Изобретение относится к эпигенетической регуляции in situ биосинтеза белка путем формирования музыкальных последовательностей, соответствующих аминокислотным последовательностям белков, для использования, в частности, в области земледелия и здравоохранения.
Этот способ основан на использовании регулирующего воздействия на биосинтез белков посредством масштабного резонанса, транспозиций в звук временных последовательностей квантовых осцилляций, ассоциированных с их элонгацией. Такое воздействие может способствовать либо увеличению скорости синтеза с одновременной регуляцией его ритма, либо снижению этой скорости, в зависимости от того, используется ли модуляция вибрационных частот в фазе или в противофазе, по отношению к их элонгации (это справедливо для квантовых осцилляций, а также для их транспозиции в звук). Полученный результат стабилизировался воздействием, вновь посредством масштабного резонанса, транспозиций в окрашенный свет групповых квантовых осцилляций, возникающих в результате пространственной конформации белков, образующихся в процессе их элонгации.
Этот способ может быть специфически применен к каждому белку известной структуры. Однако использование этого способа будет тем эффективней, чем более зависим будет синтез этого белка от эпигенетических факторов, то есть, иначе говоря, чем более он будет удален от ДНК-системы, к которой он принадлежит, а в рассматриваемом случае эффективность способа будет зависеть от акустических и электромагнитных факторов. Кроме того, для своей практической реализации этот способ требует оценки метаболического агонизма и антагонизма этих белков вследствие явлений масштабного резонанса, естественным образом связанных с их биосинтезом. Другой отличительной особенностью изобретения является определение этих белков по их связанным метаболическим субпопуляциям (что позволяет определить пределы их метаболической роли с использованием их аминокислотной последовательности).
Идентификация белков, синтез которых регулируется в соответствии с изобретением, включает в себя и другие критерии, такие, как соответствие между акустическими и электромагнитными явлениями, воздействие которых может наблюдаться на живых организмах, и транспонированные белковые последовательности.
Демонстрация музыкальных свойств элементарных частиц (J.Sternheimer, С. R. Acad. Sc. Paris, 297, 829, 1983), а также необходимость в соответствующей теории послужили поводом предположить, что масштаб, при котором происходит это явление, играет важную роль и является независимым измерением по отношению к пространству и времени. Более поздние исследования (J.Sternheimer, Colloque International "Louis Brogle, Physicien et Penser", Ancienne Ecole Polytechnique, Paris, November 5-6, 1987) привели к заключению о физическом существовании квантовых волн, связанных с частицами и распространяющихся сами по себе, не только в пространственно-временных координатах, но также и в масштабном измерении, связывая таким образом последовательные уровни организации материи. Эти волны, описываемые уравнениями распространения, способствуют воздействию одного масштаба на другой между явлениями, которые являются достаточно схожими для того, чтобы с математической точки зрения составлять гармоники общего основного тона (J.Sternheimer, Ondes d'echelle, I. Partie Physique, 1992, II.Partie Biologique).
Теоретическое обоснование существования масштабных волн, а также экспериментальное подтверждение этого факта, сделанное на основании различных свойств этих волн, позволяют рассматривать масштабные волны как универсальное явление, функция которых заключается прежде всего в обеспечении когерентности между различными масштабами квантовой системы, и эта функция принимает форму и может быть описана в процессе биосинтеза белков. Удлинение пептидной цепи происходит в результате последовательного присоединения аминокислот, которые были введены в рибосому посредством специфического переноса РНК (тРНК). Если аминокислота, будучи изначально в свободном состоянии, начинает сама присоединяться к своей тРНК, то в этом случае она уже является достаточно стабилизированной в отношении интенсификации теплового движения; и в то же время эта аминокислота сохраняет относительную автономию, поскольку она связана с тРНК лишь одной степенью свободы. Что же касается ее волны де Бройля, то она достигает величины порядка размера этой аминокислоты. Это придает ей волновые свойства и интерференцию между масштабной волной, связанной с этой аминокислотой, и масштабными волнами, аналогичным образом сформированными другими аминокислотами, приводит к синхронизации (спустя очень короткий промежуток времени порядка 10-12,5 с) собственных частот, связанных с этими аминокислотами, в соответствии с одним и тем же музыкальным строем, который, таким образом, будет с более высокой точностью зависеть от популяции переносимых РНК. Однако в пределах аппроксимации темперированного строя этот строй является универсальным благодаря, в частности, очень специфическому распределению масс аминокислот, которое является уже очень близким этому строю. (Аналогичным образом ДНК-нуклеотиды настраиваются на тот же музыкальный строй, как можно легко видеть по их массам.)
Рассматриваемое явление может быть представлено более наглядно, если тРНК, несущая аминокислоты, сама прикрепляется к рибосоме. Именно в этот момент, то есть до переноса, благодаря которому она закрепляется на пептидной цепи, стабилизация по отношению к тепловой интенсификации становится такой, что длина волны аминокислоты перерастает ее размер на целый порядок его величины. Испускаемая затем масштабная волна интерферирует, в масштабе белка в структуре, с аналогичными волнами, ранее сформированными другими аминокислотами. Это приводит к ограничениям "музыкального типа" для временной последовательности собственных частот, связанных с этими волнами, так что масштабные волны могут (если обобщить предыдущую ситуацию) продолжать свой путь, обеспечивая тем самым когерентность и связь между различными уровнями организма, например лишь последовательность этих волн имеет значение для минимизации диссонанса (гармонический интервал) и частотных интервалов (представляемых мелодическим интервалом) между последовательными аминокислотами; и даже более того, поскольку каждая масштабная волна представляет собой суперпозицию волн, связывающих два данных уровня (а значит, и уровни каждой аминокислоты с уровнем белка), которая в два, три и т.д. раза превышает самую быструю волну, то это свидетельствует о существовании периодов минимизации именно гармонических интервалов, указывая на интервалы во временной последовательности частот; причем другие уровни будут завершаться корреляциями, гораздо более многочисленными и заметными, и сами будут оказывать влияние на синтез белков. В соответствии с этим имеются основания предположить, что белки должны обладать (соответственно последовательности собственных квантовых частот, связанных с последовательностью их аминокислот) музыкальными свойствами, которые будут тем точнее и тоньше, чем чувствительнее будет их биосинтез к эпигенетическим факторам в целом; и наоборот, они должны иметь возможность действовать на биосинтез эпигенетически и специфическим образом для каждого белка.
Наблюдения белковых последовательностей, описанных в литературе (M.C. Dayhoff, Atlas of protein sequence and Structure, v. 5 and supplement, N.B. R. F. (Washington) 1972-73), позволяют подтвердить вышеуказанные факты. Не только все белки обладают музыкальными свойствами в последовательности своих аминокислот, но и сами свойства проявляются тем больше, чем эпигенетически более чувствительными являются белки. Кроме того, акустическая транспозиция серий собственных частот, соответствующих формированию масштабных волн в фазе с удлинением данного белка, оказывает стимулирующее действие на биосинтез этого белка in vivo и коррелированным образом, наоборот, оказывает подавляющее действие на масштабные волны в противофазе. Эти действия, которые воспроизводят в соответствии с данным масштабом аналогичные действия, происходящие уже в квантовом масштабе между белками в процессе их синтеза (а это значит, что они играют важную роль в их метаболизме, то есть белки, которые являются музыкально схожими, являются тем самым метаболическими агонистами), наблюдаются фактически у всех живых организмов, которые чувствительны к звуковым колебаниям.
В случае животных, обладающих нервной системой (по крайней мере для животных, у которых фактически наблюдались "микрофонные потенциалы", точно воспроизводящие форму приложенной волны), можно дать следующее описание рассматриваемых явлений: звуковая волна преобразуется в электромагнитные импульсы той же частоты непосредственно из начальной точки слухового нерва. Эти импульсы благодаря масштабной инвариантности масштабно-волновых уравнений, применяемых к фотону (обобщенные уравнения Максвелла), оказывают затем непосредственное действие путем масштабного резонанса на их квантовые транспозиции; и поскольку квадраты квантовых амплитуд пропорциональны числу одновременно синтезируемых белков, то явление резонанса в случае масштабных волн в фазе приводит к увеличению скорости синтеза и к регулированию его ритма, а в случае противофазы - к уменьшению этой скорости. Как можно заметить, именно благодаря тому, что микрофонные потенциалы имеют место в слуховом нерве до начала реального нервного импульса (P.Buser и M.Imbert, Audition, Hermann ed., Paris, 1987), механизм, рассматриваемый в изобретении, не требует на этой стадии мозгового анализа этих импульсов. Что касается растений, то их (механическая) чувствительность к звукам достаточно заметна (с помощью интерферометрии), и теоретически масштабные волны ведут себя в растениях аналогичным образом.
Решение уравнения масштабных волн, которое фактически свидетельствует о существовании масштабных волн, имеющих диапазон, близкий числу Авогадро (как и в случае вышеупомянутых транспозиций), также дает основание ожидать аналогичных свойств для масштабных волн, распространяемых в результате пространственного распределения аминокислот (волна де Бройля которых сравнима с их размером) внутри белка после его синтезирования (но на этот раз с диапазоном, аппроксимирующим квадратный корень из этого числа); например, наблюдения их структур третьего порядка подтверждает существование гармонии в пределах вибрационных частот аминокислот, пространственно близко расположенных внутри белка (особенно у их поверхности, что можно ожидать исходя из их длины волны); и в то же время можно наблюдать посредством цветных транспозиций этих частот заметную стабилизацию эффектов, полученных с использованием музыкальных транспозиций.
Задачей изобретения является создание способа формирования музыкальных последовательностей, соответствующих аминокислотным последовательностям белков, обеспечивающего повышение эффективности регулирования биосинтеза белков.
Указанный результат достигается тем, что в способе формирования музыкальных последовательностей, соответствующих аминокислотным последовательностям белков, в соответствии с изобретением декодируют и транспонируют в звук временные ряды квантовых осцилляций, связанных с их элонгацией, для эпигенетического регулирования биосинтеза этих белков посредством масштабного резонанса, при этом упомянутое декодирование осуществляют в виде последовательности следующих четырех этапов, причем на первом этапе каждую аминокислоту данного белка ассоциируют с основным тоном, частота которого определяется с использованием кода, полученного исходя из собственных частот аминокислот в их свободном состоянии, пропорциональных их массам, путем минимизации общего гармонического расстояния между частотами этих аминокислот для всех возможных пар аминокислот с учетом их соотношений в окружающей популяции транспортных РНК, при условии, что смещение начальной частоты в сторону синхронизованного значения составляет менее чем половину интервала между двумя синхронизованными частотами по обе стороны от указанной начальной частоты, и последующей транспозиции полученных таким образом частот в область звуковых частот, при этом полученный код используют для стимуляции биосинтеза данного белка, а другой код, который соответствует ингибированию белка, получают из вышеуказанного кода путем симметризации логарифмов полученных частот по отношению к их центральному значению, рассматриваемому как начальное значение. На втором этапе определяют музыкальные периоды путем распознавания сходных последовательностей. На третьем этапе определяют длину нот путем совместного уточнения периодов, полученных отдельно на втором этапе, осуществляемого посредством подстройки фразировки к определенному размеру. На четвертом этапе определяют качество тона посредством обратного действия всех аминокислот целого белка на гармоническую структуру каждой из них, осуществляемого путем подстрой распределения тонов белка к среднему распределению этих тонов для целого белка, в результате чего определяют, какие гармоники должны быть усилены, а какие снижены для того, чтобы получить качество звука, соответствующее данному белку, и в соответствии с этим получают музыкальные последовательности, обеспечивающие регулирование биосинтеза белка.
При этом код, полученный на первом этапе, предназначенный для стимуляции биосинтеза белка и универсальный в пределах аппроксимации темперированного звукоряда, будучи ассоциированным с различными аминокислотами, состоит из следующих основных тонов: Gly = низкое A; Ala = C; Ser = E; Pro, Val, Thr, Cys = F; Leu, Ile, Asn, Asp = G; Gln, Lys, Glu, Met = A; His = B-бемоль, Phe, a также Sec = B, Arg, Tyr = C-диез; Trp = D-диез, а код, полученный на первом этапе, предназначенный для ингибирования и универсальный в пределах аппроксимации темперированного звукоряда, получают из тонов темперированного ряда, являющихся симметричными по отношению к тонам кода, предназначенного для стимуляции биосинтеза, относительно их центрального значения G.
Кроме того, каждую звуковую транспозицию квантовых осцилляций, ассоциированных с биосинтезом данного белка, дополняют цветовой транспозицией квантовых осцилляций, ассоциированных со зрелым белком, после пространственного складчатого сворачивания этого белка в соответствии с кодом, предназначенным для стабилизации этого белка или ингибирования его биосинтеза и полученным с помощью сформированной музыкальной последовательности, причем указанный код формируют из кода, полученного на первом этапе, с использованием следующей формулы:
ν = νoArgch(e(f/fo)Logch1),
где f,fо - музыкальные частоты;
ν,νo - частоты цветов;
индекс "о" обозначает центральные значения;
Argch - инверсная функция гиперболического косинуса;
chl - гиперболический косинус единицы, причем код, предназначенный для стабилизации белков, стимулированных сформированными музыкальными последовательностями, с использованием кода, предназначенного для стимуляции биосинтеза белка, будучи ассоциированным с различными аминокислотами, состоит из следующих цветов: Gly = темно-красный; Ala = ярко-красный; Ser = оранжевый; Pro, Val, Thr, Cys = коричнево-желтый; Leu, Ile, Asn, Asp = лимонно-желтый; Gln, Glu, Lys, Met = зеленый; His = изумрудный; Phe = синий; Arg, Tyr = индиго; Trp = пурпурный.
При этом предпочтительно осуществляют запись сформированных музыкальных последовательностей и/или транспонированных цветов соответственно в форме нотной записи, цветовой интерпретации и/или звукозаписи на соответствующих носителях.
Кроме того, в способе характеризации белковых последовательностей, на основе определения эпигенетических взаимодействий данного белка с другими белками, для их регулирования, согласно изобретению, используют любые упомянутые записи, при осуществлении декодирования вышеописанным способом для выявления музыкального сходства или различия белков относительно данных белков, гармонические суперпозиции с другими белковыми мелодиями либо комбинацию этих факторов, на основании чего делают выводы относительно агонизма или антагонизма данных белков с другими белками.
При этом характеризацию регулируемых белковых последовательностей уточняют путем биполярного дифференциального сравнения с положительными или отрицательными эффектами, полученными с использованием упомянутых записей, или путем идентификации с использованием выявления музыкального сходства или различия белков, обусловивших получение положительных или отрицательных эффектов, связанных с акустическими или электромагнитными явлениями, формирующими различные серии частот.
Для декодирования белков в соответствии с изобретением использовался следующий способ.
1. Серии частот определялись следующим образом: каждой аминокислоте соответствует основной тон, точная частота которого определялась из соответствующих частот аминокислот в их свободном состоянии (пропорциональных их массам) путем минимизации глобального гармонического расстояния ΣijPiPj logsup(pi,qj), вычисленного для всех возможных пар нот, где (pi/qj) - гармонические интервалы, наиболее близкие соответствующим отношениям собственных частот с учетом их соответствующих соотношений pi, pj в окружающей популяции транспортных РНК; при условии δf < Δf/2, где δf - смещение начальной частоты в сторону синхронизированного значения, а Δf - интервал между двумя синхронизированными последовательными частотами полученной гаммы, которая включает в себя указанную начальную частоту, и с последующей транспозицией этих частот в поле звуковых частот (в соответствии со способом, описанным в патенте Франции N 8302122).
Таким образом, в пределах аппроксимации темперированного строя был получен универсальный код для стимуляции синтеза белков: Gly = низкое ля (A); Ala = до (C); Ser = ми (E); Pro, Val, Thr, Cys = фа (F); Leu, Ile, Asn, Asp = соль (G); Gln, Glu, Lys, Met = ля (A); His = си-бемоль (B); Phe, так же, как SeC, = си (B); Arg, Tyr = до (C); Trp = ре-диез (D-диез) и другой код для их ингибирования (подавления), который был выведен из предыдущего кода путем симметризации логарифмов частот относительно их центрального значения: Trp = до (C); Arg, Tyr = ре (D); Phe, SeC = ми-бемоль (Е-бемоль); His = ми (E); Gln, Glu, Lys, Met = фа (F); Leu, Ile, Asn, Asp = соль (G); Pro, Val, Thr, Cys = ля (A); Ser = си-бемоль (В-бемоль); Ala = ре-диез (D-диез); Gly = фа-диез (F- диез); причем применение этих кодов в процессе синтеза белков дает масштабные волны в фазе и в противофазе по отношению к волнам, формируемым в процессе синтеза, соответственно.
Термин "универсальный код" означает, что этот код является идентичным для всех белков в пределах аппроксимации темперированного строя; низкое A (для центральной частоты на 76 октав ниже центра тяжести начальной частоты лейцина, изолейцина и аспарагина) составляет 220 Гц. Определение гармонического интервала, приведенное выше, раскрывает и развивает определение, предложенное Y.Hellegouarch С.R.Math.Rep.Acad.Sci., Canada, т.4, c. 227, 1982). Более конкретно точные значения зависят от соотношения групп вышеупомянутых аминокислот среди переносимых тРНК-популяций, находящихся рядом во время синтеза белка; и эти значения могут быть вычислены для каждой конкретной ситуации.
2. Определение периода (или периодов) молекулы осуществлялось следующим образом.
Само существование таких периодов является прямым результатом существования масштабных волн. О наличии по меньшей мере некоторых из них свидетельствует присутствие очевидных каденций (таких, как CG, F-S, то есть после F непосредственно следует S; каденция, заканчивающая пептидный сигнал, если он присутствует (для стимуляции); серии R или Y (для ингибирования), за исключением соответствующих пауз, индуцированных гармоническими вариациями, которые в противном случае были бы слишком простыми; и во всех случаях каденции, выражающие возврат к тонике), формирующих "пунктуацию" музыкального развития. Затем более точно определяются сходные пассажи, либо путем прямого повторения нот (в этом случае период определяется путем простого вычисления автокорреляций нот, либо более точно путем минимизации частотного различия между нотами с помощью числа, которое минимизирует среднее для белка от мелодических интервалов между нотами, расположенными друг от друга на целое число интервалов), либо путем мелодического движения (период тогда определяют вычислением автокорреляцией сигнатур - или знаков частотных вариаций - от одной ноты к следующей ноте); либо более точно путем вычисления автокорреляции мелодических интервалов от одной ноты до другой с учетом знаков этих интервалов, т. е. умноженных на соответствующие сигнатуры, либо еще более точно с помощью числа, которое минимизирует методом постепенного приближения среднее для белка вариаций мелодических интервалов, до целого числа интервалов; при этом повторение мелодических контуров может быть уточнено путем вычисления автокорреляций пар или даже предпочтительнее триплетов сигнатур), либо с помощью логики гармонического движения, которое воспроизводит ноты или мелодическое движение в ближайшей простой гармонической транспозиции (в основном на октаву, квинту, или кварту, при этом период определяется числом, которое минимизирует среднее для белка значение гармонических расстояний между нотами, разнесенных друг от друга на целое число интервалов). Иногда также, если имеется возможность провести сравнительный анализ первичной структуры сходных последовательностей (особенно между различными образцами), период проявляется в добавлениях или исключениях некоторых из этих последовательностей. В результате должна получиться мелодически и гармонически когерентная последовательность. Для того чтобы получить такую последовательность, учитывался тот факт, что последние ноты каждого периода или части фразы (обычно второй половины, а предпочтительно последняя нота), также приходящиеся на сильную долю, являются наиболее важными для этой последовательности. Конечный результат тогда наиболее значителен с учетом всех указанных критериев; причем обычно, один из них является более значимым, чем другие, как это имеет место при определении путем расчета пространственной укладки молекул; но тем не менее случаи, аналогичные аллостерии, существуют и имеют биологическое значение (стимуляция или ингибирование такой молекулы или других молекул в процессе метаболизма), но более часто влияют на положение тактов, а не на период.
3. При необходимости можно уточнить тот или иной период, в частности, так, чтобы мелодические пассажи, которые являются связанными (то есть которые повторяются или следуют друг за другом), могли находиться в одном и том же месте внутри такта; это обеспечивается с использованием длительности нот. Эту операцию по подстройке фазировки к определенному размеру можно сравнить с хорошо известным явлением удлинения гласных вокального текста.
На практике операции, описанные в пунктах 2 и 3, могут быть легко осуществлены с помощью клавиатуры, такой, как CasioTM, снабженной устройством "игры в одном ключе", либо с помощью компьютера со специально разработанной для этой цели программой, в памяти которой предварительно заложена последовательность нот, полученная, как описано в пункте 1, и которая позволяет затем непосредственно проиграть эту последовательность нот, что дает возможность контролировать и корректировать эти операции. Однако эти методы требуют некоторой предосторожности. В частности, при декодировании одинаковых молекул или молекул, музыкально схожих друг с другом, в направлении ингибирования (или в любом направлении, противоположном исходному), следует принять во внимание тот факт, что очень часто молекулы имеют преимущественное направление декодирования; в частности, для пар молекул, которые явно обладают одними и теми же свойствами, следует найти молекулу, наиболее музыкальную в отношении ингибирования, и другую молекулу, наиболее музыкальную в отношении стимуляции (особенно в случае иммунного и аутоиммунного метаболизма). В этом случае присутствие и распределение каденций (которые отличаются по стимуляции и ингибированию) обычно позволяют распознавать эти молекулы с самого начала и, следовательно, сохранить.
4. Ритмический стиль контролируется посредством распределения оснований ДНК: сначала с помощью их автокорреляций (если молекула является достаточно музыкальной, то период этих автокорреляций соответствует периоду данного белка; в этом случае определяют размерные такты, последовательности кодонов оснований - или более точно, оснований в третьем положении этих кодонов, - для которых пики автокорреляций являются наиболее высокими, что соответствует наиболее акцентированным нотам), а затем путем сравнения с кодонами известных молекул (уже декодированных или более регулярных во избежание излишней трудоемкости процедуры), имеющих тот же самый предполагаемый ритмический стиль, т. е. стиль музыкального ритма (который при ограничении акцентирования нот влияет на выбор оснований в третьем положении), определяет (по меньшей мере приблизительно) совершенно однозначно использование кодона, поскольку молекулы одного и того же стиля должны (вполне определенно) использовать один и тот же кодон. При необходимости соотносительно корректируется декодирование некоторых пассажей.
5. Затем определяется тембр звука. В основном он отличается для каждой молекулы и в любом случае для каждого распределения нот. Теоретически тембр звука зависит главным образом от самой молекулы, однако он может также зависеть и от всех уровней организма, оказывающих обратное действие на гармоническую структуру аминокислотных вибраций. В первом приближении его определяют путем приведения распределения основных тонов молекул в соответствие с теоретической кривой распределения (которая может быть получена из уравнения масштабных волн, а также по наблюдениям, полученным в среднем на весь белок). Это дает возможность выяснить (как в патенте Франции N 8302122), какие именно гармоники усиливаются, а какие смягчаются в нужном тоне. Затем выбирается наиболее близкий тембр тона из данной палитры. Для большей точности следует выделить три ситуации:
распределение нот постоянно по всей молекуле (в этом случае имеется относительно стационарная гармоническая структура);
постоянные изменения в распределении (в этом случае имеются различные последовательные тоны инструмента, например, для цитохрома C, с несколькими регистрами органа);
прогрессирующие изменения в распределении (в этом случае распределение воспроизводит временное развитие гармонической структуры одного основного тона, например, для миозина, где это развитие явно иллюстрирует тембр звука трубы).
Что касается определения темпа, то оно не связано с какими-либо техническими проблемами и непосредственно вытекает из определения ритмического стиля, как описано выше. Причем в основном, чем быстрее темп, тем большее значение имеет избыточность в белковой последовательности, как в случае фибриллярных белков.
6. Затем определяется цвет путем использования кода (который в первом приближении также является универсальным и который был получен, исходя из вибрационных частот отдельных аминокислот) по следующей формуле (на основании теории масштабных волн) :
ν ≅ νoArgch(e(f/fo)Logch1),
где f, fо - собственные квантовые частоты, связанные с аминокислотами,
ν,νo - частоты цветов, где индекс "о" означает центральные значения.
Это уравнение дает возможность вывести код, связанный со стабилизацией белков, синтезированных in situ (этот код, связанный со стабилизацией их ингибирования, выведен, как описано в пункте 1, путем симметризации логарифмов частот по отношению к центральному желто-лимонному цвету): Gly = темно-красный; Ala = ярко-красный; Ser = оранжевый; Pro, Val, Thr, Cys = коричневато-желтый; Leu, Ile, Asn, Asp = лимонно-желтый; Gln, Glu, Lys, Met = зеленый;
His = изумрудный; Phe = синий; Arg, Tyr = индиго; Trp = пурпурный.
Эти частоты затем смещаются в направлении к красному или пурпурному цвету в соответствии с общим перераспределением молекулярных частот аналогично тому, как это описано выше для тембра звука. В этом случае пространственное расположение цветов аналогично расположению аминокислот в трехмерном пространственном представлении молекул.
Примеры
Ниже представлены примеры "музыкального" и "цветного" декодирования последовательностей белка. (В этих примерах использованы однобуквенные обозначения аминокислот: Gly = G, Ala = A, Ser = S, Pro, Val, Thr, Cys = P, V, T, C соответственно; Leu, Ile, Asn, Asp = L, I, N, D; Gln, Glu, Lys, Met = Q, F, K, M; His = H; Phe = F; Arg, Tyr = R, Y; Trp=W).
1. Пример белка, регулярного от начала до конца.
После развернутого анализа первичной структуры белка, который был особенно хорошо изучен (цитохром C), было установлено, что у белков животного происхождения по сравнению с растительными белками наблюдается постоянная делеция восьми аминокислот (иногда семь). Наблюдение автокорреляций тонов и мелодических контуров подтвердило первоначальное значение музыкального периода. Действительно, если подсчитать встречаемость одной и той же ноты, а также одно и то же направление изменения высоты тона, имеющее место три раза в ряду (один и тот же триплет сигнатур), которые отстоят друг от друга на целое число k нот, то получим следующий результат:
Значение k 1 2 3 4 5 6 7 8 9 10 11 12
Автокорреляция тона 19 15 15 20 19 15 17 21 14 17 18 13
Автокорреляция мелодического контура 1 7 4 6 5 10 8 13 5 4 4 4
Всего 20 22 19 26 24 25 25 34 19 21 22 17
где пик при k = 8 имеет значение около 2,5 стандартных отклонений (по сравнению с ожидаемой величиной 22,3 ± 4,7, определенной из распределения тонов молекулы). Значимость этого пика еще больше увеличивается при использовании мелодических интервалов, как описано в пункте 2, превышая 3 стандартных отклонения, если применять автокорреляции мелодических интервалов, принимая за величину мелодического расстояния между двумя нотами абсолютное значение разницы порядковых номеров их темперированных частот, расположенных в восходящем порядке на шкале, полученной в пункте 1. Это определение дается, исходя из стандартной номенклатуры, принятой для нот, а именно: вторая, третья и т.п. Вторичный пик при k = 7 является низко значимым, что соответствует относительной длительности седьмой ноты, которая тяготеет к тонике, а пик при k = 4 усиливается, если использовать гармонические интервалы, как описано в пункте 2, поскольку он соответствует пространственной укладке молекулы.
Наблюдение каденций также подтверждает это значение, а также внутреннюю схожесть (так, например, последние пять нот первой, второй и третьей группы из восьми вместе дают точную гармоническую суперпозицию, т. е., иначе говоря, канон для трех голосов). Более конкретно эти две последние оценки показывают прежде всего более высокую относительную важность седьмой (F-S каденция во втором периоде) и восьмой нот (возврат к тонике A-минора) для каждого периода; причем последняя нота доминирует над седьмой (совершенная каденция F-Q на шестнадцатой ноте доминирует над предыдущей F-S- каденцией с восстановлением первоначальной тональности). Деление периодов (как можно более экономичное, если принять во внимание предыдущие ограничения) дает шесть шестнадцатых, одну восьмую, одну четвертую (что означает относительную длину 1-1-1-1-1-1-2-4 с ритмом 6:8; см. фиг.1). Следует отметить, что при когерентности мелодической прогрессии (которая в основном дает наблюдаемую регулярность), а также при обогащенности гармонической прогрессии, A - минорная тональность сопровождается модуляциями в E - минор (второй такт), G - минор (восьмой такт) и F - мажор (третий и девятый такт).
Если рассматривать распределение ДНК-оснований, то можно увидеть, что первая и седьмая ноты каждого периода явно благоприятствуют аденину и тимину в третьем положении соответственно, а третья и восьмая ноты благоприятствуют цитозину и гуанину. Кроме того, в соответствии с вышеуказанным делением для периода и относительной длиной нот (т. е. седьмая и восьмая ноты имеют длительность, соответственно в два и в четыре раза превышающую длительность первой ноты), было показано, что в AT-обогащенном участке сильные доли приходятся на первую и седьмую ноты, а поэтому такты начинаются с первой ноты, а в CG-обогащенном участке музыкальная последовательность начинается с затакта (поскольку сильная доля приходится на третью и восьмую долю, то основной такт размера начинается с третьей ноты). Отсюда можно сделать вывод, что этот белок должен обладать различными метаболическими функциями в зависимости от его окружения. И действительно, диапазон метаболического действия белка был продемонстрирован степенью его музыкального развития (например, по сравнению с последовательностью Euglena gracilis, где в трех первых тактах можно наблюдать 56%-ное повышение мелодического уровня (регулярности) и 16%-ное повышение гармонического уровня (регулярности), определенного в результате минимизации соответственно мелодических и гармонических интервалов между последовательными нотами). При этом исследования музыкальной схожести с другими белками показали, с одной стороны, возможность наложения цитохрома C на эндозепин, где музыкальный кадр считывания совпадает с тактом, приходящимся на первую ноту, что характерно для AT-обогащенной молекулы, и указывает на антидепрессивную роль цитохрома (и его музыкальный строй) посредством фактического дезингибирования нейротрансмиссии; и с другой стороны, музыкальную связь (начало с затакта) с цитохром-оксидазой, для которой характерна CG-обогащенность и которая заканчивает респираторную цепь, что указывает на другую метаболическую роль цитохрома C, расположенного непосредственно перед цитохром-оксидазой в этой цепи.
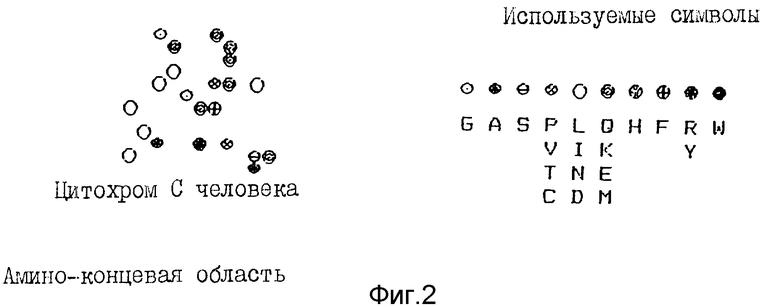
Что касается звукового тембра, то в рассматриваемой A - минорной тональности почти что отсутствие кварты (D) и относительная слабость квинты (E) по сравнению с явной доминантностью тоники и излишком октавы (низкое A - среднее A) ведет к преобладанию гармоник 1 и 2 в ущерб последующим, что придает звуку органный тембр со слегка отличающимися регистрами в соответствии с конкретными пассажами. И наконец, цвета эффективно группируются, образуя цветную окраску на зрелом белке (см. фиг. 2), и продуцируя, как в случае музыки, заметные гармонические отклики. (Следует заметить, что цветное определение использовали для подтверждения музыкального декодирования, поскольку некоторые автокорреляции нот транслируются не в музыкальный период, а в пространственную укладку молекулы: в этом случае их необходимо в итоге вычесть, если хотим определить таким способом музыкальные периоды; при этом вторичный пик таких автокорреляций (k = 4 благодаря α - спирали начала, показанного на фиг. 2) соответствует пространственной укладке белка. И наоборот, музыкальное декодирование может дать информацию относительно пространственной структуры белка).
2. Пример контроля декодирования белка, обнаруживающего ритмические вариации.
Декодирование белка может быть проконтролировано на различных уровнях, включая декодирование молекул, известных как метаболические агонисты, и включая выводы, которые можно сделать относительно метаболического уровня, исходя из наблюдаемых музыкальных соответствий. Таким образом, можно шаг за шагом восстановить крупные участки метаболизма в молекулярном масштабе. Как можно видеть из предыдущего примера, для облегчения декодирования "ритмическая формула" цитохрома C может быть транскрибирована следующим образом: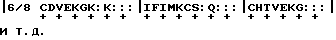
где "+" обозначает сильные доли; обозначает тактовую черту;
обозначает тактовую черту;
":" указывает длительность нот.
И напротив, в субъединице III цитохром-оксидазы, которая музыкально связана в одну цепь с цитохромом C, начало ясно представляет собой формулу четырехдольного размера, как легко видеть из внутреннего сходства (так, например, ноты 7-22, которые по своему стилю напоминают Баха, разделяются на группы из четырех нот, каждая из которых накладывается на последующую). После десятого такта имеется другой такт, который не только накладывается во всех его сильных долях на первый такт цитохрома C, но фактически даже идентичен третьему такту этого цитохрома.
В соответствии с этим можно сказать, что длительности от восьмого такта и дальше (каденции, наблюдаемые в конце этого такта, говорят сами за себя) выражают шестидольный размер (фиг. 1):
Такое изменение ритма (от 4/8 до 6/8) также хорошо прослеживается на автокорреляциях оснований ДНК, где на этой стадии явный пик наблюдается от четвертого до шестого триплета оснований (несмотря на то, что трехдольный ритм оснований, который обычно превалирует в автокорреляциях оснований кодирующих частей ДНК, в данном случае является несколько менее выраженным). (На фиг. 1 начало последовательности приходится на затакт, что, как показано выше, особо выделяет сильную долю на третьей ноте ввиду того, что к цепи присоединен CG - обогащенный ритмический вариант цитохрома C.)
3. Пример восстановления метаболической цепи, включая стимуляцию и ингибирование. В данном разделе приводится другой пример постепенного восстановления метаболической цепи. Декодирование гистона 4 осуществляется особенно легко: период 7 особенно отчетливо прослеживается в начале последовательности молекулы; повторение G внутри двух аминокислотных интервалов указывает на двудольный ритм; а каденции GS, которыми заканчиваются два первых периода, однозначно указывают на четырехдольный ритм: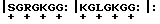
Эта картина продолжается вплоть до конца последовательности, за исключением последнего такта, где используется синкопа (посредством внутреннего повтора) для возвращения к ритму первых двух тактов (фиг.3). Общее распределение нот указывает на гармоническую структуру, соответствующую звуку флейты, а "скачки нот", повторяющиеся в начале, позволяют предположить динамическое напряжение звука типа атаки и даже определить тембр звука, похожий на звук флейты Пана.
Гистон 4 является одним из наиболее консервативных белков, встречающихся в природе. Однако это не означает, что его метаболическое действие, которое, очевидно, является главным, не нуждается иногда в темперировании; так, например, тема двух первых его тактов появляется снова - при ингибировании и транспозиции на кварту, - в консервативной части начала последовательности халконсинтазы, которая представляет собой фермент, ответственный за пигментацию многих цветущих растений (фиг. 3). Его функцию можно сравнить с предполагаемой ролью хроматина (частью которого является гистон 4) в процессе фиксации магния, т. е. весной, когда растения нуждаются в большом количестве магния для фотосинтеза, его фиксацию необходимо стимулировать (например, пением птиц, вызывающим развитие этой темы молекулы); и в этом случае халконсинтаза ингибируется, тогда как осенью более слабая стимуляция гистона дезингибирует халконсинтазу, что приводит к смене зеленой окраски листьев на характерные краски этого сезона. Это явление становится более понятным благодаря описанному эпигенетическому фактору.
Практически при прослушивании музыкальной транспозиции гистона 4 у слушателей возникало ощущение о их "принуждении съесть шоколад", который содержит магний. Это явление дискомфорта, очевидно, возникает у людей, имеющих более или менее высокий уровень холестерина в крови. И кроме того, музыкальное декодирование халконизомеразы (метаболического агониста халконсинтазы, но "музыкально более эффективно" при стимуляции) включает в себя серии тем и вариаций, последовательность которых репродуцирует (у цветущих растений) темы регулирования полного метаболического пути холестерина у человека: т. е. прослушивание этого антагониста гистона 4 "второй степени" позволяет (в соответствии с методом, обычно используемым в данных ситуациях) по возможности корректировать вышеупомянутый вторичный эффект. Кроме того, частота восходящих кварт имеет тенденцию к аппроксимации, что наблюдается в легкой цепи щелочного миозина млекопитающих, стимулирующего сокращение мышц (тогда как магний, как известно, действует как деконтрактант). Поэтому его прослушивание также побуждает к физическим упражнениям, которые, как известно, являются другим способом регуляции холестерина. Это последний пример фактически иллюстрирует важность этого почти универсального явления, которое представляет собой эпигенетическую кооперацию различных факторов, стимулирующих синтез белков; а если рассматривать музыкальную последовательность миозина с точки зрения ее семантики и информативности, то "прослушивание" миозина создает впечатление военного марша.
4. Пример биохимического анализа эпигенетической кооперации, включая гармонические суперпозиции.
Биохимический анализ эпигенетической кооперации может значительно облегчить декодирование. Так, например, хорошо известно, что другим способом эпигенетической стимуляции расслабления мышц является воздействие теплом, которое обычно используется при ревматизме. Это тепловое действие обычно сопровождается совместным синтезом группы белков, вызываемого "тепловым шоком". Это дает основание предположить, что эти белки должны обнаруживать гармонические суперпозиции: и действительно, hsp 27, который оказался наиболее музыкальным, совмещается с началом hsp 70, который является наиболее избыточным, и, очевидно, в данном случае играет роль исходного материала. Эти две молекулы совмещаются в свою очередь, с началом тропонина C, который регулирует кальций при мышечных сокращениях и для которого отводилась наиболее важная роль как антиревматического агента, поскольку его музыкальный уровень достаточно высок (фиг. 4). Однако следует отметить, что многие другие молекулы, также обладающие высоким музыкальным уровнем и эпигенетической чувствительностью, могут тем не менее быть связанными с заболеванием этого типа в результате стимуляции пролактина и бета-липотропина (предшественника бета-эндоффина) и ингибирования эстрогенового рецептора, включая ингибирование IgE и интерлейкина 1-бета.
Эти несколько примеров наглядно иллюстрируют способ постадийного восстановления больших участков пути метаболизма с использованием различных методов контроля для проверки соответствия полученных результатов, что позволяет еще более уточнить музыкальное декодирование рассматриваемых белков.
Для реализации изобретения можно использовать переложение либо в виде нотной записи, либо в виде звукозаписи полученных музыкальных последовательностей, а также в виде цветового пространственного представления белков вместе или отдельно, на всех типах носителей таких, как пластинки, компакт-диски, гибкие диски, аудио- или видеокассеты, бумага, ткани и другие носители, предназначенные для цветовой визуализации.
Звукозапись музыкальных последовательностей может быть реализована с нотных записей с использованием известных методов (см., например, В.Н.Repp. J. Acoust. Soc. Am. 88, c. 622, 1990).
В области сельского хозяйства и текстильной промышленности изобретение может быть использовано для стимуляции специфического синтеза некоторых белков, связанных, например, с лактацией у коров, с ферментацией пивных дрожжей, с приданием соответствующего вкуса некоторым фруктам и животным или растительным волокнам (кератин овечьей шерсти, фиброин тутового шелкопряда и т.п.), а также белков, специфичных для некоторых лекарственных трав. В области охраны окружающей среды стимуляция биосинтеза некоторых белков может быть осуществлена в целях поглощения производственных отходов. Наблюдение коров, которым регулярно в течение 15 дней и во время дойки давали прослушивать записи музыкальных транскрипций аминокислотных последовательностей коровьего пролактина, лактоглобулина и лактальбумина, показало снижение (в 3 раза) относительного количества молочной сыворотки и значительное увеличение содержания белков в молоке и соответственно в полученном из этого молока остром сыре. Аналогично в эксперименте с томатами, которые во время их культивирования подвергались воздействию "коктейля" из музыкальных транспозиций различных белков, таких, как специфические ингибиторы вирусов, различные экстенсины (в данном случае фермент цветения LAT 52), антибактериальный белок (от которого ожидалось благодаря его музыкальной схожести с тауматином увеличение процента сахара (P 23)) и, наконец, ингибиторы ферментов размягчения плодов (пектинэстераза и полигалактуроназа), наблюдалось значительное увеличение размеров и количества плодов (в целом в отношении примерно 3,5), а также ощутимое увеличение сладкого вкуса у значительного количества плодов, особенно подвергнутых воздействию P 23. Однако эти результаты должны восприниматься с осторожностью, поскольку имеются некоторые противопоказания к избыточной стимуляции, например, пролактина; и животноводы, использующие указанные методы, должны принимать во внимание, что и сами животные могут быть также ослаблены. Так, например, в хорошо известных экспериментах, проводимых в Израиле на коровах с использованием музыки Моцарта (коровий пролактин, независимо от его "музыкального уровня", который является действительно высоким, что можно легко определить математически), наблюдалось возрастание частоты заболевания маститом у коров; и в этом случае "прослушивание" пролактина следует завершить прослушиванием альфа-1-трипсина, который также является музыкально эффективным и метаболизм которого также является дополняющим фактором на этой стадии. Аналогично для томатов, подвергающихся внешней стимуляции, очень важно не прерывать цикл слишком резко.
Что касается лечения и профилактики заболеваний, то в этой связи можно отметить, что многие заболевания характеризуются специфической метаболической недостаточностью, а поэтому лечение и предупреждение этих заболеваний может быть с успехом осуществлено с использованием изобретения. Поскольку минимальная длина музыкально активной последовательности составляет порядка длины сигнального пептида (от нескольких аминокислот до нескольких десятков аминокислот), то действие этой последовательности может быть очень быстрым и проявляться уже через несколько секунд или несколько минут. Тем не менее полная интеграция продуцируемого эффекта, который представляет собой метаболический комплекс, может занимать несколько более продолжительное время или даже требовать в случае более жестких условий определенной начальной тренировки, но, как правило, этот эффект довольно быстро достигает желаемой цели.
Поэтому для эффективного использования рассматриваемого способа важно точно определить метаболическую роль данной молекулы. Это является одним из наиболее интересных и важных факторов музыкального декодирования белков, ассоциированных с соответствующими цветами, который позволил бы путем систематического выявления сходства и несходства мелодий (и цветов), продуцируемых последовательностями белков, имеющихся в банке данных, выделить или отобрать белки, которые являются метаболическими агонистами и антагонистами данного белка и для которых степень музыкальной выразительности также является важным показателем их метаболической роли.
В связи с этим следует напомнить, что среди животных и растительных белков, особенно среди наиболее музыкальных белков, наблюдаются последовательности мелодических фрагментов метаболических цепочек человека, в результате чего можно сделать вывод, что транспозиции, которые являются активными у человека, не ограничиваются молекулами человека. Напротив, метаболизм некоторых видов является до некоторой степени более "специализированным" в отношении продуцирования определенных молекул, и поэтому наиболее музыкальные белки имеют особо важное значение. Само собой разумеется, что соответствия между различными видами облегчают определение границ метаболической рели белков и декодирование белковых последовательностей.
В этой связи важно отметить, что музыкальность молекулы сама по себе означает, что ее эпигенетическая стимуляция в основном предпочтительна для терапевтического применения (вследствие определенных пределов метаболических взаимодействий) в целях ее прямого поглощения, т. е. "наиболее музыкальными" молекулами являются в основном такие молекулы, для которых либо продуцирование с помощью генной инженерии, либо терапевтическое использование в результате такого продуцирования сталкивается с определенными проблемами, такими, как доставка к месту действия, стабильность или побочные действия, связанные с дозами, которые должны быть гораздо более значительными для получения нужного эффекта, чем дозы, имеющиеся в организме, поскольку в этом случае масштабные волны, ассоциированные с их продуцированием, больше не присутствуют. И это особенно относится к ингибированию белков, если природный ингибитор является, например, гораздо более сильным или если продуцирование необходимо уменьшить в данное время или систематическим образом.
И наконец, что касается использования транскрипций белковых последовательностей, то их чрезвычайно быстрое действие дает возможность посредством дифференциального, а особенно биполярного сравнения их положительных и отрицательных эффектов уточнить, какие из них являются наиболее приемлемыми в данной ситуации. Указанная идентификация может быть облегчена путем сравнения с транскрипциями акустических или электромагнитных явлений, генерирующих различные серии частот, для последовательностей известных белков и для некоторых эффектов, которые могут наблюдаться в аналогичной ситуации.


Изобретение относится к области эпигенетической регуляции биосинтеза белков. При формировании музыкальных последовательностей декодируют и транспонируют в звук временные ряды квантовых осцилляций. Декодирование осуществляют в четыре этапа. В результате декодирования получают музыкальные последовательности, обеспечивающие регулирование биосинтеза белка. При характеризации белковых последовательностей используют результаты декодирования. 2 с. и 5 з. п. ф-лы, 4 ил.
ν ≅ νoArgch(e(f/fo)Logch1),
где f, fo - музыкальные частоты;
ν,νo - частоты цветов;
индекс "o" обозначает центральные значения;
Argch - инверсная функция гиперболического косинуса;
ch1 - гиперболический косинус единицы.
| J.Sternheimer | |||
| Musigue des particules elemetaires | |||
| C.R.Acad | |||
| Sc.Paris, 19 83, N 12, p | |||
| АВТОМАТИЧЕСКОЕ УСТРОЙСТВО ДЛЯ ПОДАЧИ УГЛЯ К ТОПКАМ | 1920 |
|
SU297A1 |

