Настоящее изобретение относится к способу получения целевого вещества с помощью микроорганизма. Типичные примеры микроорганизмов, используемых в настоящем изобретении, включают микроорганизмы, принадлежащие к роду Escherichia и коринеформные бактерии, которые часто применяются для получения различных веществ. Целевые вещества, получаемые согласно настоящему изобретению, включают L-аминокислоты, антибиотики, витамины, факторы роста, физиологически активные вещества и другие вещества, обычно продуцируемые микроорганизмами. В настоящей заявке описан способ, благодаря которому повышается выход конечного целевого вещества в процессе его получения с помощью микроорганизмов.
Уровень техники
Ферментация L-аминокислот представляет собой типичный пример хорошо известного способа получения целевого вещества с помощью микроорганизмов. L-аминокислоты используются не только в качестве пищевых добавок и продуктов питания, но также в качестве компонентов различных медицинских питательных смесей, добавок к животным кормам, агентов в фармацевтической и химической промышленности и в качестве факторов роста при получении L-аминокислот, таких как L-лизин и L-гомосерин, с помощью микроорганизмов. Коринеформные бактерии, микроорганизмы, принадлежащие к родам Escherichia, Bacillus и Serratia и т. п., являются известными микроорганизмами, и их можно использовать для ферментационного получения L-аминокислот.
Для ферментационного получения L-аминокислот можно использовать бактерии "дикого типа" (штаммы "дикого типа"), ауксотрофные штаммы, индуцированные из штаммов "дикого типа", мутанты с нарушенной регуляцией метаболизма, индуцированные из штаммов "дикого типа" как мутанты, устойчивые к различным лекарственным препаратам, а также штаммы, обладающие свойствами как ауксотрофных штаммов, так и мутантов с нарушенной регуляцией метаболизма. Для получения ауксотрофных штаммов используют различные вещества в зависимости от того, какой именно штамм требуется, ауксотрофные штаммы, для получения которых используются идентичные вещества, могут иметь различную степень ауксотрофии. Точно также разнообразны мутанты с нарушенной регуляцией метаболизма, полученные в качестве мутантов, устойчивых к различным лекарственным препаратам.
В последнее время для ферментационного получения L-аминокислот стали применять технологию рекомбинантной ДНК. Теоретическое обоснование этой технологии основано на усилении системы биосинтеза L-аминокислот в микроорганизме-хозяине посредством обогащения гена (генов), кодирующих фермент (ферменты) биосинтеза L-аминокислот. Более подробно данная технология описана, например, в работе "Amino Acid Fermentation, Society Press Japan (1986)".
Однако микроорганизм, обычно используемый в ферментационном получении L-аминокислот, подвергается биосинтетическим путям метаболизма, включая биосинтетические пути L-аминокислот и биосинтетические пути коферментов, идентичные биосинтетическим путям у микроорганизмов "дикого типа". Микроорганизмы, продуцирующие L-аминокислоты, были выведены путем десенсибилизации подавления конечным продуктом или его аналогом, существующим в пути биосинтеза L-аминокислот. В качестве средства, используемого в качестве такого введения, например, для придания клетке ауксотрофных свойств или устойчивости к лекарственному препарату или для амплификации гена(генов), кодирующих биосинтетические фермент(ы), применили технологию рекомбинантной ДНК.
Посредством ферментации с использованием микроорганизмов можно получить множество других веществ помимо L-аминокислот. В качестве примеров таких веществ можно упомянуть антибиотики и витамины. Известные самые разнообразные антибиотики и самые различные вещества используются в качестве биосинтетических предшественников таких антибиотиков. Например, могут быть использованы сахара, аминокислоты, уксусная кислота, пропионовая кислота и мевалоновая кислота. Целевой антибиотик получают из таких предшественников путем превращения различных метаболитов, не являющихся предшественниками. Этот же механизм применяют для получения витаминов и других биогенных веществ.
Продуцирование вышеуказанных веществ происходит в процессе биосинтеза в клетке микроорганизма. Одним из коферментов, играющих важную роль для эффективного функционирования соответствующих ферментов в системе биосинтеза является восстановленный никотинамидадениндинуклеотидфосфат (который далее в тексте настоящей заявки будет называться NADPH). Однако опубликованные данные о связи между NADPH и получением веществ с помощью микроорганизмов отсутствуют.
Никонитамиддинуклеотидтрансгидрогеназа (которая далее в тексте настоящей заявки будет именоваться "трансгидрогеназа") известна в качестве одного из ферментов, ответственных за образование NADPH. Известно, что этот фермент присутствует в различных организмах, включая микроорганизмы рода Escherichia. В Escherichia coli, типичном микроорганизме, принадлежащем к роду Escherichia, произвели очистку трансгидрогеназы (см. David M.Clarke and Philip D.Bragg, Eur. J. Biochem., 149, 517-523 (1985)), клонирование кодирующего ее гена, подтвертив тем самым существование этого фермента (см. David M.Clarke and Philip D.Bragg, J.Bacteriology, 162, 367-373 (1985)), а также определение нуклеотидной последовательности этого гена (см. David M.Clarke, Tip W. Loo, Shirley Gillam, and Philip D. Bragg, Eur. J. Biochem. 158, 647-653 (1986)), благодаря чему существование этого фермента стало очевидным. Однако физиологическая функция данного фермента до сих пор почти не известна. Об этом свидетельствует тот факт, что варианты, дефектные по этому ферменту, не проявляют какой-либо фенотипической экспрессии.
Раскрытие изобретения
Цель настоящего изобретения состоит в том, чтобы повысить продуктивность микроорганизма по целевому веществу, включая этапы культивирования микроорганизма в культуральной среде до получения целевого вещества, которое образуется и накапливается в культуральной среде, а также этап извлечения целевого вещества из культуральной среды.
Продуктивность по целевому веществу обычно увеличивают с помощью десенсибилизации регуляции синтеза конечным продуктом (продуктами) или аналогичными продуктами, продуцируемыми биосинтетическим путем целевого вещества и кофермента, необходимого для синтеза целевого вещества, который присутствует в клетках микроорганизмов. Конкретная цель настоящего изобретения - разработать средство повышения выхода целевого вещества на основе совершенно новой теории, отличающейся от вышеописанной.
В ходе биосинтеза веществ, таких как L-аминокислоты, в живых организмах происходит множество восстановительных реакций. Во многих случаях кофермент NADPH физиологически используется как прижизненный восстановитель.
Например, для глютаматдегидрогеназы NADPH требуется в качестве кофермента в процессе биосинтеза L-глютаминовой кислоты. Для аспартат-полуальдегиддегидрогеназы и дигидродипиколат-редуктазы NADPH требуется в качестве кофермента в процессе биосинтеза L-лизина.
NADPH играет важную роль в качестве кофермента в биосинтезе других L-аминокислот. Помимо этого L-глутаминовая кислота играет важную роль в качестве донора аминогрупп в процессе биосинтеза многих L-аминокислот, поэтому NADPH требуется также в качестве источника аминогрупп в биосинтезе L-аминокислот.
NADPH обычно получают в процессе метаболизма глюкозы в пентозофосфатном цикле, который включает в себя глюкозо-6-фосфат-дегидрогеназу и фосфоглюконат-дегидрогеназу. Можно вычислить, что эффективность продуцирования NADPH в пентозофосфатном цикле составляет 12 молекул на одну молекулу глюкозы, поскольку высвобождается двуокись углерода.
С другой стороны, восстановленный никотинамидадениндинуклеотид (далее в тексте настоящей заявки называемый NADH) представляет собой молекулу, очень похожую на NADPH, однако в большинстве случаев ее нельзя использовать в качестве кофермента в биосинтезе L-аминокислот. NADH синтезируется через цикл трикарбоновых кислот (ЦТК) и в клетках обычно присутствует в достаточных количествах.
Что касается путей биосинтеза L-аминокислот, то те прижизненные компоненты, которые невозможно эффективно использовать, часто образуются из глюкозы в процессе биосинтеза нужных L-аминокислот. Предполагается, что такие компоненты обычно окисляются в ЦТК, что приводит к образованию большого количества NADH.
Авторы настоящего изобретения разработали гипотезу, согласно которой затрагивается большое количество NADPH в процессе образования целевого вещества с помощью микроорганизмов, для получения этого NADPH неизбежно потребляется глюкоза и, следовательно, снижается выход целевого вещества в пересчете на использованный сахар (гипотеза 1).
Далее авторы разработали гипотезу, согласно которой прижизненные компоненты, которые невозможно эффективно использовать в процессе образования целевого вещества, неизбежно аккумулируются в процессе получения целевого вещества с помощью микроорганизмов. Эти компоненты участвуют в метаболизме через ЦТК, что приводит к увеличению концентрации NADH в клетке (гипотеза 2).
На основании гипотез 1 и 2 можно предположить, что если внутриклеточный NADH удастся эффективно превратить в NADPH, то можно будет сэкономить сахар, который требуется для биосинтеза NADPH микроорганизмами, и целевое вещество можно будет получать с большей эффективностью. Одновременно предполагается, что трансгидрогеназу можно использовать в качестве средства для превращения NADH, полученного в ЦТК, в NADPH.
На основании вышеописанных концепций авторы провели ряд исследований. Интенсивные исследования позволили получить фрагмент ДНК, содержащий ген трансгидрогеназы, из бактерий рода Escherichia, а способность микроорганизмов продуцировать восстановленный никотинамидадениндинуклеотидфосфат повышается при использовании этого фрагмента ДНК. Авторы далее обнаружили, что выход целевого вещества повышается у вышеуказанных микроорганизмов, обладающих повышенной способностью продуцировать восстановленный никотинамидадениндинуклеотидфосфат.
Конкретно, настоящее изобретение обладает следующими признаками:
(1) способ получения целевого вещества с помощью микроорганизмов, включающий в себя следующие этапы:
- микроорганизм культивируют в среде культивирования до образования и накопления в ней целевого вещества,
- извлекают целевое вещество из среды культивирования, причем продуктивность микроорганизмов в отношении восстановленного никотинамидадениндинуклеотидфосфата повышена,
(2) способ по пункту (1), в котором целевое вещество представляет собой L-аминокислоту,
(3) способ по пункту (2), в котором L-аминокислоту выбирают из группы, в которую входят L-треонин, L-лизин, L-глутаминовая кислота, L-лейцин, L-изолейцин, L-валин и L-фенилаланин,
(4) способ по пункту (1) или (2), в котором микроорганизм представляет собой микроорганизм, принадлежащий к роду Escherichia,
(5) способ по пункту (1) или (2), в котором микроорганизм представляет собой коринеформную бактерию,
(6) способ по пунктам (1)-(5), в котором повышение продуктивности микроорганизма в отношении восстановленного никотинамидадениндинуклеотидфосфата достигается за счет увеличения ферментативной активности никотинамиднуклеотидтрансгидрогеназы в клетке микроорганизма,
(7) способ по пункту (6), в котором повышение продуктивности микроорганизма в отношении восстановленного никотинамидадениндинуклеотидфосфата достигается за счет увеличения уровня экспрессии гена, кодирующего никотинамиднуклеотидтрансгидрогеназу в клетке микроорганизма,
(8) способ по пункту (7), в котором повышение продуктивности микроорганизма в отношении восстановленного никотинамидадениндинуклеотидфосфата достигается за счет увеличения числа копий гена, кодирующего никотинамиднуклеотидтрансгидрогеназу в клетке микроорганизма.
Целевое вещество, которое продуцируют микроорганизмы в соответствии с настоящим изобретением, представляет собой различные L-аминокислоты, такие как L-треонин, L-лизин, L-глутаминовая кислота, L-лейцин, L-изолейцин L-валин и L-фенилаланин. Помимо вышеперечисленных кислот настоящее изобретение может применяться в отношении других кислот, полученных до настоящего момента с помощью микроорганизмов, при условии, что для их биосинтеза требуется NADPH, в числе таких кислот нуклеиновые кислоты, например гуаниловая кислота и инозиновая кислота, витамины, антибиотики, факторы роста и биологически активные вещества.
Далее, нет необходимости упоминать, что даже в том случае, когда вещество до настоящего времени не производилось с помощью микроорганизмов, настоящее изобретение будет относиться и к этому веществу, при условии, что для его биосинтеза требуется NADPH.
В биосинтезе стрептомицина, например, NADPH используют для синтеза дТДФ-4-оксо-4,6-дидеокси-D-глюкозы из дТДФ-глю-козы. Кроме того, аминокислоты служат предшественниками в случае пептидных антибиотиков, и, таким образом, NADPH, естественно, нужен для их биосинтеза. Далее, предшественниками пенициллина, бета-лактамного антибиотика являются L-валин, L-цистеин и L-альфа-аминоадипиновая кислота, поэтому для их биосинтеза также требуется NADPH.
Если требуется выяснить для биосинтеза какого вещества требуется NADPH, то это будет видно из пути биосинтеза этого вещества, если он известен.
Настоящее изобретение не ограничивается каким-либо определенным видом микроорганизмов, при условии что микроорганизм принадлежит к числу тех микорорганизмов, которые до настоящего времени использовались для получения веществ, например бактерии, принадлежащие роду Escherichia, коринеформные бактерии, бактерии, принадлежащие роду Bacillus, и бактерии, принадлежащие роду Serratia. Предпочтительными являются такие микроорганизмы, в которых фрагмент ДНК, содержащий точку начала репликации плазмиды, был получен для данного микорорганизма, а функции гена трансгидрогеназы и количество копий гена трансгидрогеназы могут быть увеличены. С другой стороны, наиболее предпочтительным будет штамм, который изначально обладал высокой способностью продуцировать целевое вещество, и именно такой штамм лучше всего использовать в способе повышения продуктивности по настоящему изобретению. Это объясняется тем, что в штамме с высокой продуктивностью эффект повышения продуктивности по настоящему изобретению будет более заметен, поскольку более ярко будет выражено действие факторов, описанных в вышеизложенных гипотезах 1 и 2. Примером такого штамма является штамм Escherichia coli B-3996 и подобные ему штаммы, применяемые для получения L-треонина, штамм Escherichia coli AJ12604 (FERM BP-3579) и подобные ему штаммы, применяемые для получения L-фенилаланина, Escherichia coli AJ12624 (FERM ВР-3853) и подобные ему штаммы, применяемые для получения L-глутаминовой кислоты, а также Brevibacterium lactofermentum AJ3990 (FERM P-3387, АТСС 31269) и подобные ему штаммы, применяемые для получения L-лизина.
При получении целевого вещества способом по изобретению можно использовать известные культуральные среды, выбор среды будет зависеть от типа используемого микроорганизма. Конкретно, может быть использована обыкновенная культуральная среда, содержащая источник углерода, источник азота, неорганические ионы и, при желании, другие органические компоненты. Для осуществления настоящего изобретения специальная культуральная среда не требуется.
В качестве источника углерода можно использовать сахара, такие как глюкоза, лактоза, галактоза, фруктоза и гидролизат крахмала, спирты, такие как глицерин и сорбит, органические кислоты, такие как фумаровая кислота, лимонная кислота и янтарная кислота и т.п.
В качестве источника азота можно использовать неорганические соли аммония, такие как сульфат аммония, хлорид аммония и фосфат аммония, органические соединения азота, такие как гидролизат соевых бобов, газообразный аммиак, водный аммиак.
В качестве источника питательных органических веществ, применяемых в микроколичествах, требуются такие вещества, как витамин В1, L-гомосерин и L-тирозин или экстракт дрожжей, которые должны присутствовать в подходящих количествах.
Помимо вышеуказанных веществ при желании можно добавить небольшие количества фосфата калия, сульфата магния, ионы железа, ионы марганца и т.п.
Культивирование осуществляют в известных условиях, применяемых в аналогичных случаях, определенные условия зависят от типа используемого микроорганизма. Конкретно, предпочтительно производить культивирование в течение 16-120 часов в аэробных условиях. Температуру культивирования поддерживают на уровне от 25 до 45oС, а значения рН - от 5 до 8. В некоторых случаях для подведения рН можно использовать неорганические или органические кислые или щелочные вещества, а также газообразный аммиак и т.п.
Извлечение продуктов метаболизма из культуральной жидкости после окончания культивирования осуществляют обычными способами. Конкретно, в рамках способа по изобретению может применяться комбинация обычных хорошо известных способов, таких как способ с применением ионообменной смолы, способ осаждения и т.п.
Повысить продуктивность микроорганизмов в отношении NADPH можно, например, посредством увеличения ферментативной активности трансгидрогеназы в микробных клетках.
Повысить ферментативную активность трансгидрогеназы можно, например, посредством увеличения уровня экспрессии гена трансгидрогеназы в микробных клетках. Далее, другим средством повышения ферментативной активности трансгидрогеназы является модификация гена трансгидрогеназы и создание трансгидрогеназы с повышенной активностью.
Повысить уровень экспрессии гена трансгидрогеназы в микробных клетках можно, например, посредством увеличения числа копий гена трансгидрогеназы в микробных клетках.
Чтобы повысить число копий гена трансгидрогеназы, необходим фрагмент ДНК, содержащий вышеупомянутый ген. В некоторых случаях ген трансгидрогеназы был клонирован в штамме Escherichia coli К-12, который использовался в качестве одного из штаммов, принадлежащих к роду Escherichia, и его нуклеотидная последовательность уже определена (см. D.М. Clarke et al., Eur. J. Biochem., 158, 647-653 (1986)). Таким образом, выделение фрагмента ДНК, содержащего вышеупомянутый ген, было достигнуто с помощью способа, описанного в работе D.М. Clarke et al. Нужный фрагмент ДНК можно получить также способом гибридизации, используя синтетический ДНК-зонд, полученный со ссылкой на нуклеотидную последовательность, описанную в работе D.M. Clarke и др., а также методом ПЦР, используя синтетические ДНК затравки, полученные со ссылкой на вышеуказанную нуклеотидную последовательность. Если фрагмент ДНК, содержащий ген трансгидрогеназы, лигирован с векторной ДНК, которая может автономно воспроизводиться в целевом микорорганизме, и введен в вышеуказанный микроорганизм, то можно повысить число копий гена трансгидрогеназы.
ДНК-затравка, которая используется при клонировании гена трансгидрогеназы из бактерии, принадлежащей к роду Escherichia, с использованим метода ПЦР, может быть изготовлена подходящим образом на основании, например, последовательности, известной у Escherichia coli (D.M. Clarke и др., Eur. J. Biochem., 158, 647-653 (1986)). Так как трансгидрогеназа включает две субъединицы, то, может быть, необходимо амплифицировать оба гена pntA и pntB каждой из них. Для этого подходят две затравки
5'-ЦТГАТТТТТГГАТЦЦАГАТЦАЦАГ-3' (Посл-сть 1Д: 1) и
5'-ЦГТТЦТГТТААГЦТТТЦТЦААТАА-3' (Посл-сть 1Д: 2),
которые могут амплифицировать участок из 3 т.п.н., содержащий оба гена pntA и pntB. Последовательности этих затравок незначительно отличаются от той последовательности, которая сообщается D.M. Clarke и др. Однако благодаря этому изменению последовательности возможно ввести сайт расщепления BamHl перед обоими генами pntA и pntB и сайт расщепления HindIII после обоих генов pntA и pntB. Ни сайта BamHl, ни сайта HindIII нет ни в обоих генах, ни поблизости от них. Таким образом, это сайты удобны при клонировании фрагмента амплифицированной ДНК с использованием этих ферментов рестрикции, а также при переносе в другую векторную ДНК. Синтез ДНК-затравки можно выполнить в соответствии с обычным методом, используя синтезатор ДНК модели 380В, выпускаемый фирмой Applied Biosystems, а также используя фосфоамидитовый метод (см. Tetrahedron Letters, 22, 1859 (1981)). ПЦР можно выполнить с использованием прибора "DNA Thermal Cycler" модели PJ2000, выпускаемого фирмой "Takara Shuzo Co., Ltd.", а также с использованием ДНК-полимеразы Tag в соответствии с методом, указанным производителем.
Фрагмент ДНК, содержащий ген трансгидрогеназы, может быть получен из микроорганизмов, которые отличаются от бактерий, принадлежащих к роду Escherichia. Желаемый фрагмент ДНК можно получить с использованием метода гибридизации, применяя синтетический ДНК-зонд, который изготавливается со ссылкой на нуклеотидную последовательность, описанную D.M. Clarke и др. или же с использованием метода ПЦР, применяя синтетические ДНК-затравки, которые изготавливаются со ссылкой на вышеупомянутую нуклеотидную последовательность, как на метод их изготовления.
ДНК-зонд, который следует использовать в методе гибридизации или ДНК-затравки, которые следует использовать при клонировании гена с применением метода ПЦР, могут быть изготовлены подходящим образом на основании, например, последовательности, известной у Escherichia coli (D.M. Clarke и др., Eur. J. Biochem., 158, 647-653 (1986)). Установлено, что нуклеотидная последовательность гена различна у каждого микроорганизма. Таким образом, желательно изготовить синтетические ДНК, совпадающие по своим последовательностям с участками, консервативными в отношении трансгидрогеназ, которые происходят из каждого представляющего интерес микроорганизма.
Ген трансгидрогеназы, амплифицированный методом ПЦР, лигируется с векторной ДНК, которая способна к автономной репликации к клетке бактерии, принадлежащей к роду Escherichia, при введении в бактерию, принадлежащую к роду Escherichia, и вводится в клетки бактерии, принадлежащей к роду Escherichia.
В случае введения полученного фрагмента ДНК, содержащего ген трансгидрогеназы, в микроорганизм, который отличается от бактерий, принадлежащих к роду Escherichia, вышеуказанный фрагмент ДНК лигируется с векторной ДНК, которая способна к автономной репликации в клетке микроорганизма, подвергнутого введению вышеуказанного фрагмента ДНК, и вводится в вышеуказанные клетки.
В качестве векторной ДНК, пригодной для использования в настоящем изобретении, предпочтительной является плазмидная векторная ДНК, примерами которой могут служить pUC19, pUC18, pBR322, pHSG299, pHSG399, RSF1010 и им подобные. Векторы из фаговой ДНК, отличающиеся от указанных выше, также являются пригодными. Для того, чтобы добиться эффективной экспрессии трансгидрогеназы, также приемлемо использовать промотор, дееспособный в микроорганизмах, такой как lac, trp и PL. В дополнение к этому, для того, чтобы увеличить число копий гена трансгидрогеназы, ДНК, содержащая вышеуказанный ген, может быть интегрирована в хромосому посредством метода, использующего транспозон (Berg, D.Е. и Berg, С.М., Bio/Technol., 1, 417 (1983)), фаг Мu (Японский Патент (выложенная заявка) 2-109985) или гомологичную рекомбинацию (Эксперименты в молекулярной генетике. Cold Spring Harbor Lab. (1972)).
Когда микроорганизмом, в который вводится ген, является коринеформная бактерия, то векторной ДНК, которая может быть использована в настоящем изобретении, является плазмидный вектор, способный к автономной репликации в коринеформной бактерии, такой как рАМ330 (см. Публикация японского патента 1-11280) и рНМ1519) и рНМ1519 (см. Японский Патент (выложенная заявка) 58-77895).
Для того, чтобы отобрать штамм, имеющий действительно повышенную ферментативную активность трансгидрогеназы среди штаммов-кандидатов, возможно имеющих повышенную ферментную активность трансгидрогеназы, в качестве метода для подтверждения повышения ферментной активности трансгидрогеназы можно использовать известный метод, например (D.M. Clarke и Р.D. Bragg, J. Bacteriology, 162, 367-373 (1985)).
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
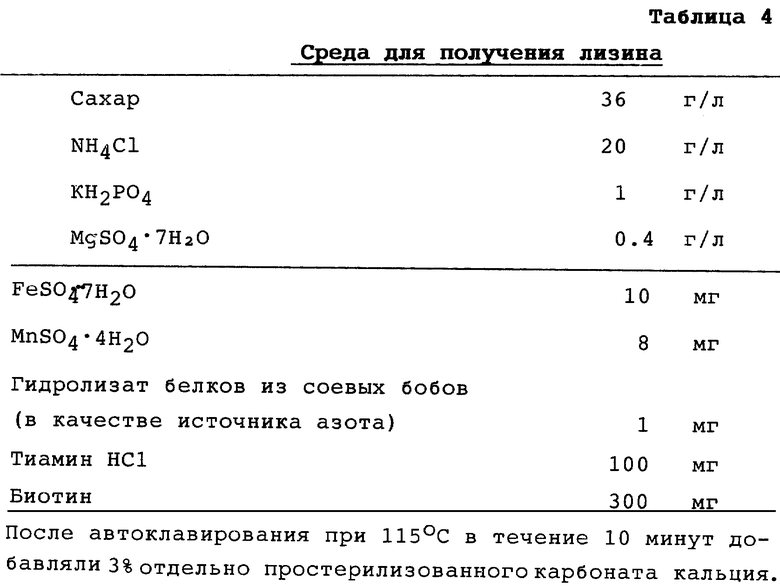
Фиг. 1 показывает изготовление плазмиды pMW::THУ.
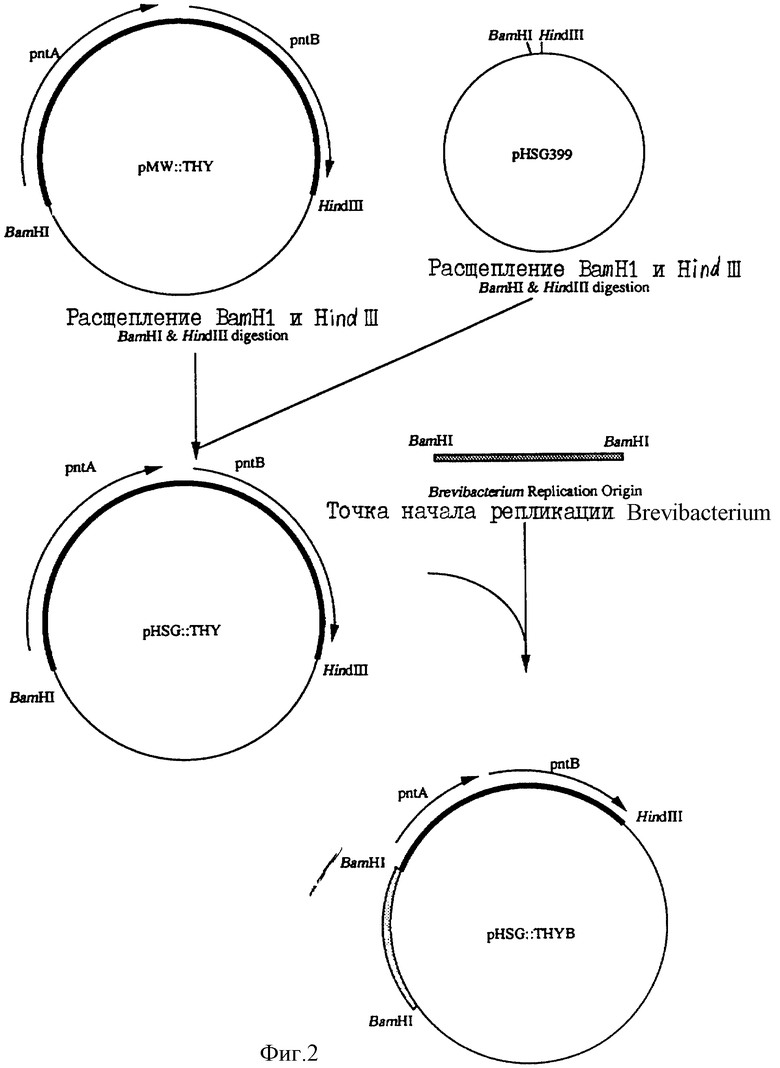
Фиг. 2 показывает изготовление плазмиды рНSG::ТНУВ.
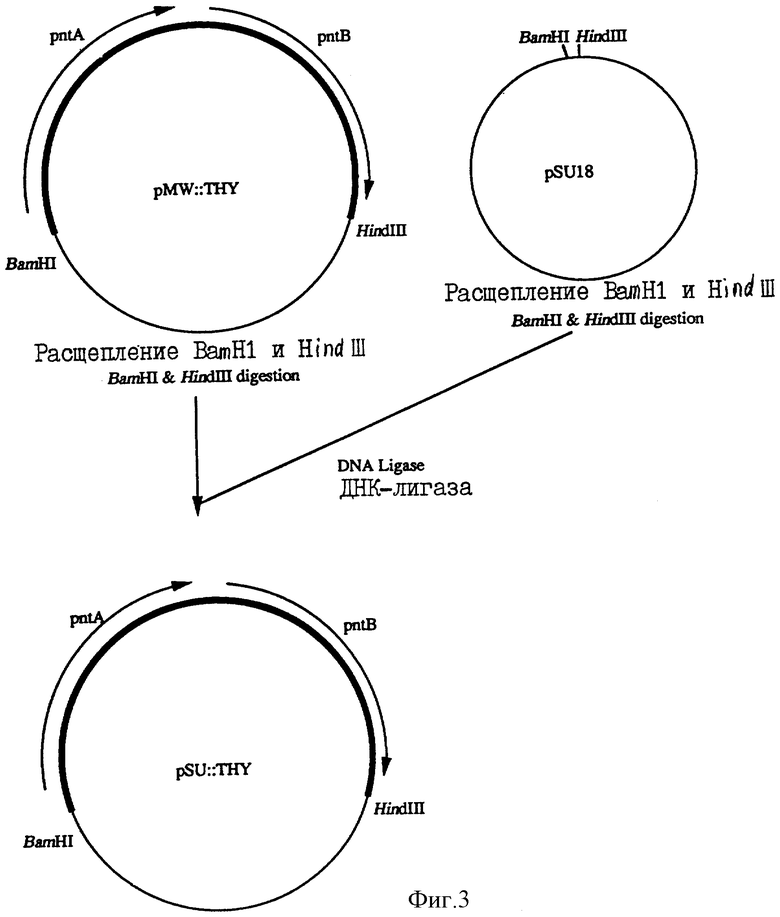
Фиг. 3 показывает изготовление плазмиды рSU::ТНУ.
Настоящее изобретение будет объяснено ниже более конкретно со ссылкой на примеры.
Пример 1: клонирование гена трансгидрогеназы
Нуклеотидные последовательности генов pntA и pntB, которые кодируют трансгидрогеназу у Escherichia coli, были определены предварительно (D.M. Clarke и др. , Eur. J. Biochem., 158, 647-653 (1986)), сообщалось, что pntA и pntB кодируют белки из 502 и 462 аминокислотных остатков, соответственно. В добавление к этому, известно также, что для экспрессии ферментативной активности трансгидрогеназы требуются оба белка, указанные выше, оба гена pntA и pntB расположены на хромосомной ДНК последовательно и благодаря этому оба гена могут быть клонированы на одном фрагменте ДНК.
Для удобства последующих операций авторы настоящего изобретения одновременно клонировали не только оба гена pntA и pntB, но и участок, расположенный перед этими генами, который обладает промоторной активностью. Конкретно, для клонирования фрагмента ДНК, содержащего оба гена и промоторный участок, его амплифицировали при помощи ПЦР.
В соответствии с общепринятым методом, два синтетических олигонуклеотида, имеющих последовательности
5'-ЦТГАТТТТТГГАТЦЦАГАТЦАЦАГ-3' (Посл-ть 1Д :1) и
5'-ЦГТТЦТГТТААГЦТТТЦТЦААТАА-3' (Посл-ть 1Д :2)
были синтезированы в качестве затравок для ПЦР. Тотальная геномная ДНК Escherichia coli K-12 МС1061 была получена в соответствии с методом Saitoh и Miura (Biochem. Biophys. Acta., 72, 619 (1963)) в качестве матричной ДНК для ПЦР. Целевой фрагмент ДНК был амплифицирован путем ПЦР, используя два затравочных олигонуклеотида и матричную хромосомную ДНК в соответствии с методом Erlich и др. (Технология ПЦР, Stockton press (1989)). Синтетические ДНК, использованные в качестве затравок, имели нуклеотидные последовательности, которые немного отличались в соответствующих центральных участках фрагментов синтетической ДНК от нуклеотидной последовательности, сообщенной D.M. Clarke и др. Это отличие спланировано для введения сайта расщепления BamH1 и сайта расщепления HindIII в проект синтетических олигонуклеотидов. Эти сайты расщепления ферментами рестрикции требуются для вставки фрагмента амплифицированной ДНК в векторную ДНК. Введение сайтов рестрикции вызывает несовпадение между затравками и хромосомной ДНК в процессе ПЦР. Однако это несовпадение не влияло на амплификацию ДНК путем ПЦР, так как эти сайты рестрикции были расположены в центральных участках фрагмента синтетической ДНК. Амплификация фрагмента ДНК из 3 т. п.н. была подтверждена электрофорезом на агарозном геле.
Фрагмент ДНК с амплификацией из 3 т.п.н. и плазмидный вектор pMW118, имеющий маркер устойчивости к ампициллину (выпускаемый фирмой "Nippon Gene Inc."), расщеплялись BamHl и HindIII. Расщепленные фрагменты ДНК и векторная ДНК были лигированы с использованием ДНК-лигазы для получения рекомбинантной ДНК. Полученная рекомбинантная плазмида была названа pMW::ТНУ (см. фиг. 1).
Штамм Escherichia coli JM109 (выпускаемый фирмой "Таkara Shuzo Co., Ltd. ") был трансформирован плазмидой pMW::THУ, в результате был получен трансформант Escherichia coli JM109 (pMW::ТНУ). Ферментативная активность трансгидрогеназы, осуществляющаяся в каждом из клеточных экстрактов штаммов Escherichia coli JM109 и Escherichia coli JM109 (pMW::ТНУ), была измерена в соответствии с известным методом (D.M. Clarke и Ph. D. Bragg, J. Bacteriology, 162, 367-373 (1985)). Результаты показаны в таблице 1 (см. в конце описания).
На основании данных таблицы 1 очевидно, что Escherichia coli JM109 (pMW: : THУ) обладает более высокой ферментативной активностью трансгидрогеназы, чем Escherichia coli JM109. Исходя из этого результата было доказано, что фрагмент ДНК, вставленный в плазмиду pMW::THУ, включает ген трансгидрогеназы. Штамм Escherichia coli, содержащий плазмиду pMW::THУ, был обозначен как штамм AJ12929. Штамм AJ12929 помещен на хранение в Национальный институт бионауки и технологии человека, Агенства производственной науки и технологии, Министерства международной торговли и промышленности (1-3, Higashi 1-chome, Tsukuba-shi, Ibaraki-ken, Япония; почтовый код 305) 4 октября 1993 года под номером хранения FERM Р-13890, перенесен из первоначального хранения в международное хранение на основании Будапештского договора от 14 сентября 1994 года и помещен на хранение под номером хранения FERM BP-4798.
Плазмидная ДНК pMW::THУ была приготовлена в соответствии с общепринятым методом и расщеплена BamHl и HindIII для выделения фрагмента ДНК из т.п.н., содержащего ген трансгидрогеназы. Плазмидный вектор pHSG399, имеющий маркер устойчивости к хлорамфениколу (выпускаемый фирмой "Takara Shuzo Co., Ltd."), был расщеплен BamHl и HindIII, и больший его фрагмент был выделен. Затем фрагмент ДНК, содержащий ген трансгидрогеназы, и больший фрагмент BamHl HindIII плазмиды pHSG399 были лигированы с использованием ДНК-лигазы для получения плазмиды pHSG::THУ.
Плазмида pHSG::THУ может автономно реплицироваться в клетках микроорганизмов, принадлежащих к роду Escherichia, но не сохраняется стабильно в клетках коринеформных бактерий. Поэтому в плазмиду pHSG::THУ была введена точка начала репликации, полученная из автономно реплицирующей плазмиды, происходящей из коринеформной бактерии.
Плазмида рНМ1519, способная к автономной репликации в клетках коринеформных бактерий (см. Японский патент (вылож. заявка) 58-77895), была расщеплена ферментом рестрикции BamHl для получения фрагмента из 3 т.п.н., содержащего точку начала репликации. С другой стороны, плазмиду pHSG::THУ расщепили BamHl для получения фрагмента ДНК. Оба фрагмента ДНК были лигированы ДНК-лигазой для изготовления плазмиды pHSG::THУB (фиг.2). Штамм Escherichia coli, содержащий плазмиду pHSG::THУВ, был обозначен как штамм AJ12872. Этот штамм AJ12872 был помещен на хранение в Национальный институт бионауки и технологии человека, Агенства производственной науки и технологии, Министерства международной торговли и промышленности 4 октября 1993 года под номером хранения FERM Р-13889, перенесен из первоначального хранения в международное хранение на основании Будапештского договора от 14 сентября 1994 года и помещен на хранение под номером хранения FERM BP-4797.
Далее, плазмида pSUl8, которая способна к автономной репликации в клетках бактерий, принадлежащих к роду Escherichia, и имеет маркер устойчивости к канамицину (Borja, Bartolome и др.. Gene, 102, 75-78 (1991)), была расщеплена ферментами рестрикции BamHl и HindIII для получения ее большего фрагмента. Этот больший фрагмент был лигирован с фрагментом ДНК из 3 т.п.н., содержащим ген трансгидрогеназы, описанный выше, с использованием ДНК-лигазы для изготовления плазмиды pSU::THУ (фиг.3). Штамм Escherichia coli, содержащий pSU::THУ, был обозначен как штамм AJ12930. Этот штамм AJ12930 был помещен на хранение в Национальный институт бионауки и технологии человека, Агенства селекции промышленных микроорганизмов, Агенства производственной науки и технологии, Министерства международной торговли и промышленности 4 октября 1993 года под номером хранения FERM Р-13891, перенесен из первоначального хранения в международное хранение на основании Будапештского договора от 14 сентября 1994 года и помещен на хранение под номером хранения FERM ВР-4799.
Было подтверждено, каким образом продуктивность по различным L-аминокислотам в клетках бактерии, принадлежащей к роду Escherichia, или коринеформной бактерии была затронута увеличением активности трансгидрогеназы в обеих бактериях.
Пример 2: ферментационное получение L-треонина при помощи штамма с введенной трансгидрогеназой
Среди штаммов Escherichia coli, образующих L-треонин, которые известны в настоящее время, наибольшей продуцирующей способностью обладает штамм В-3996 (см. Японский патент (вылож. заявка) 3-501682 (РСТ)). Поэтому штамм В-3996 был использован в качестве штамма-хозяина для оценки повышения активности трансгидрогеназы. Штамм В-3996 содержит плазмиду pVIC40 (International Publicati on Pamphlet WO 90/04636), которая была получена путем вставки оперона треонина в векторную плазмиду рАУС32 с широким спектром хозяев, имеющую маркер устойчивости к стрептомицину (для сравнения см. Чистосердов А.Ю. и Цыганков Ю.Д., Plasmid, 1986, 16, 161-167). Штамм В-3996 помещен на хранение в Исследовательском институте генетики и селекции промышленных микроорганизмов под регистрационным номером RIA1867.
Плазмида pMW::THУ была выделена из штамма Escherichia coli AJ12929, полученного в Примере 1, при помощи метода Маниатис и др. (Сэмбрук Дж., Фритч И. Ф., Маниатис Т., Молекулярное клонирование. Gold Spring Harbor Laboratory Press, 1. 21 (1898)). Полученную плазмиду rMW::THУ ввели в штамм В-3996 при помощи метода Моррисона (Методы в энзимологии, 68, 326, (1979)). Штамм В-3996, трансформированный плазмидой pMW: :THУ, был обозначен как штамм В-3996(pMW: : THУ). Штамм В-3996 и штамм В-3996(pMW::THУ) культивировали в следующих условиях.
Культивирование проводили в течение 38 часов при температуре 37oС с перемешиванием 114-116 об/мин, используя среду, которая имела состав, представленный в таблице 2. Указанные в таблице 2 компоненты А, В и С готовились отдельно, стерилизовались и охлаждались; затем их смешивали, используя 16/20 объема компонента А, 4/20 объема компонента В и 30 г/л компонента С. Результаты культивирования показаны в таблице 3. Было обнаружено, что продуктивность по L-треонину повысилась путем увеличения внутриклеточной активности трансгидрогеназы у бактерии, образующей L-треонин, которая принадлежит к роду Escherichia.
Пример 3: ферментационное получение L-лизина с помощью штамма с введенной трансгидрогеназой
Была проведена проверка того, повышается или нет продуктивность по L-лизину путем увеличения внутриклеточной активности трансгидрогеназы у коринеформной бактерии, образующей L-лизин. В качестве бактерии, образующей L-лизин и принадлежащей к коринеформным бактериям, использовали штамм Brevibacterium lactofermentum AJ3990. Штамм AJ3990 помещен на хранение в Национальный институт бионауки и технологии человека, Агенства производственной науки и технологии, Министерства международной торговли и промышленности с присвоенным ему номером хранения FERM P-3387. Плазмида рНSG::ТНУВ была выде-лена из штамма Escherichia coli AJ12872, полученного в Примере 1, при помощи метода Маниатис и др. (Сэмбрук Дж., Фритч И.Ф., Маниатис Т., Молекулярное клонирование. Cold Spring Harbor Laboratory Press, 1. 21 (1989)). Полученная плазмида была введена в штамм AJ3390 при помощи метода трансформации с использованием электрического импульса (см. Японский патент (выложенная заявка) 2-207791). Штамм AJ3990, трансформированный плазмидой pHSG::THУB, был обозначен как штамм AJ3390 (pHSG::ТНУВ). Штамм AJ3390 и штамм AJ3990(pHSG::ТНУВ) культивировали в следующих условиях.
Культивирование проводили в течение 72 часов при температуре 31,5oС с перемешиванием 114-116 об/мин, используя среду, которая имела состав, представленный в таблице 4. В качестве сахара были использованы три различных сахара глюкоза, сахароза и фруктоза. Результаты культивирования представлены в таблице 5. Было обнаружено, что продуктивность по L-лизину была повышена путем увеличения внутриклеточной активности трансгидрогеназы у коринеформной бактерии, образующей L-лизин.
Пример 4: ферментационное получение L-фенилаланина с помощью штамма с введенной трансгидрогеназой
Была проведена проверка того, повышается или нет продуктивность по L-фенилаланину путем увеличения внутриклеточной активности трансгидрогеназы у бактерии, образующей L-фенилаланин и принадлежащей к роду Escherichia. В качестве бактерии, образующей L-фенилаланин и принадлежащей к роду Escherichia, использовали штамм Escherichia coli AJ12604. Штамм AJ12604 содержит плазмиду pBR-aroG4, которая была получена путем вставки мутантного гена aroG в векторную плазмиду pBR322, имеющую маркер устойчивости к ампициллину, и плазмиду рАСМАВ, которая была получена путем вставки мутантного гена pheA в векторную плазмиду рАСУС184, имеющую маркер устойчивости к хлорамфениколу (см. Японский патент (вылож. заявка) 5-236947). Штамм AJ12604 был помещен на хранение в Национальный институт бионауки и технологии человека, Агентства производственной науки и технологии, Министерства международной торговли и промышленности 28 января 1991 года под номером хранения FERM P-11975, перенесен из первоначального хранения в международное хранение на основании Будапештского договора от 26 сентября 1991 года и помещен на хранение как номер хранения FERM ВР-3579. Плазмида рSU::ТНУ была выделена из штамма Escherichia coli AJ12930, полученного в Примере 1, при помощи метода Маниатис и др. (Сэмбрук Дж., Фритч И.Г., Маниатис Т., Молекулярное клонирование, Cold Spring Harbor Laboratory Press, 1. 21 (1989)). Полученная плазмида была введена в штамм AJ12604 при помощи метода Д.М. Моррисона (Методы в энзимологии, 68, 326 (1979)). Штамм AJ12604, трансформированный плазмидой рSU::ТНУ, был обозначен как штамм AJ12604 (pSU::ТНУ). Штамм AJ12604 и штамм AJ12604 (рSU:: ТНУ) культивировали в следующих условиях.
Культивирование проводили в течение 16 часов при температуре 37oС с перемешиванием 114-116 об/мин, используя среду, которая имела состав, показанный в таблице 6. Результаты культивирования представлены в таблице 7. Было обнаружено, что продуктивность по L-фенилаланину была повышена путем увеличения внутриклеточной активности трансгидрогеназы у бактерии, образующей L-фенилаланин и принадлежащей к роду Escherichia.
(1) СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
(1) ОБЩАЯ ИНФОРМАЦИЯ:
(i) ЗАЯВИТЕЛЬ; Ajinomoto Co. Inc.
(ii) НАЗВАНИЕ ИЗОБРЕТЕНИЯ: Способ получения веществ
(iii) ЧИСЛО ПОСЛЕДОВАТЕЛЬНОСТЕЙ: 2
(iv) АДРЕС ДЛЯ КОРРЕСПОНДЕНЦИИ:
(A) АДРЕСАТ:
(B) УЛИЦА:
(C) ГОРОД:
(D) ШТАТ:
(Е) СТРАНА:
(F) ПОЧТОВЫЙ КОД:
(vi) ДАННЫЕ ПО НАСТОЯЩЕЙ ЗАЯВКЕ:
(A) НОМЕР ЗАЯВКИ:
(B) ДАТА ПОДАЧИ ЗАЯВКИ:
(C) КЛАССИФИКАЦИЯ:
(vii) ДАННЫЕ ПО ПЕРВОЙ ЗАЯВКЕ:
(А) НОМЕР ЗАЯВКИ:
(В) ДАТА ПОДАЧИ ЗАЯВКИ:
(viii) ИНФОРМАЦИЯ О ПАТЕНТНОМ ПОВЕРЕННОМ:
(A) ИМЯ:
(B) РЕГИСТРАЦИОННЫЙ НОМЕР:
(C) НОМЕР ПО РЕЕСТРУ ДЛЯ ССЫЛКИ:
(ix) ТЕЛЕКОММУНИКАЦИОННАЯ ИНФОРМАЦИЯ:
(A) ТЕЛЕФОН:
(B) ТЕЛЕФАКС:
(2) ИНФОРМАЦИЯ О ПОСЛЕДОВАТЕЛЬНОСТИ 1Д :1:
(i) ХАРАКТЕРИСТИКИ ПОСЛЕДОВАТЕЛЬНОСТИ:
(A) ДЛИНА: 24
(B) ТИП: нуклеиновая кислота
(C) ЧИСЛО НИТЕЙ: однонитевая
(D) ТОПОЛОГИЯ: линейная
(ii) МОЛЕКУЛЯРНЫЙ ТИП: другая синтетическая ДНК
(iii) ГИПОТЕТИЧЕСКИЙ: нет
(xi) ОПИСАНИЕ ПОСЛЕДОВАТЕЛЬНОСТИ: Посл-сть 1Д :1:
ЦТГАТТТТТГ ГАТЦЦАГАТЦ АЦАГ 24
(2) ИНФОРМАЦИЯ О ПОСЛЕДОВАТЕЛЬНОСТИ 1Д :2:
(i) ХАРАКТЕРИСТИКИ ПОСЛЕДОВАТЕЛЬНОСТИ:
(A) ДЛИНА: 24
(B) ТИП: нуклеиновая кислота
(C) ЧИСЛО НИТЕЙ: однонитевая
(D) ТОПОЛОГИЯ: линейная
(ii) МОЛЕКУЛЯРНЫЙ ТИП: другая синтетическая ДНК
(iii) ГИПОТЕТИЧЕСКИЙ: нет
(xi) ОПИСАНИЕ ПОСЛЕДОВАТЕЛЬНОСТИ: Посл-ть 1Д :2:
ЦГТТЦТГТТА АГЦТТТЦТЦА АТАА 24)






Способ предусматривает проведение ферментации с использованием микроорганизма, у которого повышена способность продуцировать в клетке восстановленный никотинамидадениндинуклеотидфосфат. Используют микроорганизмы, относящиеся к Escherichia или Corynebacterium. Указанное свойство микроорганизма обеспечивается в результате целенаправленного повышения активности никотинамиднуклеотидтрансгидрогеназы. Изобретение обеспечивает повышение выхода целевого вещества. 6 з.п. ф-лы, 3 ил., 7 табл.
| Способ получения тетрахлоралкил- и тетрахлоралкенилтрихлорбензолов | 1960 |
|
SU136359A1 |
| ТЕПЛООБМЕННОЕ УСТРОЙСТВО ДЛЯ ПЕЧЕЙ КИПЯЩЕГО СЛОЯ | 0 |
|
SU179338A1 |
| ИЗМЕРИТЕЛЬНОЕ УСТРОЙСТВО ДЛЯ ГЕОЭЛЕКТРОРАЗВЕДКИ ПО МЕТОДУ ПЕРЕХОДНЫХ ПРОЦЕССОВ | 0 |
|
SU379903A1 |
| Eur.J.Biochem, 1986, v.158, № 3, p.647-653 | |||
| Eur | |||
| J.Biochem, 1985, v.149, № 3, p.517-523 | |||
| J.Bacteriol, 1985, v.162, № 1, p.367-373 | |||
| Arch | |||
| Biochem, Biophys., 1976, v.176, № 1, p.136-143. | |||
