ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение относится к гликозилированным полипептидам на основе белка млекопитающего, вызывающим пониженный иммунный ответ или повышенную иммунную толерантность благодаря модулированному связыванию SIGLEC, и к способам лечения с использованием этого гликозилированного полипептида. Кроме того, изобретение относится к белковым комплексам, вызывающим пониженный иммунный ответ или повышенную иммунную толерантность благодаря модулированному связыванию SIGLEC, и к способам лечения с использованием этих белковых комплексов.
УРОВЕНЬ ТЕХНИКИ
Гемофилия представляет собой группу наследственных генетических заболеваний, при которых нарушается способность организма контролировать свертывание крови или коагуляцию. При наиболее распространенной форме, гемофилии А, дефицитным является фактор свертываемости VIII (FVIII), причем гемофилия А встречается с частотой примерно 1 случай на 5000-10000 рожденных мальчиков. Белок FVIII является неоходимым кофактором свертываемости крови с множеством функций. Дефицит FVIII может быть восполнен концентратами FVIII, полученными из плазмы, или рекомбинантно продуцированным FVIII. Лечение концентратами FVIII привело к нормализации жизни пациентов с гемофилией. Исторически гемофилию лечили FVIII, получаемым из плазмы крови человека. В плазме крови при нормальных условиях молекула FVIII всегда связана со своим кофактором, фактором фон Виллебранда (vWF), который обеспечивает стабильность молекулы FVIII к различным видам деградации.
Было описано множество способов очистки фактора VIII из плазмы или культуральной массы, которая рекомбинантно продуцирует фактор VIII (rFVIII), без и с фактором фон Виллебранда. В 90-х годах первые поступившие в продажу рекомбинантные FVIII (rFVIII) подразделялись на два типа: полноразмерные молекулы rFVIII, имитирующие основную форму FVIII в плазме крови, и молекулы rFVIII с удаленным В-доменом (Eriksson et al., 2001), в которых один неактивный участок (B-домен) был удален, причем оба типа имели высокую степень чистоты (все без vWF).
Пациенты с гемофилией А получают лечение с помощью FVIII по мере необходимости или в качестве профилактической терапии несколько раз в неделю. Для профилактического лечения, FVIII в количестве 15-25 МЕ/кг массы тела вводят три раза в неделю, что необходимо из-за постоянной потребности в FVIII и его короткого времени полужизни в кровотоке, которое для человека составляет только около 11 часов. (Ewenstein et al., 2004).
Частое хроническое введение экзогенного FVIII вызывает ответную реакцию иммунной системы пациента (Saenko et al., Haemophilia 8:1-11 (2002)), что является серьезным ограничением для терапии.
В настоящее время наиболее распространенная опция для достижения иммунной толерантности у пациентов с гемофилией А (врожденной недостаточностью FVIII) и ингибиторами представляет собой индукцию иммунной толерантности (ITI), когда высокие дозы FVIII вводят в течение длительного времени. Однако лечение может занимать до двух лет, остается безуспешным приблизительно у 30% пациентов, является чрезвычайно дорогостоящим и не может быть использовано в профилактических целях для подавления первичного возникновения ингибирующих антител.
Поэтому, необходимы подходы для ослабления иммунного ответа. Одним из перспективных подходов является оптимизация гликозилирования либо FVIII, либо его связывающего партнера - vWF.
Например, в WO 2014/176125 A1 описаны иммунные конъюгаты для индукции антиген-специфичной иммунной толерантности к FVIII. Иммунными конъюгатами являются белки FVIII, конъюгированные с определенными гликановыми лигандами, мишенью которых являются молекулы SIGLEC, экспрессирующиеся на В-клетках, а именно SIG-1 или SIG-10 (или ортолог SIG-G). Гликаны-лиганды, в частности, связаны с липосомами, в которые вводится FVIII.
Связывающие сиаловую кислоту иммуноглобулиновые лектины (SIGLEC) представляют собой семейство из 15 человеческих и 9 мышиных рецепторов на клеточной поверхности, которые экспрессируются на различных белых кровяных клетках иммунной системы за исключением большинства Т-клеток у мышей и у человека. SIGLEC локализованы на различных типах клеток и связываются с различными гликанными структурами (см. обзор в Paulson et al. 2012). Например, было продемонстрировано связывание vWF и FVIII с SIG-5 (Pegon 2012). Однако механизм связывания остается неизвестным.
Другой подход описан в WO 2014/179184 А1. Авторы предполагают снижение нежелательного гуморального иммунного ответа и индукцию иммунной толерантности в отношении факторов свертываемости крови, таких как FVIII, путем добавления SIGLEC-лигандов. SIGLEC-лиганды выбирают из 9-N-бифенилкарбоксил-NeuAca2-6Gal~l-4GlcNAc (6'-BPCNeuAc), NeuAca2-6Galwl-4GlcNAc и NeuAca2-6Galwl-4(6-сульфо)GlcNAc. SIGLEC-лиганд связан с фактором свертываемости через водорастворимый полимер.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Настоящее изобретение, среди прочего, основано на открытии того факта, что гликановая структура, в природе встречающаяся на плазматических белках, в частности vWF, позволяет взаимодействие с группой молекул SIGLEC, в частности SIG-5, SIG-7, SIG-8 и SIG-9. Кроме того, авторы изобретения обнаружили, что путем модификации гликановой структуры на белке может быть увеличено взаимодействие с SIGLEC, такими как SIG-5, SIG-7, SIG-8 И SIG-9. Это увеличение приводит к снижению иммунного ответа и/или повышению иммунной толерантности у пациента, которому вводят белок.
Таким образом, в соответствии с первым аспектом изобретение относится к гликозилированному полипептиду, содержащему аминокислотную последовательность, идентичную или гомологичную по меньшей мере фрагменту белка млекопитающего, предпочтительно, человека, где указанный гликозилированный полипептид содержит один или несколько сиалилированных О-гликанов, и где полипептид имеет повышенную аффинность связывания с одним или несколькими SIGLEC, выбранными из группы, состоящей из SIG-5, SIG-7, SIG-8, и SIG-9, по сравнению с белком млекопитающего или его фрагментом.
Авторы настоящего изобретения, в частности, определили гликановую структуру, которая, с одной стороны, необходима для взаимодействия с SIGLEC, но также добавление которой приводит к повышенному связыванию с SIGLEC. Ответственными за это являются сиалилированные коровые О-гликаны 2-го типа и/или удлиненные коровые О-гликаны 1-го типа.
Таким образом, гликозилированный полипептид в соответствии с первым аспектом может быть также описан как гликозилированный полипептид, содержащий аминокислотную последовательность, идентичную или гомологичную по меньшей мере фрагменту белка млекопитающего, предпочтительно, человека, где указанный гликозилированный полипептид содержит один или несколько сиалилированных О-гликанов, и где объединенное количество сиалилированных коровых O-гликанов 2-го типа и сиалилированных удлиненных коровых O-гликанов 1-го типа гликозилированного полипептида превышает объединенное количество сиалилированных коровых O-гликанов 2-го типа и сиалилированных удлиненных коровых O-гликанов 1-го типа в белке млекопитающего или его фрагменте.
Белок с гликановой композицией, включающей сиалилированные коровые O-гликаны 2-го типа и/или сиалилированные удлиненные коровые O-гликаны 1-го типа, в частности, сиалилированные коровые 2 O-гликаны, может быть использован для модификации иммунного ответа пациента на терапевтический белок при совместном введении.
Таким образом, согласно второму аспекту изобретение относится к применению гликозилированного полипептида, содержащего один или несколько сиалилированных О-гликанов и связывающегося с одним или несколькими SIGLEC, выбранными из SIG-5, SIG-7, SIG-8, и SIG-9, для снижения иммунного ответа или повышения иммунной толерантности пациента к терапевтическому белку.
Используя модификацию гликозилированного полипептида, аффинно связывающегося с SIGLEC, можно не только напрямую модифицировать аффинность связывания с SIGLEC у самого модифицированного полипептида, но также и у белкового комплекса или композиции, частью которых является гликозилированный полипептид, например, комплекса фактора VIII (FVIII) и фактора фон Виллебранда (vWF).
Таким образом, согласно третьему аспекту настоящее изобретение относится к белковой композиции, содержащей первый и второй полипептиды, где первый полипептид представляет собой гликозилированный полипептид, содержащий один или несколько сиалилированных О-гликанов, и второй полипептид содержит аминокислотную последовательность, гомологичную или идентичную второму белку млекопитающего, в частности, человека, где (по сравнению со вторым полипептидом) композиция имеет повышенную аффинность связывания с одним или несколькими SIGLEC, выбранными из SIG-5, SIG-7, SIG-8, и SIG-9. Предпочтительно, что первый и второй полипептид композиции согласно третьему аспекту образуют белковый комплекс.
Согласно четвертому аспекту настоящее изобретение относится к выделенному полинуклеотиду, который содержит последовательность нуклеиновой кислоты, кодирующую гликозилированный полипептид по первому аспекту изобретения. В пятом аспекте изобретение также относится к экспрессионному вектору, содержащему полинуклеотид по четвертому аспекту изобретения.
Гликозилированный полипептид по первому аспекту, в частности vWF или FVIII, и композиция по третьему аспекту, в частности, комплекс FVIII и vWF, благодаря сниженному иммунному ответу, являются особенно полезными в терапии.
Таким образом, соглано четвертому аспекту, настоящее изобретение относится к гликозилированному полипептиду по первому аспекту или к композиции по третьему аспекту для применения в лечении или профилактике нарушений свертываемости крови.
ФИГУРЫ
На фиг.1 показаны результаты теста связывания vWF с различными SIGLEC. Поглощение при 492 нм пропорционально количеству vWF, связавшемуся с соответственно указанными SIGLEC или контролями. SIG-2, SIG-5, SIG-7, SIG-F, SIG-9 и SIG-10 иммобилизовали с помощью белка А на микротитровочном планшете при концентрации 500 нг/лунка. Биотинилированный vWF добавляли в концентрации от 0 до 0,8 мкг/мл, и после промывания связывание визуализировали с использованием конъюгированного с HRP стрептавидина и измеряя поглощение при 492 нм. Для контроля использовали анти-vWF и антитела к куриным IgY.
На фиг.2 схематически представлена структура домена vWF, включающая сайты N- и О-гликозилирования, расщепления протеазой V8, и фрагменты, полученные после расщепления протеазой V8.
На фиг.3 показаны результаты теста связывания N-концевого и C-концевого фрагмента vWF с SIGLEC - SIG-5, SIG-7, SIG-F и SIG-9. Оптическая плотность при 492 нм пропорциональна количеству фрагмента vWF, связавшегося с соответственно указанными SIGLEC или контролем. SIG-5, SIG-7, SIG-F И SIG-9 иммобилизовали с помощью белка А на микротитровочном планшете при концентрации 500 нг/лунка. Биотинилированный N-концевой VWF-фрагмент (темно-серые столбики) и С-концевой VWF-фрагмент (светло-серые столбики)) добавляли в концентрации 1 мкг/мл, и после промывания связывание визуализировали с использованием конъюгированного с HRP стрептавидина и измеряя поглощение при 492 нм. В качестве отрицательного контроля использовали антитела к куриным IgY.
На фиг.4 показаны результаты теста на связывание N-концевого фрагмента vWF в десиалилилированной, де-N-гликозилированной и необработанной форме. Поглощение при 492 нм пропорционально количеству N-концевого фрагмента vWF, связавшегося с соответственно указанным SIGLEC или контролем. N-концевой фрагмент VWF до расщепления представлен белыми столбиками, де-N-гликозилированный ферментом PNGaseF фрагмент представлен серыми столбиками, и десиалилированный фрагмент представлен черными столбиками. SIG-5, SIG-7, SIG-F И SIG-9 иммобилизовали с помощью белка А на микротитровочном планшете при концентрации 500 нг/лунка. Биотинилированные N-концевые фрагменты vWF (до расщепления, расщепленные фермнентами PNGaseF или Сиалидазой A) добавляли в концентрации 8 мкг/мл, и после промывания связывание визуализировали с использованием конъюгированного с HRP стрептавидина и измеряя поглощение при 492 нм. В качестве отрицательного контроля использовали антитела к куриным IgY.
На фиг. 5 показаны результаты теста на связывание кластера I и кластера II O-гликозилирования с SIGLEC. Поглощение при 492 нм пропорционально количеству фрагмента кластера I (светло-серые столбики) или фрагмента кластера II (темно-серые столбики), связавшегося с соответственно указанным SIGLEC или контролем. SIG-5, SIG-7, SIG-F, SIG-9 и SIG-10 иммобилизовали с помощью белка А на микротитровочном планшете при концентрации 500 нг/лунка. Биотинилированный кластер I и кластер II добавляли в концентрации 4 мкг/мл, и после промывания связывание визуализировали с использованием конъюгированного с HRP стрептавидина и измеряя поглощение при 492 нм. В качестве отрицательного контроля использовали антитела к куриным IgY.
На фиг.6 показаны результаты теста на связывание кластера II О-гликозилирования с SIGLEC до и после обработки сиалидазой А. Поглощение при 492 нм пропорционально количеству необработанного фрагмента кластера II (светло-серые столбики) или фрагмента кластера II, расщепленного Сиалидазой А (темно-серые столбики), связавшегося с соответственно указанными SIGLEC или контролем. SIG-5, SIG-7, SIG-F И SIG-9 иммобилизовали с помощью белка А на микротитровочном планшете при концентрации 500 нг/лунка. Биотинилированный кластер II до расщепления (светло-серые столбики) и гидролизованный Сиалидазой А добавляли в концентрации 2 мкг/мл, и после промывания связывание визуализировали с использованием конъюгированного с HRP стрептавидина и измеряя поглощение при 492 нм. В качестве отрицательного контроля использовали антитела к куриным IgY.
На фиг.7 показано схематическое представление рекомбинантно экспрессированных фрагментов фрагментов vWF - Seq11 и Seq12.
На фиг.8 показаны полученные методом MALDI масс-спектры O-гликопептида, выделенного из Seq11 после расщепления трипсином/химотрипсином; расщепления Сиалидазой А и обогащения на лектине. Идентифицированная пептидная последовательность представляет собой KVTLNPSDPEHCQICHCDVVNLTCEACQEPGGLVVPPTDAPVSPTTLYVEDISEPPLHGSAW (SEQ ID NO: 6), где последние четыре аминокислоты (подчеркнуты) соответствуют тэгу, присоединенному к С-концевой последовательности. На верхнем спектре показан полноразмерный O-гликозилированный гликопептид, на нижнем спектре показан тот же гликопептид после расщепления О-гликозидазой.
На фиг.9 показаны полученные методом MALDI масс-спектры O-гликопептида, выделенного из Seq12 после расщепления трипсином/химотрипсином; расщепления сиалидазой А и обогащения на лектине. Идентифицированная пептидная последовательность представляет собой KVTLNPSDPEHCQICHCDVVNLTCEACQEPGGLVVPPTDAPVSPTTLYVEDISEPPLHQEPGGLVVPPTDAPVSPTTLYVEDISEPPLHQEPGGLVVPPTDAPVSPTTLYVEDISEPPLHGSAW (SEQ ID NO:7), где последние четыре аминокислоты (подчеркнуты) соответствуют тэгу, присоединенному к С-концевой последовательности. На верхнем спектре показан полноразмерный O-гликозилированный гликопептид, на нижнем спектре показан тот же гликопептид после расщепления О-гликозидазой.
На фиг.10 показаны результаты теста на связывание рекомбинантных полипептидов Seq11 и Seq12 с SIGLEC. Поглощение при 492 нм пропорционально количеству Seq11 (темно-серые столбики) или Seq12 (светло-серые столбики), связавшихся с соответственно указанными SIGLEC или контролем. SIG-5, SIG-7, SIG-F и SIG-9 иммобилизовали с помощью белка А на микротитровочном планшете при концентрации 500 нг/лунка. Последовательности, несущие метку Strep-Tag, вносили в планшет при равных молярных концентрациях 42 нМ, и после промывания связывание визуализировали с использованием конъюгированного с HRP Strep-Tactin и измеряя поглощение при 492 нм. В качестве отрицательного контроля использовали антитела к куриным IgY, а анти-vWF-pAb использовали в качестве положительного контроля.
На фиг.11 показаны результаты теста на связывание рекомбинантных полипептидов Seq11 и Seq12 после обработки Сиалидазой А. Поглощение при 492 нм пропорционально количеству Seq11 (темно-серые столбики)) или Seq12 (светло-серые столбики), связавшихся с соответственно указанными SIGLEC или контролем. SIG-5, SIG-7, SIG-F И SIG-9 иммобилизовали с помощью белка А на микротитровочном планшете при концентрации 500 нг/лунка. Последовательности, несущие метку Strep-Tag, были ферментативно десиалилированы и внесены в планшет при равных молярных концентрациях 42 нМ, и после промывания связывание визуализировали с использованием конъюгированного с HRP Strep-Tactin и измеряя поглощение при 492 нм. В качестве отрицательного контроля использовали антитела к куриным IgY, а анти-vWF-pAb использовали в качестве положительного контроля.
На фиг.12 показаны результаты зависимого от концентрации связывания рекомбинантного полипептида Seq11 с SIGLEC, и Скетчардовский анализ кривых удельного связывания.
На фиг.13 показаны результаты зависимого от концентрации связывания рекомбинантного полипептида Seq12 с SIGLEC, и Скетчардовский анализ кривых удельного связывания.
На фиг.14 показаны итоговые значения KD, полученных из Скетчардовского анализа, выполненного для кривых, представленных на фиг.12 и фиг.13.
На фиг.15 показаны значения константы диссоциации (KD), рассчитанные для связывания Seq11, Seq12 и полноразмерного плазматического VWF с рекомбинантным FVIII. Данные были получены с помощью SPR.
На фиг.16 показано влияние N- и С-концевых фрагментов VWF на IL-12p70 и IFN-γ. moDC культивировали с различными концентрациями фрагментов VWF либо без добавления (левая колонка колонка), либо с добавление (правая колонка) 0,1 мкг/мл LPS. Внеклеточные уровни цитокинов определяли одновременно с помощью цитометрического анализа на шариках. Уровни IL-12p70 и IFN-γ в нестимулированных клетках были для большинства доноров ниже предела обнаружения (bd, 0,6 пг/мл для IL-12p70 и 1,8 мкг/мл для IFN-γ). Данные представлены как среднее ± SEM, причем каждая точка представляет собой данные от одного донора.
На фиг.17 показаны результаты фосфорилирования SIGLEC-адапторных молекул SHP-1 и SHP-2, участвующих в SIGLEC-сигнальном пути после стимуляции moDC в течение 10 минут 500 нМ N-концевого фрагмента VWF. Клетки, стимулированные тем же объемом 100 мМ NaCl, служили в качестве контроля. Анализ фосфорилирования иммунорецепторов в клеточных лизатах проводили с использованием набора Proteome Profiler Human Phosphosphoreceptor Array Kit. Результаты показаны как средняя плотность пикселей ± SEM для 2-4 отдельных экспериментов.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Для ясного и точного понимания описания и формулы изобретения, включая объем, определяемый такими терминами, ниже приведены определения.
Определения
Используемый в настоящем описании термин «пептид» может состоять из любого числа аминокислот любого типа, предпочтительно, природных аминокислот, которые, предпочтительно, связаны пептидными связями. В частности, пептид содержит по меньшей мере 3 аминокислоты, предпочтительно, по меньшей мере 5, по меньшей мере 7, по меньшей мере 9, по меньшей мере 12 или по меньшей мере 15 аминокислот. Кроме того, отсутствует верхний предел длинеы пептида. Однако предпочтительно, чтобы пептид по изобретению не превышал в длину 500 аминокислот, более предпочтительно, не превышал в длину 300 аминокислот, еще более предпочтительно, не превышал в длину 250 аминокислот.
Таким образом, термин «пептид» включает «олигопептиды», которые обычно относятся к пептидам длиной от 2 до 10 аминокислот, и «полипептиды», которые обычно относятся к пептидам длиной более 10 аминокислот.
Используемый в настоящем описании термин «белок» относится к пептиду из по меньшей мере 60, по меньшей мере 80, по меньшей мере 80, предпочтительно, по меньшей мере 100 аминокислот. Термины «полипептид» и «белок» используются взаимозаменяемо. Полипептиды и белки в контексте настоящего изобретения включают химически синтезированные белки, а также синтезируемые в природе белки, которые кодируются генами. Полипептиды или белки могут быть получены из природного источника, такого как человеческая кровь, или могут быть получены в клеточной культуре в виде рекомбинантных белков.
Используемый в настоящем описании термин «белок млекопитающего» относится к природному белку млекопитающего, то есть к белку, естественным образом экспрессируемому организмом млекопитающего. Поэтому белок млекопитающего имеет природную аминокислотную последовательность и природные посттрансляционные модификации, такие как гликозилирование. В соответствии с настоящим изобретением термины «белок млекопитающего» и «природный белок млекопитающего» могут использоваться взаимозаменяемо.
Используемый в настоящем описании термин «белок человека» относится к природному белку человека, то есть к белку, естественным образом экспрессируемому организмом человека. Таким образом, белок человека имеет природную аминокислотную последовательность и природные посттрансляционные модификации, такие как гликозилирование. В соответствии с настоящим изобретением термины белок человека и природный белок человека используются взаимозаменяемо.
«Рекомбинантные белки» или «рекомбинантные полипептиды» в контексте настоящего изобретения представляют собой те, которые кодируются трансгенами, введенными в клетки методами молекулярной биологии. Белки могут быть модифицированы химическими способами или ферментами в посттрансляционных процессах.
Термин «слитый белок» согласно изобретению относится к белкам, созданным путем соединения двух или нескольких генов, кДНК или последовательностей, которые изначально кодируют отдельные белки/пептиды. В природе гены могут находиться в одном организме или в различных организмах или могут представлять собой синтетические полинуклеотиды.
Термин «терапевтический белок», используемый в настоящем описании, относится к белкам или полипептидам с терапевтическим действием, а именно к белкам, используемым в качестве действующего фармацевтического ингредиента.
Связь между двумя аминокислотными последовательностями или между двумя нуклеотидными последовательностями описывается параметром «идентичность последовательностей». Для целей настоящего изобретения степень идентичности последовательностей между двумя аминокислотными последовательностями определяют с использованием алгоритма Нидлмана-Вунша (Needleman and Wunsch, 1970, J. Mol. Biol. 48: 443-453), реализованного в программе Needle в пакете EMBOSS (EMBOSS: The European Molecular Biology Open Software Suite, Rice et al., 2000, Trends Genet. 16: 276-277), предпочтительно, версии 3.0.0 или более поздней. Используемыми необязательными параметрами являются штраф за введение разрыва - 10, штраф за удлинение разрыва 0,5, и матрица замен EBLOSUM62 (EMBOSS-версия матрицы BLOSUM62). Данные на выходе программы Needle, указанные как «наиболее продолжительная идентичность» (полученные с использованием опции No brief) используют в качестве процента идентичности и вычисляют следующим образом: (идентичные остатки × 100)/(длина выравнивания - общее количество разрывов в выравнивании).
Переходный термин «содержащий», который является синонимом «включающий», «включающий в себя» или «отличающийся тем, что» является является неисключающим или открытым и не исключает дополнительных, неуказанных элементов или стадий способа. Переходная фраза «состоящий из» исключает любой элемент, стадию, или ингредиент, не указанный в формуле изобретения, за исключением примесей, обычно связанных с ними. Когда фраза «состоит из», появляется в пункте формулы изобретения, а не сразу после преамбулы, она ограничивает только элемент, изложенный в этом пункте; другие элементы не исключены из формулы изобретения в целом. Переходная фраза «состоящий по существу из» ограничивает объем утверждения (пункта формулы) указанными материалами или стадиями «и теми, которые не оказывают существенного влияния на основные и новые свойства (или свойство)» заявленного изобретения. Пункт формулы «состоящий по существу из» занимает промежуточное положение между закрытым пунктом формулы объекта, выраженным в формате «состоящий из» и полностью открытым пунктом формулы, выраженным в формате «содержащий».
Термин «гомологичный», используемый в настоящем описании, означает, что соответствующая нуклеотидная последовательность аминокислотной последовательности имеет заданную степень идентичности с эталонной аминокислотной последовательностью и заданными нуклеотидными последовательностями. Предполагают, что гомологичная последовательность включает аминокислотную последовательность, которая по меньшей мере на 75%, 80%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или даже на 99% идентична заданной последовательности с использованием обычной программы для выраванивания последовательностей Clustal V с параметрами по умолчанию. Как правило, гомологи включают в себя одинаковые остатки в активных сайтах с заданной аминокислотной последовательностью, хотя могут включать любое число консервативных аминокислотных замен. Термин «идентичный», используемый в настоящем описании, относится к 100%-й идентичности по аминокислотной или нуклеотидной последовательности с эталонной последовательностью.
Термин «рекомбинантный» при использовании в отношении рассматриваемых клетки, нуклеиновой кислоты, белка или вектора, указывает на то, что субъект был модифицирован путем введения гетерологичной нуклеиновой кислоты или белка, или изменения природных нуклеиновой кислоты или белка, или на то, что клетка получена из клетки, модифицированной таким образом. Например, рекомбинантные клетки экспрессируют гены, которые не обнаруживаются в природной (не рекомбинантной) форме клетки, или экспрессируют природные гены на другом уровне или при других условиях, чем в природе.
Используемые в настоящем описании термины «трансформированный», «стабильно трансформированный» и «трансгенный», используемые в отношении клетки означает, что клетка содержит неприродную (например, гетерологичную) нуклеотидную последовательность, встроенную в ее геном или перенесенную в виде эписомы, которая поддерживается в множестве поколений.
Термин «фрагмент», используемый в настоящем изобретении, относится к полипептиду, который имеет на амино- и/или карбоксильном конце делецию одной или нескольких аминокислот по сравнению с нативным белком или белком дикого типа, но остальная аминокислотная последовательность является идентичной с соответствующими позициями в аминокислотной последовательности, выведенной из полноразмерной кДНК. Фрагменты как правило составляют по меньшей мере 50 аминокислот в длину.
Термин «гликозилирование», используемый в настоящем описании, относится к присоединению гликанов к молекулам, например, к белкам. Гликозилирование может быть ферментативной реакцией. Присоединение может происходить через ковалентные связи. Соответственно, гликозилированный полипептид в контексте настоящего изобретения представляет собой полипептид, к которому присоединены гликаны. Выражение «высоко гликозилированный» относится к молекуле, такой как фермент, который гликозилирован по всем или почти всем доступным сайтам гликозилирования, например, по сайтам О- или N-связанного гликозилирования.
Термин «гликан», используемый в настоящем описании, относится к полисахариду или олигосахариду, или к углеводному фрагменту гликопротеина или гликозилированного полипептида. Гликаны могут представлять собой гомо- или гетерополимеры моносахаридных остатков. Они могут представлять собой линейные или разветвленные молекулы. Гликаны, как правило, содержат по меньшей мере три сахара, и могут быть линейными или разветвленными. Гликан может включать природные сахара (например, глюкозу, N-ацетилглюкозамин, N-ацетилнейраминовую кислоту, галактозу, маннозу, фукозу, гексозу, арабинозу, рибозу, ксилозу и т.д.) и/или модифицированные сахара (например, 2'-фторрибозу, 2'-дезоксирибозу, фосфоманнозу, 6'-сульфо-N-ацетилглюкозамин и т.д.)
Термин «О-гликаны», используемый в настоящем описании, относится к гликанам, которые обычно являются ковалентно связанными с остатками серина и треонина в гликопротеинах млекопитающих. О-гликаны могут быть α-связаны через N-ацетилгалактозаминовый (GalNAc) фрагмент с ОН-группой серина или треонина через O-гликозидную связь. Другие связи включают гликаны α-связанную О-фукозу, β-связанную О-ксилозу, α-связанную O-маннозу, β-связанный O-GlcNAc (N-ацетилглюкозамин), α- или β-связанную О-галактозу и α- или β-связанные О-глюкозу.
Термин «сиалилированный», используемый в настоящем описании, относится к молекулам, в частности гликанам, которые вступили в реакцию с сиаловой кислотой или ее производными.
Термины «аффинность связывания» или «сродство», используемые в настоящем описании, указывают на прочность связи между двумя молекулами, в частности между лигандом и белком-мишенью. На аффинность связывания влияют нековалентные межмолекулярные взаимодействия между двумя молекулами, такие как водородные связи, электростатические взаимодействия, гидрофобные взаимодействия и силы ван-дер-Ваальса.
Иммунный ответ в контексте настоящего описания относится к адаптивному или врожденному иммунному ответу. Врожденный иммунный ответ относится к неспецифическим защитным механизмам, которые активируются немедленно или в течение нескольких часов после попадания чужеродного антигена в организм. Эти механизмы включают физические барьеры, такие как кожа, химические вещества в крови и клетки иммунной системы, которые атакуют чужеродные клетки в организме. Врожденный иммунный ответ активируется химическими свойствами антигена. Адаптивный иммунный ответ относится к антигенспецифичному иммунному ответу. Для этого антиген должен быть обработан и распознан. После распознавания антигена, адаптивная иммунная система создает большое число иммунных клеток, специально созданных для атаки этого антигена.
Используемый в настоящем описании термин «иммунная толерантность» (или просто «толерантность»)) представляет собой процесс, с помощью которого иммунная система не атакует антиген. Этот процесс существует в трех формах: центральная толерантность, периферическая толерантность и приобретенная толерантность. Толерантность может являться «природной» или «самотолерантностью», когда организм не вырабатывает иммунный ответ на аутоантигены, либо «индуцированной толерантностью», когда толерантность к антигенам может быть выработана путем манипулирования иммунной системой.
Гликозилированный полипептид
Согласно первому аспекту изобретение относится к гликозилированному полипептиду, содержащему аминокислотную последовательность, идентичную или гомологичную по меньшей мере фрагменту белка млекопитающего, предпочтительно, человека, где указанный гликозилированный полипептид содержит один или несколько сиалилированных О-гликанов, и где полипептид обладает повышенной аффинностью связывания с одним или несколькими SIGLEC по сравнению с белком млекопитающего или его фрагментом.
Гликозилированный полипептид по изобретению основан на белке млекопитающего, т.е. содержит аминокислотную последовательность, идентичную или гомологичную белку млекопитающего. Белком млекопитающего является, в частности, белок человека. Белком человека, с которым гомологична или идентична аминокислотная последовательность гликозилированного пептида, предпочтительно, является гликозилированный белок.
Более предпочтительно, чтобы белок человека представлял собой белок крови человека. Белок крови человека может представлять собой фактор свертываемости крови человека, транспортный белок, ингибитор протеаз, иммуноглобулин, клеточный белок плазмы, аполипопротеины, фактор комплемента, ростовой фактор, антиангиогенный белок, высокогликозилированный белок, факторы крови или другой белок крови человека.
Фактор свертываемости крови человека выбирают, в частности, из группы, состоящей из фибриногена, мономера фибрина, протромбина, тромбина, FV, FX, FIX, FVII, FVIII, FXI, FXII и FXIII, фактора фон Виллебранда и ADAMTS13.
Понятно, что факторы свертываемости FV, FX, FIX, FVII, FVIII, FXI, FXII и FXIII имеют неактивную и актвированную форму. Таким образом, в контексте изобретения ссылка на FV, FX, FIX, FVII, FVIII, FXI, FXII и FXIII включает активированные формы FVa, FXa, FIXa, FVIIa, FVIIIa, FXIa, FXIIa и FXIIIa, соответственно, если не указано иное, или из контекста активированная форма может быть логически не включена. Таким образом, например, в этом контексте FV, FX, FIX, FVII, FVIII, FXI, FXII и FXIII могут читаться как FV/FVa, FX/FXa, FIX/FIXa, FVII/FVIIa, FVIII/FVIIIa, FXI/FXIa, FXII/FXIIa, FXIII/FXIIIa.
Транспортный белок может быть выбран из альбумина, трансферрина, церулоплазмина, гаптоглобина, гемоглобина и гемопексина.
Потенциальными ингибиторами протеаз являются, например, β-антитромбин, α-антитромбин, окисленный-антитромбин, 2-макроглобулин, C1-ингибитор, ингибитор пути тканевого фактора (TFPI), кофактор гепарина II, ингибитор белка С (PAI-3), белок С, белок S и белок Z.
Примеры иммуноглобулинов явлются, например, поликлональные антитела (IgG), моноклональные антитела, IgG1, IgG2, IgG3, IgG4, IgA, IgA1, IgA2, IgM, IgE, IgD и белок Бенс-Джонса.
Клеточным белком плазмы может являться, например, фибронектин, тромбоглобулин, тромбоцитарный фактор 4. Примерами аполипопротеинов являются apo A-I, apo A-II и apo E.
Факторами комплемента согласно изобретению являются, например, фактор В, фактор D, фактор H, фактор I, C3b-инактиватор, пропердин, C4-связывающий белок и т.д.
Примеры ростовых факторов включают фактор роста, полученный из тромбоцитов (PDGF), эпидермальный фактор роста (EGF), трансформирующий фактор роста-альфа (TGF-α), трансформирующий фактор роста-бета (TGF-β), фактор роста фибробластов (FGF) и фактор роста гепатоцитов.
Антиангиогенные белки включают в себя латентный антитромбин, прелатентный антитромбин, окисленный антитромбин и плазминоген.
Примерами высоко гликозилированных белков являются альфа-1-кислотный гликопротеин, антихимотрипсин, интер-α-трипсин-ингибитор, α-2-HS-гликопротеин, С-реактивный белок. Факторами крови могут быть, например, эритропоэтин, интерферон, опухолевые факторы, tPA, gCSF.
Другие белки крови человека включают богатый гистидином гликопротеин, маннан-связывающий лектин, С4-связывающий белок, фибронектин, GC-глобулин, плазминоген/плазмин, α-1-микроглобулин, C-реактивный белок.
Белок человека, в частности, выбирают из vWF, FVIII, FVII, FIX, ADAMTS13.
Фактор VIII у человека кодируется геном F8, который содержит 187000 пар оснований в шести экзонах. Транскрибированная мРНК имеет длину 9029 пар оснований и транслируется в белок из 2351 аминокислот, из которых в результате посттрансляционной модификации удаляются 19 аминокислот. Молекула FVIII у человека гликозилирована по 31-аминокислотным остаткам (25 гликозилирований по азоту и 6 гликозилирований по атому O).
После трансляции аминокислотная цепь расщепляется специфичными протеазами в определеных сайтах, что приводит к образованию тяжелой цепи примерно 200 кДа и легкой цепи примерно 80 кДа. Доменная организация обычно характеризуется как A1-A2-B-A3-C1-C2. Легкая цепь состоит из доменов A3-C1-C2. Тяжелая цепь в основном состоит из доменов А1-А2-В. Тяжелые цепи, находящиеся в плазме, имеют гетерогенный состав с молекулярным весом в диапазоне от 90 до 200 кДа. Причиной этого является гетерогенность гликозилирования, существование сплайсинговых вариантов и протеолитических продуктов, таких как тяжелая цепь A1 A2 без В-домена. Аминокислотная последовательность полноразмерного FVIII представлена аминокислотами с 20-й по 2351-ю в последовательности Р00451 (Swisssrot, 21 июля 1986 г.).
Предпочтительно, чтобы белок человека являлся полноразмерным FVIII, представленным аминокислотами с 20-й по 2351-ю в последовательности Р00451 (Swisssrot, 21 июля 1986 г.), FVIII с делецией B-домена или белком FVIII, в котором часть В-домена заменена линкером.
vWF представляет собой мультимерный адгезивный гликопротеин, присутствующий в плазме млекопитающих, который обладает множественными физиологическими функциями. Во время первичного гемостаза vWF действует как посредник между специфическими рецепторами на поверхности тромбоцитов и компонентами внеклеточного матрикса, такими как коллаген. Кроме того, vWF служит носителем и стабилизирующим белком для прокоагулянтного фактора VIII. vWF синтезируется в эндотелиальных клетках и мегакариоцитах в виде 2813-аминокислотной молекулы-предшественника. Предшественник полипептида, пре-про-vWF, состоит из 22-аминокислотного сигнального пептида, 741-аминокислотного пропептида и 2050-аминокислотного полипептида в зрелом плазматическом факторе фон Виллебранда (Fischer et al., 1994). Полноразмерный vWF имеет номер P04275 в базе данных Uniprot.
После секреции в плазму vWF циркулирует в ней в виде молекул с различной молекулярной массой. Эти молекулы фактора vWF состоят из олиго- и мультимеров зрелой 2050-аминокислотной субъединицы. vWF обычно находится в плазме в виде мультимеров с размером в диапазоне от 500 до 20000 кДа (Furlan et al. 1996). В частности, аминокислотная последовательность vWF соотстветствует любой из последовательностей с номером P04275 в Uniprot. Более предпочтительно, чтобы белок vWF имел последовательность SEQ ID NO: 1.
SLSCRPPMVKLVCPADNLRAEGLECTKTCQNYDLECMSMGCVSGCLCPPGMVRHENRCVALERCPCFHQGKEYAPGETVKIGCNTCVCQDRKWNCTDHVCDATCSTIGMAHYLTFDGLKYLFPGECQYVLVQDYCGSNPGTFRILVGNKGCSHPSVKCKKRVTILVEGGEIELFDGEVNVKRPMKDETHFEVVESGRYIILLLGKALSVVWDRHLSISVVLKQTYQEKVCGLCGNFDGIQNNDLTSSNLQVEEDPVDFGNSWKVSSQCADTRKVPLDSSPATCHNNIMKQTMVDSSCRILTSDVFQDCNKLVDPEPYLDVCIYDTCSCESIGDCACFCDTIAAYAHVCAQHGKVVTWRTATLCPQSCEERNLRENGYECEWRYNSCAPACQVTCQHPEPLACPVQCVEGCHAHCPPGKILDELLQTCVDPEDCPVCEVAGRRFASGKKVTLNPSDPEHCQICHCDVVNLTCEACQEPGGLVVPPTDAPVSPTTLYVEDISEPPLHDFYCSRLLDLVFLLDGSSRLSEAEFEVLKAFVVDMMERLRISQKWVRVAVVEYHDGSHAYIGLKDRKRPSELRRIASQVKYAGSQVASTSEVLKYTLFQIFSKIDRPEASRITLLLMASQEPQRMSRNFVRYVQGLKKKKVIVIPVGIGPHANLKQIRLIEKQAPENKAFVLSSVDELEQQRDEIVSYLCDLAPEAPPPTLPPDMAQVTVGPGLLGVSTLGPKRNSMVLDVAFVLEGSDKIGEADFNRSKEFMEEVIQRMDVGQDSIHVTVLQYSYMVTVEYPFSEAQSKGDILQRVREIRYQGGNRTNTGLALRYLSDHSFLVSQGDREQAPNLVYMVTGNPASDEIKRLPGDIQVVPIGVGPNANVQELERIGWPNAPILIQDFETLPREAPDLVLQRCCSGEGLQIPTLSPAPDCSQPLDVILLLDGSSSFPASYFDEMKSFAKAFISKANIGPRLTQVSVLQYGSITTIDVPWNVVPEKAHLLSLVDVMQREGGPSQIGDALGFAVRYLTSEMHGARPGASKAVVILVTDVSVDSVDAAADAARSNRVTVFPIGIGDRYDAAQLRILAGPAGDSNVVKLQRIEDLPTMVTLGNSFLHKLCSGFVRICMDEDGNEKRPGDVWTLPDQCHTVTCQPDGQTLLKSHRVNCDRGLRPSCPNSQSPVKVEETCGCRWTCPCVCTGSSTRHIVTFDGQNFKLTGSCSYVLFQNKEQDLEVILHNGACSPGARQGCMKSIEVKHSALSVELHSDMEVTVNGRLVSVPYVGGNMEVNVYGAIMHEVRFNHLGHIFTFTPQNNEFQLQLSPKTFASKTYGLCGICDENGANDFMLRDGTVTTDWKTLVQEWTVQRPGQTCQPILEEQCLVPDSSHCQVLLLPLFAECHKVLAPATFYAICQQDSCHQEQVCEVIASYAHLCRTNGVCVDWRTPDFCAMSCPPSLVYNHCEHGCPRHCDGNVSSCGDHPSEGCFCPPDKVMLEGSCVPEEACTQCIGEDGVQHQFLEAWVPDHQPCQICTCLSGRKVNCTTQPCPTAKAPTCGLCEVARLRQNADQCCPEYECVCDPVSCDLPPVPHCERGLQPTLTNPGECRPNFTCACRKEECKRVSPPSCPPHRLPTLRKTQCCDEYECACNCVNSTVSCPLGYLASTATNDCGCTTTTCLPDKVCVHRSTIYPVGQFWEEGCDVCTCTDMEDAVMGLRVAQCSQKPCEDSCRSGFTYVLHEGECCGRCLPSACEVVTGSPRGDSQSSWKSVGSQWASPENPCLINECVRVKEEVFIQQRNVSCPQLEVPVCPSGFQLSCKTSACCPSCRCERMEACMLNGTVIGPGKTVMIDVCTTCRCMVQVGVISGFKLECRKTTCNPCPLGYKEENNTGECCGRCLPTACTIQLRGGQIMTLKRDETLQDGCDTHFCKVNERGEYFWEKRVTGCPPFDEHKCLAEGGKIMKIPGTCCDTCEEPECNDITARLQYVKVGSCKSEVEVDIHYCQGKCASKAMYSIDINDVQDQCSCCSPTRTEPMQVALHCTNGSVVYHEVLNAMECKCSPRKCSK (SEQ ID NO: 1)
Гликозилированный полипептид может, например, содержать фрагмент vWF согласно WO 2015/185758 А2. Как показано в WO 2015/185758 А2, комплекс FVIII и фрагментов vWF согласно изобретению демонстрирует меньшее связывание с фосфолипидными мембранами по сравнению с одним FVIII, а также пониженное связывание с коллагеном III и гепарином по сравнению с комплексом FVIII и полноразмерным vWF.
Вышеуказанный фрагмент vWF представляет собой, в частности, фрагмент, начиная с аминокислоты 1 в SEQ ID NO: 1. Аминокислоты 1-272 в SEQ ID NO: 1 содержит домен связывания FVIII с vWF.
Фрагмент vWF, предпочтительно начинающийся с 1-й аминокислоты в SEQ ID NO 1, предпочтительно, заканчивается аминокислотой из SEQ ID NO: 1 в диапазоне от 1142 до 1390. Более предпочтительно, чтобы фрагмент заканчивался аминокислотой в диапазоне от 1267 до 1390. Более предпочтительно, чтобы фрагмент vWF заканчивался аминокислотой из SEQ ID NO: 1 в диапазоне от 1337 до 1390.
Следует понимать, что гликозилированный полипептид имеет повышенную аффинность связывания по сравнению с белком или фрагментом млекопитающего, определяемыми аминокислотной последовательностью, содержащейся в гликозилированном пептиде. Таким образом, если гликозилированный полипептид содержит аминокислотную последовательность полноразмерного белка млекопитающего, то тогда гликозилированный полипептид имеет более высокое сродство к SIGLEC по сравнению с полноразмерным белком млекопитающих.
С другой стороны, если гликозилированный полипептид содержит фрагмент белка млекопитающего, определяемый субпоследовательностью белка млекопитающего, то гликозилированный полипептид обладает повышенной аффинностью связывания по сравнению с идентичным фрагментом из природного белка. Например, если аминокислотная последовательность в гликозилированном полипептиде идентична или гомологична фрагменту vWF, то гликозилированный полипептид по первому аспекту обладает повышенной аффинностью связывания с одним или несколькими SIGLEC по сравнению с тем же фрагментом, полученным в результате фрагментации vWF, полученного из плазмы.
Как показано в примерах, была определена гликановая структура vWF, которая специфично связывается по меньшей мере с SIGLEC SIG-5, SIG-7, SIG-8 и SIG-9 (см. пример 2). Таким образом, согласно одному варианту осуществления первого аспекта один или несколько SIGLEC выбирают из группы SIG-5, SIG-7, SIG-8 и SIG-9.
Авторы изобретения неожиданно обнаружили, что в vWF человека О-гликаны ответственны за связывание с SIG-5, SIG-7, SIG-8 и SIG-9. В отличие от этого, N-гликаны не связываются с этими SIGLEC (см. пример 2). Это особенно удивительно, потому что до сего момента как раз было показано, что N-гликаны взаимодействуют с SIGLEC (Lai et al, 2015).
Авторы изобретения также определили, что О-гликаны не только должны быть сиалилированы для связывания с SIG-5, SIG-7, SIG-8, SIG-9, но также должен присутствовать минимальный процент коровых O-гликанов 2-го типа (см. пример 4).
Поэтому, связывание SIGLEC и, следовательно, пониженный иммунный ответ основаны на повышенном количестве или процентном содержании сиалилированных коровых O-гликанов 2-го типа в гликозилированном белке по сравнению с числом сиалилированных коровых O-гликанов 2-го типа в белке млекопитающего или его фрагменте.
Из-за структурной аналогии предполагается, что сиалилированные удлиненные коровые O-гликаны 1-го типа обладают тем же эффектом, что и сиалилированные коровые O-гликаны 2-го типа. Таким образом, для повышения аффинности связывания с указанными выше SIGLEC предпочтительно, чтобы увеличивалось объединенное количество или процентное содержание сиалилированных коровых O-гликанов 2-го типа и удлиненных коровых O-гликанов 1-го типа.
Таким образом, согласно одному варианту осуществления изобретения количество сиалилированных коровых O-гликанов 2-го типа и/или удлиненных коровых O-гликанов 1-го типа в гликозилированном полипептиде превышает количество сиалилированных коровых O-гликанов 2-го типа и/или удлиненных коровых O-гликанов 1-го типа в белке млекопитающего или его фрагменте. Соответственно, также увеличивается процентное содержание сиалилированных коровых O-гликанов 2-го типа и/или удлиненных коровых O-гликанов 1-го типа по сравнению с процентным содержанием коровых O-гликанов 2-го типа и/или удлиненных коровых O-гликанов 1-го типа в белке млекопитающего.
Это означает, что объединенное количество сиалилированных коровых O-гликанов 2-го типа и сиалилированных удлиненных коровых O-гликанов 1-го типа гликозилированного полипептида превышает объединенное количество сиалилированных коровых O-гликанов 2-го типа и сиалилированных удлиненных коровых O-гликанов 1-го типа в белке млекопитающего или его фрагменте.
В альтернативном варианте может быть увеличено только количество сиалилированных коровых O-гликанов 2-го типа. Соответственно, также увеличивается процентное содержание сиалилированных коровых O-гликанов 2-го типа по сравнению с процентным содержание сиалилированных коровых O-гликанов 2-го типа в белке млекопитающего.
Молекулы SIGLEC, с которыми было показано связывание, участвуют в иммунном ответе у человека и мышей. SIGLEC имеют в общем N-концевой иммунгоглобулиновый V-домен, который связывает лиганды, содержащие сиаловую кислоту, и различное число иммуноглобулиновых С2-доменов, которые удаляют сай связывания лиганда от поверхности мембраны.
Кроме того, многие SIGLEC имеют цитоплазматические тирозиновые мотивы, включая иммунорецепторный тирозиновый ингибирующий мотив (ITIM) и ITIM-подобные мотивы, обычно присутствующие в корецепторах, участвующих в регуляции клеточной сигнализации. Другие SIGLEC не содержат тирозиновых мотивов, но содержат положительно заряженную трансмембранную область, которая позволяет связывание с белками-адаптерами. SIGLEC не распознают связанные опасностью молекулярные сигналы (danger associated molecular patterns, DAMPS), а наоборот распознают детерминанты «самоидентификации».
SIGLEC связываются с сиалилированными лигандами самоидентификации (самолигандами) на той же клетке в «цис»-манере и на соседних клетках в «транс»-манере. SIGLEC человека обычно обозначаются с SIG-1 по SIG-14. Мышиные SIGLEC - SIG-E, SIG-F и SIG-G, соответственно представляют собой ортологи SIG-9, SIG-8 и SIG-10 человека.
Молекулы SIG-1 - SIG-4 также именуются соответственно Сиалоатезином, CD22, CD33 и MAG. CD22 и SIG-10 присутствуют в В-клетках, SIG-5 в нейтрофилах и моноцитах, SIG-7 в NK-клетках, SIG-8 в эозинофилах, SIG-9 в моноцитах, нейтрофилах и дендритных клетках (Paulsen et al 2012).
SIGLEC связываются с различными гликановыми структурами. Каждый из SIG-2, SIG-5, SIG-7, SIG-8, SIG-9 и SIG-10, предпочтительно, связывается с различными гликанами (Paulson et al., 2012). SIGLEC играют роль в врожденном и адаптивном иммунитете. В частности, SIG-2 и SIG-10, которые присутствуют на B-клетках человека и мышей.
Согласно Paulsen и др., SIG-2 и SIG-10, по-видимому, оказывают синергическое воздействие на толерантность периферических В-клеток. Кроме того, SIGLEC действуют как ингибирующие корецепторы для Toll-подобных рецепторов (TRL). В связи с этим было показано, что сшивающие SIG-7 или SIG-9 с активирующими рецепторами приводят к ингибированию цитолитической активности NK-клеток в отношении опухолевых клеток и, соответственно, к высвобождению химических медиаторов из тучных клеток.
Более того, сшивание SIG-E (SIG-9) и SIG-11 иммобилизованным антителом приводит к ингибированию продукции цитокинов в макрофагах в ответ на LPS.
Следует отметить, что экспрессия SIG-5 и SIG-9 в клеточной макрофагальной линии ингибирует продукцию TNF-альфа и увеличивает продукцию IL-10 в ответ на пептидный гликан, лиганд ATLR2, LPS и CpG. Кроме того, показано, что LPS-индуцированная экспрессия SIG-E (SIG-9) в макрофагах влияет на передачу сигнала от TRL. Кроме того, сиалилированные патогены ослабляют иммунный ответ через SIGLEC. Например, стрептококк группы B экпрессирует остаток Neu-Acα-1 Galβ-1 4GlcNAc на полисахаридах оболочки и рекрутирует SIGLEC-9 на нейтрофилах, что приводит к подавлению микробицидной функции нейтрофилов.
Соответственно, без привязки к какой-либо конкретной теории, считается, что связывание с SIGLEC на антигенпредставляющих клетках (например, дендритных клетках) приводит к снижению экспрессии провоспалительных рецепторов и повышению экспрессии иммуносупрессивных рецепторов на поверхности клетки. Кроме того, связывание приводит к повышению продукции противовоспалительных цитокинов, снижает продукцию провоспалительных цитокинов и, как следствие, приводит к ингибированию пролиферации Т-клеток и продукции антител. Таким образом, связывание SIG-5, SIG-7, SIG-8 и SIG-9 приводит к снижению иммунного ответа или повышенной иммунной толерантности при введении пациенту гликозилированного полипептида.
Таким образом, гликозилированный полипептид в соответствии с первым аспектом также может быть описан как гликозилированный полипептид, содержащий аминокислотную последовательность, идентичную или гомологичную по меньшей мере фрагменту белка млекопитающего, предпочтительно, человека, где указанный гликозилированный полипептид содержит один или несколько сиалилированных O-гликанов, и где по сравнению с белком млекопитающего или его фрагментом:
- снижается иммунный ответ человека на гликозилированный полипептид; и/или
- повышается иммунная толерантность человека к гликозилированному полипептиду.
С другой стороны, более структурное определение первого аспекта изобретения представляет собой следующее: гликозилированный полипептид, содержащий аминокислотную последовательность, идентичную или гомологичную по меньшей мере фрагменту белка млекопитающего, предпочтительно, человека, где указанный гликозилированный полипептид содержит один или несколько сиалилированных O-гликанов, и где объединенное количество сиалилированных коровых O-гликанов 2-го типа и сиалилированных удлиненных коровых O-гликанов 1-го типа гликозилированного полипептида превышает объединенное количество сиалилированных коровых O-гликанов 2-го типа и сиалилированных удлиненных коровых O-гликанов 1-го типа в белке млекопитающего или его фрагменте.
Предпочтительно, чтобы гликозилированный полипептид, содержащий аминокислотную последовательность, был идентичен или гомологичен по меньшей мере фрагменту белка млекопитающего, предпочтительно, человека, где указанный гликозилированный полипептид содержит один или несколько сиалилированных O-гликанов, и где количество сиалилированных коровых O-гликанов 2-го типа гликозилированного полипептида превышет объединенное количество сиалилированных коровых O-гликанов 2-го типа в белке млекопитающего или его фрагменте.
Для соединения O-гликанов с аминокислотной последовательностью гликозилированного полипептида, он содержит один или несколько сайтов O-гликозилирования. Сайты О-гликозилирования в гликозилированном полипептиде могут представлять собой стандартные сайты О-гликозилирования - серин (Ser) и треонин (Thr). Однако в контексте изобретения также было описано связывание O-гликанов с тирозином (Tyr), гидроксилизином (Hydroxy-Lys) или гидроксипролином (Hydroxy-Pro). Таким образом, один или несколько сайтов O-гликозилирования в гликозилированном полипептиде могут быть выбраны из Ser, Thr, Tyr, Hydroxy-Lys и Hydroxy-Pro в любом другом возможном месте O-гликозилирования. Предпочтительно, чтобы сайты O-гликозилирования были выбраны из Ser и Thr.
Стандартные сайты гликозилирования Ser и Thr, как правило, в наибольшей степени заняты О-гликанами. Таким образом, согласно одному варианту осуществления, один или несколько сайтов гликозилирования выбирают из Ser и Thr.
В контексте изобретения по практическим соображениям термин «гликозилированный полипептид» используется в одной форме. Обычно же гликозилированный полипептид будет представлен в форме композиции полипептидов одного типа. В этом отношении гликозилированные полипептиды в первоначальной форме представляют собой композицию гликозилированных полипептидов, имеющих одну аминокислотную последовательность, однако отличия в гликозилировании. Например, не все индивидуальные молекулы композиции могут быть гликозилированы на 100 процентов. Кроме того, могут присутствовать отличия в гликанах, связанных с конкретным сайтом O-гликозилирования. Соответственно, настоящее изобретение также относится к композиции, содержащей, по меньшей мере молекулы гликозилированного полипептида первого типа, где аминокислотная последовательность белковых молекул первого типа идентична или гомологична по меньшей мере фрагменту белка млекопитающего, предпочтительно, человека, и молекулы белка содержат один или несколько сайтов гликозилирования.
Предпочтительно, чтобы полипептид содержал один или несколько кластеров сайтов гликозилирования. Хотя единственный сайт гликозилирования может быть достаточным для связывания SIGLEC, предполагается, что формирование кластера сайтов O-гликозилирования приводит к улучшению связывания с SIGLEC. Образование кластера сайтов гликозилирования часто наблюдается в белках млекопитающих, примерами являются IgA человека, содержащие кластерные O-гликаны в шарнирной области (см. Franc et al. 2013), и муцин человека (см. Guzman-Aranguez and Argüeso 2010).
В этим контексте уже два близкорасположенных сайта O-гликозилирования рассматриваются как кластер O-гликозилирования. Таким образом, один или несколько кластеров сайтов O-гликозилирования содержат по меньшей мере два сайта O-гликозилирования. Кластеры сайтов O-гликозилирования могут иметь разное количество сайтов O-гликозилирования. Например, гликозилированный полипептид может содержать один кластер с двумя и второй кластер с тремя сайтами гликозилирования. Между сайтами O-гликозилирования в кластере могут присутствовать также сайты N-гликозилирования. Предпочтительно, чтобы в кластере O-гликозилирования не было сайтов N-гликозилирования.
Кроме того, один кластер может содержать три сайта O-гликозилирования, а другой - четыре сайта O-гликозилирования. Ряд из трех сайтов O-гликозилирования дает три расположенных рядом O-гликана, которые могут взаимодействовать с SIGLEC и, следовательно, приводят к увеличению эффекта. Согласно одному варианту осуществления один или несколько кластеров сайтов O-гликозилирования, предпочтительно, содержат по меньшей мере три сайта O-гликозилирования.
В vWF присутствуют два кластера с четырьмя сайтами O-гликозилирования в каждом. Таким образом, предпочтительно, чтобы полипептид содержал один или несколько кластеров по меньшей мере с четырьмя сайтами O-гликозилирования. В настоящее время предполагается, что чем выше число сайтов O-гликозилирования, тем выше аффинность связывания.
Кластер сайтов O-гликозилирования может характеризоваться наличием двух или нескольких сайтов O-гликозилирования на близком расстоянии друг от друга в аминокислотной последовательности. Эти кластеры также указываютя как «последовательные кластеры». Однако из-за трехмерной структуры гликозилированного полипептида кластер O-гликозилирования может также включать сайты O-гликозилирования, которые находятся на большом расстоянии в аминокислотной последовательности, но после укладки располагаются в непосредственной близости. Этот тип кластеров также называется «структурным кластером».
В vWF 2-й кластер O-гликозилирования содержит 4 сайта O-гликозилирования в пределах 20 аминокислот, которые расположены в виде бета-поворота. Соответственно, расстояние между O-гликанами или сайтами O-гликозилирования составляет от 27,2 до 34,0 Å, что дает среднее расстояние от 6,8 до 8,5 Å. Таким образом, среднее расстояние между двумя сайтами O-гликозилирования в кластере может находиться в диапазоне от 4,0 до 15,0 Å. При расстоянии меньше 4,0 Å могут возникать стерические затруднения между O-гликанами, в частности, может быть невозможно гликозилирование обоих сайтов O-гликозилирования. При среднем расстоянии между двумя аминокислотами более 15,0 Å, вероятно, нет никакого комбинированного эффекта O-гликанов. Комбинированным эффектом является, например, взаимодействие с SIGLEC на одной клетке. Предпочтительно, чтобы среднее расстояние между двумя сайтами O-гликозилирования в кластере находилось в диапазоне от 5,0 до 12,0 Å. Более предпочтительно, чтобы среднее расстояние между двумя сайтами O-гликозилирования в кластере находилось в диапазоне от 6,0 до 9,0 Å.
Структурные кластеры могут охватывать аминокислотные последовательности из более чем 100 аминокислот. Однако предпочтительно, чтобы пространственное расположение кластера не превышало 80 Å. Если сайты O-гликозилирования отделены друг от друга более чем на 80 Å, предполагается, что O-гликаны не проявляют комбинированного эффекта. Комбинированный эффект O-гликанов в кластере является самым сильным, если O-гликаны расположены в области с диаметром 50 Å. Таким образом, более предпочтительно, чтобы пространственная компоновка кластера не превышала 50 Å.
Согласно одному варианту осуществления один или несколько кластеров, например, последовательных кластеров, содержат по меньшей мере один сайт O-гликозилирования в десяти аминокислотах. Если сайты O-гликозилирования расположены на большем расстоянии, считается, что O-гликаны, связанные с сайтом O-гликозилирования, возможно, не могут взаимодействовать совместно с одной и той же клеткой, содержащей SIGLEC.
Предпочтительно, чтобы один или несколько кластеров содержали по меньшей мере один сайт O-гликозилирования в четырех аминокислотах. При среднем расстоянии между сайтами O-гликозилирования в четыре аминокислоты существует высокая вероятность того, что сайты O-гликозилирования также являются близкорасположенными после укладки. Пространственная близость позволяет совместное взаимодействие гликанов в кластере с SIGLEC на одной и той же клетке.
Более предпочтительно, чтобы один или несколько кластеров содержали по меньшей мере один сайт О-гликозилирования в трех аминокислотах. Как показано в примерах, тестируемый vWF-пептид содержит кластеры с одним сайтом O-гликозилирования в двух аминокислотах. Таким образом, согласно предпочтительному варианту осуществления один или несколько кластеров содержат по меньшей мере один сайт гликозилирования в двух аминокислотах.
Как показано в примерах, уже один из таких кластеров O-гликозилирования достаточен для сильного взаимодействия с SIGLEC. Более того, как также было показано путем сравнения двух полипептидов vWF, большее количество кластеров сайтов O-гликозилирования приводит к увеличению аффинности связывания пептида с SIGLEC, в частности, с SIG-5, SIG-7, SIG-8 и SIG-9. Таким образом, гликозилированный полипептид предпочтительно содержит по меньшей мере два кластера гликозилирования, более предпочтительно, по меньшей мере три кластера гликозилирования.
Без привязки к какой-либо конкретной теории, аффинность связывания гликозилированного полипептида с SIGLEC выше, чем ближе расположены кластеры. В этом контексте, если присутствуют два кластера, то их предпочтительно разделяют менее 100 аминокислот. Расстояние менее 100 аминокислот позволяет совместное действие кластеров гликанов при связывании SIGLEC. Более предпочтительно, чтобы два кластера были разделены менее чем 50 аминокислотами. Наиболее предпочтительно, чтобы два кластера были разделены менее чем 30 аминокислотами.
Согласно одному варианту осуществления изобретения, расстояние между любыми двумя соседними кластерами составляет менее 100 аминокислот, предпочтительно, расстояние между любыми двумя кластерами в гликозилированном полипептиде составляет менее 50 аминокислот. Более предпочтительно, расстояние между любыми двумя соседними кластерами составляет менее 30 аминокислот.
Гликозилированный полипептид, предпочтительно, содержит по меньшей мере один дополнительный кластер сайтов O-гликозилирования по сравнению с белком человека, с которым эта последовательность является гомологичной или идентичной.
Как описаноы выше, подобно всем гликозилированным полипептидам, гликозилированный полипептид по изобретению представляет собой композицию гликозилированных полипептидных молекул. Эти молекулы имеют определенную степень гетерогенности в гликозилировании, в частности - не все сайты гликозилирования обязательно заняты O-гликанами. Занятость O-гликанами зависит, в частности, от клетки-хозяина, в которой продуцируется рекомбинантный гликозилированный полипептид. Предпочтительно, чтобы клетка-хозяин, то есть система экспрессии, была выбрана таким образом, чтобы процент занятости сайтов O-гликозилирования был выше 70%. При занятости ниже 70% может присутствовать недостаточно О-гликанов для связывания SIGLEC. Предпочтительно, чтобы более 80% сайтов O-гликозилирования были заняты O-гликанами. Более предпочтительно, чтобы более 90% сайтов O-гликозилирования были заняты O-гликанами. Согласно предпочтительному варианту осуществления более 95% сайтов O-гликозилирования должны быть заняты O-гликанами.
Композиция гликанов, присоединенных к гликозилированному полипептиду, зависит от способа получения. O-гликаны могут быть природными или синтетическими гликанами. Природные O-гликаны представляют собой, например, гликаны со следующей коровой структурой:
Коровый O-гликан 1-го типа: Galβ1→3GalNAcα1→Ser/Thr
Удлиненый коровый O-гликан 1-го типа: Galβ1→4GlcNAcβ1→3Galβ1→3GalNAcα1→Ser/Thr
Коровый O-гликан 2-го типа: Galβ1→3(Galβ1→3GlcNAcβ1→6)GalNAc α1→Ser/Thr
Известно, что SIGLEC связываются с сиаловой кислотой. В соответствии с этим в примерах показано, что десиалилирование отменяет связывание с SIGLEC (см. пример 3). Таким образом подтверждается, что сиалилирование O-гликанов является обязательным для связывания. Соответственно, высокий процент сиалилирования O-гликанов в гликозилированном полипептиде является предпочтительным.
Соответственно, O-гликаны гликозилированного полипептида, предпочтительно, являются сиалилированными, то есть содержат по меньшей мере одну сиаловую кислоту в составе молекулы гликана.
Предпочтительно, чтобы сиалилированные О-гликаны содержали по меньшей мере две молекулы сиаловой кислоты в альфа-2-3 гликозидной связи. В альтернативном варианте, сиалилированные О-гликаны могут содержать две молекулы сиаловой кислоты в альфа-2-8 гликозидной связи. Сиалилированные O-гликаны также могут включать альфа-2-3 и альфа-2-8 гликозидную связь. Согласно одному варианту осуществления сиалилированные О-гликаны содержат по меньшей мере три молекулы сиаловой кислоты в 2-3 и/или 2-8 гликозидных связях. Сиалилированными O-гликанами в гликозилированном полипептиде являются, в частности, коровые 1 или коровые 2 O-гликаны. Ниже представлены общие структуры O-гликанов: коровых 1-го типа, удлиненных коровых 1-го типа и коровых 2-го типа:
Сиалилированный коровый O-гликан 1-го типа: NeuNAcα2→3Galβ1→3GalNAcα1→Ser/Thr
Сиалилированный удлиненный коровый O-гликан 1-го типа: NeuNAcα2→3Galβ1→4GlcNAcβ1→3Galβ1→3GalNAcα1→Ser/Thr
Сиалилированный коровый O-гликан 2-го типа: NeuNAcα2→3Galβ1→4GlcNAcβ1→6(NeuNAcα2→3Galβ1→3)GalNAcα1→Ser/ Thr
Возможно, что на гликозилированном полипептиде должны присутствовать О-гликаны как коровый 1-го типа, так и коровый 2-го типа и/или удлиненный коровый 1-го типа. Согласно одному варианту осуществления гликозилированный полипептид содержит сиалилированные коровые O-гликаны 1-го типа, а также сиалилированные коровые 2-го типа и/или удлиненные коровые O-гликаны 1-го типа. Как показано в примере 4, процентное содержание коровых гликанов 2-го типа, составляющее 2,5% от общего количества O-гликанов, недостаточно для взаимодействия с SIGLEC. Таким образом, согласно одному варианту осуществления гликозилированного полипептида, процент коровых O-гликанов 2-го типа от количества O-гликанов составляет по меньшей мере 5%. В том же примере показано, что кластер 2 с процентным содержанием коровых O-гликанов 2-го типа 10,78% от количества O-гликанов дает сильное взаимодействие с SIGLEC. Таким образом, согласно предпочтительному варианту осуществления изобретения, процентное содержание коровых O-гликанов 2-го типа от количества O-гликанов в гликозилированном полипептиде составляет по меньшей мере 8%. Более предпочтительно, чтобы процентное содержание коровых O-гликанов 2-го типа от количества O-гликанов составляло по меньшей мере 10%.
В примере 7 было определено, что около 80% гликопептидных молекул рекомбинантно продуцируемых vWF-пептидов содержат либо коровый O-гликан 2-го типа, либо удлиненный коровый О-гликан 1-го типа. Соответственно, процентное содержание сиалилированных коровых О-гликанов 2-го типа и/или удлиненных коровых О-гликанов 1-го типа от общего количества O-гликанов составляет по меньшей мере 20%. Таким образом, согласно одному варианту осуществления концентрация коровых О-гликанов 2-го типа и/или удлиненных коровых О-гликанов 1-го типа от общего количества O-гликанов в гликозилированном полипептиде составляет по меньшей мере 15%, более предпочтительно, по меньшей мере 18% и, наиболее предпочтительно, по меньшей мере 20%.
Количество или процентное содержание коровых O-гликанов 2-го типа в гликозилированном полипептиде, в частности, количество или процентное содержание индивидуальных молекул гликозилированного полипептида, несущих коровый O-гликан 2-го типа, может быть увеличено с помощью любой из следующих стратегий:
Одна из стратегий заключается в использовании фермента β-1,6-N-ацетилглюкозаминилтрансферазы. Этот фермент участвует в образовании коровых O-гликанов 2-го типа. Таким образом, увеличение числа несущих коровые O-гликаны 2-го типа молекул и/или числа коровых O-гликанов 2-го типа на молекулу полипептида может быть получено экспрессией гликозилированного полипептида в клеточной линии, которая сверхэкспрессирует фермент β-1,6-N-ацетилглюкозаминилтрансферазу.
Другим вариантом является экспрессия гликозилированного полипептида в экспрессионной клеточной линии, полученной из линии раковых клеток. Показано, что часто раковые клеточные линии продуцируют гликозилированные белки с большим количеством коровых O-гликанов 2-го типа.
Одна из стратегий увеличения процентного содержания удлиненных коровых O-гликанов 1-го типа заключается в использовании фермента β-1,3-N-ацетилглюкозаминилтрансферазы. Этот фермент участвует в образовании удлиненных коровых O-гликанов 1-го типа. Таким образом, увеличение числа молекул, несущих коровые удлиненые О-гликаны 1-го типа, и/или числа удлиненных коровых O-гликанов 1-го типа на молекулу полипептида может быть получено экспрессией гликозилированного полипептида в клеточной линии, которая сверхэкспрессирует фермент β-1,6-N-ацетилглюкозаминилтрансферазу.
Концентрация сиалилированных коровых O-гликанов 2-го типа и/или удлиненных коровых O-гликанов 1-го типа может быть увеличена путем химического синтеза гликанов.
Соответственно, на основе идей изобретения специалист может адаптировать гликаны для создания синтетических гликанов с высокой аффинностью связывания. Согласно предпочтительному варианту осуществления O-гликаны являются природными гликанами.
Согласно одному варианту осуществления, по меньшей мере часть сиалилированных коровых O-гликанов 2-го типа в гликозилированном полипептиде содержит сульфатную группу, присоединенную к галактозе (Gal) или N-ацетилглюкозамину (GlcNAc), или фукозу, присоединенную к GlcNAc в коровом O-гликане 2-го типа, который представляет собой высокоаффинный связывающий лиганд для SIGLEC 7, 8 и 8 (Paulson et al., 2012).
Специалисту в данной области известны различные способы создания дополнительных сайтов O-гликозилирования. Гликозилированный полипептид может иметь увеличенное количество сиалилированных коровых O-гликанов 2-го типа из-за увеличения количества сайтов O-гликозилирования по сравнению с белком человека или его фрагментом.
В этом случае гликозилированный полипептид может представлять собой слитый белок, где вторая аминокислотная последовательность, содержащая один или несколько сайтов O-гликозилирования, ковалентно связана с аминокислотной последовательностью, идентичной или гомологичной белку или фрагменту белка человека (первая аминокислотная последовательность). Вторая аминокислотная последовательность может быть расположена в N-конце по отношению к первой аминокислотной последовательности. В альтернативном варианте вторая аминокислотная последовательность может быть расположена в С-конце относительно первой аминокислотной последовательности. Гликозилированный полипептид может содержать дополнительные аминокислотные последовательности - как на N-, так и на С-конце относительно первой аминокислотной последовательности.
Соответственно, в таком слитом белке могут присутствовать вторая аминокислотная последовательность и, необязательно, дополнительные аминокислотные последовательности, которые в основном содержат сайты О-гликозилирования, в частности, кластеры О-гликозилирования. Кластер сайтов O-гликозилирования может быть основан на аминокислотной последовательности от известного млекопитающего, в частности, на гликозилированном белке человека.
Вторая аминокислотная последовательность может содержать один или несколько из следующих кластеров О-гликозилирования:
VVPPTXAPVXPTTXYVXXXSXPP (SEQ ID NO: 8),
VVPPTDAPVSPTTLYVEDISEPP (SEQ ID NO: 9),
PPPTXPPXXAXVTVXPXXXXVSTXXP (SEQ ID NO: 10),
PPPTLPPDMAQVTVGPGLLGVSTLGP (SEQ ID NO: 11),
VSSTSXXXXSTXPSXXXAAXTXXTSSXXPPSXPVXXXSXXXTTXXXX (SEQ ID NO: 12),
VSSTSNNLISTIPSDNLAAGTDDTSSLGPPSMPVHYDSQLDTTLFGK (SEQ ID NO: 13),
XXXATTXPXXXXXXTXPXXX (SEQ ID NO: 14),
QFNATTIPENDIEKTDPWFA (SEQ ID NO: 15),
XXTTAATXXX (SEQ ID NO: 16),
LGTTAATELK (SEQ ID NO: 17),
XXPTPXXXSXSXXXEAX (SEQ ID NO: 18),
QSPTPHGLSLSDLQEAK (SEQ ID NO: 19);
VXXXXXXXXXTXTSXXSPXXXXXVXXSXXXXTXXAXX (SEQ ID NO: 20) и
VHIYQKDLFFTETSDGSPGHLDLVEGSLLQGTEGAIK (SEQ ID NO: 21).
В последовательностях SEQ ID NO: 8,10,12, 14, 16, 18 и 20, Х означает любую из природных аминокислот.
SEQ ID NO: 9 и 11 обнаружены в vWF, а SEQ ID NO: 13,15,17, 19 и 21 получены из B-домена FVIII.
Вторая аминокислотная последовательность может содержать одну или несколько последовательностей, выбранных из SEQ ID NO: 8, 10, 12, 14, 16, 18 и 20. Вторая аминокислотная последовательность может содержать комбинации последовательностей. Вторая аминокислотная последовательность, предпочтительно, содержит множество копий одной из последовательностей SEQ ID NO: 8, 10, 12, 14, 16, 18 и 20. Вторая аминокислотная последовательность может дополнительно содержать комбинации множества копий SEQ ID NO: 8, 10, 12, 14, 16, 18 и 20
Вторая аминокислотная последовательность может содержать одну или несколько последовательностей, выбранных из SEQ ID NO: 9, 11, 13, 15, 17, 19 и 21. Вторая аминокислотная последовательность может содержать комбинации последовательностей. Вторая аминокислотная последовательность, предпочтительно, содержит несколько копий одной из последовательностей SEQ ID NO: 9, 11, 13, 15, 17, 19 и 21. Вторая аминокислотная последовательность может дополнительно содержать комбинации множества копий SEQ ID NO: 9, 11, 13, 15, 17, 19 и 21.
Согласно предпочтительному варианту осуществления вторая аминокислотная последовательность содержит одну или несколько копий SEQ ID NO: 8.
Кроме того, вторая аминокислотная последовательность может иметь некоторый процент идентичности с последовательностью природного гликозилированного белка. Уровень идентичности с природным белком, предпочтительно, составляет 80%, более предпочтительно, по меньшей мере 90%.
В альтернативном варианте, аминокислотная последовательность сайтов O-гликозилирования во второй аминокислотной последовательности может быть полностью синтетической. Полностью синтетическая аминокислотная последовательность, используемая в настоящем изобретении, представляет собой последовательность, которая не основана на известном белке, в частности, на белке млекопитающего.
Согласно одному варианту осуществления настоящего изобретения, ковалентный линкер, соединяющий вторую аминокислотную последовательность с аминокислотной последовательностью, идентичной или гомологичной белку человека или его фрагменту в гликозилированном полипептиде, выбирают из пептидной связи, химического линкера или гликозидной связи. Химическими линкерами, подходящими для этого, являются:
- Амин с аминными линкерами, такими как бисмалеимидоэтан, 1,8-бисмалеимидо-диэтиленгликоль,
- Амин с сульфгидрильными линкерами, такими как сукцинимидилиодацетат, N-α-малеимидоацетамидо-оксисукцинимидный эфир,
- Карбоксил с аминными линкерами дициклогексилкарбодиимид, гидрохлорид 1-этил-3-(3-диметиламинопропил)карбодиимида и
- Сульфгидрил с углеводными линкерами, такими как гидразид N-β-малеимидопропионовой кислоты, гидразид N-ε-малеимидокапроновой кислоты.
Линкер слитого белка по изобретению может быть образован спейсерной пептидной последовательностью, разделяющей первую и вторую аминокислотную последовательность, которые образуют слитый белок. Спейсерная пептидная последовательность может способствовать правильной укладке отдельных частей белка или пептида и может обеспечить большую вероятность сохранения индивидуальных функциональных свойств для индивидуальных частей белка или пептида. Спейсерные пептидные последовательности могут быть вставлены в последовательности ДНК слитого белка во время сборки с сохранением рамки считывания отдельных фрагментов ДНК, которые составляют полную последовательность ДНК слитого белка, например, во время перекрывающейся ПЦР или лигирования ДНК.
Преимущество пептидной связи состоит в том, что полный гликозилированный полипептид может сразу экспрессироваться в виде слитого белка.
Вторая аминокислотная последовательность может быть добавлена к первой аминокислотной последовательности с помощью химического линкероа и, следовательно, после экспрессии белка.
Как показано в примерах, в частности, vWF и его фрагменты, содержащие кластеры 1 и/или 2 O-гликозилирования, связываются с SIGLEC.
Таким образом, согласно одному варианту осуществления белок человека представляет собой FVIII или его фрагмент. Соответственно, идентичность последовательности с FVIII в гликозилированном полипептиде составляет по меньшей мере 90%, более предпочтительно, по меньшей мере 95% и, наиболее предпочтительно, по меньшей мере 98%. Согласно предпочтительному варианту осуществления первая аминокислотная последовательность идентична или гомологична аминокислотам с 1 по 505 из SEQ ID No: 1.
Длина второй аминокислотной последовательности, предпочтительно, находится в интервале от 5 до 100 аминокислот, более предпочтительно, от 10 до 80 аминокислот, наиболее предпочтительно, от 20 до 70 аминокислот.
Согласно одному варианту осуществления вторая аминокислотная последовательность по меньшей мере на 98% гомологична аминокислотам 475-505 из SEQ ID NO: 1. Предпочтительно, чтобы вторая аминокислотная последовательность была идентична аминокислотам 475-505 из SEQ ID NO: 1. В соответствии с более предпочтительным вариантом осуществления вторая аминокислотная последовательность по меньшей мере на 98% гомологична двум последовательным копиям аминокислот 475-505 из SEQ ID NO: 1. Предпочтительно, чтобы вторая аминокислотная последовательность была идентична двум последовательным копиям аминокислот 475-505 из SEQ ID NO: 1.
Примером химерного белка в соответствии с настоящим изобретением является Seq12. Seq12 имеет следующую аминокислотную последовательность (SEQ ID NO: 2):
SLSCRPPMVKLVCPADNLRAEGLECTKTCQNYDLECMSMGCVSGCLCPPGMVRHENRCVALERCPCFHQGKEYAPGETVKIGCNTCVCQDRKWNCTDHVCDATCSTIGMAHYLTFDGLKYLFPGECQYVLVQDYCGSNPGTFRILVGNKGCSHPSVKCKKRVTILVEGGEIELFDGEVNVKRPMKDETHFEVVESGRYIILLLGKALSVVWDRHLSISVVLKQTYQEKVCGLCGNFDGIQNNDLTSSNLQVEEDPVDFGNSWKVSSQCADTRKVPLDSSPATCHNNIMKQTMVDSSCRILTSDVFQDCNKLVDPEPYLDVCIYDTCSCESIGDCACFCDTIAAYAHVCAQHGKVVTWRTATLCPQSCEERNLRENGYECEWRYNSCAPACQVTCQHPEPLACPVQCVEGCHAHCPPGKILDELLQTCVDPEDCPVCЕVAGRRFASGKKVTLNPSDPEHCQICHCDVVNLTCEACQEPGGLVVPPTDAPVSPTTLYVEDISEPPLHQEPGGLVVPPTDAPVSPTTLYVEDISEPPLHQEPGGLVVPPTDAPVSPTTLYVEDISEPPLH
Следующая последовательность (SEQ ID NO: 3) представляет Seq12 с дополнительным 22-аминокислотным сигнальным пептидом (выделен полужирным шрифтом и подчеркнут). Экспрессия этого пептида обеспечивает мономерную форму Seq12. Сигнальный пептид отщепляется ферментативно.
MIPARFAGVLLALALILPGTLC SLSCRPPMVKLVCPADNLRAEGLECTKTCQNYDLECMSMGCVSGCLCPPGMVRHENRCVALERCPCFHQGKEYAPGETVKIGCNTCVCQDRKWNCTDHVCDATCSTIGMAHYLTFDGLKYLFPGECQYVLVQDYCGSNPGTFRILVGNKGCSHPSVKCKKRVTILVEGGEIELFDGEVNVKRPMKDETHFEVVESGRYIILLLGKALSVVWDRHLSISVVLKQTYQEKVCGLCGNFDGIQNNDLTSSNLQVEEDPVDFGNSWKVSSQCADTRKVPLDSSPATCHNNIMKQTMVDSSCRILTSDVFQDCNKLVDPEPYLDVCIYDTCSCESIGDCACFCDTIAAYAHVCAQHGKVVTWRTATLCPQSCEERNLRENGYECEWRYNSCAPACQVTCQHPEPLACPVQCVEGCHAHCPPGKILDELLQTCVDPEDCPVCЕVAGRRFASGKKVTLNPSDPEHCQICHCDVVNLTCEACQEPGGLVVPPTDAPVSPTTLYVEDISEPPLHQEPGGLVVPPTDAPVSPTTLYVEDISEPPLHQEPGGLVVPPTDAPVSPTTLYVEDISEPPLH (SEQ ID NO: 3)
Дополнительный пример слитого белка по изобретению представляет собой Pro-Seq12, включающий Seq12 и пропептид (выделен полужирным шрифтом) с сигнальным пептидом (выделен полужирным шрифтом и подчеркнут). Pro-Seq12 приведен как SEQ ID NO: 4:
MIPARFAGVLLALALILPGTLCAEGTRGRSSTARCSLFGSDFVNTFDGSMYSFAGYCSYLLAGGCQKRSFSIIGDFQNGKRVSLSVYLGEFFDIHLFVNGTVTQGDQRVSMPYASKGLYLETEAGYYKLSGEAYGFVARIDGSGNFQVLLSDRYFNKTCGLCGNFNIFAEDDFMTQEGTLTSDPYDFANSWALSSGEQWCERASPPSSSCNISSGEMQKGLWEQCQLLKSTSVFARCHPLVDPEPFVALCEKTLCECAGGLECACPALLEYARTCAQEGMVLYGWTDHSACSPVCPAGMEYRQCVSPCARTCQSLHINEMCQERCVDGCSCPEGQLLDEGLCVESTECPCVHSGKRYPPGTSLSRDCNTCICRNSQWICSNEECPGECLVTGQSHFKSFDNRYFTFSGICQYLLARDCQDHSFSIVIETVQCADDRDAVCTRSVTVRLPGLHNSLVKLKHGAGVAMDGQDVQLPLLKGDLRIQHTVTASVRLSYGEDLQMDWDGRGRLLVKLSPVYAGKTCGLCGNYNGNQGDDFLTPSGLAEPRVEDFGNAWKLHGDCQDLQKQHSDPCALNPRMTRFSEEACAVLTSPTFEACHRAVSPLPYLRNCRYDVCSCSDGRECLCGALASYAAACAGRGVRVAWREPGRCELNCPKGQVYLQCGTPCNLTCRSLSYPDEECNEACLEGCFCPPGLYMDERGDCVPKAQCPCYYDGEIFQPEDIFSDHHTMCYCEDGFMHCTMSGVPGSLLPDAVLSSPLSHRSKRSLSCRPPMVKLVCPADNLRAEGLECTKTCQNYDLECMSMGCVSGCLCPPGMVRHENRCVALERCPCFHQGKEYAPGETVKIGCNTCVCQDRKWNCTDHVCDATCSTIGMAHYLTFDGLKYLFPGECQYVLVQDYCGSNPGTFRILVGNKGCSHPSVKCKKRVTILVEGGEIELFDGEVNVKRPMKDETHFEVVESGRYIILLLGKALSVVWDRHLSISVVLKQTYQEKVCGLCGNFDGIQNNDLTSSNLQVEEDPVDFGNSWKVSSQCADTRKVPLDSSPATCHNNIMKQTMVDSSCRILTSDVFQDCNKLVDPEPYLDVCIYDTCSCESIGDCACFCDTIAAYAHVCAQHGKVVTWRTATLCPQSCEERNLRENGYECEWRYNSCAPACQVTCQHPEPLACPVQCVEGCHAHCPPGKILDELLQTCVDPEDCPVCEVAGRRFASGKKVTLNPSDPEHCQICHCDVVNLTCEACQEPGGLVVPPTDAPVSPTTLYVEDISEPPLHQEPGGLVVPPTDAPVSPTTLYVEDISEPPLHQEPGGLVVPPTDAPVSPTTLYVEDISEPPLH (SEQ ID NO: 4)
Экспрессия Pro-Seq12 приводит к образованию димеров. Пептидные димеры остаются также после расщепления пропептида.
Согласно одному варианту осуществления изобретения, гликозилированные полипептиды содержат первую аминокислотную последовательность, которая по меньшей мере на 98% идентична аминокислотам 1-505 из SEQ ID NO: 1. Вторая аминокислотная последовательность по меньшей мере на 98% гомологична двум последовательным копиям аминокислот 475-505 из SEQ ID NO: 1.
Согласно одному варианту осуществления изобретения, гликозилированный полипептид получают путем экспрессии в линии клеток человека. Обычно для экспрессии гликозилированного полипептида подходит любая линия клеток человека. Пептид с желательным гликозилированием, в частности, получают с использованием клеточных линий HEK.
Примерами клеточных линий HEK для получения гликозилированного полипептида являются HEK 293 F, Flp-In™-293 (Invitrogen, R75007), 293 (ATCC® CRL-1573), 293 EBNA, 293 H (ThermoScientific 11631017), 293S, 293T (ATCC® CRL-3216™), 293T/17 (ATCC® CRL11268™), 293T/17 SF (ATCC® ACS4500™), HEK 293 STF (ATCC® CRL 3249™), HEK-293.2sus (ATCC® CRL-1573™). Предпочтительной клеточной линией для получения полипептида является HEK 293 F.
Другие клеточные линии, подходящие в качестве клеток-хозяев для экспрессии, включают клеточные линии, происходящие из клеток миелоидного лейкоза человека. Конкретными примерами клеток-хозяев являются K562, NM-F9, NM-D4, NM-H9D8, NM-H9D8-E6, NM H9D8-E6Q12, GT-2X, GT-5s и клетки, полученные из любых из указанных клеток-хозяев. К562 представляет собой клеточную линию миелоидного лейкоза человека, содержащуюся в Американской коллекции типовых культур (АТСС CCL-243). Остальные клеточные линии получены из клеток К562 и были отобраны по определенным особенностям гликозилирования.
Согласно альтернативному варианту осуществления изобретения, один или несколько сайтов гликозилирования находятся в пределах аминокислотной последовательности, гомологичной или идентичной белку млекопитающего или его фрагменту. Следует понимать, что сайты O-гликозилирования, которые не присутствуют в аминокислотной последовательности белка млекопитающего или его фрагмента, находятся в гомологичной или идентичной аминокислотной последовательности в гликозилированном полипептиде.
Один или несколько сайтов О-гликозилирования в аминокислотной последовательности, гомологичной или идентичной белку млекопитающего или его фрагменту, могут быть встроены в последовательность. В альтернативном варианте одним или несколькими сайтами O-гликозилирования можно заменить аминокислоты белка млекопитающего. Аминокислотная замена является предпочтительной, поскольку она не изменяет размер полипептидной цепи и, следовательно, менее вероятно повлияет на трехмерную структуру белка.
Один или несколько сайтов O-гликозилирования в аминокислотной последовательности, предпочтительно, находятся в частях последовательности, которые не образуют сайтов связывания или активных центров белка. Кроме того, один или несколько сайтов O-гликозилирования, предпочтительно, добавляют в аминокислотную позицию, которая будет экспонироваться на поверхности свернутого белка. Чтобы добиться наименьшего влияния на активность или целостность белка, один или несколько сайтов O-гликозилирования могут быть добавлены в гибкую петлю белка.
В случае белков FVIII, один или несколько сайтов O-гликозилирования, предпочтительно, добавляют в В-домен или замещая В-домен.
В одном варианте осуществления гликозилированный полипептид модифицируют путем присоединения одного или нескольких биосовместимых полимеров для увеличения, например, времени полужизни или стабильности. Подходящие биосовместимые полимеры включают полиалкиленоксиды, такие как, без ограничения, полиэтиленгликоль (ПЭГ), декстраны, коломиновые кислоты или другие полимеры на основе углеводов, полимеры аминокислот, производные биотина, поливиниловый спирт (PVA), поликарбоксилаты, поливинилпирролидон, сополимер полиэтилен/ангидрид малеиновой кислоты, сополимер полистирол/ангидрид яблочной кислоты, полиоксазолин, полиакрилоилморфолин, гепарин, альбумин, целлюлозы, гидролизаты хитозана, крахмалы, такие как гидроксиэтилкрахмалы и гидроксипропилкрахмалы, гликоген, агарозы и их производные, гуаровая камедь, пуллулан, инулин, ксантановая камедь, каррагенан, пектин, гидролизаты альгиновой кислоты, другие биополимеры и любые их эквиваленты. В одном варианте осуществления полимер представляет собой полиэтиленгликоль (ПЭГ). В другом варианте осуществления полимер представляет собой метоксиполиэтиленгликоль (мПЭГ). Другими полезными соединениями полиалкиленгликоля являются полипропиленгликоли (PPG), полибутиленгликоли (PBG), PEG-глицидиловые эфиры (Epox-PEG), PEG-оксикарбонилимидазол (CDI-PEG), разветвленные полиэтиленгликоли, линейные полиэтиленгликоли, раздвоенные полиэтиленгликоли и мультиразветвленные или «суперразветвленные» полиэтиленгликоли (звездчатые ПЭГ). Биосовместимый полимер, предпочтительно, связан с полипептидом через один из следующих остатков: -SH, OH, -COOH.
Согласно одному варианту осуществления гликозилированный полипептид способен образовывать димеры или мультимеры. Образование димеров и, в частности, мультимеров увеличивает количество близкорасположенных O-гликанов или кластеров O-гликанов. Таким образом, большее количество O-гликанов может взаимодействовать с SIGLEC на одной клетке. Более того, O-гликаны могут взаимодействовать с несколькими экспрессирующими SIGLEC клетками, которые расположены близко друг к другу, тем самым повышая иммунную толерантность.
Мультимеризация может являться результатом присутствия домена мультимеризации в аминокислотной последовательности белка млекопитающего, на которой основан гликозилированный полипептид. В альтернативном варианте, мультимеры гликозилированного полипептида могут быть образованы путем введения доменов мультимеризации в аминокислотную последовательность гликозилированного полипептида.
Примером гибридного белка по изобретению, который образует мультимеры, является Pro-Seq12-Mult, включающий Seq12 и пропептид (выделены полужирным шрифтом) с сигнальным пептидом (выделен полужирным шрифтом и подчеркнут), а также мультимеризующуюся последовательность (домен «цистеиновый узел») из vWF (подчеркнута). Pro-Seq12-Mult указан как SEQ ID NO: 5:
MIPARFAGVLLALALILPGTLCAEGTRGRSSTARCSLFGSDFVNTFDGSMYSFAGYCSYLLAGGCQKRSFSIIGDFQNGKRVSLSVYLGEFFDIHLFVNGTVTQGDQRVSMPYASKGLYLETEAGYYKLSGEAYGFVARIDGSGNFQVLLSDRYFNKTCGLCGNFNIFAEDDFMTQEGTLTSDPYDFANSWALSSGEQWCERASPPSSSCNISSGEMQKGLWEQCQLLKSTSVFARCHPLVDPEPFVALCEKTLCECAGGLECACPALLEYARTCAQEGMVLYGWTDHSACSPVCPAGMEYRQCVSPCARTCQSLHINEMCQERCVDGCSCPEGQLLDEGLCVESTECPCVHSGKRYPPGTSLSRDCNTCICRNSQWICSNEECPGECLVTGQSHFKSFDNRYFTFSGICQYLLARDCQDHSFSIVIETVQCADDRDAVCTRSVTVRLPGLHNSLVKLKHGAGVAMDGQDVQLPLLKGDLRIQHTVTASVRLSYGEDLQMDWDGRGRLLVKLSPVYAGKTCGLCGNYNGNQGDDFLTPSGLAEPRVEDFGNAWKLHGDCQDLQKQHSDPCALNPRMTRFSEEACAVLTSPTFEACHRAVSPLPYLRNCRYDVCSCSDGRECLCGALASYAAACAGRGVRVAWREPGRCELNCPKGQVYLQCGTPCNLTCRSLSYPDEECNEACLEGCFCPPGLYMDERGDCVPKAQCPCYYDGEIFQPEDIFSDHHTMCYCEDGFMHCTMSGVPGSLLPDAVLSSPLSHRSKRSLSCRPPMVKLVCPADNLRAEGLECTKTCQNYDLECMSMGCVSGCLCPPGMVRHENRCVALERCPCFHQGKEYAPGETVKIGCNTCVCQDRKWNCTDHVCDATCSTIGMAHYLTFDGLKYLFPGECQYVLVQDYCGSNPGTFRILVGNKGCSHPSVKCKKRVTILVEGGEIELFDGEVNVKRPMKDETHFEVVESGRYIILLLGKALSVVWDRHLSISVVLKQTYQEKVCGLCGNFDGIQNNDLTSSNLQVEEDPVDFGNSWKVSSQCADTRKVPLDSSPATCHNNIMKQTMVDSSCRILTSDVFQDCNKLVDPEPYLDVCIYDTCSCESIGDCACFCDTIAAYAHVCAQHGKVVTWRTATLCPQSCEERNLRENGYECEWRYNSCAPACQVTCQHPEPLACPVQCVEGCHAHCPPGKILDELLQTCVDPEDCPVCEVAGRRFASGKKVTLNPSDPEHCQICHCDVVNLTCEACQEPGGLVVPPTDAPVSPTTLYVEDISEPPLHQEPGGLVVPPTDAPVSPTTLYVEDISEPPLHQEPGGLVVPPTDAPVPTTLYVEDISEPPLHEEPECNDITARLQYVKVGSCKSEVEVDIHYCQGKCASKAMYSIDINDVQDQCSCCSPTRTEPMQVALHCTNGSVVYHEVLNAMECKCSPRKCSK
В альтернативном варианте мультимеризация гликозилированного полипептида может быть достигнута путем связывания с полимером или липосомой.
Применение гликозилированного полипептида
Как описано выше, авторы изобретения обнаружили, что белок с гликановой композицией, включающий в себя сиалилированные коровые O-гликаны 2-го типа и/или сиалилированные удлиненные коровые O-гликаны 1-го типа, взаимодействует с определенными SIGLEC и, следовательно, влияет на клетки иммунной системы млекопитающих. В частности, гликозилированный полипептид вызывает пониженный иммунный ответ. Поэтому, такой гликозилированный полипептид при введении с вторым белком может влиять на иммунный ответ пациента на второй белок. Поэтому, гликозилированный полипептид с сиалилированными коровыми O-гликанами 2-го типа и/или сиалилированными удлиненными коровыми О-гликанами 1-го типа может быть использован для модификации, в частности, для уменьшения иммунного ответа пациента на белок, в частности, терапевтический белок при совместном введении.
Таким образом, согласно второму аспекту изобретение относится к применению гликозилированного полипептида, содержащего один или несколько сиалилированных O-гликанов, и к связыванию с одним или несколькими SIGLEC, выбранными из SIG-5, SIG-7, SIG-8 и SIG-9, для снижения иммунного ответа на терапевтический белок.
Предпочтительно, чтобы гликозилированный полипептид, используемый для уменьшения иммунного ответа, содержал сиалилированные коровые O-гликаны 2-го типа и/или сиалилированные удлиненные короые O-гликаны 1-го типа. Более предпочтительно, чтобы гликозилированный полипептид представлял собой гликозилированный полипептид по первому аспекту.
Применение также может быть описано как способ лечения пациента терапевтическим белком, где способ включает введение гликозилированного полипептида, содержащего один или несколько сиалилированных O-гликанов, и связывание с одним или несколькими SIGLEC, выбранными из SIG-5, SIG-7, SIG-8 и SIG-9, для снижения иммунного ответа на терапевтический белок.
Композиция и белковый комплекс
Соответственно, концепция изобретения, а именно, пониженный иммунный ответ на белок человека, например, путем добавления одного или нескольких сиалилированных коровых O-гликанов 2-го типа может быть достигнута не только получением слитого белка или вставкой или заменой аминокислот сайтами O-гликозилирования, но также путем добавления дополнительного полипептида, как описано в применении по второму аспекту. Это приводит к образованию композиции гликозилированного полипептида и второго полипептида, иммунный ответ на который должен быть снижен.
Соответственно, в третьем аспекте настоящее изобретение также относится к композиции, содержащей первый и второй полипептиды, где первый полипептид представляет собой гликозилированный полипептид, содержащий один или несколько сиалилированных О-гликанов, а второй полипептид содержит аминокислотную последовательность, гомологичную или идентичную второму белку млекопитающего, в частности, человека, где по сравнению со вторым полипептидом композиция обладает повышенной аффинностью связывания с SIGLEC, выбранным из одного или нескольких SIGLEC, выбранных из SIG-5, SIG-7, SIG-8 и SIG-9.
Также можно создать связывающего партнера - полипептид, который обладает повышенной аффинностью связывания с SIGLEC, так что комплекс двух полипептидов будет обладать повышенной аффинностью связывания с SIGLEC по сравнению с полипептидом.
Таким образом, в соответствии с третьим аспектом изобретение относится к композиции, содержащей первый и второй полипептиды, причем первый полипептид представляет собой гликозилированный полипептид, содержащий один или несколько сиалилированных О-гликанов, а второй полипептид содержит аминокислотную последовательность, гомологичную или идентичную второму белку млекопитающего, в частности, человека, где по сравнению со вторым полипептидом:
- композиция обладает повышенной аффинностью связывания с SIGLEC, выбранным из; и/или
- иммунный ответ человека на комплекс снижен; и/или
- иммунная толерантность человека к комплексу повышена.
Второй белок человека, предпочтительно, представляет собой белок крови человека. Белок крови человека может представлять собой фактор свертываемости крови человека, транспортный белок, ингибитор протеаз, иммуноглобулин, клеточный белок плазмы, аполипопротеины, фактор комплемента, фактор роста, антиангиогенный белок, высокогликозилированный белок, факторы крови или другой белок крови человека
Фактор свертываемости крови человека выбирают, в частности, из группы, состоящей из фибриногена, фибринового мономера, протромбина, тромбина, FV/FVa, FX/FXa, FIX/FIXa, FVII/FVIIa, FVIII/FVIIIa, FXI/FXIa, FXII/FXIIa, FXIII/FXIIIa, фактора фон Виллебранда и ADAMTS13.
Транспортный белок может быть выбран из альбумина, трансферрина, церулоплазмина, гаптоглобина, гемоглобина и гемопексина.
Потенциальными ингибиторами протеаз являются, например, β-антитромбин, α-антитромбин, окисленный-антитромбин, 2-макроглобулин, C1-ингибитор, ингибитор пути тканевого фактора (TFPI), кофактор гепарина II, ингибитор белка С (PAI-3), белок С, белок S и белок Z.
Примеры иммуноглобулинов являются, например, поликлональные антитела (IgG), моноклональные антитела, IgG1, IgG2, IgG3, IgG4, IgA, IgA1, IgA2, IgM, IgE, IgD и белок Бенс-Джонса.
Клеточным белком плазмы может быть, например, фибронектин, тромбоглобулин, тромбоцитарный фактор 4. Примерами аполипопротеинов являются apo A-I, apo A-II и apo E.
Факторами комплемента согласно изобретению являются, например, фактор В, фактор D, фактор H, фактор I, C3b-инактиватор, пропердин, C4-связывающий белок и т.д.
Примеры ростовых факторов включают фактор роста, полученный из тромбоцитов (PDGF), эпидермальный фактор роста (EGF), трансформирующий фактор роста-альфа (TGF-α), трансформирующий фактор роста-бета (TGF-β), фактор роста фибробластов (FGF) и фактор роста гепатоцитов.
Антиангиогенные белки включают в себя латентный антитромбин, прелатентный антитромбин, окисленный антитромбин и плазминоген.
Примерами высоко гликозилированных белков являются альфа-1-кислотный гликопротеин, антихимотрипсин, интер-α-трипсин-ингибитор, α-2-HS-гликопротеин, С-реактивный белок. Факторами крови могут быть, например, эритропоэтин, интерферон, опухолевые факторы, tPA, gCSF.
Другие белки крови человека включают богатый гистидином гликопротеин, маннан-связывающий лектин, С4-связывающий белок, фибронектин, GC-глобулин, плазминоген/плазмин, α-1-микроглобулин, C-реактивный белок.
Второй белок человека, в частности, выбирают из vWF, FVIII, FVII/FVIIa, FIX, ADAMTS13.
Композиция согласно третьему аспекту представляет собой, в частности, белковый комплекс первого и второго полипептида.
Первый полипептид, предпочтительно, является гликозилированным и содержит один или несколько сиалилированных О-гликанов. Второй полипептид содержит аминокислотную последовательность, идентичную белку млекопитающего, в частности, человека. Для уменьшения иммунного ответа человека на второй полипептид первый полипептид образует комплекс со вторым полипептидом.
Для образования белкового комплекса первый полипептид, в частности, содержит связывающий домен, который обеспечивает связывание со вторым полипептидом, и домен гликозилирования. Домен гликозилирования, в частности, содержит один или несколько сайтов O-гликозилирования, предпочтительно, кластер O-гликозилирования.
В соответствии с одним вариантом осуществления изобретения второй полипептид представляет собой белок FVIII, а первый полипептид содержит домен связывания FVIII с vWF и один или несколько сайтов O-гликозилирования, с которыми связаны сиалилированные коровые O-гликаны 2-го типа.
Предпочтительным примером композиции является комплекс белка FVIII с аминокислотной последовательностью на 95% идентичной последовательности, определяемой аминокислотами с 20-й по 2351-ю из р00451, и первого полипептида в качестве связывающего партнера, содержащего аминокислотную последовательность по меньшей мере на 95% идентичную аминокислотам 1-172 из SEQ ID NO: 1.
Согласно одному варианту белкового комплекса по третьему аспекту первый полипептид представляет собой полипептид по первому аспекту.
Согласно еще одному варианту осуществления второй полипептид может быть выбран, например, из FVIII, FVII, FIX и ADAMTS13
В одном варианте осуществления третьего аспекта, первый полипептид содержит по меньшей мере фрагмент vWF человека, а второй полипептид представляет собой белок FVIII, в частности, полноразмерный белок FVIII, белок FVIII с удаленным В-доменом или белок FVIII, в котором часть В-домена заменена линкером. Согласно одному варианту осуществления первый полипептид определяется аминокислотами 1-505 из SEQ ID NO: 1, который продуцируется в клетках HEK, в частности, в клетках HEK 293F.
Согласно еще одному варианту осуществления изобретения, первый полипептид определяется аминокислотами 475-505 из SEQ ID NO: 1, и одной копией аминокислот 475-505 из SEQ ID NO: 1. Кроме того, первый полипептид может определяться аминокислотной последовательностью, выбранной из SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 4 и SEQ ID NO: 5.
Как показано в примерах, белок с повышенной аффинностью связывания с SIGLEC, в частности, с SIG-5, SIG-7, SIG-8 и/или SIG-9, может быть получен в клеточной линии HEK 293F. Таким образом, в соответствии с дополнительным вариантом осуществления белкового комплекса первый и второй полипептиды продуцируются с помощью рекомбинантной экспрессии в линии клеток человека, предпочтительно, в клеточной линии HEK. Примерами клеточных линий НЕК для получения гликозилированного полипептида являются HEK 293 F, Flp-In™-293, 293, 293 EBNA, 293 H, 293S, 293T, 293T/17, 293T/17 SF, HEK 293 STF и HEK-293.2sus. Предпочтительной клеточной линией для получения полипептида является HEK 293 F.
Первый и второй полипептиды могут быть получены путем раздельной рекомбинантной экспрессии и после этого соединены. В альтернативном варианте первый и второй полипептиды рекомбинантно экспрессируются в одной клетке. Для этого первый и второй полипептиды могут быть закодированы в одном векторе или в двух различных векторах.
Полинуклеотид
В соответствии с четвертым аспектом настоящее изобретение относится к выделенному полинуклеотиду, который содержит последовательность нуклеиновой кислоты, кодирующую гликозилированный полипептид согласно первому аспекту изобретения.
Выделенный полинуклеотид может представлять собой молекулу ДНК или молекулу РНК. Выделенный полинуклеотид, предпочтительно, представляет собой молекулу ДНК, в частности, молекулу кДНК. Методы, используемые для выделения или клонирования полинуклеотида, кодирующего пептид, известны в данной области и включают выделение из геномной ДНК, получение из кДНК или их комбинацию. Клонирование полинуклеотидов из такой геномной ДНК может быть осуществлено, например, с использованием хорошо известной полимеразной цепной реакции (ПЦР) или скрининга экспрессирующих библиотек антител для обнаружения клонированных фрагментов ДНК с общими структурными особенностями (см., например, Innis et al, 1990 PCR: A Guide to Methods and Application, Academic Press, New York). Могут быть использованы другие процедуры амплификации нуклеиновых кислот, такие как лигазная цепная реакция (LCR), активированная лигированием транскрипция (LAT) и амплификация на основе полинуклеотидов (NASBA).
Выделенный полинуклеотид может представлять собой молекулу ДНК, кодирующую гликозилированный полипептид с аминокислотной последовательностью, аналогичной или идентичной последовательности, выбранной из группы, состоящей из SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 4, SEQ ID NO: 5.
В частности, выделенный полинуклеотид может представлять собой молекулу ДНК, кодирующую гликозилированный полипептид, имеющий аминокислотную последовательность с идентичностью по меньшей мере на 90%, предпочтительно, по меньшей мере на 95%, более предпочтительно, по меньшей мере на 98%, наиболее предпочтительно, на 100% с SEQ ID NO: 2. Кроме того, выделенный полинуклеотид может представлять собой молекулу ДНК, кодирующую гликозилированный полипептид, имеющий аминокислотную последовательность с идентичностью по меньшей мере на 90%, предпочтительно, по меньшей мере на 95%, более предпочтительно, по меньшей мере на 98%, наиболее предпочтительно, на 100% с SEQ ID NO: 3. Согласно одному варианту осуществления изобретения, выделенный полинуклеотид представляет собой молекулу ДНК, кодирующую гликозилированный полипептид, имеющий аминокислотную последовательность с идентичностью по меньшей мере на 90%, предпочтительно, по меньшей мере на 95%, более предпочтительно, по меньшей мере на 98%, наиболее предпочтительно, на 100% с SEQ ID NO: 4. Согласно одному варианту осуществления изобретения, выделенный полинуклеотид представляет собой молекулу ДНК, кодирующую гликозилированный полипептид, имеющий аминокислотную последовательность с идентичностью по меньшей мере на 90%, предпочтительно, по меньшей мере на 95%, более предпочтительно, по меньшей мере на 98%, наиболее предпочтительно, на 100% с SEQ ID NO: 5.
Экспрессионный вектор
В пятом аспекте изобретение также относится к экспрессионному вектору, содержащему полинуклеотид согласно четвертому аспекту изобретения.
Предпочтительно, чтобы экспрессионный вектор дополнительно содержал регуляторные элементы, такие как промотор, и сигналы остановки транскрипции и трансляциии. Полинуклеотид согласно четвертому аспекту и регуляторные элементы могут быть соединены вместе для полученого рекомбинантный экспрессионного вектора, который может включать один или несколько сайтов рестрикции, чтобы позволить введение или замещение полинуклеотида, кодирующего полипептид по таким сайтам. Полинуклеотид может быть вставлен в соответствующий экспрессионный вектор для экспрессии. При создании экспрессионного вектора кодирующая последовательность расположена в экспрессионном векторе таким образом, что кодирующая последовательность функционально связана с соответствующими регуляторными последовательностями для экспрессии.
Рекомбинантным экспрессионным вектором может быть любой вектор (например, плазмида или вирус), который подходит для методов рекомбинантных ДНК и может обеспечивать экспрессию полинуклеотида по четвертому аспекту изобретения. Выбор экспрессионного вектора обычно зависит от совместимости экспрессионного вектора с клеткой-хозяином, в которую должен быть введен экспрессионный вектор. Экспрессионные векторы могут представлять собой линейную или замкнутую кольцевую плазмиду.
Предпочтительно, чтобы экспрессионный вектор был адаптирован для экспрессии в клетках млекопитающих. Экспрессионный вектор может представлять собой автономно реплицирующийся вектор, то есть вектор, который существует в качестве внехромосомного объекта, репликация которого не зависит от хромосомной репликации, например, в виде плазмиды, внехромосомного элемента, минихромосомы или искусственной хромосомы. Для автономной репликации вектор может дополнительно содержать точку инициации репликации, позволяющую вектору реплицироваться автономно в соответствующей клетке-хозяине. Точкая инициации репликации может представлять собой любой плазмидный репликон, опосредующий автономную репликацию, которая функционирует в клетке. Термины «точка инициации репликации» или «плазмидный репликон» означают полинуклеотид, который позволяет плазмиде или вектору реплицироваться in vivo.
Вектор, предпочтительно, представляет собой вектор, который при введении в клетку-хозяина, встраивается в геном и реплицируется вместе с хромосомой (хромосомами), в которую он встроился. Для встраивания в геном клетки-хозяина в экспрессионном векторе может использоваться любой другой элемент экспрессионного вектора для встраивания в геном путем гомологичной или негомологичной рекомбинации. В альтернативном варианте вектор может содержать дополнительные полинуклеотиды для направленного встраивания с помощью гомологичной рекомбинации в геном клетки-хозяина в точное местоположение на хромосоме.
Векторы по настоящему изобретению, предпочтительно, содержат один или большее число (например, несколько) селективных маркеров, которые позволяют легко отбирать трансформированные, трансфицированные, трансдуцированные или аналогично модифицированные клетки. Селективный маркер представляет собой ген, продукт которого обеспечивает биоцидную или вирусную резистентность, устойчивость к тяжелым металлам, прототрофию по ауксотрофам и т.п.
Методики, используемые для лигирования описанных выше элементов для конструирования рекомбинантных экспрессионных векторов по настоящему изобретению, хорошо известны специалистам в данной области (см., например, Sambrook et al., 1989, выше).
Согласно одному варианту осуществления настоящего изобретения каркас вектора согласно пятому аспекту выбирают из pCDNA3, pCDNA3.1, pCDNA4, pCDNA5, pCDNA6, pCEP4, pCEP-puro, pCET1019, pCMV, pEF1, pEF4, pEF5, pEF6, pExchange, pEXPR, pIRES и pSCAS.
Вектор согласно пятому аспекту может быть временно или постоянно трансформирован в клетку-хозяина. Клетка-хозяин может представлять собой любую из перечисленных выше клеток. Предпочтительно, чтобы клеткой-хозяином являлась HEK 293F.
Медицинское применение и способ лечения
Как описано выше, гликозилированный полипептид и композиция, в частности, белковый комплекс по изобретению обладают преимуществом сниженного иммунного ответа у пациентов, в частности, у человека. Таким образом, гликозилированный полипептид и белковый комплекс особенно полезны в качестве действующих ингредиентов для лечения.
Согласно шестому аспекту изобретение относится к гликозилированным полипептидам по первому аспекту для применения в терапии. В качестве альтернативы, согласно шестому аспекту изобретение относится к композиции по третьему аспекту для применения в терапии. Предпочтительно, для лечения или профилактики нарушения свертываемости крови.
Таким образом, шестой аспект изобретения также относится к способу лечения или профилактики нарушения свертываемости крови у пациента, причем указанный способ включает введение указанному пациенту гликозилированного полипептида по первому аспекту или композиции, в частности, белкового комплекса по третьему аспекту.
Используемый в настоящем описании термин «нарушение свертываемости крови» относится к заболеванию или состоянию, которое нарушает нормальный гемостаз. Нарушением свертываемости, может являться, например, гемофилия А, гемофилия В, недостаточность фактора VIII, недостаточность фактора XI, болезнь фон Виллебранда, тромбастения Гланцмана, синдром Бернара Сулье, идиопатическая тромбоцитопеническая пурпура, интрацеребральное кровоизлияние, травма, травматическое повреждение головного мозга и т.п.
Используемый в настоящем описании термин «гемофилия» относится к группе нарушений свертываемости крови, связанных с увеличением времени образования сгустка крови по сравнению с временем образования сгустка крови у здоровых людей без гемофилии. «Гемофилия» относится как к гемофилии А, так и к расстройству, приводящему к продукции дефектного фактора VIII, и к гемофилии B, которая является заболеванием, приводящим к продукции дефектного фактора IX.
Нарушением свертываемости крови, предпочтительно, является гемофилия. Лечение может представлять собой, например, лечение гемофилии у ранее не подвергавшихся лечению пациентов (PUP) или индукцию иммунотолерантности (ITI).
В соответствии с альтернативным вариантом осуществления третьего аспекта изобретение относится к белковому комплексу по второму аспекту для применения в лечении или профилактике нарушений свертываемости крови.
Лечение, предпочтительно, включает введение пациенту эффективного количества гликозилированного полипептида или композиции, в частности, белкового комплекса.
Гликозилированный полипептид или композиция, в частности, белковый комплекс, описанные в настоящем документе, могут быть введены отдельно или в форме фармацевтических композиций. Фармацевтические композиции по изобретению могут содержать эффективное количество конъюгатов, в комбинации по меньшей мере с одним фармацевтически приемлемым носителем. Фармацевтические композиции в соответствии с вариантами осуществления могут быть получены и введены субъекту любыми способами, хорошо известными в области фармацевтики. См., например, Goodman & Gilman's The Pharmacological Basis of Therapeutics, Hardman et al., eds., McGraw-Hill Professional (10th ed., 2001); Remington: The Science and Practice of Pharmacy, Gennaro, ed., Lippincott Williams & Wilkins (20th ed., 2003); и Pharmaceutical Dosage Forms and Drug Delivery Systems, Ansel et al. (eds), Lippincott Williams & Wilkins (7th ed., 1999). Кроме того, фармацевтические композиции по настоящему изобретению также могут быть составлены таким образом, чтобы включать другие терапевтически полезные лекарственные средства или биологические агенты. Фармацевтическая композиция обычно содержит терапевтически эффективное количество гликозилированного полипептида или белкового комплекса в сочетании с фармацевтически приемлемым носителем. Фармацевтически приемлемый носитель представляет собой любой носитель, известный или известный в данной области. Примеры фармацевтически приемлемых носителей включают стерильную апирогенную воду и стерильный апирогенный солевой раствор. Другие формы фармацевтически приемлемых носителей, которые могут быть использованы для настоящих вариантов осуществления, включают связующие вещества, способствующие распадаемости вещества, поверхностно-активные вещества, ускорители абсорбции, удерживающие влагу агенты, абсорбенты, смазывающие вещества, наполнители, увеличители объема, придающие влажность агенты, консерванты, стабилизаторы, эмульгаторы, солюбилизирующие агенты, соли для регуляции осмотического давления и разбавляющие агенты, такие как буферы и вспомогательные вещества, обычно используемые в зависимости от формы применения состава. Их необязательно выбирают и используют в зависимости от лекарственной формы получаемого состава.
Для применения in vivo гликозилированный полипептид, белковый комплекс или фармацевтическая композиция могут вводиться пациенту любым обычным способом введения, например, перорально, парентерально или путем ингаляции. Парентеральное введение включает внутривенную инъекцию, подкожную инъекцию, интраперитонеальную инъекцию, внутримышечные инъекции и внутрибрюшинные инъекции, жидкие агенты, суспензии, эмульсии и вводимые капельно агенты. Для парентерального введения гликозилированный полипептид, белковый комплекс или фармацевтическая композиция должны являться инъекционным агентом, таким как жидкий агент или суспензия.
В других вариантах осуществления изобретения гликозилированный полипептид, белковый комплекс или фармацевтическую композицию вводят пациенту перорально. В этих вариантах осуществления форма лекарственного средства включает твердые составы, такие как таблетки, таблетки с покрытием, порошкообразные агенты, гранулы, капсулы и пилюли, жидкие композиции, такие как жидкие агенты (например, глазные капли, капли для носа и капли), суспензии, эмульсии и сиропы, ингаляционные агенты, такие как аэрозольные агенты, распылители и небулайзеры, и включенные в липосомы агенты. В некоторых других вариантах осуществления гликозилированный полипептид, белковый комплекс или фармацевтическую композицию вводят путем ингаляции в дыхательный канал пациента для направленного введения в трахею и/или легкое субъекта.
Согласно одному варианту осуществления шестого аспекта гликозилированный полипептид или композиция, в частности, белковый комплекс предназначены для применения, причем применение включает внутривенную или невнутривенную инъекцию. Невнутривенная инъекция, предпочтительно, представляет собой подкожную инъекцию.
Далее изобретение будет описано с помощью следующих неограничивающх примеров.
ПРИМЕРЫ
Пример 1 - Связывание фактора фон Виллебранда (vWF) с SIGLEC:
1.1. Методика эксперимента
Рекомбинантные SIG-2, SIG-5, SIG-7, SIG-F (мышиный эквивалент SIG-8 человека), SIG-9 и SIG-10 были получены в виде слитых с Fc белков от компании R&D Systems. Сначала планшет покрывали белком А (SERVA Feinbiochemica GmbH & Co) в концентрации 0,5 мкг/лунку при 4°С в течение ночи (O/N). После блокирования и промывки буфером для промывки (20 мМ HEPES, 125 мМ NaCl, 1 мМ EDTA, 1% BSA), Fc-слитые SIGLEC или контрольные антитела связывались с белком А путем 1-часовой инкубации при 37°С в концентрации 5 мкг/мл. В качестве положительного контроля иммобилизовали анти-vWF-pAb (Dako, #A0082), а в качестве отрицательного контроля иммобилизовали антитела против куриных IgY (Sigma Aldrich, # C2288) через Fc-часть антитела.
Плазматический vWF (pdVWF)) биотинилировали с использованием набора для биотинилирования EZ-Link™ Sulfo-NHS-Biotin (Thermo Fisher Scientific). Серийные разведения биотинилированного фактора vWF вносили в лунки в концентрациях от 0 до 0,8 мкг/мл. После пятикратной промывки буфером для промывки в лунки добавляли HRP-связанный стрептавидин (Thermo Fisher Scientific, #31001) и инкубировали в течение 1 часа при 37°С с последующей пятикратной промывкой.
Для визуализации связанного биотинилированного pdvWF лунки инкубировали с субстратом дигидрохлоридом о-фенилендиамина (SIGMAFAST™ OPD, #P9187, Sigma Aldrich). Затем измеряли поглощение при 492 нм.
1.2 Результаты
Как показано на фиг.1, поглощение при 492 нм увеличивается с начальной концентрацией vWF в экспериментах по связыванию с SIG-5, SIG-7, SIG-F и SIG-9. Значения являются несколько выше (SIG-F И SIG-9) или ниже (SIG-5 и SIG-7) значений для положительного контроля (анти-vWF). Напротив, поглощение в экспериментах по связыванию с SIG-2, SIG-10 и служащими отрицательным контролем антителами к куриным IgY составляло примерно 0 независимо от концентрации.
Соответственно, vWF связывается с SIG-5, SIG-7, SIG-F И SIG-9 в зависимости от концентрации. С другой стороны, vWF не связывается с SIG-2 и SIG-10.
Пример 2 Связывание фрагментов vWF с SIGLEC
2.1 Методика эксперимента
С- и N-концевые фрагменты vWF получали расщеплением протеазой V8 (Thermo Fisher Scitific, #201959), осуществляемым в течение 3 часов при 37°С, качании при 300 об/мин с использованием весового соотношения фермента и белка 1:100 в 50 мМ Трис-HCl, 150 мМ NaCl, рН 7,8, и очищали с помощью анионообменной хроматографии на колонке MonoQ 5/50 GL (GE Healthcare # 17-5166-01). В качестве рабочего буфера использовали 20 мМ Tris-HCl, pH 7,4, и буфер для элюирования - 20 мМ Tris-HCl, 500 мМ NaCl, pH 7,4. Фрагменты далее очищали и обессоливали с помощью эксклюзионной хроматографии на колонке Superose 6 10/300 GL (GE Healthcare № 17-5172-01) с использованием 100 мМ NaCl в качестве рабочего буфера.
Полученные фрагменты (С-конец и N-конец) схематически изображены на фиг. 2 с указанием доменов и сайтов гликозилирования.
Очищенные С-концевой фрагмент vWF и N-концевой фрагмент vWF биотинилировали с использованием набора для биотинилирования EZ-Link™ Sulfo-NHS-Biotin (Thermo Fisher Scientific), и связывание с SIGLEC измеряли, как описано в примере 1, причем концентрации C-концевого фрагмента vWF и N-концевого фрагмента vWF составляли 1 мкг/мл.
2.2 Результаты
Исходя из значений поглощения, показанных на фиг. 3, N-концевой фрагмент vWF, который содержит большинство сайтов О-гликозилирования и 2 О-гликановых кластера (кластер 1 и кластер 2), связывается с SIG-5, SIG-7, SIG-F и SIG-9. Напротив, в случае C-концевого фрагмента vWF наблюдалось небольшое или совсем не наблюдалось поглощение. Соответственно, последний фрагмент не связывается с SIGLEC.
Пример 3 - Связывание N-концевой части vWF с SIGLEC:
3.1 Методика эксперимента
Одну аликвоту N-концевого фрагмента vWF, полученного в примере 2, подвергали ферментативному десиалилированию с использованием Сиалидазы A. Инкубацию проводили при 37°С в течение 3 часов в 50 мМ фосфате натрия, рН 6,0, с использованием 2 мкл фермента на 100 мкг фрагмента VWF (Sialidase A™, #GK80040, Prozyme).
Вторую аликвоту N-концевого фрагмента vWF де-N-гликозилировали. Инкубацию проводили в течение ночи при 37°С в 50 мМ фосфате натрия, рН 7,5, с использованием 1 мкл фермента на 20 мкг фрагмента VWF (PNGaseF, #P0704, New England Biolabs).
Образцы десиалилированного, де-N-гликозилированного и необработанного N-концевого фрагмента vWF тестировали на связывание с SIG-5, SIG-7, SIG-8, и SIG-9. Эксперимент по связыванию проводили, как описано в примере 1, с концентрацией N-концевых фрагментов vWF, составляющей 8 мкг/мл.
3.2 Результаты:
Как показано на фиг.4, значения поглощения, определяемые для де-N-гликозилированного N-концевого фрагмента vWF и необработанного N-концевого фрагмента vWF, отличаются лишь незначительно. Таким образом, де-N-гликозилирование не влияет на связывание, демонстрируя, что связывание опосредовано О-гликанами.
Десиалилирование О-гликанов сильно снижает или отменяет связывание N-концевого фрагмента vWF с SIGLEC, как показано на фиг. 4. Связывание N-концевого фрагмента vWF опосредовано сиаловой кислотой, присоединенной к O-гликановым цепям.
Пример 4 - Связывание с SIGLEC пептидов, содержащих О-гликановые кластеры 1 и 2
4.1 Методика эксперимента
vWF содержит два кластера полностью занятых сайтов O-гликозилирования (см. Solecka et al., 2016), схематически изображенных на фиг.2. Оба кластера различаются по относительному количеству коровых структур 2-го типа. Только 4,9% гликопептидных молекул содержат коровые структуры 2-го типа в кластере 1. Соответственно, процентное содержание сиалилированных коровых О-гликанов 2-го типа от общего количества О-гликанов в кластере 1 составляет 1,25%.
Напротив, 34,86% гликопептидных молекул содержат коровые структуры 2-го типа в кластере 2 (см. Solecka et al., 2016). В соответствии с этим, процентное содержание сиалилированных коровых О-гликанов 2-го типа от общего количества О-гликанов в кластере 2 составляет 10,78%.
Для измерения независимого связывания двух кластеров N-концевой фрагмент vWF обрабатывали трипсином, получая следующие фрагменты: фрагмент кластера 1, охватывающий аминокислоты 449-511, и фрагмент кластера 2, охватывающий аминокислоты 674-728/729 из последовательности VWF (SEQ ID NO: 1). Фрагменты очищали с помощью обращеннофазовой ВЭЖХ. Кратко, pdVWF восстанавливали, а свободные цистеины блокировали малеимид-PEG2-биотином в соответствии с инструкцией производителя (EZ-Link™ Maleimide-PEG2-Biotin, #21901BID, Thermo Fisher Scientific). После расщепления трипсином при 37°С в течение ночи высокомолекулярные пептиды концентрировали с использованием центрифужного фильтра 10 кДа (Millipore). После этого пептиды разделяли на колонке Jupiter 5 микрон 300Å C18 (Phenomenex). Подвижная фаза: А - 0,1% трифторуксусной кислоты (ТФУ) в Н2О; B - 0,085% ТФУ в ацетонитриле, и скорость потока составляла 0,3 мл/мин. Элюируемые пептиды/гликопептиды детектировали по поглощению в ультрафиолетовой обалсти при длине волны 215 нм. Представляющие интерес фракции собирали, лиофилизировали и затем ресуспендировали в 10 мкл H2O. Поскольку оба кластера содержат цистеины, в оба был добавлен биотин.
Эксперимент по связыванию проводили, как описано в Примере 1, с использованием SIGLEC SIG-5, SIG-7, SIG-F, SIG-9 и SIG-10, и концентрации фрагментов кластера 1 и кластера 2 составляли 4 мкг/мл.
Кроме того, образцы фрагментов кластера 1 и кластера 2 подвергали десиалилированию и после этого исследовали в эксперименте по связыванию, как описано в примере 1, с концентрацией фрагментов десиалилированного кластера 1 и 2, составляющей 2 мкг/мл.
4.2 Результаты:
Исходя из значений поглощения, приведенных на фиг. 5, фрагмент кластера 2 связывался с SIG-5, SIG-7, SIG-F, SIG-9,. Отсуствие поглощения или небольшое поглощение детектировалось с использованием SIG-10, подтверждая результаты из других примеров.
Следовательно, высокий процент коровых структур 2-го типа в О-гликановых кластерах является необходимым для связывания с SIG-5, SIG-7, SIG-F, SIG-9.
Пример 5 - Зависимость связывания кластера 2 с SIGLEC от сиаловой кислоты
5.1 Методика эксперимента
Образец кластера 2, полученный, как описано в примере 4, десиалилировали. Десиалилированный фрагмент кластера 2 и необработанные фрагменты кластера 2 затем тестировали в эксперименте по связыванию, как описано в примере 1, с использованием SIG-5, SIG-7, SIG-F и SIG-9 и концентрации десиалилированных фрагментов кластера 1 и 2, составляющей 2 мкг/мл.
5.2 Результаты
Значения поглощения, детектируемые для необработанного фрагмента кластера 2, подтвердили результаты, полученные в примере 4 (см. фиг.6). Десиалилированный фрагмент кластера 2 демонстрирует отсутствие связывания или только небольшое связывание. Таким образом, сиалилирование коровых структур гликанов 2-го типа является необходимым для связывания с SIG-5, SIG-7, SIG-F, SIG-9.
Пример 6 - Рекомбинантная экспрессия фрагментов VWF с O-связанными гликановыми повторами, содержащими сайт связывания FVIII:
Два рекомбинантных фрагмента vWF были проэкспрессированы в линии клеток НЕК 293 F. Первый фрагмент, Seq11, включает аминокислоты 1-505 из SEQ ID NO: 1 и содержит кластер 1 с сайтами O-гликозилирования в позициях 485, 492, 493, 500.
Второй фрагмент, Seq 12, включает аминокислоты 1-505 из SEQ ID NO: 1 и 2 дополнительных повтора аминокислот 475-505 (AA1-505+2×475-505) и, таким образом, две дополнительные копии О-гликановых кластерных повтора из кластера 1.
Seq 11 и Seq 12 были транзиентно проэкспрессированы в клеточной линии HEK293 с C-концевым Strep-Tag и очищены аффинной хроматографией на StrepTactin сефарозе (IBA GmbH). Для этого, гены, кодирующие Seq 11 и 12, были синтезированы в GeneArt (Thermo Fisher Scientific) и клонированы в экспрессионный вектор pDSG (IBA GmbH), содержащий Twin-Strep-tag. Клетки E.coli TOP10 (IBA gmbH) были протрансформированы конструкциями, и индивидуальные клоны были отобраны после инкубации в течение ночи при 37°С на ампициллин-содержащих чашках с LB-агаром. Выделение плазмидной ДНК проводили с использованием набора QIAamp DNA-Mini или Maxi (Qiagen) в соответствии с рекомендациями производителя. Правильная ориентация и целостность клонированных конструкций была проверена путем секвенирования. Для эукариотической экспрессии обоих фрагментов vWF клетки MEXi-293 (IBA GmbH), выращенные в трансфекционной среде MEXi (IBA GmbH), трансфицировали 1,5 мг/л конструкций с использованием 4,5 мг/мл 25 кДа линейного полиэтиленимина. После 2-4-часовой инкубации при 37°С, 5% СО2 и качании 100-150 об/мин культуру разбавляли 1:2 трансфекционной средой MEXi, и культивирование продолжали до достижения клетками 75%-й конфлюентности. Затем супернатант отделяли от клеток центрифугированием при 4°С и 300×g. Чтобы минимизировать ингибирующее действие биотина в клеточной культуральной среде и для регуляции рН, в супернатант добавляли 0,1 объема буфера (1М Трис-HCl, 1,5 мМ NaCl, 10 мМ ЭДТА, рН 8,0) и 0,09 об.% раствор BioLock (IBA GmbH) и инкубировали в течение 20 мин при 4°С. После центрифугирования супернатант наносили на колонку Strep-Tactin XT (IBA GmbH), промывали пять раз промывочным буфером (100 мМ Трис-HCl, 150 мМ NaCl, 1 мМ ЭДТА, pH 8,0) и связавшиеся белки с Strep-tag элюировали элюирующим буфером (100 мМ Трис-HCl, 150 мМ NaCl, 1 мМ ЭДТА, 10 мМ дестиобиотин, pH 8,0).
Оба фрагмента, Seq11 и Seq12, схематически изображены на фиг.7
Пример 7 - Анализ O-гликозилирования фрагментов vWF, Seq11 и Seq12
7.1 Методика эксперимента
О-гликозилирование фрагментов Seq11 и Seq12, полученных в соответствии с примером 6, анализировали с помощью масс-спектрометрии.
Для этого, Seq11 и Seq12 сначала восстанавливали и алкилировали инкубированием в течение ночи с 50 мМ дитиотреитолом при 60°С и затем в течение 20 минут с 100 мМ иодацетамидом. После расщепления трипсином и химотрипсином полученные пептиды ресуспендировали в буфере для расщепления Сиалидазой А и десиалилировали в течение ночи с использованием условий, описанных в примере 3. O-гликопептиды были специально обогащены аффинной хроматографией на Якалине (лектине из Artocarpus integrifolia) с использованием иммобилизованного на агарозе лектина (Vector Laboratories). Якалин-агарозой набивали самотечную колонку, и хроматографию проводили в соответствии с инструкциями производителя. Элюированные O-гликопептиды очищали для анализа MALDI-MS с использованием наконечников от пипетки C4 Ziptip (Millipore) и анализировали в режиме линейной положительной ионизации с использованием 25 мг/мл матрицы Super-DHB, растворенной в 50%-м ацетонитриле/0,1%-й трифторуксусной кислоте.
Аликвоту обогащенных гликопептидов дополнительно обрабатывали О-гликозидазой (эндо-α-N-ацетилгалактозаминидазой, #P0733, New England Biolabs). Кратко, пептиды инкубировали с ферментом в течение 2 часов при 37°С с использованием 1 мкМ фермента на 10 мкМ гликопротеин. Так как О-гликозидаза специфична к коровым О-гликанам 1-го типа (Galβ1→3GalNAcα1→Ser/Thr дисахарид), то она оставляет коровые О-гликаны 2-го типа и/или удлиненные коровые О-гликаны 1-го типа, присоединенными к пептидной цепи.
7.2 Результаты
Результаты приведены на фиг.8 и 9.
О-гликопептиды идентифицировали с помощью MALDI в PSD-режиме (распад после источника). Пептидная последовательность идентифицированного фрагмента Seq11 представляет собой KVTLNPSDPEHCQICHCDVVNLTCEACQEPGGLVVPPTDAPVSPTTLYVEDISEPPLHGSAW (SEQ ID NO: 6). Последние четыре аминокислоты (подчеркнуты) соответствуют C-концевому Strep-Tag. Этот пептид содержит четыре сайта О-гликозилирования.
На верхнем спектре на фиг. 8 показан полностью O-гликозилированный гликопептид, на нижнем спектре на фиг. 8 показан тот же гликопептид после расщепления O-гликозидазой. Массовый сдвиг на 1460,3 Да после расщепления О-гликозидазой (отмеченный стрелкой), соответствует четырем коровым О-гликанам 1-го типа, каждый из которых имеет массу 365 Да. Сдвиги по 365 Да, наблюдаемые в верхнем спектре, соответствуют разным гликоформам одного и того же пептида, поскольку каждое дополнительное добавление массы по 365 Да соответствует дисахариду Galβ1→4GlcNAc, образующему структуру корового О-гликана 2-го типа. После обработки O-гликозидазой наблюдаются полностью дегликозилированная форма пептида (8358,6 Да) и гликоформы, содержащие коровые структуры 2-го типа (Galβ1→4GlcNAcβ1→6(Galβ1→3)GalNAc, 730 Да) и/или удлиненные коровые структуры 1-го типа (Galβ1→4GlcNAcβ1→3Galββ1→3GalNAc, 730 Да).
Идентифицированная пептидная последовательность фрагмента Seq12 представляет собой KVTLNPSDPEHCQICHCDVVNLTCEACQEPGGLVVPPTDAPVSPTTLYVEDISEPPLHQEPGGLVVPPTDAPVSPTTLYVEDISEPPLHQEPGGLVVPPTDAPVSPTTLYVEDISEPPLHGSAW (SEQ ID NO: 7). Последние четыре аминокислоты (подчеркнуты) соответствуют C-концевому Strep-Tag.
На верхнем спектре на фиг. 9 показан полностью O-гликозилированный гликопептид, на нижнем спектре на фиг. 9 показан тот же гликопептид после расщепления O-гликозидазой. Массовый сдвиг на 4380,3 Да после расщепления О-гликозидазой, соответствует двенадцати коровым О-гликанам 1-го типа, каждый из которых имеет массу 365 Да, что подтверждает, что все двенадцать сайтов О-гликозилирования в Seq12 заняты О-гликанами. Аналогично наблюдаемому для Seq11, расстояния в 365 Да и 730 Да, наблюдаемые в спектрах, соответствуют дисахариду Galβ1→4GlcNAc или коровой структуре 2-го типа и/или удлиненной коровой структуре 1-го типа, соответственно.
Количественное определение коровых О-гликанов 1-го и 2-го типа основано на относительном количестве гликопептидов с присоединенными к ним О-гликанами. Количественную оценку различных гликоформ данного пептида осуществляют путем оценки интенсивности сигнала в MALDI-спектре.
Для гликопептида Seq11 количественную оценку проводят на основе интенсивности сигнала MALDI от различных гликоформ одного пептида. Общая интенсивность пиков всех гликоформ этого пептида (8358 Да, 8724 Да, 9089 Да, 9454 Да, 9819 Да, 10181 Да, 10543 Да, 10910 Да, 11277 Да) равна 41275 a.u., что составляет 100%.
Гликоформа, содержащая только коровые гликаны 1-го типа с массой 8358 Да, имеет интенсивность 8410 a.u., что соответствует 20% от общей интенсивности. Таким образом, все другие гликоформы (80%) содержат по меньшей мере один присоединенный коровый гликан 2-го типа и/или удлиненный коровый гликан 1-го типа. В этом анализе коровый гликан 2-го типа и удлиненный коровый гликан 1-го типа неразличимы. Соответственно, процентное содержание коровых О-гликанов 2-го типа и/или удлиненных коровых О-гликанов 1-го типа от общего количества О-гликанов составляет по меньшей мере 20%.
Соответственно, четыре сайта О-гликозилирования в Seq11 и все двенадцать сайтов О-гликозилирования в Seq12, рекомбинантно продуцируемых в клеточной линии HEK, заняты коровыми О-гликанами 1-го типа и также в высокой степени коровыми О-гликанами 2-го типа и/или удлиненными коровыми О-гликанами 1-го типа. Принимая во внимание большое количество коровых O-гликанов 2-го типа, обе последовательности могут быть хорошими лигандами для связывания SIGLEC. Добавление двух дополнительных повторов последовательностей, содержащих четыре сайта О-гликозилирования, позволило успешно получить белок с двенадцатью кластерными и полностью занятыми О-гликанами сайтами О-гликозилирования.
Пример 8. Анализ связывания SIGLEC последовательностями Seq11 и Seq12
8.1 Методика эксперимента
Несущие Strep-Tag рекомбинантные белки, Seq11 и Seq12, тестировали в ELISA на связывания SIGLEC, как описано в примере 1, за исключением стратегии детекции; для выявления белков, меченных Strep-Tag, вместо Стрептавидина-HRP использовали конъюгат Strep-Tactin-HRP (#2-1502-001, IBA GmbH). Концентрация нанесенного Strep-Tactin-HRP составляла 0,25 мкг/мл. Оба белка, Seq11 и Seq12, тестировали в равной молярной концентрации - 42 нМ.
8.2 Результаты
Как показано на фиг.10, оба полипептида Seq11 и Seq12 связывались с SIG-5, SIG-7, SIG-F и SIG-9. Измеренное поглощение для обоих полипептидов Seq11 и Seq12 со всеми четырьмя SIGLEC было таким же, как и связывание Seq11 и Seq12 с анти-vWF.
Напротив, поглощение для Seq11 и Seq12 в эксперименте с SIG-2 и SIG-10 находилось в одном диапазоне с экспериментом с отрицательным контролем (антителом против куриных IgY). Таким образом, ни Seq11, ни Seq12, не связываются с SIG-2 или с SIG-10 (см. фиг.10).
Пример 9. Зависимость связывания SIGLEC последовательностями Seq11 и Seq12 от сиаловой кислоты
9.1 Методика эксперимента
Повторяли экспериментальный протокол из примера 8 с добавлением расщепления Сиалидазой Strep-меченных полипептидов Seq11 и Seq12. Десиалилирование проводили, как описано в примере 3.
9.2 Результаты
Результаты экспериментов по связыванию с десиалилированными Sеq11 и Seq12 показаны на фиг.11. Связывание с SIG-7, SIG-F и SIG-9 полностью блокируется, т.е. поглощение находится на том же уровне, что и при связывании с SIG-2 и SIG-10. Таким образом, связывание обоих полипептидов Seq11 и Seq12 с SIG-5, SIG-7, SIG-F и SIG-9 зависит от сиаловой кислоты.
Пример 10 - Сравнение связывания Seq11 and Seq12 с SIGLEC
10.1 Методика эксперимента
Для того чтобы измерить и сравнить кажущуюся аффинность связывания Seq11 и Seq12, проводили твердофазный ИФА (ELISA) на связывание с SIGLEC с анализом Скэтчарда кривых связывания. ELISA проводили, как описано в примерах 8 и 9.
10.2 Результаты
Кривые связывания для Seq11 и Seq12 и соответствующие графики Скэтчарда показаны на фиг. 12 и 13. Кажущаяся аффинность связывания (KD), выведенная из графиков Скэтчарда, приведена на фиг. 14. Увеличение числа О-гликановых повторов в Seq12 оказывает значительное влияние на аффинность связывания SIGLEC. Аффинность к SIGLEC 5 может быть увеличена с 0,494 мкМ для Seq 11 до 0,14 мкМ для Seq12. Аффинность к SILEC 7 может быть увеличена с 0,371 мкМ для Seq11 до 0,005 мкМ для Seq12. Аффинность к SIGLEC 8 может быть увеличена с 1,027 мкМ для Seq11 до 0,015 мкМ для Seq12. Наконец, аффинность к SIGLEC 9 может быть увеличена с 0,591 мкМ для Seq11 до 0,041 мкМ для Seq12. Утверждение в тексте: «из этого эксперимента ELISA будут рассчитаны константы диссоциации для всех взаимодействий Seq11 и 12 с SIGLEC».
Пример 11 - Определение аффинности связывания Seq11 и Seq12 с FVIII
11.1 Методика эксперимента
Связывание FVIII с обеими последовательностями оценивали с помощью поверхностного плазменного резонанса (SPR). Анализ проводили с использованием прибора Biacore 3000 (GE Healthcare). Полипептиды Seq11 и Seq12 иммобилизовали на чипе CM5 с использованием набора для присоединения по аминогруппам (GE Healthcare). В качестве положительного контроля иммобилизовали полноразмерный плазматический VWF (Wislat, Ocpharma). После этого, над сенсорной поверхностью чипа пропускали серии концентарций FVIII (Nuwiq, Octapharma) (0,2 нМ, 0,6 нМ, 1,7 нМ, 5,0 нМ, 15 нМ, 45 нМ). Рабочий буфер представлял собой 150 мМ HEPES, 150 мМ NaCl, 5 мМ CaCl2, 0,05%.
11.2 Результаты
Результаты измерений методом SPR показали, что аффинность связывания FVIII (KD) у обеих последовательностей составляет 1,4 нМ, таким образом, дополнительные О-гликановые повторы, не оказывают влияния на аффинность связывания с FVIII.
Пример 12 - N-концевой, но не С-концевой фрагмент VWF, уменьшает внеклеточный уровень IL-12p70 и IFN-γ
12.1 Теоретическое обоснование
SIGLEC экспрессируются на различных клетках иммунной системы, включая моноциты и дендритные клетки, и играют роль в адгезии клеток, эндоцитозе и модуляции сигнальных путей адаптивного и врожденного иммунитета (Macauley et al. 2014). Большинство SIGLEC содержат иммунорецепторный тирозиновый ингибирующий мотив (ITIM) или ITIM-подобный мотив в своем цитоплазматическом домене, который, как было показано, действует, ослабляя воспалительный ответ путем ингибирования пролиферации и активации клеток (Vitale et al. 1999; Ikehara et al. 2004), индуцируя апоптоз (Nutku et al. 2003) и подавляя продукцию цитокинов (Erdmann et al. 2009; Chen et al. 2013).
Для того чтобы определить, меняются ли уровни воспалительных цитокинов, продуцируемых moDC, в присутствии связывающегося с SIGLEC фрагмента vWF, проводили одновременный анализ количества IL-12p70 и IFN-γв супернатанте стимулированных незрелых полученных из моноцитов дендритных клеток (moDC) с помощью проточной цитометрии.
12.2 Схема эксперимента
Моноциты от здоровых доноров обогащали в градиенте фикола, а затем выделяли CD14+ моноциты с помощью магнитного сортинга. Для получения moDC, CD14+ моноциты культивировали в течение 5-6 дней в среде RPMI, дополненной 10% фетальной телячьей сыворотки, 1000 Ед/мл интерлейкина-4 и 1000 Ед/мл гранулоцитарно-макрофагального колониестимулирующего фактора. Профиль цитокинов, секретируемых moDC, анализировали через 24 часа после стимуляции соответствующими фрагментами vWF с помощью цитометрического анализа на шариках CBA Flex (BD), определяющего IL-12p70 и IFN-γ, в соответствии с рекомендациями производителя. Клетки, обработанные тем же объемом 100 мМ NaCl, служили в качестве контроля. Образцы анализировали на проточном цитометре FACSVerse. Окончательный анализ и расчет концентрации цитокинов проводили с использованием программного обеспечения FCAP Array (BD).
12.3 Результаты
На фиг.16 показана концентрация цитокинов после инкубации клеток с двумя фрагментами фактора vWF с стимуляцией LPS и без стимуляции LPS. В соответствии с этими результатами N-концевая часть, но не С-концевая часть vWF, снижает продукцию провоспалительных цитокинов, синтезируемых в ответ на стимуляцию LPS. Без стимуляции LPS не было обнаружено влияния фрагментов vWF на секрецию провоспалительных цитокинов.
Пример 13 - Анализ фосфорилирования SIGLEC и их адапторных молекул
13.1 Теоретическое обоснование
При связывании содержащего сиаловую кислоту лиганда ITIM и ITIM-подобные мотивы в SIGLEC фосфорилируются тирозинкиназами SRC-семейства, что приводит к рекрутированию содержащих SH2-домен (2-й гомологичный SRC домен) тирозинфосфатаз SHP-1 и SHP-2. После активации эти фосфатазы могут дефосфорилировать клеточные субстраты, таким образом, контролируя активацию различных сигнальных путей (Crocker Et al. 2007). В то время как SHP-1 считалась участвующей в ингибирующих сигнальных путях, SHP-2 усиливает передачу сигнала в большинстве сигнальных путей, но также по литературным данным участвует в отрицательном регулировании процессов внутриклеточной передачи сигналов (An et al., 2006; Avril et al., 2004; Boyd et al., 2009; Qu, 2000; Salmond and Alexander, 2006).
13.2 Методика эксперимента
Клетки moDC получали, как описано в примере 12. Для определения влияния N-концевого vWF-фрагмента на тирозиновое фосфорилирование SIGLEC и их адапторных молекул SHP-1 и SHP-2, 6×106 moDC инкубировали в течение 10 минут с 500 нМ N-концевым фрагментом vWF. Клетки, стимулированные тем же объемом 100 мМ NaCl, служили в качестве контроля. Анализ фосфорилирования иммунорецепторов проводили с помощью набора Proteome Profiler Human Phospho-Immunoreceptor Array Kit (R&D Systems) в соответствии с рекомендациями производителя с использованием 500 мкг клеточного лизата.
13.3 Результаты
Результаты, полученные в Phospho-Immunoreceptor Array, показаны на фиг.17. В соответствии с измеренной плотностью пикселей N-концевой фрагмент vWF специфично изменяет фосфорилирование SHP-1, SHP-2, SIG-5 и SIG-7 по сравнению с контролем. Фосфорилирование SIG-2 и SIG-10 не наблюдается, что прямо коррелирует с отсутствием связывания N-концевым фрагментом vWF этих SIGLEC.
Имеющим преимущество в виде идеи изобретения, представленной в приведенном выше описании и соответствующих чертежах, специалистам в области техники, к которой относится изобретение, изложенное в настоящем описании, будут очевидны многие модификации и другие варианты осуществления изобретения. Таким образом, следует понимать, что изобретение не ограничивается описанными конкретными вариантами осуществления, и подразумеватеся, что модификации и другие варианты осуществления включены в объем прилагаемой формулы изобретения. Хотя в настоящем описании используются конкретные термины, они используются только в общем и описательном смысле, а не в целях ограничения.
СПИСОК ЛИТЕРАТУРЫ
An,H., Zhao,W., Hou,J., Zhang,Y., Xie,Y., Zheng,Y., Xu,H., Qian,C., Zhou,J., Yu,Y., Liu,S., Feng,G., and Cao,X. (2006). SHP-2 phosphatase negatively regulates the TRIF adaptor protein-dependent type I interferon and proinflammatory cytokine production. Immunity. 25, 919-928.
Avril,T., Floyd,H., Lopez,F., Vivier,E., and Crocker,P.R. (2004). The membrane-proximal immunoreceptor tyrosine-based inhibitory motif is critical for the inhibitory signaling mediated by Siglecs-7 and -9, CD33-related Siglecs expressed on human monocytes and NK cells. J. Immunol. 173, 6841-6849.
Boyd,C.R., Orr,S.J., Spence,S., Burrows,J.F., Elliott,J., Carroll,H.P., Brennan,K., Ni,G.J., Coulter,W.A., Jones,C., Crocker,P.R., Johnston,J.A., and Jefferies,C.A. (2009). Siglec-E is up-regulated and phosphorylated following lipopolysaccharide stimulation in order to limit TLR-driven cytokine production. J. Immunol. 183, 7703-7709.
Chen, Weilin; Han, Chaofeng; Xie, Bin; Hu, Xiang; Yu, Qian; Shi, Liyun et al. (2013): Induction of Siglec-G by RNA viruses inhibits the innate immune response by promoting RIG-I degradation. In: Cell 152 (3), S. 467-478. DOI: 10.1016/j.cell.2013.01.011.
Crocker,P.R., Paulson,J.C., and Varki,A. (2007). Siglecs and their roles in the immune system. Nat. Rev. Immunol. 7, 255-266.
Ewenstein BM, Collins P, Tarantino MD, Negrier C, Blanchette V, Shapiro AD, Baker D, Spotts G, Sensel M, Yi SE, Gomperts ED. Hemophilia therapy innovation development of an advanced category recombinant factor VIII by a plasma/albumin-free method Proceedings of a Special Symposium at the XIXth Congress of the International Society on Thrombosis and Haemostasis; 2004, vol. 41, pg. 1-16.
Erdmann, Hanna; Steeg, Christiane; Koch-Nolte, Friedrich; Fleischer, Bernhard; Jacobs, Thomas (2009): Sialylated ligands on pathogenic Trypanosoma cruzi interact with Siglec-E (sialic acid-binding Ig-like lectin-E). In: Cellular microbiology 11 (11), S. 1600-1611. DOI: 10.1111/j.1462-5822.2009.01350.x.
Franc V, Řehulka P, Raus M, Stulík J, Novak J, Renfrow MB, Šebela M. Elucidating heterogeneity of IgA1 hinge-region O-glycosylation by use of MALDI-TOF/TOF mass spectrometry: role of cysteine alkylation during sample processing. Journal of Proteomics, 2013, Oct 30; vol. 92, pg. 299-312
Guzman-Aranguez A, Argüeso P. Structure and Biological Roles of Mucin-type O-glycans at the Ocular Surface. The ocular surface. 2010; vol. 8 (1) pg- 8-17.
Ikehara, Yuzuru; Ikehara, Sanae Kabata; Paulson, James C. (2004): Negative regulation of T cell receptor signaling by Siglec-7 (p70/AIRM) and Siglec-9. In: The Journal of biological chemistry 279 (41), S. 43117-43125. DOI: 10.1074/jbc.M403538200.
Lai JD Georgescu MT, Hough C, Lillicrap D. To clear or to fear: An innate perspective on factor VIII immunity, Cellular Immunology, 2016 Mar; vol. 301, pg. 82-89.
Nutku, Esra; Aizawa, Hideyuki; Hudson, Sherry A.; Bochner, Bruce S. (2003): Ligation of Siglec-8: a selective mechanism for induction of human eosinophil apoptosis. In: Blood 101 (12), S. 5014-5020. DOI: 10.1182/blood-2002-10-3058.
Pegon JN, Kurdi M, Casari C, Odouard S, Denis CV, Christophe OD, Lenting PJ. Factor VIII and von Willebrand factor are ligands for the carbohydrate-receptor Siglec-5, Haematologica. 2012 Dec; 97(12):1855-63.
Paulson JC, Macauley MS, and Kawasaki N. Siglecs as sensors of self in innate and adaptive immune responses. Ann N Y Acad Sci. 2012 April; 1253(1): 37-48.
Qu,C.K. (2000). The SHP-2 tyrosine phosphatase: signaling mechanisms and biological functions. Cell Res. 10, 279-288.
Salmond,R.J., and Alexander,D.R. (2006). SHP2 forecast for the immune system: fog gradually clearing. Trends Immunol. 27, 154-160.
Solecka BA, Weise C, Laffan MA, Kannicht C, Site-specific analysis of von Willebrand factor O-glycosylation, J Thromb Haemost. 2016 Jan 19.
Vitale, C.; Romagnani, C.; Falco, M.; Ponte, M.; Vitale, M.; Moretta, A. et al. (1999): Engagement of p75/AIRM1 or CD33 inhibits the proliferation of normal or leukemic myeloid cells. In: Proceedings of the National Academy of Sciences of the United States of America 96 (26), S. 15091-15096.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Octpharma AG
<120> ПОЛИПЕПТИДЫ, МОДУЛИРУЮЩИЕ SIGLEC-ЗАВИСИМЫЕ ИММУННЫЕ ОТВЕТЫ
<130> 4830/P/1095-WO
<160> 21
<170> PatentIn version 3.5
<210> 1
<211> 2050
<212> БЕЛОК
<213> Homo sapiens
<400> 1
Ser Leu Ser Cys Arg Pro Pro Met Val Lys Leu Val Cys Pro Ala Asp
1 5 10 15
Asn Leu Arg Ala Glu Gly Leu Glu Cys Thr Lys Thr Cys Gln Asn Tyr
20 25 30
Asp Leu Glu Cys Met Ser Met Gly Cys Val Ser Gly Cys Leu Cys Pro
35 40 45
Pro Gly Met Val Arg His Glu Asn Arg Cys Val Ala Leu Glu Arg Cys
50 55 60
Pro Cys Phe His Gln Gly Lys Glu Tyr Ala Pro Gly Glu Thr Val Lys
65 70 75 80
Ile Gly Cys Asn Thr Cys Val Cys Gln Asp Arg Lys Trp Asn Cys Thr
85 90 95
Asp His Val Cys Asp Ala Thr Cys Ser Thr Ile Gly Met Ala His Tyr
100 105 110
Leu Thr Phe Asp Gly Leu Lys Tyr Leu Phe Pro Gly Glu Cys Gln Tyr
115 120 125
Val Leu Val Gln Asp Tyr Cys Gly Ser Asn Pro Gly Thr Phe Arg Ile
130 135 140
Leu Val Gly Asn Lys Gly Cys Ser His Pro Ser Val Lys Cys Lys Lys
145 150 155 160
Arg Val Thr Ile Leu Val Glu Gly Gly Glu Ile Glu Leu Phe Asp Gly
165 170 175
Glu Val Asn Val Lys Arg Pro Met Lys Asp Glu Thr His Phe Glu Val
180 185 190
Val Glu Ser Gly Arg Tyr Ile Ile Leu Leu Leu Gly Lys Ala Leu Ser
195 200 205
Val Val Trp Asp Arg His Leu Ser Ile Ser Val Val Leu Lys Gln Thr
210 215 220
Tyr Gln Glu Lys Val Cys Gly Leu Cys Gly Asn Phe Asp Gly Ile Gln
225 230 235 240
Asn Asn Asp Leu Thr Ser Ser Asn Leu Gln Val Glu Glu Asp Pro Val
245 250 255
Asp Phe Gly Asn Ser Trp Lys Val Ser Ser Gln Cys Ala Asp Thr Arg
260 265 270
Lys Val Pro Leu Asp Ser Ser Pro Ala Thr Cys His Asn Asn Ile Met
275 280 285
Lys Gln Thr Met Val Asp Ser Ser Cys Arg Ile Leu Thr Ser Asp Val
290 295 300
Phe Gln Asp Cys Asn Lys Leu Val Asp Pro Glu Pro Tyr Leu Asp Val
305 310 315 320
Cys Ile Tyr Asp Thr Cys Ser Cys Glu Ser Ile Gly Asp Cys Ala Cys
325 330 335
Phe Cys Asp Thr Ile Ala Ala Tyr Ala His Val Cys Ala Gln His Gly
340 345 350
Lys Val Val Thr Trp Arg Thr Ala Thr Leu Cys Pro Gln Ser Cys Glu
355 360 365
Glu Arg Asn Leu Arg Glu Asn Gly Tyr Glu Cys Glu Trp Arg Tyr Asn
370 375 380
Ser Cys Ala Pro Ala Cys Gln Val Thr Cys Gln His Pro Glu Pro Leu
385 390 395 400
Ala Cys Pro Val Gln Cys Val Glu Gly Cys His Ala His Cys Pro Pro
405 410 415
Gly Lys Ile Leu Asp Glu Leu Leu Gln Thr Cys Val Asp Pro Glu Asp
420 425 430
Cys Pro Val Cys Glu Val Ala Gly Arg Arg Phe Ala Ser Gly Lys Lys
435 440 445
Val Thr Leu Asn Pro Ser Asp Pro Glu His Cys Gln Ile Cys His Cys
450 455 460
Asp Val Val Asn Leu Thr Cys Glu Ala Cys Gln Glu Pro Gly Gly Leu
465 470 475 480
Val Val Pro Pro Thr Asp Ala Pro Val Ser Pro Thr Thr Leu Tyr Val
485 490 495
Glu Asp Ile Ser Glu Pro Pro Leu His Asp Phe Tyr Cys Ser Arg Leu
500 505 510
Leu Asp Leu Val Phe Leu Leu Asp Gly Ser Ser Arg Leu Ser Glu Ala
515 520 525
Glu Phe Glu Val Leu Lys Ala Phe Val Val Asp Met Met Glu Arg Leu
530 535 540
Arg Ile Ser Gln Lys Trp Val Arg Val Ala Val Val Glu Tyr His Asp
545 550 555 560
Gly Ser His Ala Tyr Ile Gly Leu Lys Asp Arg Lys Arg Pro Ser Glu
565 570 575
Leu Arg Arg Ile Ala Ser Gln Val Lys Tyr Ala Gly Ser Gln Val Ala
580 585 590
Ser Thr Ser Glu Val Leu Lys Tyr Thr Leu Phe Gln Ile Phe Ser Lys
595 600 605
Ile Asp Arg Pro Glu Ala Ser Arg Ile Thr Leu Leu Leu Met Ala Ser
610 615 620
Gln Glu Pro Gln Arg Met Ser Arg Asn Phe Val Arg Tyr Val Gln Gly
625 630 635 640
Leu Lys Lys Lys Lys Val Ile Val Ile Pro Val Gly Ile Gly Pro His
645 650 655
Ala Asn Leu Lys Gln Ile Arg Leu Ile Glu Lys Gln Ala Pro Glu Asn
660 665 670
Lys Ala Phe Val Leu Ser Ser Val Asp Glu Leu Glu Gln Gln Arg Asp
675 680 685
Glu Ile Val Ser Tyr Leu Cys Asp Leu Ala Pro Glu Ala Pro Pro Pro
690 695 700
Thr Leu Pro Pro Asp Met Ala Gln Val Thr Val Gly Pro Gly Leu Leu
705 710 715 720
Gly Val Ser Thr Leu Gly Pro Lys Arg Asn Ser Met Val Leu Asp Val
725 730 735
Ala Phe Val Leu Glu Gly Ser Asp Lys Ile Gly Glu Ala Asp Phe Asn
740 745 750
Arg Ser Lys Glu Phe Met Glu Glu Val Ile Gln Arg Met Asp Val Gly
755 760 765
Gln Asp Ser Ile His Val Thr Val Leu Gln Tyr Ser Tyr Met Val Thr
770 775 780
Val Glu Tyr Pro Phe Ser Glu Ala Gln Ser Lys Gly Asp Ile Leu Gln
785 790 795 800
Arg Val Arg Glu Ile Arg Tyr Gln Gly Gly Asn Arg Thr Asn Thr Gly
805 810 815
Leu Ala Leu Arg Tyr Leu Ser Asp His Ser Phe Leu Val Ser Gln Gly
820 825 830
Asp Arg Glu Gln Ala Pro Asn Leu Val Tyr Met Val Thr Gly Asn Pro
835 840 845
Ala Ser Asp Glu Ile Lys Arg Leu Pro Gly Asp Ile Gln Val Val Pro
850 855 860
Ile Gly Val Gly Pro Asn Ala Asn Val Gln Glu Leu Glu Arg Ile Gly
865 870 875 880
Trp Pro Asn Ala Pro Ile Leu Ile Gln Asp Phe Glu Thr Leu Pro Arg
885 890 895
Glu Ala Pro Asp Leu Val Leu Gln Arg Cys Cys Ser Gly Glu Gly Leu
900 905 910
Gln Ile Pro Thr Leu Ser Pro Ala Pro Asp Cys Ser Gln Pro Leu Asp
915 920 925
Val Ile Leu Leu Leu Asp Gly Ser Ser Ser Phe Pro Ala Ser Tyr Phe
930 935 940
Asp Glu Met Lys Ser Phe Ala Lys Ala Phe Ile Ser Lys Ala Asn Ile
945 950 955 960
Gly Pro Arg Leu Thr Gln Val Ser Val Leu Gln Tyr Gly Ser Ile Thr
965 970 975
Thr Ile Asp Val Pro Trp Asn Val Val Pro Glu Lys Ala His Leu Leu
980 985 990
Ser Leu Val Asp Val Met Gln Arg Glu Gly Gly Pro Ser Gln Ile Gly
995 1000 1005
Asp Ala Leu Gly Phe Ala Val Arg Tyr Leu Thr Ser Glu Met His
1010 1015 1020
Gly Ala Arg Pro Gly Ala Ser Lys Ala Val Val Ile Leu Val Thr
1025 1030 1035
Asp Val Ser Val Asp Ser Val Asp Ala Ala Ala Asp Ala Ala Arg
1040 1045 1050
Ser Asn Arg Val Thr Val Phe Pro Ile Gly Ile Gly Asp Arg Tyr
1055 1060 1065
Asp Ala Ala Gln Leu Arg Ile Leu Ala Gly Pro Ala Gly Asp Ser
1070 1075 1080
Asn Val Val Lys Leu Gln Arg Ile Glu Asp Leu Pro Thr Met Val
1085 1090 1095
Thr Leu Gly Asn Ser Phe Leu His Lys Leu Cys Ser Gly Phe Val
1100 1105 1110
Arg Ile Cys Met Asp Glu Asp Gly Asn Glu Lys Arg Pro Gly Asp
1115 1120 1125
Val Trp Thr Leu Pro Asp Gln Cys His Thr Val Thr Cys Gln Pro
1130 1135 1140
Asp Gly Gln Thr Leu Leu Lys Ser His Arg Val Asn Cys Asp Arg
1145 1150 1155
Gly Leu Arg Pro Ser Cys Pro Asn Ser Gln Ser Pro Val Lys Val
1160 1165 1170
Glu Glu Thr Cys Gly Cys Arg Trp Thr Cys Pro Cys Val Cys Thr
1175 1180 1185
Gly Ser Ser Thr Arg His Ile Val Thr Phe Asp Gly Gln Asn Phe
1190 1195 1200
Lys Leu Thr Gly Ser Cys Ser Tyr Val Leu Phe Gln Asn Lys Glu
1205 1210 1215
Gln Asp Leu Glu Val Ile Leu His Asn Gly Ala Cys Ser Pro Gly
1220 1225 1230
Ala Arg Gln Gly Cys Met Lys Ser Ile Glu Val Lys His Ser Ala
1235 1240 1245
Leu Ser Val Glu Leu His Ser Asp Met Glu Val Thr Val Asn Gly
1250 1255 1260
Arg Leu Val Ser Val Pro Tyr Val Gly Gly Asn Met Glu Val Asn
1265 1270 1275
Val Tyr Gly Ala Ile Met His Glu Val Arg Phe Asn His Leu Gly
1280 1285 1290
His Ile Phe Thr Phe Thr Pro Gln Asn Asn Glu Phe Gln Leu Gln
1295 1300 1305
Leu Ser Pro Lys Thr Phe Ala Ser Lys Thr Tyr Gly Leu Cys Gly
1310 1315 1320
Ile Cys Asp Glu Asn Gly Ala Asn Asp Phe Met Leu Arg Asp Gly
1325 1330 1335
Thr Val Thr Thr Asp Trp Lys Thr Leu Val Gln Glu Trp Thr Val
1340 1345 1350
Gln Arg Pro Gly Gln Thr Cys Gln Pro Ile Leu Glu Glu Gln Cys
1355 1360 1365
Leu Val Pro Asp Ser Ser His Cys Gln Val Leu Leu Leu Pro Leu
1370 1375 1380
Phe Ala Glu Cys His Lys Val Leu Ala Pro Ala Thr Phe Tyr Ala
1385 1390 1395
Ile Cys Gln Gln Asp Ser Cys His Gln Glu Gln Val Cys Glu Val
1400 1405 1410
Ile Ala Ser Tyr Ala His Leu Cys Arg Thr Asn Gly Val Cys Val
1415 1420 1425
Asp Trp Arg Thr Pro Asp Phe Cys Ala Met Ser Cys Pro Pro Ser
1430 1435 1440
Leu Val Tyr Asn His Cys Glu His Gly Cys Pro Arg His Cys Asp
1445 1450 1455
Gly Asn Val Ser Ser Cys Gly Asp His Pro Ser Glu Gly Cys Phe
1460 1465 1470
Cys Pro Pro Asp Lys Val Met Leu Glu Gly Ser Cys Val Pro Glu
1475 1480 1485
Glu Ala Cys Thr Gln Cys Ile Gly Glu Asp Gly Val Gln His Gln
1490 1495 1500
Phe Leu Glu Ala Trp Val Pro Asp His Gln Pro Cys Gln Ile Cys
1505 1510 1515
Thr Cys Leu Ser Gly Arg Lys Val Asn Cys Thr Thr Gln Pro Cys
1520 1525 1530
Pro Thr Ala Lys Ala Pro Thr Cys Gly Leu Cys Glu Val Ala Arg
1535 1540 1545
Leu Arg Gln Asn Ala Asp Gln Cys Cys Pro Glu Tyr Glu Cys Val
1550 1555 1560
Cys Asp Pro Val Ser Cys Asp Leu Pro Pro Val Pro His Cys Glu
1565 1570 1575
Arg Gly Leu Gln Pro Thr Leu Thr Asn Pro Gly Glu Cys Arg Pro
1580 1585 1590
Asn Phe Thr Cys Ala Cys Arg Lys Glu Glu Cys Lys Arg Val Ser
1595 1600 1605
Pro Pro Ser Cys Pro Pro His Arg Leu Pro Thr Leu Arg Lys Thr
1610 1615 1620
Gln Cys Cys Asp Glu Tyr Glu Cys Ala Cys Asn Cys Val Asn Ser
1625 1630 1635
Thr Val Ser Cys Pro Leu Gly Tyr Leu Ala Ser Thr Ala Thr Asn
1640 1645 1650
Asp Cys Gly Cys Thr Thr Thr Thr Cys Leu Pro Asp Lys Val Cys
1655 1660 1665
Val His Arg Ser Thr Ile Tyr Pro Val Gly Gln Phe Trp Glu Glu
1670 1675 1680
Gly Cys Asp Val Cys Thr Cys Thr Asp Met Glu Asp Ala Val Met
1685 1690 1695
Gly Leu Arg Val Ala Gln Cys Ser Gln Lys Pro Cys Glu Asp Ser
1700 1705 1710
Cys Arg Ser Gly Phe Thr Tyr Val Leu His Glu Gly Glu Cys Cys
1715 1720 1725
Gly Arg Cys Leu Pro Ser Ala Cys Glu Val Val Thr Gly Ser Pro
1730 1735 1740
Arg Gly Asp Ser Gln Ser Ser Trp Lys Ser Val Gly Ser Gln Trp
1745 1750 1755
Ala Ser Pro Glu Asn Pro Cys Leu Ile Asn Glu Cys Val Arg Val
1760 1765 1770
Lys Glu Glu Val Phe Ile Gln Gln Arg Asn Val Ser Cys Pro Gln
1775 1780 1785
Leu Glu Val Pro Val Cys Pro Ser Gly Phe Gln Leu Ser Cys Lys
1790 1795 1800
Thr Ser Ala Cys Cys Pro Ser Cys Arg Cys Glu Arg Met Glu Ala
1805 1810 1815
Cys Met Leu Asn Gly Thr Val Ile Gly Pro Gly Lys Thr Val Met
1820 1825 1830
Ile Asp Val Cys Thr Thr Cys Arg Cys Met Val Gln Val Gly Val
1835 1840 1845
Ile Ser Gly Phe Lys Leu Glu Cys Arg Lys Thr Thr Cys Asn Pro
1850 1855 1860
Cys Pro Leu Gly Tyr Lys Glu Glu Asn Asn Thr Gly Glu Cys Cys
1865 1870 1875
Gly Arg Cys Leu Pro Thr Ala Cys Thr Ile Gln Leu Arg Gly Gly
1880 1885 1890
Gln Ile Met Thr Leu Lys Arg Asp Glu Thr Leu Gln Asp Gly Cys
1895 1900 1905
Asp Thr His Phe Cys Lys Val Asn Glu Arg Gly Glu Tyr Phe Trp
1910 1915 1920
Glu Lys Arg Val Thr Gly Cys Pro Pro Phe Asp Glu His Lys Cys
1925 1930 1935
Leu Ala Glu Gly Gly Lys Ile Met Lys Ile Pro Gly Thr Cys Cys
1940 1945 1950
Asp Thr Cys Glu Glu Pro Glu Cys Asn Asp Ile Thr Ala Arg Leu
1955 1960 1965
Gln Tyr Val Lys Val Gly Ser Cys Lys Ser Glu Val Glu Val Asp
1970 1975 1980
Ile His Tyr Cys Gln Gly Lys Cys Ala Ser Lys Ala Met Tyr Ser
1985 1990 1995
Ile Asp Ile Asn Asp Val Gln Asp Gln Cys Ser Cys Cys Ser Pro
2000 2005 2010
Thr Arg Thr Glu Pro Met Gln Val Ala Leu His Cys Thr Asn Gly
2015 2020 2025
Ser Val Val Tyr His Glu Val Leu Asn Ala Met Glu Cys Lys Cys
2030 2035 2040
Ser Pro Arg Lys Cys Ser Lys
2045 2050
<210> 2
<211> 567
<212> БЕЛОК
<213> Homo sapiens
<400> 2
Ser Leu Ser Cys Arg Pro Pro Met Val Lys Leu Val Cys Pro Ala Asp
1 5 10 15
Asn Leu Arg Ala Glu Gly Leu Glu Cys Thr Lys Thr Cys Gln Asn Tyr
20 25 30
Asp Leu Glu Cys Met Ser Met Gly Cys Val Ser Gly Cys Leu Cys Pro
35 40 45
Pro Gly Met Val Arg His Glu Asn Arg Cys Val Ala Leu Glu Arg Cys
50 55 60
Pro Cys Phe His Gln Gly Lys Glu Tyr Ala Pro Gly Glu Thr Val Lys
65 70 75 80
Ile Gly Cys Asn Thr Cys Val Cys Gln Asp Arg Lys Trp Asn Cys Thr
85 90 95
Asp His Val Cys Asp Ala Thr Cys Ser Thr Ile Gly Met Ala His Tyr
100 105 110
Leu Thr Phe Asp Gly Leu Lys Tyr Leu Phe Pro Gly Glu Cys Gln Tyr
115 120 125
Val Leu Val Gln Asp Tyr Cys Gly Ser Asn Pro Gly Thr Phe Arg Ile
130 135 140
Leu Val Gly Asn Lys Gly Cys Ser His Pro Ser Val Lys Cys Lys Lys
145 150 155 160
Arg Val Thr Ile Leu Val Glu Gly Gly Glu Ile Glu Leu Phe Asp Gly
165 170 175
Glu Val Asn Val Lys Arg Pro Met Lys Asp Glu Thr His Phe Glu Val
180 185 190
Val Glu Ser Gly Arg Tyr Ile Ile Leu Leu Leu Gly Lys Ala Leu Ser
195 200 205
Val Val Trp Asp Arg His Leu Ser Ile Ser Val Val Leu Lys Gln Thr
210 215 220
Tyr Gln Glu Lys Val Cys Gly Leu Cys Gly Asn Phe Asp Gly Ile Gln
225 230 235 240
Asn Asn Asp Leu Thr Ser Ser Asn Leu Gln Val Glu Glu Asp Pro Val
245 250 255
Asp Phe Gly Asn Ser Trp Lys Val Ser Ser Gln Cys Ala Asp Thr Arg
260 265 270
Lys Val Pro Leu Asp Ser Ser Pro Ala Thr Cys His Asn Asn Ile Met
275 280 285
Lys Gln Thr Met Val Asp Ser Ser Cys Arg Ile Leu Thr Ser Asp Val
290 295 300
Phe Gln Asp Cys Asn Lys Leu Val Asp Pro Glu Pro Tyr Leu Asp Val
305 310 315 320
Cys Ile Tyr Asp Thr Cys Ser Cys Glu Ser Ile Gly Asp Cys Ala Cys
325 330 335
Phe Cys Asp Thr Ile Ala Ala Tyr Ala His Val Cys Ala Gln His Gly
340 345 350
Lys Val Val Thr Trp Arg Thr Ala Thr Leu Cys Pro Gln Ser Cys Glu
355 360 365
Glu Arg Asn Leu Arg Glu Asn Gly Tyr Glu Cys Glu Trp Arg Tyr Asn
370 375 380
Ser Cys Ala Pro Ala Cys Gln Val Thr Cys Gln His Pro Glu Pro Leu
385 390 395 400
Ala Cys Pro Val Gln Cys Val Glu Gly Cys His Ala His Cys Pro Pro
405 410 415
Gly Lys Ile Leu Asp Glu Leu Leu Gln Thr Cys Val Asp Pro Glu Asp
420 425 430
Cys Pro Val Cys Glu Val Ala Gly Arg Arg Phe Ala Ser Gly Lys Lys
435 440 445
Val Thr Leu Asn Pro Ser Asp Pro Glu His Cys Gln Ile Cys His Cys
450 455 460
Asp Val Val Asn Leu Thr Cys Glu Ala Cys Gln Glu Pro Gly Gly Leu
465 470 475 480
Val Val Pro Pro Thr Asp Ala Pro Val Ser Pro Thr Thr Leu Tyr Val
485 490 495
Glu Asp Ile Ser Glu Pro Pro Leu His Gln Glu Pro Gly Gly Leu Val
500 505 510
Val Pro Pro Thr Asp Ala Pro Val Ser Pro Thr Thr Leu Tyr Val Glu
515 520 525
Asp Ile Ser Glu Pro Pro Leu His Gln Glu Pro Gly Gly Leu Val Val
530 535 540
Pro Pro Thr Asp Ala Pro Val Ser Pro Thr Thr Leu Tyr Val Glu Asp
545 550 555 560
Ile Ser Glu Pro Pro Leu His
565
<210> 3
<211> 589
<212> БЕЛОК
<213> Homo sapiens
<400> 3
Met Ile Pro Ala Arg Phe Ala Gly Val Leu Leu Ala Leu Ala Leu Ile
1 5 10 15
Leu Pro Gly Thr Leu Cys Ser Leu Ser Cys Arg Pro Pro Met Val Lys
20 25 30
Leu Val Cys Pro Ala Asp Asn Leu Arg Ala Glu Gly Leu Glu Cys Thr
35 40 45
Lys Thr Cys Gln Asn Tyr Asp Leu Glu Cys Met Ser Met Gly Cys Val
50 55 60
Ser Gly Cys Leu Cys Pro Pro Gly Met Val Arg His Glu Asn Arg Cys
65 70 75 80
Val Ala Leu Glu Arg Cys Pro Cys Phe His Gln Gly Lys Glu Tyr Ala
85 90 95
Pro Gly Glu Thr Val Lys Ile Gly Cys Asn Thr Cys Val Cys Gln Asp
100 105 110
Arg Lys Trp Asn Cys Thr Asp His Val Cys Asp Ala Thr Cys Ser Thr
115 120 125
Ile Gly Met Ala His Tyr Leu Thr Phe Asp Gly Leu Lys Tyr Leu Phe
130 135 140
Pro Gly Glu Cys Gln Tyr Val Leu Val Gln Asp Tyr Cys Gly Ser Asn
145 150 155 160
Pro Gly Thr Phe Arg Ile Leu Val Gly Asn Lys Gly Cys Ser His Pro
165 170 175
Ser Val Lys Cys Lys Lys Arg Val Thr Ile Leu Val Glu Gly Gly Glu
180 185 190
Ile Glu Leu Phe Asp Gly Glu Val Asn Val Lys Arg Pro Met Lys Asp
195 200 205
Glu Thr His Phe Glu Val Val Glu Ser Gly Arg Tyr Ile Ile Leu Leu
210 215 220
Leu Gly Lys Ala Leu Ser Val Val Trp Asp Arg His Leu Ser Ile Ser
225 230 235 240
Val Val Leu Lys Gln Thr Tyr Gln Glu Lys Val Cys Gly Leu Cys Gly
245 250 255
Asn Phe Asp Gly Ile Gln Asn Asn Asp Leu Thr Ser Ser Asn Leu Gln
260 265 270
Val Glu Glu Asp Pro Val Asp Phe Gly Asn Ser Trp Lys Val Ser Ser
275 280 285
Gln Cys Ala Asp Thr Arg Lys Val Pro Leu Asp Ser Ser Pro Ala Thr
290 295 300
Cys His Asn Asn Ile Met Lys Gln Thr Met Val Asp Ser Ser Cys Arg
305 310 315 320
Ile Leu Thr Ser Asp Val Phe Gln Asp Cys Asn Lys Leu Val Asp Pro
325 330 335
Glu Pro Tyr Leu Asp Val Cys Ile Tyr Asp Thr Cys Ser Cys Glu Ser
340 345 350
Ile Gly Asp Cys Ala Cys Phe Cys Asp Thr Ile Ala Ala Tyr Ala His
355 360 365
Val Cys Ala Gln His Gly Lys Val Val Thr Trp Arg Thr Ala Thr Leu
370 375 380
Cys Pro Gln Ser Cys Glu Glu Arg Asn Leu Arg Glu Asn Gly Tyr Glu
385 390 395 400
Cys Glu Trp Arg Tyr Asn Ser Cys Ala Pro Ala Cys Gln Val Thr Cys
405 410 415
Gln His Pro Glu Pro Leu Ala Cys Pro Val Gln Cys Val Glu Gly Cys
420 425 430
His Ala His Cys Pro Pro Gly Lys Ile Leu Asp Glu Leu Leu Gln Thr
435 440 445
Cys Val Asp Pro Glu Asp Cys Pro Val Cys Glu Val Ala Gly Arg Arg
450 455 460
Phe Ala Ser Gly Lys Lys Val Thr Leu Asn Pro Ser Asp Pro Glu His
465 470 475 480
Cys Gln Ile Cys His Cys Asp Val Val Asn Leu Thr Cys Glu Ala Cys
485 490 495
Gln Glu Pro Gly Gly Leu Val Val Pro Pro Thr Asp Ala Pro Val Ser
500 505 510
Pro Thr Thr Leu Tyr Val Glu Asp Ile Ser Glu Pro Pro Leu His Gln
515 520 525
Glu Pro Gly Gly Leu Val Val Pro Pro Thr Asp Ala Pro Val Ser Pro
530 535 540
Thr Thr Leu Tyr Val Glu Asp Ile Ser Glu Pro Pro Leu His Gln Glu
545 550 555 560
Pro Gly Gly Leu Val Val Pro Pro Thr Asp Ala Pro Val Ser Pro Thr
565 570 575
Thr Leu Tyr Val Glu Asp Ile Ser Glu Pro Pro Leu His
580 585
<210> 4
<211> 1330
<212> БЕЛОК
<213> Homo sapiens
<400> 4
Met Ile Pro Ala Arg Phe Ala Gly Val Leu Leu Ala Leu Ala Leu Ile
1 5 10 15
Leu Pro Gly Thr Leu Cys Ala Glu Gly Thr Arg Gly Arg Ser Ser Thr
20 25 30
Ala Arg Cys Ser Leu Phe Gly Ser Asp Phe Val Asn Thr Phe Asp Gly
35 40 45
Ser Met Tyr Ser Phe Ala Gly Tyr Cys Ser Tyr Leu Leu Ala Gly Gly
50 55 60
Cys Gln Lys Arg Ser Phe Ser Ile Ile Gly Asp Phe Gln Asn Gly Lys
65 70 75 80
Arg Val Ser Leu Ser Val Tyr Leu Gly Glu Phe Phe Asp Ile His Leu
85 90 95
Phe Val Asn Gly Thr Val Thr Gln Gly Asp Gln Arg Val Ser Met Pro
100 105 110
Tyr Ala Ser Lys Gly Leu Tyr Leu Glu Thr Glu Ala Gly Tyr Tyr Lys
115 120 125
Leu Ser Gly Glu Ala Tyr Gly Phe Val Ala Arg Ile Asp Gly Ser Gly
130 135 140
Asn Phe Gln Val Leu Leu Ser Asp Arg Tyr Phe Asn Lys Thr Cys Gly
145 150 155 160
Leu Cys Gly Asn Phe Asn Ile Phe Ala Glu Asp Asp Phe Met Thr Gln
165 170 175
Glu Gly Thr Leu Thr Ser Asp Pro Tyr Asp Phe Ala Asn Ser Trp Ala
180 185 190
Leu Ser Ser Gly Glu Gln Trp Cys Glu Arg Ala Ser Pro Pro Ser Ser
195 200 205
Ser Cys Asn Ile Ser Ser Gly Glu Met Gln Lys Gly Leu Trp Glu Gln
210 215 220
Cys Gln Leu Leu Lys Ser Thr Ser Val Phe Ala Arg Cys His Pro Leu
225 230 235 240
Val Asp Pro Glu Pro Phe Val Ala Leu Cys Glu Lys Thr Leu Cys Glu
245 250 255
Cys Ala Gly Gly Leu Glu Cys Ala Cys Pro Ala Leu Leu Glu Tyr Ala
260 265 270
Arg Thr Cys Ala Gln Glu Gly Met Val Leu Tyr Gly Trp Thr Asp His
275 280 285
Ser Ala Cys Ser Pro Val Cys Pro Ala Gly Met Glu Tyr Arg Gln Cys
290 295 300
Val Ser Pro Cys Ala Arg Thr Cys Gln Ser Leu His Ile Asn Glu Met
305 310 315 320
Cys Gln Glu Arg Cys Val Asp Gly Cys Ser Cys Pro Glu Gly Gln Leu
325 330 335
Leu Asp Glu Gly Leu Cys Val Glu Ser Thr Glu Cys Pro Cys Val His
340 345 350
Ser Gly Lys Arg Tyr Pro Pro Gly Thr Ser Leu Ser Arg Asp Cys Asn
355 360 365
Thr Cys Ile Cys Arg Asn Ser Gln Trp Ile Cys Ser Asn Glu Glu Cys
370 375 380
Pro Gly Glu Cys Leu Val Thr Gly Gln Ser His Phe Lys Ser Phe Asp
385 390 395 400
Asn Arg Tyr Phe Thr Phe Ser Gly Ile Cys Gln Tyr Leu Leu Ala Arg
405 410 415
Asp Cys Gln Asp His Ser Phe Ser Ile Val Ile Glu Thr Val Gln Cys
420 425 430
Ala Asp Asp Arg Asp Ala Val Cys Thr Arg Ser Val Thr Val Arg Leu
435 440 445
Pro Gly Leu His Asn Ser Leu Val Lys Leu Lys His Gly Ala Gly Val
450 455 460
Ala Met Asp Gly Gln Asp Val Gln Leu Pro Leu Leu Lys Gly Asp Leu
465 470 475 480
Arg Ile Gln His Thr Val Thr Ala Ser Val Arg Leu Ser Tyr Gly Glu
485 490 495
Asp Leu Gln Met Asp Trp Asp Gly Arg Gly Arg Leu Leu Val Lys Leu
500 505 510
Ser Pro Val Tyr Ala Gly Lys Thr Cys Gly Leu Cys Gly Asn Tyr Asn
515 520 525
Gly Asn Gln Gly Asp Asp Phe Leu Thr Pro Ser Gly Leu Ala Glu Pro
530 535 540
Arg Val Glu Asp Phe Gly Asn Ala Trp Lys Leu His Gly Asp Cys Gln
545 550 555 560
Asp Leu Gln Lys Gln His Ser Asp Pro Cys Ala Leu Asn Pro Arg Met
565 570 575
Thr Arg Phe Ser Glu Glu Ala Cys Ala Val Leu Thr Ser Pro Thr Phe
580 585 590
Glu Ala Cys His Arg Ala Val Ser Pro Leu Pro Tyr Leu Arg Asn Cys
595 600 605
Arg Tyr Asp Val Cys Ser Cys Ser Asp Gly Arg Glu Cys Leu Cys Gly
610 615 620
Ala Leu Ala Ser Tyr Ala Ala Ala Cys Ala Gly Arg Gly Val Arg Val
625 630 635 640
Ala Trp Arg Glu Pro Gly Arg Cys Glu Leu Asn Cys Pro Lys Gly Gln
645 650 655
Val Tyr Leu Gln Cys Gly Thr Pro Cys Asn Leu Thr Cys Arg Ser Leu
660 665 670
Ser Tyr Pro Asp Glu Glu Cys Asn Glu Ala Cys Leu Glu Gly Cys Phe
675 680 685
Cys Pro Pro Gly Leu Tyr Met Asp Glu Arg Gly Asp Cys Val Pro Lys
690 695 700
Ala Gln Cys Pro Cys Tyr Tyr Asp Gly Glu Ile Phe Gln Pro Glu Asp
705 710 715 720
Ile Phe Ser Asp His His Thr Met Cys Tyr Cys Glu Asp Gly Phe Met
725 730 735
His Cys Thr Met Ser Gly Val Pro Gly Ser Leu Leu Pro Asp Ala Val
740 745 750
Leu Ser Ser Pro Leu Ser His Arg Ser Lys Arg Ser Leu Ser Cys Arg
755 760 765
Pro Pro Met Val Lys Leu Val Cys Pro Ala Asp Asn Leu Arg Ala Glu
770 775 780
Gly Leu Glu Cys Thr Lys Thr Cys Gln Asn Tyr Asp Leu Glu Cys Met
785 790 795 800
Ser Met Gly Cys Val Ser Gly Cys Leu Cys Pro Pro Gly Met Val Arg
805 810 815
His Glu Asn Arg Cys Val Ala Leu Glu Arg Cys Pro Cys Phe His Gln
820 825 830
Gly Lys Glu Tyr Ala Pro Gly Glu Thr Val Lys Ile Gly Cys Asn Thr
835 840 845
Cys Val Cys Gln Asp Arg Lys Trp Asn Cys Thr Asp His Val Cys Asp
850 855 860
Ala Thr Cys Ser Thr Ile Gly Met Ala His Tyr Leu Thr Phe Asp Gly
865 870 875 880
Leu Lys Tyr Leu Phe Pro Gly Glu Cys Gln Tyr Val Leu Val Gln Asp
885 890 895
Tyr Cys Gly Ser Asn Pro Gly Thr Phe Arg Ile Leu Val Gly Asn Lys
900 905 910
Gly Cys Ser His Pro Ser Val Lys Cys Lys Lys Arg Val Thr Ile Leu
915 920 925
Val Glu Gly Gly Glu Ile Glu Leu Phe Asp Gly Glu Val Asn Val Lys
930 935 940
Arg Pro Met Lys Asp Glu Thr His Phe Glu Val Val Glu Ser Gly Arg
945 950 955 960
Tyr Ile Ile Leu Leu Leu Gly Lys Ala Leu Ser Val Val Trp Asp Arg
965 970 975
His Leu Ser Ile Ser Val Val Leu Lys Gln Thr Tyr Gln Glu Lys Val
980 985 990
Cys Gly Leu Cys Gly Asn Phe Asp Gly Ile Gln Asn Asn Asp Leu Thr
995 1000 1005
Ser Ser Asn Leu Gln Val Glu Glu Asp Pro Val Asp Phe Gly Asn
1010 1015 1020
Ser Trp Lys Val Ser Ser Gln Cys Ala Asp Thr Arg Lys Val Pro
1025 1030 1035
Leu Asp Ser Ser Pro Ala Thr Cys His Asn Asn Ile Met Lys Gln
1040 1045 1050
Thr Met Val Asp Ser Ser Cys Arg Ile Leu Thr Ser Asp Val Phe
1055 1060 1065
Gln Asp Cys Asn Lys Leu Val Asp Pro Glu Pro Tyr Leu Asp Val
1070 1075 1080
Cys Ile Tyr Asp Thr Cys Ser Cys Glu Ser Ile Gly Asp Cys Ala
1085 1090 1095
Cys Phe Cys Asp Thr Ile Ala Ala Tyr Ala His Val Cys Ala Gln
1100 1105 1110
His Gly Lys Val Val Thr Trp Arg Thr Ala Thr Leu Cys Pro Gln
1115 1120 1125
Ser Cys Glu Glu Arg Asn Leu Arg Glu Asn Gly Tyr Glu Cys Glu
1130 1135 1140
Trp Arg Tyr Asn Ser Cys Ala Pro Ala Cys Gln Val Thr Cys Gln
1145 1150 1155
His Pro Glu Pro Leu Ala Cys Pro Val Gln Cys Val Glu Gly Cys
1160 1165 1170
His Ala His Cys Pro Pro Gly Lys Ile Leu Asp Glu Leu Leu Gln
1175 1180 1185
Thr Cys Val Asp Pro Glu Asp Cys Pro Val Cys Glu Val Ala Gly
1190 1195 1200
Arg Arg Phe Ala Ser Gly Lys Lys Val Thr Leu Asn Pro Ser Asp
1205 1210 1215
Pro Glu His Cys Gln Ile Cys His Cys Asp Val Val Asn Leu Thr
1220 1225 1230
Cys Glu Ala Cys Gln Glu Pro Gly Gly Leu Val Val Pro Pro Thr
1235 1240 1245
Asp Ala Pro Val Ser Pro Thr Thr Leu Tyr Val Glu Asp Ile Ser
1250 1255 1260
Glu Pro Pro Leu His Gln Glu Pro Gly Gly Leu Val Val Pro Pro
1265 1270 1275
Thr Asp Ala Pro Val Ser Pro Thr Thr Leu Tyr Val Glu Asp Ile
1280 1285 1290
Ser Glu Pro Pro Leu His Gln Glu Pro Gly Gly Leu Val Val Pro
1295 1300 1305
Pro Thr Asp Ala Pro Val Ser Pro Thr Thr Leu Tyr Val Glu Asp
1310 1315 1320
Ile Ser Glu Pro Pro Leu His
1325 1330
<210> 5
<211> 1423
<212> БЕЛОК
<213> Homo sapiens
<400> 5
Met Ile Pro Ala Arg Phe Ala Gly Val Leu Leu Ala Leu Ala Leu Ile
1 5 10 15
Leu Pro Gly Thr Leu Cys Ala Glu Gly Thr Arg Gly Arg Ser Ser Thr
20 25 30
Ala Arg Cys Ser Leu Phe Gly Ser Asp Phe Val Asn Thr Phe Asp Gly
35 40 45
Ser Met Tyr Ser Phe Ala Gly Tyr Cys Ser Tyr Leu Leu Ala Gly Gly
50 55 60
Cys Gln Lys Arg Ser Phe Ser Ile Ile Gly Asp Phe Gln Asn Gly Lys
65 70 75 80
Arg Val Ser Leu Ser Val Tyr Leu Gly Glu Phe Phe Asp Ile His Leu
85 90 95
Phe Val Asn Gly Thr Val Thr Gln Gly Asp Gln Arg Val Ser Met Pro
100 105 110
Tyr Ala Ser Lys Gly Leu Tyr Leu Glu Thr Glu Ala Gly Tyr Tyr Lys
115 120 125
Leu Ser Gly Glu Ala Tyr Gly Phe Val Ala Arg Ile Asp Gly Ser Gly
130 135 140
Asn Phe Gln Val Leu Leu Ser Asp Arg Tyr Phe Asn Lys Thr Cys Gly
145 150 155 160
Leu Cys Gly Asn Phe Asn Ile Phe Ala Glu Asp Asp Phe Met Thr Gln
165 170 175
Glu Gly Thr Leu Thr Ser Asp Pro Tyr Asp Phe Ala Asn Ser Trp Ala
180 185 190
Leu Ser Ser Gly Glu Gln Trp Cys Glu Arg Ala Ser Pro Pro Ser Ser
195 200 205
Ser Cys Asn Ile Ser Ser Gly Glu Met Gln Lys Gly Leu Trp Glu Gln
210 215 220
Cys Gln Leu Leu Lys Ser Thr Ser Val Phe Ala Arg Cys His Pro Leu
225 230 235 240
Val Asp Pro Glu Pro Phe Val Ala Leu Cys Glu Lys Thr Leu Cys Glu
245 250 255
Cys Ala Gly Gly Leu Glu Cys Ala Cys Pro Ala Leu Leu Glu Tyr Ala
260 265 270
Arg Thr Cys Ala Gln Glu Gly Met Val Leu Tyr Gly Trp Thr Asp His
275 280 285
Ser Ala Cys Ser Pro Val Cys Pro Ala Gly Met Glu Tyr Arg Gln Cys
290 295 300
Val Ser Pro Cys Ala Arg Thr Cys Gln Ser Leu His Ile Asn Glu Met
305 310 315 320
Cys Gln Glu Arg Cys Val Asp Gly Cys Ser Cys Pro Glu Gly Gln Leu
325 330 335
Leu Asp Glu Gly Leu Cys Val Glu Ser Thr Glu Cys Pro Cys Val His
340 345 350
Ser Gly Lys Arg Tyr Pro Pro Gly Thr Ser Leu Ser Arg Asp Cys Asn
355 360 365
Thr Cys Ile Cys Arg Asn Ser Gln Trp Ile Cys Ser Asn Glu Glu Cys
370 375 380
Pro Gly Glu Cys Leu Val Thr Gly Gln Ser His Phe Lys Ser Phe Asp
385 390 395 400
Asn Arg Tyr Phe Thr Phe Ser Gly Ile Cys Gln Tyr Leu Leu Ala Arg
405 410 415
Asp Cys Gln Asp His Ser Phe Ser Ile Val Ile Glu Thr Val Gln Cys
420 425 430
Ala Asp Asp Arg Asp Ala Val Cys Thr Arg Ser Val Thr Val Arg Leu
435 440 445
Pro Gly Leu His Asn Ser Leu Val Lys Leu Lys His Gly Ala Gly Val
450 455 460
Ala Met Asp Gly Gln Asp Val Gln Leu Pro Leu Leu Lys Gly Asp Leu
465 470 475 480
Arg Ile Gln His Thr Val Thr Ala Ser Val Arg Leu Ser Tyr Gly Glu
485 490 495
Asp Leu Gln Met Asp Trp Asp Gly Arg Gly Arg Leu Leu Val Lys Leu
500 505 510
Ser Pro Val Tyr Ala Gly Lys Thr Cys Gly Leu Cys Gly Asn Tyr Asn
515 520 525
Gly Asn Gln Gly Asp Asp Phe Leu Thr Pro Ser Gly Leu Ala Glu Pro
530 535 540
Arg Val Glu Asp Phe Gly Asn Ala Trp Lys Leu His Gly Asp Cys Gln
545 550 555 560
Asp Leu Gln Lys Gln His Ser Asp Pro Cys Ala Leu Asn Pro Arg Met
565 570 575
Thr Arg Phe Ser Glu Glu Ala Cys Ala Val Leu Thr Ser Pro Thr Phe
580 585 590
Glu Ala Cys His Arg Ala Val Ser Pro Leu Pro Tyr Leu Arg Asn Cys
595 600 605
Arg Tyr Asp Val Cys Ser Cys Ser Asp Gly Arg Glu Cys Leu Cys Gly
610 615 620
Ala Leu Ala Ser Tyr Ala Ala Ala Cys Ala Gly Arg Gly Val Arg Val
625 630 635 640
Ala Trp Arg Glu Pro Gly Arg Cys Glu Leu Asn Cys Pro Lys Gly Gln
645 650 655
Val Tyr Leu Gln Cys Gly Thr Pro Cys Asn Leu Thr Cys Arg Ser Leu
660 665 670
Ser Tyr Pro Asp Glu Glu Cys Asn Glu Ala Cys Leu Glu Gly Cys Phe
675 680 685
Cys Pro Pro Gly Leu Tyr Met Asp Glu Arg Gly Asp Cys Val Pro Lys
690 695 700
Ala Gln Cys Pro Cys Tyr Tyr Asp Gly Glu Ile Phe Gln Pro Glu Asp
705 710 715 720
Ile Phe Ser Asp His His Thr Met Cys Tyr Cys Glu Asp Gly Phe Met
725 730 735
His Cys Thr Met Ser Gly Val Pro Gly Ser Leu Leu Pro Asp Ala Val
740 745 750
Leu Ser Ser Pro Leu Ser His Arg Ser Lys Arg Ser Leu Ser Cys Arg
755 760 765
Pro Pro Met Val Lys Leu Val Cys Pro Ala Asp Asn Leu Arg Ala Glu
770 775 780
Gly Leu Glu Cys Thr Lys Thr Cys Gln Asn Tyr Asp Leu Glu Cys Met
785 790 795 800
Ser Met Gly Cys Val Ser Gly Cys Leu Cys Pro Pro Gly Met Val Arg
805 810 815
His Glu Asn Arg Cys Val Ala Leu Glu Arg Cys Pro Cys Phe His Gln
820 825 830
Gly Lys Glu Tyr Ala Pro Gly Glu Thr Val Lys Ile Gly Cys Asn Thr
835 840 845
Cys Val Cys Gln Asp Arg Lys Trp Asn Cys Thr Asp His Val Cys Asp
850 855 860
Ala Thr Cys Ser Thr Ile Gly Met Ala His Tyr Leu Thr Phe Asp Gly
865 870 875 880
Leu Lys Tyr Leu Phe Pro Gly Glu Cys Gln Tyr Val Leu Val Gln Asp
885 890 895
Tyr Cys Gly Ser Asn Pro Gly Thr Phe Arg Ile Leu Val Gly Asn Lys
900 905 910
Gly Cys Ser His Pro Ser Val Lys Cys Lys Lys Arg Val Thr Ile Leu
915 920 925
Val Glu Gly Gly Glu Ile Glu Leu Phe Asp Gly Glu Val Asn Val Lys
930 935 940
Arg Pro Met Lys Asp Glu Thr His Phe Glu Val Val Glu Ser Gly Arg
945 950 955 960
Tyr Ile Ile Leu Leu Leu Gly Lys Ala Leu Ser Val Val Trp Asp Arg
965 970 975
His Leu Ser Ile Ser Val Val Leu Lys Gln Thr Tyr Gln Glu Lys Val
980 985 990
Cys Gly Leu Cys Gly Asn Phe Asp Gly Ile Gln Asn Asn Asp Leu Thr
995 1000 1005
Ser Ser Asn Leu Gln Val Glu Glu Asp Pro Val Asp Phe Gly Asn
1010 1015 1020
Ser Trp Lys Val Ser Ser Gln Cys Ala Asp Thr Arg Lys Val Pro
1025 1030 1035
Leu Asp Ser Ser Pro Ala Thr Cys His Asn Asn Ile Met Lys Gln
1040 1045 1050
Thr Met Val Asp Ser Ser Cys Arg Ile Leu Thr Ser Asp Val Phe
1055 1060 1065
Gln Asp Cys Asn Lys Leu Val Asp Pro Glu Pro Tyr Leu Asp Val
1070 1075 1080
Cys Ile Tyr Asp Thr Cys Ser Cys Glu Ser Ile Gly Asp Cys Ala
1085 1090 1095
Cys Phe Cys Asp Thr Ile Ala Ala Tyr Ala His Val Cys Ala Gln
1100 1105 1110
His Gly Lys Val Val Thr Trp Arg Thr Ala Thr Leu Cys Pro Gln
1115 1120 1125
Ser Cys Glu Glu Arg Asn Leu Arg Glu Asn Gly Tyr Glu Cys Glu
1130 1135 1140
Trp Arg Tyr Asn Ser Cys Ala Pro Ala Cys Gln Val Thr Cys Gln
1145 1150 1155
His Pro Glu Pro Leu Ala Cys Pro Val Gln Cys Val Glu Gly Cys
1160 1165 1170
His Ala His Cys Pro Pro Gly Lys Ile Leu Asp Glu Leu Leu Gln
1175 1180 1185
Thr Cys Val Asp Pro Glu Asp Cys Pro Val Cys Glu Val Ala Gly
1190 1195 1200
Arg Arg Phe Ala Ser Gly Lys Lys Val Thr Leu Asn Pro Ser Asp
1205 1210 1215
Pro Glu His Cys Gln Ile Cys His Cys Asp Val Val Asn Leu Thr
1220 1225 1230
Cys Glu Ala Cys Gln Glu Pro Gly Gly Leu Val Val Pro Pro Thr
1235 1240 1245
Asp Ala Pro Val Ser Pro Thr Thr Leu Tyr Val Glu Asp Ile Ser
1250 1255 1260
Glu Pro Pro Leu His Gln Glu Pro Gly Gly Leu Val Val Pro Pro
1265 1270 1275
Thr Asp Ala Pro Val Ser Pro Thr Thr Leu Tyr Val Glu Asp Ile
1280 1285 1290
Ser Glu Pro Pro Leu His Gln Glu Pro Gly Gly Leu Val Val Pro
1295 1300 1305
Pro Thr Asp Ala Pro Val Pro Thr Thr Leu Tyr Val Glu Asp Ile
1310 1315 1320
Ser Glu Pro Pro Leu His Glu Glu Pro Glu Cys Asn Asp Ile Thr
1325 1330 1335
Ala Arg Leu Gln Tyr Val Lys Val Gly Ser Cys Lys Ser Glu Val
1340 1345 1350
Glu Val Asp Ile His Tyr Cys Gln Gly Lys Cys Ala Ser Lys Ala
1355 1360 1365
Met Tyr Ser Ile Asp Ile Asn Asp Val Gln Asp Gln Cys Ser Cys
1370 1375 1380
Cys Ser Pro Thr Arg Thr Glu Pro Met Gln Val Ala Leu His Cys
1385 1390 1395
Thr Asn Gly Ser Val Val Tyr His Glu Val Leu Asn Ala Met Glu
1400 1405 1410
Cys Lys Cys Ser Pro Arg Lys Cys Ser Lys
1415 1420
<210> 6
<211> 62
<212> БЕЛОК
<213> Homo sapiens
<400> 6
Lys Val Thr Leu Asn Pro Ser Asp Pro Glu His Cys Gln Ile Cys His
1 5 10 15
Cys Asp Val Val Asn Leu Thr Cys Glu Ala Cys Gln Glu Pro Gly Gly
20 25 30
Leu Val Val Pro Pro Thr Asp Ala Pro Val Ser Pro Thr Thr Leu Tyr
35 40 45
Val Glu Asp Ile Ser Glu Pro Pro Leu His Gly Ser Ala Trp
50 55 60
<210> 7
<211> 124
<212> БЕЛОК
<213> Homo sapiens
<400> 7
Lys Val Thr Leu Asn Pro Ser Asp Pro Glu His Cys Gln Ile Cys His
1 5 10 15
Cys Asp Val Val Asn Leu Thr Cys Glu Ala Cys Gln Glu Pro Gly Gly
20 25 30
Leu Val Val Pro Pro Thr Asp Ala Pro Val Ser Pro Thr Thr Leu Tyr
35 40 45
Val Glu Asp Ile Ser Glu Pro Pro Leu His Gln Glu Pro Gly Gly Leu
50 55 60
Val Val Pro Pro Thr Asp Ala Pro Val Ser Pro Thr Thr Leu Tyr Val
65 70 75 80
Glu Asp Ile Ser Glu Pro Pro Leu His Gln Glu Pro Gly Gly Leu Val
85 90 95
Val Pro Pro Thr Asp Ala Pro Val Ser Pro Thr Thr Leu Tyr Val Glu
100 105 110
Asp Ile Ser Glu Pro Pro Leu His Gly Ser Ala Trp
115 120
<210> 8
<211> 23
<212> БЕЛОК
<213> Homo sapiens
<220>
<221> Прочие признаки
<222> (6)..(6)
<223> Xaa может представлять собой любую природную аминокислоту
<220>
<221> Прочие признаки
<222> (10)..(10)
<223> Xaa может представлять собой любую природную аминокислоту
<220>
<221> Прочие признаки
<222> (14)..(14)
<223> Xaa может представлять собой любую природную аминокислоту
<220>
<221> Прочие признаки
<222> (17)..(19)
<223> Xaa может представлять собой любую природную аминокислоту
<220>
<221> Прочие признаки
<222> (21)..(21)
<223> Xaa может представлять собой любую природную аминокислоту
<400> 8
Val Val Pro Pro Thr Xaa Ala Pro Val Xaa Pro Thr Thr Xaa Tyr Val
1 5 10 15
Xaa Xaa Xaa Ser Xaa Pro Pro
20
<210> 9
<211> 23
<212> БЕЛОК
<213> Homo sapiens
<400> 9
Val Val Pro Pro Thr Asp Ala Pro Val Ser Pro Thr Thr Leu Tyr Val
1 5 10 15
Glu Asp Ile Ser Glu Pro Pro
20
<210> 10
<211> 26
<212> БЕЛОК
<213> Homo sapiens
<220>
<221> Прочие признаки
<222> (5)..(5)
<223> Xaa может представлять собой любую природную аминокислоту
<220>
<221> Прочие признаки
<222> (8)..(9)
<223> Xaa может представлять собой любую природную аминокислоту
<220>
<221> Прочие признаки
<222> (11)..(11)
<223> Xaa может представлять собой любую природную аминокислоту
<220>
<221> Прочие признаки
<222> (15)..(15)
<223> Xaa может представлять собой любую природную аминокислоту
<220>
<221> Прочие признаки
<222> (17)..(20)
<223> Xaa может представлять собой любую природную аминокислоту
<220>
<221> Прочие признаки
<222> (24)..(25)
<223> Xaa может представлять собой любую природную аминокислоту
<400> 10
Pro Pro Pro Thr Xaa Pro Pro Xaa Xaa Ala Xaa Val Thr Val Xaa Pro
1 5 10 15
Xaa Xaa Xaa Xaa Val Ser Thr Xaa Xaa Pro
20 25
<210> 11
<211> 26
<212> БЕЛОК
<213> Homo sapiens
<400> 11
Pro Pro Pro Thr Leu Pro Pro Asp Met Ala Gln Val Thr Val Gly Pro
1 5 10 15
Gly Leu Leu Gly Val Ser Thr Leu Gly Pro
20 25
<210> 12
<211> 47
<212> БЕЛОК
<213> Homo sapiens
<220>
<221> Прочие признаки
<222> (6)..(9)
<223> Xaa может представлять собой любую природную аминокислоту
<220>
<221> Прочие признаки
<222> (12)..(12)
<223> Xaa может представлять собой любую природную аминокислоту
<220>
<221> Прочие признаки
<222> (15)..(17)
<223> Xaa может представлять собой любую природную аминокислоту
<220>
<221> Прочие признаки
<222> (20)..(20)
<223> Xaa может представлять собой любую природную аминокислоту
<220>
<221> Прочие признаки
<222> (22)..(23)
<223> Xaa может представлять собой любую природную аминокислоту
<220>
<221> Прочие признаки
<222> (27)..(28)
<223> Xaa может представлять собой любую природную аминокислоту
<220>
<221> Прочие признаки
<222> (32)..(32)
<223> Xaa может представлять собой любую природную аминокислоту
<220>
<221> Прочие признаки
<222> (35)..(37)
<223> Xaa может представлять собой любую природную аминокислоту
<220>
<221> Прочие признаки
<222> (39)..(41)
<223> Xaa может представлять собой любую природную аминокислоту
<220>
<221> Прочие признаки
<222> (44)..(47)
<223> Xaa может представлять собой любую природную аминокислоту
<400> 12
Val Ser Ser Thr Ser Xaa Xaa Xaa Xaa Ser Thr Xaa Pro Ser Xaa Xaa
1 5 10 15
Xaa Ala Ala Xaa Thr Xaa Xaa Thr Ser Ser Xaa Xaa Pro Pro Ser Xaa
20 25 30
Pro Val Xaa Xaa Xaa Ser Xaa Xaa Xaa Thr Thr Xaa Xaa Xaa Xaa
35 40 45
<210> 13
<211> 47
<212> БЕЛОК
<213> Homo sapiens
<400> 13
Val Ser Ser Thr Ser Asn Asn Leu Ile Ser Thr Ile Pro Ser Asp Asn
1 5 10 15
Leu Ala Ala Gly Thr Asp Asp Thr Ser Ser Leu Gly Pro Pro Ser Met
20 25 30
Pro Val His Tyr Asp Ser Gln Leu Asp Thr Thr Leu Phe Gly Lys
35 40 45
<210> 14
<211> 20
<212> БЕЛОК
<213> Homo sapiens
<220>
<221> Прочие признаки
<222> (1)..(3)
<223> Xaa может представлять собой любую природную аминокислоту
<220>
<221> Прочие признаки
<222> (7)..(7)
<223> Xaa может представлять собой любую природную аминокислоту
<220>
<221> Прочие признаки
<222> (9)..(14)
<223> Xaa может представлять собой любую природную аминокислоту
<220>
<221> Прочие признаки
<222> (16)..(16)
<223> Xaa может представлять собой любую природную аминокислоту
<220>
<221> Прочие признаки
<222> (18)..(20)
<223> Xaa может представлять собой любую природную аминокислоту
<400> 14
Xaa Xaa Xaa Ala Thr Thr Xaa Pro Xaa Xaa Xaa Xaa Xaa Xaa Thr Xaa
1 5 10 15
Pro Xaa Xaa Xaa
20
<210> 15
<211> 20
<212> БЕЛОК
<213> Homo sapiens
<400> 15
Gln Phe Asn Ala Thr Thr Ile Pro Glu Asn Asp Ile Glu Lys Thr Asp
1 5 10 15
Pro Trp Phe Ala
20
<210> 16
<211> 10
<212> БЕЛОК
<213> Homo sapiens
<220>
<221> Прочие признаки
<222> (1)..(2)
<223> Xaa может представлять собой любую природную аминокислоту
<220>
<221> Прочие признаки
<222> (8)..(10)
<223> Xaa может представлять собой любую природную аминокислоту
<400> 16
Xaa Xaa Thr Thr Ala Ala Thr Xaa Xaa Xaa
1 5 10
<210> 17
<211> 10
<212> БЕЛОК
<213> Homo sapiens
<400> 17
Leu Gly Thr Thr Ala Ala Thr Glu Leu Lys
1 5 10
<210> 18
<211> 17
<212> БЕЛОК
<213> Homo sapiens
<220>
<221> Прочие признаки
<222> (1)..(2)
<223> Xaa может представлять собой любую природную аминокислоту
<220>
<221> Прочие признаки
<222> (6)..(8)
<223> Xaa может представлять собой любую природную аминокислоту
<220>
<221> Прочие признаки
<222> (10)..(10)
<223> Xaa может представлять собой любую природную аминокислоту
<220>
<221> Прочие признаки
<222> (12)..(14)
<223> Xaa может представлять собой любую природную аминокислоту
<220>
<221> Прочие признаки
<222> (17)..(17)
<223> Xaa может представлять собой любую природную аминокислоту
<400> 18
Xaa Xaa Pro Thr Pro Xaa Xaa Xaa Ser Xaa Ser Xaa Xaa Xaa Glu Ala
1 5 10 15
Xaa
<210> 19
<211> 17
<212> БЕЛОК
<213> Homo sapiens
<400> 19
Gln Ser Pro Thr Pro His Gly Leu Ser Leu Ser Asp Leu Gln Glu Ala
1 5 10 15
Lys
<210> 20
<211> 37
<212> БЕЛОК
<213> Homo sapiens
<220>
<221> Прочие признаки
<222> (2)..(10)
<223> Xaa может представлять собой любую природную аминокислоту
<220>
<221> Прочие признаки
<222> (12)..(12)
<223> Xaa может представлять собой любую природную аминокислоту
<220>
<221> Прочие признаки
<222> (15)..(16)
<223> Xaa может представлять собой любую природную аминокислоту
<220>
<221> Прочие признаки
<222> (19)..(23)
<223> Xaa может представлять собой любую природную аминокислоту
<220>
<221> Прочие признаки
<222> (25)..(26)
<223> Xaa может представлять собой любую природную аминокислоту
<220>
<221> Прочие признаки
<222> (28)..(31)
<223> Xaa может представлять собой любую природную аминокислоту
<220>
<221> Прочие признаки
<222> (33)..(34)
<223> Xaa может представлять собой любую природную аминокислоту
<220>
<221> Прочие признаки
<222> (36)..(37)
<223> Xaa может представлять собой любую природную аминокислоту
<400> 20
Val Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Thr Xaa Thr Ser Xaa Xaa
1 5 10 15
Ser Pro Xaa Xaa Xaa Xaa Xaa Val Xaa Xaa Ser Xaa Xaa Xaa Xaa Thr
20 25 30
Xaa Xaa Ala Xaa Xaa
35
<210> 21
<211> 37
<212> БЕЛОК
<213> Homo sapiens
<400> 21
Val His Ile Tyr Gln Lys Asp Leu Phe Phe Thr Glu Thr Ser Asp Gly
1 5 10 15
Ser Pro Gly His Leu Asp Leu Val Glu Gly Ser Leu Leu Gln Gly Thr
20 25 30
Glu Gly Ala Ile Lys
35
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ГЛИКОЗИЛИРОВАННЫЕ СЛИТЫЕ БЕЛКИ VWF С УЛУЧШЕННОЙ ФАРМАКОКИНЕТИКОЙ | 2017 |
|
RU2782212C2 |
| КОНСТРУКЦИИ, ИМЕЮЩИЕ SIRP-АЛЬФА ДОМЕН ИЛИ ЕГО ВАРИАНТ | 2016 |
|
RU2740672C2 |
| СПОСОБЫ ЛЕЧЕНИЯ ГЕМОФИЛИИ A | 2019 |
|
RU2812863C2 |
| САЙТ-СПЕЦИФИЧЕСКАЯ КОНЪЮГАЦИЯ АНТИТЕЛО-ЛЕКАРСТВЕННОЕ СРЕДСТВО ПОСРЕДСТВОМ ГЛИКОИНЖЕНЕРИИ | 2014 |
|
RU2831409C2 |
| АНТИТЕЛА ПРОТИВ SIGLEC-15 И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2759334C2 |
| АНТИТЕЛО ПРОТИВ MUC1 | 2019 |
|
RU2792347C2 |
| САМОСОБИРАЮЩИЕСЯ НАНОСТРУКТУРНЫЕ ВАКЦИНЫ | 2019 |
|
RU2811439C2 |
| МУТАЦИИ В ГЕНАХ OAS1 | 2006 |
|
RU2465328C2 |
| СКОНСТРУИРОВАННЫЕ PH-ЗАВИСИМЫЕ АНТИТЕЛА К CD3, А ТАКЖЕ СПОСОБЫ ИХ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ | 2020 |
|
RU2832079C2 |
| СПОСОБЫ ЛЕЧЕНИЯ ТТП ИММУНОГЛОБУЛИНОВЫМИ ОДИНОЧНЫМИ ВАРИАБЕЛЬНЫМИ ДОМЕНАМИ И ИХ ПРИМЕНЕНИЕ | 2015 |
|
RU2807602C2 |
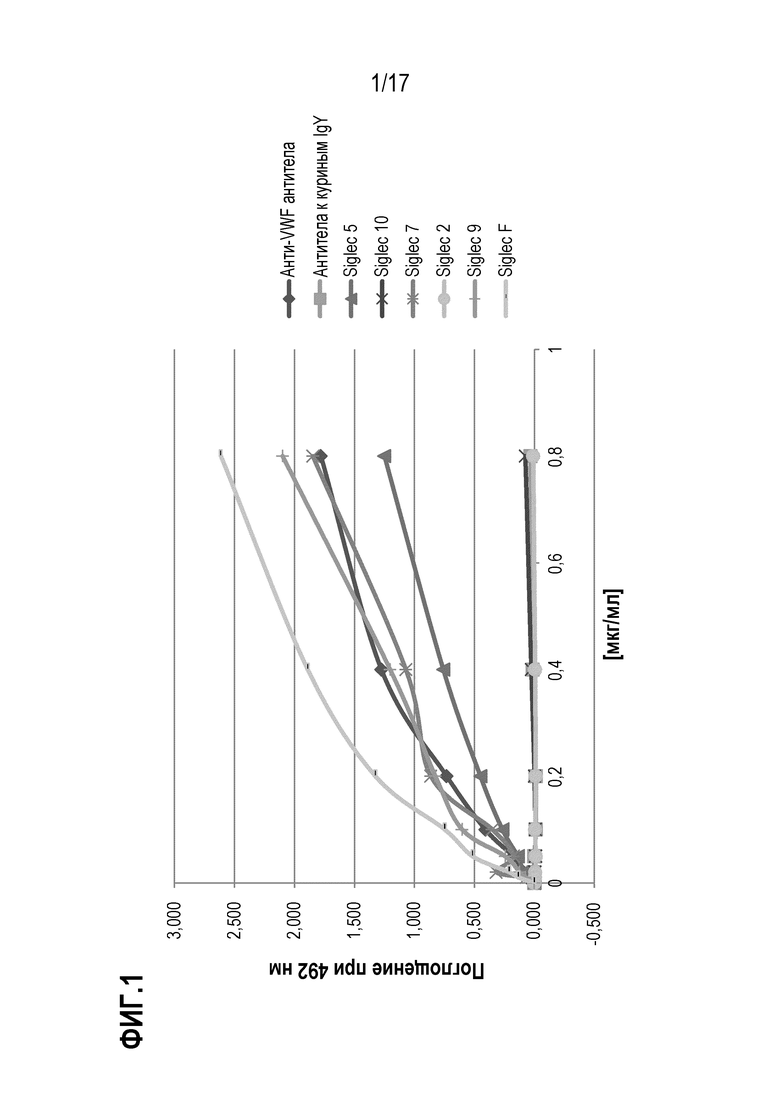
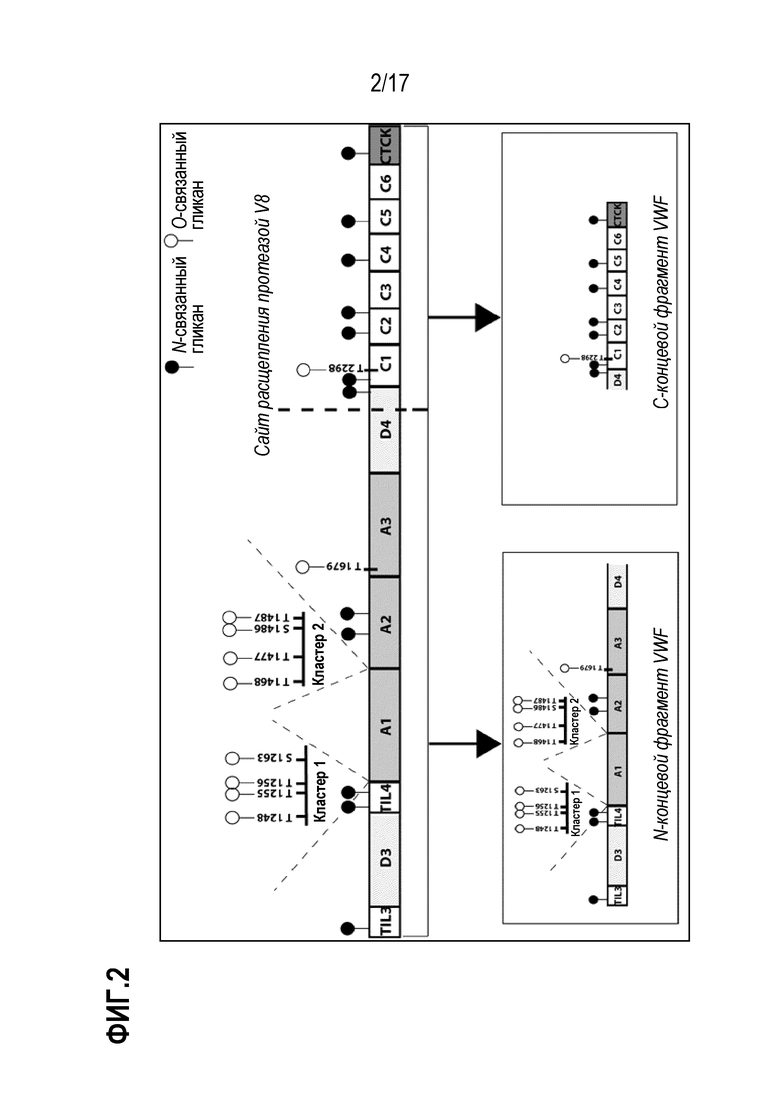
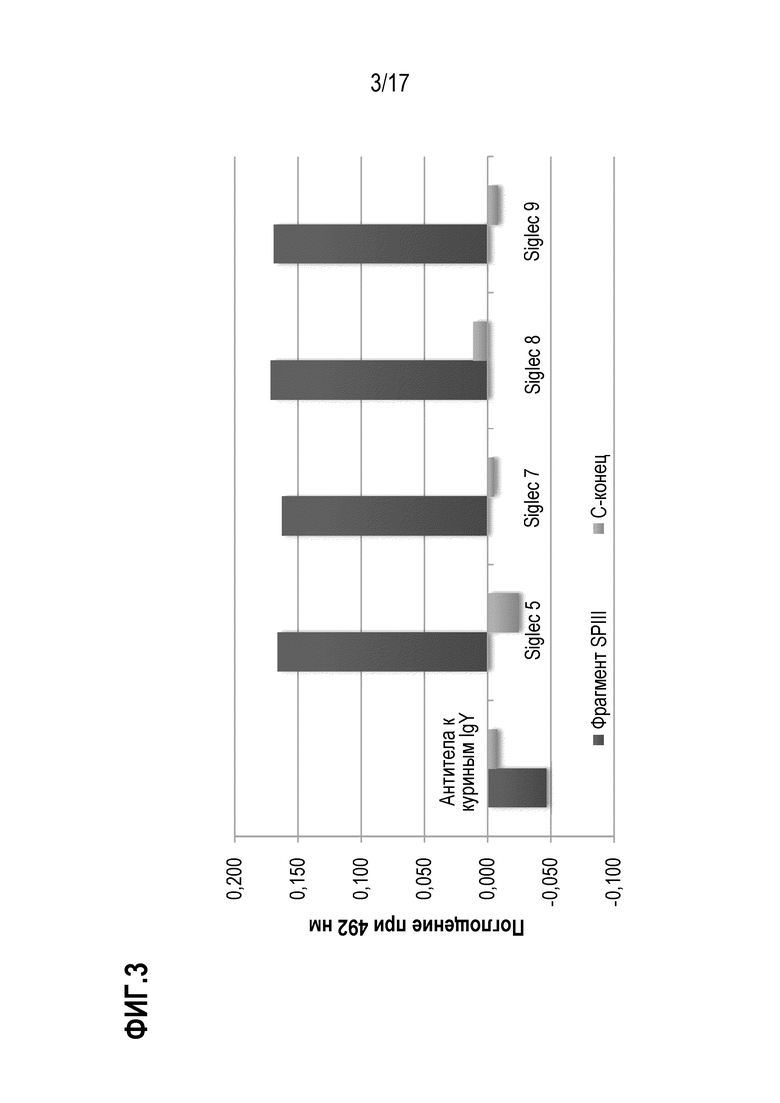
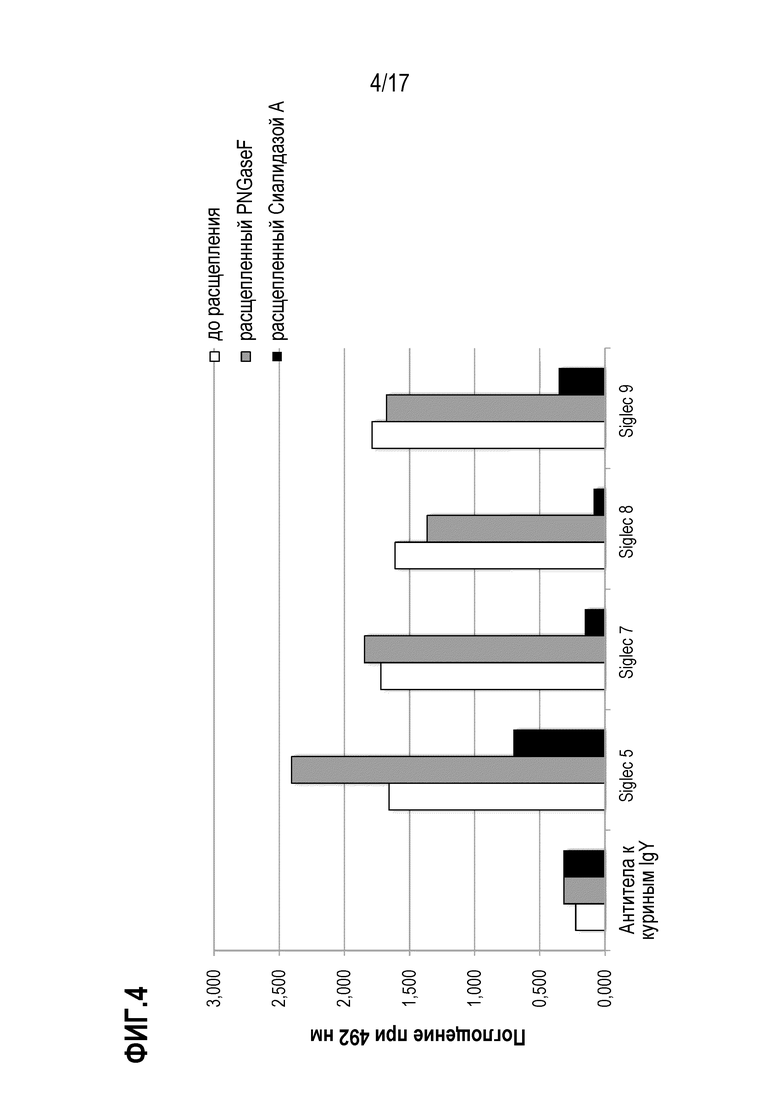
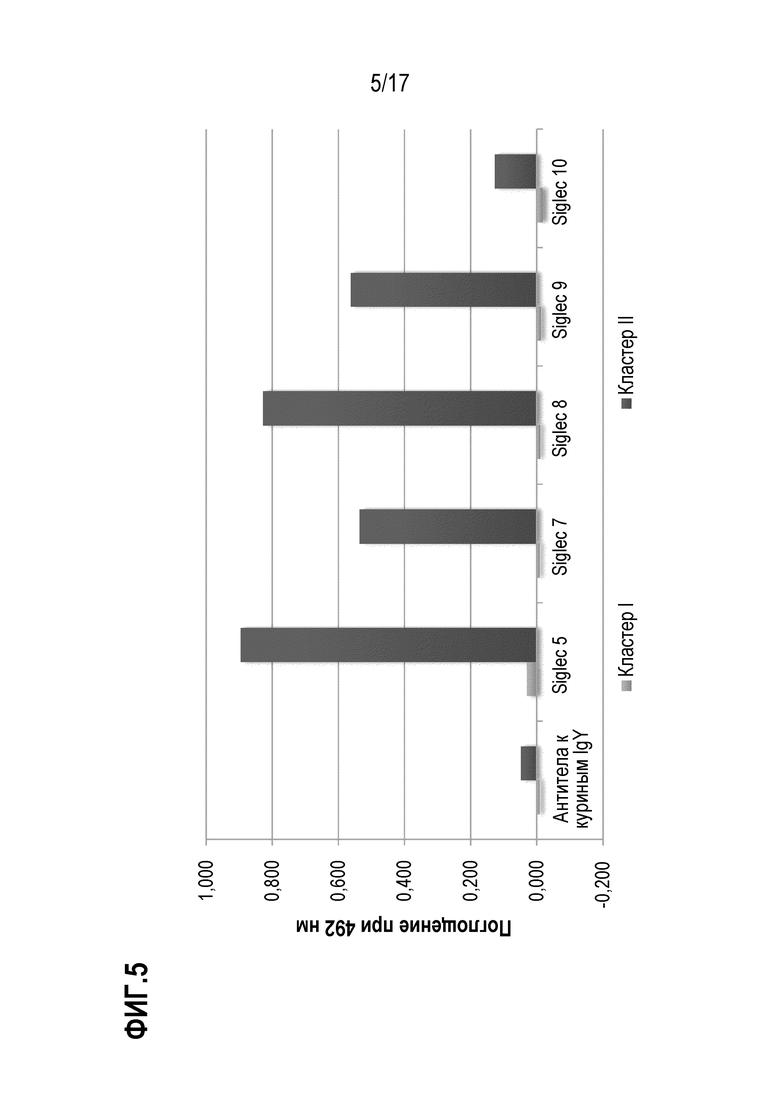
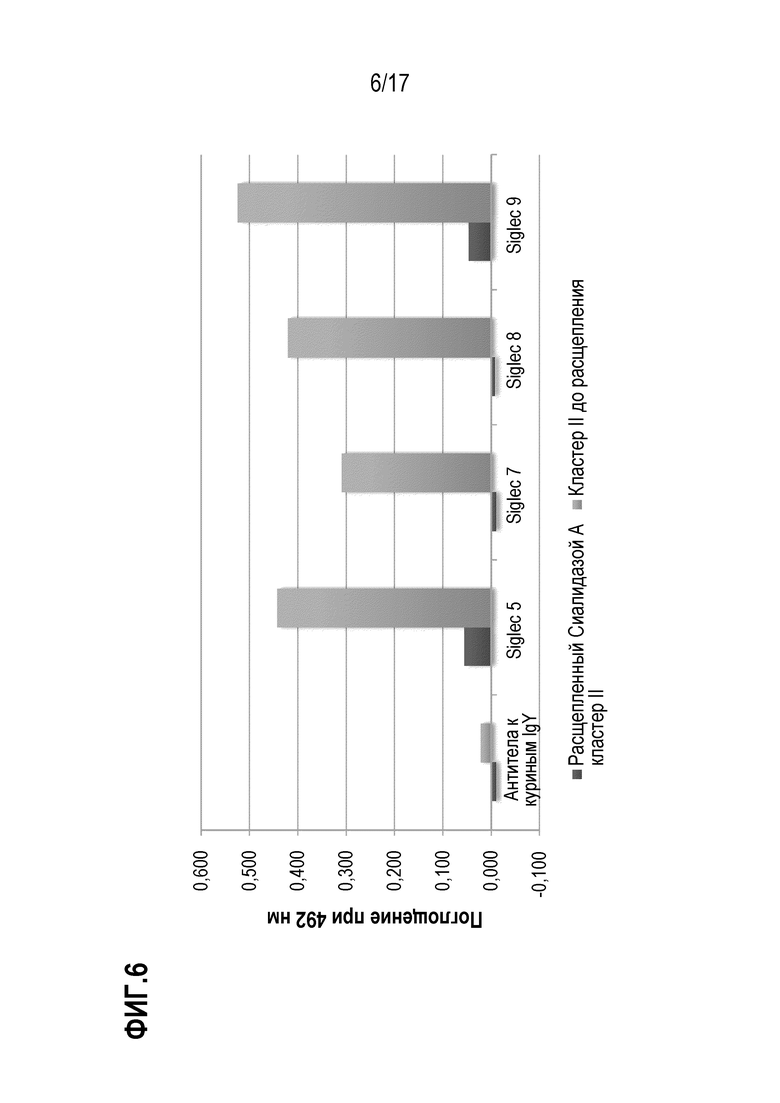
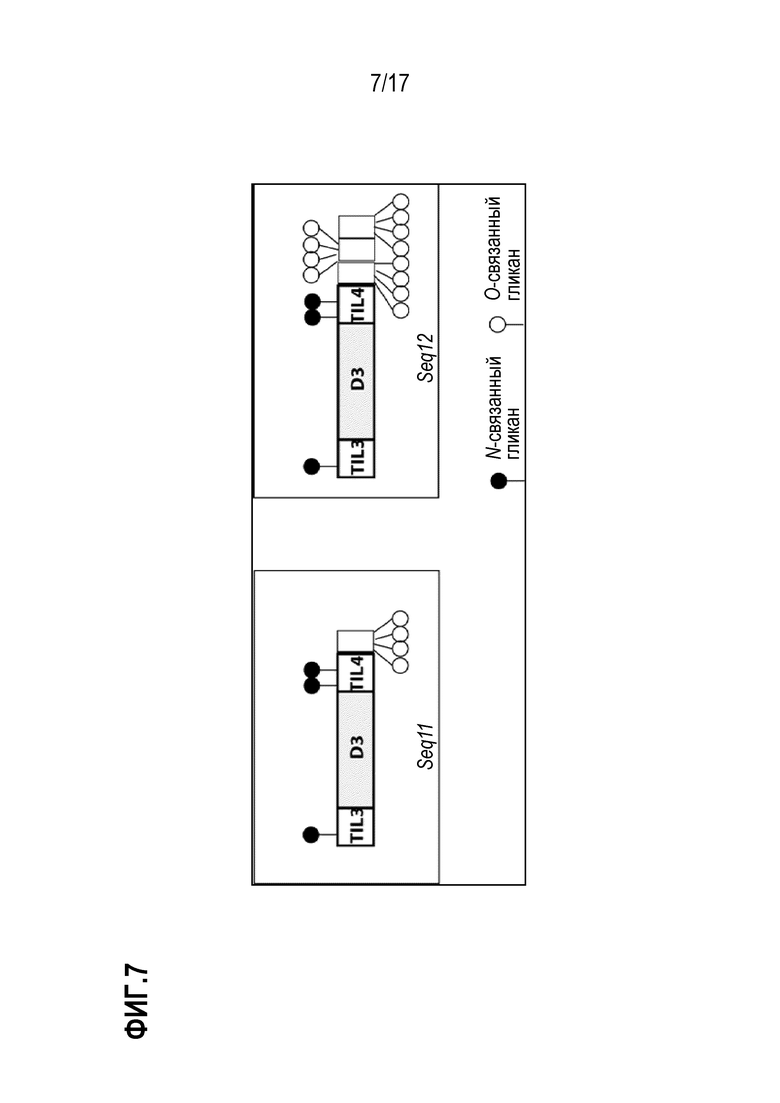
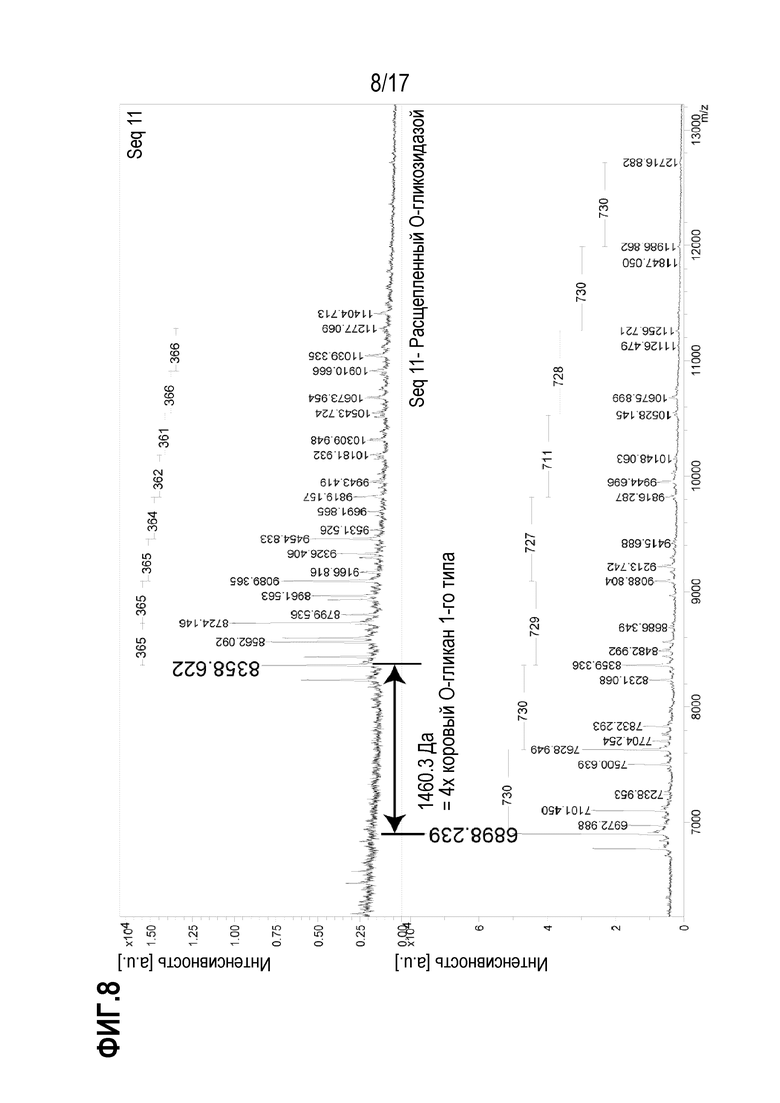
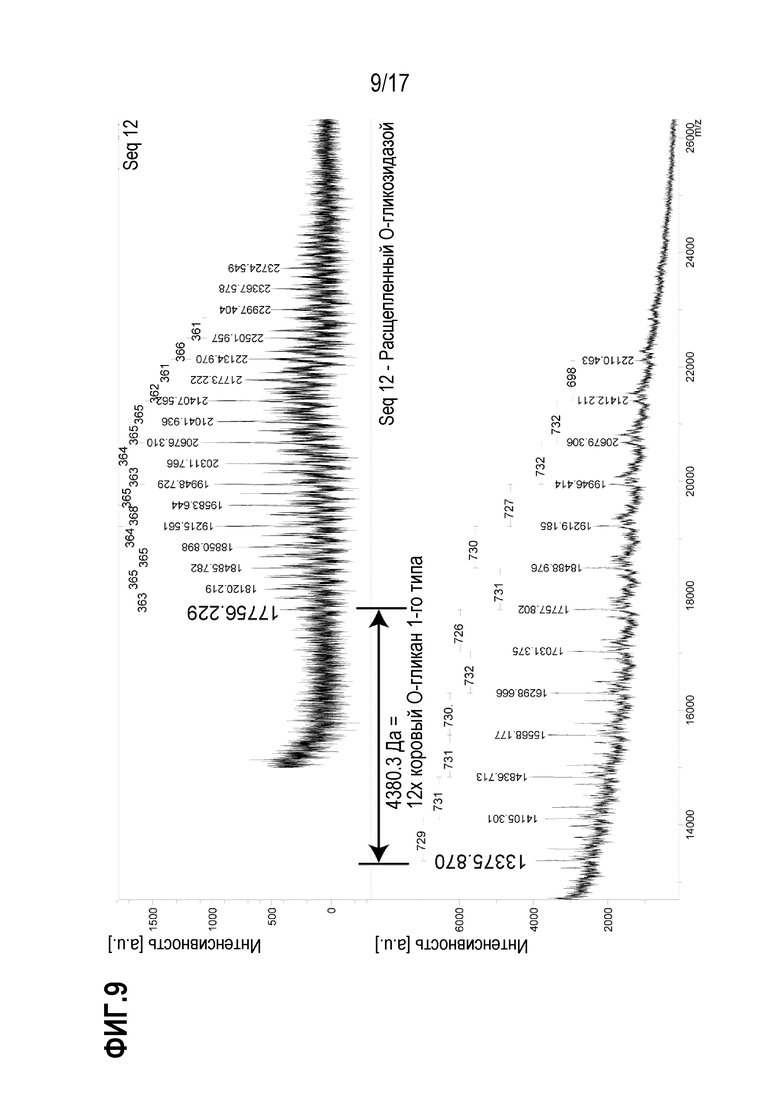
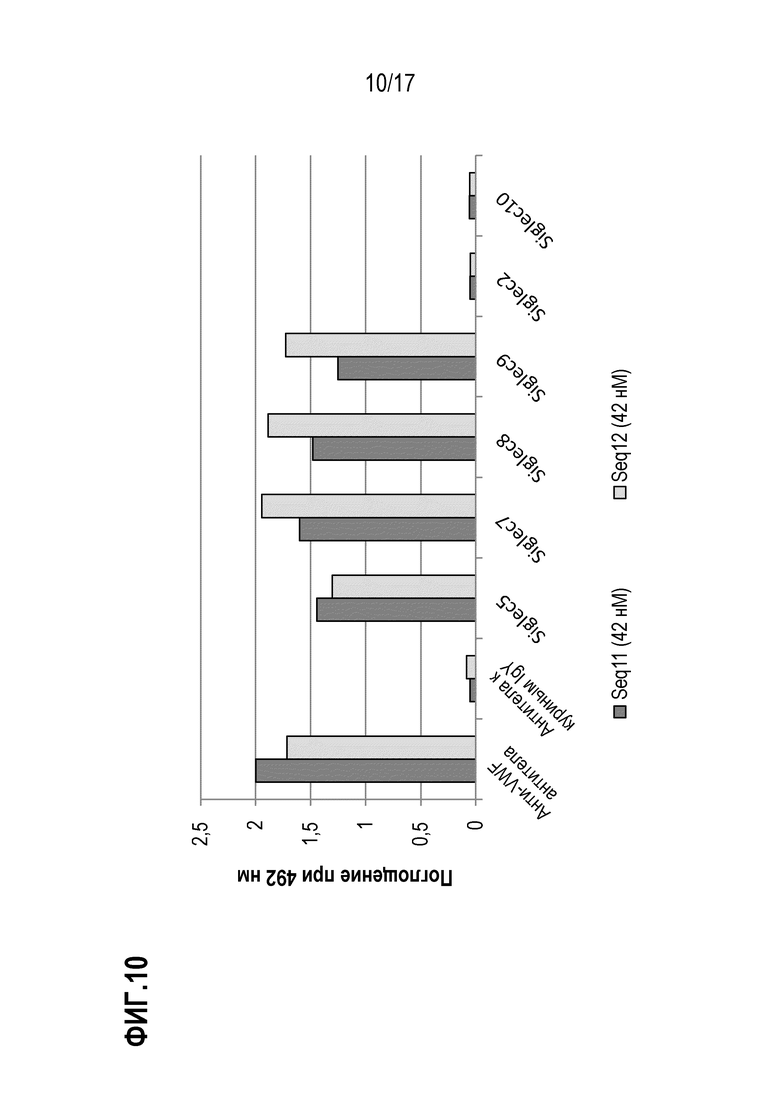
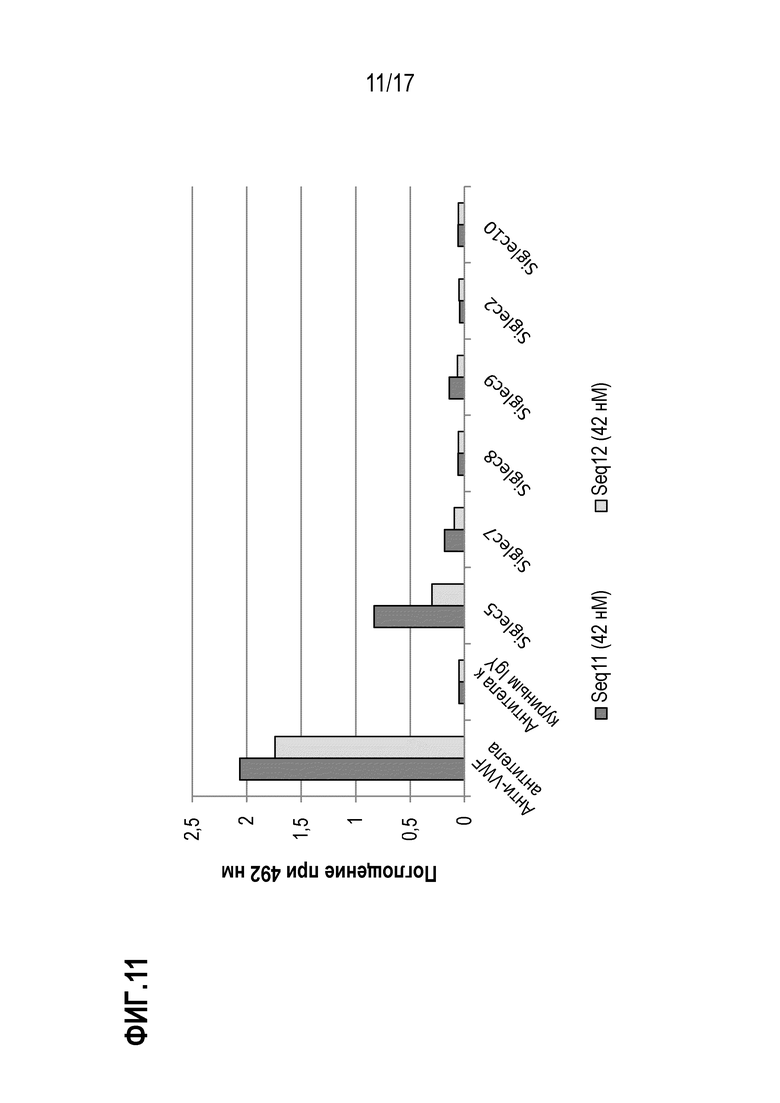
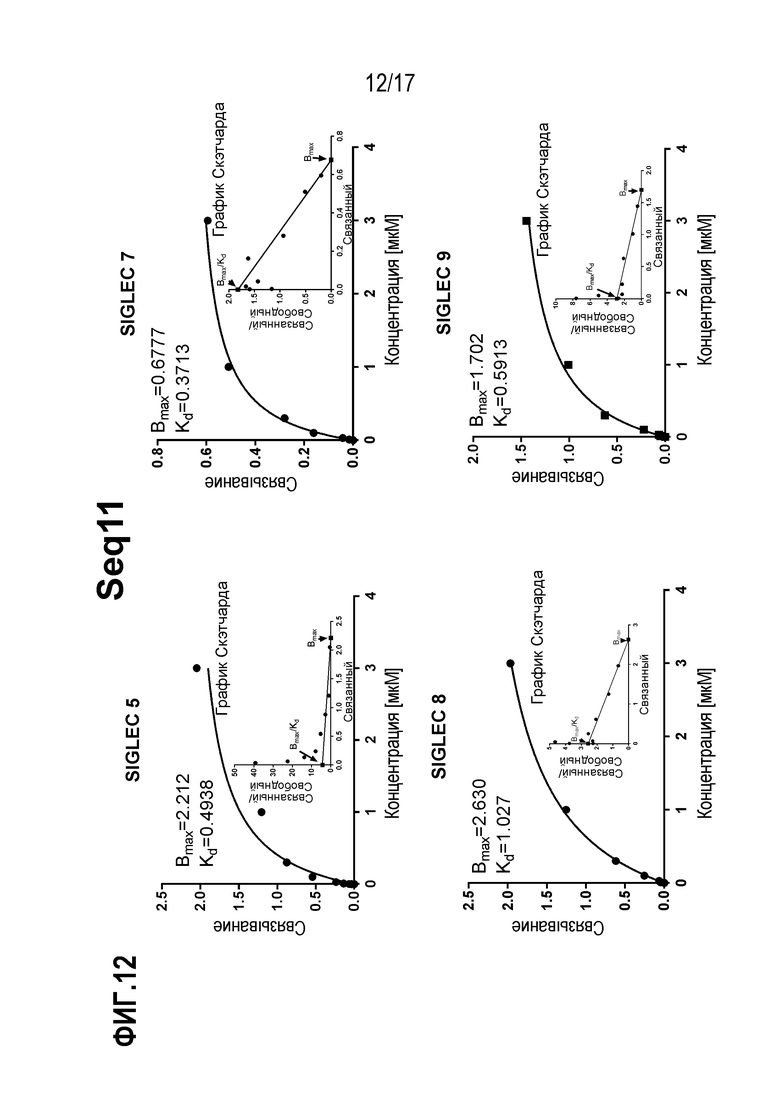
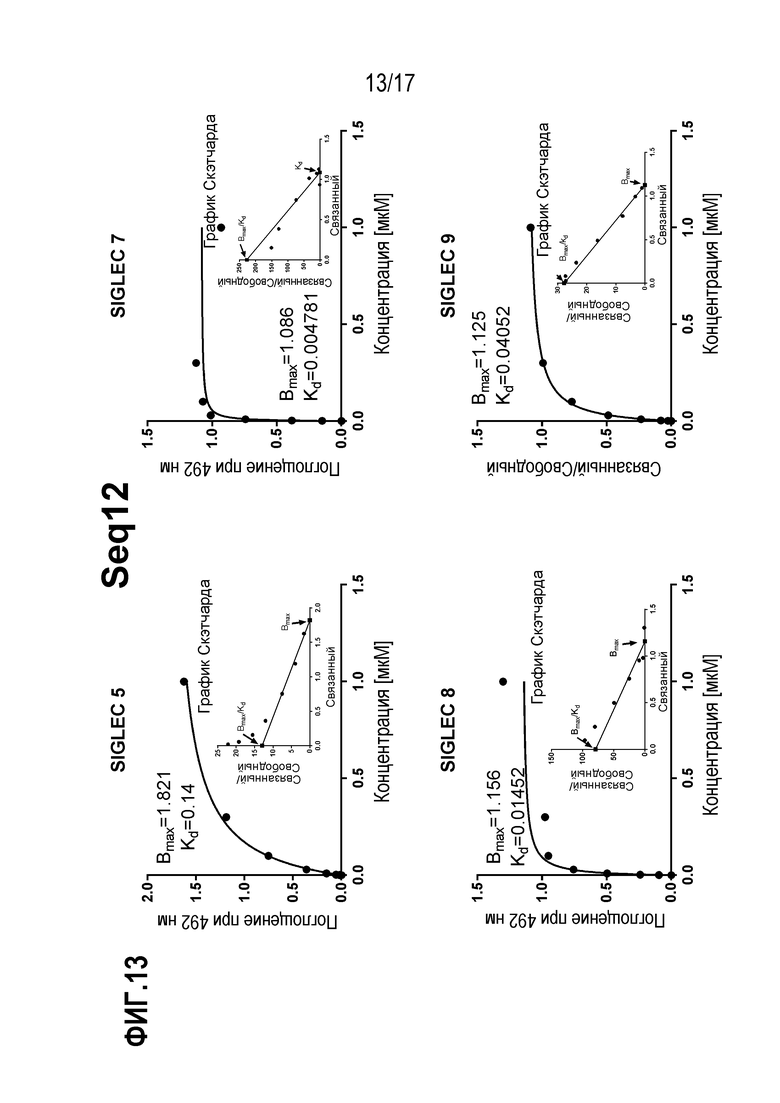
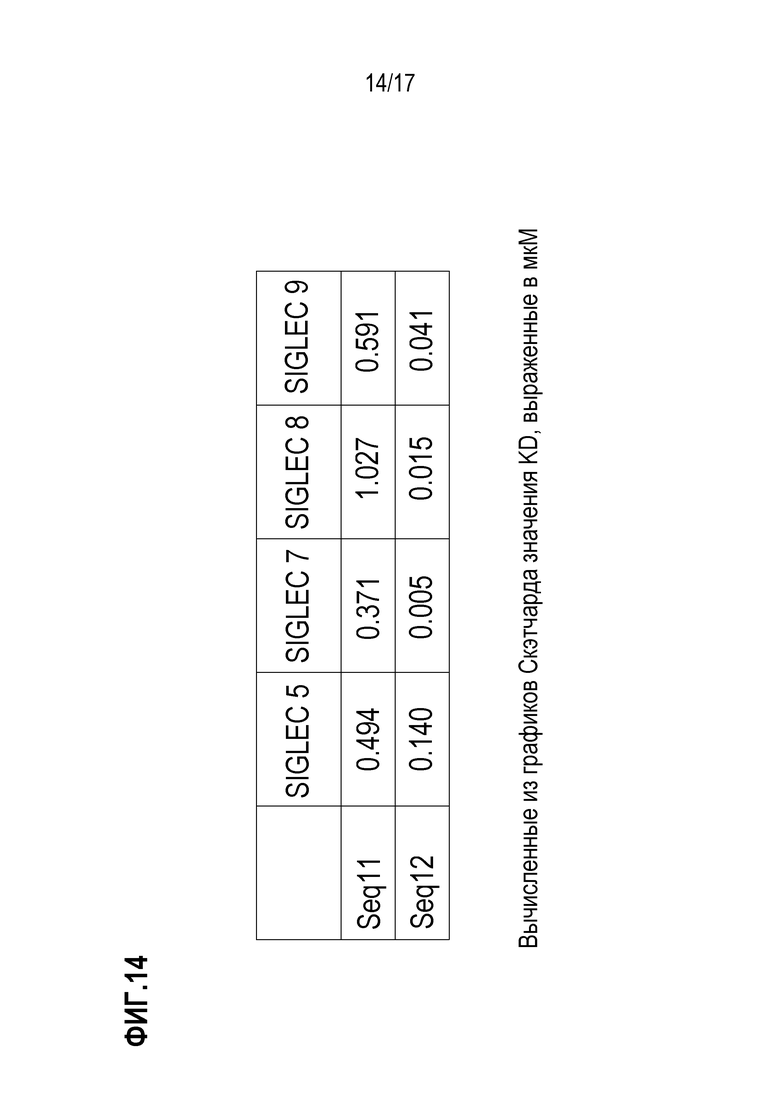

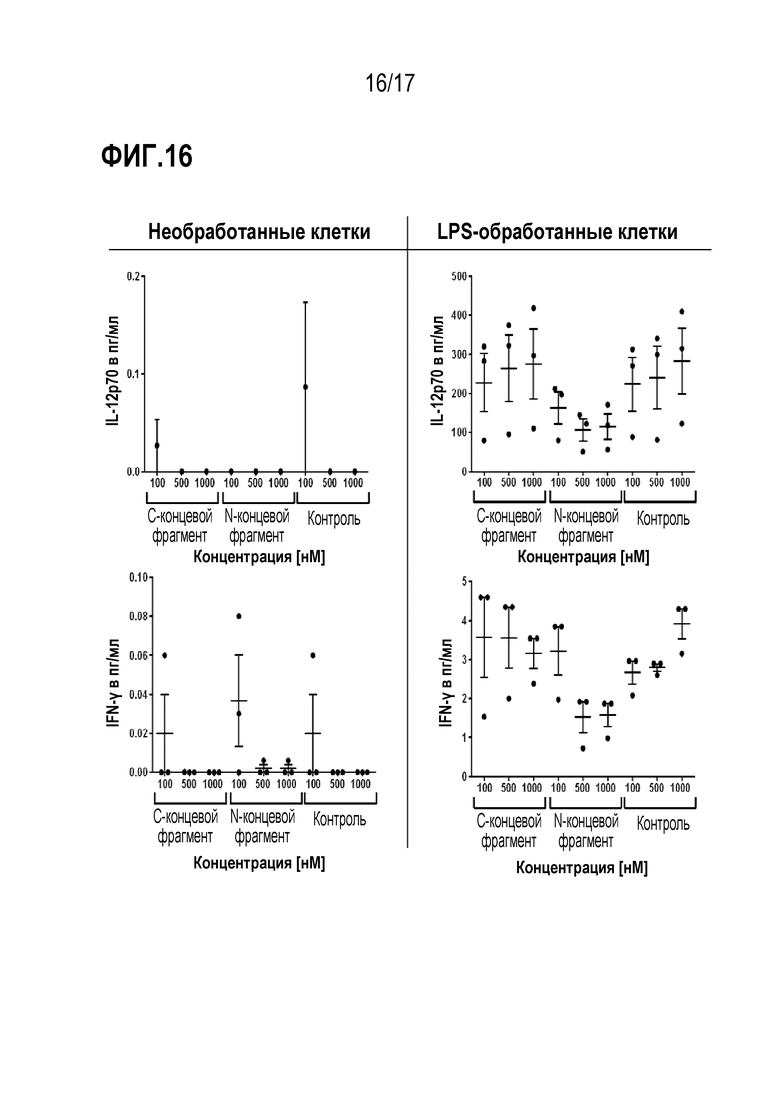

Группа изобретений относится к биотехнологии. Представлены: гликозилированный полипептид, содержащий аминокислотную последовательность, имеющую по меньшей мере 90% идентичность по меньшей мере фрагменту фактора фон Виллебранда человека (vWF), а также его применение для снижения иммунного ответа на второй полипептид, в частности терапевтический белок, композиция для лечения или профилактики нарушения свертываемости крови у пациента, содержащая первый и второй полипептиды в эффективном количестве, выделенный полинуклеотид, кодирующий гликозилированный полипептид, вектор для получения гликозилированного полипептида и клетка-хозяин. Новая молекула, содержащая аминокислотную последовательность, имеющую по меньшей мере 90% идентичность по меньшей мере фрагменту фактора фон Виллебранда человека (vWF), позволяет применять её для эффективного лечения и профилактики нарушения свертываемости крови у пациента. 6 н. и 18 з.п. ф-лы, 17 ил., 13 пр.
1. Гликозилированный полипептид для лечения или профилактики нарушения свертываемости крови у пациента, содержащий аминокислотную последовательность, имеющую по меньшей мере 90% идентичнсть по меньшей мере фрагменту фактора фон Виллебранда человека (vWF), имеющего аминокислотную последовательность SEQ ID NO: 1, где указанный гликозилированный полипептид содержит один или несколько кластеров сайтов O-гликозилирования, где указанные кластеры содержат по меньшей мере два сайта O-гликозилирования и один или несколько сиалилированных O-гликанов, где объединенное количество сиалилированных коровых O-гликанов 2-го типа и сиалилированных удлиненных коровых O-гликанов 1-го типа гликозилированного полипептида превышает количество сиалилированных коровых O-гликанов 2-го типа и сиалилированных удлиненных коровых O-гликанов 1-го типа vWF человека или его фрагмента, и где гликозилированный полипептид имеет повышенную аффинность связывания с одним или несколькими SIGLEC, выбранными из SIG-5, SIG-7, SIG-8 и SIG-9, по сравнению с природным vWF человека.
2. Гликозилированный полипептид по п. 1, причем гликозилированный полипептид имеет повышенное связывание с SIGLEC SIG-5, SIG-7, SIG-8 и SIG-9.
3. Гликозилированный полипептид по любому из предшествующих пунктов, где полипептид содержит по меньшей мере три сайта O-гликозилирования и наиболее предпочтительно по меньшей мере четыре сайта O-гликозилирования.
4. Гликозилированный полипептид по п. 1, где один или несколько кластеров содержат по меньшей мере один сайт О-гликозилирования в десяти аминокислотах, предпочтительно по меньшей мере один сайт О-гликозилирования в четырех аминокислотах, более предпочтительно по меньшей мере один сайт О-гликозилирования в трех аминокислотах и наиболее предпочтительно один сайт О-гликозилирования в двух аминокислотах.
5. Гликозилированный полипептид по любому из пп. 1-4, содержащий по меньшей мере два кластера, более предпочтительно по меньшей мере три кластера.
6. Гликозилированный полипептид по любому из пп. 1-5, где кластеры разделены менее чем 100 аминокислотами, предпочтительно менее чем 50 аминокислотами, более предпочтительно менее чем 30 аминокислотами.
7. Гликозилированный полипептид по любому из предшествующих пунктов, где сиалилированные О-гликаны содержат по меньшей мере две сиаловые кислоты в α2-3 и/или в α2-8 гликозидной связи, более предпочтительно по меньшей мере три сиаловые кислоты.
8. Гликозилированный полипептид по п. 7, где процентное содержание коровых О-гликанов 2 типа от количества сиалилированных О-гликанов составляет по меньшей мере 5%, более предпочтительно по меньшей мере 10%, более предпочтительно по меньшей мере 35%, наиболее предпочтительно по меньшей мере 50%.
9. Гликозилированный белок по любому из предшествующих пунктов, содержащий домен мультимеризации.
10. Гликозилированный полипептид по любому из пп. 1-9, где один или несколько сайтов O-гликозилирования расположены в аминокислотной последовательности VWF человека, в частности, заменяя аминокислоты VWF человека.
11. Гликозилированный полипептид по любому из пп. 1-10, где гликозилированный полипептид представляет собой слитый белок, где вторая аминокислотная последовательность, содержащая один или несколько сайтов O-гликозилирования, ковалентно связана с аминокислотной последовательностью VWF человека.
12. Гликозилированный полипептид по п. 11, где ковалентный линкер выбирают из пептидной связи, химического линкера или гликозидной связи.
13. Гликозилированный полипептид по п. 11 или 12, где вторая аминокислотная последовательность по меньшей мере на 95% гомологична и предпочтительно идентична аминокислотам 475-505 из SEQ ID NO: 1 или двум последовательным копиям аминокислот 475-505 из SEQ ID NO: 1.
14. Гликозилированный полипептид по любому из пп. 1-13, предпочтительно продуцируемый путем экспрессии в линии клеток человека.
15. Применение гликозилированного полипептида по любому из пп. 1-14, содержащего один или несколько сиалилированных O-гликанов и связывающегося с одним или несколькими SIGLEC, выбранными из SIG-5, SIG-7, SIG-8 и SIG-9, для снижения иммунного ответа на второй полипептид, в частности терапевтический белок.
16. Композиция для лечения или профилактики нарушения свертываемости крови у пациента, содержащая первый и второй полипептиды в эффективном количестве, где первый полипептид представляет собой гликозилированный полипептид по любому из пп. 1-14, а второй полипептид содержит аминокислотную последовательность, гомологичную или идентичную второму белку млекопитающего, в частности человека, где первый и второй полипептиды формируют белковый комплекс, где по сравнению со вторым полипептидом композиция имеет повышенную аффинность связывания с одним или несколькими SIGLEC, выбранными из SIG-5, SIG-7, SIG-8 и SIG-9.
17. Композиция по п. 16, где второй полипептид выбран из белка FVIII, FVII, FIX, ADAMTS13, в частности полноразмерного белка FVIII, белка FVIII с удаленным B-доменом или белка FVIII, в котором часть B-домена была заменена линкером.
18. Композиция по п. 16 или 17, где первый полипептид содержит по меньшей мере фрагмент vWF человека, в частности фрагмент vWF, по меньшей мере на 95% идентичный последовательности, ограниченной аминокислотами 1-505 из SEQ ID NO: 1, по меньшей мере на 95% идентичный SEQ ID NO: 2, по меньшей мере на 95% идентичный SEQ ID NO: 3, по меньшей мере на 95% идентичный SEQ ID NO: 4 или по меньшей мере на 95% идентичный SEQ ID NO: 5, и второй полипептид представляет собой белок FVIII.
19. Выделенный полинуклеотид, кодирующий гликозилированный полипептид по любому из пп. 1-14.
20. Выделенный полинуклеотид по п. 19, где гликозилированный полипептид имеет аминокислотную последовательность с идентичностью по меньшей мере на 90%, предпочтительно по меньшей мере на 95%, более предпочтительно по меньшей мере на 98%, наиболее предпочтительно на 100% с последовательностью, выбранной из SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 4, SEQ ID NO: 5.
21. Вектор для получения гликозилированного полипептида по пп. 1-14, где вектор содержит векторный каркас и полинуклеотид по п. 19 или 20, где векторный каркас предпочтительно выбирают из pCDNA3, pCDNA3.1, pCDNA4, pCDNA5, pCDNA6, pCEP4, pCEP-puro, pCET1019, pCMV, pEF1, pEF4, pEF5, pEF6, pExchange, pEXPR, pIRES и pSCAS.
22. Клетка-хозяин для получения гликозилированного полипептида по пп. 1-14, где клетка-хозяин содержит полинуклеотид по п. 19 или 20 или вектор по п. 21, где хозяин предпочтительно представляет собой ранее установленную линию клеток млекопитающего, предпочтительно линию клеток человека, более предпочтительно линию клеток почки человека, наиболее предпочтительно линию клеток почки эмбриона человека, в частности клеточную линию HEK293, такую как HEK293F.
23. Гликозилированный полипептид по любому из пп. 1-14, или композиция по любому из пп. 16-18 для применения в лечении или профилактике нарушения свертываемости крови, предпочтительно выбранного из лечения ранее не подвергавшихся лечению пациентов (PUP) или индукции иммунотолерантности (ITI).
24. Гликозилированный полипептид или композиция для применения по п. 23, где применение включает внутривенную или невнутривенную инъекцию, предпочтительно подкожную инъекцию.
| Кипятильник для воды | 1921 |
|
SU5A1 |
| Бесколесный шариковый ход для железнодорожных вагонов | 1917 |
|
SU97A1 |
| Способ гальванического снятия позолоты с серебряных изделий без заметного изменения их формы | 1923 |
|
SU12A1 |
| Цилиндрический прибор для взятия проб жидкостей | 1923 |
|
SU1855A1 |
| S0LECKA В A ET AL.: "Site-specific analysis of von Willebrand factor O-glycosylation.", JOURNAL OF THROMBOSIS AND HAEMOSTASIS, vol | |||
| Паровоз для отопления неспекающейся каменноугольной мелочью | 1916 |
|
SU14A1 |
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |

