Изобретение относится к области молекулярной биологии и биотехнологии, а именно к способам клонирования нуклеиновых кислот. Преимущественной областью использования является получение индивидуальных нуклеиновых кислот.
В настоящее время клонирование нуклеиновых кислот осуществляют in vivo, то есть в живых клетках. Процедура клонирования включает ряд стадий, таких как встраивание (легирование) фрагментов нуклеиновых кислот в клонирующие векторы (например, плазмиды или вирусные геномы), трансформация живых клеток полученными рекомбинантными молекулами, и получение колоний трансформированных клеток или вирусных "бляшек" на поверхности питательного агара [1]. Здесь используется способность живых клеток и вирусов к экспоненциальному размножению, благодаря чему можно быстро получить большое потомство каждой трансформированной клетки, несущей желательную нуклеиновую кислоту. Эту процедуру часто называют молекулярным клонированием, что не вполне верно, так как клонируют клетки или вирусы, а не молекулы. Кроме того, рекомбинантные нуклеиновые кислоты размножаются в клетках наряду с клеточными ДНК и РНК, что требует их последующей очистки.
В настоящее время известен ряд способов экспоненциального размножения нуклеиновых кислот в бесклеточных системах, таких как размножение РНК вирусными РНК-зависимыми полимеразами (ВРП), размножение ДНК в полимеразной цепной реакции (ПЦР) и изотермическая трехферментная система самоподдерживающегося размножения (3СР). Реакции экспоненциального размножения нуклеиновых кислот in vitro проводят в жидких средах, содержащих компоненты бесклеточной ферментной системы, в том числе реакционный буфер, соответствующие ферменты, нуклеотидные субстраты и, когда необходимо, полимеризационные затравки. Благодаря экспоненциальной природе описанных реакций размножения каждая из них могла бы быть использована для быстрого получения многочисленного потомства одиночных молекул нуклеиновых кислот. Это дало бы возможность клонировать нуклеиновые кислоты in vitro, то есть, дало бы альтернативу традиционному клонированию генов в живых клетках.
В ВРП реакции экспоненциальный синтез достигается благодаря тому, что как матричная РНК, так и синтезированная РНК остаются однонитевыми, и обе служат одинаково эффективными матрицами в последующих циклах репликации. Эта реакция не требует затравок. Вирусные РНК полимеразы, такие как Qβ репликаза, демонстрируют узкую матричную специфичность, основанную на узнавании ферментом особых структур, которые присутствуют в специфических матричных РНК, но отсутствуют в других РНК. РНК, содержащие такие структуры, называют реплицирующимися РНК. Другие РНК, не являющиеся реплицирующимися, могут быть размножены в ВРП реакции, если они встроены в цепочку реплицирующейся РНК [2] . Такие рекомбинантные реплицирующиеся РНК были использованы для внеклеточного размножения мРНК, кодирующих белки [3].
ПЦР используют для внеклеточного размножения ДНК. Для осуществления этой реакции необходимы две олигонуклеотидных затравки, которые комплементарны участкам цепочек двунитевой ДНК, ограничивающим размножаемый фрагмент (мишень). Эти затравки отжигаются с ДНК и наращиваются во взаимно встречных направлениях ДНК полимеразой. В отличие от ВРП реакции, матричная и синтезируемая цепочки ДНК оказываются в дуплексе, который необходимо расплавить при повышенной температуре, чтобы разрешить последующие циклы репликации, в которых каждая из цепочек служит матрицей. Процесс многократно повторяется путем циклической смены температур, при которых происходит отжиг затравки и плавление дуплекса ДНК, что приводит к экспоненциальному размножению ДНК-мишени [4] . В настоящее время ПЦР проводят с использованием термоустойчивых ДНК-полимераз, которые выдерживают многократную смену температур без потери активности [5] . Необходимость циклической смены температур требует наличия специального оборудования и делает ПЦР на порядок медленнее ВРП реакции. В то же время, любая ДНК-мишень может быть размножена в ПЦР при наличии пары специфических затравок, без надобности в изготовлении рекомбинантной молекулы.
Способ размножения нуклеиновых кислот в трехферментной системе самоподдерживающегося размножения (3СР) соединяет преимущества ПЦР и ВРП реакции. 3СР основана на совместном действии трех ферментов: ДНК- зависимой РНК-полимеразы, обратной транскриптазы и РНКазы H [6]. В качестве стартовой матрицы может служить как фрагмент двунитевой ДНК, несущий на каждом конце промотор какой-либо РНК-полимеразы, так и однонитевая РНК. РНК-полимераза (например, РНК-полимераза фага Т7) использует двунитевую ДНК для синтеза многочисленных однонитевых РНК- транскриптов с последовательности ДНК, лежащей вслед за промотором на каждой цепочке ДНК. Обратная транскриптаза (например, обратная транскриптаза вируса миелобластоза птиц) синтезирует двунитевые кДНК-копии РНК-транскриптов, используя затравки, комплементарные 3'-концам транскриптов и содержащие последовательность промотора РНК-полимеразы для восстановления этой последовательности на каждом конце кДНК. РНКаза H разрушает РНК-матрицу, вовлеченную в РНК/ДНК гетеродуплекс после синтеза первой цепи кДНК, тем самым позволяя синтезироваться второй цепи кДНК. Действие РНК-полимеразы и РНКазы H приводит к образованию однонитевых матриц, что позволяет экспоненциально размножать нуклеиновые кислоты в отсутствие циклического изменения температуры. Скорость 3СР реакции сравнима со скоростью ВРП реакции, причем подобно ПЦР использование 3СР не требует создания рекомбинантых нуклеиновых кислот. Продуктом 3СР реакции является смесь молекул двунитевых ДНК и однонитевых РНК.
В качестве прототипа настоящего изобретения может быть рассмотрена работа Левисона и Спигельмана [7], заявивших, что им удалось клонировать молекулы РНК, используя Qβ репликазную систему размножения. Они наблюдали синтез РНК в примерно половине образцов после добавления РНК-матрицы, разбавленной до такой степени, чтобы в среднем получить 0,5 молекулы РНК на образец. Очевидно, использованный прием не может гарантировать получения потомства единственной молекулы, так как из-за случайности распределения в образце может оказаться две, три и более матричных молекул. Более того, в этих экспериментах продукт синтеза не был идентифицирован, а вывод авторов был поставлен под сомнение наблюдением, что синтез РНК в Qβ репликазной системе может происходить даже в отсутствие добавленной матрицы [8]. Недавно было показано, что этот спонтанный синтез РНК вызывается реплицирующимися РНК, загрязняющими окружающее пространство, и что в среднем образец содержит до 100 посторонних молекул реплицирующихся РНК [9]. Проблема загрязнений также присуща ПЦР и ЗСР реакции, хотя она и не столь серьезна, как в случае ВРП реакции, поскольку размножение нуклеиновых кислот находится под контролем олигонуклеотидных затравок, комплементарных мишени. Более существенной в этом случае является ограниченная специфичность затравок. Из-за наличия неправильного спаривания оснований и гетерогенности олигонуклеотидных затравок возможен отжиг затравки на чужеродной матрице, присутствующей в образце, который начинает заметно конкурировать с правильным отжигом при понижении доли специфических матриц в образце ниже определенного предела. По крайней мере, 100 копий специфической матрицы должно присутствовать в образце для надежного запуска ПЦР [10]. Таким образом, ни один из существующих методов внеклеточного размножения нуклеиновых кислот не позволяет достичь истинного клонирования in vitro, то есть получения однородного потомства единственной молекулы в бесклеточной системе.
Настоящее изобретение основано на обнаружении нами того факта, что нуклеиновые кислоты подобно микроорганизмам могут быть выращены в виде колоний в иммобилизованной среде. Предлагаемый способ позволяет клонировать молекулы нуклеиновых кислот in vitro, несмотря на загрязнение образца посторонними матрицами и на неспецифический отжиг затравок на чужеродных матрицах. Возможно использование любых бесклеточных систем экспоненциального размножения нуклеиновых кислот, таких как ВРП реакция, ПЦР или 3СР реакция. В отличие от указанного прототипа и других известных способов размножения нуклеиновых кислот в жидких средах потомство (клон) каждой одиночной молекулы нуклеиновой кислоты образует колонию молекул в ограниченной зоне вокруг родительской матрицы. Разные колонии занимают, как правило, разные зоны иммобилизованной среды, что позволяет наблюдать индивидуальные колонии и манипулировать с ними по-отдельности.
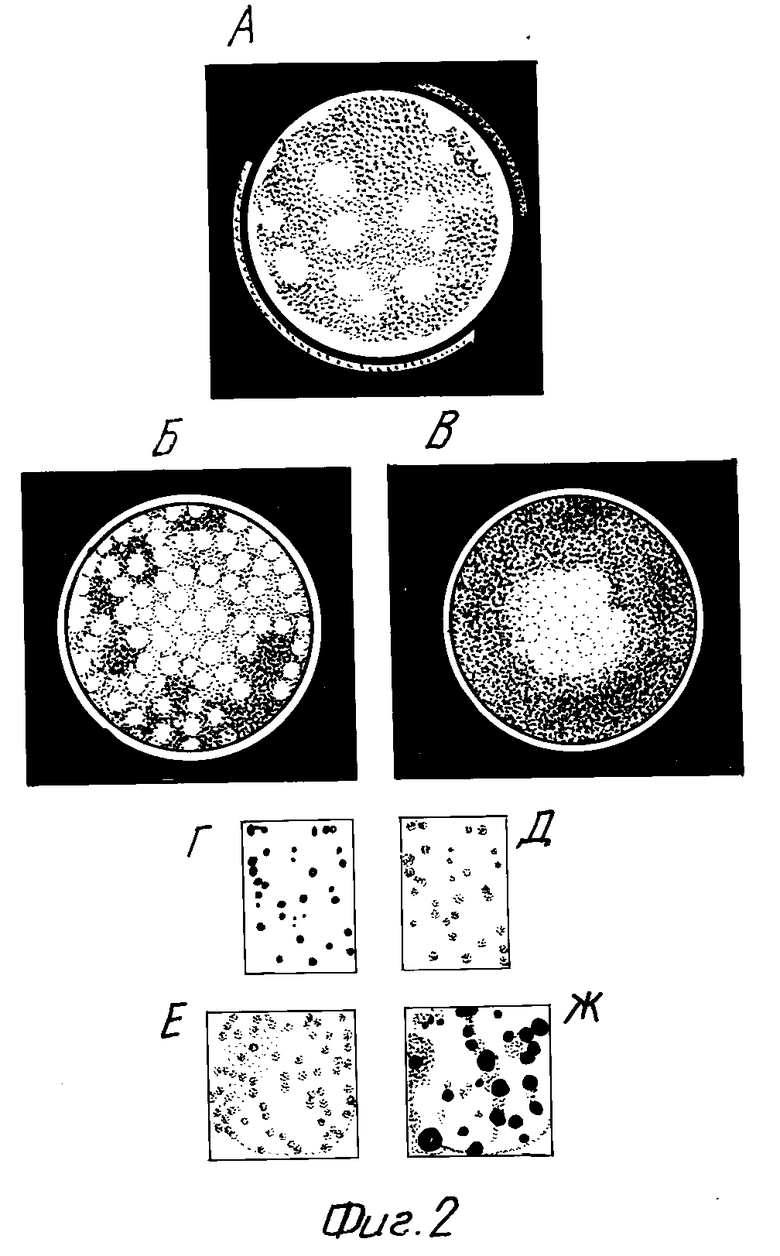
На фиг. 1 схематически изображает рост колоний нуклеиновых кислот в иммобилизованной среде; на фиг. 2 колонии РНК, выросшие в тонком слое иммобилизованной среды, содержащей компоненты Qβ репликазной системы.
Иммобилизованная среда для клонирования нуклеиновых кислот включает жидкую фазу, заключенную в твердой основе. Твердая основа иммобилизованной среды может иметь разнообразную структуру, например пористую, волокнистую, сетчатую, капиллярную, слоистую, складчатую. Основным требованием к твердой основе является наличие развитых поверхностей, пронизывающих жидкую фазу, среднее расстояние между которыми обеспечивает ту или иную степень иммобилизации жидкости за счет ее трения о поверхность и структурирования воды вблизи поверхности. Предпочтительно использовать твердую основу со средним расстоянием между поверхностями (размером пор) от 100 мкм (крупнопористая основа) до 5 нм (мелкопористая основа). Верхний предел размера пор должен быть меньше расстояния, на которое размножаемые нуклеиновые кислоты способны диффундировать за время инкубации (порядка 1 мм за час инкубации при комнатной температуре для нуклеиновых кислот длиной около 100 нуклеотидов, ср. фиг. 2A - 2B). Нижний предел размера пор должен быть несколько больше линейных размеров нуклеиновых кислот и/или ферментов, чтобы обеспечить их подвижность, необходимую для протекания реакции размножения нуклеиновых кислот. В качестве твердой основы может служить трехмерная сеть, образуемая отдельными молекулами полимеров или их агрегатами; склеенные, спеченные или спрессованные водонерастворимые порошки, мелкие гранулы или кристаллы; губчатые материалы; плотно упакованные волокна, капилляры или пленки (например, полимерная пленка или металлическая фольга). Основными требованиями к материалу, из которого изготавливают твердую основу, является его химическая инертность в условиях инкубации, гидрофильность формируемой им поверхности, а также отсутствие физических воздействий на ферменты (например, их необратимая сорбция на поверхности), влекущих их инактивацию. Подходящими материалами для изготовления твердой основы являются различные органические или неорганические носители, используемые в биотехнологии для хроматографии и электрофореза биополимеров, для иммобилизации ферментов, а также для выращивания микроорганизмов, клеток и вирусов. Примерами таких носителей являются агароза, полиакриламид, желатин, альгинат, карагеннан, целлюлоза, силикагель, губчатый титан, оксиапатит, химически сшитые агароза, декстран и полиэтиленгликоль, а также их комбинации и производные.
Согласно изобретению, иммобилизованная среда для клонирования нуклеиновых кислот содержит также компоненты системы размножения, включающей бесклеточную ферментную систему, способную к экспоненциальному размножению нуклеиновых кислот. Примеры такой бесклеточной системы рассмотрены выше. Ферменты, входящие в состав бесклеточной системы, могут быть либо растворены в жидкой фазе, либо иммобилизованы на твердой основе. Реакционный буфер, размножаемые нуклеиновые кислоты, субстраты и (где необходимо) затравки вводят в жидкую фазу.
При размножении в иммобилизованной среде нуклеиновые кислоты образуют колонии, как схематически показано на фиг. 1. На верхней диаграмме представлен фрагмент 11 иммобилизованной среды, образуемой сетью твердой основы 12 и водным раствором 13, содержащим молекулы фермента 14, нуклеиновокислотные матрицы 15, а также субстраты и реакционный буфер. В результате инкубации иммобилизованной среды в течение определенного времени при температурном режиме, подходящем для размножения нуклеиновых кислот, образуются колонии нуклеиновых кислот 16 в зонах локализации родительских матриц.
Предпочтительно, чтобы твердая основа иммобилизованной среды, используемой для клонирования, могла обратимо взаимодействовать с нуклеиновыми кислотами. Это позволяет существенно подавить диффузию молекул нуклеиновых кислот и получить колонии меньшего размера, с более четкими краями и большей концентрацией нуклеиновых кислот; иными словами, значительно увеличить разрешающую способность метода. Большинство гидрофильных полимеров способно к взаимодействию с нуклеиновыми кислотами благодаря образованию водородных связей. Эта способность может быть усилена путем химической модификации твердой основы положительно заряженными группами, способными к ионным взаимодействиям с фосфатными остатками нуклеиновых кислот, и/или умеренно гидрофобными группами, способными к взаимодействиям с пуриновыми или пиримидиновыми кольцами. Например, твердая основа может быть модифицирована слабокатионными группами, такими как этиламиноэтильными или диэтиламиноэтильными, или интеркалирующими красителями, такими как этидиум или пропидиум. Интеркалирующие красители способны к гидрофобным взаимодействиям со стекингованными основаниями нуклеиновых кислот и при этом несут положительный заряд. Более того, присутствие интеркалирующих красителей в иммобилизованной среде позволяет обнаруживать колонии нуклеиновых кислот с помощью флуоресценции (см. ниже).
Чтобы предотвратить распространение нуклеиновых кислот в иммобилизованной среде в процессе роста колоний, очень важно не допускать наличие неиммобилизованной жидкости. В частности, нельзя допускать конденсации воды на поверхности иммобилизованной среды. В то же время нельзя допускать и пересыхания иммобилизованной среды, что может приводить к инактивации ферментов. Пересыхание иммобилизованной среды можно предотвратить путем проведения инкубаций в плотно закрытой камере, кассете или запаянном пластиковом пакете, а также оборачиванием среды водонепроницаемой пленкой или наслаиванием минерального масла.
Предпочтительно, чтобы реакция размножения не начиналась до полной иммобилизации среды. Это условие легко выполнимо в случае ПЦР, так как в этом случае реакция запускается циклическим изменением температуры. В случае реакций экспрессии или изотермического размножения, таких как ВРП или 3СР реакция, это условие выполняется либо путем приготовления среды при или около 0oC, когда скорость реакции мала, либо путем помещения ферментов и субстратов в разные зоны иммобилизованной среды, после чего субстраты диффундируют в ферментную зону, либо путем использования защищенных субстратов (химически недоступных ферменту) с последующим удалением защиты и высвобождением нормального субстрата. Примером защищенного субстрата является светочувствительное производное АТР, в котором γ -фосфат модифицирован 1-(2-нитро)фенилэтильной группой [11], и которое может быть превращено в АТР фотолизом для запуска реакции размножения.
Для клонирования предпочтительно использовать иммобилизованную среду, приготовленную в виде тонкого слоя. В этом случае растущие колонии располагаются в двух измерениях, что облегчает их наблюдение, анализ и пересев, а также позволяет изготавливать реплики, например, путем частичного переноса колоний на мембранный фильтр. Реплики можно использовать для анализа колоний, для их пересева на новую среду, а также хранить в течение длительного времени.
Малая толщина слоя уменьшает температурные градиенты в поперечном направлении (что особенно важно в случае ПЦР) и облегчает диффузию субстратов в ферментный слой при использовании сэндвичевых сред с раздельными ферментным и субстратным слоями (см. ниже). Предпочтительно, чтобы толщина слоя не превышала размера колоний (который определяется, главным образом, скоростью линейной диффузии нуклеиновых кислот), то есть была в пределах нескольких миллиметров. Наиболее предпочтительно использовать очень тонкие слои, так как это повышает разрешающую способность метода. Нижний предел толщины слоя зависит от механической прочности твердой основы, от способности предотвратить пересыхание слоя при манипуляциях и в процессе реакции размножения, а также от чувствительности метода, используемого для анализа колоний. На практике удовлетворительные результаты получаются при толщине слоя от 1 мм до 50 мкм. Конечно, могут быть использованы более толстые (например, 10 мм) и более тонкие (до 1 мкм) слои. Если используют более толстые слои, то предпочтительно помещать ферменты и субстраты в разные слои, которые накладывают друг на друга, и/или вводить нуклеиновокислотные матрицы на поверхность слоя или между слоями, чтобы ограничить реакцию размножения узкой зоной вблизи поверхностей слоев. Более тонкие слои предпочтительно иммобилизовывать на подложке из прочного материала, такой как стеклянная, металлическая или пластиковая пластинка или пленка.
Выбор техники приготовления тонких слоев зависит от свойств твердой основы, от температурного режима реакции, от свойств ферментов, используемых для реакции (как например, их способность переносить условия формирования полимерной основы), а также от того, используют ли однослойную или многослойную иммобилизованную среду.
Однослойные среды удобно использовать, когда все компоненты ферментной системы могут быть смешаны без того, чтобы началась реакция (как в случае ПЦР). В то же время, однослойные среды могут быть использованы и тогда, когда предпочтительно держать ферменты отдельно от субстратов до полной иммобилизации среды. В этом случае ферментный слой, содержащий также нуклеиновокислотные матрицы, приготавливают без субстратов и покрывают полупроницаемой (например, диализной) мембраной, или же для приготовления ферментного слоя используют твердую основу, способную удерживать нуклеиновые кислоты и ферменты (например, модифицированную положительно заряженными группами). Для запуска реакции размножения такой слой приводят в контакт с субстратным раствором.
Во многих случаях предпочтительно использовать многослойные (сэндвичевые) среды, в которых разные слои приводятся в непосредственный контакт друг с другом в нужный момент. Например, ферменты и субстраты могут быть разнесены в разные слои, и реакция запускается путем соприкосновения слоев благодаря диффузии субстратов в ферментный слой. Использование сэндвичевых сред может быть предпочтительным и в том случае, если разносить ферменты и субстраты не требуется, как, например, в ПЦР. Например, вторым слоем может быть мембранный фильтр, используемый для анализа колоний, на который нуклеиновые кислоты переносятся одновременно с их синтезом. Второй слой может также содержать компоненты системы экспрессии нуклеиновых кислот и наслаиваться на первый слой по завершении ПЦР для анализа колоний по продукту экспрессии. В этом случае может быть использован и третий слой, в качестве которого может служить мембранный фильтр для переноса продуктов экспрессии или же мембранный фильтр, пропитанный субстратами ферментов, синтезируемых в результате экспрессии, для анализа колоний по активности кодируемых ферментов.
Тонкие слои могут быть изготовлены разными способами, такими как пропитывание сухой твердой основы растворами, содержащими ферменты и/или субстраты, или включение ферментов и/или субстратов в процессе гелеобразования или синтеза (полимеризации) твердой основы.
Гелеобразующие растворы, такие, как основанные на агарозе, желатине, карагенане, смешивают с ферментами и/или субстратами и заливают, например, в чашки Петри или между двумя ровными поверхностями. В последнем случае можно получить очень тонкие слои геля и избежать образование мениска. Ферменты и/или субстраты могут быть также включены в слой в процессе синтеза твердой основы, например в процессе полимеризации акриламида [12] или фотоиндуцируемой полимеризации этиленгликоля [13] . В тех случаях, когда формирование твердой основы происходит в условиях, слишком жестких для ферментов и/или субстратов, сначала приготавливают твердую основу, а затем пропитывают ее соответствующим раствором. Например, повысить термоустойчивость агарозного геля (чтобы его можно было использовать в ПЦР) можно путем сшивания агарозы эпихлоргидрином или 2,3- дибромпропанолом; термоустойчивый гель можно также получить путем сшивания эпихлоргидрином или N,N'-метиленбисакриламидом декстрана [14] . Однако, сшивание происходит в условиях, ведущих к инактивации ферментов и субстратов реакции размножения. Поэтому тонкий слой геля сначала формируют в отсутствии ферментов и субстратов и затем пропитывают раствором, содержащим ферменты и/или субстраты. Таким же образом могут быть приготовлены слои из геля на основе полиакриламида или смеси полиакриламида и агарозы. Волокнистые слои (например, основанные на целлюлозе или нейлоне) или пористые слои (например, основанные на силикагеле или губчатом титане) могут быть приготовлены просто пропитыванием соответствующих сухих листов, мембранных фильтров или пластинок раствором, содержащим компоненты системы размножения.
Нуклеиновые кислоты, предназначенные для клонирования, вводят в жидкую фазу твердой среды путем включения в ферментный и/или субстратный раствор перед его иммобилизацией или путем нанесения на слой или между слоями.
Реакцию размножения запускают помещением иммобилизованной среды в соответствующие условия: повышение температуры среды, циклическое изменение температуры, воздействие света, приведение в соприкосновение слоев, содержащих ферменты и субстраты. Среду инкубируют в течение промежутка времени, позволяющего продуктам синтеза накопиться до детектируемого уровня. Например, при размножении нуклеиновых кислот необходимо, чтобы в каждой колонии образовалось, по крайней мере, 106 копий нуклеиновой кислоты (то есть, должно произойти, по крайней мере, 20 циклов репликации при условии, что в каждом цикле число матриц удваивается), что соответствует нижнему пределу чувствительности существующих методов обнаружения нуклеиновых кислот [15].
По окончании реакции размножения колонии нуклеиновых кислот можно обнаружить различными способами, такими как окрашивание интеркалирующими красителями (этидиум бромид, пропидиум йодид), авторадиографией (если для размножения использованы субстраты, меченые радиоизотопом), а также неизотопными методами, такими как использующими мечение субстратов флуоресцентными группами и их модификацию биотином или дигоксигенином. Колонии могут быть также анализированы на предмет обладания каким-либо признаком, таким как присутствие определенной нуклеотидной последовательности (путем гибридизации со специфическим меченым зондом) или способность кодировать определенный белок (путем внеклеточной экспрессии содержащихся в нуклеиновых кислотах генов). Часто обнаружение колоний удобно проводить после их частичного переноса на мембранный фильтр. В этом случае мембранный фильтр предпочтительно накладывать на ростовый слой перед размножением, чтобы перенос происходил по мере роста колоний. Обнаруженные клоны можно далее размножить, используя известные способы размножения нуклеиновых кислот in vitro.
Экспрессию нуклеиновых кислот с целью анализа колоний можно проводить в жидкой среде, однако предпочтительно осуществлять экспрессию в иммобилизованной среде, сформованной, по крайней мере, в один тонкий слой. Это позволяет параллельно анализировать большое число колоний в одной реакции. При экспрессии в иммобилизованной среде продукты экспрессии накапливаются в тех зонах, где присутствуют колонии соответствующих нуклеиновых кислот. При использовании изотермических реакций размножения (таких как ВРП или 3СР), компоненты системы экспрессии могут быть введены в тот же слой (или те же слои), что и компоненты системы размножения. Если для размножения используют ПЦР, то система экспрессии может быть включена в ту же среду, но в отдельный слой, который наслаивают по завершении ПЦР, или же в отдельную среду. В последнем случае, колонии ДНК переносят в экспрессирующую среду, например, с помощью мембранного фильтра. Для экспрессии нуклеиновые кислоты получают в форме РНК (так, как они образуются в реакциях ВРП или 3СР) и транслируют в белки в среде, содержащей бесклеточную систему трансляции [16-18]. Если же нуклеиновые кислоты получены в форме ДНК (как при размножении в ПЦР), то экспрессию осуществляют в среде, включающей сопряженную бесклеточную систему транскрипции/трансляции [19-22]. Синтезированные полипептиды можно идентифицировать in situ или на фильтре-реплике разнообразными способами, такими как использующими иммунохимические реакции, или способность синтезированных белков осуществлять специфические ферментативные реакции или связывать специфические лиганды.
Способ клонирования нуклеиновых кислот in vitro может быть использован в тех же целях, что и клонирование in vivo [1]. В то же время он обладает рядом преимуществ: клонирование in vitro быстрее; оно позволяет получить клон любой нуклеиновой кислоты без каких-либо ограничений, накладываемых клеткой, и позволяет получать гены, свободные от загрязнений хромосомной ДНК или клеточной РНК. Полученные клоны нуклеиновых кислот можно использовать для трансформации клеток, в терапевтических целях (например, для лечения с использованием антисмысловых РНК или ДНК), или для использования в качестве матриц в белоксинтезирующих биореакторах [23].
Из-за чрезвычайной чувствительности способа важно максимально уменьшить возможность загрязнения образца посторонними нуклеиновыми кислотами до стадии размножения. В связи с этим предпочтительно проводить стадии, предшествующие размножению, и последующие стадии (анализ колоний) в разных лабораториях. Предпочтительно выращивать колонии в тонких слоях, помещенных в герметичные кассеты, которые вскрывают только в аналитической лаборатории.
Предложенный способ иллюстрируется следующими примерами его осуществления.
Пример 1. Выращивание колоний РНК с помощью ВРП реакции.
Этот пример иллюстрирует способ размножения РНК в тонком слое иммобилизованной среды на примере RQ-РНК, являющихся природными матрицами Qβ репликазы. Порошок низкотемпературной агарозы (ultra-low gelling temperature agarose type IX, Sigma Chemical Company) расплавляют нагреванием в автоклавированном буфере А (80 мМ Трис-HCl pH 7,8, 2 мМ MgCl2, 1 мМ ЭДТА, 20%-ный глицерин), охлаждают до ≈40oC и тщательно перемешивают на роторном встряхивателе с концентрированным раствором Qβ репликазы, очищенной, как описано Блюменталем [24] . Конечные концентрации агарозы и Qβ репликазы - 2% и 35 мкг/мл соответственно. Затем приготавливают сэндвичевую среду одним из нижеописанных способов. Когда необходимо, разведения матричной РНК вводят в субстратный слой или в промежуток между ферментным и субстратным слоями в виде небольшой аликвоты.
(а) 1,5 мл вышеуказанного раствора наслаивают на слой 1%-ной агарозы, содержащий субстраты (по 1 мМ ATP, GTP, CTP и UTP) и буфер Б (80 мМ Трис-HCl pH 7,8, 25 мМ MgCl2, 2 мМ ЭДТА, 10%-ный глицерин), и приготовленный в 35-миллиметровых пластиковых чашках Петри. После заливки каждого слоя чашку выдерживают на льду для застывания агарозы. Для размножения РНК чашку инкубируют при комнатной температуре или при З7oC в течение 1 ч. (на термостатированном столике). Колонии РНК проявляют окрашиванием раствором этидиум бромида, как показано на фиг. 2,А-Б. На фиг. 2,А показан "спонтанный" рост колоний РНК, наблюдаемый, когда верхний (ферментный) слой заливают сразу после застывания субстратного слоя. На фиг. 2,Б видно большое число колоний, выросших после того, как чашка с субстратным слоем была оставлена в лаборатории открытой в течение 1 ч. перед заливкой ферментного слоя. Этот эксперимент демонстрирует эффективность предложенного способа в предотвращении распространения "шумового" роста на весь реакционный объем, а также потенциальную опасность со стороны наружных (в частности, воздушных) загрязнений, если размножение нуклеиновых кислот проводится традиционным способом в жидкой среде. Краситель может быть включен в субстратный слой при его заливке; в этом случае колонии РНК можно наблюдать в процессе их роста (фиг. 2,В).
(б) 130 мкл раствора, содержащего фермент и агарозу, заливают между покровным стеклом, используемом в микроскопии, и большей по размеру пластиковой пластинкой, между которыми имеется зазор 0,4 мм. Покровное стекло осторожно снимают и на агарозу накладывают нейлоновый мембранный фильтр, пропитанный субстратным раствором (по 4 мМ ATP, GTP, CTP и UTP в буфере Б) и высушенный. Для обнаружения колоний РНК в субстратный раствор добавляют [ [α-32P]- ]-меченый нуклеотид. После инкубации среды в течение 20-40 мин для размножения РНК мембранный фильтр снимают с агарозы и отмывают от невключившихся субстратов. Если размножение проводили в присутствии меченого субстрата, делают радиоавтограф мембранного фильтра. На фиг. 2,Г показан негатив радиоавтографа исходного (субстратного) мембранного фильтра, а на фиг. 2,Д - негатив радиоавтографа фильтра-реплики. Колонии на фиг. 2,Г и 2,Д значительно менее диффузные, чем на фиг. 2,А-В. Разница объясняется наличием в нейлоновом мембранном фильтре положительно заряженных и гидрофобных групп, способных к взаимодействию с РНК. В отсутствие меченого субстрата колонии можно обнаружить с помощью окрашивания агарозы этидиум бромидом, как описано выше, или с помощью гибридизации мембранного фильтра со специфическими зондами, как описано ниже.
РНК пришивают к мембранному фильтру под действием ультрафиолета и гибридизуют с радиоактивно-меченым зондом путем инкубации в запаянном пластиковом пакете между листами фильтровальной бумаги [25]. Температуру гибридизации определяют по известной формуле [26] и оптимизируют в предварительных экспериментах. После гибридизации мембранный фильтр отмывают от негибридизовавшегося зонда и делают радиоавтограф. Для повторной гибридизации того же мембранного фильтра с другим зондом первый зонд отмывают кипячением мембранного фильтра в 0,1%-ном растворе Na-додецилсульфата. На фиг. 2,Е и 2,Ж показаны негативы радиоавтографов, полученных в результате последовательной гибридизации одного и того же мембранного фильтра с радиоактивно-мечеными зондами, специфичными к RQ87-3 РНК и RQ135-1 РНК соответственно. Смесь этих РНК была введена в субстратный слой в количестве около 100 копий каждой из них. Этот эксперимент демонстрирует, что разные колонии содержат разные виды РНК и что число колоний соответствует количеству добавленных матриц. Иными словами, имеет место истинное клонирование молекул РНК.
Пример 2. Выращивание колоний нуклеиновых кислот с помощью 3СР реакции.
Раствор низкотемпературной агарозы в буфере смешивают с ферментами и нуклеиновой кислотой, отожженной с олигонуклеотидными затравками при 65oC, и заливают ферментный слой, как описано в примере 1 (б). В качестве мишени используют РНК, ДНК или их смесь. Используют реакционный буфер, ферменты, затравки и условия инкубации согласно рекомендациям Гуателли и др. [6]. На ферментный слой наслаивают нейлоновый мембранный фильтр, пропитанный субстратным раствором, содержащим по 4 мМ dATP, dGTP, dCTP, dTTp и по 16 мМ ATP, GTP, CTP, UTP в реакционном буфере. После инкубации колонии нуклеиновых кислот идентифицируют при помощи гибридизации мембранного фильтра с меченым зондом, комплементарным внутренней области мишени, как описано в примере 1.
Пример 3. Выращивание колоний ДНК с помощью ПЦР.
Все компоненты реакции, включая буфер, термоустойчивую ДНК-полимеразу, образец ДНК, затравки и субстраты, смешивают с дегазированным раствором мономеров (смесь акриламида и N, N'-метиленбисакриламида) и катализаторами полимеризации (персульфат аммония и N,N,N',N'-тетраметилендиамин) и заливают тонкий слой геля. По завершении полимеризации на гель наслаивают нейлоновый мембранный фильтр, смоченный в реакционном буфере, оборачивают термостойкой пленкой и помещают гель на металлический термостатируемый столик, температура которого контролируется в требуемом для ПЦР диапазоне автоматическим устройством, включающим два [27] или три [28] водяных термостата и соединенным шлангами с термостатируемым столиком. Используют реакционные компоненты и условия ПЦР, рекомендованные Сэйкай и др. [5]. После 20 - 35 температурных циклов колонии ДНК обнаруживают, как описано выше. Если в качестве образца используют РНК, ее сначала превращают в кДНК [1].
Пример 4. Выделение индивидуальных клонов из сложных смесей нуклеиновых кислот.
В этом примере описана процедура, основанная на использовании ПЦР. Конечно, могут быть также использованы 3СР и ВРП реакции. При использовании ВРП нуклеиновые кислоты переводят в форму реплицирующейся РНК путем встраивания клонируемых последовательностей в молекулу RQ РНК [2].
В обычной ПЦР желательная нуклеотидная последовательность из смеси нуклеиновых кислот может быть избирательно размножена путем использования двух уникальных олигонуклеотидных затравок, специфичных к этой последовательности, что обуславливает необходимость синтеза новой пары олигонуклеотидов для выделения каждой новой последовательности. Предлагаемый способ позволяет клонировать любую нуклеотидную последовательность с использованием одной и той же пары универсальных затравок. Процедура клонирования включает получение смеси нуклеиновых кислот, чей размер не превышает размера, который может быть размножен в ПЦР (то есть, до 10 кб); навешивание на них универсальных концов; и использование для их размножения пары затравок, комплементарных универсальным концам. В этом случае каждая нуклеиновая кислота, присутствующая в смеси, может быть размножена и образует отдельную колонию при проведении ПЦР в тонком слое иммобилизованной среды. Следующим шагом является идентификация колонии (или колоний), содержащей потомство определенной нуклеиновой кислоты, и отбор этой колонии для дальнейшего размножения и использования.
Исходной смесью нуклеиновых кислот может быть, например, набор фрагментов, получаемый перевариванием геномной ДНК какой-либо рестриктазой [29], или смесь кДНК, полученных из цитоплазматических мРНК [30]. На фрагменты навешиваются универсальные концы либо легированием фрагментов с олигонуклеотидами, либо наращиванием гомополинуклеотидных последовательностей. Предпочтительно, чтобы последовательности, вводимые на 3' и 5' концах фрагментов, отличались друг от друга и не были комплементарны друг другу. Этого можно добиться, например, путем удлинения 3' концов за счет легирования двунитивых ДНК-фрагментов с 5'- фосфорилированными олигодезоксирибо-нуклеотидными адапторами, совместимыми с концами фрагментов, или путем добавления гомополинуклеотида с помощью дезоксинуклеотидил-трасферазы [1], и удлинения 5' концов за счет легирования с олигорибонуклеотидами [31].
После размножения фрагментов в тонком слое иммобилизованной среды для получения одиночных колоний как описано в примере 3, колонии анализируют на присутствие определенной последовательности (гена) подходящим для этого методом. Это может быть либо гибридизация со специфическим зондом, комплементарным внутренней области гена (см. выше), если известна хотя бы частичная нуклеотидная последовательность гена или его гомолога из другого источника, либо обнаружение гена по продуктам его экспрессии в системе транскрипции/трансляции in vitro.
Пример 5. Анализ клонов нуклеиновых кислот путем экспрессии в тонком слое иммобилизованной среды.
Если клонирование осуществляют с использованием ПЦР, то включают последовательность промотора РНК-полимеразы, такой как SP6 или Т7 полимераза, в один из универсальных концов, вводимых в клонируемые фрагменты, как описано в примере 4. Выросшие колонии приводят в контакт с экспрессирующим слоем. Этого достигают либо наслаиванием экспрессирующего слоя поверх слоя, в котором проводили ПЦР, либо переносом колоний на экпрессирующий слой с помощью мембранного фильтра. Экспрессирующий слой приготавливают, как описано в примере 1, с той разницей, что вместо компонентов Qβ репликазной системы в агарозу вводят компоненты подходящей бесклеточной системы трансляции/транскрипции [19-22] . Если для клонирования используют изотермическую систему размножения, то компоненты подходящей бесклеточной системы экспрессии [16-22] могут быть введены в тот же слой, что и компоненты системы размножения. Продукты экспрессии идентифицируют in situ по их способности выполнять определенные ферментативные реакции или по их способности связывать определенные лиганды или антитела.
Например, клоны, несущие ген фосфатазы, можно обнаружить с помощью мембранного фильтра, смоченного раствором 5-бром-4-хлор-3- индолилфосфата и нитросинего тетразолиума [32]. В тех местах, где синтезировалась фосфатаза, появляется пурпурное окрашивание. Гены, кодирующие фотобелки, такие как апообелин из Obelia geniculata, можно обнаружить по люминесценции [33]. В этом случае простетическая группа белка колентеразин вводится в экспрессирующий слой, залитый на прозрачной подложке. После инкубации экспрессирующего слоя для синтеза белков, его помещают подложкой на фотопленку, а с другой стороны на него наслаивают мембранный фильтр, смоченный раствором Ca++. Диффундируя в экспрессирующий слой и связываясь с обелином, ионы Ca++ индуцируют люминесценцию и, следовательно, засветку пленки в тех местах, где синтезировался апообелин.
Можно также идентифицировать продукты экспрессии с помощью разнообразных иммунохимических методов. Наиболее предпочтительны очень чувствительные методы, основанные на хемилюминесцентных реакциях, катализируемых щелочной фосфатазой и пероксидазой хрена. Можно использовать соответствующие наборы, продаваемые, соответственно фирмами United States Biochemical Corporation и Amersham International plc.
Список литературы.
1. Sambrook, J., Fritsch, E. F. and Maniatis, T. (1989). Molecular Cloning: A Laboratory Manual, 2nd edition. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N. Y.
2. Miele, A. A., Mills, D. R. and Kramer, F. R. (1983). Autocatalytic Replication of a Recombinant RNA. J. Mol. Biol. 171, 281-295.
3. Wu, Y. , Zhang, D. and Kramer, F. R. (1992). Amplifiable Messenger RNA. Proc. Natl. Acad. Sci. U.S.A., in press.
4. Saiki, R. K., Scharf, S., Faloona, F., Mullis, K. B., Horn, G. T., Erlich, H. A. and Arnheim, N. (1985). Enzymatic Amplification of Qβ - Globin Genomic Sequence and Restriction Site Analysis for Diagnosis of Sickle Cell Anemia. Science 230, 1350-1354.
5. Saiki, R. K., Gelfand, D. H., Stoffel, S., Scharf, S. J., Higuchi, R. , Horn, G. T., Mullis, K. B. and Eriich, H. A. (1988). Primer-directed Enzymatic Amplification of DNA with a Thermostable DNA Polymerase. Science 239, 487-491.
6. Guatelli, J. C. , Whitfield, K. M., Kwoh, D. Y., Barringer, K. J., Richman, D. D. and Gingeras, T. R. (1990). Isothermal, in vitro Amplification of Nucleic Acids by a Multienzyme Reaction Modeled after Retroviral Replication. Proc. Natl. Acad. Sci. U.S.A. 87, 1874-1878.
7. Levisohn, R. and Spiegelman, S. (1968). The Cloning of a Self-replicating RNA Molecule. Proc. Natl. Acad. Sci. U.S.A. 60, 866-872 (прототип).
8. Sumper, M. and Luce, R. (1975). Evidence for de novo Production of Self-replicating and Environmentally Adapted RNA Structures by Bacteriophage Qβ Replicase. Proc. Natl. Acad. Sci. U.S.A. 72, 162-166.
9. Chetverin, A. B., Chetverina, H. V. and Munishkin, A. V. (1991). On the Nature of Spontaneous RNA Synthesis by Qβ Replicase. J. Mol. Biol. 222, 3-9.
10. Myers, T. W. and Gelfand, D. H. (1991). Reverse Transcription and DNA Amplification by a Thermus thermophilus DNA Polymerase. Biochemistry 30, 7661-7666.
11. Kaplan, J. H. , Forbush, B., III and Hoffman, J. F. (1978). Rapid Photolytic Release of Adenosine 5'-Triphosphate from a Protected Analogue: Utilization by the Na:K Pump of Human Red Blood Cell Ghosts. Biochemistry 17, 1929-1935.
12. O'Driscoll, K. F. (1976). Techniques, of Enzyme Entrapment in Gels. Methods Enzymol. 44, 169-183.
13. Nojima, S. and Yamada, T. (1987). Large-scale Production of Photo-cross-linkable Resin-immobilized Yeast and Its Application to Industrial Ethanol Production. Methods Enzymol. 136, 380-394.
14. Osterman, L. A. (1986). Methods of Protein and Nucleic Acid Research, Vol. 3. Springer-Verlag, New York.
15. Landegren, U. , Kaiser, R., Caskey, C. T. and Hood, L. (1988). DNA Diagnostics - Molecular Techniques and Automation. Science 242, 229-237.
16. Anderson, C. W., Straus, J. W. and Dudock, B. S. (1983). Preparation of a Cell-free Protein-synthesizing System from Wheat Germ. Methods Enzymol. 101, 635-644.
17. Chambliss, G. H., Henkin, T. M. and Leventhal, J. M. (1983). Bacterial in vitro Protein-synthesis Systems. Methods Enzymol. 101, 598-605.
18. Merrick, W. C. (1983). Translation of Exogenous mRNAs in Reticulocyte Lysates. Methods Enzymol. 101, 606-615.
19. Chen, H.-Z. and Zubay, G. (1983). Prokaryotic Coupled Transcription-translation. Methods Enzymol. 101, 674-690.
20. Bujard, H., Gentz, R., Lanzer, M., Stueber, D., Mueller, M., Ibrahimi, I. , Haeuptle, M. -T. and Dobberstein, B. (1987). A T5 Promoter-based Transcription-translation System for the Analysis of Proteins in vitro and in vivo. Methods Enzymol. 155, 416-433.
21. Tymms, M. J. and McInnes, B. (1988). Efficient in vitro Expression of Interferon Analogs Using SP6 Polymerase and Rabbit Reticulocyte Lysate. Gene Anal. Tech. 5, 9-15.
22. Baranov, V. I., Morozov, I. Yu., Ortlepp, S. A. and Spirin, A. S. (1989). Gene Expression in a Cell-free System on the Preparative Scale, Gene 84, 463-466.
23. Spirin, A. S. (1992). Cell-free Protein Synthesis Bioreactor. In Frontiers in Bioprocessing II, ed. by P. Todd, S. K. Sikdar and M. Bier. American Chemical Society, Washington, DC, pp. 31- 43.
24. Blumenthal, T. (1979). Qβ RNA Replicase and Protein Synthesis Elongation Factors EF-Tu and EF-Ts. Methods Enzymol. 60, 628-638.
25. Jones, R. W. and Jones, M. J. (1992). Simplified Filter Paper Sandwich Blot Provides Rapid, Background-free Northern Blots. BioTechniques 12, 685-688.
26. Bodkin, D. K. and Knudson, D. L. (1985). Sequence Relatedness of Palyam Virus Genes to Cognates of the Palyam Serogroup Viruses by RNA-RNA Blot Hybridization. Virology 143, 55-62.
27. Weier, H. U. and Gray, J. W. (1988). A Programmable System to Perform the Polymerase Chain Reaction. DNA 7, 44-47.
28. Torgensen, H. , Blaas, D. and Skern, T. (1989). Low Cost Apparatus for Primer-directed DNA Amplification Using Thermus aquaticus-DNA Polymerase. Analyt. Biochem. 176, 33-35.
29. Kessler, C. and Holtke, H. J. (1986). Specificity of Restriction Endonucleases and Methylases -- A Review. Gene 47, 1-153.
30. Williams, J. G. (1981). The Preparation and Screening of a cDNA Clone Bank. In Genetic Engineering l (ed. by R. Williamson), pp. 1-59, Academic Press, London.
31. Higgins, N. P., Gebale, A. P. and Cozzarelli, N. R. (1979). Addition of Oligonucleotides to the 5'-Terminus of DNA by T4 RNA Ligase. Nucleic Acids Res. 6, 1013-1024.
32. Leary, J. J., Brigati, D. J. and Ward, D. C. (1983). Rapid and Sensitive Colometric Method for Visualizing Biotin-labeled DNA Probes Hybridized to DNA or RNA immobilized on Nitrocellulose: Bio-blots. Proc. Natl. Acad. Sci. U.S.A. 80, 4045-4049.
33. Campbell, A. K., Patel, A. K., Razavi, Z. S. and McCapra, F. (1988). Formation of the Ca -activated Photoprotein Obelin from Apo-obelin and mRNA Inside Human Neurophils. Biochem. J. 252, 143-149.
Использование: биотехнология, молекулярная биология. Сущность изобретения: клонирование нуклеиновых кислот осуществляют контактированием in vitro нуклеиновых кислот с компонентами среды, содержащей бесклеточную ферментную систему и субстраты, необходимые для экспоненциального размножения нуклеиновых кислот, далее жидкую фазу среды иммобилизируют путем ее заключения в твердую основу, обладающую развитой поверхностью со средним размером пор от 100 мкм до 5 нм, проводят инкубацию в условиях, обеспечивающих экспоненциальное размножение нуклеиновых кислот, и осуществляют обнаружение и анализ полученных колоний нуклеиновых кислот. 19 з.п.ф-лы, 2 ил.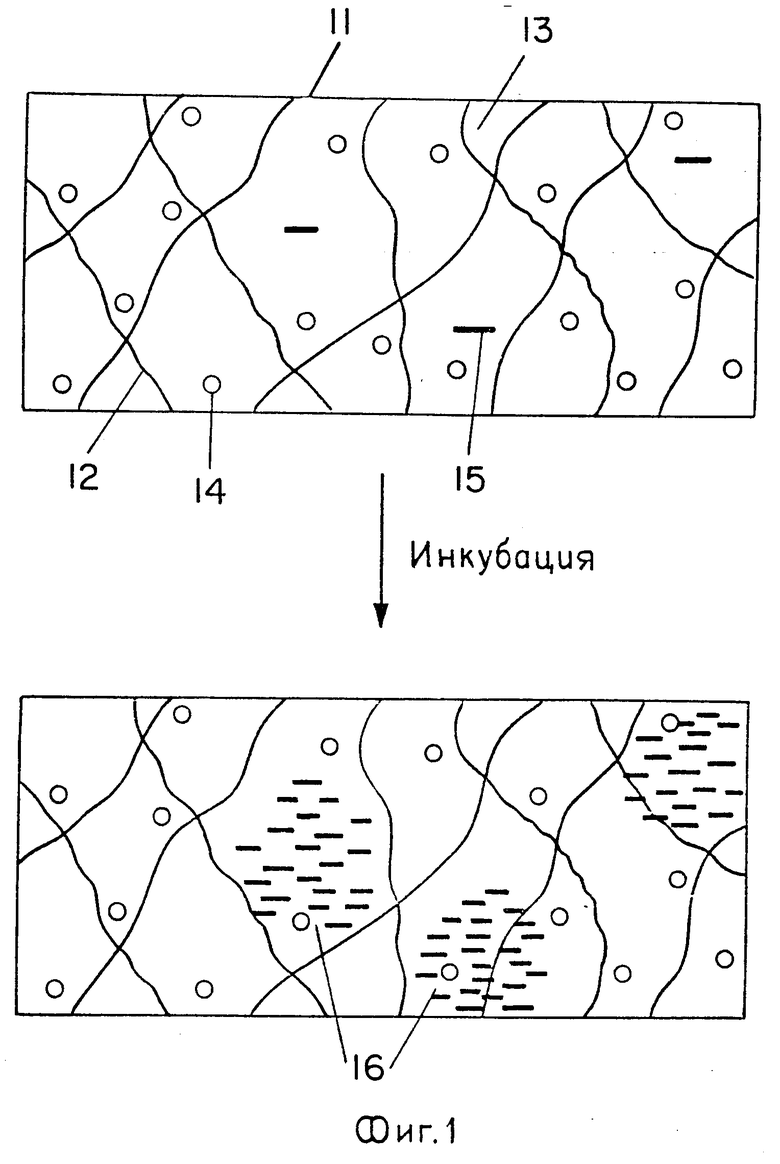
| National Academy of Sciences, 1968, USA, 60, 866-872. |
