Изобретение относится к области биотехнологии (к генетической инженерии) и представляет собой экспрессионный вектор "pCAT-3'", который найдет применение для изучения чужеродной ДНК в системе с репортерным геном. В частности, данный вектор позволяет изучать сигналы экспрессии, расположенные в 3'- нетранслируемых областях (3'-НТО) различных эукариотических генов.
Для изучения регуляторных областей ДНК, влияющих на экспрессию эукариотических генов, обычно используются векторы с репортерным геном. Полученные конструкции экспрессируются в соответствующих клеточных культурах либо транзиентно, либо после стабильной интеграции в геном. Для клонирования в них чужеродной ДНК эти векторы имеют полилинкерные участки, содержащие ряд сайтов различных рестрикционных эндонуклеаз (рестриктаз). Однако, практически все коммерчески доступные векторы имеют полилинкерные участки в областях, расположенных "выше" 5'-конца репортерного гена. Это связано с тем, что до недавнего времени обычно изучались регуляторные области, лежащие в геноме в 5'-областях генов и "выше" - такие как энхансеры транскрипции и промоторы.
Отсутствие подобных полилинкерных участков в областях, расположенных "ниже" 3'-конца репортерного гена, является определенным недостатком всех известных экспрессионных эукариотических векторов. Задача клонирования фрагментов ДНК в такие области вектора возникает, в частности, при изучении 3'-НТО генов.
Ряд известных экспрессионных векторов, которые можно было бы использовать для решения таких задач, имеет обычно один уникальный сайт (в лучшем случае два) той или иной рестриктазы, расположенный "ниже" 3'-конца репортерного гена. Это такие векторы, как: "pSEAP-Control", "pβgal-Control" [1], "pGL3-Control", "pRL-TK", "pRL-CMV", "pRL-SV40", "pGL2", "рСАТ3-Control" [2] . Они содержат различные репортерные гены - человеческой плацентарной щелочной фосфатазы, бактериальных β- -галактозидазы и хлорамфениколацетилтрансферазы (CAT), люциферазы светлячка (насекомые).
Известна также плазмида "pCAT-Control" [2, 3 4], содержащая в качестве репортерного гена ген CAT и предназначенная к использованию в качестве позитивного контроля при экспрессии этого гена в других конструкциях. Экспрессия репортерных генов в перечисленных векторах находится под контролем различных вирусных регуляторных элементов. В общем случае, клонирование фрагментов ДНК в указанные сайты возможно лишь в несколько стадий - с применением специфических олигонуклеотидных адапторов, либо же клонирования по "тупым концам". Такое клонирование сравнительно трудоемко, и поэтому достаточно неудобно.
В основу изобретения положена задача создания эукариотического экспрессионного вектора, имеющего полилинкерный участок с рядом сайтов общеупотребительных рестриктаз в области, примыкающей к 3'-концу репортерного гена, что позволяло бы клонировать в этот участок разнообразные фрагменты ДНК одностадийно. Организация этих сайтов в указанном полилинкере должна также удовлетворять условию возможности получения систематических делеций клонированного фрагмента ДНК с обеих его сторон. Необходимость в подобном векторе возникла при изучении 3'-НТО одного из генов человека.
Задача решена введением в 3'-концевую область репортерного гена в плазмиде "pCAT-Control" полилинкерного участка, удовлетворяющего поставленному условию. Из всех коммерчески доступных векторов плазмида "pCAT-Control", как исходная конструкция, была наиболее удобной, ввиду необходимого отсутствия в ней ряда сайтов определенных рестриктаз.
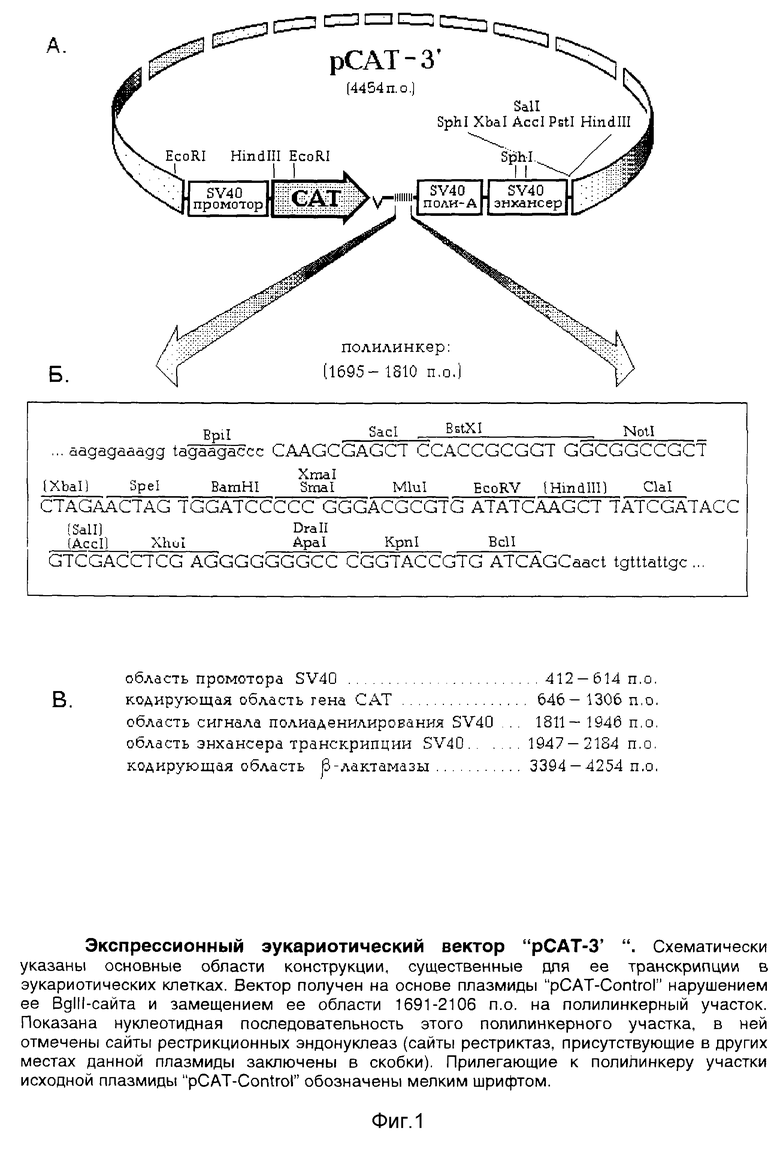
Изобретением предлагается вектор "pCAT-3'", сконструированный на основе известной плазмиды "pCAT-Control", содержащей в качестве репортерного гена бактериальный ген хлорамфениколацетилтрансферазы (CAT). В данной плазмиде ее область, находящаяся "ниже" гена CAT и соответствующая BpiI / Hpal-рестриктному фрагменту, была заменена на участок, составленный из двух синтетических олигонуклеотидных адапторов и участков полилинкера известной плазмиды "pBluescript II SK+" [5]. Указанный участок содержит 14 уникальных сайтов общеупотребительных рестриктаз и позволяет легко клонировать в него различные фрагменты ДНК. Организация этих сайтов делает возможным в составе данной конструкции получение в клонированном фрагменте систематических делеций с обеих его сторон с помощью экзонуклеазы ExoIII.
Экспрессионный вектор "pCAT-3'" размером 4454 п.о., принципиальная схема которого приведена на фиг. 1, A, содержит:
все основные области плазмиды "pCAT-Control", обеспечивающие его репликацию в бактериальных клетках и экспрессию в эукариотических, в том числе:
а) область промотора SV40: 412-614 п.о.;
б) кодирующую область гена CAT: 646-1306 п.о.;
в) область сигнала полиаденилирования SV40: 1811-1946 п.о.;
г) область энхансера транскрипции SV40: 1947-2184 п.о.;
д) кодирующую область гена β-лактамазы (ген устойчивости к ампициллину, обеспечивающий селекцию данной конструкции в клетках E.coli): 3394-4254 п.о. (представлено на фиг. 1, B).
полилинкерный участок, расположенный в области 1695-1810 п.о. (соответствующий области 1691-2106 п.о. исходной плазмиды "pCAT- Control) с нуклеотидной последовательностью
5'- CAAGCGAGCT CCACCGCGGT GGCGGCCGCT CTAGAACTAG TGGATCCCCC GGGACGCGTG ATATCAAGCT TATCGATACC GTCGACCTCG AGGGGGGGCC CGGTACCGTG ATCAGC
Данный полилинкер содержит 14 уникальных сайтов рестрикции, расположенных в следующей последовательности
5'- BpiI, SacI, Bst XI, NotI, SpeI, BamHI, SmaI (XmaI), MluI, EcoRV, ClaI, XhoI, ApaI, KpnI, BclI
(представлено на фиг. 1, Б; сайты рестриктаз, присутствующих в других местах данной плазмиды, на указанном рисунке заключены в скобки);
- рестрикционный сайт BgIII, присутствующий в исходной плазмиде "pCAT-Control", нарушен.
Емкость встраивания чужеродной ДНК по крайней мере не менее 3 т.п.о.
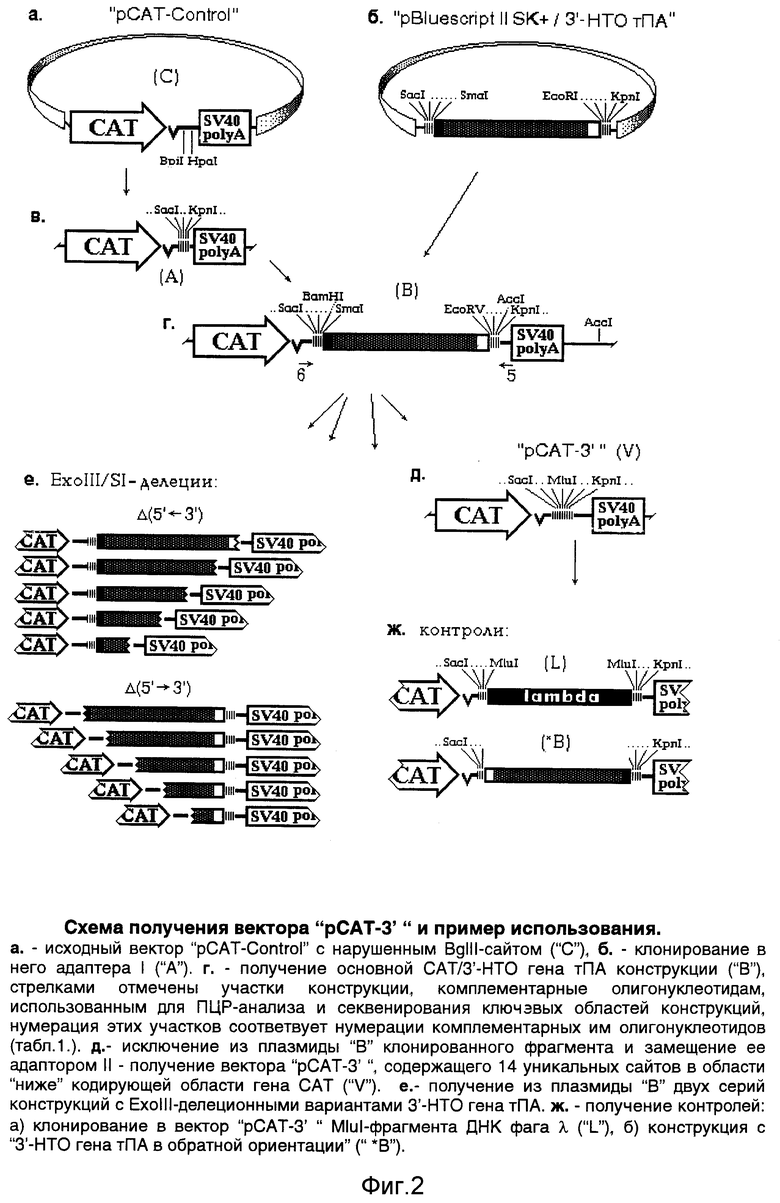
Экспрессионный вектор "рСАТ-3'" сконструирован следующим образом:
- в плазмиде "pCAT-Control" удаляют сайт BgIII - после ее разрезания этой рестриктазой, достраивания липких концов до тупых и их лигирования (все "генноинженерные" манипуляции здесь и далее выполняют, как это описано в соответствующих разделах руководства Сэмбрука и соавт. [6]);
- в полученной плазмиде (конструкция "C", фиг.2, а) участок, расположенный между сайтами рестриктаз BpiI и HpaI (соответствующий области 1691-2106 п. о. исходlной плазмиды "pCAT-Control"), замещают на адаптор 1, полученный отжигом олигонуклеотидов "1" и "2" (таблица). Полученная плазмида "A", таким образом, приобретает в указанной области сайты рестриктаз SacI и KpnI (фиг.2; в);
- по этим сайтам (SacI и KpnI) в конструкцию "A" субклонируют ~950 п.о. SacI / KpnI - фрагмент ДНК (конструкция "B", фиг.2, г). Указанный фрагмент ДНК получают вырезанием его по этим сайтам из плазмиды "pBluescript II SK+" (фиг. 2, б), в полилинкер которой его предварительно переклонируют по сайтам XmaI и EcoRI из другой конструкции (см. ниже). Таким образом, данный SacI / KpnI - фрагмент ДНК имеет по своим краям следующие участки полилинкера плазмиды "pBluescript II SK+": с 5'-конца: SacI - SmaI (XmaI)-участок, с 3'-конца: EcoRI - KpnI-участок. Этот фрагмент ДНК фактически является "переносчиком" указанных участков полилинкера плазмиды "pBluescript II SK+" в конечную конструкцию;
- в полученной конструкции "В" (фиг.2, г) указанный фрагмент ДНК затем замещают после его вырезания по сайтам XmaI и EcoRV на адаптор II, образованный отжигом олигонуклеотидов "3" и "4" (таблица). Этот адаптор соединяет участки полилинкера, при этом восстанавливая сайты XmaI и EcoRV и внося между ними сайт MluI (фиг.2, д). В результате, длина сформированного таким образом полилинкерного участка составила 116 п.о.
Нуклеотидную последовательность полилинкерной области в полученной конструкции "рСАТ-3'" (она обозначена, как "V" на фиг.2, д) проверяют секвенированием по Сэнгеру [6] этого участка (по обеим цепям ДНК) с использованием олигонуклеотидов "5" и "6" (таблица) в качестве праймеров (эти олигонуклеотиды комплементарны участкам вектора, примыкающим к данному полилинкеру, как это отмечено на фиг.2 г).
Клонированный в данный полилинкер вектора "pCAT-3'" фрагмент ДНК оказывается в составе транскрипта данной конструкции, при ее трансфекции в соответствующие клетки. С 5'-конца этого транскрипта находится кодирующая область гена CAT, с 3'-конца - транскрибируемый участок "области полиаденилирования SV40" (если этот "фрагмент" не содержит собственной "области полиаденилирования"). В том случае, если указанный фрагмент ДНК содержит в себе регуляторные участки посттранскрипционного контроля экспрессии какого-либо гена (соответствуя, например, 3'-НТО его транскрипта), он может оказывать влияние и на экспрессию гена CAT - увеличивая, либо уменьшая ее. Проводя целенаправленные изменения в нуклеотидной последовательности данного фрагмента ДНК в составе этой конструкции и измеряя при этом активность гена CAT, можно изучать указанные регуляторные участки.
В частности, организация сайтов рестриктаз в данном полилинкере позволяет в составе этой же конструкции получать систематические делеции в клонированном фрагменте ДНК - как с одной его стороны, так и с другой. Для этого плазмиду линеаризуют совместной рестрикцией либо с помощью рестриктаз SacI и какой-нибудь другой, сайт которой находится в этом же полилинкере с дистальной стороны клонированного фрагмента, либо Kpnl и какой-нибудь из рестриктаз, сайт которой находится с проксимальной стороны клонированного фрагмента. SacI- и KpnI-концы при этом устойчивы к экзонуклеазе Exolll и создают от нее "защиту" области вектора, область же "фрагмента" оказывается "незащищенной". Поэтому, после стандартной обработки такой плазмиды экзонуклеазой Exolll, Sl-нуклеазой и лигазой можно получать указанные делеции клонированного фрагмента ДНК.
Изобретение иллюстрируется следующим примером.
Клонированный в конструкцию "А" (фиг.2, в) ~950 п.о. SacI / KpnI - фрагмент ДНК (имевший в качестве концевых участков области полилинкера плазмиды "pBluescript II SK+") представляет собой 3'-НТО гена тканевого активатора плазминогена (тПА) человека, субклонированную в указанную плазмиду по сайтам XmaI и EcoRI (фиг. 2, б) из ранее полученного нами рекомбинантного фага, выделенного из библиотеки ДНК человека и содержавшего соответствующий участок гена тПА [7, 8]. Получившуюся конструкцию ("B", фиг.2, г) можно формально рассматривать, как продукт клонирования в вектор "pCAT-3'" этого же фрагмента, но по сайтам XmaI и EcoRV.
Далее получают "контроли": а) указанный фрагмент ДНК вырезают из конструкции "В" по сайтам рестриктаз SmaI и EcoRV (образующих "тупые концы") и вновь в нее вставляют в обратной ориентации (конструкция "*B" фиг,2, ж); б) в MluI-сайт вектора "pCAT-3'" клонируют MluI-фрагмент ДНК фага λ длиной 960 п.о. (контроль "длины вставки", конструкция "L", фиг.2, ж).
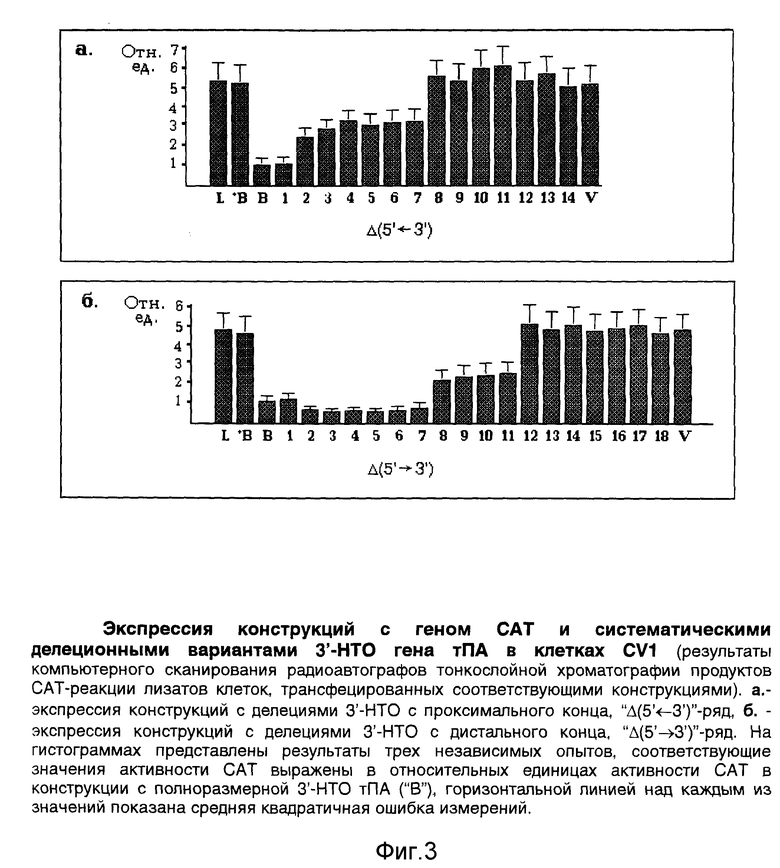
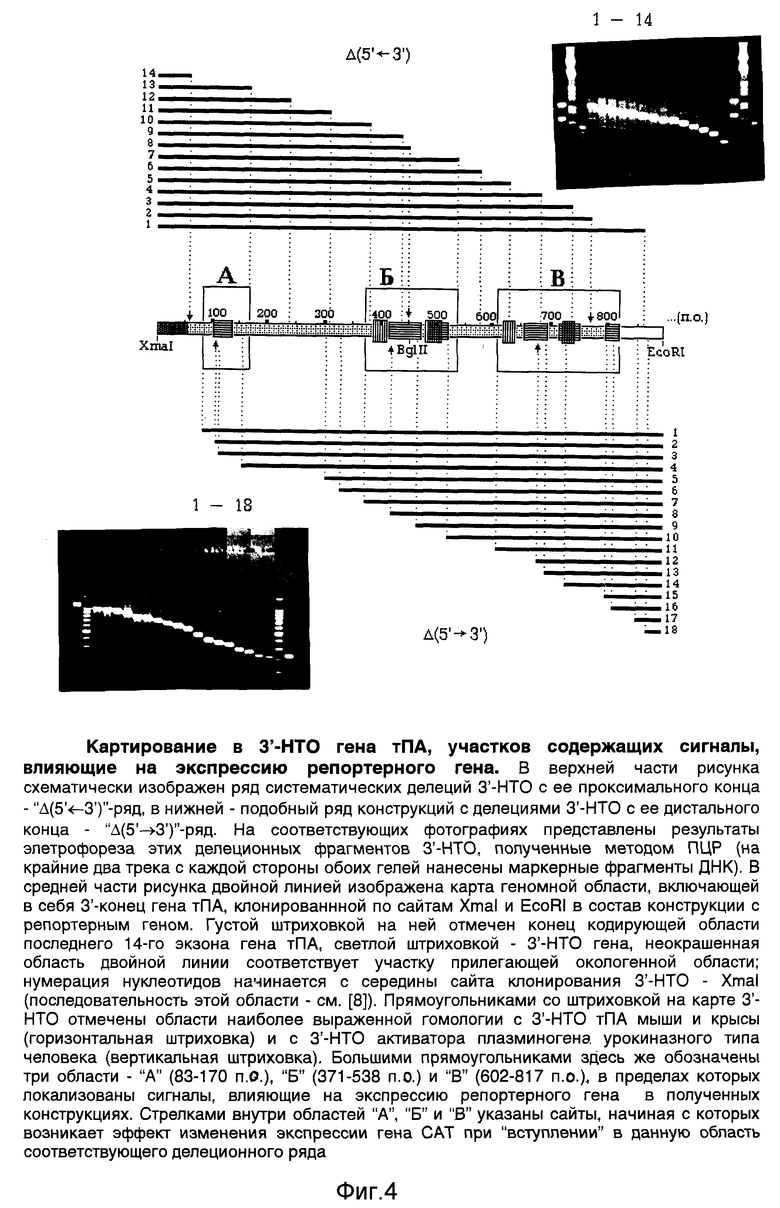
Наконец, на основе плазмиды "B" по стандартной методике с использованием экзонуклеазы Exolll [2] получают два ряда конструкций с делециями 3'-НТО гена тПА с обеих ее сторон (фиг.2, е). Для делеций 3'-НТО с дистальной стороны конструкцию "B" предварительно расщепляют рестриктазами SacI и SmaI, для делеций 3'-НТО с проксимальной стороны - рестриктазами KpnI и EcoRV. Размеры делеций предварительно определяют методом ПЦР [6], затем уточняют секвенированием сайтов делеций с использованием вышеуказанных праймеров (олигонуклеотиды "5" и "6", таблица). Эти же праймеры используют для секвенирования ключевых областей всех остальных конструкций.
Полученные конструкции транзиентно экспрессируют в различных культурах эукариотических клеток с использованием стандартной процедуры кальций-фосфатной трансфекции, как это описано в работе [6]. На фиг.3 представлены результаты измерения активности гена CAT [6] в обеих панелях делеционных конструкций, трансфецированных в клетки CV1 (клетки эпителия почки зеленой мартышки) в трех независимых экспериментах. По сравнению с контролями, 3'-НТО гена тПА ингибирует активность гена CAT в 6-8 раз. На фиг.4 показана схема картирования в 3'-НТО (с помощью ее делеций) трех участков, непосредственно ответственных за указанный эффект. Эти участки, обозначены, как "А" (83-170 п.о.), "Б" (371-538 п.о.) и "В" (602-817 п.о.) на карте 3'-НТО в центральной части фиг.4. Их отдельный вклад в экспрессию гена CAT можно оценить примерно, как следующий: участок "А", внося наиболее слабый вклад, увеличивает экспрессию в 1,5-2 раза, участки "Б" и "В" - уменьшают ее, каждый в 2-3 раза.
Литература
1. Clontech. Catalog. 1996. Clontech Laboratories Inc. p. 153-185.
2. Promega. "Biological Research Products" Catalog. 1997. Promega Corporation, p. 221- 250.
3. Technical Bulletin N 81, "Promega". 1996.
4. GenBanK AC.X65321.
5. Fermentas. Catalog. 1996-1997. MBI Fermentas. p. 208.
6. Sambrook J., Fritch E.F., Maniatis T. Molecular Cloning: A Laboratory Manual. N.Y.; Cold Spring Laboratory Press. 1989.
7. Сарафанов А.Г., Тимофеева М.Я., Алешков С.Б., Куприянова Н.С., Банников В. М. , Захарьев В.М., Баев А.А. "Выделение и характеристика фрагментов ДНК области гена тканевого активатора плазминогена и прилегающих к нему участков генома человека."// Молекуляр. биология. 1994. Т. 28. С. 790-795.
8. GenBank, АС.77531.

Изобретение относится к биотехнологии и генетической инженерии. Представляет собой эукариотический экспрессионный вектор, позволяющий клонировать разнообразные фрагменты ДНК одностадийно. Изобретением предлагается вектор "рСАТ-3", сконструированный на основе известной плазмиды "рСАТ - Control". Вектор содержит в качестве репортерного гена бактериальный ген хлорамфениколацетилтрансферазы (САТ). В данной плазмиде область Bpi I/Hpal, находящаяся ниже гена САТ, была заменена на участок из двух синтетических олигонуклеотидных адапторов и участков полилинкера плазмиды "pBluescript//SK+". Указанный участок содержит 14 уникальных сайтов общеупотребительных рестриктаз и позволяет легко клонировать в него различные фрагменты ДНК. 4 ил., 1 табл.
Экспрессионный эукариотический вектор "рСАТ-3" размером 4454 п.о. для клонирования фрагментов чужеродной ДНК в область, примыкающую к 3'-концу репортерного гена, и изучения содержащихся в них сигналов посттранскрипционного контроля экспрессии генов, содержащий:
все основные области плазмиды "рСАТ-Control", обеспечивающие его репликацию в бактериальных клетках и экспрессию в эукариотических, в том числе:
а) область промотора SV 40: 412 - 614 п.о.;
б) кодирующую область гена САТ: 646 - 1306 п.о.;
в) область сигнала полиаденилирования SV 40: 1811 - 1946 п.о.;
г) область энхансера транскрипции SV 40: 1947 - 2184 п.о.;
д) кодирующую область гена β-лактамазы (ген устойчивости к ампициллину, обеспечивающий селекцию данной конструкции в клетках E.coli): 3394 - 4254 п. о.;
полилинкерный участок, расположенный в области 1695 - 1810 п.о. с нуклеотидной последовательностью
5' - CAAGCGAGCT CCACCGCGGT GGCGGCCGCT CTAGAACTAG TGGATCCCCC GGGACGCGTG ATATCAAGCT TATCGATACC GTCGACCTCG AGGGGGGGCC CGGTACCGTG ATCAGC
и содержащий 14 уникальных сайтов рестрикции, расположенных в следующей последовательности
5' - BpiI, SacI, Bst XI, NotI, SpeI, BamHI, SmaI (XmaI), MluI, EcoRV, ClaI, XhoI, ApaI, KphI, BclI
рестрикционный сайт BglII, присутствующий в исходной плазмиде "рСАТ-Control", нарушен;
емкость встраивания чужеродной ДНК по крайней мере не менее 3 т.п.о.
| Сарафанов А.Г | |||
| и др | |||
| Выделение и характеристика фрагментов ДНК области гена тканевого активатора плазминогена и прилегающих к нему участков генома человека | |||
| Молекуляр | |||
| Биология, 1994, т.28, с.790 - 795. |

