Изобретение относится к биотехнологии, в частности к генетической инженерии, и представляет собой сконструированную in vitro рекомбинантную плазмидную ДНК, содержащую синтетический ген гранулоцитарного колониестимулирующего фактора человека, тандем промоторов триптофанового оперона Escherichia coli и синтетический участок - усилитель трансляции, обусловливающие биосинтез полипептида со свойствами гранулоцитарного колониестимулирующего фактора человека, а также штамм Escherichia coli - продуцент этого полипептида.
Гранулоцитарный колониестимулирующий фактор человека (ГКСФ) представляет собой гликопротеид с молекулярной массой 19 000 Да. ГКСФ, продуцируемый моноцитами-макрофагами, фибробластами и эндотелиальными клетками, индуцирует пролиферацию колоний нейтралов и дифференциацию клеток-предшественников в нейтрофилы, а также стимулирует активность зрелых нейтрофилов [1, 2, 3, 4].
ГКСФ снижает продолжительность нейтропении при химиотерапии злокачественных опухолей и трансплантации костного мозга, укорачивает период гранулоцитопении, индуцированной облучением, что обусловливает широкое медицинское применение ГКСФ в онкологии и при лечении ряда инфекционных заболеваний.
Известны способы получения ГКСФ, основанные на использовании культуральной жидкости клеточных линий, продуцирующих ГКСФ [5, 6, 7], а также на экспрессии гена ГКСФ в трансформированных вектором клетках высших эукариот COS, CHO, C-127 [1, 7, 8]. Недостатком этих способов является чрезвычайно низкий выход целевого продукта и как следствие высокая стоимость препаратов ГКСФ. Поэтому значительно более перспективным является способ получения ГКСФ микробиологическим синтезом, который обеспечивает возможность получения продукта с более высоким выходом из сравнительно недорогого исходного сырья. Использование при этом химического подхода позволяет создать оптимальные для бактериальной экспрессии варианты структурного гена, а также регуляторных элементов, контролирующих его экспрессию. К тому же известно, что полисахаридная цепь гликопротеида ГКСФ не является необходимой для его биологической активности [9].
Известны способы получения ГКСФ микробиологическим синтезом [3, 7], показывающие, что присутствие дополнительного метионинового остатка на N-конце молекулы рекомбинантного полипептида не влияет на его биологические свойства. Недостатком этих способов является то, что ген ГКСФ в рекомбинантных плазмидах находится под контролем индуцибельных промоторов. Необходимость процесса индукции чаще всего труднодоступными дорогими индукторами снижает технологичность процесса получения рекомбинантного полипептида.
Наиболее близким к заявляемому техническому решению (прототипом) является способ, описанный в работе [10]. Рекомбинантная плазмидная ДНК содержит ген ГКСФ человека под контролем термоиндуцибельного P1 промотора фага λ . Уровень экспрессии рекомбинантного ГКСФ в штамме-продуценте E.coli достигает 3-5% от суммы клеточных белков.
Недостатком способа-прототипа является относительно низкий уровень синтеза кодируемого полипептида ГКСФ (3-5% от суммарного клеточного белка) и необходимость применения температурной индукции при 42oC.
Технической задачей изобретения является получение полипептида со свойствами гранулоцитарного колониестимулирующего фактора человека путем конститутивного синтеза и увеличение уровня его биосинтеза.
Поставленная задача решается путем конструирования рекомбинантной плазмидной ДНК pGGF8, кодирующей конститутивный синтез полипептида со свойствами ГКСФ человека, и штамма Escherichia coli SG20050/pGGF8, обеспечивающего синтез этого полипептида с уровнем экспрессии не ниже 10% суммарного клеточного белка. Конститутивный высокий уровень синтеза целевого полипептида обеспечивается тем, что плазмида pGGF8 содержит тандем промоторов триптофанового оперона E.coli и синтетический усилитель трансляции гена 10 бактериофага 17.
Рекомбинантная плазмидная ДНК pGGF8, кодирующая полипептид со свойствами гранулоцитарного колониестимулирующего фактора человека, характеризуется следующими признаками:
имеет молекулярную массу 2,88 Md (4,360 т.п.о.);
кодирует аминокислотную последовательность зрелого гранулоцитарного колониестимулирующего фактора человека [11];
состоит из: XhoI/EcoRI - фрагмента ДНК плазмиды pGM280 [12], содержащего тандем промоторов триптофанового оперона E.coli, терминатор транскрипции фага лямбда, ген bla β -лактамазы и участок ori инициации репликации; EcoRI/SalI - фрагмента промежуточной плазмиды TroТEGF (b) размером 0,640 т.п.о. , включающий участок TREN [13] для эффективной трансляции гена gcsf, и ген gcsf, кодирующий аминокислотную последовательность зрелой формы ГКСФ [11];
содержит: тандем промоторов триптофанового оперона E.coli; синтетический усилитель трансляции (TNEN) гена 10 бактериофага 17; синтетический ген gcsf; в качестве генетического маркера ген bla-лактамазы, определяющий устойчивость трансформированных плазмидой pGGF8 клеток к ампициллину; уникальные сайты узнавания рестрикционными эндонуклеазами, имеющими следующие координаты:
EcоRI - 280, KpnI - 292, ClaI - 386,
PvuII - 602, BamHI - 1130, NcoI - 1766,
PstI - 782 и 3610, HindIII - 512 и 959,
XbaI - 1961 и 2064.
Существенным отличием предложенной плазмидной конструкции является то, что ген gcsf находится под контролем тандема промоторов триптофанового оперона E.coli, а для усиления трансляции используется синтетический усилитель трансляции, что в совокупности обеспечивает конститутивный синтез целевого белка с высоким выходом.
Для получения штамма-продуцента полипептида со свойствами гранулоцитарного колониестимулирующего фактора человека трансформируют компетентные клетки Escherichia coli SG20050 рекомбинатной плазмидной ДНК pGGF8.
Полученный штамм Escherichia coli SG20050/pGGF8 характеризуется следующими признаками.
Морфологические признаки. Клетки мелкие палочковидной формы, грамотрицательные, неспороносные, 1 х 3-5 мкм, подвижные.
Культуральные признаки. При росте на плотной среде LA колонии круглые, гладкие, полупрозрачные, блестящие, серые, край ровный; диаметр колоний 1-3 мм; консистенция пастообразная. Рост в жидких средах (LB, минимальная среда с глюкозой) характеризуется ровным помутнением с образованием легкого осадка.
Физико-биохимические признаки. Клетки растут при температуре 4 - 42oC при оптимуме pH 6,8 - 7,2. В качестве источника азота используют как минеральные соли в аммонийной форме, так и органические соединения в виде пептона, триптона, дрожжевого экстракта, аминокислот. В качестве источника углерода используют аминокислоты, глицерин, углеводы.
Устойчивость к антибиотикам. Клетки проявляют устойчивость к ампициллину (до 300 мкг/мл), обусловленную наличием в плазмиде гена бета-лактамазы, и устойчивость к тетрациклину (до 50 мкг/мл), связанную с наличием транспозона [14].
Штамм E.coli SG20050/GGF8 обеспечивает конститутивный синтез полипептида со свойствами ГКСФ в количестве не ниже 10% от суммарного клеточного белка, что в два раза выше, чем в прототипе, и освобождает в отличие от прототипа проведение операции индукции в ходе культивирования штамма.
Полученный штамм депонирован в НИИ коллекция культур микроорганизмов ГНЦ ВБ "Вектор" под номером В663.
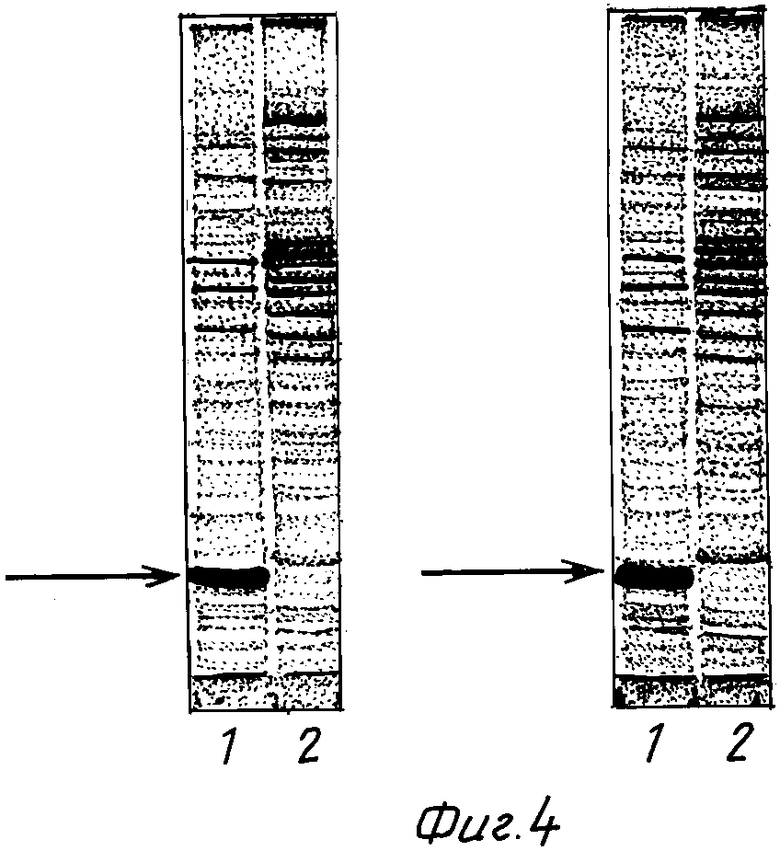
На фиг. 1 представлена физическая карта рекомбинантной плазмиды pGGF8; на фиг. 2 - нуклеотидная последовательность гена ГКСФ с прилегающими регуляторными элементами: тандем промоторов Ptrp (7-278 п.о.), усилитель трансляции TREN (293-386 п.о.), ген ГКСФ (390-914 п.о.); инициирующий и терминирующий кодоны подчеркнуты; в рамки взяты сайты рестриктаз: EcorI, KpnI, ClaI, и XhoI; на фиг. 3 - аминокислотная последовательность полипептида ГКСФ, кодируемого рекомбинантной плазмидой pGGF8; на фиг. 4 - электрофореграмма лизатов клеток штамма-продуцента E.coli SG20050/pGGF8 (дорожка 1) и штамма-реципиента E. coli SG20050 (дорожка 2) в 12% полиакриламидном геле. Стрелкой указан полипептид ГКСФ.
Изобретение иллюстрируется следующими примерами.
Пример 1. Конструирование промежуточной рекомбинантной плазмидной ДНК pTrcTEGF(b).
10 мкг плазмидной ДНК pGCSF(b) [11] обрабатывают последовательно рестриктазами ClaI и SalGI в соответствии с методикой, описанной в работе [15], и из полученного гидролизата выделяют в 1% геле легкоплавной агарозы фрагмент длиной 0,530 т.п.о., содержащий ген gcsf.
10 мкг плазмидной ДНК TREN5 [13] обрабатывают последовательно рестриктазами KpnI и ClaI и из полученного гидролизата выделяют в 1% геле легкоплавной агарозы фрагмент длиной 0,094 т.п.о., представляющий собой синтетический усилитель трансляции.
10 мкг плазмидной ДНК pTrc 99A [16] обрабатывают последовательно рестриктазами KpnI и SalGI и из полученного гидролизата выделяют векторную часть плазмиды.
Полученные фрагменты и векторную часть плазмиды pTrc 99A сшивают при помощи лигазной реакции в 50 мкл буфера для лигирования [15]. 10 мкг реакционной смеси используют для трансформации компетентных клеток E.coli C600 [17] . Трансформанты высевают на LB-агар, содержащий 75 мкг/мл ампициллина. Из выросших клонов выделяют плазмидную ДНК и анализируют рестрикционным анализом. Отбирают ДНК, содержащую нужный набор рестрикционных фрагментов. Плазмида pTrc TEGF(b) содержит Trc-промотор, вслед за ним EcoRI-сайт, KpnI-сайт, далее идет синтетический усилитель трансляции и структурный ген gcsf, фланкированный сайтами ClaI и SalI. Клетки E.coli, трансформированные плазмидой pTrc TEGF(b), способны синтезировать полипептид ГКСФ при индукции pTrc-промотора изопропилтиогалактозидом (ИПТГ).
Пример 2. Конструирование рекомбинантной плазмидной ДНК pGGF8.
10 мкг плазмидной ДНК pTrcTEGF(b) обрабатывают последовательно эндонуклеазами рестрикции EcoRI и SalGI в соответствии с методикой, описанной в работе [15] , и из полученного гидролизата выделяют в 1% геле легкоплавкой агарозы фрагмент ДНК длиной 0,64 т.п.о. 10 мкг плазмидной ДНК pGM280 обрабатывают последовательно эндонуклеазами рестрикции EcoRI и XhoI в соответствии с методикой, приведенной в работе [15], и из полученного гидролизата выделяют в 1% геле легкоплавкой агарозы векторную ДНК длиной 3,72 т.п. о. Полученные фрагмент и векторную ДНК соединяют при помощи лигазной реакции в 50 мкл буфера для лигирования [13]. 5 мкг реакционной смеси используют для трансформации компетентных клеток C600 [17]. Трансформанты высевают на LB-агар, содержащий 75 мкг/мл ампициллина. Из выросших клонов выделяют целевую плазмидную ДНК pGGF8 и анализируют ее путем обработки набором эндонуклеаз рестрикции HaeIII, HindIII, PstI, KpnI и RcoRI с последующим электрофоретическим анализом длин рестрикционных фрагментов в 6% полиакриламидном геле. Из 10 проанализированных клонов 5 показали нужный набор рестрикционных фрагментов. Целевая рекомбинантная плазмида pGGF8 содержит уникальные сайты узнавания рестрикционными эндонуклеазами, имеющие следующие координаты:
EcoRI - 280, KpnI - 292, ClaI - 386, PvuII - 602,
BamHI - 1130, NcoI - 1766, PstI - 782 и 3610,
HindIII - 512 и 959, XbaI - 1961 и 2064 (фиг. 1).
Окончательно структуру рекомбинантной ДНК pGGF8 подтверждают определением нуклеотидной последовательности в районе встроенного фрагмента, содержащего синтетический ген ГКСФ человека (фиг. 2).
Пример 3. Получение штамма-продуцента полипептида со свойствами гранулоцитарного колониестимулирующего фактора человека.
Рекомбинантной плазмидной ДНК pGGF8 трансформируют компетентные клетки Escherichia coli SG20050 [14] и получают штамм-продуцент полипептида со свойствами гранулоцитарного колониестимулирующего фактора человека.
Пример 4. Определение продуктивности штамма-продуцента полипептида со свойствами гранулоцитарного колониестимулирующего фактора человека.
Клетки E.coli SG20050/pGGF8 выращивают при 30oC в 20 мл жидкой среды LB, содержащей 75 мкг/мл ампициллина, в течение 16 ч на качалке при скорости вращения 120 об/мин. Отбирают пробу 2 мл и центрифугируют 5 мин при скорости 8000 об/мин, после чего клетки суспендируют в 200 мкл буфера, содержащего 125 мМ Трис-HCl, pH 6,8, 20% глицерин, 3% додецилсульфат натрия, 3% меркаптоэтанол, 0.005% бромфеноловый синий, инкубируют 10 мин на кипящей водяной бане, образцы 7,5 мкл анализируют электрофорезом в 12,5% полиакриламидном геле с додецилсульфатом натрия [18]. Гель окрашивают Кумасси R-250 (фиг. 4) и сканируют на лазерном сканиметре Ultroscan XL (LKB). По данным сканирования полипептид ГКСФ составляет 10 - 17% суммарного клеточного белка.
Пример 5. Выделение и характеризация рекомбинантного полипептида со свойствами гранулоцитарного колониестимулирующего фактора человека.
5 г биомассы клеток E.coli SG20050/pGGF8, выращенных в 4 х 250 мл жидкой среды LB в условиях, описанных в примере 4, суспендируют в 50 объемах буфера A (20 мМ Трис-HCl, pH 7,9; 70 мМ меркаптоэтанол, 1 мкг/мл лизоцим) при 20oC, обрабатывают ультразвуком при частоте 22 кГц 2 х 15 с. После центрифугирования осадок промывают 0,1% раствором дезоксихолиевой кислоты в буфере A, затем буфером A. Растворение осадка проводят в буфере B (20 мМ Трис-HCl, pH 8,9; 8 М мочевина, 0,5 мМ ФМСФ, 2 мМ трилон Б). Для хроматографической очистки полученного экстракта телец включения используют схему, описанную в работе [10]. В результате из 5 г биомассы выделено 5,2 мг электрофоретически гомогенного препарата.
N-концевую аминокислотную последовательность определяют на секвинаторе 477A и ФТГ анализаторе 120A фирмы "Applied Biosystems", США. Препарат рекомбинантного ГКСФ имеет следующую структуру N-конца молекулы: Met+Thr ProLeuGly..., т.е. соответствует структуре N-конца природного ГКСФ человека с дополнительным метиониновым остатком, не влияющим на его биологические свойства [3, 7].
Пример 6. Определение биологической активности рекомбинантного полипептида.
Клетки-предшественники грануло- и грануломоноцитопоэза клонируют в полувязкой культуральной среде следующего состава: 79% RPM1-1640, 1% метилцеллюлозы, 20% эмбриональной сыворотки телят, 280 мг/л L-глютамина, 4 мкМ 2-меркаптоэтанола, 50 мг/л гентамицина, 1 мл тест-системы содержит 105 неадгезирующих миелокариоцитов. Исследуемый препарат рекомбинантного ГКСФ добавляют в культуральную среду в концентрациях 0,5, 1,0, 5,0, 10, 20, 30, 50 нг/мл. По 0,5 мл приготовленной клеточной взвеси помещают в 24-луночные планшеты ("Corning", США) и культивируют в течение 7 сут. в CO2-инкубаторе при 37oC, 5% CO2 и 100% влажности воздуха. Под колонией подразумевают очаг гемопоэза, содержащий 50 и более клеток. С целью морфологического изучения колоний готовят препараты, которые окрашивают азуром-11-эозином.
Результаты экспериментов обрабатывают методом вариационной статистики с использованием t-критерия Стьюдента. За единицу активности принимают количество препарата, необходимое для образования одной колонии.
Препарат рекомбинантного ГКСФ, полученный в примере 5, обладает активностью 107 ед. акт./мг белка.
Полученные данные по характеристике и биологической активности продукта экспрессии искусственного гена ГКСФ в клетках штамма E.coli SG20050/pGGF8 свидетельствуют о соответствии исследуемого полипептида его природному аналогу ГКСФ.
Таким образом, заявляемое техническое решение позволяет получить полипептид со структурой и свойствами, идентичными структуре и свойствам природного ГКСФ человека; биосинтез полипептида конститутивен, и уровень его составляет не ниже 10% от суммарного клеточного белка за счет того, что ген ГКСФ находится под контролем тандема промоторов триптофанового оперона E. coli, а трансляция белка усиливается за счет синтетического усилителя трансляции. Все это позволяет по сравнению с прототипом упростить процесс получения рекомбинантного ГКСФ за счет исключения стадии индукции процесса биосинтеза и увеличить выход целевого продукта в 2 раза.
Источники информации.
1.Nagata S., Tsuchiya M., Asano S., Kaziro Y., Yamazaki T., Yamamoto O., Hirata Y. , Kubota N., Oheda M., Nomura H., Ono M. // Nature, 1986, v.319, No.6052, p.415-418.
2. Nagata S., Tsuchiya M., Asano S., Yamamoto O., Hirata Y., Jubota N., Oheda M., Nomura H., Yamazaki T. // EMBO J., 1986, v.5, p.575-581.
3. Souza L.M., Boone T.C., Gabrilove J., Lai P.H., Zsebo K.M., Mudrock D.C., Chazin V.R., Bruszewski J., Kenneth H.L., Chen K.K., Barendt J., Platzer E. , Moore M.A.S., Mertelsmann R., Welte K. // Scince, 1986, v.232, No. 4746, p.61-65.
4. Cohen A.M., Zsebo K.M., Inoue H., Hines D., Boone T.C., Chazin V.R., Tsai L., Ritch T., Soyza L.M. // Proc. Natl. Acad. Sci, USA, 1987, v.87, No. 8, p.2484-2288.
5. Патент США N 4833127, кл. A 61 K 37/02, 1989.
6. Nomura H., Imazeki I., Oheda M., Kubota N., et al. // EMBO J., 1986, v. 5, p. 871-876.
7. Европейский патент N 215126, кл. C 12 N 15/00, 1987.
8. Европейский патент N 220520, кл. C 12 N 15/00, 1987.
9. Oheda M., Hasegawa M., Hattori K., Kuboniwa H., Kojima T., Orita T., Tomonou K. , Yamazaki T., Ochi N. // J.Biol.Chem., 1990, v.265, No.20, p. 11432-11435.
10. Патент США N 4810643, кл. C 12 N 15/00, 1989.
11. Кашьяп С.П., Быстров Н.С., Болдырева Е.Ф., Полякова И.А., Северцова И.В., Коробко В.Г. // Биоорган. химия, 1992, т. 18, N 1, c. 71-77.
12. Заявка на изобретение N 94009371/13 от 16.03.94 г. "Рекомбинантная плазмидная ДНК p280GM, кодирующая полипептид со свойствами GM-CSF человека, и штамм E.coli - продуцент полипептида со свойствами GM-CSF человека".
13. Кашьяп С.П. Синтез и экспрессия искусственного гена гранулоцитарного колониестимулирующего фактора человека: Дис... канд. биол. наук: ИБХ РАН. -М., 1992.
14. Trisler P. , Gottesman S. // J.Bacteriol., 1984. v.160, No.2, p. 184-191.
15. Sambrook J. , Fritsch E.F., Maniatis T. // Molecular Cloning. A. Laboratory Manual. 2bd ed. Cold Spring Harbor, NY, 1989.
16. Amann E., Ochss B., Abel K.-J. // Gene, 1988, v. 69, p. 301-315.
17. Hanahan J. // J.Mol.Biol., 1983, v. 166, No. 4, p. 557-580.
18. Laemmli U.K. // Nature, 1970, v. 227, p. 680.


Изобретение относится к биотехнологии, в частности, к генетической инженерии. Может быть использовано для получения гранулоцитарного колониестимулирующего фактора (ГКСФ) человека. Рекомбинантная плазмидная ДНК содержит синтетический ген ГКСФ человека, тандем промоторов триптофанового оперона E. coli и синтетический усилитель трансляции гена 10 бактериофага 17. Получен штамм Escherichia coli - продуцент этого полипептида. Биосинтез полипептида конститутивен, и уровень его составляет не ниже 10% от суммарного клеточного белка. 2 с. п. ф-лы, 4 ил.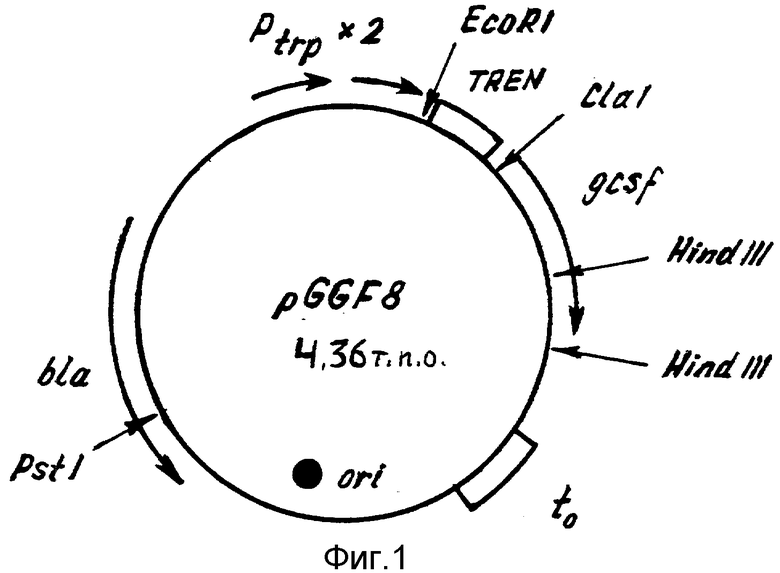
EcoRI 280; Kpn I - 292; ClaI - 386;
Pvu II - 602; BamHI - 1130; NcoI - 1766;
Pst I -782 и 3610; Hind III - 512 и 959; XbaI - 1961 и 2064.
| US, патент, 4810643, кл | |||
| Способ гальванического снятия позолоты с серебряных изделий без заметного изменения их формы | 1923 |
|
SU12A1 |

