Изобретение относится к молекулярной биологии и может найти применение в медицине и молекулярной биологии для анализа экспрессии генов, диагностики и выявления механизмов патологий на генетическом уровне.
Дифференциальная экспрессия генов, реализуемая путем синтеза различных наборов матричных РНК (мРНК), лежит в основе разнообразия фенотипов и функций клеток живых организмов. В этой связи понятна важность методов, которые бы позволяли идентифицировать и изучать дифференциально экспрессирующиеся мРНК, т. е. такие мРНК, содержание которых в пуле клеточной мРНК различается в двух или более типах клеток или изменяется в зависимости от функционального состояния клеток.
В настоящее время описан ряд методов детекции и клонирования дифференциально экспрессирующихся мРНК, которые в большинстве случаев сводятся к применению метода гибридизации нуклеиновых кислот.
В методе дифференциального скрининга [1] путем переноса на нитроцеллюлозные или нейлоновые фильтры реплики высеянных на чашки библиотек комплементарной ДНК (кДНК) из популяции клеток А гибридизуют поочередно с мечеными препаратами кДНК из популяции клеток А, Б, В и так далее. Поскольку гибридизационный сигнал, получаемый от отдельных клонов, теоретически должен быть пропорционален представленности соответствующей клонированной последовательности в препарате кДНК, можно по уровню сигнала, получаемого от разных препаратов, идентифицировать клоны, соответствующие генам, экспрессирующимся дифференциально в популяциях клеток А, Б и т.д.
Существенным недостатком данного подхода является низкая чувствительность, поскольку детектировать таким образом удается лишь высокопредставленные (как правило, составляющие не менее 0,1% мРНК) последовательности. Для более редких последовательностей гибридизационный сигнал не превышает фона. Кроме того, в данном подходе удается просмотреть небольшое число (от нескольких тысяч до нескольких десятков тысяч) клонов.
Еще один недостаток состоит в том, что информация об индивидуальности клона (описание его местоположения на чашке или реплике), теряется по окончании работы с данными репликами и не может быть использована в последующих экспериментах. Более того, библиотеки кДНК из разных типов клеток не сравнимы в том плане, что одинаковые последовательности занимают разные места на фильтрах в разных библиотеках.
Таким образом, переход от одних типов клеток к другим практически невозможен. В последнее время делаются попытки создать упорядоченные библиотеки кДНК, содержащие отдельные клоны, распределенные в ячейках 96-луночных планшетов. При этом, однако, не устраняется главный недостаток - низкая чувствительность. Кроме того, наборы клонов, полученные из одного типа клеток, по-прежнему не сравнимы с наборами клонов из другого типа клеток.
Для того чтобы преодолеть низкую чувствительность метода дифференциального скрининга, применяется метод вычитающей гибридизации [1], при котором препарат кДНК из клеток А гибридизуется с препаратом кДНК или мРНК из клеток Б, после чего образовавшиеся гибриды удаляют тем или иным способом. Полученный препарат обогащен последовательностями, специфичными для препарата А. Такой "вычтенный" препарат можно использовать как для создания библиотек, так и для дифференциального скрининга, что позволяет как увеличить вероятность нахождения дифференциально экспрессирующихся последовательностей, так и повысить чувствительность детекции слабо представленных последовательностей.
Применение такой процедуры, однако, делает подход длительным и трудоемким, кроме того, выигрыш в чувствительности все равно недостаточен для детекции редких последовательностей и сопровождается потерей возможности непосредственно сравнивать уровень экспрессии гена в разных клетках.
В последнее время была предложена модификация метода дифференциального скрининга, при котором вместо клонированных последовательностей для иммобилизации на твердой основе используются большие упорядоченно расположенные наборы олигонуклеотидов, например, всевозможных тетрадекануклеотидов, содержащих общую последовательность ААТААА, встречающуюся в большинстве мРНК [2]. Этот способ теоретически обеспечивает перебор гораздо большего, чем при стандартном дифференциальном скрининге, набора последовательностей мРНК.
Недостатком этого подхода, как и других способов дифференциального скрининга, является тот факт, что в условиях гибридизации с чрезвычайно сложной смесью молекул кДНК, частоты представленности которых могут варьировать в сотни и тысячи раз, вероятность появления ложных сигналов за счет кросс-гибридизации становится ощутимой, что делает весьма проблематичным надежную детекцию и количественный анализ редких последовательностей мРНК.
Дополнительной трудностью является необходимость синтеза и иммобилизации на небольшой площади от нескольких десятков до сотни тысяч олигонуклеотидов, что требует больших начальных инвестиций.
Кроме того, такой подход сам по себе не дает возможность клонирования сколько-нибудь протяженных участков дифференциально экспрессирующихся мРНК, что требует отдельной работы.
Наиболее близким к изобретению является метод, описанный Liang, Рагdee [3] , и получивший название "дифференциальный дисплей мРНК. В данном методе кДНК, синтезированная на матрице мРНК, используется для создания набора фрагментов кДНК дискретной длины с помощью полимеразной цепной реакции (ПЦР), при этом для синтеза первой цепи кДНК используются в качестве затравки олиго(дТ) праймеры, содержащие на 3' конце два дополнительных основания. Поскольку существует 12 вариантов таких праймеров, возможен синтез двенадцати различных вариантов первых цепей кДНК. Затем на матрице одного из вариантов первой цепи происходит амплификация с помощью ПЦР с использованием соответствующего олиго(дТ) праймера и праймера произвольной последовательности. При этом начальные циклы амплификации проводятся при относительно невысокой температуре, т. е. в нестрогих условиях, способствующих затравливанию и амплификации не одной, как это обычно предполагается в стандартной ПЦР, а целой группы последовательностей кДНК. Амплифицируемые участки прилежат к 3' концу (поли(А) хвосту) мРНК. В ходе реакции ПЦР происходит также включение радиоактивной метки в образовавшиеся фрагменты. Набор дискретных фрагментов кДНК разделяют электрофорезом в акриламидном геле и сравнивают с набором фрагментов, полученным в идентичных условиях из другого типа клеток. Фрагменты с дифференциальной интенсивностью можно элюировать из геля, реамплифицировать и клонировать в подходящий вектор. Используя комбинации двенадцати олиго(дТ) праймеров и различных произвольных праймеров, можно, по крайней мере теоретически, большинству видов мРНК сопоставить один или несколько дискретных фрагментов кДНК. Кроме того, существуют также незначительные модификации этого подхода [4 и 5].
Существенным недостатком подхода является то, что в одной реакции амплифицируется не более нескольких десятков последовательностей. Таким образом, для перебора 10-15 тыс. последовательностей РНК, экспрессирующихся в отдельных типах клеток или тканях, потребуется проведение как минимум нескольких сотен (вероятно, до нескольких тысяч) реакций. Кроме того, выборка последовательностей, амплифицирующихся в одной реакции, случайна, поэтому при переборе значительной части последовательностей клеточной мРНК будет происходить весьма существенное накопление избыточной информации. Так, после просмотра 50% всех последовательностей, в каждой следующей реакции только 1/2 амплифицируемых последовательностей будет являться новой; при глубине просмотра 90% доля новых последовательностей снизится до 1/10.
Вероятно, наиболее значительным недостатком подхода является его низкая воспроизводимость, связанная с тем, что при слабо селективных условиях затравливания небольшие колебания в исходных условиях, в качестве препаратов РНК, праймеров и других параметрах вызывают заметные количественные изменения в спектре амплифицируемых фрагментов кДНК. Так, при оптимальной температуре отжига 42oC смещение температуры отжига вниз или вверх на 2oC приводило к появлению значительного фона (т.е. дополнительных полос), или к исчезновению некоторых полос соответственно [5]. Во-вторых, по данным самих авторов, при проведении параллельных экспериментов воспроизводимыми оказываются около 95% полос [4]. Таким образом, уровень невоспроизводимости (5%) сопоставим или даже превышает уровень различий между популяциями мРНК.
По данным авторов при применении данного подхода количество воспроизводимых дифференциальных полос обычно меньше количества невоспроизводимых полос. Вероятно, чувствительность подхода к качеству препаратов РНК, праймеров, температуре и времени отжига и другим параметрам может в значительной степени препятствовать сравнению получаемых результатов с уже известными данными.
Поскольку для исчерпывающего анализа популяций мРНК требуется проводить сотни или тысячи отдельных реакций амплификации, количество требующегося клеточного материала в таком случае становится значительным. Таким образом, использование метода при наличии только небольшого количества клеток (сортированные с помощью проточной цитофлюорометрии клетки, например) либо невозможно, либо ограничено весьма небольшой глубиной просмотра, что является дополнительным недостатком подхода. Наконец, картины разделения фрагментов в геле после проведения амплификации демонстрируют значительный фон. Полосы, интенсивность которых равна или слабее фона, неразличимы, поэтому чувствительность метода может быть понижена за счет такого эффекта.
Задача - обеспечение возможности прямого качественного и количественного сравнения спектров мРНК, синтезируемых в разных типах клеток одного организма, возможности детекции дифференциально экспрессируемых РНК, в том числе и с низким уровнем представленности, возможности соотнесения таких РНК с уже известными, а также возможности клонирования фрагментов таких РНК.
Задача решается тем, что набор матричных РНК, экспрессирующихся в клетке путем синтеза комплементарной ДНК (кДНК) с последующей фрагментацией кДНК частощепящими рестрикционными эндонуклеазами (имеющими четырех- или пятибуквенные сайты узнавания) и использованием только фрагментов кДНК, соответствующим в зависимости от конкретного варианта, 3' или 5' концам мРНК, представляется в виде набора фрагментов кДНК дискретной длины (не более одного фрагмента на каждый вид мРНК) таким образом, что по крайней мере один из концов фрагментов несет метку или необходимую для детекции группу. Генерируемые фрагменты кДНК, происходящие из 3'-концевых (прилежащих к поли(А) хвосту) или 5'-концевых областей мРНК, обеспечивают представление (в зависимости от конкретной схемы осуществления изобретения) от 90 до практически 100% последовательностей мРНК. Поскольку получаемая в итоге популяция фрагментов содержит от десяти до двадцати-тридцати тысяч различных фрагментов кДНК, что делает затруднительной детекцию редких мРНК, существенной чертой подхода является разбиение набора фрагментов кДНК на более простые непересекающиеся поднаборы фрагментов, легче поддающиеся анализу. Для увеличения чувствительности метода концы фрагментов, образованные расщеплением рестрикционной эндонуклеазой, лигируют с олигонуклеотидным адаптором, что позволяет проводить амплификацию всего набора фрагментов кДНК с помощью ПЦР. Наборы или поднаборы фрагментов разделяются одно- или двумерным гель-электрофорезом и производится детекция метки и сравнение картин разделения, при этом интенсивность сигнала от данного фрагмента кДНК варьирует для препаратов мРНК из разных клеток пропорционально представленности соответствующей ему последовательности в пуле мРНК. При необходимости на стадии детекции путем переноса разделенных фрагментов на мембрану и последовательной гибридизации с набором олигонуклеотидов, частично перекрывающих общую концевую последовательность фрагментов, возможен дополнительный анализ разделенной картины. Разделенные фрагменты кДНК после дополнительных операций могут быть амплифицированы и исследованы путем проведения рестрикционного и гибридизационного анализа, а также секвенирования, и клонированы в плазмидный или фаговый вектор.
Таким образом, в предлагаемом изобретении, в отличие от метода дифференциального дисплея мРНК, одна из границ фрагментов задается положением рестрикционного сайта. Кроме того, в предлагаемом подходе сначала создается один набор фрагментов кДНК, представляющий подавляющее большинство последовательностей мРНК, который затем разбивается на более простые непересекающиеся поднаборы. Этим предлагаемый подход принципиально отличается от метода "дифференциального дисплея мРНК", в котором в ходе отдельных реакций ПЦР создаются простые наборы фрагментов кДНК, которые только в своей совокупности могут более или менее полно представлять популяцию последовательностей мРНК. Использование реакции расщепления ДНК рестрикционными нуклеазами, характеризующейся высокой специфичностью, полнотой расщепления и малой чувствительностью к колебаниям температурных условий, позволяет сделать подход заметно более воспроизводимым, и, следовательно, позволит надежное сравнение с проведенными независимо экспериментами. Такие характеристики вместе с высокой разрешающей способностью метода позволят приступить к созданию баз данных гелевых координат экспрессируемых последовательностей. Предлагаемый метод позволяет устранить избыточность информации, характерную для метода с произвольными праймерами. Кроме того, наличие на начальных стадиях одной общей популяции фрагментов кДНК позволяет проводить такие операции, как выравнивание частот путем самогибридизации популяции фрагментов кДНК [6], что позволит значительно снизить уровень представленности наиболее часто встречающихся видов мРНК и поможет выявлению очень редких последовательностей мРНК за счет уменьшения интерференции от часто встречающихся последовательностей.
Изобретение поясняется фиг. 1 - 6, на которых:
фиг. 1 - схема варианта осуществления изобретения путем образования набора 3'-концевых меченых фрагментов кДНК, разбиения его на поднаборы фрагментов с помощью иммобилизации на твердом носителе и последовательной обработки серией рестрикционных нуклеаз, и разделения полученных поднаборов электрофорезом;
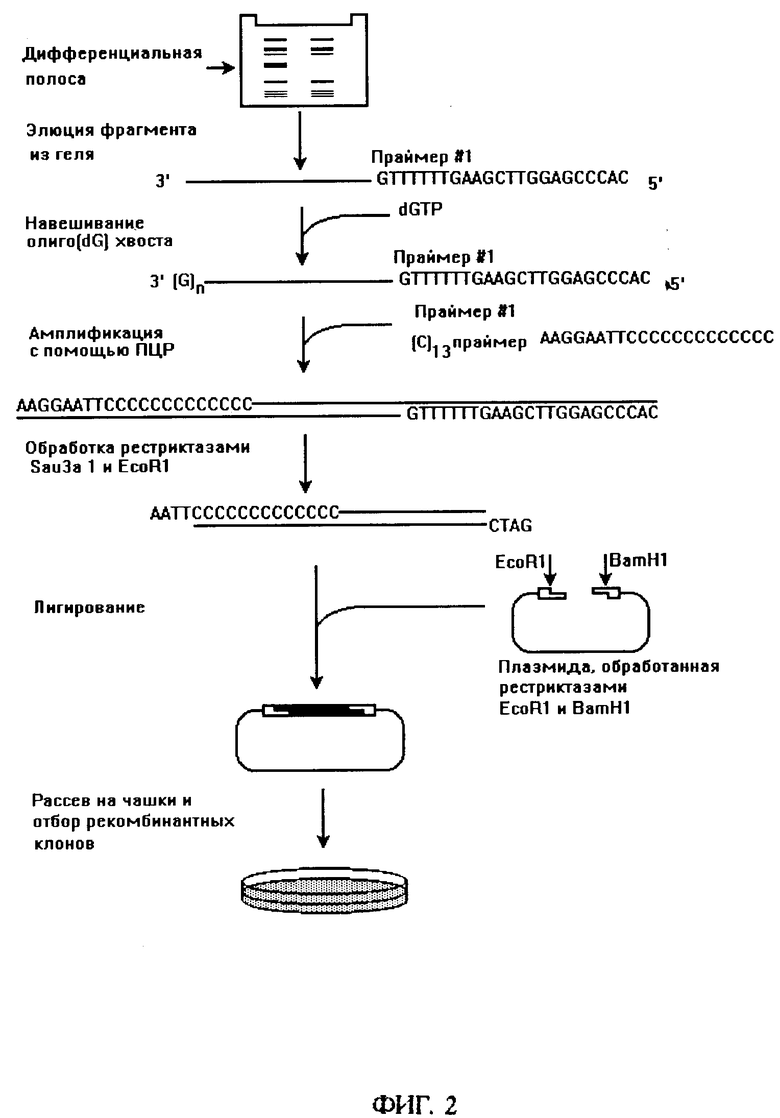
фиг. 2 - схема амплификации и клонирования фрагментов кДНК, полученных и разделенных в геле с помощью способа, изображенного на фиг. 1;
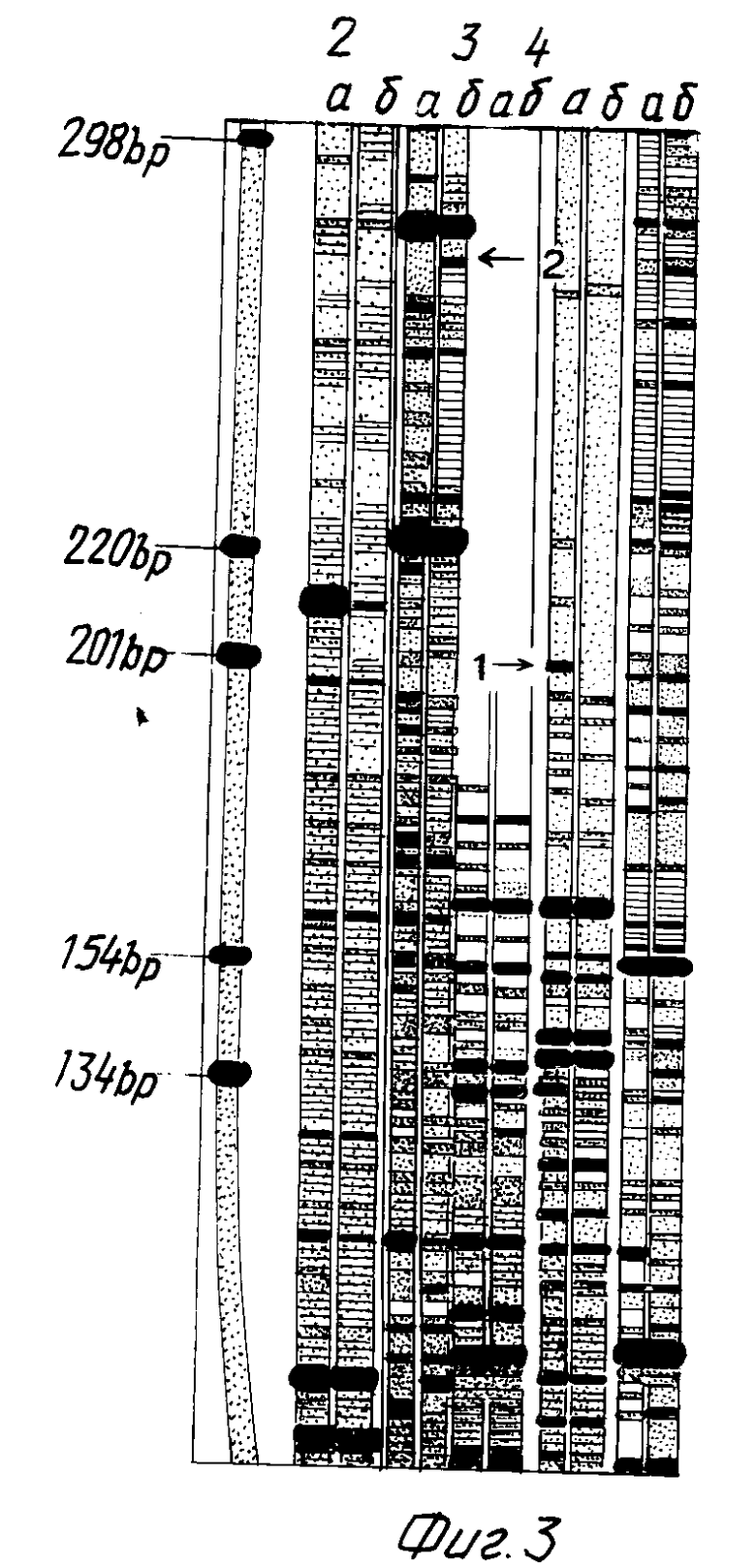
фиг. 3 - картина разделения в денатурирующем полиакриламидном гель-электрофорезе 32P-меченых фрагментов кДНК из препаратов РНК тимуса (а) и селезенки мышей (б), полученных с помощью способа, изображенного на фиг. 1 (первичное расщепление проводили ферментами Sau3A(2-5) и BamHI (6), вторичное - ферментами EcoRV (2), PstI (3,6), MspI (4) и Hin PI (5); слева (1) нанесены маркерные фрагменты ДНК и указаны их длины в нуклеотидах);
фиг. 4 - гибридизация клонированных фрагмента 1 (а) и фрагмента 2 (б) с разделенными электрофорезом и перенесенными на мембрану препаратами поли(А) РНК из селезенки (1) и тимуса (2) мышей (картина гибридизации подтверждает специфичность экспрессии, следующую из фиг. 3; справа (3) нанесены маркерные одноцепочечные фрагменты ДНК и указаны их длины в нуклеотидах);
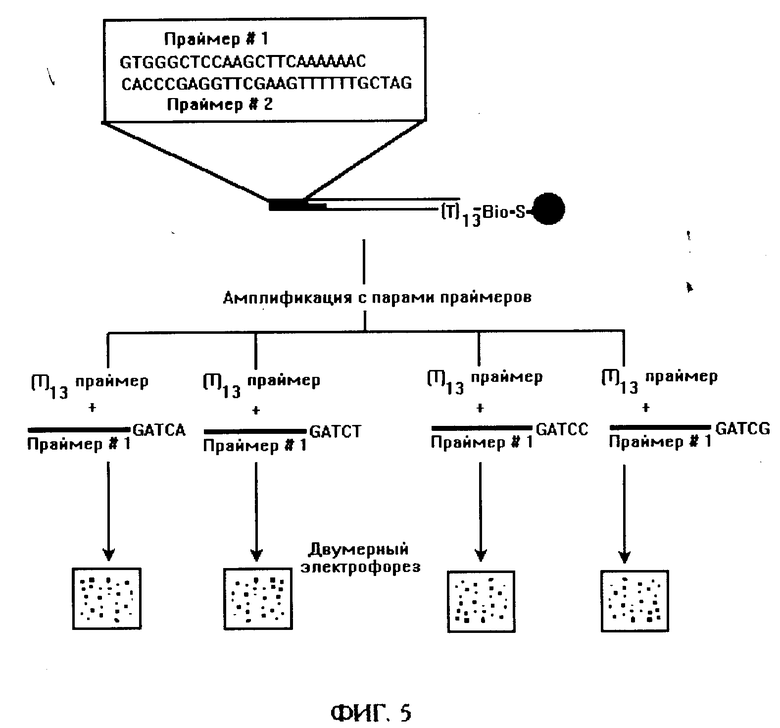
фиг. 5 - альтернативный способ разбиения набора З'-концевых фрагментов кДНК, полученных по способу, изображенному на фиг. 1, на 4 поднабора путем амплификации набора фрагментов полимеразной цепной реакцией с адапторными праймерами, содержащими добавочное основание на 3' концеза пределами общей для всех фрагментов части;
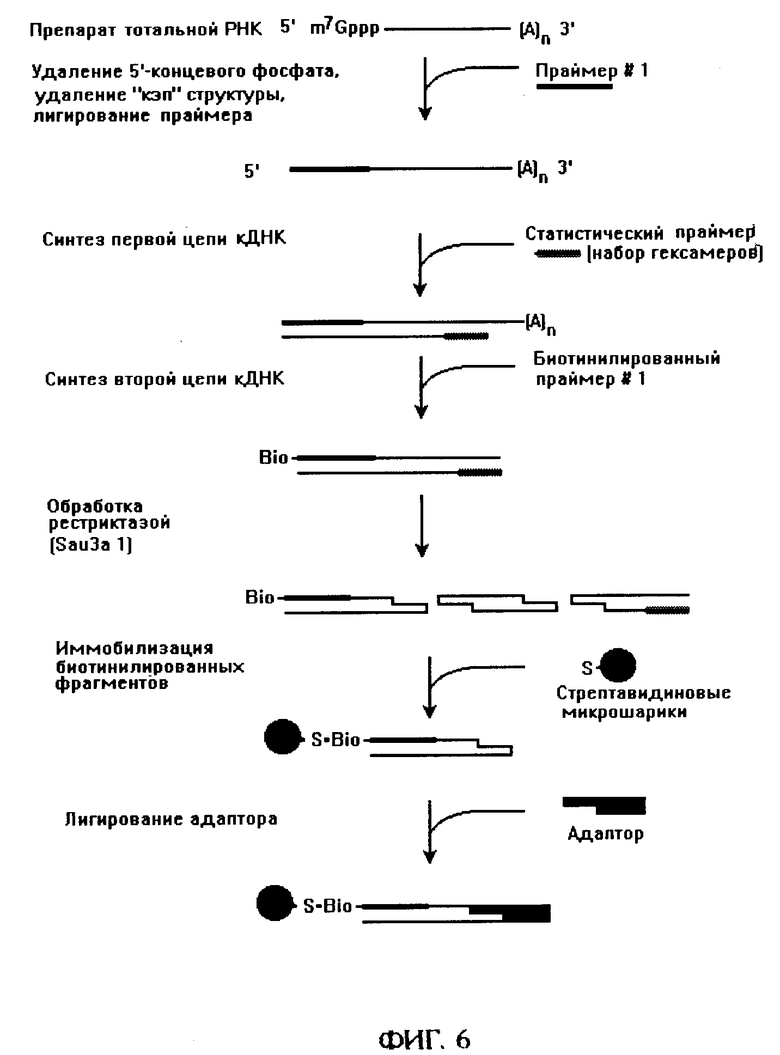
фиг. 6 - способ создания набора 5'-концевых фрагментов кДНК.
Ниже более подробно излагается один из вариантов реализации заявленного способа.
Синтез первой цепи кДНК осуществляют энзиматическим путем с помощью обратной транскриптазы и олиго(дТ) праймера, иммобилизованного на микрогранулированном твердом носителе, или олиго(дТ)-содержащего праймера, несущего на 5'конце биотиновую группу. Для синтеза второй цепи кДНК проводится предварительно навешивание олиго(дГ) хвоста на 3'конец первой цепи кДНК с помощью терминальной трансферазы. Синтез второй цепи проводится модифицированной ДНК-полимеразой фага T7 Sequenase (United States Biochemicals, США) с использованием олиго(дЦ)-содержащего праймера в качестве затравки. Проводится полный гидролиз кДНК рестрикционной эндонуклеазой, имеющей четырехбуквенной сайт узнавания (например, Sau3A), отмывка высвобожденных продуктов расщепления и лигирование адаптора, комплементарного сайту расщепления. В случае использования биотинилированного праймера для синтеза первой цепи, кДНК до или после расщепления рестриктазой связывают со стрептавидин-содержащим твердым носителем (Streptavidin MagneSphere, Promega, США). В этой операции используется свойство стрептавидина быстро и прочно связывать биотиновую группу. Для увеличения количества материала производится амплификация фрагментов кДНК путем ПЦР с использованием праймера, входящего в состав адаптора, и модифицированного олиго(дТ) праймера, содержащего биотиновую группу на 5'-конце. После иммобилизации фрагментов на содержащем стрептавидин твердом носителе свободные цепи снимаются обработкой 100 mM NaОН, затем на иммобилизованных цепях отжигается праймер, в 3'-конец праймера включается метка путем реакции удлинения цепи ДНК-полимеразой с использованием радиоактивного α-32P дАТФ или α-33P дАТФ, после чего добавляют немеченые дезоксинуклеотидтрифосфаты и достраивают полные цепи кДНК. Последовательность праймера и отжигающегося с ним участка кДНК, а также условия реакции мечения таковы, что область мечения ограничена последовательностью адаптора. После включения метки проводится последовательное исчерпывающее расщепление 8 - 10 рестриктазами, причем вначале используются рестриктазы, имеющие 6-буквенный сайт узнавания, затем 5-буквенный, и наконец 4-буквенный сайт узнавания. Высвобожденные меченые фрагменты кДНК после каждой реакции собирают отдельно. Разделение фрагментов проводят с помощью одной из систем электрофореза в полиакриламидном или родственном геле (Hydrolink, AT Biochem, США). Предпочтительной для наиболее высокоразрешающего разделения является система двумерного электрофореза ДНК, в которой в первом направлении производится разделение двухцепочечной ДНК по длине, а во-втором - по составу (точнее, по профилю плавления) с помощью денатурирующего градиента [7]. По литературным данным, даже после переноса на мембрану, что заметно ухудшает разрешение, подобная система позволяет разделить по меньшей мере 625 фрагментов ДНК на одном геле [8] . По всей видимости, верхний предел разрешения составляет 1 - 1,5 тыс. фрагментов ДНК. При разбиении популяции фрагментов кДНК на 8 - 12 непересекающихся наборов, можно разделить до 10 - 15 тыс. фрагментов, т.е. практически все мРНК, экспрессирующиеся в клетках одного типа. Сигнал от разделенных фрагментов детектируют путем авторадиографии на рентгеновскую пленку. Картины, полученные от препаратов РНК из разных типов клеток, сравнивают между собой, по положению в одно- или двумерном электрофорезе устанавливают соответствие между отдельными фрагментами кДНК из разных клеток и идентифицируют фрагменты, дающие дифференциальный сигнал. Для того чтобы выяснить, какой из уже известных последовательностей может соответствовать то или иное пятно на двумерной электрофореграмме, нужно сначала исходя из клонированных последовательностей мРНК, имеющихся в банке, определить, какие последовательности могут давать фрагменты соответствующей длины, затем, используя алгоритм для предсказания подвижностей фрагментов ДНК в денатурирующем градиенте [9] , затем выбрать из таких последовательностей такую, профиль плавления которой наиболее точно соответствует месту, занимаемому фрагментом во втором направлении.
Для того, чтобы получить нужный фрагмент в количестве, достаточном для проведения анализа и клонирования, фрагмент элюируют из геля, навешивают олиго(дГ) хвост с помощью терминальной трансферазы и затем амплифицируют путем ПЦР с использованием адапторного праймера и олиго(дЦ) праймера (фиг. 2). Идентичность фрагмента известной последовательности может быть проверена путем рестрикционного анализа или путем определения нуклеотидной последовательности непосредственно на амплифицированном фрагменте. Кроме того, амплифицированный фрагмент может быть легко клонирован в плазмидный или фаговый вектор.
Проведенный авторами компьютерный анализ 659 клонированных последовательностей мРНК мыши, имеющих секвенированную прилежащую к поли(А) хвосту область, показал, что при использовании для расщепления кДНК рестрикционной эндонуклеазы Sau3A (сайт узнавания: ГАТЦ) 93% всех мРНК будет расщеплено ферментом и, следовательно будет участвовать в анализе. Доля последовательностей мРНК, участвующих в анализе, может быть доведена до 99 - 99,5%, если проводить расщепление второй рестриктазой, имеющей другой сайт узнавания, тех последовательностей кДНК, которые не были расщеплены первой рестриктазой. Компьютерный анализ показывает также, что вторичное расщепление иммобилизованных фрагментов кДНК набором рестрикционных эндонуклеаз может затронуть подавляющее большинство фрагментов (96% при использовании десяти 4-буквенных рестриктаз). Таким образом, от 90 до 96% всех последовательностей мРНК клетки будет представлено в виде дискретных фрагментов кДНК и будет участвовать в анализе.
Предлагаемый метод обладает высокой чувствительностью. При использовании только 1 мкг фрагментов кДНК для мечения легко получить включение до 10 расп. /мин. Такой уровень мечения позволяет детектировать последовательности мРНК, составляющие от 0,001 до 0,0001% мРНК клетки.
Кроме вышеописанного существуют другие варианты осуществления изобретения. Синтез второй цепи кДНК можно осуществлять другими известными способами, в частности используя в качестве затравок для синтеза частично расщепленную с помощью РНКазы H цепь РНК в гибриде кДНК-РНК, или используя самозатравливание первой цепи кДНК после гидролиза цепи РНК [10].
Наиболее существенной модификацией является использование альтернативного способа разбиения набора 3'-концевых фрагментов кДНК на непересекающиеся поднаборы фрагментов путем амплификации с помощью 12 различных пар праймеров, включающих:
а) 4 варианта модифицированного адапторного праймера, включающие общую для всех фрагментов кДНК последовательность адапторного конца и содержащие каждый одно дополнительное основание на 3'-конце;
б) 3 варианта олиго(дТ)-содержащего праймера, содержащие одно дополнительное основание на 3'-конце.
При этом используют свойство Taq ДНК-полимеразы удлинять только те праймеры, которые содержат полностью спаренное 3'-концевое основание [11]. В таком варианте может не требоваться проведение второго расщепления набором рестриктаз, поэтому доля последовательностей мРНК, участвующих в анализе, может возрасти практически до 100%.
Еще одной важнейшей разновидностью подхода является создание набора фрагментов кДНК, соответствующих 5'-концу мРНК (фиг. 6). Для этого применяют специфическое мечение 5'-конца мРНК олигонуклеотидным праймером (праймер 1) согласно процедуре [12]. После этого синтезируют первую цепь с помощью набора случайных гексамерных праймеров, синтезируют вторую цепь с помощью содержащего на 5'-конце биотиновую группу праймера 1, иммобилизуют кДНК на стрептавидиновых микрогранулах и далее проводят гидролиз кДНК рестриктазой, лигирование адаптора и прочие процедуры, как указано выше. Разбиение набора 5'-концевых фрагментов кДНК на поднаборы осуществляют либо путем иммобилизации меченых фрагментов и последовательной обработкой рестриктазами, либо с помощью раздельных реакций амплификации с концевыми праймерами, содержащими добавочное основание на 3'-конце, как указано выше. Кроме двумерного электрофореза с использованием денатурирующего градиента во втором направлении, возможны также системы двумерного электрофореза, использующие во втором направлении разделение одноцепочечной ДНК в неденатурирующих условиях за счет ее конформационного полиморфизма [13], или расщепление ДНК после первого направления частощепящими рестриктазами [14] и разделение во втором направлении по длине.
Кроме использования для детекции радиоактивной 32P или 32P метки с последующей радиоавтографией, возможно осуществление и нерадиоактивного варианта детекции, при котором для мечения фрагментов кДНК используют биотиновые группы или другие химические группировки, которые после переноса фрагментов на мембрану и иммобилизации детектируют с помощью одной из известных коммерческих систем нерадиоактивной детекции с использованием хемилюминесценции.
Детекцию разделенных немеченых фрагментов кДНК можно осуществлять путем переноса фрагментов на мембрану и гибридизации с меченым адапторным праймером. При этом путем гибридизации с адапторными праймерами, содержащими добавочные основания на 3'-конце, возможна избирательная визуализация поднаборов фрагментов, разделенных в данном геле.
Для подтверждения возможности идентификации и клонирования дифференциально экспрессирующихся мРНК проводили эксперименты, изложенные в нижеприведенном примере.
Пример. Идентификация и клонирование мРНК, дифференциально экспрессирующихся в тимусе и селезенке мыши.
Из тимуса и селезенки мышей выделяют параллельно препараты тотальной РНК с помощью экстракции кислым фенолом [15]. Синтез первой цепи кДНК осуществляют с использованием следующих условий: 37oC; 60 мин; объем реакции 20 мкл, в состав реакционной смеси входят 5 мкг тотальной РНК, 200 ед. обратной транскриптазы Superscript (Gibсo-BRL, США), 10 пмоль (T)-праймера, биотинилированного по 5'-концу (последовательность биотин-ГГГАГГЦЦЦ(Т)), 30 ед. ингибитора РНКаз из плаценты человека, дАТФ, дГТФ, дЦТФ, дТТФ (1 мМ каждый), 1 • буфер обратной транскриптазы согласно рекомендациям производителя. Удаление праймера проводят с помощью переосаждения цетилтриметиламмоний бромидом [16], после чего проводят дополнительную очистку на колонках Wizard(Promega, США) согласно рекомендациям производителя. Очищенный препарат осаждают 3 объемами этанола с использованием 2 мкг гликогена (Boehringer-Mannheim, Германия) в качестве носителя. Навешивание олиго(дГ) хвоста проводят в следующих условиях: 37oC; 20 мин; объем реакции 20 мкл; в состав реакционной смеси входят гибрид мРНК-первая цепь кДНК, 20 ед. терминальной трансферазы (Gibco-BRL, США), 0,02 мМ дГТФ, 1 • буфер терминальной трансферазы согласно рекомендациям производителя. Синтез второй цепи кДНК проводят в следующих условиях: денатурация 98oC; 1,5 мин; отжиг 60oC; 2 мин; элонгация 72oC; 20 мин, ; объем реакции 25 мкл; в состав реакционной смеси входят гибрид мРНК-кДНК, 10 пмоль (Ц)-праймера (последовательность 5' ААГГААТТ(Ц)); дАТФ, дГТФ, дЦТФ, дТТФ (0,1 мМ каждый); 1,5 ед. ДНК-полимеразы Bio-Taq (Биомастер, Россия); 1 • буфер Bio-Taq согласно рекомендациям производителя. Расщепление кДНК эндонуклеазой рестрикции проводят в следующих условиях: 37oC, 60 мин; объем реакции 20 мкл; 4 ед. эндонукл. рестрикции Sau3A (New England Biolabs, США), 1 • буфер фермента (согласно рекомендациям производителя). После гидролиза реакцию останавливают добавлением ЭДТА до 20мМ и 3'-концевые фрагменты кДНК иммобилизуют на стрептавидиновые микрошарики (Promega, США) согласно рекомендации производителя. При этом добавляют адаптор: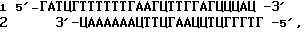 ,
,
и лигируют при 12oC в течение ночи. Реамплификацию фрагментов кДНК с помощью ПЦР проводят с использованием Био-(T) праймера и праймера 1 в следующих условиях: денатурация 95oC, 1,2 мин; отжиг 55oC, 1,5 мин; элонгация 72oC, 3 мин; объем реакции 100 мкл; в состав реакционной смеси входят 30 пмоль Био-(T)-праймера, 30 пмоль праймера 1, 2,5 ед. ДНК-полимеразы Bio-Taq; смесь дезоксинуклеотидтрифосфатов, 0,1 мМ каждый; 1 • буфер Bio-Taq. После 15 циклов ПЦР фрагменты иммобилизуют на стрептавидиновые микрошарики (Promega, США), затем снимают свободную цепь обработкой 100 мМ NaOH в течение 10 мин, промывают буфером состава 40 mM Tris-Cl pH 7,0, 20 mM MgCl, 50 NaCl, отжигают праймер 3 (5' ГТГГГЦТЦЦААГЦТТЦ), добавляют радиоактивный α - 32P дАТФ и включают метку в течение 5 мин с помощью модифицированной ДНК-полимеразы фага T7 Sequenase (United States Biochemicals, США), после чего добавляют смесь дНТФ (до 0,2 мМ каждый) и достраивают полные цепи. Отщепление фрагментов проводят последовательно эндонуклеазами рестрикции EcoRV, PstI, MspI, Hin PI (New England Biotabs, США). После каждой обработки эндонуклеазой фрагменты собирают, денатурируют и наносят на секвенирующий полиакриламидный гель (5% акриламид, 0,25% метиленбисакриламид, 7 М мочевина, 1 • буфер TBE). Разделение фрагментов проводят в стандартных условиях, после чего гель фиксируют в 10%-ной уксусной кислоте в течение 30 мин, сушат и авторадиографируют. Дифференциально экспрессируемые в тимусе и селезенке последовательности мРНК идентифицируют путем прямого сравнения наборов полос кДНК, полученных из тимуса и селезенки, и разделенных в соседних дорожках. Для клонирования дифференциально экспрессирующихся последовательностей соответствующие полосы геля вырезают и фрагменты элюируют инкубацией в 150 mM NaCl, 50 mM Tris-Cl pH 8,0, 10 mM EDTA в течение ночи. Затем проводят осаждение фрагментов 3 объемами 96%-ного этанола с использованием гликогена в качестве носителя. На 3'-конец фрагмента навешивают олиго(дГ) хвост с помощью терминальной трансферазы, как описано выше, и обработанный таким образом фрагмент амплифицируют путем ПЦР с использованием (Ц)-праймера и праймера 1. Амплифицированный фрагмент очищают с помощью электрофореза в агарозном геле, переводят в легкоплавкую агарозу и выделяют в чистом виде с помощью фенольной экстракции [17]. Для клонирования фрагментов их обрабатывают рестриктазой Sau3A и EcoRI и лигируют в плазмидный вектор pUC18, расщепленный рестриктазой BamHI и EcoRI, после чего трансформируют компетентные бактерии. Наличие рекомбинантных клонов проверяют с помощью амплификации вставок с помощью ПЦР. Проверку специфичности клонированных фрагментов проводят путем гибридизации 32P-меченых вставок из клонов с блотами амплифицированной кДНК и поли(А)+ РНК из соответствующих органов. Приведенные на фиг. 3 данные показывают, что указанный способ приводит к образованию множества меченых дискретных фрагментов кДНК, при этом сравнение картин разделения гель- электрофорезом кДНК из двух разных типов клеток выявляет полосы, соответствующие дифференциально экспрессируемым мРНК. Примером дифференциальных полос являются полосы, обозначенные номерами 1 и 2 на фиг. 3. Клонирование фрагментов кДНК, соответствующих полосам 1 и 2 фиг. 3, провели согласно методу, указанному выше. Гибридизация клонированных фрагментов 1 и 2 с перенесенными на мембрану препаратами поли(А) РНК (фиг. 4) доказала, что клонированные фрагменты кДНК действительно представляют дифференциально экспрессирующиеся в тимусе и селезенке мыши мРНК.
Источники информации
1. Sargent, T.D. Isolation of differentially expressed genes. Methods in Enzymol. 1987, 152, 423 - 432.
2. PCT/GB 89/00460, 1989.
3. Liang, P., Pardee, A.B. Differential display of eukaryotic messenger RNA by means of the polymerase chain reaction. Science, 1992, 257, 967 - 971.
4. Liang, P. , Averboukh, L., Pardee, A.B. Distribution and cloning of eukaryotic mRNAs by means of differential display: refinements and optimization. NucL Acids Res. 1993, 21, 3269 - 3275.
5. Bauer, D., Muller, H., Reich, J., Riedel, H., Ahrenkiel, V., Warthoe, P. , Strauss, M. Identification of differentially expressed mRNA species by an improved display technique (DDRT-PCR). Nucl.Acids Res. 1993, 21, 4272 - 4280.
6. Sasaki, Y.F., lwasaki, T., Kobayashi, H., Tsuji, S., Ayusawa, D., Oishi, M. Construction of an equalized cDNA library from human brain by semi-solid self-hybridization system. DNA Research. 1994, 1, 91 - 96.
7. Fischer,S.G.,Lerman,L.S. Length-independent separation of DNA restriction fragments in two-dimensional gel-electrophoresis. Cell. 1979, 16, 191 - 200.
8. Uitterlinden, A. G., Slagboom, P., Knook, D.L., Vijg, J. Two-dimensional DNA fingerprinting of human individuals. Proc.Natl.Acad.Sci.USA. 1989, 86, 2742 - 2746.
9. Lerman, L.S., Silverstein, K. Computational simulation of DNA melting and its application to denaturing gradient gel electrophoresis. Methods in Enzymol. 1987, 155, 482 - 501.
10. Kimmel, A.R., Berger, S.L. Preparation of cDNA and the generation of cDNA libraries : Overview. Methods in Enzymol. 1987, 152, 307 - 316.
11. Newton, C.R., Graham, A., Heptinstall, L.E., Powell, S.J., Summers, C., Kalsheker, N., Smith, J.C., Markham, A.F. Analysis of any point mutation in DNA. The amplification refractory mutation system (ARMS). Nucl.Acids Res. 1989, 17, 2503 - 2516.
12. Maruyama, K., Sugano, S. Oligo-capping: a simple method to replace the cap structure of eukaryotic mRNAs with oligoribonucleotides. Gene, 1994, 138, 171 - 174.
13. Orita, M., lwahana. H., Kanazawa, H., Hayashi, K., Sekiya, T. Detection of polymorphism of human DNA by gel electrophoresis as single-strand conformation polymorphisms. Proc.Natl.Acad.Sci.USA, 1989, 86, 2766 - 2770.
14. Hatada, l., Hayashizaki, Y., Hirotsune, S., Komatsubara, H., Mukai, T. A genomic scanning method for higher organisms using restriction sites as landmarks. Proc.Natl.Acad.Sci.USA. 1991, 88, 9523 - 9527.
15. Chomczynski, P., Sacchi, N. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal.Biochem. 1987, 162, 156 - 159.
16. Belyavsky,A.V., Vinogradova, T.V., Rajewsky, K. PCR- based cDNA library construction: General cDNA libaries at the level of a few cells. Nucl. Acids Res. 1989, 17, 2919 - 2932.
17. Sambrook, J., Pritsch, E.F., Maniatis, T., eds. Molecular Cloning. A laboratory manual. Second ed. (1989). Cold Spring Harbor Laboratory Press, New York.

Изобретение предназначено для использования в медицине при диагностике заболеваний и в научно-исследовательской практике при изучении их этиологии на генетическом уровне. Для детекции дэмРНК выделяют мРНК из сравниваемых типов клеток, синтезируют соответствующие им кДНК, обрабатывают кДНК частощепящими рестриктазами, отбирают в каждом отдельном наборе рестрикционных фрагментов соответствующие либо только 3'-концевым, либо только 5'-концевым областям мРНК каждого типа клеток, метят полученные наборы и поздразделяют их на непересекающиеся поднаборы кДНК-фрагментов, которые анализируют методом электрофореза в геле, сравнивают картины из разделения и выявляют фрагменты кДНК с различающейся интенсивностью сигнала. Для клонирования фрагменты кДНК, соответствующие дэмРНК, после детекции изолируют из геля, амплифицируют методом ПЦР и встраивают в подходящий для клонирования вектор. 2 с. и 14 з.п. ф-лы, 6 ил.
| Liang, P., Pardee, A.B | |||
| Differential display of eukaryotic messenger RNA by means of the polymerase chain reaction | |||
| Пуговица для прикрепления ее к материи без пришивки | 1921 |
|
SU1992A1 |
