту с N 696, и два цистеина, содержащиеся в последовательности, заключенной в скобки, соединены через дисульфидную связь.
Предметом изобретения является способ продуцирования какого-либо из описанных выше полипептидов, который включает в себя трансформацию бактериальной клетки с получением плазмиды экспрессии, кодирующей полипептид, выращивание результирующей бактериальной клетки с продуцированием полипептида, закодированного плазмидой, и выделение полипептида, продуцированного указанным способом.
Предметом изобретения является фармацевтический состав, включающий в себя некоторое количество какого-либо из описанных выше полипептидов, являющихся эффективными в отношении торможения процесса агрегации тромбоцитов, и фармацевтически приемлемый носитель. Предметом изобретения является также способ торможения процесса агрегации тромбоцитов, который включает в себя контактирование тромбоцитов с некоторым количеством какого-либо из описанных выше полипептидов, являющихся эффективными в отношении торможения процесса агрегации тромбоцитов.
Предметом изобретения являются способы лечения, устраняющие или подавляющие расстройства, такие как цереброваскулярные или сердечно-сосудистые расстройства или тромбоз, включающие в себя введение субъекту некоторого количества какого-либо из описанных выше полипептидов, эффективных в плане лечения или устранения таких расстройств.
Предметом изобретения является также способ выделения очищенного биологически активного описанного выше полипептида, который включает в себя:
а) продуцирование в бактериальной клетке первого полипептида, обладающего аминокислотной последовательностью полипептида, но не содержащего дисульфидную связь;
б) разрушение бактериальной клетки с продуцированием лизата, содержащего первый полипептид;
в) обработку лизата до получения телец включений, содержащих первый полипептид;
г) контактирование телец включений, полученных на стадии в), с получением первого полипептида в растворимой форме;
д) обработку результирующего первого полипептида с образованием биологически активного полипептида;
е) извлечение образовавшегося биологически активного полипептида;
ж) очистку извлеченного биологически активного полипептида.
На фиг. 1 показана конструкция плазмиды pvWIP. Отделяли ряд кДНК-клонов фактора vWF в λ gtII (выделенных из кДНК-совокупности эндотелиальной человеческой клетки). Один кДНК-клон, охватывающий всю связующую область GPIb, субклонировали в EcoRI-сайт плазмиды (фиг. 9). Результирующая плазмида pvWIP содержит кДНК-вставку из 2500 оснований.
На фиг. 2 показана конструкция плазмиды pvWF-VAI. Синтетический олигомер, содержащий ATG-инициирующий кодон, расположенный перед аминокислотой gIu-437 (т. е. перед 437-й аминокислотой в белке фактора vWF) (фиг. 12-19), присоединяли к плазмиде pvWIP, подвергнутой расщеплению при воздействии NdeI и Bsu 361. Результирующая плазмида была обозначена как pvWF-VAI и внесена в штамм S ⊘ 930 культуры E. coli под инвентарным номером 68530 (Американская коллекция типовых культур микроорганизмов).
На фиг. 3 показана конструкция плазмиды pvWF-VBI. Синтетический олигомер, содержащий ATG-инициирующий кодон, расположенный перед аминокислотой phe-443 (фиг. 12 - 19), присоединяли к плазмиде pvWIp, подвергнутой расщеплению при воздействии NdeI и Bsu361. Результирующая плазмида была обозначена как pvWF-VBI.
На фиг. 4 показана конструкция плазмиды pvWF-VA2. Синтетический олигомер, содержащий TAA-завершающий кодон, расположенный после аминокислоты Iys-728 (фиг. 12 - 19), присоединяли к плазмиде pvWF-VA1, подвергнутой расщеплению при воздействии Hind 111 и Xmal. Результирующая плазмида была обозначена как pvWF-VA2.
На фиг. 5 показана конструкция плазмиды pvWF-VB2. Синтетический олигомер, содержащий TAA-завершающий кодон, был присоединен к плазмиде pvWF-VB1, подвергнутой расщеплению при воздействии Hind 111 и Xmal. Результирующая плазмида была обозначена как pvWF-VB2.
На фиг. 6 показана конструкция плазмиды pvWF-VA3. Из плазмиды pvWF-VA2 выделяли фрагмент NdeI-EcoRV и присоединяли к плазмиде pMF-945, подвергнутой расщеплению при воздействии NdeI и Rvu 11. Полученную плазмиду обозначали как pvWF-VA3. Плазмида экспрессировала образование VA, полипептид связующей области GPIb фактора vWF, который включает в себя аминокислоты от 437 до 728 (фиг. 12 - 19), что находится под управлением промотора deo P1P2.
На фиг. 7 показана конструкция плазмиды pvWF-VB3. Из плазмиды pvWF-VB2 выделяли фрагмент NdeI-EcoRV и присоединяли к плазмиде pWF-945, подвергнутой расщеплению при воздействии NdeI и Pvu 11. Полученную плазмиду обозначали как pvWF-VB3. Плазмида экспрессировала VB, полипептид связующей области GPIb фактора vWF, который включает в себя аминокислоты от 443 до 728, что находится под управлением промотора deo P1P2.
На фиг. 8 показана конструкция плазмиды pvWF-VC3. Синтетическую сцепку присоединяли к плазмиде pvWF-VA3, подвергнутой расщеплению при воздействии NdeI и TthIIII. Полученную плазмиду обозначали как pvWF-VC3, и она была внесена в Американскую коллекцию типовых культур микроорганизмов под инвентарным номером 68241 (Американская коллекция типовых культур микроорганизмов). Плазмида экспрессировала VC (это образование фигурирует также как VCL или VC3), полипептид связующей области GPIb фактора vWF, который включает в себя аминокислоты от 504 до 728 (фиг. 12 - 19), что находится под управлением промотора deo P1P2.
На фиг. 9 показана конструкция плазмиды pvWF-VD3. Синтетическую сцепку присоединяли к плазмиде pvWF-VA3, подвергнутой расщеплению при воздействии NdeI и TthIIII. Полученную плазмиду обозначали как pvWF-VD3. Плазмида экспрессировала образование VD, полипептид связующей области GPIb фактора vWF, который включает в себя аминокислоты от 513 до 728 (фиг. 12 - 19), что находится под управлением промотора deo P1P2.
На фиг. 10 показано относительное положение плазмид, экспрессирующих полипептиды связующей области vWF-GPIb. Кроме того, показанные верхние две линии характеризуют кДНК фактора vWF и положение связующей области GPIb, кодирующей область в пределах кДНК.
На фиг. 11 показана конструкция плазмиды pMF-945. Плазмиду pEFF-920 (у штамма S ⊘ 930 культуры Escherichia coli, инвентарный номер 67706 у Американской коллекции типовых культур микроорганизмов) подвергали расщеплению при воздействии BgIII и NdeI и выделяли большой фрагмент. Этот фрагмент присоединяли к небольшому фрагменту из 540 пар оснований, полученному расщеплением плазмиды pMF-5534 (инвентарный номер 67703 у Американской коллекции типовых культур микроорганизмов) при воздействии BgIII и NdeI. В результате этого продуцировали плазмиду pMF-945, которая содержала PAR-последовательность и в порядке расположения 5' и 3' промоторные последовательности deo P1P2, модифицированный deo-рибосомальный связующий сайт с усилительной последовательностью, аналоговую кодирующую последовательность pGH и транскрипционные концевые последовательности T1T2. Плазмида pMF-945 представляет собой образование, которое на высоком уровне экспрессирует аналоговый белок pGH.
На фиг. 12 - 19 показана транслированная кДНК-последовательность зрелого человеческого фактора фон Виллебранда. Эта последовательность была составлена с использованием данных, которые привели Веруидж и др. (Verweij, C. L., et al., EMBO journal 5: 1839-1847 / 1986// и Садлер и др. /Sadler, J. E., et al. , Proc. NatI. Acad. Sci. 82: 6394 - 6398 /1985/). Эта нуклеотидная последовательность начинается с нуклеотида с номером 2519 (где нуклеотид 1 приходится на начало кодирующей последовательности для сигнального пептида) и завершается нуклеотидом 8668 при общем числе нуклеотидов 6150, кодирующих зрелый vWF-фактор, состоящий из 2050 аминокислот. Транслированная аминокислотная последовательность начинается с аминокислоты с номером 1 и завершается аминокислотными обозначениями, используемыми во всей этой заявке.
На фиг. 20 показана транслированная последовательность у полипептида связующей области GPIb-фактора фон Виллебранда, выраженного плазмидами pvWF-VC3 (инвентарный номер 68241 в Американской коллекции типовых культур микроорганизмов) и pvWF-VCl (инвентарный номер 68242 в Американской коллекции типовых культур микроорганизмов).
Первый кодон ATG, кодирующий трансляционный метионин кодона инициирования, был добавлен к нуклеотидной последовательности, соответствующей нуклеотидам от 4028 до 4702 последовательности, показанной на фиг. 12-19. Эта последовательность кодирует полипептид, содержащий 225 аминокислот (плюс метионин инициирования), которые соответствуют аминокислотам, начиная от аминокислоты Leu/лейцин/ с номером 504 и кончая аминокислотой Lys/лизин/ с номером 728 (фиг. 12-19), т.е. всего оказывается 226 аминокислот.
На фиг. 21 показана конструкция плазмиды pvWF-VCL. Плазмида pvWF-VC3 была подвергнута расщеплению при воздействии Hind 111 и Sty 1 и был выделен фрагмент с числом пар оснований 860. Этот фрагмент был соединен с большим фрагментом, выделенным из продукта расщепления плазмиды pMLK-100 при воздействии Hind 111 и Sty 1. Результирующая плазмида была обозначена как pvWF-VCL и внесена в штамм 4300 /F-/ культуры E. coli Американской коллекции типовых культур микроорганизмов под инвентарным номером 68242 (Американская коллекция типовых культур микроорганизмов). Эта плазмида экспрессирует образование VCL, отвечающее тому же полипептиду связующей области GPIb фактора vWF, что и в случае плазмиды pvWF-VC3 (метионин плюс аминокислоты 504-728); однако происходит это под управлением λ Pl - промотора и deo-рибосомального связующего сайта.
На фиг. 22 показана конструкция плазмиды pvWF-VE2. Плазмида pvWF-VA2 была подвергнута расщеплению при использовании Nde1 и Pst1 и был выделен большой фрагмент. Синтетические олигомеры N 2 и N 3 (фиг. 23) были обработаны полинуклеотидной киназой T4. Большой фрагмент плазмиды pvWF-VA2 был затем соединен с синтетическими олигомерами N 1 и N 4 (фиг. 23) и с подвергнутыми воздействию киназы олигомерами N 2 и N 3. Результирующая плазмида была обозначена как pvWF-VE2.
На фиг. 23 показаны четыре синтетические связки (с номерами 1-4), использованные при конструкции плазмиды pvWF-VE2.
На фиг. 24 показана конструкция плазмиды pvWF-VE3. Плазмида pvWF-VE2 была подвергнута расщеплению при использовании Ndel и HindIII; был выделен небольшой фрагмент с числом пар оснований 770, и он был присоединен к большому фрагменту, выделенному из продукта расщепления плазмиды pMLK-7891 при воздействии Ndel и HindIII. Результирующая плазмида была обозначена как pvWF-VE3.
На фиг. 25 показана конструкция плазмиды pvWF-VEL. Плазмида pvWF-VE3 была подвергнута расщеплению при использовании XmnI, обработана бактериальной щелочной фосфатазой (BAP) и затем подвергнута расщеплению при воздействии Ndel и HindIII. Плазмида pMLK-100 была подвергнута расщеплению при использовании NDeI и HindIII и обработана бактериальной щелочной фосфатазой. Два продукта расщепления были смешаны и сшиты с образованием плазмиды pvWF-VEL, которая экспрессировала ДНК-последовательность, соответствующую аминокислотам 469-728 vWF-фактора, что находилось под управлением λ PL-промотора и рибосомального связующего сайта cII.
На фиг. 26 показано влияние образования VCL на BJV-индуцированную агрегацию в человеческой плазме, богатой тромбоцитами. На этой фигуре показаны результаты, полученные при проведении стандартизованного теста по оценке vWF-зависимой агрегации (зависимой от фактора фон Виллебранда), который был проведен с использованием человеческой плазмы, богатой тромбоцитами.
На фиг. 27 показано влияние образования VCL на BJV-индуцированную агрегацию в крысиной плазме, богатой тромбоцитами. На этой фигуре показаны результаты, полученные при проведении стандартизованного теста по оценке vWF-зависимой агрегации (зависимой от фактора фон Виллебранда), который был проведен с использованием крысиной плазмы, богатой тромбоцитами.
Плазмиды pvWF-VC3, pvWF-VCL и pvWF-VAI были внесены в культуру Escherichia coli согласно и в соответствии с требованиями Будапештского договора по международному признанию вклада микроорганизмов для целей патентной практики, который был заключен с организацией в лице Американской коллекции типовых культур микроорганизмов (12301 Parklawn Drive, Rockville, Maryland 20852), под инвентарными номерами в Американской коллекции типовых культур микроорганизмов соответственно 68241, 68242 и 68530.
Это изобретение касается негликозилированного биологически активного полипептида со следующей аминокислотной последовательностью:
X-A-[Cys Ser Arg Ley Leu Asp Leu Val Phe Leu Leu Asp Gly
Ser Ser Arg Leu Ser Glu Ala Glu Phe Glu Val Leu Lys Ala
Phe Val Val Asp Met Met Glu Arg Leu Arg Ile Ser Gln Lys
Trp Val Arg Val Ala Val Val Glu Tyr His Asp Gly Ser His
Ala Tyr Ile Gly Leu Lys Asp Arg Lys Arg Pro Ser Glu Leu
Arg Arg Ile Ala Ser Gln Val Lys Tyr Ala Glu Ser Gln Val
Ala Ser Thr Ser Glu Val Leu Lys Tyr Thr Leu Phe Gln Ile
Phe Ser Lys Ile Asp Arg Pro Glu Ala Ser Arg Ile Ala Leu
Leu Leu Met Ala Ser Gln Glu Pro Gln Arg Met Ser Arg Asn
Phe Val Arg Tyr Val Gln Gly Leu Lys Lys Lys Lys Val Ile
Val Ile Pro Val Gly Ile Gly Pro His Ala Asn Leu Lys Gln
Ile Arg Leu Ile Glu Lys Gln Ala Pro Glu Asn Lys Ala Phe
Val Leu Ser Ser Val Asp Glu Leu Glu Gln Gln Arg Asp Glu
Ile Val Ser Tyr Ley Cys]-B-COOH
где
X - группа NH2-метионин- или NH2-;
A - последовательность, состоящая по крайней мере из одной аминокислоты, но при этом содержащая менее 35 аминокислот, которая присутствует в vWF-факторе естественного происхождения и у которой карбоксиконцевая аминокислота представляет собой тирозин с N 508, показанный на фиг. 12-19;
B - последовательность, состоящая по крайней мере из одной аминокислоты, но при этом содержащая менее 211 аминокислот, которая присутствует в vWF-факторе естественного происхождения и у которой концевая аминогруппа аминокислоты представляет собой аспарагиновую кислоту с N 696, показанную на фиг. 12-19, и два цистеина, содержащиеся в последовательности, заключенной в скобки, соединены через дисульфидную связь. Заключенная в скобки последовательность включает в себя аминокислоты с номерами 509-695, как это показано на фиг. 12-19.
В одном из вариантов этот полипептид обладает следующей аминокислотной последовательностью:
X-[Leu His Asp Phe Tyr Cys Ser Arg Leu Leu Asp Leu Val
Phe Leu Leu Asp Giy Ser Ser Arg Leu Ser Glu Ala Glu Phe
Glu Val Leu Lys Ala Phe Val Val Asp Met Met Glu Arg Leu
Arg Ile Ser Gln Lys Trp Val Arg Val Ala Val Val Glu Tyr
His Asp Gly Ser His Ala Tyr Ile Gly Leu Lys Asp Arg Lys
Arg Pro Ser Glu Leu Arg Arg Ile Ala Ser Gln Val Lys Tyr
Ala Gly Ser Gln Val Ala Ser Thr Ser Glu Val Leu Lys Tyr
Thr Leu Phe Gln Ile Phe Ser Lys Ile Asp Arg Pro Gly Ala
Ser Arg Ile Ala Leu Leu Leu Met Ala Ser Gln Glu Pro Gln
Arg Met Ser Arg Asn Phe Val Arg Tyr Val Gln Gly Leu Lys
Lys Lys Lys Val Ile Val Ile Pro Val Gly Ile Gly Pro His
Ala Asn Leu Lys Gln Ile Arg Leu Ile Glu Lys Gln Ala Pro
Glu Asn Lys Ala Phe Val Leu Ser Ser Val Asp Glu Leu Glu
Gln Gln Arg Asp Glu Ile Val Ser Tyr Leu Cys Asp Leu Ala
Pro Glu Ala Pro Pro Pro Thr Leu Pro Pro Asp Met Ala Gln
Val Thr Val Gly Pro Gly Leu Leu Gly Val Ser Thr Leu Gly
Pro Lys]-COOH
где X - группа NH2 - или NH2 - метионин-; желательно, чтобы это была группа NH2-метионин-.
В заключенную в скобки последовательность входят аминокислоты с номерами 504-728, показанные на фиг. 12-19.
Специалисты, работающие в этой области техники, к которым относится предмет изобретения, могут легко воссоздать такие полипептиды, используя методики с применением ракомбинантной или нерекомбинантной ДНК.
Полипептиды могут быть сконструированы по способу с использованием рекомбинантной ДНК. Сказанное означает, что для получения полипептидов необходимо экспрессировать нуклеиновую кислоту, кодирующую полипептиды, что делается в подходящем хозяине, таком как бактериальная, дрожжевая клетка или клетка млекопитающего, при использовании способов, хорошо известных в этой области техники, с выделением полипептида, экспрессированного в таком хозяине.
Примерами векторов, которые могут быть использованы для экспрессии нуклеиновой кислоты, кодирующей полипептиды, являются вирусы, такие как бектериофаги (такие как фаг лямбда), космиды, плазмиды и другие рекомбинантные векторы. Молекулы нуклеиновой кислоты вставляют в векторные геномы, используя способы, хорошо известные в этой области техники. Например, используя обычные выделенные ферментами сайты, вставку и векторную ДНК- и то и другое - подвергают воздействию фермента ограничения с целью образования комплементарных концов у обеих молекул, которые образуют пару оснований друг с другом и которые затем сшиваются при использовании лигазы. Или же сцепки могут быть сшиты с ДНК-вставкой, которая соответствует сайту ограничения у векторной ДНК; она затем подвергается расщеплению под воздействием фермента ограничения, который режет молекулу по такому сайту. Известны также и другие способы.
Векторы, составляющие кислоту, кодирующую полипептиды, могут быть адаптированы для экспрессии в бактериальной клетке, дрожжевой клетке или в клетке млекопитающего, которая дополнительно включает в себя регуляторные элементы, необходимые для обеспечения экспрессии нуклеиновой кислоты в бактериальной клетке, дрожжевой клетке или в клетке млекопитающего, причем они занимают такое положение относительно нуклеиновой кислоты, кодирующей полипептид, при котором обеспечивается экспрессия такового. Регуляторные элементы, требуемые для обеспечения экспрессии, включают в себя промоторные последовательности, необходимые для связывания РНК-полимеразы, и последовательности инициирования транскрипции, необходимые для обеспечения рибосомного связывания. Например, бактериальный экспрессионный вектор может включать в себя промотор, такой как λ PL-промотор или deo-промотор, и для обеспечения инициирования транскрипции содержать C11- или deo-рибосомальные связующие сайты. Такие векторы могут быть приобретены у других изготовителей или скомпонованы из последовательностей описанными способами, хорошо известными в этой области техники, например, способами, описанными выше применительно к конструированию векторов вообще.
Кроме того, для получения описанных выше полипептидов могут быть использованы нерекомбинантные способы, такие как способы с применением химического синтеза, синтетической ДНК или кДНК. Один из способов выделения полипептида сводится к прощупыванию человеческих геномных материалов, образующих библиотеку данных, при использовании природной или искусственно сконструированной ДНК-пробы, что проводится способами, хорошо известными в этой области техники. Молекулы ДНК и кДНК, которые кодируют полипептиды, могут быть использованы для получения комплементарной геномной ДНК, кДНК или РНК из источников человеческого, млекопитающего или иного животного происхождения или для выделения родственной кДНК или геномных клонов посредством проверки кДНК или библиотечных геномных материалов.
Предметом изобретения является далее фармацевтический состав, включающий в себя некоторое количество какого-либо из описанных выше полипептидов, эффективно противодействующих агрегации тромбоцитов, и фармацевтически приемлемый носитель.
Как это используется в заявке, термин "фармацевтически приемлемый носитель" распространяется на любые стандартные фармацевтические носители. Такие носители являются хорошо известными в этой области техники, и к ним могут быть отнесены, но, разумеется, не только они, любые стандартные фармацевтические носители, такие как солевые растворы с фосфатным буфером, вода, эмульсии, такие как водомасляная эмульсия, и различные типы смачивающих веществ. К другим носителям могут быть также отнесены стерильные растворы, таблетки, таблетки с покрытием и капсулы.
В типичном случае такие носители содержат наполнители, такие как крахмал, молоко, сахар, некоторые типы глины, желатин, стеариновую кислоту или соли таковых, стеараты магния или кальция, тальк, растительные жиры или масла, камеди, гликоли или другие известные наполнители. Такие носители могут также содержать душистые и окрашивающие добавки или другие ингредиенты. Составы с такими носителями образуют хорошо известными общепринятыми способами.
Состав берется в таком количестве, чтобы его было достаточно для образования концентрации в крови величиной от 0,06 до 58 мкМ, желательно в области примерно от 0,06 до 29 мкМ, например, в области от 0,23 до 23 мкМ. В ином выражении количество должно составлять от 0,1 до 100 мг на 1 кг массы тела, желательно от 0,1 до 50 мг на 1 кг массы тела, например, от 0,4 до 40 мг на 1 кг массы тела.
Введение состава может быть осуществлено любым из хорошо известных способов, включая, но не ограничиваясь ими, внутривенный, внутримышечный, подкожный и оральный способы.
Изобретение касается также способа торможения агрегации тромбоцитов, который включает в себя контактирование тромбоцитов с некоторым количеством какого-либо из описанных выше полипептидов, эффективных в отношении торможения агрегации тромбоцитов, чем обеспечивается торможение агрегации тромбоцитов.
Изобретение касается также экспрессии плазмид, кодирующих описанные выше полипептиды. В одном из вариантов реализации изобретения плазмида экспрессии, кодирующая полипептид в виде заключенной в скобки последовательности, т. е. с аминокислотами под номерами 504-728 на фиг. 12-19 обозначена как плазмида pvWF-VC3, и она внесена в Американскую коллекцию типовых культур микроорганизмов под инвентарным номером 68241. В еще одном варианте реализации изобретения плазмида экспрессии, кодирующая полипептид в виде заключенной в скобки последовательности, т.е. с аминокислотами под номерами 504-728 на фиг. 12-19, обозначена как плазмида pvWF-VCL, и она внесена в Американскую коллекцию типовых культур микроорганизмов под инвентарным номером 68242.
Плазмиды экспрессии, отвечающие изобретению, содержат далее регуляторные элементы, расположенные в пределах плазмиды и имеющие отношение к ДНК, кодирующей полипептид, чем обеспечивается экспрессия полипептида в надлежащей клетке хозяина, которые являются такими, как промотор и операторы, например deo P1P2 и λ PLOL, рибосомальные связующие сайты, например, сайты deo и C11, и репрессоры. К другим приемлемым регуляторным элементам относятся, например, 1ас-, trp-, tac- и 1pp-промоторы (Европейский патент, номер заявочной публикации 0303972, публикация произведена 22 февраля 1989 г.).
Подходящие регуляторные элементы располагаются в пределах плазмиды, имеющей отношение к ДНК, кодирующей полипептид, чем обеспечивается экспрессия полипептида в подходящей клетке хозяина. В предпочтительных вариантах реализации изобретения регуляторные элементы располагаются вблизи и у верхней части ДНК, кодирующей полипептид.
Плазмиды экспрессии, отвечающие изобретению, могут быть введены в подходящие клетки хозяина, желательно в бактериальные клетки хозяина. Предпочтительными бактериальными клетками хозяина являются клетки культуры Escherichia coli. Примерами подходящих клеток культуры Escherichia coli являются штаммы S ⊘ 930 или 4300, но для плазмид в качестве клеток хозяина могут быть также использованы и иные штаммы культуры Escherichia coli и другие бактерии. К таким бактериям относятся Pseudomonas aeruginosa и Bacillus subtilis.
Бактерии, используемые в качестве хозяина, могут представлять собой любой штамм, включая ауксотрофный (такой как А1645), прототрофный (такой как А4255) и литический штаммы, штаммы F+ и F-, штаммы с cl857 - респрессорной последовательностью λ - профага (такие как А1645 и А4255) и штаммы с дилецией в отношении deo - репрессоров и deo - гена (см. публикацию Европейской патентной заявки N 0303972, опубликованную 22 февраля 1989 г.). Штамм А4255 /F+/ культуры Escherichia coli был внесен в Американскую коллекцию типовых культур микроорганизмов под инвентарным номером 53468; и штамм А1 645 культуры Escherichia coli - под инвентарным номером 67829.
Изобретение касается бактериальной клетки, которая включает в себя эти плазмиды экспрессии. В случае одного из вариантов бактериальная клетка представляет собой клетку культуры Escherichia coli. В предпочтительных вариантах реализации изобретение касается клетки культуры Escherichia coli, содержащей плазмиду, обозначенную как pvWF-VAL и внесенную в штамм S ⊘ 930 культуры E. coli Американской коллекции типовых культур микроорганизмов под инвентарным номером 68530, плазмиду с обозначением pvWF-VA3, плазмиду с обозначением pvWF-VB3, плазмиду с обозначением pvWF-VC3, внесенную в штамм S ⊘ 930 культуры E. coli Американской коллекции типовых культур микроорганизмов под инвентарным номером 68241, плазмиду с обозначением pvWF-VCL, внесенную в штамм 4300 /F-/ культуры E. coli Американской коллекции типовых культур микроорганизмов под инвентарным номером 68242.
Все штаммы хозяина культуры E. coli, описанные выше, могут быть "заготовлены" из плазмид, их составляющих, способами, хорошо известными в этой области техники, например, по способу с бромидом этидия, который описал Новик (R.P. Novick) в журнале Bacteriol. Review 33, 210 (1969).
Кроме того, предметом изобретения является способ продуцирования любого из описанных выше полипептидов, который включает в себя трансформацию бактериальной клетки с плазмидой экспрессии, кодирующей полипептид, культивирование результирующей бактериальной клетки с продуцированием полипептида, закодированного плазмидой, и извлечение полипептида, продуцированного таким способом.
К тому же, изобретение касается способа лечения субъекта с цереброваскулярным расстройством, который включает в себя введение субъекту некоторого количества какого-либо из полипептидов, отвечающих изобретению и эффективных в отношении торможения процесса агрегации тромбоцитов.
Кроме того, дается способ лечения субъекта с сердечно-сосудистым расстройством, который включает в себя введение субъекту некоторого количества полипептида, эффективного в отношении торможения процесса агрегации тромбоцитов. К примерам сердечно-сосудистых расстройств, приемлемых для лечения, относятся острый инфаркт миокарда или стенокардия.
Далее предметом изобретения является способ ингибирования агрегации тромбоцитов у субъекта до, во время или после того, как субъект подвергся пластической операции на сосудах, тромболитической терапии или коронарной операции с искусственным кровообращением, который включает в себя введение субъекту некоторого количества полипептида, отвечающего изобретению, который является эффективным в отношении торможения процесса агрегации тромбоцитов.
Изобретение также касается способа обеспечения проходимости кровеносных сосудов у субъекта до, во время или после того, как субъект подвергся коронарной операции с искусственным кровообращением, который включает в себя введение субъекту некоторого количества полипептида, отвечающего изобретению, который является эффективным в отношении торможения процесса агрегации тромбоцитов.
Изобретение также касается способа терапии субъекта с карциномой, который включает в себя введение субъекту некоторого количества полипептида, отвечающего изобретению, который является эффективным в отношении торможения развития опухолевого метастаза.
Изобретение также касается способа ингибирования тромбообразования у субъекта, который включает в себя введение субъекту некоторого количества полипептида, отвечающего изобретению, который является эффективным в отношении торможения процесса тромбообразования. Тромбоз может быть связан с реакцией на воспалительный процесс.
Кроме того, предметом изобретения является полипептид, присоединенный к твердой матрице, который отвечает изобретению.
Изобретение также касается способа терапии субъекта, страдающего от прилипания тромбоцитов к поврежденным васкулярным поверхностям, который включает в себя введение субъекту некоторого количества полипептида, отвечающего изобретению, который является эффективным в отношении торможения прилипания тромбоцитов к поврежденным васкулярным поверхностям.
Изобретение также касается способа предотвращения адгезии тромбоцитов к протезному материалу и устройству у субъекта, который включает в себя введение субъекту некоторого количества полипептида, отвечающего изобретению, который является эффективным в отношении предотвращения адгезии тромбоцитов к материалу или устройству.
Изобретение также касается способа ингибирования реокклюзии у субъекта после проведения пластической операции на сосудах или тромболиза, который включает в себя введение субъекту некоторого количества полипептида, отвечающего изобретению, который является эффективным в отношении торможения окклюзии.
Изобретение также касается способа предотвращения возникновения вазоокклюзивных кризов у субъекта, страдающего серповидноклеточной анемией, который включает в себя введение субъекту некоторого количества полипептида, отвечающего изобретению, который является эффективным в отношении предотвращения возникновения вазоокклюзивных кризов.
Изобретение также касается способа предотвращения возникновения артериосклероза у субъекта, который включает в себя введение субъекту некоторого количества полипептида, отвечающего настоящему изобретению, который является эффективным в отношении предотвращения развития артериосклероза.
Изобретение также касается способа тромболитической терапии тромбосодержащих агрегатов с большим числом тромбоцитов у субъекта, который включает в себя введение субъекту некоторого количества полипептида, отвечающего изобретению, который является эффективным в отношении воздействия на тромбосодержащие агрегаты с большим числом тромбоцитов.
Изобретение также касается способа предотвращения тромбоцитной активации и образования тромба вследствие действия высоких касательных сил у субъекта со стенозированными или частично перекрытыми артериями, который включает в себя введение субъекту некоторого количества полипептида, отвечающего изобретению, который является эффективным в отношении предотвращения активации тромбоцитов и образования тромба.
Изобретение также касается способа предотвращения активации тромбоцитов под воздействием тромбина у субъекта, который включает в себя введение субъекту некоторого количества полипептида, отвечающего изобретению, который является эффективным в отношении предотвращения инициированной тромбином тромбоцитной активации.
Изобретение также касается способа предотвращения стеноза, вызванного пролифирацией гладкой мышцы после васкулярной травмы, у субъекта, который включает в себя введение субъекту некоторого количества полипептида, отвечающего изобретению, который является эффективным в отношении предотвращения развития стеноза.
Изобретение также касается способа извлечения очищенного биологически активного полипептида, отвечающего изобретению, который включает в себя:
а) продуцирование в бактериальной клетке первого полипептида с аминокислотной последовательностью у полипептида, но с отсутствующей дисульфидной связью;
б) разрушение бактериальной клетки с продуцированием лизата, содержащего первый полипептид;
в) обработку лизата с получением тел включения, содержащих первый пептид;
г) контактирование тел включения, полученных на стадии в), с получением первого пептида в растворимой форме;
д) обработку результирующего первого пептида с образованием биологически активного полипептида;
е) извлечение полученного биологически активного полипептида;
ж) очистку извлеченного биологически активного полипептида.
Стадия д) может включать в себя контактирование полипептида с тиолсодержащим соединением и дисульфидом, чем обеспечиваются перегибание и переокисление полипептида. Желательно, чтобы тиосодержащие соединения представляли собой глутатион, тиоредоксин, β -меркаптоэтанол или цистеин.
Стадия контактирования г) может быть проведена в присутствии денатурирующего вещества, такого как хлористоводородный гуанидин или мочевина.
Извлечение полипептида на стадии е) может проводиться одновременно с удалением денатурирующего вещества посредством диализа.
На стадии ж) биологически активный пептид может быть очищен проведением катионообменной хроматографии.
Первый полипептид может быть также очищен проведением катионообменной хроматографии после стадии г).
Примеры, которые приведены ниже, способствуют пониманию изобретения, и если не направлены они на какое-либо ограничение предмета изобретения и не должны рассматриваться в этом плане. Примеры не содержат детальные описания общепринятых способов, применяемых при конструировании векторов, при введении генов, кодирующих представляющих интерес полипептиды в такие векторы и при введении результирующих плазмид в бактериальных хозяев. Такие способы являются хорошо известными специалистам, работающим в этой области техники, и они описаны в многочисленных публикациях, включая руководство Самбрука и др. (Sambrook Fritisch and Maniatis, Molecular Cloning : A Laboratory Manual, and Edition, Cold Spring Harbor Laboratory Press, USA, (1989)).
Все ссылки на картированные положения соответствуют идентичным образом перенумерованным положениям по транслированной нуклеотидной последовательности у фактора фонд Вилленбранда для взрослого человека, что показано на фиг. 12 - 19
Пример 1. Клонирование и экспрессия полипептидов связующей области GPIb у фактора фонд Виллебранда vWF.
кДНК-Клонирование связующей области GPIb человеческого фактора vWF.
Человеческая эндотелиальная библиотека кДНК-данных (полученная у фирмы "КЛОНТЕХ лабораториз, инк.") для λ gtll была проверена на наличие положительных последовательностей у человеческого фактора vWF при использовании синтетических ДНК-проб. Пробы синтезировали в согласии с опубликованной ДНК-последовательностью (Sadler et al., Proc. Nat., Acad. Sci. 82: 6394-8 (1985) and Verweij et al. , EMBO J. 3: 1829-47 (1986)) человеческого vWF-фактора (известно, что фланговые 5'-конец и 3'-конец области vWF-фактора связаны с GPIb-рецептором) (фиг. 12 - 19).
Синтетические пробы обладали следующими последовательностями:
Последовательность - Нуклеотиды
AAATCTGGCAGTGCTCAGGGTCACTGGGATTCAAGGTGAC - 3863-3902
CCAGGACGAACGCCACATCCAGAACCATGGAGTTCCTCTT - 4700-4739
Ряд кДНК-клонов vWF-фактора, охватывающих всю связующую область GPIb, был идентифицирован и выделен, кДНК-фрагменты были субклонированы по EcoRI-сайту плазмиды (фиг. 26) (фирма "Нью Ингланд биолаб., инк."). Один из субклонов, обозначенный как pvWIP (фиг.1), содержит вставку в 2500 оснований. Эта вставка в 2500 оснований охватывает всю связующую область GPIb, простирающуюся вверх от GPIb-связующего сайта на 550 пар оснований и вниз от GPIb-связующего сайта на 1100 оснований (Субклон pvWIP был также обозначен как pvWF-IP).
Манипулирование ДНК-кодированием для связующей области GPIb фактора vWF
Чтобы получить экспрессию связующей области GPIb у культуры Escherichia coli при регулирующем воздействии промотора deo P1P2, КДНК-фрагмент vWF-фактора, выделенный из плазмиды pvWIP, использован при последующих манипулированиях так, как это описано ниже. Как отмечали ранее, фрагмент триптического расщепления vWF-фактора, который связывает GPIb-рецептор, простирается от аминокислоты Val 499 (валин) до аминокислоты Lys 728 (лизин).
A. Субклонирование 5'-конца связующей области GPIb фактора vWF и добавление ATG-кодона инициирования трансляции.
У плазмиды pvWIP имеются два подходящих сайта ограничения на 5'-конце; BSu 361, в результате чего происходит рассечение ДНК-последовательности, соответствующее аминокислоте Ser (445, серия), и TthIIII, в результате чего рассечение приходится на аминокислоту Asp (514, аспарагин). Конструировали синтетические фрагменты различного размера, в которые вставляли ATG-кодон, инициировали трансляции на 5'-конце, а также дополнительные аминокислоты. Это, прежде всего, делали для достижения максимальной вероятности получения высоких уровней экспрессии. Во-вторых, они являлись первым этапом на пути понижения размера пептида связующей области GPIb у vWF-фактора до минимальной величины, необходимой, возможно, для исключения коллагенового и гепаринового связующих сайтов, которые, в конечном итоге, могут воздействовать на функциональное назначение продукта.
A1. Аминокислота Glu (глутаминовая кислота) 437 на 5'-конце.
Синтетические олигомеры с последовательностями вида
5' - T 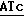 G-AGCTGCCTCG-CCGGCGTTTGCC - 3'
G-AGCTGCCTCG-CCGGCGTTTGCC - 3'
3' - 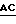 CTCCACCGACCGGCCGCAAAAC
CTCCACCGACCGGCCGCAAAAC 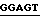 - 5'
- 5'
Ndel Bsu 361
присоединяли к плазмиде pvWF-IP, подвергнутой расщеплению при воздействии Ndel и Bsu 361 (фиг.2). Полученную плазмиду обозначали как pvWF-VAl. Плазмида pvWF-VAl находилась в штамме S ⊘ 930 культуры E. coli и была внесена в Американскую коллекцию типовых культур микроорганизмов под инвентарным номером 68530.
A2. Аминокислота Phe (Фенилаланин) 443 на 5'-конце.
Синтетические олигомеры с последовательностями вида
5' - T 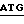 TTTGCC - 3'
TTTGCC - 3'
3' - ACAAA CGGAGT - 5'
присоединяли к плазмиде pvWIP, подвергнутой расщеплению при воздействии Ndel и Bsu 361 (фиг.3). Полученную плазмиду обозначали как pvWF-VBl.
B. Субклонирование 3'-конца связующей области GPIb фактора vWF, введение кодона остановки трансляции.
B1. Введение стоп-кодона в плазмиду pvWF-VAl
Синтетический олигомер с последовательностью вида
5' - CCGGGGCTCTTGGGGGTTTCGACCCTGGGGGCCCAAG 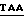 GATATCA - 3'
GATATCA - 3'
3' - CCGAGAACCCCCCAAAGCTGGGACCCCGGGTTCATTCTATAGTTCGA - 5'
присоединяли к плазмиде pvWF-VAl (фиг.4), подвергнутой расщеплению при воздействии Xmal и Hind III. Полученную плазмиду обозначали как pvWF-VA2. Эта вновь сконструированная плазмида содержит кодон окончания трансляции TAA, примыкающий к аминокислоте 728 (Lys (лизин) и EcoRV-сайту.
B2. Введение трансляционного стоп-кодона в плазмиду pvWF-VBl.
Синтетический олигомер с последовательностью вида
5' - CCGGGGCTCTTGGGGTTTCGACCCTGGGGCCCAAG 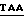 GATATCA - 3'
GATATCA - 3'
3' - CCGAGAACCCCAAAGCTGGGACCCCGGGTTCATTCTATAGTTCGA - 5'
присоединяли к плазмиде pvWF-VBl (фиг.4), подвергнутой расщеплению при воздействии Xmal и Hind III. Полученную плазмиду обозначали как pvWF-VB2 (фиг. 5).
Экспрессия связующей области GPIb факторов vWF у Escherichia coli.
Для получения экспрессии связующей области GPIb фактора vWF конструировали различные плазмиды экспрессии, основываясь на deo P1P2-составляющей промоторной системе.
1. Экспрессия полипептида связующей области GPIb фактора vWF, включающей аминокислоты от аминокислоты GI и 437 (глутаминовая кислота) до аминокислоты Lys 728 (лизин) (основанная на плазмиде pvWF-VA2).
Используя NdeI и EcoRV, из плазмиды pvWF-VA2 выделяли фрагмент и присоединяли к плазмиде pMF-945 (фиг.11), подвергнутой расщеплению с использованием NdeI и Pvu11 (фиг. 6). Полученная плазмида была обозначена как pvWF-VA3 и была встроена в штамм S ⊘ 930 культуры Escherichia coli.
Экспрессия полипептида связующей области GPIb фактора vWF, включающей аминокислоты от аминокислоты Phe 443 (фенилаланин) до аминокислоты Lys 728 (лизин) (основанная на плазмиде pvWF-VB2).
Используя Ndel и EcoRV, из плазмиды pvWF-VB2 выделяли фрагмент и вводили в плазмиду pMF-945, подвергнув расщеплению с использованием Ndel и PvuII (фиг. 7). Полученную плазмиду обозначали как pvWF-VB3, и она была встроена в штамм S ⊘ 930 культуры Escherichia coli.
3. Экспрессия полипептида связующей области GPIb фактора vWF, включающей аминокислоты от аминокислоты Leu 504 (лейцин) до аминокислоты Lys 728 (лизин) (основанная на экспрессии плазмиды PcWF-VA3).
Синтетический олигомер с последовательностью вида
5' - T 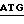 TTGCACGATTTCTACTGCAGCAGGCTACTGGACC - 3'
TTGCACGATTTCTACTGCAGCAGGCTACTGGACC - 3'
3' - 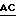 AACGTGCTAAAGATGACGTCGTCCGATGAC
AACGTGCTAAAGATGACGTCGTCCGATGAC 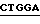 - 5'
- 5'
Ndel TthIIII
присоединяли к плазмиде pvWF-VA3, подвергнутой расщеплению с использованием Ndel и TthIIII. Полученную плазмиду обозначали как pvWF-VC3 (фиг. 8). Плазмида pvWF-VC3 была встроена в штамм S ⊘ 930 культуры Escherichia coli и была внесена в Американскую коллекцию типовых культур микроорганизмов под инвентарным номером 68241 (фиг. 20).
4. Экспрессия полипептида связующей области GPIb фактора vWF, включающей аминокислоты от аминокислоты Leu 513 (лейцин) до аминокислоты Lys 728 (лизин) (основанная на экспрессии плазмиды pvWF-VA3).
Синтетический олигомер с последовательностью вида
5' - T 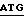 CTGGATCC - 3'
CTGGATCC - 3'
3' - 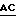 GAC
GAC 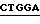 - 5'
- 5'
Ndel TthIIII
присоединяли к плазмиде pvWF-VA3, подвергнутой расщеплению с использованием Ndel и TthIIII. Полученную плазмиду обозначали как pvWF-VD3 (фиг. 9). Плазмида pvWF-VD3 была встроена в штамм S ⊘ 930 культуры Escherichia coli.
Экспрессия полипептидов связующей области GPIb фактора vWF.
Относительно последовательное расположение плазмид экспрессии показано на фиг. 10. Плазмиды pvWF-VA3, pvWF-VB3, pvWF-VC3 и pvWF-VD3 у штамма S ⊘ 930 культуры Escherichia coli были использованы для проверки анализа уровней экспрессии у различных пептидов связующей области GPIb фактора vWF. Концы, полученные таким способом, выращивали в LB-среде, содержащей ампициллин (100 мг/мл), что делали при 37oC в течение 48 ч.
По истечении 48 ч роста бактериальные клетки собирали и центрифугировали в течение 2 мин при скорости 10000 об/мин. Остатки от центрифугирования растворяли в 1/10 объема трис- HCl с концентрацией 50 мМ при pH величиной 8,0. Добавляли некоторое количество буферной смеси (содержащей додецилсульфат натрия и β -меркаптоэтанол). Образцы кипятили в течение 10 мин и вносили в полиакриламидный гель, содержащий додецилсульфат натрия в количестве 10%. Экспрессия полипептидов связующей области GPIb фактора vWF в клонах pvWF-VA3, pvWF-VB3 и pvWF-VD3 была низкой в сравнении с бактериальными всеми белками.
Полипептиды фактора vWF из этих клонов могут быть обнаружены при проведении капельного анализа Уэстерна (Western) при использовании промышленно выпускаемого поликлонального vWF-антитела (акц. общество "Декопаттс/", Глосруп, Дания). Однако клоны, проистекающие из штамма S ⊘ 930 культуры Escherichia coli, трансформированного плазмидой pvWF-VС3, экспрессированной полипептидом связующей области GPIb фактора vWF (от аминокислоты Leu 504 (лейцин) до аминокислоты Lys 728 (лизин), включая метионин), являются с высокой степенью обнаруживаемыми окрашиванием по Кумасси (Coomassie).
Штамм S ⊘ 930 культуры Escherichia coli, содержащий плазмиду pvWF-VC3, был внесен в Американскую коллекцию типовых культур микроорганизмов под инвентарным номером 68241. Затем была сконструирована искусственная плазмида, которая содержала ту же кодирующую область фактора vWF, что и плазмида pvWF-VC3, выраженную под управлением λ PL-промотора и deo-рибосомального связующего сайта (фиг. 21). Эта новая плазмида, обозначенная как pvWF-VCL, является, как оказалось, сильным экспрессором VCL, полипептида связующей области GPIb фактора vWF (метионин и аминокислоты от аминокислоты Leu 504 (лейцин) до аминокислоты Lys 728 (лизин). Эта плазмида, находящаяся в штамме 4300 культуры Escherichia coli, была внесена в Американскую коллекцию типовых культур микроорганизмов под инвентарным номером 68242.
Штамм 4300 культуры Escherichia coli, сконструированный из штамма культуры Escherichia coli, фиксирующего в Американской коллекции типовых культур микроорганизмов под инвентарным номером 12435, зависимый от биотина штамм дикого типа F-, содержащий чувствительный к температуре λ с 1857-репрессор (Третья плазмидная конструкция, содержащая ту же vWF-кодирующую область, находящуюся под управлением λ -промотора и cII-рибосомального связующего сайта, не экспрессирует какой-либо vWF-пептид, обнаруживаемый окрашиванием по Кумасси (Coomassie)).
Ndel-HindIII-Вставка плазмиды pcWF-VCL может быть легко субклонирована в другие векторы экспрессии, такие как промышленно выпускаемая плазмида (фиг. 26), используемая для продуцирования ряда полипептидов; которые включают в себя ту же аминокислотную последовательность в пределах от аминокислоты 509 (цистин) до аминокислоты 695 (цистин) и обладают такой же биологической активностью.
Пример 2. Ферментация бактерий, экспрессирующих полипептиды связующей области GPIb фактора vWF.
При увеличении числа ферментаций клона pvWF-VC3 было обнаружено, что хозяин обладает тенденцией терять плазмиду из-за нестабильности. Потеря плазмид сопровождается понижением экспрессии у полипептида связующей области GPIb фактора vWF. Было найдено необходимым непрерывно поддерживать давление отбора (т.е. непрерывно добавлять ампициллин), чтобы сохранять число плазмидных копий и поддерживать уровни экспрессии. Крупномасштабную ферментацию проводили в течение 12 ч.
Ферментацию проводили в следующей ростовой среде, г:
N - Z-амино-AS - 20
Дрожжевой экстракт - 10
NaCl - 5
K2HPO4 - 2,5
MgSO4•7H2O - 1,0
Пеногаситель - 0,4
К ростовой среде добавляли фруктозу (50%) при конечной концентрации 451 мл/л, и в ферментер непрерывно вводили ампициллин (100 мг/мл при общем количестве 8 мл/л). Ферментацию проводили в течение 12 ч при 37oC.
Очистка полипептидов.
Клетки собирали через 12 ч после начала ферментации и центрифугировали. Полученный от центрифугирования бактериальный осадок вновь суспендировали в буферном растворе (50 мМ трис-кислоты при pH величиной 8,0, 50 мМ NaCl, 1 мМ этилендиаминтетрауксусной кислоты, 1 мМ дитиотреитола, 1 мМ фенилметилсульфонилхлорида и 10% глицерола). После дополнительного центрифугирования и разрушения клеточной взвеси ультразвуком полипептид связующей области GPIb фактора vWF находили в осадке от центрифугирования.
Полипептид связующей области GPIb фактора vWF подвергали дальнейшей очистке посредством солюбилизации осадка от центрифугирования в мочевине с концентрацией 8 М, содержащей 10 мМ дитиотреитола, 25 мМ трис-кислоты при pH величиной 8 и 1 мМ этилендиаминтетрауксусной кислоты, что делали при комнатной температуре. Солюбилизированный осадок от центрифугирования фракционировали, провод ионообменную колоночную хроматографию на (диэтиламино)этилцеллюлозе. (Использовали ту же элюирующую буферную смесь, что и выше, но не содержащую 0,5 мМ дитиотреитола).
Полипептид связующей области GPIb фактора vWF элюировали при содержании хлорида натрия 150 мМ. После разбавления до содержания хлорида натрия 50 мМ (в указанной выше буферной смеси) частично очищенный пептид подавали в колонку с Q-сефарозой. Элюирование из колонки с Q-сефарозой проводили при разных концентрациях хлорида натрия (ступенчатое элюирование).
Пептид связующей области GPIb фактора vWF был представлен четырьмя пиками, которые появлялись при элюировании хлоридом натрия при содержаниях 100, 200, 250 и 500 мМ. В случае всех четырех пиков диализ проводили при содержаниях хлорида натрия 150 мМ и трис-смеси 50 мМ с величиной pH 8, что делали в течение 8 ч. При проведении диализа концентрацию мочевины в диализном растворе понижали по линейному закону до отсутствия мочевины, начиная с содержания в 6 М.
Пример 3. Биологическая активность пептидов связующей области GPIb фактора vWF.
Анализ агрегации тромбоцитов.
Приготовление vWF-фактора.
Человеческий извлеченный из плазмы vWF-фактор очищали от известной совокупности банковских компонентов человеческой плазмы согласно Лоскалцо и Хандину (J. Loscalzo and R.I.Handin, Biochemistry, 23: 3880-3886 (1984)). Очищенный vWF-фактор, выделенный из плазмы, концентрировали на фильтрующей мембране из амикона с отсечкой 100000 до конечной концентрации 0,25 мг/мл.
Приготовление асиало-vWF-фактора.
Очищенный человеческий vWF-фактор, выделенный из плазмы, подвергали десиализации согласно ДеМарко и Шапиро (L. DeMarco and S.Shapiro, J.Clin. Invst, 68: 321-328 (1981)) при следующих изменениях.
1. Использованная нейраминидаза была взята из культуры Vibrio cholera типа II (сигма).
2. Реакционная смесь содержала 0,2 ед. фермента на 1 мг ингибиторов белка и протеазы при следующих концентрациях: бензамидин - 20 мМ, лейпептин - 15 мг/мл и апротинин - 20 ед., на 1 мл. Асиало - vWF-фактор использовали для проведения агрегации тромбоцитов для дальнейшей очистки.
Агрегация тромбоцитов, индуцированная асиало- vWF-фактором.
Как говорили выше, растворимый vWF-фактор не связывается с тромбоцитами через GPIb-рецептор. Асиало-vWF-фактор, полученный проведением нейраминидазной обработки с целью удаления остатков сиаловой кислоты, легко прилипает к тромбоцитам через G-PIb. Десиалирование, видимо, понижает полный отрицательный заряд vWF-фактора, в результате чего оказывается возможным связывание с ним отрицательно заряженного GPIb-рецептора. Асиало-vWF-связывание с тромбоцитами сопровождается активацией, высвобождением аденозиндифосфата и GPIIb/IIIa-нейромедиаторной агрегацией. Тромбоцитную агрегацию, индуцированную асиало- vWF-фактором, проводили, используя 200 мкл богатой тромбоцитами плазмы (Fujimura Y., et al., J.Biol. Chem. 261: 381-385 (1986)) и 39 мкг/мл асиало- vWF-фактора (конечная концентрация) с применением измерителя агрегации Луми (Lumi). Результаты по торможению тромбоцитной агрегации с воздействием VC-полипептида связующей области GPIb фактора vWF, суммированы в табл. 1.
Полипептид VC (также обозначаемый как VCL или VC3) представляет собой полипептид связующей области GPIb фактора vWF, который содержит метионин и аминокислоты 504-728 (фиг. 12-19).
Агрегация тромбоцитов, индуцированная vWF-ристоцетином.
Индуцированную ристоцетином тромбоцитную агрегацию в присутствии очищенного человеческого интактного vWF-фактора проводили с вымыванием человеческих тромбоцитов согласно Фуджимуре и др. (Fujimuera Y. et al., Biol. Chem. 261: 381-385 (1986)).
Результаты ингибирования тромбоцитной агрегации, индуцированной ристоцетином в присутствии интактного vWF-фактора, суммированы в табл. 2. Дополнительные результаты, полученные при проведении этих анализов, описаны в примере 5.
Пример 4. Улучшенный способ получения чистого, окисленного, сложенного и биологически активного полипептида связующей области GPIb фактора vWF.
В примере 2 была описана ферментация клеток, содержащих плазмиду pvWF-VC3. Затем, как это описано в примере 1, была сконструирована предпочтительная плазмида pvWF-VCL, и она находилась в штамме A4300 культуры E. coli. Эта система из хозяина и плазмиды была подвергнута ферментации, что делали так, как обычно поступают в этой области техники в случае факторов, содержащих ген, выраженный под управлением λPL-промотора; см., например, совместную патентную публикацию Европейского патентного ведомства N 173280, опубликованную 5 марта 1986 г., пример 5, с.73-74 (без добавления биотина, тиамина, следовых элементов и ампициллина). В этом улучшенном способе очистки полипептида связующей области GPIb фактора vWF использовали клеточный осадок от центрифугирования, полученный после проведения описанной выше ферментации системы A4300/pvWF-VCL.
В этом улучшенном способе получается более чистый и более активный полипептид, чем получаемый по способу, раскрытому в примере 2. Общая поэтапная схема сводится к проведению следующих этапов от A до H.
A. Разрушение клеток и суспендирование осадка от центрифугирования. Осадок от центрифугирования, содержащий полипептид связующей области GPIb фактора vWF, получали так, как это описано в примере 2, проводя разрушение клеточной взвеси ультразвуком и центрифугирование клеточной суспензии в смеси, содержащей 50 мМ трис-HCl при pH величиной 8, 50 мМ хлорида натрия, 1 мМ этилендиаминтетрауксусной кислоты, 1 мМ дитиотреитола, 1 мМ фтористого фенилметилсульфонила и 10% глицерола.
Осадок от центрифугирования, содержащий тела включения, растворяли в примерно 10%-ном (по массе на объем) растворе такого состава, в случае которого конечная концентрация после растворения оказывалась следующей: 8 М мочевины, 20 мМ OTT (дитиотреитола), 20 мМ HEPES при pH с величиной 8 и 100 мМ хлорида натрия. Результирующий раствор мог быть подвергнут дальнейшей очистке использованием ионообменной хроматографии, как это описано ниже. Или же тела включения могли быть солюбилизированы в буферной смеси, содержащей 6 М хлористого гунидина, с последующей заменой буферной смеси на мочевину. Тела включения могли быть также растворены при разных концентрациях мочевины, хлористого гуанидина или какого-либо иного денатурирующего вещества или же при отсутствии денатурирующих веществ, например, при крайних значениях величины pH.
B. Катионообменная хроматография. На этом этапе происходит удаление большинства загрязняющих веществ и получается полипептид связующей области GPIb фактора vWF с чистотой, превышающей 90%. На этом этапе может быть использован любой способ проведения катионного обмена (например карбоксиметильный), но предпочтение следует отдавать CM-сефарозной высокопроточной (фирма "Фармация") хроматографии. В качестве функциональной группы может выступать карбоксиметильная группа, фосфогруппа или сульфоновые группы, такие как сульфопропильная группа. В основе матрицы могут быть неорганические соединения, синтетические смолы, полисахариды или органические полимеры; приемлемыми матрицами являются агароза, целлюлоза, трисакрил, декстран, стеклянные бусинки, оксиранакриловые бусинки, акриламид, агароза/полиакриламидный сополимер (ультрогель) или гидрофильный виниловый полимер (фактогель). В конкретном варианте реализации настоящего изобретения полипептид подают в CM-сефарозную высокопроточную колонку, уравновешенную со смесью, содержащей 8 М мочевины, 1 мМ DTT (дитиотреитол), 20 мМ HEPES при pH с величиной 8, 100 мМ хлорида натрия. Чистый полипептид элюировали в смесь, содержащую 8 М мочевины, 1 мМ DTT (дитиотреитол), 20 мМ HEPES при pH с величиной 8 и 200 мМ хлорида натрия. На 1 мл CM-сефарозы FF может быть подано примерно до 30 ед. OD280 солюбилизированных тел включения. При этом соотношении концентрация у элюированного полипептида в типичном случае равна 4-5 OD280/мл.
C. Окисление и новое изгибание. Раствор полипептида, элюированного при проведении описанной выше стадии катионного обмена, обрабатывали 6 М раствором хлористого гуанидина, что делали для разрушения агрегатов. Полипептид затем разбавляли до концентрации 0,05 OD280/мл, делая это смесью, содержащей 2 М хлористого гуанидина, желательно, 20 мМ HEPES при pH с величиной 8, 0,1 мМ GSSG (глутатионин, окисленная форма). Эту смесь оставляли на ночь при комнатной температуре. Продукты анализировали гель-фильтрацией, проводя быструю белковую жидкостную хроматографию, типа проводимой с использованием суперозы 12, что делали перед тем, как продолжать методику дальше. Из результатов анализа следовало, что при этой белковой концентрации воспроизводимо содержится примерно 30% надлежащим образом окисленных мономеров. Чем выше белковая концентрация, тем выше абсолютный выход надлежащим образом окисленных мономеров, но при этом оказывается пониженным процентное содержание, что обусловлено усиленным образованием связанных через серу димеров и мультимеров. Например, при белковой концентрации величиной 0,1 OD280/мл получается лишь 20% надлежащим образом окисленных мономеров. При понижении концентрации до 0,025 OD280/мл выход надлежащим образом окисленных мономеров составляет 35-40%, но при этом оказывается пониженным абсолютный выход в расчете на 1 л окисляемого вещества. Операцию по окислению можно также проводить в мочевине вместо хлористого гуанидина или в каком-либо ином денатурирующем веществе, или при отсутствии денатурирующих веществ в надлежащих буферных условиях, например величина pH, ионная сила и гидрофобность. Предпочтительная концентрация мочевины лежит в области от 0,5 до 10 М, желательно, чтобы она составляла 4 М; и предпочтительным окислителем является глутатион, находящийся в окисленной форме и присутствующий в концентрациях от 0,01 до 5 мМ, причем желательно, чтобы концентрация составляла 0,1 мМ. Могут быть использованы и другие окисляющие вещества, такие как дихлорид меди, или же окисляющее вещество может не добавляться, чем обеспечивается протекание окисления только за счет воздуха. При крупномасштабных операциях на стадии окисления предпочтительным раствором является раствор мочевины с концентрацией 4 М.
D. Концентрация. Окисленные продукты концентрировали, доводя, желательно, до концентрации порядка OD280 = 1, чего достигали, используя тангенциальную проточную ультрафильтрационную систему с мембраной, имеющей отсечку в 30000, такой как система "MINITAN" или "PELLICON" с материалом миллипор. Фильтрат был вполне прозрачным, поскольку материал был сравнительно прозрачным и поскольку большинство загрязнений были достаточно большими, что мешало их прохождению через мембрану с отсечкой в 30000. Этим обеспечивалась возможность повторного использования фильтрата для проведения окислений. При этом достигалась значительная экономия, поскольку хлористый гуанидин, используемый в больших количествах при концентрации 2 М, является довольно дорогим. По результатам проведения анализа методом быстрой белковой жидкостной хроматографии не обнаруживали различия у продуктов окисления, полученных в операциях окисления с повторным использованием фильтрата в сравнении со свежеприготовленным 2 М хлористым гуанидином.
E Диализ. Необходимо понизить концентрацию хлористого гуанидина или мочевины до величины, составляющей менее 10 мМ. Этого достигали, проводя диализ относительно смеси, содержащей 20 мМ HEPES при pH величиной 8 и 10 мМ хлорида натрия.
Диализ проводили в трубчатом диализаторе, заменяя 3-3 раза буферную смесь; но можно с той же целью проводить диафильтрацию относительно той же буферной смеси, делая это в тангенциальной проточной ультрафильтрационной системе с мембраной, отсекающей образования с мол.мас. выше 10000.
При проведении диализа по мере падения концентрации хлористого гуанидина (или мочевины или иного денатурирующего вещества) происходит образование белого осадка. Этот осадок содержит примерно 80% белка, полученного на стадии D, который содержит связанные через серу димеры (S-S-димеры), восстановленный и неправильно окисленный мономер и некоторые загрязнения, которые претерпели коэлюирование на стадии катионного обмена. Надосадочная жидкость почти на 100% представляет собой правильно окисленный и вновь изогнутый мономер с концентрацией 0,2 OD280/мл, который примерно на 20% представляет собой белок, полученный на стадии D. Это избирательное осаждение загрязнений и нежелательных образований белка, являющееся следствием проведения диализа, являлось поразительным и не могло быть предсказано. Выход надлежащим образом окисленного мономера может быть значительно повышен посредством извлечения из осадка. Этого достигают следующим образом: раствор осветляют центрифугированием. Надосадочную жидкость сохраняют, и осадок от центрифугирования обрабатывают дитиотреитолом, что ведет к восстановлению S-S-связей и реокислению, как это говорили выше. Осадок от центрифугирования растворяют в минимальном количестве смеси, содержащей 6М хлористого гуанидина, 20 мл HEPES при pH с величиной 8, 150 мМ хлорида натрия и 20 мМ дитиотреитола. Раствор пропускают через сефадекс G25 в буферную смесь, аналогичную растворяющей буферной смеси, но при этом содержащей лишь 1 мМ дитиотреитола (вместо 20 мМ). Элюат затем разбавляют до достижения концентрации величиной 0,05 OD280/мл и обрабатывают так, как это делали выше на стадиях C, D и E. Эта методика может быть повторена несколько раз до получения дополнительно очищенного мономера. Все надосадочные жидкости соединяют.
F. Катионный обмен. Объединенную надосадочную жидкость диализата, полученную на стадии E концентрируют, подвергая воздействию CM сеферозы в смеси, содержащей 20 мМ HEPES при pH величиной 8 и 100 мМ хлорида натрия. Элюирование проводят, используя смесь, содержащую 20 мМ HEPES при pH величиной 8 и 400 мМ хлорида натрия. Элюат является исключительно мономерным несмотря на высокую концентрацию соли. По этому способу получали концентрации, доходящие до 3 мг/мл, и такая концентрация не являлась верхним пределом. Или же эта стадия сегодня могла быть проведена с использованием гепарин-сефарозы, что также сопровождается связыванием очищенного мономера, находящегося в смеси, содержащей 10 мМ трис-реактива с pH величиной 7,4 и 150 мМ хлорида натрия. Элюирование из гепарин-сефарозы проводили, используя смесь, содержащую 10 мМ трис-HCl с pH величиной 7,4 и 500 мМ хлорида натрия.
G. Диализ. Продукт, полученный на предыдущем этапе, подвергали диализу относительно смеси, содержащей 20 мМ HEPES при pH величиной 8 и 150 мМ хлорида натрия.
H. Хранение. На этом этапе очищенный полипептид связующей области GPIb фактора vWF может быть подвергнут лиофилизации. При воссоздании в объеме воды, равном объему, бывшему до лиофилизации, результирующий раствор содержит исключительно мономерный белок и согласно результатам проведения быстрой белковой жидкостной хроматографии не содержит следы димеров или иных мультимеров.
В специальном варианте реализации предлагаемого способа использовали следующую методику.
A. Тела включения, взятые в количестве 10 г (с суммарной массой 0,43 г по сухому веществу), растворяли в смеси, в конечном объеме которой содержалось (100 мл) 8 М мочевины, 20 мМ OTT (дитиотреитола), 20 мМ HEPES при pH величиной 8 и 100 мМ хлорида натрия
B. Белок подавали в колонку с CM-сефарозой, уравновешенной относительно смеси, содержащей 8М мочевины, 1 мМ OTT (дитиотреитола), 20 мМ HEPES при pH величиной 8 и 100 мМ хлорида натрия. Белок элюировали при содержании хлорида натрия 200 мМ, находящегося в смеси, содержащей 8 М мочевины, 20 мМ HEPES при pH величиной 8 и 1 мМ OTT (дитиотреитола), и сохраняли.
C. Сохраненный элюат, полученный на предыдущем этапе, обрабатывали 6 М хлористым гуанидином, чем устраняли всякие агрегаты, и затем разбавляли до концентрации 0,05 OD280/мл смесью, содержащей 2 М хлористого гуанидина, 20 мМ HEPES при pH величиной 8 и 0,1 мМ GSSG (глутатион, окисленная форма). Окисление вели на протяжении ночи при комнатной температуре (следует заметить, что стадия окисления может быть проведена в присутствии мочевины, а не хлористого гуанидина).
D. Продукты окисления концентрировали до концентрации 1 OD280, проводя ультрафильтрацию на устройстве "MINITAN", содержащем мембрану с отсечкой в 30000.
Е. Концентрат, полученный на предыдущей стадии, подвергали диализу при трехкратной замене буферной смеси, что делали относительно смеси, содержащей 20 мМ HEPES при pH величиной 8 и 100 мМ хлорида натрия. При проведении диализа по мере падения концентрации хлористого гуанидина образовывался белый осадок, который удаляли центрифугированием, и еще раз повторяли процесс, проводя его, как описано выше. Надосадочные жидкости соединяли.
F. Соединенные надосадочные жидкости концентрировали, проводя связывание с СМ сефарозой, что делали в смеси, содержащей 20 мМ HEPES при pH величиной 8 и 100 мМ хлорида натрия. Полипептид элюировали в смесь, содержащую 20 мМ HEPES при pH величиной 8 и 400 мМ хлорида натрия и хранили при 4oC.
G. Сохраненный элюат, полученный на предыдущей стадии, подвергали диализу при 4oC, делая это относительно смеси, содержащей 20 мМ HEPES при pH величиной 8 и 150 мМ хлорида натрия.
H. После диализа очищенный полипептид связующей области GPIb фактора vWF, обозначенный как VCL, подвергали лиофилизации.
Анализ полипептида VCL.
1. Из результатов анализа на аминокислотную последовательность полипептида VCL, очищенного так, как это было описано выше, следовательно, что N-концевая последовательность представляет собой последовательность Met-Leu-His-Asp-Phe (последовательность метионин-лейцин-гистидин-аспаргиновая кислота-фенилаланин), которая является ожидаемой последовательностью в соответствии с фиг. 12-19 с добавлением N-концевого метионинового остатка.
2. Из анализа полипептида VCL, проведенного на полиакриламидных гелях, следует, что полипептид VCL претерпевает электрофорез при пониженной кажущейся молекулярной массе в невосстанавливающих условиях в сравнении с восстанавливающими условиями (бетамеркаптоэтанол). Этот сдвиг от плотной к менее плотной конфигурации находится в согласии с представлением о восстановлении дисульфидной связи. Такая внутримолекулярная связь образуется между цистинами, находящимися в положениях 509 и 695 (смещение у молекулярной массы не является столь большим, чтобы можно было говорить о восстановлении межмолекулярной связи).
Пример 5. Биологическая активность полипептида VCL, представляющего собой полипептид связующей области GPIb фактора vWF.
Полипептид связующей области GPIb фактора vWF, полученный, как это было описано в примере 4, обозначали как VCL и анализировали на биологическую активность, как это описано ниже.
1. Индуцированная ристоцетином агрегация тромбоцитов.
Индуцированную ристоцетином агрегацию тромбоцитов исследовали так, как это описано в примере 3, что делали в реакционной смеси, содержащей 2•108 тромбоцитов в 1 мл, 1 мгк/мл фактора vWF плазмы и 1 мг/мл ристоцетина. При тестировании брали ряд концентраций полипептида VCL, и величина IC50 у полипептида VCL, отвечающая трем анализам, была найдена равной 0,2-0,3 мкМ. Торможение в 100% достигалось при концентрации порядка 1 мкМ по полипептиду VCL.
2. Асиало- vWF-индуцированная агрегация тромбоцитов.
Исследование асиало-vWF-индуцированной агрегации тромбоцитов проводили так, как это описано в примере 3, это делали в измерителе агрегации Луми (Lumi), используя 200 мкл богатой тромбоцитами плазмы и 10 мкг/мл асиало- vWF-фактора. Тестированию подвергали ряд концентраций полипептида VCL, и величина IC50 у полипептида VCL, использованного при проведении этого анализа, была найдена равной 0,15 мкМ с достижением полного ингибирования при величине 0,5 мкМ.
3. Влияние полипептида VCL на уже образовавшиеся агрегаты.
Тестировали эффект от влияния полипептида VCL на агрегаты, ранее образовавшиеся в условиях индуцированной ристоцетином агрегации тромбоцитов. Агрегацию проводили по п.1, приведенному выше, при отсутствии полипептида VCL. При добавлении полипептида VCL в концентрации 0,5 мкМ агрегаты сразу же разрушались.
4. Ингибирование индуцированной тромбином агрегации тромбоцитов.
Опыт с индуцированной тромбином агрегаций тромбоцитов проводили, используя 0,025 ед./мл тромбина и тромбоциты, полученные с использованием страктана. Был тестирован ряд концентраций полипептида VCL, и в этом тесте величина IC50 у полипептида VCL была найдена равной 0,3 мкМ. Эффект является поразительным, поскольку в параллельном эксперименте полипептид VCL не был эффективным в отношении ингибирования прямого связывания тромбина с меткой в виде иода-125 с тромбоцитами.
5. Влияние на осаждение тромбоцитов в условиях течения.
В модельной системе, состоящей из вывернутой денудированной человеческой умбиликальной артерии, находящейся в проточной ячейке, может быть изучено осаждение тромбоцитов. Неизмененная человеческая кровь протекает через кусок артерии. По прошествии 10-15 мин поток останавливают и микроскопически определяют осаждение тромбоцитов. В этой системе величина IC50 у полипептида VCL найдена примерно равной 1 мкМ.
Все приведенные выше результаты суммированы в табл. 3.
Ингибиторная активность полипептида VCL в отношении инициированной ристоцетином или асиало- vWF-инициированной агрегации тромбоцитов, индуцированного ристоцитином vWF-связывания и тромбоцитной адгезии, пропадает при восстановлении дисульфидной связи, действующей между цистеинами. Полипептид VCL выпадал из раствора.
Пример 6. Конструкция плазмиды pvWF-V EL.
Было решено сконструировать плазмиду, которая экспрессирует несколько большую часть связующей области GPIb фактора vWF, чем это делает плазмида pvWF-VCL. Конструкция показана на фиг. 22-25, и описание содержится в кратком описании фигур.
А. Конструкция плазмиды pvWF-VE2.
Плазмида pvWF-VA2 (сконструированная, как это показано на фиг. 4) была подвергнута расщеплению с использованием NdeI и PstI, и был выделен большой фрагмент. Были приготовлены четыре синтетические олигомера, показанных на фиг. 23. Олигомеры с номерами 2 и 3 были обработаны Т4-полинуклеотидной киназой, чем обеспечивалось добавление 5'-фосфата. Упомянутый выше большой фрагмент плазмиды pvWF-VA2 был соединен, как это показано на фиг. 22, с четырьмя олигомерами (с двумя подвергнутыми воздействию киназы и двумя не подвергнутыми воздействию киназы). Результирующая плазмида, показанная на фиг. 22, была обозначена как плазмида pvWF-VE2.
В. Конструкция плазмиды pvWF-VE3.
Плазмида pvWF-VE2 была подвергнута расщеплению с использованием NdeI и HindIII, и был выделен фрагмент из 770 пар оснований, содержащий связующую область GPIb фактора vWF. Плазмида pMLK-7891 была подвергнута расщеплению с использованием NdeI и HindIII, и был выделен большой фрагмент. Результирующая плазмида, показанная на фиг.24, была обозначена как pvWF-VE3.
С. Конструкция плазмиды pvWF-VEl.
Плазмида pvWF-VE3 была подвергнута расщеплению с использованием XmnI, дефосфорилирована бактериальной щелочной фосфатазой и затем подвергнута расщеплению с использованием NdeI и HindIII. Плазмида pMLK-100 была подвергнута расщеплению с использованием NdeI и HindIII и дефосфорилирована бактериальной щелочной фосфатазой. Два продукта расщепления были затем соединены с образованием плазмиды pvWF-VEL, как это показано на фиг. 25. Эта плазмида экспрессирует ДНК-последовательность, соответствующую аминокислотам 469-728 зрелого F-фактора при управлении λ PL-промотора и с N-рибосомального связующего сайта. Белок, по-видимому, включает также добавочный N-концевой метиониновый остаток. Консервативный переход оснований был введен в аланин с номером 473, что сопровождалось изменением GCC на GCA и что также кодирует аланин. Этим вводился SphI-сайт в ген за счет изменения GCCTGC на GCATGC.
Экспрессируя плазмиду pvWF-VEL в штамме 4300 (F-) культуры E. coli, получали белок в 29000 ед. Дальтона, который сильно взаимодействует с моноклональным анти- vWF-антителом и который обозначается здесь как VEL.
Пример 7. Фармацевтические применения полипептида связующей области GPIb фактора vWF.
В примерах 1 и 4 описаны продуцирование и очистка ранее неизвестного полипептида связующей области GPIb фактора vWF, обозначенного как VCL. Ниже описаны некоторые возможные применения полипептида VCL или других полипептидов связующей области GPIb фактора vWF. Фармацевтические составы, содержащие VCL или также иные полипептиды, могут быть приготовлены с использованием надлежащего фармацевтически приемлемого носителя при применении способов и носителей, хорошо известных в этой области техники.
1. Полипептид VCL описанного выше состава может быть использован для предотвращения адгезии тромбоцитов к поврежденным васкулярным поверхностям (см. пример 5, подраздел 5).
2. Полипептид VCL описанного выше состава может быть использован для разрушения агрегатов с высоким содержанием тромбоцитов (см. пример 5, подраздел 3).
1. Полипептид VCL описанного выше состава может быть использован для предотвращения реокклюзии после проведения пластической операции на сосудах или тромболиза (см. работу Беллинджера "Предотвращение возникновения обтурирующего коронарного артериального тромбоза воздействием муринового моноклонального антитела на свиной фактор фон Виллебранда" (Bellinger et al., PNAS, USA, 84: 8100-8104 (1987), Prevention of occlusive coronary artery thrombosis by a murine monoclonal antibody to porcine von Willebrand Factor)).
4. Полипептид VCL описанного выше состава может быть использовано для предотвращения тромбоцитной активации и образования тромба под воздействием высоких касательных сил, типа существующих в стенозированных или частично закупоренных артериях или при артериальных бифуркациях (см. работу Питерсона "Необходимость воздействия фактора фон Виллебранда и тромбоцитных мембранных гликопротеидов Ib и II - IIIa при тромбоцитной агрегации, индуцированной касательными силами" (Peterson et al., Blood 2: 625 - 628 (1987), Shear-induced platelet aggregation reguires von Willebrand Fact or and platelet membrane glycoproteins Ib and III - IIIa)).
5. Полипептид VCL описанного выше состава может быть использован для предотвращения возникновения тромбоза и реокклюзии после проведения пластической операции на сосудах и тромболиза, вызванных тромбиновой активацией тромбоцитов (см. работу Фастера "Антитромботическая терапия после миокардиальной реперфузии при остром инфаркте миокарда" (Fuster et al., J, Am. Coll. Cardiol. 12: 78A-84A (1988), Antithrombotic therapy after myocardial reperfusion in acute myocardial infarction).
6. Полипептид VCL описанного выше состава может быть использован для предотвращения тромбоцитной адгезии и агрегации на протезных материалах (см. работу Бейдимона "Взаимодействие тромбоцитов с протезными материалами - роль фактора фон Виллебранда во взаимодействии тромбоцитов с политетрафторэтиленом" (Badimon et al., J. of Biomaterials Applications, 5: 27-48 (1990), Platelet interaction to prostetic materials - role of von Willebrand Factor in Platelet Interaction to PTFE))
7. Полипептид VCL описанного выше состава может быть использован для предотвращения интрамиокардиальной тромбоцитной агрегации у пациентов с нестабильной стенокардией (см. работу Дейвиса "Интрамиокардиальная тромбоцитная агрегация у пациентов с нестабильной стенокардией, страдающих внезапным ишемическим кардиальным омертвением" (Davies et al., Circulation 73: 418-427 (1986), Intramyocardial platelet aggregation in patients with unstable angina suffering sudden is chemic cardiac death)).
8. Полипептид VCL описанного выше состава может быть использован для предотвращения возникновения вазоспазмы и вазоконстрикции после артериального нарушения, вызванного проведением пластической операции на сосудах, тромболизом или иными причинами (см. работу Лама "Связан ли вазоспазм с осаждением тромбоцитов? " (Lam et al., Circulation 75: 243-248 (1987), is vasospasm related to platelet deposition?)).
9. Полипептид VCL описанного выше состава может быть использован для предотвращения возникновения вазоокклюзивных кризов при серповидноклеточной анемии (см. работу Уика "Способность необычайно больших мультимеров фон Виллебранда повышать адгезию дрепаноцитов к человеческим эндотелиальным клеткам в условиях управляемого течения". (Wick et al., J. Clin. Invest. 80: 905-910 (1987), Unusually large von Willenbrand multimers increase adhesion of sickle erythrocytes to human endothelial cell under controlled flow)).
10. Полипептид VCL описанного выше состава может быть использован для предотвращения возникновения тромбоза, вызванного протеканием воспалительного процесса (см. работу Эзмона "Регуляция путей движения природного антикоагулянта" (Esmon, Science 235: 1348-1352 (1987), The regulation of natural anticoagulant pathways)).
11. Полипептид VCL описанного выше состава может быть использован для предотвращения появления артериосклероза (см. работу Фастера "Артериосклероз у обычных свиней и свиней фон Виллебранда" (Fuster et al., Circulation Res. 51: 587-593 (1982), Arteriosclerosis in normal and vao Willebrand pigs)).
12. Полипептид VCL описанного выше состава может быть использован в качестве антиметастатического вещества (см. работу Китагава "Участие тромбоцитного мембранного гликопротеида Ib и IIb/III-a-комплекса в зависимой от тромбина и независимой от тромбина агрегации тромбоцитов, индуцированной раковыми клетками" (Kitagawa et al., Cancer Res. 49: 537-541 (1989), Involvemen of platelet membrane glycoprotein Ib and IIb/IIIa complex in thrombin-dependent and independent platelet aggregation induced by tumor cells)).
Пример 8. 1. Исследования in vitro.
При проведении этих исследований полипептид VCL или полипептид управления носителем готовили свежим в стерильной воде (штамм при концентрации 2,2 мг/мл).
А. Агрегация тромбоцитов (богатая тромбоцитная плазма).
Эксперимент представляет собой тест на зависимую от фактора фонд Виллебранда vWF агрегацию, который проводят с использованием человеческой или крысиной плазмы с высоким содержанием тромбоцитов. Добавление в разных концентрациях нефракционированного яда Bothrops jararaca (BJV), который содержит ботроцетин и еще тромбиноподобный компонент, или ристоцетин, ведет к возникновению агрегации при отсутствии какого-либо иного вещества. При использовании в качестве агониста ристоцетина (1,5 мг/мл) полипептид VCL, взятый в концентрации 43 мкг/мл, устраняет агрегацию в человеческой плазме, богатой тромбоцитами. При концентрациях ристоцетина, доходящих до 5,0 мг/мл, не происходит заметная агрегация в крысиной плазме, богатой тромбоцитами. При использовании этой тест-системы с применением яда BJV в качестве агониста полипептид VCL, взятый в концентрации 83 мкг/мл, несколько ингибирует ответную реакцию у человеческой плазмы, богатой тромбоцитами (фиг. 27), но не ингибирует у крысиной плазмы, богатой тромбоцитами (фиг. 20).
Был сделан вывод, что ристоцетин не является подходящим агонистом для индуцирования vWF-зависимой агрегации у крысы. Кроме того, нельзя проконтролировать эффекты от воздействия полипептида VCL ex vivo, прибегая к использованию BJV-индуцирования агрегации в крысиной плазме, богатой тромбоцитами. Однако полипептид VCL эффективно ингибирует vWF-зависимую агрегацию в человеческой плазме с большим содержанием тромбоцитов in vitro.
B. Тромбоцитный тромбиновый рецепторный тест.
В этом тесте оценивают ингибирование индуцированной тромбином тромбоцитной прокоагулянтной экспрессии, и он кратко описан ниже. Человеческие промытые тромбоциты инкубируют в буферной смеси, которая содержит CaCl2, фактор Xa, протромбин и человеческий альфа-тромбин, что делают в течение 60 мин при 28oC. По истечении этого времени аликвоту переносят в буферную смесь, содержащую S 2238 и этилендиаминтетрауксусную кислоту (для устранения всякой последующей тромбиновой генерации). Взаимодействие вещества S 2238 останавливают по истечении 15 мин при комнатной температуре уксусной кислотой, и измеряют спектральную поглощательную способность на длине волны 405 нм. Количество S 2238-расщепленного вещества, прямо связанное с количеством добавленного человеческого альфа-тромбина, оценивают, проводя контрольный опыт без использования протромбина; и эту величину вычитают из всех результатов. Полипептид VCL тестировали этим способом при конечной концентрации 0,1 мг/мл.
Этот тест является чувствительным как к тромбоцитам ингибиторам, так и к тромбоцитным рецепторным антагонистам. В присутствии полипептида VCL тромбиновая генерация составляла 114% от контрольной величины (n = 2).
Был, следовательно, сделан вывод, что полипептид VCL не является тромбиновым рецепторным антагонистом в этой системе.
2. Исследования in vitro.
Модель артериального тромбоза (крыса).
Этот способ представляет собой существенную модификацию модели, предложенной Шандом и др. (Shand et al., Thromb, Res. 45 505-515 (1987)). Способ, обычно используемый заявителями, описан ниже.
Крыс метили тромбоцитами с индием-III и фибриногеном с иодом-125. На 1 мин пережимали дорсальную аорту, используя модифицированный зажим Спенсера-Уэллса (Spencer-Wells). По прошествии 45 мин реперфузии поврежденный сосуд удаляли, промывали в цитрате и производили счет. Результаты выражали в единицах, эквивалентных миллиграмма крови. Находили разность в радиологическом накоплении между плацебо и животными, обработанными лекарственным препаратом, и ее выражали в виде процентного ингибирования.
С целью оценки пути введения полипептида VCL была произведена болюсная внутривенная инъекция. Полипептид VCL использовали в дозах 2 мг/кг (n = 5) и 4 мг/кг (n = 3). Препарат вводили за 1 мин до пережимания. Сосуд затем подвергали реперфузии в течение 20 мин. Антитромботический эффект оценивали в конце 20-минутной реперфузии. С целью сохранения соединения прибегали к сокращению времени реперфузии (в сравнении с обычным временем). Оценивали надлежащую управляющую способность носителя (при n = 5 в случае обеих доз).
Можно видеть, что в этих условиях полипептид VCL ингибирует образование тромба в этой модели (табл. 4). Ингибирование просматривается в случае тромбоцитных компонентов (с индием-III) тромба при дозе в 4 мг/кг. Другие изменения не носят статистического смысла; таким образом, полипептид VCL обнаруживает антитромботическую эффективность в этой крысиной артериальной модели.
Следует сказать, что полипептид VCL проявляет антитромботический эффект в этой крысиной модели артериального тромбоза, который может оказаться независящим от дозы.
3. Обсуждение. Результаты табл. 4 выражены в виде среднего процентного ингибирования с прибавленной или вычтенной стандартной ошибкой. В табл. 4 указано число экспериментов, проведенных в обработанных группах; и все случаи сопоставлены с группой из пяти контрольных животных. Статистический анализ проведен на первичных данных до их преобразования в процентное ингибирование. Обозначение NS означает, что статистический смысл отсутствует.
Из представленных данных следует, что полипептид VCL взаимодействует с человеческим тробоцитом vWF-рецептором, и, следовательно, ингибирует тромбоцитную агрегацию в человеческой плазме, богатой тромбоцитами. Наблюдается, однако, заметное различие между образованиями (крыса в сравнении с человеком), когда сравнивают ингибирование тромбоцитной агрегации. Не проводили в этом эффекте и действующего механизма. В практическом отношении это означает, что не удалось осуществить анализ образцов ex vivo с целью установления корреляции эффектов от воздействия полипептида VCL на агрегацию и артериальный тромбоз. Следовательно, анализ и интерпретация эффективности действия полипептида VCL in vivo, используемого в качестве антитромботического средства в модели артериального тромбоза у крыс, осложняется этим фактором.
Несмотря на наличие в GPIb-области связующих сайтов как у vWF-фактора, так и у тромбина, складывается впечатление, что любые воздействия полипептида VCL на связывание тромбина с гликопротеидом GPIb не транслируются в антагонизм у индуцированной тромбином прокоагулянтной экспрессии.
Полный полипептид VCL обнаруживает антитромботический эффект в крысиной модели артериального тромбоза. Это ингибирование может быть обусловлено его вмешательством в связывание vWF-фактора с его рецептом.
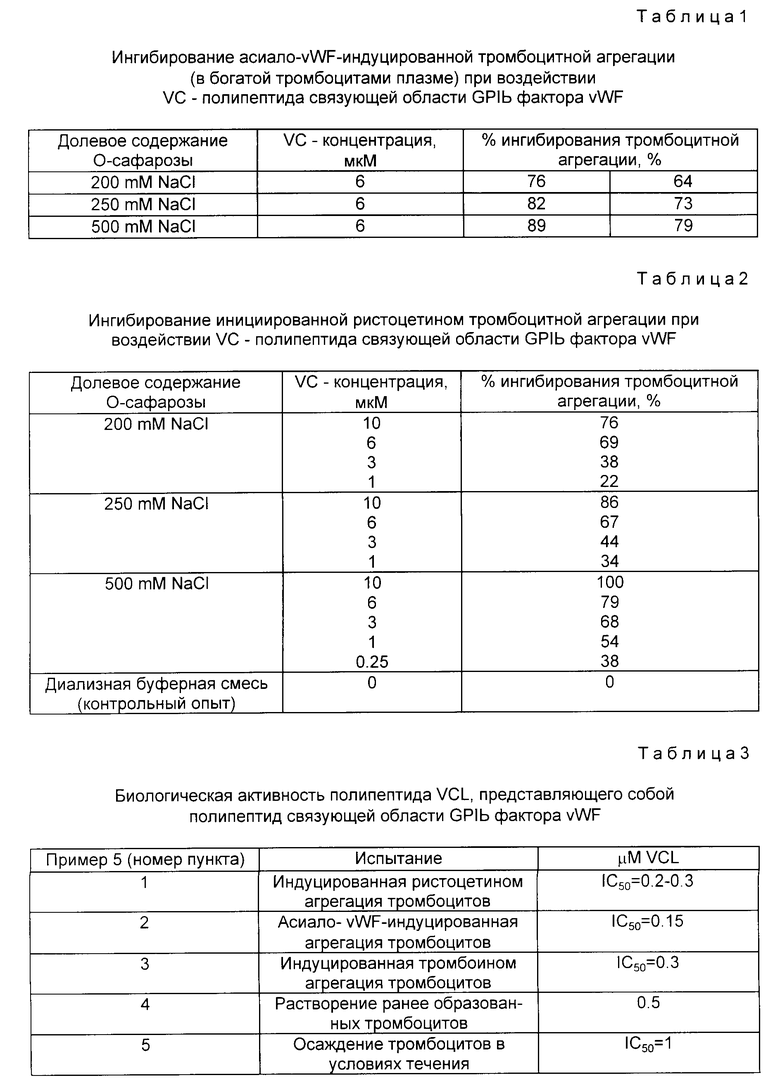

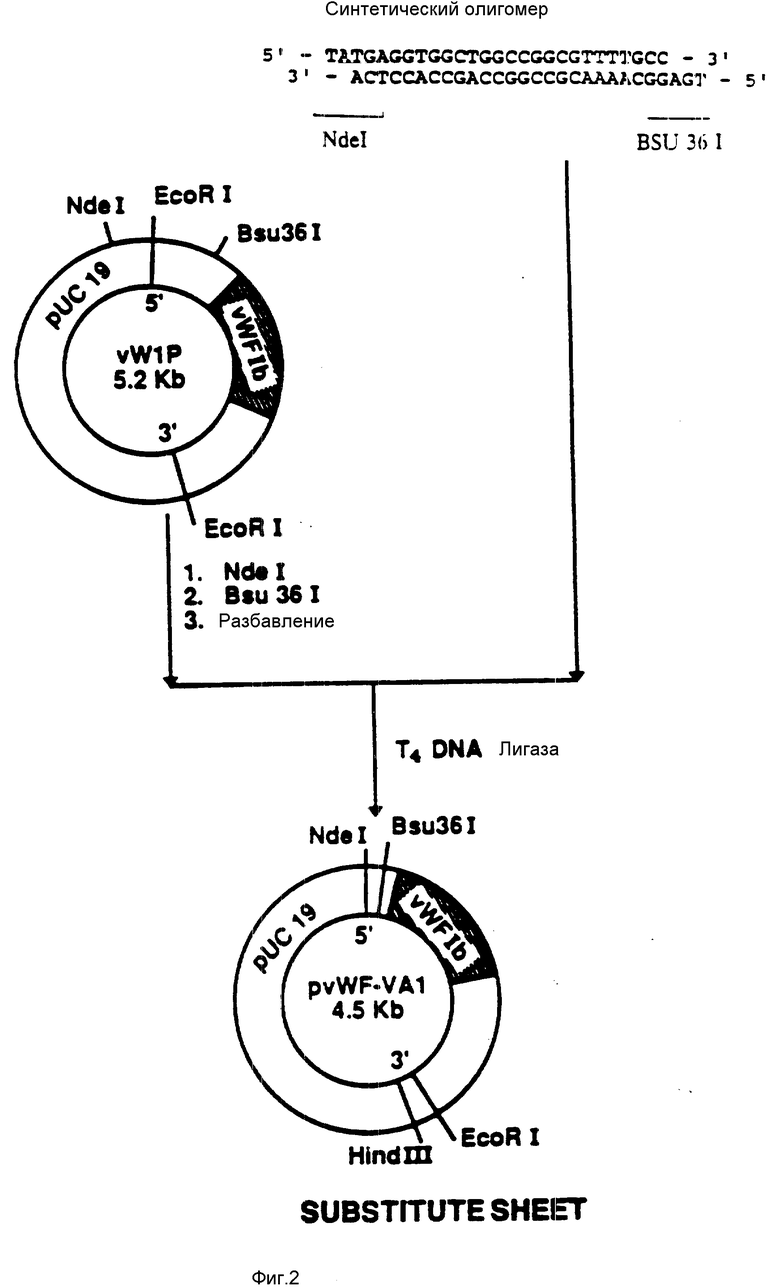
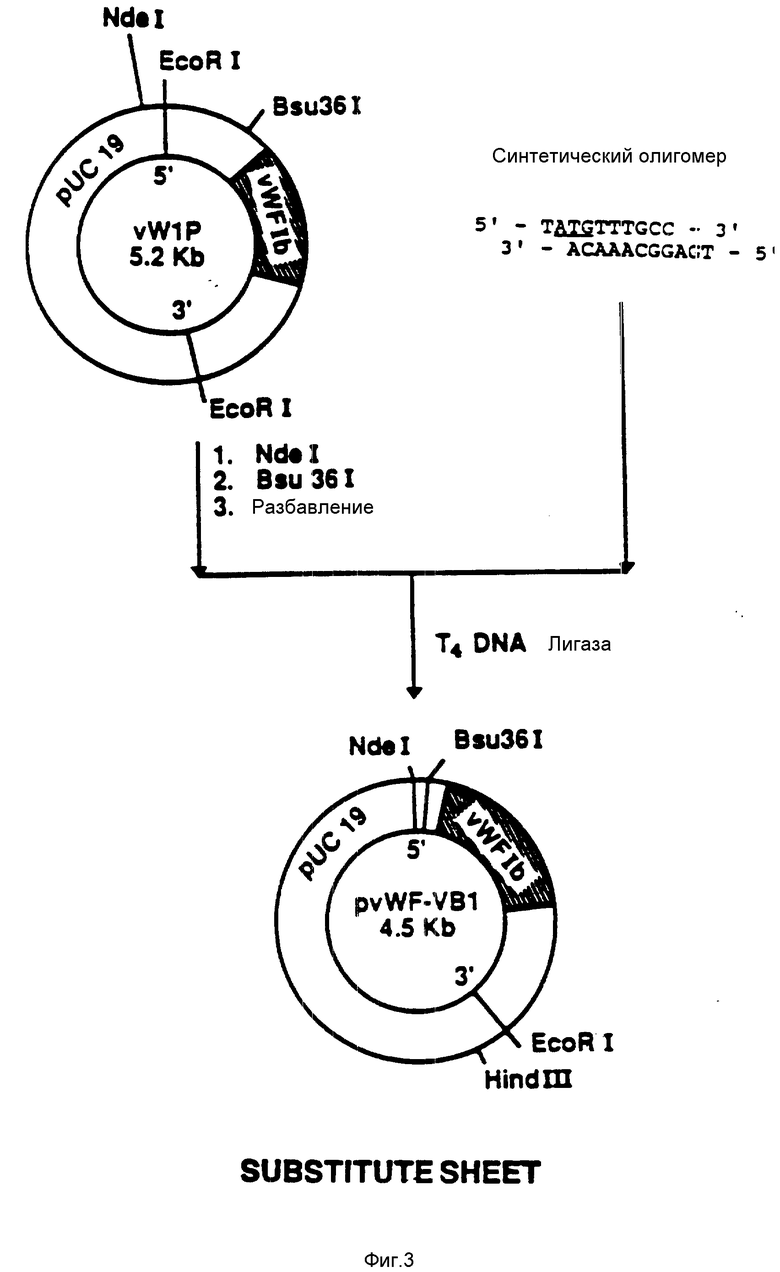
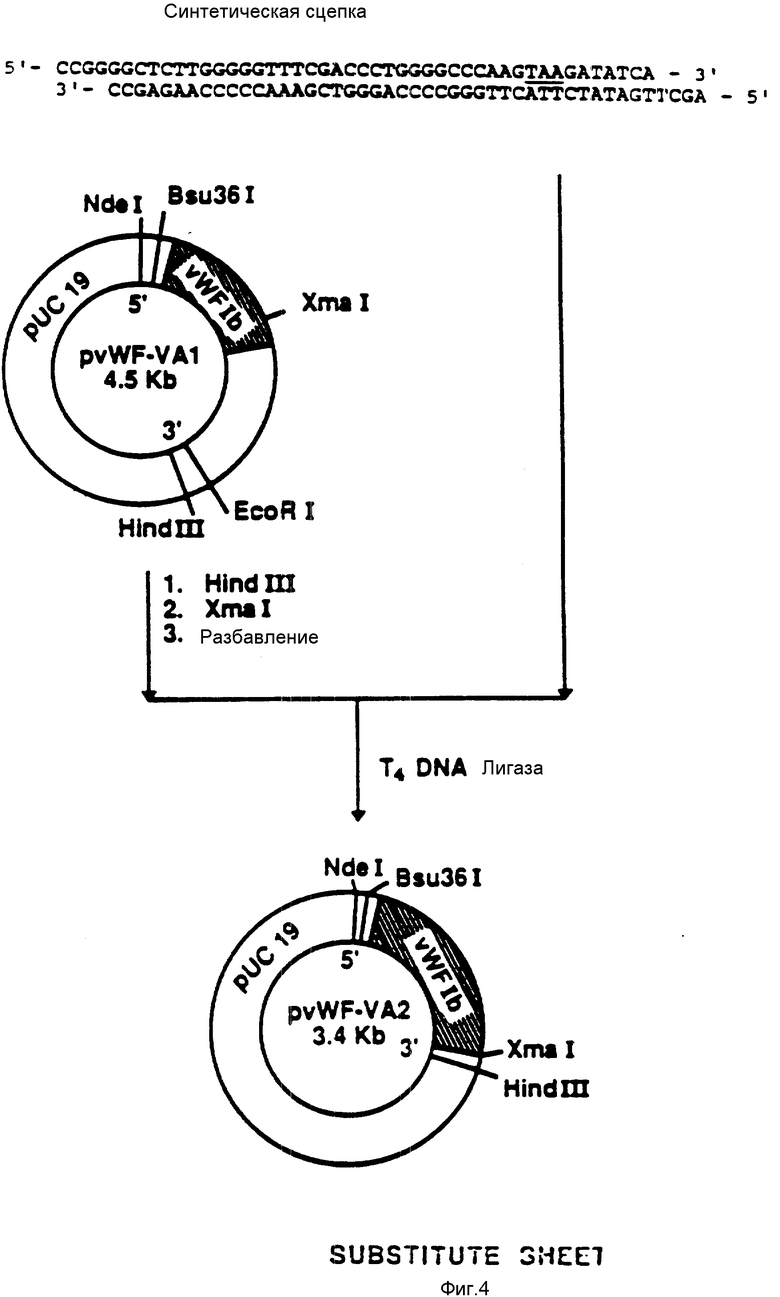
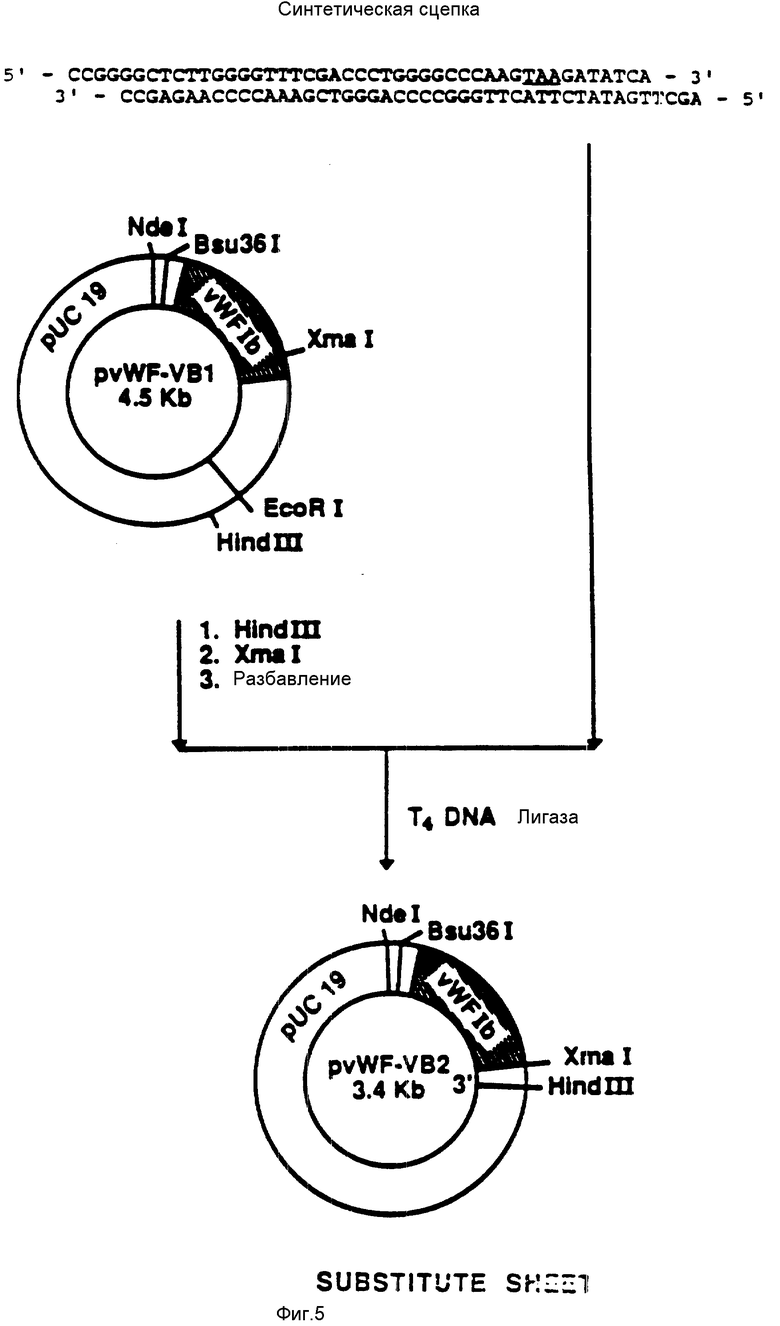
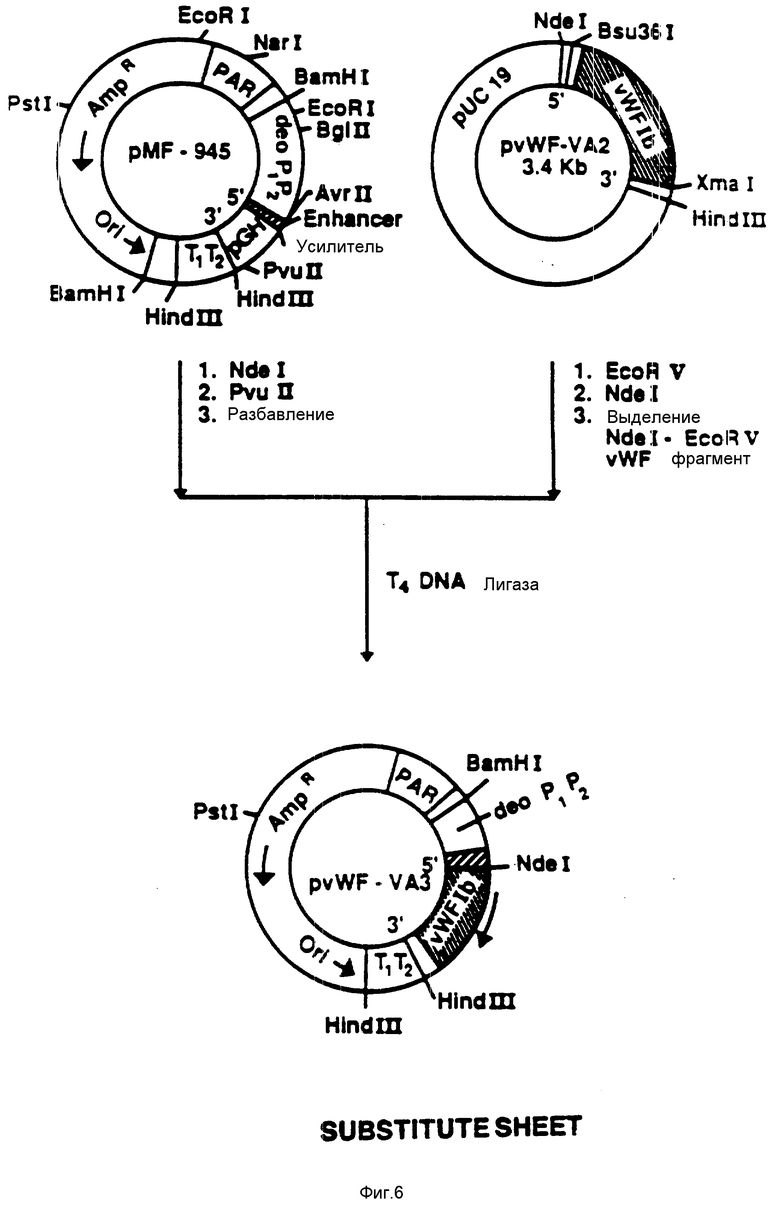
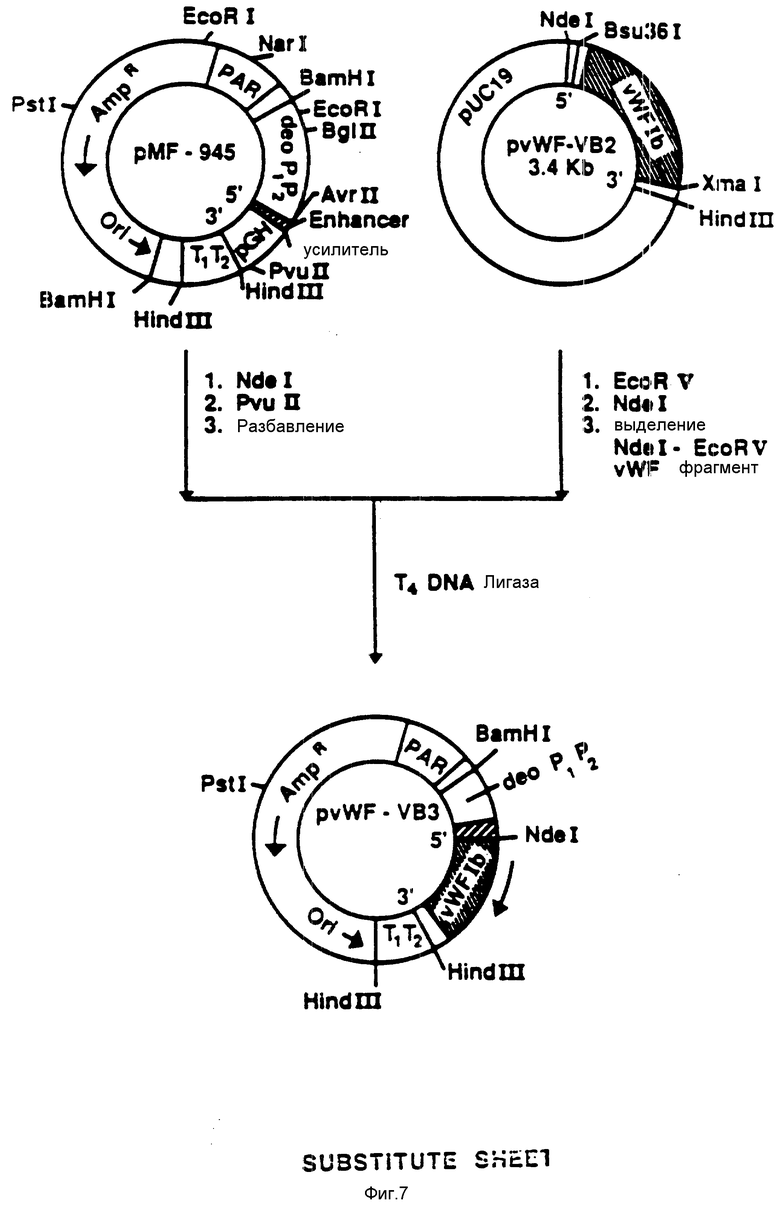
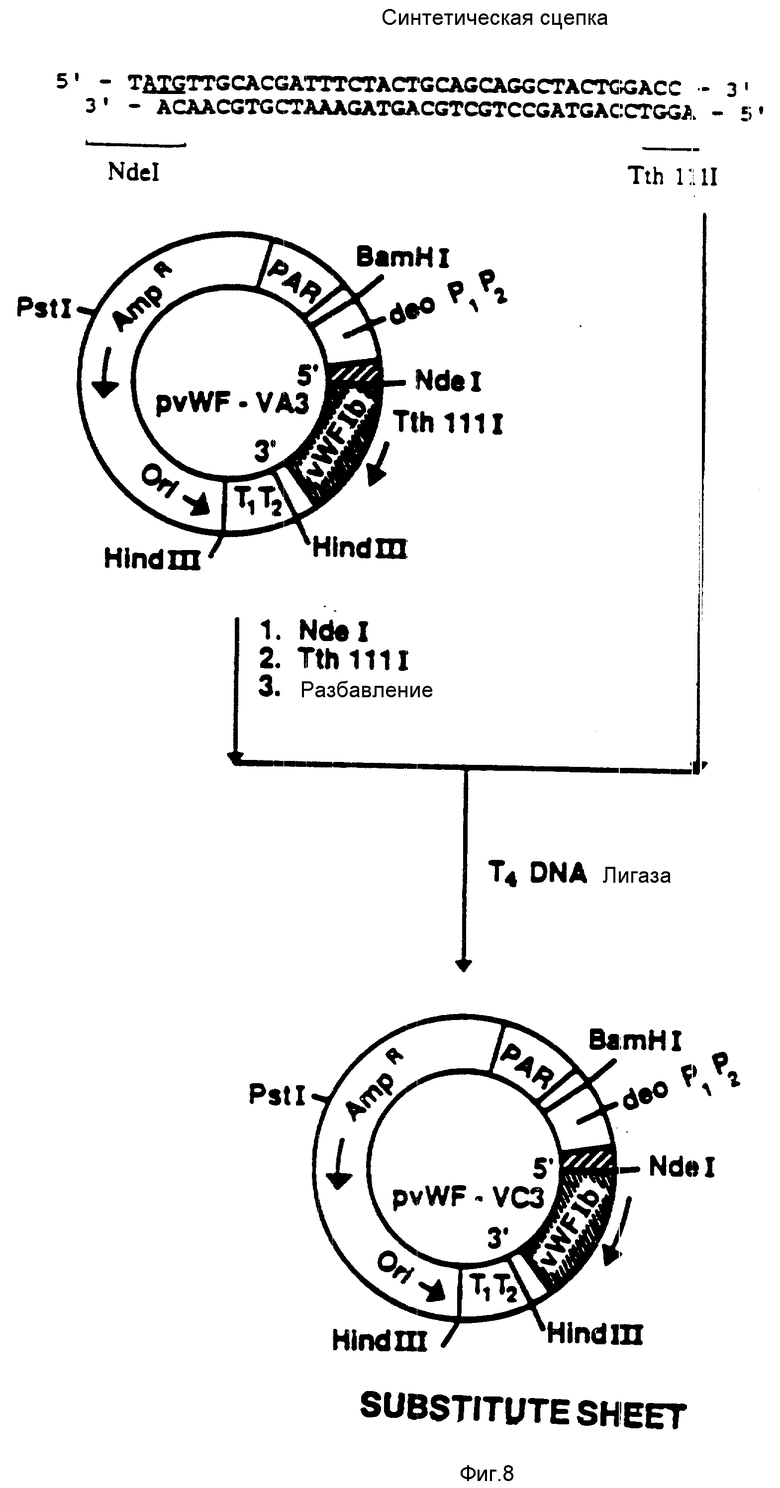
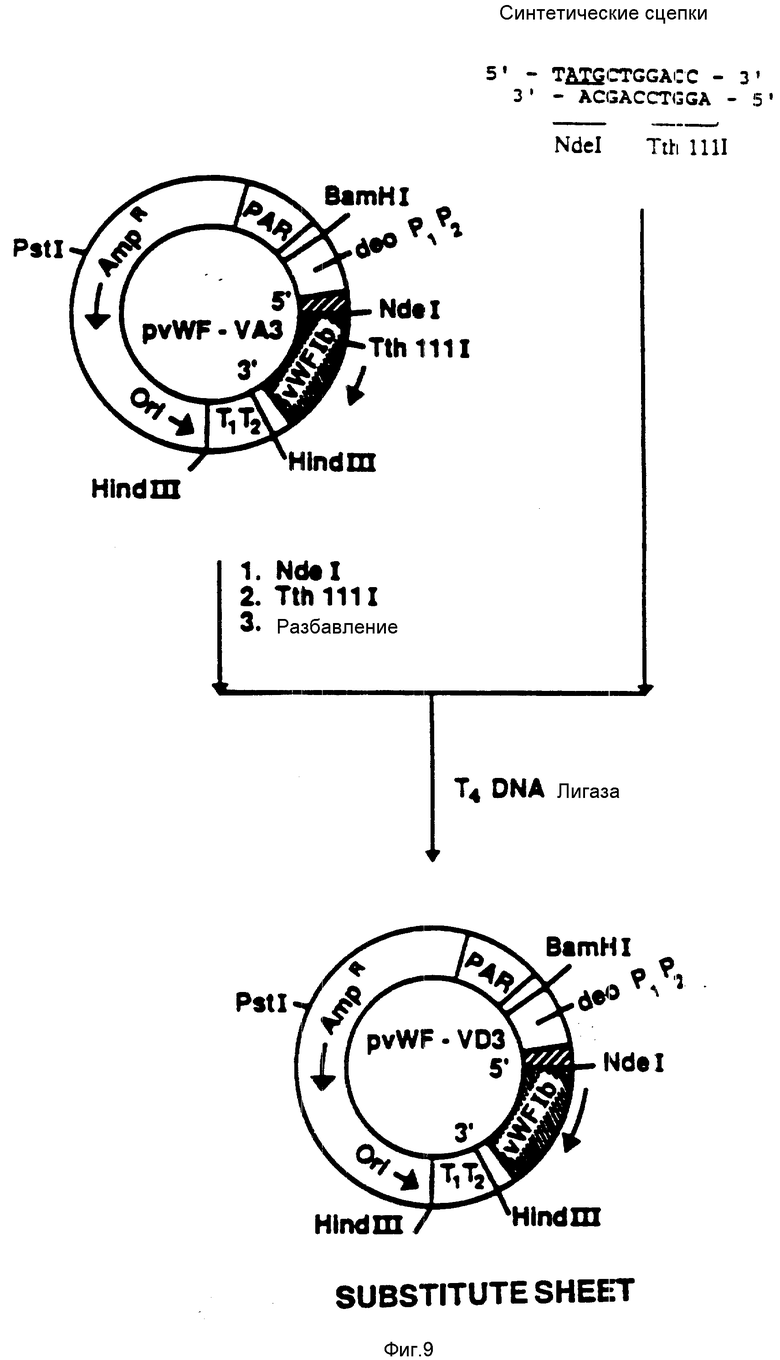
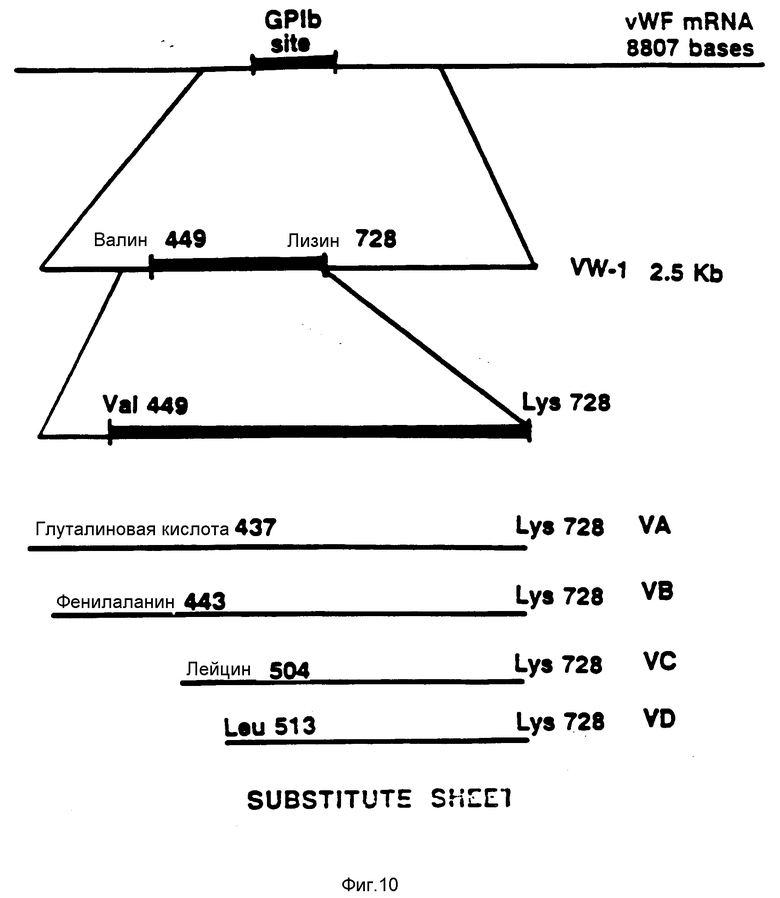
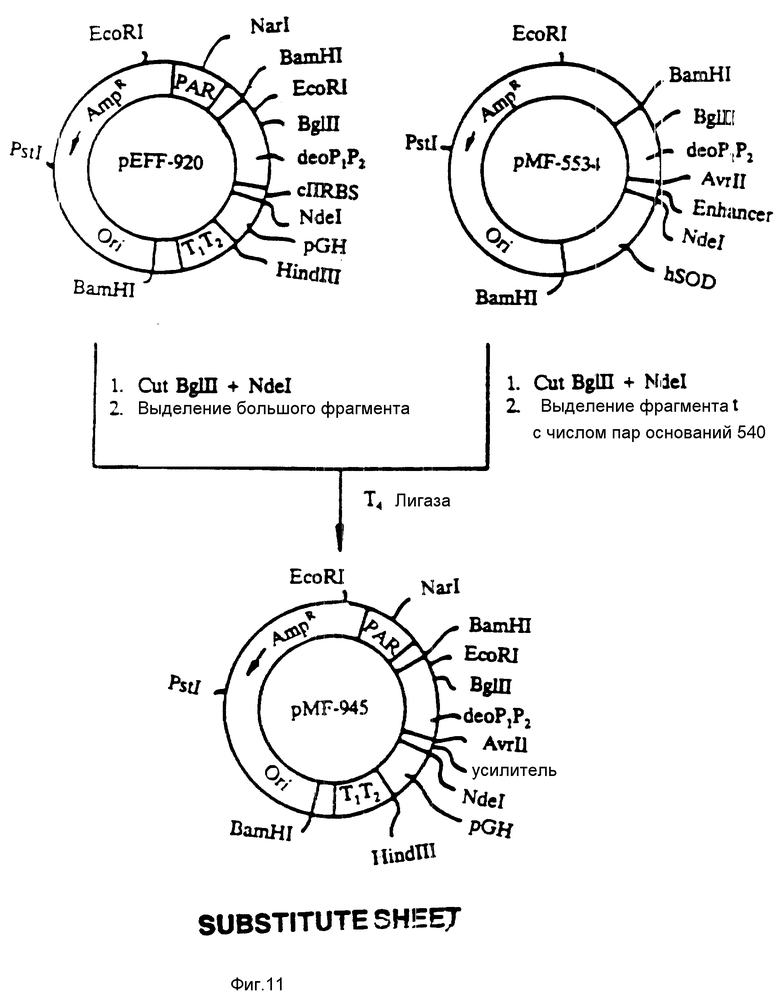









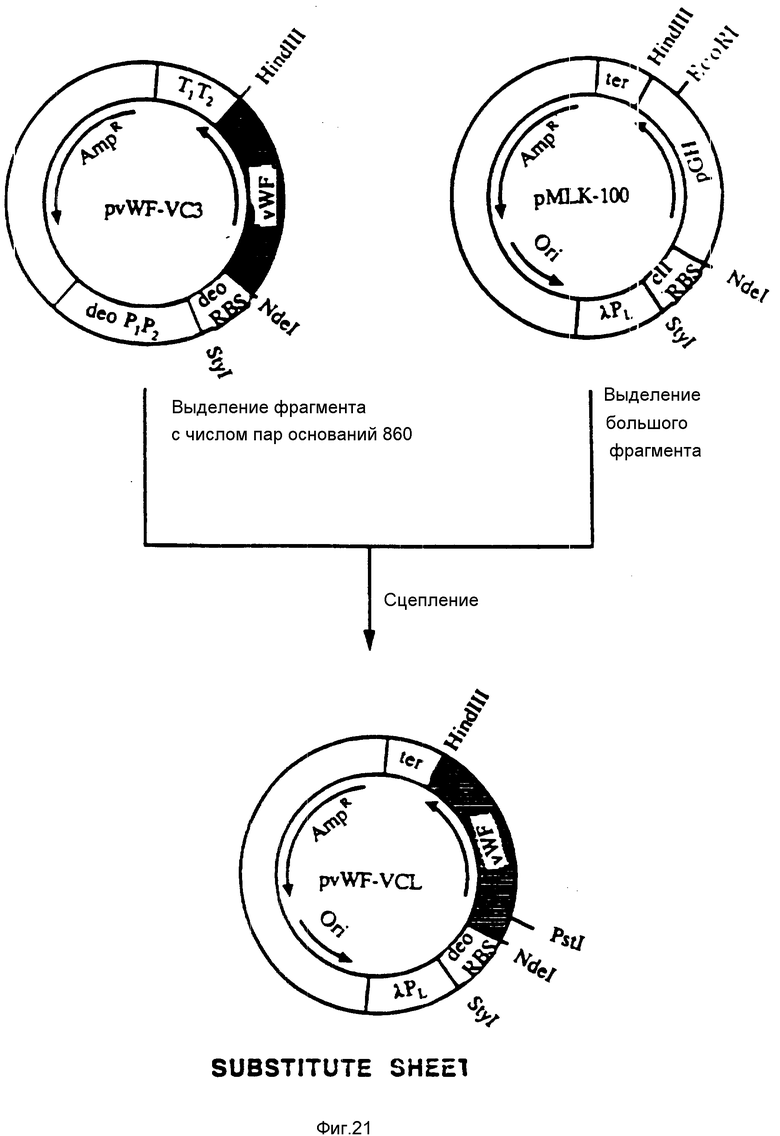
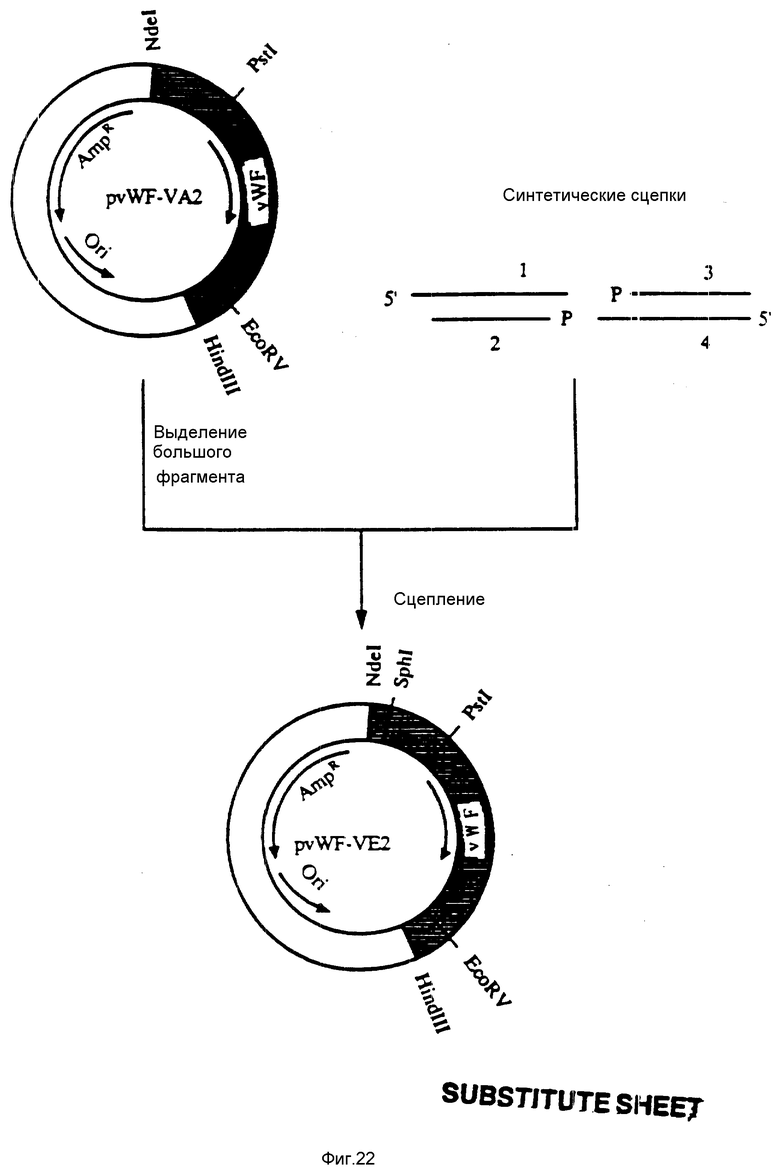
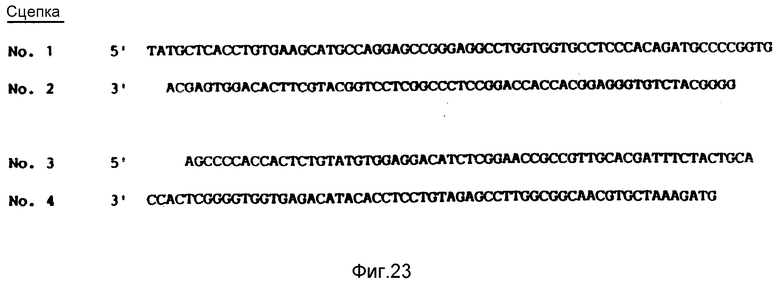
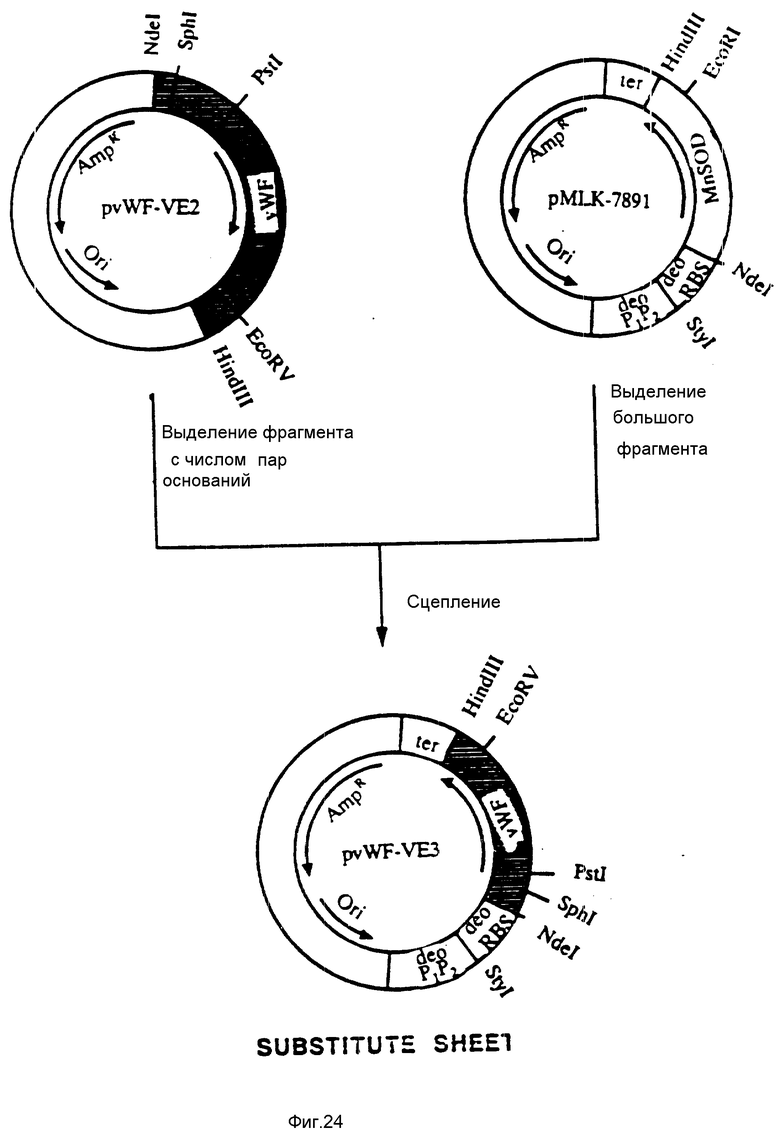
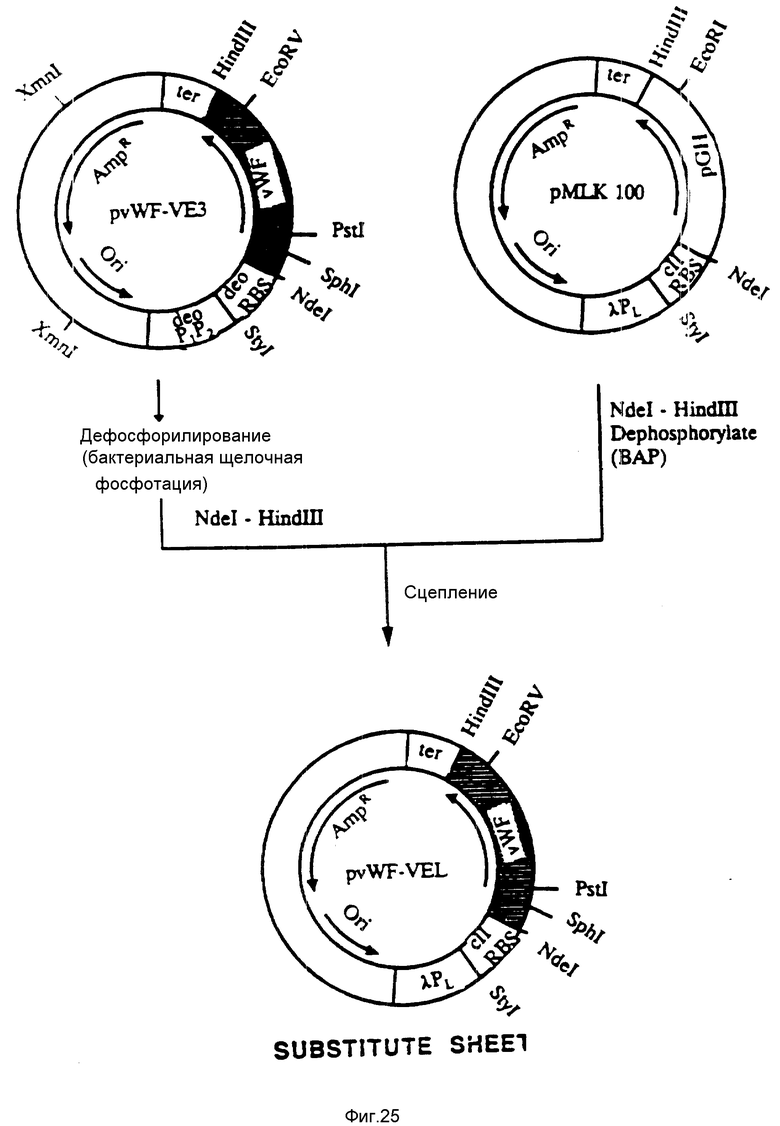
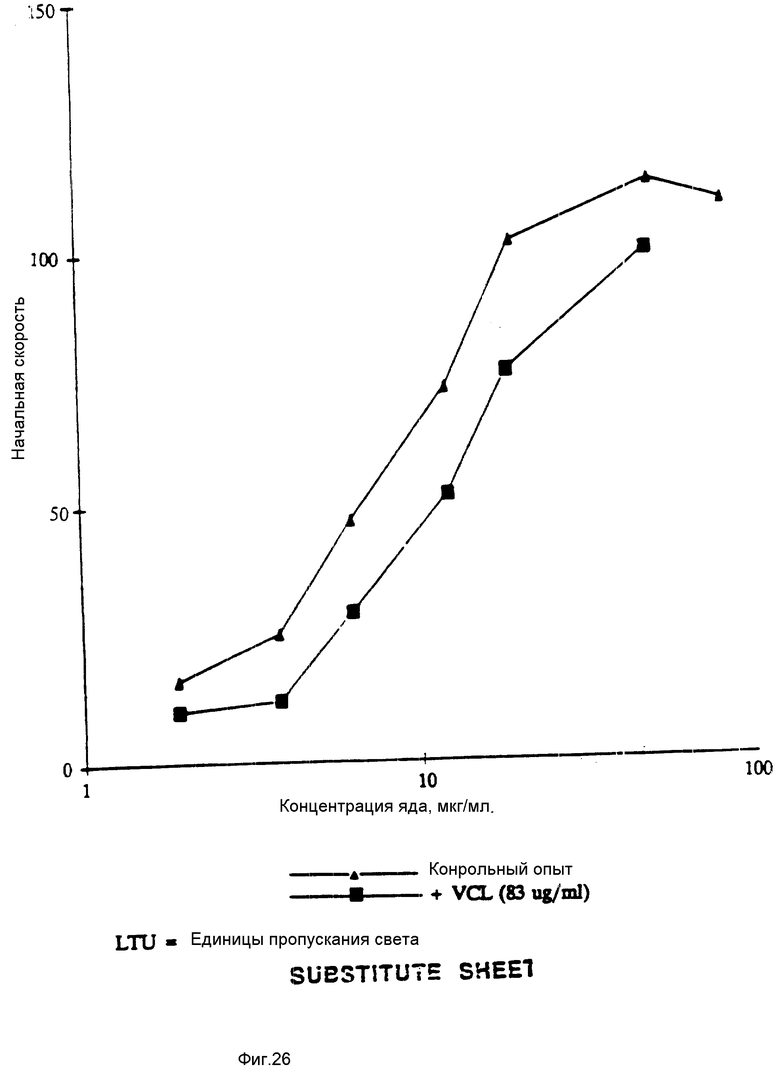
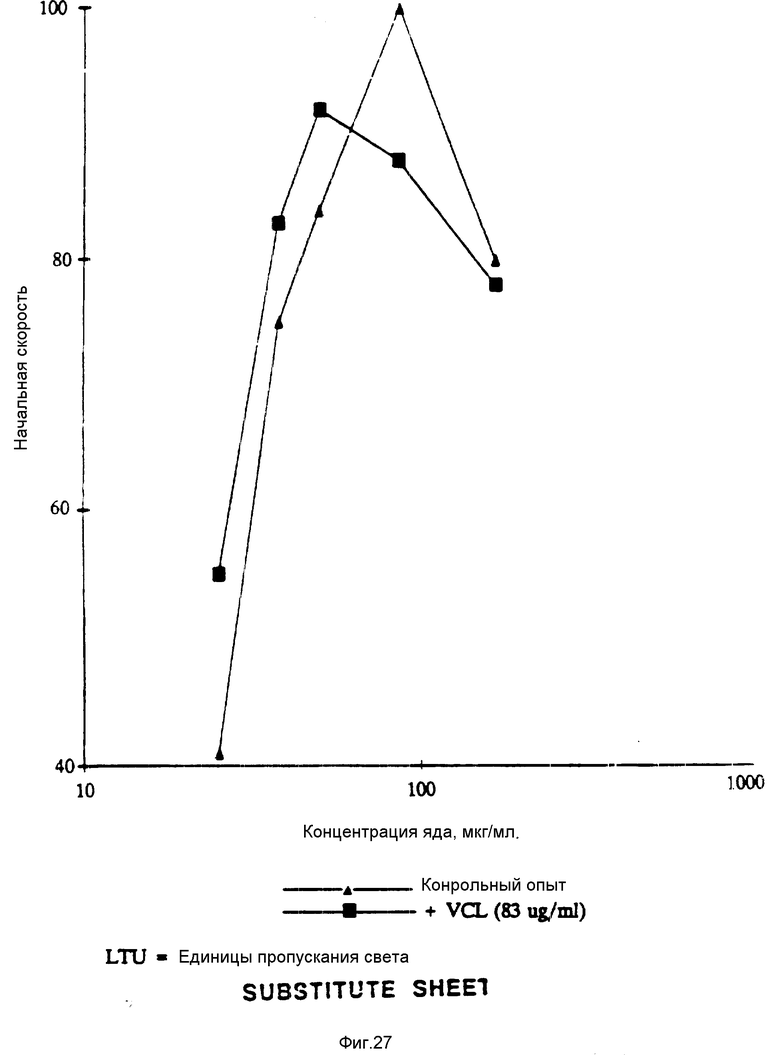
Изобретение относится к биотехнологии и позволяет получить рекомбинантный полипептид, ингибирующий слипание и агрегацию тромбоцитов при воздействии таких условий, при которых проявляются церебровескулярные расстройства и сердечно-сосудистые расстройства. Рекомбинантный полипептид содержит область связывания GPIb фактора фон Виллебранда человека и имеет определенную аминокислотную последовательность. Способ ингибирования агрегации тромбоцитов осуществляют путем контактирования тромбоцитов с эффективным количеством рекомбинантного полипептида. Для экспрессии рекомбинантного полипептида используют рекомбинантную плазмидную ДНК pvWF-VC3 или pvWF-VCL. Каждая из ДНК содержит область связывания гликопротеина Ib фактора фон Виллебранда человека. Эта область включает фрагмент ДНК, кодирующий аминокислотную последовательность полипептида, и содержит промоторную систему deo P1P2 или промотор Pl фага ламбда, deo - сайт связывания рибосом, T1T2-терминатор транскрипции и ген устойчивости к ампициллину Amp2 в качестве генетического маркера. Продуцентами полипептида являются рекомбинантный штамм E • coli/pvWF-VC3 или IpvWF-VCL. При получении полипептида трансформируют штамм E • coli рекомбинантной плазмидной ДНК, проводят отбор и культивирование трансформантов в условиях, обеспечивающих накопление полипептида. При выделении биологически активного пептида разрушают бактериальные клетки с выделением лизата. Из лизата выделяют тельца включения с помощью денатурирующего агента или гуанидингидрохлорида, или мочевины с последующей очисткой раствора полипептида катионнообменной хроматографией. Растворенный полипептид обрабатывают дисульфидом или глутатионом, или тиоредоксином, или бета-меркаптоэтанолом, или цистеином. Полученный полипептид обрабатывают диализом, а диализат -катионно-обменной хроматографией. 7 с.п.ф-лы, 4 табл., 27 ил.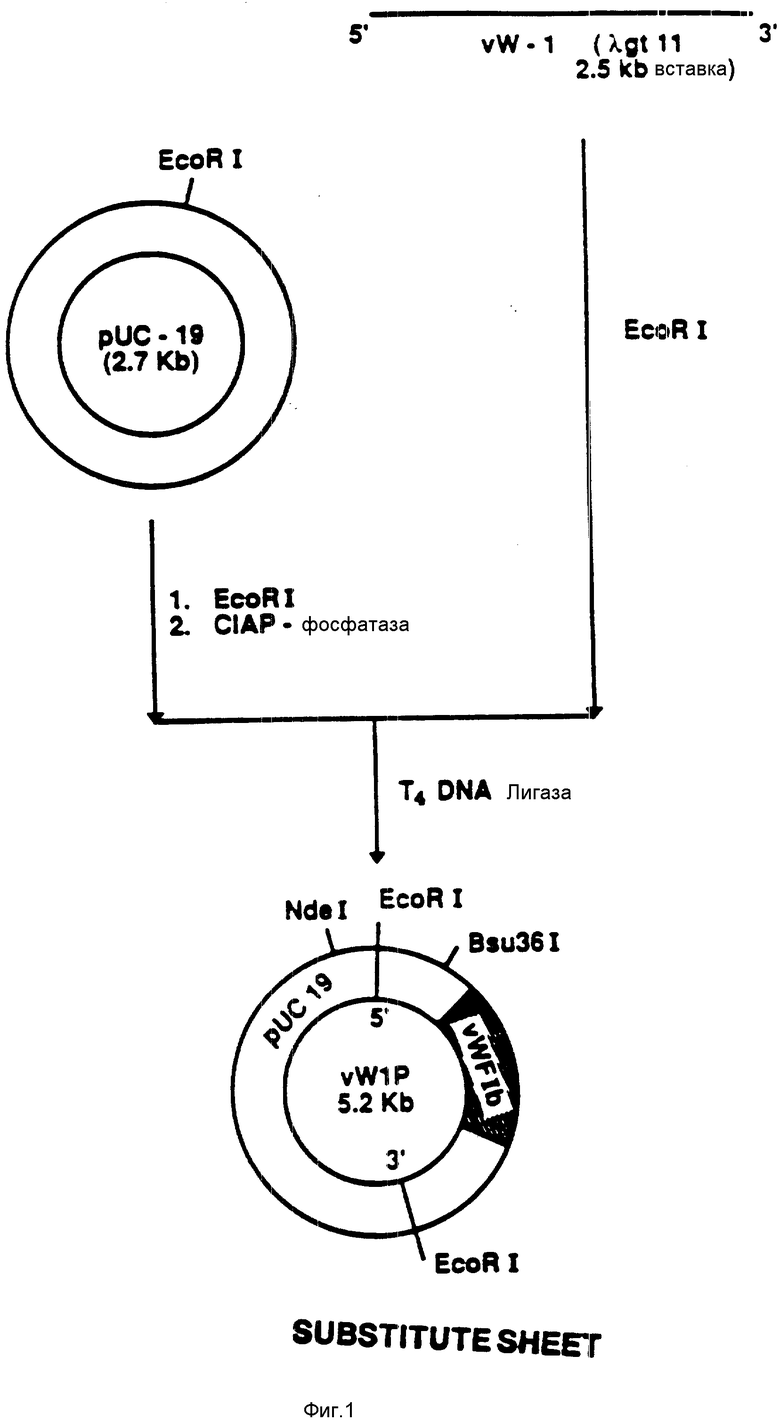
или аналогичную ей последовательность, включающую делеции, и/или инсерции, и/или замены аминокислот, не влияющие на биологическую активность, состоящую в способности рекомбинантного полипептида связывать гликопротеин Ib (GPIb) тромбоцитов.
| Gujmuna et al.journal of Biol.Chem., 261, N 1, 381-385, 1986. |
