Изобретение относится к биотехнологии, в частности к генетической инженерии, и представляет собой сконструированную in vitro рекомбинантную плазмидную ДНК, содержащую синтетический ген гранулоцитарно-макрофагального колониестимулирующего фактора человека (ГМ-КСФ), тандем промоторов (ptrpx2) триптофанового оперона E.coli и синтетические участки инициации трансляции, обуславливающие эффективный биосинтез полипептида с биологической активностью ГМ-КСФ, а также штамм Escherichia coli продуцент этого полипептида.
ГМ-КСФ глипопротеин, зрелая форма которого образуется путем отщепления от белка предшественника (144 а.о.) сигнального пептида (17 а.о.) и последующего гликозилирования. Его источниками являются Т-лимфоциты, эндотелиальные клетки и фибробласты. ГМ-КСФ стимулирует in vitro пролиферацию и дифференциацию стволовых клеток с образованием колоний нейтрофильных и эозинофильных лейкоцитов и макрофагов. Он также вовлечен во многие другие биологические и иммунологические функции организма, такие как изменение морфологии эффекторных клеток, их подвижности, цитотоксической активности и фагоцитоза [1]
Способность ГМ-КСФ стимулировать рост гранулоцитов и макрофагов делает возможным его клиническое использование для снижения побочных эффектов луче- и химеотерапии, в терапии побочных эффектов при трансплантации костного мозга.
Известны способы получения ГМ-КСФ человека, основанные на использовании экстрактов плаценты либо культуральной жидкости клеточных линий - суперпродуцентов ГМ-КСФ [2, 3] а также экспрессии гена ГМ-КСФ в COS-клетках [4] Недостатком этих подходов является чрезвычайно низкий выход целевого продукта и, как следствие, высокая стоимость препаратов ГМ-КСФ. Поэтому значительно более перспективным является способ получения ГМ-КСФ микробиологическим синтезом, который обеспечивает возможность получения целевого продукта со значительно более высоким выходом из сравнительно недорогого исходного сырья. Использование при этом химического подхода позволяет создать оптимальные для бактериальной экспрессии варианты структурного гена, а также регуляторных элементов, контролирующих его экспрессию.
Известна рекомбинантная плазмида pGMtrp [5] содержащая синтетический ген, кодирующий полипептид со свойствами ГМ-КСФ человека и экспрессирующийся под контролем промотора триптофанового оперона E. coli. Клетки E.coli SG20050, содержащие эту плазмиду обеспечивают уровень синтеза целевого продукта, не превышающий 5-7% от суммарного белка клетки, что недостаточно для получения промышленного штамма-продуцента.
Известна рекомбинантная плазмида, в которой к-ДНК-копия гена ГМ-КСФ находится под контролем индуцируемого температурой промотора фага λ [6] Уровень экспрессии ГМ-КСФ достигает 7-10% от суммарного белка клетки, а присутствие дополнительного аминокислотного остатка метионина на 5'-конце рекомбинантного полипептида не влияет на его биологические свойства. По принципу конструирования, технической сущности и достигаемому результату данная плазмида является наиболее близкой к заявляемой и выбрана в качестве прототипа. Существенным недостатком прототипа является использование индуцибельного промотора, что снижает технологичность процесса получения рекомбинантного продукта.
Задачей разработки является получение высокого конститутивного биосинтеза рекомбинантного ГМ-КСФ человека в компетентных клетках E.coli.
Поставленная задача была решена путем конструирования новой рекомбинантной плазмиды p280GM, кодирующей конститутивный синтез ГМ-КСФ, и штамма E. coli SG20050/p280GM, обеспечивающего уровень экспрессии ГМ-КСФ около 15% от суммарного клеточного белка.
Рекомбинантная плазмидная ДНК p280GM характеризуется следующими признаками:
кодирует аминокислотную последовательность зрелого ГМ-КСФ человека;
имеет молекулярную массу 2,7 Мд (3966 т.п.о.);
состоит из:
PstI/BamHI фрагмента ДНК плазмиды pDS280 (1083 п.о.), который содержит часть гена b-лактамазы и тандем промоторов триптофанового оперона E.coli;
NcoI/PstI фрагмента плазмиды pGMtrp, содержащий часть синтетического гена ГМ-КСФ, кодирующую полипептид (13-127) зрелого ГМ-КСФ человека (2834 т.п. о.), часть гена b-лактамазы и терминатор транскрипции фага l;
синтетического дезоксиолигонулеотидного дуплекса, 5'-GATCAATTCTATGGCACCAGCACGATCTCCCTCTCCTTCTACTCAAC- 3' 3'- TTAAGATACCGTGGTCGTGCTAGAGGGAGAGGAAGATGAGTTGGTAC- 5', который содержит ATG-кодон и кодирует полипептид (1-12) зрелого ГМ-КСФ человека.
содержит:
синтетический ген ГМ-КСФ человека;
в качестве генетического маркера ген b-лактамазы, детерминирующий устойчивость трансформированных плазмидой p280GM клеток E.coli к пенициллиновым антибиотикам;
уникальные сайты узнавания рестрикционными эндонуклеазами, расположенными на следующих расстояниях вправо от сайта EcoRI (279 п.о.)- BamHI 454 нуклеотида; PstI 2938 нуклеотидов.
Существенным отличием предложенной плазмидной конструкции является то, что с помощью дезоксиолигонуклеотидного адаптора заменяются 12 5'-концевых триплетов гена ГМ-КСФ. Эта замена не нарушает кодируемую им аминокислотную последовательность, но оптимизирует вторичную структуру мРНК, что приводит к существенному увеличению уровня синтеза целевого продукта.
Для получения бактериального штамма-продуцента полипептида с биологической активностью ГМ-КСФ человека компетентные клетки E.coli SG20050 трансформируют сконструируемой плазмидой p280GM.
Полученный таким образом штамм E.coli SG20050/p280GM характеризуется следующими признаками:
Морфологические признаки. Клетки мелкие? утолщенной палочковидной формы, грамотрицательные, неспороносные.
Культуральные признаки. Клетки хорошо растут на простых питательных средах. При росте на агаре "Дифко" колонии круглые, гладкие, прижаты, мутные, блестящие серые, край ровный. При росте в жидких средах (на минимальной среде с глюкозой или LB-бульоне) образуют интенсивную ровную муть.
Физико-биологические признаки. Клетки растут при температуре от 4 до 40oC при оптимуме pH от 6,8 до 7,0. В качестве источника азота используют как минеральные соли в аммонийной форме, так и органические соединения в виде пептона, триптона, дрожжевого экстракта, аминокислот и т.д. В качестве источника углерода используют аминокислоты, глицерин, углеводы.
Устойчивость к антибиотикам. Клетки проявляют устойчивость к ампициллину (до 300 мкг/мл), обусловленную наличием плазмиды, а также к тетрациклину (до 50 мкг/мл), благодаря наличию транспозона.
Существенными отличиями штамма E.coli SG20050/280GM является то, что он обусловливает конститутивный синтез полипептида со свойствами ГМ-КСФ человека с уровнем экспрессии около 15% суммарного клеточного белка, что более чем в 2 раза превосходит прототип.
Полученный штамм депонирован во Всесоюзной коллекции промышленных микроорганизмов во ВНИИ генетики под номером ВКПМ B-6613.
На фиг. 1 показаны схема получения и физическая карта рекомбинантной плазмиды P280GM.1 -выделение фрагментов и лигирование ДНК лигазой фага T4 в присутствии дезоксиолигонуклеотидного дуплекса. 2 трансформация компетентных клеток E. coli; на фиг. 2 первичная структура EcoRI BamHI фрагмента днк p280GM, содержащего синтетический ген ГМ-КСФ, и кодируемая им аминокислотная последовательность. Подчеркнуты прерывистой чертой снизу нуклеотидные замены в структурной части гена ГМ-КСФ, отличающие его от природного аналога [4] чертой снизу терминирующие кодоны; чертой сверху последовательность SD; на фиг. 3 электрофореграмма лизатов клеток E.coli SG20050/pGMtrp (дорожка 1), E.coli SG20050/p280GM (дорожка 2), E.coli SG20050 (дорожка 3) в 12% ПААГ.
Пример 1. Химический синтез олигонуклеотидов.
Синтез олигонуклеотидов выполняют твердофазным фосфорамидитным методом на ДНК-синтезаторе System 1 (Beckman) с наращиванием олигонуклеотидной цепи в направлении от 3'-конца к 5'-концу с помощью защищенных фосфамидитов - 5-диметокситритил-N-ацил-2-дезоксинуклеозид-3-О-(метоксидиизопропиламино)-фосактивированных тетразолом. Синтез проводят в масштабе 0,5-0,7 мкмоль. Используют синтетический цикл, описанный в работе [8] После окончания синтеза защитные группы удаляли последовательной обработкой тиофенолятом триэтиламмония и концентрированным аммиаком. При этом происходит отделение олигонуклеотида от носителя. 5'-Диметокситритильную группу удаляют кислотной обработкой и олигонуклеотид очищают электрофорезом в 20% ПААГ, содержащем 7М мочевину. Выход 1-5 о.е.
Пример 2. Конструирование рекомбинантной плазмидной ДНК p280GM.
Клетки бактерий E.coli G600, содержащие плазмидную ДНК pDS280 [7] выращивают при 37oC в LB-бульоне, содержащем 100 мкг/мл ампициллина, до стационарной фазы. Затем плазмидную ДНК выделяют в соответствии с процедурой, описанной в работе [9]
Аналогичным образом выделяют ДНК плазмиды pGMtrp, содержащей синтетический ген ГМ-КСФ [5] Полученные препараты плазмидных ДНК используют для конструирования плазмиды p280GM. С этой целью по 10 мкг плазмидных ДНК pDS280 и pGMtrp инкубируют смесью эндонуклеаз рестрикции BamHI и PstI, и NcoI и PstI (10 ед каждой), соответственно, в 30 мкл буфера R, содержащего 20 мм Трис-HCl, pH 7,5, 50 мМ NaCl,10 мМ MgCl2, 7 мМ меркаптоэтанол, в течение 1 ч при 37oC. После инкубации реакции останавливают двукратной экстракцией смесью фенолфлороформа (1:1) и ДНК осаждают 70% этанолом. Анализ полноты гидролиза и выделение BamHI/PstI-фрагмента плазмидой ДНК pDS280 (1028 п.о.) (фрагмент 1) и NcoI/PstI-фрагмента плазмидой ДНК pGMtrp (2834 п.о.) (фрагмента 2) проводят при помощи электрофореза в 1%-ном геле легкоплавкой агарозы. Далее фрагмент 1 (1 мкг) соединяют в присутствии дуплекса: 5'-GATCAATTCTATGGCACCAGCACGATCTCCCTCTCCTTCTACTCAAC- 3' 3'- TTAAGATACCGTGGTCGTGCTAGAGGGAGAGGAAGATGAGTTGGTAC- 5', с фрагментом 2 (0,4 мкг) с помощью 30 ед. T4 ДНК лигазы в 50 мкл буфера L в течение 6 ч при 15oC и реакционную смесь используют для трансформации компетентных клеток E. coli C600. Трансформанты высевают на агаризованную среду, содержащую ампициллин (100 мкг/мл) и полученные таким образом отдельные колонии выращивают при 37oC в LB-бульоне, содержащем 100 мкг/мл ампициллина, до стационарной фазы. Затем плазмидную ДНК выделяют в соответствии с процедурой, описанной в работе [9] и анализируют с помощью эндонуклеазы рестрикции HaeIII. Из клонов, содержащий встроенный фрагмент, выделяют плазмидную ДНК, строение которой подтверждают прямым определением нуклеотидной последовательности ДНК в районе встройки с помощью модифицированного метода Максама-Гилберта [10]
Пример 3. Получение штамма-продуцента полипептида со свойствами ГМ-КСФ человека.
Плазмидой p280GM трансформируют компетентные клетки E.coli SG20050 и получают штамм-продуцент полипептида со свойствами ГМ-КСФ человека.
Пример 4. Вестернблот-анализ.
Клетки E. coli SG20050 и E.coli SG20050/p280GM выращивают при 37oC в 20 мл LB-бульона в течение 20 ч на качалке при скорости вращения 120 об/мин, отбирают пробу объемом 2 мл и клетки центрифугируют 10 мин при 10000 об/мин. Клетки суспендируют в 100 мкл буфера, содержащего 125 мМ Трис-NCl, pH 6,8, 20% глицерин, 3% SDS, 3% меркаптоэтанол, 0,005% бромфеноловый синий, нагревают 10 мин на кипящей водяной бане и охлаждают во льду. 10 мкл образцов подвергают электрофорезу в 12,5% SDS-ПААГ. По окончании электрофореза белки переносят на нитроцеллюлозные фильтры (Schleicher @ Schuel HAWP 293 23 HA 0,45 m) с помощью прибора для электро-блот-анализа. После окончания переноса фильтр инкубируют в течение ночи в буфере PBS (10 мМ калийфосфатный буфер, pH 7,2 150 мМ NaCl), содержащем 3% (весовая концентрация) БСА при 4oC. После инкубации в растворе БСА, фильтры инкубируют в течение 2-3 ч со специфическими антителами, разведенными PBS до концентрации 5 мкг/мл. После инкубации фильтры тщательно промываются в PBS, содержащем 0,1% твин-20, и добавляют коньюгат пероксидазы хрена с антителами второго порядка (производство ВЕКТОР-БИОПРОДУКТ, Кольцово Новосибирской области), инкубируют не менее 1 ч. при комнатной температуре. Для окраски фильтра 50 мг 4-хлор-1-нафтол, растворенного в 1 мл этанола, разводят 1:100 PBS и добавляют 30%-ный пероксид водорода до концентрации 0,006% После появления окрашивания фильтр промывают PBS, высушивают и хранят в темноте. По данным вестерн-блот-анализа единственный полипептид, реагирующий с антителами к человеческому ГМ-КСФ (SIGMA, США), это полипептид с молекулярной массой около 16 кД, детерминируемый плазмидой p280GM.
Пример 5. Определение биологической активности.
Клетки E.coli SG20050/p280GM выращивают как в примере 4. Клетки суспендируют в 20 мл буфера, содержащего 10%-ную сахарозу; 0,1 М трис-HCl, pH 7,5; 5 мМ Na2EDTA, и разрушают ультразвуком на установке Soniprep 150 (MSE). Полученный лизат стерилизуют ультрафильтрацией через фильтры 0,45 mm CA/CN (SIGMA, США) и активность ГМ-КСФ определяют, используя метод клонирования гемопоетических предшественников в полутвердых агаровых культурах [6] Мононуклеары получают из крови гематологически здоровых доноров разделением в градиенте плотности фикол- верографина (SIGMA, США). Клетки в концентрации 5•105 лунка культивируют в среде Мак-Коя 5A (НПО "ВЕКТОР", Россия) в планшетах для культур тканей (SIGMA, США; 24 wells/plate) с добавлением стерильных лизатов клеток E. coli SG20050 и E.coli SG20050/p280GM. Подсчет количества колоний проводят на 7-е сутки.
По данным определения биологической активности лизат клеток E.coli SG20050/p280GM стимулирует образование колоний клеток- предшественников макрофагов и гранулоцитов; спонтанного колониеобразования (при добавлении лизатов клеток E.coli SG20050) не происходит.
Пример 6. Определение продуктивности штамма E.coli продуцента полипептида со свойствами ГМ-КСФ человека.
Клетки E.coli SG20050/p280GM выращивают как в примере 4. Содержание рекомбинантного ГМ-КСФ определяют по отношению к суммарному клеточному белку.
Клетки суспендируют 200 мкл буфера, содержащего 125 мМ Трис-HCl, pH 6,8; 20% глицерин; 3% SDS; 3% меркаптоэтанол; 0,005% бромфеноловый синий, нагревают 10 мин на кипящей водяной бане и охлаждают во льду. Образцы 2, 5 и 10 мкл подвергают электрофорезу в 12% SDS-ПААГ. По окончании электрофореза гель промывают 10%-ным раствором трихлоруксусной кислоты и прокрашивают при помощи кумасси R-250. После отмывки избытка красителя гель сканируют при 550 нм, используя лазерный сканиметр UltroScan XL (LKB, Швеция).
По данным сканирования рекомбинантный ГМ-КСФ составляет 15-20% всего белка E.coli.
Пример 7. Выделение и характеризация белка, синтезирующегося в клетках штамма E.coli продуцента полипептида со свойствами ГМ-КСФ человека.
Клетки E.coli SG20050/p280GM выращивают при 37oC в 500 мл (10 колб по 50 мл) LB-бульона как указано выше в примере 4, биомассу собирают центрифугированием 15 мин при 6000 об/мин, клеточный осадок промывают 200 мл буфера, содержащем 10% -ную сахарозу; 0,1 M трис-HCl, pH 7,5; 5 mM Na2EDTA и 0,5 mM DTT, ресуспендируют в 20 мл этого же буфера и разрушают ультразвуком с помощью ультразвуковой установки Soniprep 150 (MSE). Полученный лизат осветляют центрифугированием 30 мин при 10000 об/мин, осадок дважды промывают 30 мл раствора 2 M мочевины в буфере PBS, содержащего 150 mM NaCl, 20 mM калий фосфат, pH 7,5, 5 mM Na2EDTA, растворяют в 10 мл раствора 4 M мочевины в буфере PBS. Нерастворившиеся частицы удаляют центрифугированием 30 мин при 20000 об/мин. По данным электрофореза в 12% ПААГ чистота препарата ГМ-КСФ составляет 60%
Раствор ГМ-КСФ хроматографируют на колонке с широкопористым обращенно-фазовым носителем Полисил ОДС-300 [11] в градиенте смеси ацетонитрила, изопропанола и воды (3:1:1) со скоростью 1 мл/мин с детекцией профиля элюции при 220 нм. Белковый пик, содержащий по данным электрофореза гомогенный ГМ-КСФ, собирают и используют для анализа аминокислотного состава, N-концевого анализа по способу, описанному в работе [11] Препарат ГМ-КСФ хранят при -70oC. Данный способ выделения позволяет получить из 0,5 л культуры клеток E.coli SG20050/p280GM при плотности 109 клеток мл 20-30 мг рекомбинантного ГМ-КСФ человека.
Полученные данные о строении продукта экспрессии искусственного гена ГМ-КСФ в клетках штамма E.coli SG20050/p280GM свидетельствуют о соответствии исследуемого полипептида его природному аналогу ГМ-КСФ человека.
Таким образом, заявляемое техническое решение позволяет получать полипептид со структурой белка идентичной структуре и свойствам природного ГМ-КСФ человека; уровень биосинтеза ГМ-КСФ составляет около 15% суммарного клеточного белка E.coli за счет усовершенствования вторичной структуры полицистронной мРНК, содержащей синтетический ген ГМ-КСФ.
Источники информации
1. Clark S. K. and Kamen R.// Sceince, 1987, v.236, N. 4806, pp.1229-1237.
2. Burgess A.V. Wilson E.M.A. and Metcalf D.// Blood, 1979, v.54, p.614.
3. Gasson J.C. Weishard R.H. Kaufman S.E. Clark S.G. Wong G.C. Gold D.W. // Science, 1984, v.226, p.1339.
4. Wong G.G. Witek J.S. Temple P.A. Wilkens K.M. Leary A.C. Luxenberg D. P. Jones S.S. Brown E.L. Kay R.M. Orr E.C. Shoemarker C. Kaufman R.J. Hewick R.M. Wang E.A. Clark S.G.// Science, 1985, v.228, p.819.
5. Гилева И.П. Бондарь Т.С. Блинова Н.Н. Халдояниди С.К. Кравченко В.В. Офицеров В.И. Ястребов С.И. Коробко В.Г.// Доклады РАН, 1994, т.336, N 2, с. 254-256.
6. Burgess A.W. Begly C.G. Jonson G.R. Lopez A.F. Williamson D.J. Mermoid J.J. Simpson R.J. Schmitz A. DeLammarter J.F.// Blood, 1987, v.69, N 1, p. 43-51.
7. Кравченко В. В. Гилева И.П. Шамин В.В. Лихошвай В.А. Добрынин В.Н. Филиппов С. А. Коробко В. Г. Биоорганическая химия, 1988, т.14. N 10, с. 1372-1386.
8. Колосов М.Н. Коробко В.Г. Добрынин В.Н. Северцова И.В. Чувпило С.А. Быстров Н.С. Берлин Ю.А. Каюшин А.Л. Буткус В.В. Полякова И.А. Болдырева Е. Ф. Сандахчиев Л. С. Попов С.Г. Шубина Т.Н. Кравченко В.В. Серпинский О.И. Ямщиков В.Ф. Беликов С.И. Синяков А.Н. Сиволобова Г.Ф. Авторское свидетельство, N 1092176, опубл.в. Б.И. 1984, N 18.
9. Birnboim H.C.// in: Methods in Enzymology. Recombinant DNA. Part B. 1983, v. 100. Eds. Wu R. Grossman L. Moldave R. p.243-254. Academic Press, Inc.
10. Chuvpilo S.A. and Kravchenko V.V.//FEBS. 1984, v.179, N 1, p.34-36.
11. Акименко З. А. Зыков С.А. Офицеров В.И. Гилева И.П. Кравченко В.В. Сандахчиев Л.С.//Докл. АН СССР, 1991, т.40, N 4, с. 878-884М


Использование: биотехнология, в частности генетическая инженерия. Сущность изобретения: предложена новая рекомбинантная плазмида p280 GM, содержащая синтетический ген гранулоцитарно-макрофагального колониестимулирующего фактора (ГМ-КСФ), детерминирующий конститутивный синтез полипептида со свойствами ГМ-КСФ человека, и штамм E.coli SG 20050/p280GM, обеспечивающий уровень экспрессии ГМ-КСФ около 15% от суммарного клеточного белка при плотности 109 клеток/мл, что составляет около 20-30 мг в одном литре такой клеточной суспензии. 2 с.п. ф-лы, 3 ил.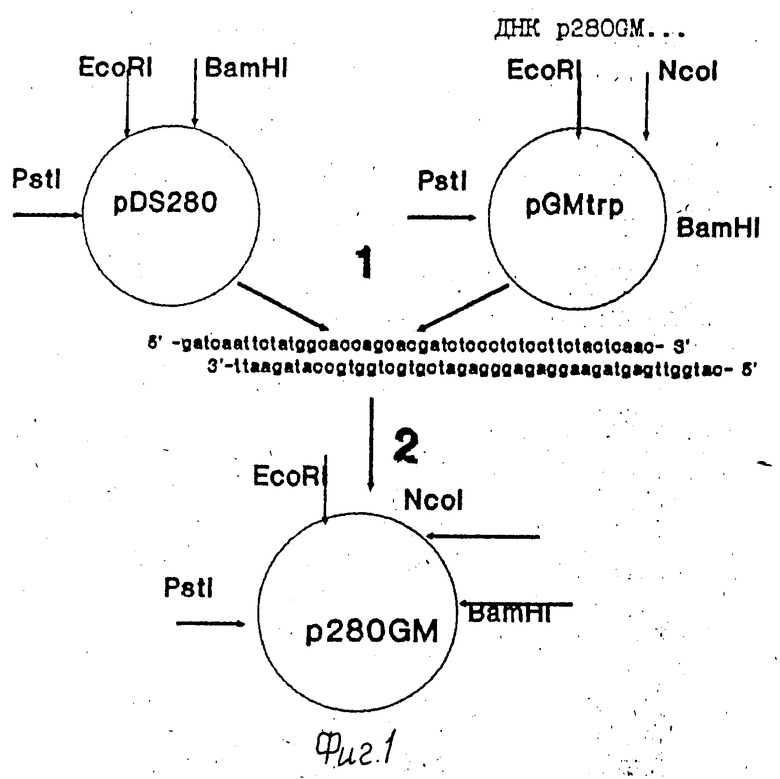
5'-GATCAATTCTATGGACCAGCATCTCCCTCTCCTTCTACTCAAC-3'
3'-TTAAGATACCGTGGTCGTAGAGGGAGAGG-AAGATGAGTTGGTAC-5',
включающий ATG-кодон и кодирующий полипептид (1 12) зрелого гранулоцитарно-макрофагального колониестимулирующего фактора человека; синтетический ген гранулоцитарно-макрофагального колониестимулирующего фактора человека; генетический маркер ген β -лактамазы, детерминирующий устойчивость трансформированных плазмидой p 280GM клеток E.coli к пенициллиновым антибиотикам; уникальные сайты узнавания рестрикционными эндонуклеазами, расположенными на следующих расстояниях вправо от сайта EcoRI (279 п.о.) - BamHI-454 нуклеотида; PstI-2938 нуклеотидов.
| Burgess A.W | |||
| et al | |||
| Blood | |||
| Кузнечная нефтяная печь с форсункой | 1917 |
|
SU1987A1 |
| Способ приготовления пищевого продукта сливкообразной консистенции | 1917 |
|
SU69A1 |
| Зубчатое колесо со сменным зубчатым ободом | 1922 |
|
SU43A1 |
