ОБЛАСТЬ ТЕХНИКИ
Изобретение относится к области молекулярной биологии и биотехнологии, а именно к способам обнаружения специфических РНК-мишеней.
УРОВЕНЬ ТЕХНИКИ
Одним из направлений молекулярной диагностики является размножение искомой молекулярной мишени с помощью подходящей бесклеточной ферментативной системы экспоненциального размножения полинуклеотидов. Как правило, для размножения специфических РНК-мишеней (например, вирусных РНК или определенных клеточных мРНК) осуществляют синтез комплементарных ДНК (кДНК) с помощью обратной транскриптазы (РНК-зависимой ДНК-полимеразы) и праймеров, способных гибридизоваться с искомыми мишенями с 3'-стороны от копируемого участка. В качестве праймеров можно использовать, например, специфические праймеры (то есть, олигонуклеотиды, комплементарные уникальным участкам копируемых мишеней), смесь случайных олигонуклеотидов (например, гексануклеотидов) или олиго(dT), если РНК-мишень содержит поли(А)-последовательность на 3'-конце. Полученную кДНК размножают до детектируемых количеств с использованием полимеразной цепной реакции (ПНР [1]) или одной из многочисленных изотермических систем размножения, таких как 3SR (Self-Sustained Sequence Replication [2]), SDA (Strand Displacement Amplification [3]), NASBA (nucleic acid sequence-based amplification [4]), RCA (rolling circle amplification [5, 6]) и LAMP (loop-mediated isothermal DNA amplification [7]). Термин "система размножения" означает полный набор реагентов и иных компонентов, необходимых для синтеза нуклеиновых кислот на определенного вида матрицах (мишенях), в том числе один или более ферментов, включающих по крайней мере одну полимеразу нуклеиновых кислот; субстраты полимеризации (рибо- и/или 2'-дезоксирибонуклеозидтрифосфаты); реакционный буфер; а также мишень-специфичные олигонуклеотидные праймеры, если синтез нуклеиновых кислот осуществляют с помощью праймер-зависимой полимеразы. Теоретически, с использованием такого подхода можно обнаружить единичные молекулы РНК данного вида. Однако на практике, при анализе клинических образцов, содержащих большое количество посторонних (отличных от мишени) нуклеиновых кислот, чувствительность оказывается в 100-1000 раз ниже вследствие неспецифического синтеза ДНК, происходящего из-за ограниченной специфичности гибридизации праймеров как на стадии обратной транскрипции, так и при последующем размножении кДНК. Размножение посторонних нуклеиновых кислот приводит также и к снижению специфичности диагностики, что в большой степени обесценивает результат анализа [8].
В технике известны различные способы понижения уровня неспецифического синтеза ДНК при определении РНК-мишеней.
Прежде всего, это снижение содержания ДНК в препарате РНК, используемом для обратной транскрипции. С этой целью используют методы выделения РНК, позволяющие удалить из препарата большую часть ДНК, такие как метод Хомчинского [9]. От остатков ДНК освобождаются, обрабатывая полученный препарат дезоксирибонуклеазой [10, 11]. Однако такая обработка не освобождает образец от многочисленных РНК, содержание которых может быть в миллионы и миллиарды раз больше, чем содержание искомой мишени.
Обогатить препарат РНК анализируемой мишенью можно с помощью аффинной очистки на носителях с иммобилизованным олиго(dT) (олиготимидилатом) [12]. Это позволяет выделить РНК, несущие поли(А)-последовательность на 3'-конце и, тем самым, освободиться от рибосомных РНК и тРНК, составляющих более 90% от суммарного препарата клеточной РНК. Однако осуществление этой стадии приводит к удорожанию процедуры анализа и связано с риском перекрестного загрязнения образцов. Кроме того, препарат оказывается загрязненным олиго(dT), который участвует в обратной транскрипции наряду со специфическим праймером [13].
Аналогичного результата можно добиться более простым способом, осуществляя обратную транскрипцию с использованием олиго(dT)-праймера [14], так как в этом случае исключается копирование РНК, лишенных поли(А)-последовательности (в частности, рибосомных РНК и тРНК). Однако у такого способа есть следующие недостатки. (1) Продуктом обратной транскрипции является сумма разнородных кДНК, из которых искомая кДНК составляет ничтожную часть. (2) Часть потенциальных РНК-мишеней лишены поли(А)-последовательности и не будут представлены в продуктах обратной транскрипции [15]. (3) Возможен синтез укороченных кДНК из-за гибридизации праймера с внутренними поли(А)-последовательностями [16].
Повысить специфичность обратной транскрипции можно используя термостабильные обратные транскриптазы и осуществляя обратную транскрипцию при повышенной температуре с целью разрушения гибридов праймера с посторонними РНК, обладающих меньшей стабильностью, чем совершенный гибрид с РНК-мишенью. Например, используя обратные транскриптазы, продаваемые фирмой Invitrogen (США) под торговыми марками Superscript™ и ThermoScript™, можно осуществлять обратную транскрипцию при температуре 50 и 65°С, соответственно. Однако и в этом случае не удается полностью исключить гибридизацию праймера с посторонними матрицами и, следовательно, неспецифический синтез ДНК.
Из вышеизложенного следует, что существует потребность в усовершенствованных способах повышения специфичности размножения РНК-мишеней. Настоящее изобретение направлено на решение данной задачи посредством избирательной деградации РНК, отличных от искомой мишени.
Наиболее близким аналогом (прототипом) данного изобретения является вышеупомянутый способ удаления примесей ДНК из препаратов РНК с помощью дезоксирибонуклеазы [10].
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Указанная техническая задача повышения специфичности обратной транскрипции реализуется посредством нового способа избирательной деградации РНК с использованием препарата полинуклеотидфосфорилазы (ПНФазы), практически свободного от эндонуклеазной и гидролазной активностей.
Одним из аспектов изобретения является то, что до начала проведения обратной транскрипции образец, который содержит хотя бы один вид подлежащей обратной транскрипции РНК (мишени) и хотя бы один вид РНК, отличной от мишени, отжигают хотя бы с одним защитным олигонуклеотидом для защиты РНК мишени от действия полинуклеотидфосфорилазы (ПНФазы). При этом олигонуклеотид гибридизуют с соответствующей РНК-мишенью на 3'-стороне от участка, подлежащего обратной транскрипции. Обработанный образец инкубируют в растворе, содержащем термостабильную ПНФазу, ее косубстрат и кофактор, при таких значениях температуры, времени инкубации, концентрации ПНФазы, косубстрата и кофактора, которые обеспечивают существенно полную сохранность защищенных РНК-мишеней и существенно полную деградацию РНК, отличных от мишеней. При этом в качестве ПНФазы используют, например, ПНФазу Thermus thermophilus.
Другой аспект изобретения состоит в том, что защитный олигонуклеотид модифицируют с целью повышения стабильности его гибрида с РНК-мишенью. Модификацию олигонуклеотида осуществляют, например, путем замены хотя бы одного из его нуклеотидов на LNA-аналог.
Другим аспектом изобретения является то, что продукты обратной транскрипции размножают с использованием бесклеточной ферментативной системы экспоненциального размножения полинуклеотидов.
КРАТКОЕ ОПИСАНИЕ ФИГУР
Фиг.1 демонстрирует зависимость фосфоролиза РНК Tth-ПНФазой от температуры.
Фиг.2 демонстрирует селективность и полноту деградации незащищенной РНК Tth-ПНФазой.
Фиг.3 демонстрирует защитный эффект олигонуклеотида, гибридизованного к внутреннему участку РНК, на ее деградацию Tth-ПНФазой.
Фиг.4 демонстрирует последовательность, защищаемую от Tth-ПНФазы олигонуклеотидом, гибридизованным к внутреннему участку РНК.
Фиг.5 демонстрирует эффективную защиту РНК олигонуклеотидом, содержащим LNA-модификации.
Фиг.6 демонстрирует кинетику и избирательность деградации Tth-ПНФазой незащищенной высокомолекулярной РНК.
Фиг.7 демонстрирует повышение специфичности ОТ-ПЦР защищенной РНК в результате обработки препарата РНК Tth-ПНФазой перед стадией обратной транскрипции.
ОПРЕДЕЛЕНИЯ ТЕРМИНОВ
Термин «полинуклеотидфосфорилаза» («ПНФаза») используется здесь для обозначения 3'→5' экзорибонуклеазы, способной катализировать реакцию фосфоролиза полирибонуклеотида, отщепляя нуклеотиды от его 3'-конца, независимо от того, способен ли этот фермент также катализировать реакцию фосфоролиза поли-2'-дезоксирибонуклеотида или иного типа нуклеотидной последовательности.
Термин «LNA-аналог нуклеотида» используется здесь для обозначения нуклеотида, рибозное кольцо которого зафиксировано в 3'-эндо-конфигурации путем введения 2'-O, 4'-С-метиленового мостика.
ОСУЩЕСТВЛЕНИЕ ИЗОБРЕТЕНИЯ
Полинуклеотидфосфорилаза (ПНФаза) - это 3'→5' экзорибонуклеаза, осуществляющая обратимый фосфоролиз полирибонуклеотида (pN)n с 3'-конца в соответствии с уравнением: (pN)n+Pi↔(pN)n-1+ppN, где Pi - неорганический ортофосфат (косубстрат фосфоролиза), а ppN - нуклеозид-5'-дифосфат [17]. Для функционирования ПНФазы необходимы кофакторы - ионы Mg2+, которые могут быть заменены другими двухвалентными катионами, такими как Со2+, Ni2+, Cd2+, Cu2+ и Zn2+, но не Са2+. Вместо Pi в качестве косубстрата ПНФаза может использовать арсенат. В этом случае реакция становится практически необратимой, так как образующиеся нуклеозид-5'-фосфориларсенаты спонтанно гидролизуются до нуклеозидмонофосфатов и арсената [18].
ПНФаза была обнаружена во всех типах бактерий [18], за исключением микоплазмы [19]. Хотя последовательностей, кодирующих ПНФазу, нет в геномах архей и дрожжей, изученных к настоящему времени (эти геномы, тем не менее, содержат гены похожего фермента - РНКазы РН), они присутствуют в высших эукариотах [20-22]. ПНФазы растений и животных кодируются ядерными генами, но локализуются, главным образом, в органеллах [23]. В растениях есть два вида ПНФазы, один из которых локализован в хлоропластах, а другой - в митохондриях [20, 24].
Ген ПНФазы (pnp) кодирует полипептид массой 75-100 кДа, состоящий из пяти эволюционно консервативных доменов: двух N-концевых «коровых» доменов, гомологичных РНКазе РН Е.coli, разделенных α-спиральным доменом, и двух С-концевых РНК-связывающих доменов - КН и S1 [19-23, 25]. РНКаза РН, как и ПНФаза, представляет собой фосфоролитическую 3'→5' экзорибонуклеазу. В клетках Е.coli РНКаза РН ответственна за процессинг 3'-концов молекул предшественников тРНК [26]. Домен КН («K-homology») был исходно идентифицирован в РНК-связывающем белке К человека [27], а домен S1 назван так из-за гомологии с РНК-связывающим белком S1 рибосом Е.coli [28]. Молекулы ПНФаз бактерий Е.coli и Streptomyces antibioticus состоят из трех идентичных субъединиц, которые образуют структуру в виде бублика с отверстием в центре, где может помещаться цепь РНК [25, 29], а ПНФаза хлоропластов состоит из двух таких бубликов, положенных один на другой [20, 30].
В дрожжах и археях функциональными и структурными аналогами ПНФазы являются экзосомы. Экзосомы состоят из субъединиц, являющихся гомологами доменов ПНФазы, и выполняют такие же функции, катализируя обратимый фосфоролиз полирибонуклеотидов. Поэтому экзосома может быть рассмотрена как разновидность ПНФазы. В пользу этого говорит также то, что трехмерные структуры бактериальной ПНФазы [25] и РНКазы РН, составляющей «кор» (основу) архейной экзосомы [31], высоко гомологичны [32].
Настоящее изобретение основано на результатах работы по клонированию гена Tth-ПНФазы, его экспрессии в клетках Е.coli и изучению биохимических свойств выделенного фермента [33, 34]. Фермент выделяли как описано в источнике [33]. В процессе исследования биохимических свойств выделенного фермента неожиданно было установлено, что выделенная Tth-ПНФаза явилась первой очищенной термостабильной ПНФазой, способной к осуществлению реакции фосфоролиза полирибонуклеотидов.
В присутствии 40 мкМ Mg2+ оптимальная температура фосфоролиза составляет 65°С, тогда как в отсутствие ПНФазы РНК остается практически целой даже при 85°С (фиг.1). Фиг.1 также демонстрирует, что рабочий диапазон температур, в котором Tth-ПНФаза проявляет существенную фосфорилазную активность, составляет 60-75°С. Это выше, чем максимальная температура, при которой проявляют фосфорилазную активность мезофильные ПНФазы, такие как ПНФаза Е.coli, - 55°С [18].
С помощью ОТ-ПЦР (обратной транскрипции и последующей ПЦР) установлено, что в оптимальных условиях (температура, концентрации Mg2+ и EDTA, соотношение фермента к РНК, время реакции) Tth-ПНФаза разрушает более 99% высокоструктурированной РНК, не повреждая РНК, 3'-конец которой защищен комплементарным олигодезоксирибонуклеотидом (фиг.2А). Этот результат указывает на то, что олигонуклеотид, гибридизованный с 3'-концом РНК, мог бы селективно защищать ее от Tth-ПНФазы. Следующий эксперимент (фиг.2Б) демонстрирует, что это действительно так. В этом эксперименте смесь двух видов РНК - 5'- и 3'-фрагментов RQ135 РНК [35, 36] - отожгли с олигодезоксирибонуклеотидом, комплементарным 3'-концу 3'-фрагмента. Последующий фосфоролиз с Tth-ПНФазой привел к полной деградации 5'-фрагмента и получению практически чистого 3'-фрагмента (если не считать сам олигонуклеотид, который можно удалить разными способами, например, с помощью хроматографии, электрофореза или обработки ДНКазой).
На фиг.3 представлены результаты деградации трех РНК, имеющих одинаковую 5'-концевую часть, но в разной мере удлиненных на 3'-конце. Все три РНК подвергли фосфоролизу Tth-ПНФазой после отжига в присутствии или в отсутствие 38-нт олигодезоксирибонуклеотида, комплементарного 3'-концевой последовательности наиболее короткой РНК («SmaI») и внутренним последовательностям более длинных РНК («EcoRI» и «PvuII»). Видно, что олигонуклеотид полностью защищает самую короткую РНК и частично защищает более длинные РНК. Деградация последних дает устойчивый продукт, подвижность которого при неденатурирующем электрофорезе несколько меньше, чем подвижность гибрида олигонуклеотида с РНК SmaI (фиг.3, слева). Такой продукт не образуется в отсутствие олигонуклеотида (фиг.3, справа).
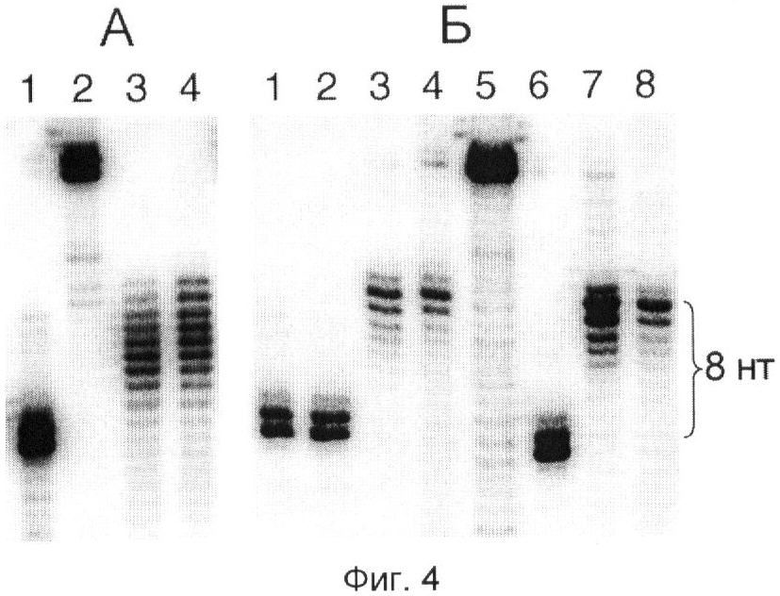
Тонкую структуру продуктов фосфоролиза анализировали с помощью электрофореза высокого разрешения в денатурирующих условиях (фиг.4). Перед фосфоролизом каждую из трех РНК дефосфорилировали и затем метили на 5'-конце с помощью [γ-32P]ATP и полинуклеотидкиназы. Из фиг.4А видно, что устойчивый продукт деградации более длинных РНК имеет довольно гетерогенный 3'-конец и до 8 нт длиннее, чем РНК с защищенной концевой последовательностью. Последняя также имеет гетерогенный 3'-конец, но это обычно для РНК, полученных с помощью транскрипции обрезанных ДНК («run-off transcription») [37]. Наблюдаемая гетерогенность продуктов фосфоролиза по большей части является следствием гетерогенности защитного олигонуклеотида, так как ее удается существенно уменьшить при использовании олигонуклеотида с точным 5'-концом. Такой защитный олигонуклеотид получали обрезая с помощью эндонуклеазы рестрикции олигонуклеотид, удлиненный на 5'-конце [33, 34]. Фосфоролиз EcoRI- и PvuII-PHK, гибридизованных с таким 5'-обрезанным олигонуклеотидом, давал продукт, чья 3'-концевая гетерогенность была сравнима с гетерогенностью продукта транскрипции, и который был на 8 нт длиннее, чем РНК SmaI с защищенной концевой последовательностью (фиг.4Б). Отсюда следует, что Tth-ПНФаза останавливается на разрушаемой РНК на расстоянии 8 нт от 5'-конца олигонуклеотида, гибридизованного с внутренней последовательностью РНК. Присутствие фосфатной группы на 5'-конце олигонуклеотида не влияло на длину защищаемого участка РНК (фиг.4Б; ср. дорожки 3 и 4 с дорожками 7 и 8).
С одной стороны, полученные результаты позволяют надеяться, что избирательную деградацию ПНФазой незащищенной РНК можно использовать для уничтожения РНК, отличных от мишени, и, следовательно, для повышения специфичности обратной транскрипции. С другой стороны, результаты, представленные на фиг.2, показывают, что олигонуклеотид, гибридизованный с внутренней последовательностью РНК, защищает ее не полностью. Действительно, в отличие РНК SmaI, гибридизованной с олигонуклеотидом своей 3'-концевой последовательностью, количество РНК EcoRI и - в еще большей степени - более длинной РНК PvuII заметно падает после обработки Tth-ПНФазой (см. полосы, отмеченные белыми стрелками). Это наблюдение существенно ограничивает применимость данного подхода для молекулярной диагностики, так как, с одной стороны, анализируемые мишени часто не имеют строго определенных 3'-концов, а, с другой стороны, последовательность мишени, представляющая интерес для диагностики, часто находится вдали от 3'-конца. В последнем случае, результат анализа будет подвержен действию даже ничтожных примесей эндонуклеаз, поскольку даже одиночный разрыв в области, лежащей между 3'-концом и анализируемой последовательностью, приведет к полному снятию защиты.
Оказалось, что простое увеличение длины защитного олигонуклеотида (и, следовательно, температуры плавления гибрида) не улучшает ситуацию. Мы предположили, что неполнота защиты является следствием двух факторов: (1) известно, что ПНФаза является высокопроцессивным ферментом (то есть, не диссоциирует от РНК, пока не разрушит ее полностью, вплоть до 5'-конца) [18], и (2) вероятно, что, даже в отсутствие полной диссоциации защитного олигонуклеотида, гибрид может "дышать": иногда 5'-конец защитного олигонуклеотида отходит от РНК. В этот момент молекула ПНФазы, остановленная на расстоянии 8 нт от 5'-конца олигонуклеотида, может занять временно освободившееся место. После ряда таких событий ПНФаза разрушит весь сегмент РНК, комплементарный защитному олигонуклеотиду, а затем и остальную часть РНК. Такой механизм помогает объяснить зависимость защитного эффекта олигонуклеотида от того, гибридизован он с 3'-концевой или с внутренней последовательностью РНК.
Если предложенный механизм верен, то защитный эффект олигонуклеотида, гибридизованного с внутренней последовательностью, можно увеличить, повысив стабильность гибрида. Известно несколько модификаций олигонуклеотидов, повышающих стабильность их гибридов с комплементарными цепями РНК и ДНК. Например, температура плавления гибридов, образованных PNA (peptide nucleic acid)-аналогами олигонуклеотидов, в которых сахарофосфатный остов заменен на полиамидный [38-40], существенно выше, чем гибридов, образованных обычными (природными) олигонуклеотидами с такой же длиной и нуклеотидной последовательностью [41]. К существенному повышению стабильности гибридов приводит также замена обычных рибозных колец нуклеотидов на некоторые их би-три- или полициклические аналоги; многие из таких модификаций раскрыты в патентах [42-44]. Наиболее популярными стали так называемые LNA (locked nucleic acid)-аналоги нуклеотидов, рибозные кольца которых зафиксированы в 3'-эндо-конфигурации, характерной для А-формы двойной спирали РНК, путем введения 2'-O, 4'-С-метиленового мостика [45]. Введение одного LNA-аналога вместо обычного нуклеотида приводит к повышению на 2-10°С температуры плавления гибрида длиной до 10 нт, причем суммарное повышение температуры плавления может достигать несколько десятков градусов [46].
Фиг.5 демонстрирует, что замена 6 из 38 нуклеотидов олигодезоксирибонуклеотида на их LNA-аналоги приводит к существенному повышению защиты комплементарной РНК от Tth-ПНФазы. В данном эксперименте приблизительно одинаковую эффективность показали как олигонуклеотид, в котором все замены были сгруппированы вблизи 5'-конца (LNA-1), так и олигонуклеотид, в котором замены были рассредоточены по длине (LNA-2).
Фиг.6 демонстрирует результаты эксперимента, в котором Tth-ПНФазой обрабатывали смесь высокомолекулярных рибосомных РНК Е.coli (длиной 1542 и 2904 нт) и мРНК маммаглобина (маркера рака молочной железы) длиной 486 нт, защищенной LNA-модифицированным олигонуклеотидом на расстоянии 18 нт от 3'-конца. Видно, что рибосомные РНК практически полностью разрушаются в течение 5 мин, в то время как защищенная РНК остается целой. При дальнейшей инкубации олигонуклеотид, в котором LNA-аналоги сгруппированы у 5'-конца (LNA-2), демонстрирует лучшую защиту.
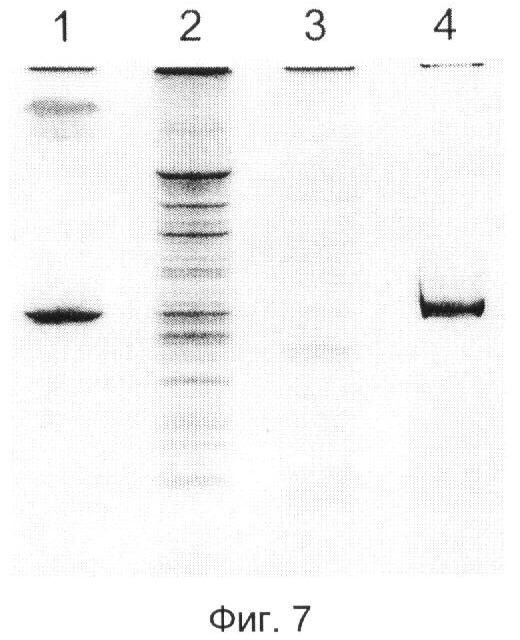
Наконец, фиг.7 демонстрирует эффективность предложенного здесь подхода в подавлении неспецифического синтеза ДНК при обратной транскрипции и последующей ПНР (ОТ-ПЦР). Осуществление ОТ-ПЦР с использованием суммарной РНК, выделенной из цельной крови, и праймеров, специфичных к мРНК теломеразы человека (hTERT, вероятного универсального маркера рака), дает гетерогенный набор неспецифических продуктов (дорожка 2), в том числе существенно большего размера, чем искомый фрагмент кДНК hTERT (203 п.о, дорожка 1). Обработка препарата суммарной РНК крови Tth-ПНФазой перед стадией обратной транскрипции резко снижает количество неспецифических продуктов (дорожка 3). В то же время, обработка фрагмента мРНК hTERT длиной 486 нт, защищенной LNA-модифицированным олигонуклеотидом на расстоянии 36 нт от 3'-конца, не снижает выход специфического продукта (дорожка 4).
Таким образом, благодаря деградации незащищенных РНК Tth-ПНФазой можно существенно подавить неспецифический синтез ДНК при обратной транскрипции и тем самым повысить специфичность размножения (а, следовательно, и чувствительность детекции) РНК-мишени, которую защищают от ПНФазы олигонуклеотидом, гибридизующимся с РНК-мишенью по 3'-сторону от участка, подлежащего обратной транскрипции. Конечно, сам защитный олигонуклеотид может быть использован в качестве праймера при обратной транскрипции, а также при возможном последующем размножении кДНК с помощью выбранной бесклеточной ферментной системы экспоненциального размножения полинуклеотидов. В то же время, для защиты от ПНФазы можно использовать отдельный олигонуклеотид, при этом предпочтительно удалять или блокировать его 3'-гидроксильную группу (например, путем фосфорилирования), чтобы исключить его последующее использование полимеразами в качестве праймера.
Хотя в приведенных примерах для защиты РНК использовали олигодезоксирибонуклеотиды, такую же защиту могут обеспечить олигорибонуклеотиды, смешанные олигонуклеотиды, в которых присутствуют дезоксирибо- и рибонуклеотидные звенья, а также аналоги олигонуклеотидов, в которых природный сахарофосфатный остов заменен на неприродный полимер, например, содержащий модифицированные фосфодиэфирные связи, неприродный сахарный компонент, химические аналоги фосфодиэфирной связи, полностью измененный остов, остов с 2'-5'-межнуклеотидными связями или составленный из α-нуклеотидных аномеров. Многочисленные примеры такого рода представлены в Патенте США №6329144 [47]. Примеры неприродных олигомеров, которые способны к правильному спариванию по Уотсону и Крику с комплементарным участком нуклеиновой кислоты-мишени, включают, но не исчерпываются такими химическими аналогами, как фосфоротиоатные, метилфосфонатные, боранфосфонатные производные нуклеиновых кислот [48, 49], уже упомянутые пептидные нуклеиновые кислоты [38-40], морфолиновые олигонуклеотиды [50, 51]. Для защитного действия олигонуклеотида или его аналога необходимо и достаточно, чтобы он был способен гибридизоваться с РНК посредством спаривания азотистых оснований по Уотсону и Крику и чтобы образующийся гибрид имел существенную стабильность при температуре реакции фосфоролиза.
Очевидно, что, помимо Tth-ПНФазы, с той же целью могут быть использованы другие термостабильные ПНФазы, например, ПНФаза T.aquaticus. Более того, учитывая большое структурное и функциональное сходство бактериальных ПНФаз [25] и экзосом архей S. solfataricus [31] и Archaeoglobus fulgidus [52], можно ожидать, что архейные экзосомы обладают биохимическими свойствами, подобными свойствам Tth-ПНФазы, и также могут быть использованы для указанной выше цели. Было показано, что РНКаза РН S. solfataricus и A. fulgidus, формирующая «кор» экзосомы, сохраняет фосфорилазную активность при 60°С [31, 52] и даже при 70°С [53].
Однако фосфорилазная активность была продемонстрирована только в отношении неструктурированных олиго(А) последовательностей, в то время как РНКаза РН S. solfataricus оказалась неспособной разрушить шпильку с черешком из 8 G:C-пap даже при инкубации в течение 30 мин при 70°С [53]. Возможно, неспособность разрушать структурированные РНК является результатом того, что в «кор» экзосомы отсутствуют субъединицы, гомологичные РНК-связывающим доменам КН и S1 ПНФазы. Можно надеяться, что полная архейная экзосома, содержащая также КН- и S1-подобные субъединицы, будет эффективно разрушать структурированные РНК при высокой температуре.
Для применения предложенного способа важно использовать термостабильную ПНФазу и осуществлять реакцию в условиях [температура, концентрация ПНФазы, ее косубстратов (помимо РНК, таких как Pi) и кофакторов (таких как Mg2+), продолжительность инкубации], при которых достигается полная деградация незащищенной РНК. Не существенно, обладает ли термостабильностью ПНФаза, выделенная из природного источника, или ее термостабильность повышена с помощью методов генетической инженерии, в результате селекции, химической модификации или иными способами, известными в технике.
Хотя в приведенных здесь примерах стадия инкубации РНК с ПНФазой предшествует стадии обратной транскрипции, возможно частичное перекрывание этих стадий во времени (то есть, начало обратной транскрипции до окончания реакции фосфоролиза), особенно при использовании термостабильной обратной транскриптазы, такой как ThermoScript™ (Invitrogen, США), способной работать при температуре, оптимальной для Tth-ПНФазы (65°С).
Предложенный способ поясняется, но не ограничивается, следующими конкретными примерами его осуществления.
Пример 1. Препараты РНК
В данной работе использовали следующие препараты РНК:
(1) CT1n1 РНК, вариант RQ135-1 РНК [35], несущий 43-нт вставку (подчеркнута), выделенный из продуктов межмолекулярной рекомбинации РНК:

Препарат CT1n1 РНК получают транскрипцией плазмиды, несущей рекомбинантную кДНК, расщепленной по сайту SmaI.
(2) 5'-фрагмент RQ135 РНК: 5'-концевой фрагмент RQ135-1 РНК [35] длиной 52 нт, несущий чужеродный довесок длиной 23 нт (подчеркнут):

Препарат 5' фрагмента получают транскрипцией плазмиды, кодирующей 5'-фрагмент [54], расщепленной по сайту BamHI.
(3) 3'-фрагмент RQ135 РНК (транскрипт SmaI): 3'-концевой фрагмент RQ135-1 RNA [35] длиной 81 нт, несущий чужеродный довесок длиной 28 нт (подчеркнут):

Препарат 3' фрагмента получают транскрипцией плазмиды, кодирующей 3'-фрагмент [54], расщепленной по сайту SmaI.
(4) Транскрипт EcoRI [3'-фрагмент RQ135 РНК, удлиненный на 3'-конце последовательностью длиной 18 нт (подчеркнута)]:

Препарат этой РНК получают транскрипцией плазмиды, кодирующей 3'-фрагмент [54], расщепленной по сайту EcoRI.
(5) Транскрипт PvuII [3'-фрагмент RQ135 РНК, удлиненный на 3'-конце последовательностью длиной 194 нт (подчеркнута)]:

Препарат этой РНК получают транскрипцией плазмиды, кодирующей 3'-фрагмент [54], расщепленной по сайту PvuII.
(6) мРНК маммаглобина (секретоглобина SCGB2A2; подчеркнута последовательность, комплементарная защитному олигонуклеотиду):

Препарат этой РНК получают транскрипцией плазмиды, кодирующей 8ССВ2А2-кДНК, расщепленной по сайту EcoRI.
(7) Фрагмент мРНК теломеразы человека [hTERT; одинарной линией подчеркнута последовательность, комплементарная защитному олигонуклеотиду; двойными линями подчеркнуты участки, соответствующие ОТ-ПЦР-праймерам]:
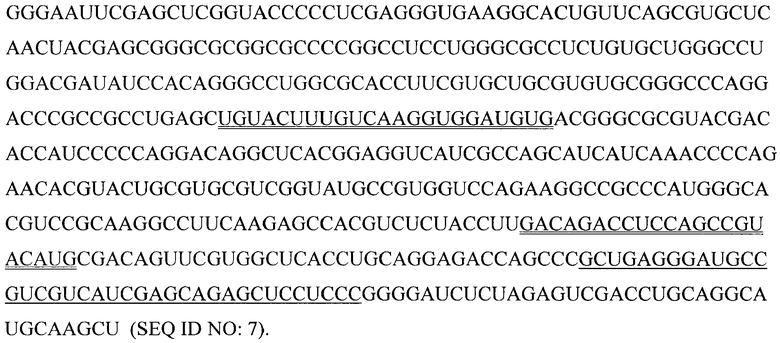
Препарат этой РНК получают транскрипцией плазмиды, кодирующей hTERT-кДНК, расщепленной по сайту HindIII.
Все препараты РНК получают с помощью Т7 РНК-полимеразы и очищают с помощью электрофореза в полиакриламидном геле, как описано ранее [54].
Пример 2. Выделение суммарной РНК из цельной крови человека
Метод является модификацией известного метода Хомчинского [9], разработанного для выделения РНК из гомогенизированных тканей. С учетом специфики крови, мы ввели в лизирующий раствор ЭДТА, дополнительно добавляли 2-меркаптоэтанол (перед размораживанием образцов), а также ввели дополнительную экстракцию фенолом из водного раствора и стадию переосаждения РНК смесью ацетона и этанола. Эти изменения повысили выход и сохранность РНК, улучшили растворимость конечного препарата, а также понизили содержание примесей, ингибирующих ОТ и ПЦР.
Если не указано иное, все операции проводят при комнатной температуре в микроцентрифужных пробирках типа "Эппендорф", а центрифугирования осуществляют на холоде (0-4°С) в центрифугах MiniSpin™ (Eppendorf, Германия) на максимальной скорости (13400 об/мин), используя для выделения аликвоты лизата, полученного следующим образом.
В пробирку с 1,5 мл консервирующего раствора [5,33 М тиоцианат гуанидина, 33,3 мМ цитрат натрия (рН 7,5), 0,67% саркозил, 17,8 мМ ЭДТА, 1,8% 2-меркаптоэтанол (конечные концентрации: 4 М тиоцианат гуанидина, 25 мМ цитрат натрия, 0,5% саркозил, 13 мМ ЭДТА, 1,3% 2-меркаптоэтанол] добавляют 0,5 мл свежесобранной крови и перемешивают. Полученную суспензию хранят при температуре -20°С в темноте.
Ниже описана детальная процедура для выделения РНК из 0,6 мл лизата; при использовании другого объема лизата объемы всех реагентов пропорционально увеличивают. Все растворы и воду автоклавируют. Все перемешивания осуществляют на встряхивателе Reax-Top™ (Heidolph, Германия) при максимальной интенсивности (2500 об/мин).
Размораживание лизата. К замороженному лизату (2 мл) добавляют 26,7 мкл 2-меркаптоэтанола (из расчета 8 мкл на 600 мкл лизата). Лизат размораживают, перемешивают, центрифугируют в течение 15 с, отбирают в чистую пробирку 608 мкл; остальное замораживают и хранят при -20°С.
Первая экстракция фенолом. Добавляют 68 мкл 2 М ацетата натрия рН 4,0 (до 200 мМ), перемешивают в течение 10 с; добавляют 600 мкл фенола (рН 4,5, насыщенного водой), перемешивают в течение 10 с; добавляют 180 мкл смеси хлороформ/изоамиловый спирт (49: 1), перемешивают в течение 1 мин; инкубируют во льду в течение 15 мин, перемешивают в течение 10 с, центрифугируют при комнатной температуре в течение 20 мин; отбирают водную фазу (верхнюю) в чистую пробирку: 3 раза по 164 мкл (всего 492 мкл).
Осаждение изопропанолом. К водной фазе добавляют 690 мкл изопропанола, охлажденного до -20°С, смешивают переворачиванием 8-10 раз, перемешивают в течение 10 с, центрифугируют при 4°С в течение 15 сек; инкубируют при -20°С в течение 60 мин, центрифугируют при 4°С в течение 20 мин, надосадочную жидкость удаляют.
Промывание осадка. К осадкам добавляют на льду 1 мл охлажденной (-20°С) смеси этанола и 0,1 мМ ЭДТА (4:1), обмывают крышечку переворачиванием пробирки 4-5 раз, центрифугируют при 4°С в течение 10 мин, надосадочную жидкость удаляют, осадок высушивают при 45°С в течение 5 мин.
Вторая экстракция фенолом. Осадок растворяют в 180 мкл 0,5% SDS, добавляют 20 мкл 3М NaCl (до 300 мМ), перемешивают в течение 10 с; добавляют 200 мкл фенола (рН 4,5, насыщенного водой), перемешивают в течение 1 мин, центрифугируют при 22°С в течение 10 мин; отбирают 175 мкл водной фазы (верхней) в чистую пробирку.
Промывание экстракта. К водной фазе добавляют 175 мкл хлороформа, перемешивают в течение 1 мин, центрифугируют при 22°С в течение 1 мин; отбирают 165 мкл водной фазы (верхней) в чистую пробирку.
Осаждение смесью ацетона и спирта. К водной фазе добавляют 990 мкл охлажденной (-20°С) смеси ацетон : этанол (1:1), смешивают переворачиванием 8-10 раз, перемешивают в течение 10 сек, центрифугируют при 4°С в течение 15 с; инкубируют при -20°С в течение 20 мин; центрифугируют при 4°С в течение 20 мин, надосадочную жидкость удаляют.
Промывание осадка. К осадкам добавляют на льду 1 мл охлажденной (-20°С) смеси ацетон : этанол (1:1), обмывают крышку пробирки переворачиванием пробирки 4-5 раз, центрифугируют при 4°С в течение 10 мин, надосадочную жидкость удаляют, осадок высушивают при 45°С в течение 10 мин.
Растворение осадка. Осадок растворяют в 30 мкл 0,1 мМ ЭДТА, инкубируют при 65°С в течение 10 мин, перемешивают в течение 5 мин, центрифугируют в течение 1 мин; нагревают в течение 3 мин при на водяной бане, быстро переносят на лед; инкубируют в течение 5 мин, центрифугируют в течение 2 мин, перемешивают 3 раза в течение 1 с.
Гель-фильтрация. Полученный образец пропускают (с помощью центрифугирования) через уравновешенную в 0,1 мМ ЭДТА 0,6-мл колонку Sephadex G-25 DNA Grade (Amersham Biosciences), приготовленную в микроцентрифужной пробирке с прорезанным бритвой дном. Перед пропусканием образца всю свободную жидкость удаляют из колонки с помощью центрифугирования в тех же условиях. Образец хранят при -70°С.
Пример 3. Исследование зависимости деградации РНК от температуры
Реакция фосфоролиза. Реакционная смесь объемом 10 мкл имеет следующий состав: 50 мМ Hepes-KOH (рН 8,1), 75 мМ КС1, 0,1 мМ MgCl2, 1 мМ дитиотрейтол, 1 мМ Na-фосфат, 0,5 пмоль 3'-фрагмента RQ135 РНК. Сверх того, в каждый образец вместе с препаратами ПНФазы и РНК вносят 0,056 мМ EDTA. Реакцию осуществляют в течение 15 мин при указанных на фиг.1 температурах в присутствии или в отсутствие 0,5 пмоль Tth-ПНФазы и останавливают добавлением 10 мкл 10мМ Na-EDTA.
Анализ продуктов реакции. Нуклеиновые кислоты выделяют с помощью фенольной экстракции [55] и разделяют с помощью электрофореза в денатурирующих условиях: в полиакриламидном геле, содержащем 7М мочевину, в буфере ТВЕ (100 мМ Tris-OH, 100 мМ борная кислота, 2 мМ Na-EDTA). После электрофореза гель окрашивают серебром [56]. Фиг.1 демонстрирует результат эксперимента.
Пример 4. Гибридизация РНК с защитным олигонуклеотидом
Расплавление образцов. Непосредственно перед экспериментом, РНК [в 0,1 мМ Na-EDTA] и олигодезоксирибонуклеотид [в 10 мМ Tris-HCl (рН 9,0), 0,01 мМ Na-EDTA] раздельно расплавляют в кипящей водяной бане в течение 2 мин, после чего быстро охлаждают на льду.
Гибридизация РНК с олигонуклеотидом. Расплавленные РНК и олигодезоксирибонуклеотид (2,5 пмоль) смешивают в 5 мкл раствора, также содержащего 80 мМ Hepes-KOH (рН 8,1), 150 мМ KCl, 0,2 мМ Na-EDTA и инкубируют в течение 15 мин при 65°С.
Пример 5. Определение полноты деградации РНК
Обработка ПНФазой. CT1n1 РНК (SEQ ID NO:1) либо ее гибрид с олигонуклеотидом R-38 [5'-GGGCTAACAGTGCGGTAACACGCACTGGA-GGTCCGAAA-3' (SEQ ID NO:8) синтезирован ЗАО "Синтол", Россия], полученный как описано в примере 4, инкубируют в присутствии или в отсутствии 2 пмоль Tth-ПНФазы в условиях реакции фосфоролиза (пример 3) при 65°С. Затем образцы экстрагируют фенолом [55], водную фазу смешивают с 6 объемами смеси этанол : ацетон (1:1 по объему) в присутствии 20 мкг линейного полиакриламида [57] и 300 мМ KCl, нуклеиновые кислоты осаждают в микроцентрифуге при 12000 g, осадок дважды промывают 80% этанолом и растворяют в 6 мкл воды. Остаточное количество CT1n1 РНК определяют с помощью ОТ-ПЦР следующим образом.
Обратная транскрипция (ОТ). Реакционная смесь объемом 10 мкл содержит 50 мМ Tris-HCl (рН 8,3), 79 мМ KCl, 3 мМ MgCl2, 10 мМ дитиотрейтол, 0,2 мМ каждого из dATP, dGTP, dTTP и dCTP, 0.5 мкМ праймера R-38 (SEQ ID NO:8), 50 нг поли(А), 50 ед обратной транскриптазы M-MLV (Amersham Biosciences) и анализируемый образец РНК. После инкубации в течение 60 мин при 45°С образцы осаждают смесью этанол: ацетон как описано выше.
ПЦР. Реакции осуществляют в ДНК-амплификаторе RapidCycler™ (Idaho Technology, США) в капиллярах объемом 10 мкл, содержащих 50 мМ Tris-HCl (рН 8,7), 2,5 мМ MgCl2, 0,2 мМ каждого из dATP, dGTP, dTTP и dCTP, 0,2 мкМ каждого из праймеров R-38 (SEQ ID NO:8) и F-39 [5'-AAGCTTAATACGACTCACTATA-GGGGTTCCAACCGGAAG-3' (SEQ ID NO:9), подчеркнут участок, совпадающий с 5'-концевой последовательностью RQ135 кДНК], 10 мкг казеина, 12 нг His6-модифицированной Taq ДНК-полимеразы [58] и часть полученного на стадии ОТ препарата кДНК, соответствующую указанному на фиг.5А стартовому числу молекул CT1n1 РНК. Препарат кДНК разводят в буфере 10 мМ Tris-HCl (рН 9,0), 0,1 мМ Na-EDTA, 0,1% полиэтиленгликоль 6000, 10нг/мкл поли(А). После пяти начальных циклов ПЦР (расплавление при 94°С в течение 10 с, отжиг при 60°С в течение 10 с, элонгация при 70°С в течение 30 с) выполняют 35 циклов, в которых температуру расплавления понижают до 88°С, а температуру отжига повышают до 65°С.
Продукты ОТ-ПЦР разделяют с помощью электрофореза в неденатурирующих условиях в буфере ТВЕ и окрашивают бромистым этидием. Результаты представлены на фиг.2А. ПНФаза присутствовала во всех образцах, кроме помеченных «-ПНФаза».
Пример 6. Селективная деградация РНК, не защищенной комплементарным олигонуклеотидом
Смесь 0,5 пмоль 3'-фрагмента и 0,5 пмоль 5'-фрагмента RQ135 РНК гибридизуют с олигонуклеотидом R-38 (SEQ ID NO:8) (пример 4), инкубируют в присутствии или в отсутствии 2 пмоль Tth-ПНФазы в условиях реакции фосфоролиза (пример 3) при 65°С и анализируют в неденатурирующих условиях, как описано в примере 5. Результат представлен на фиг.2Б.
Пример 7. Защита РНК олигонуклеотидом, гибридизованным с внутренней последовательностью
Фиг.3 демонстрирует результаты эксперимента, в котором РНК-транскрипты одной и той же плазмиды, расщепленной по сайтам рестрикции SmaI (длина транскрипта 109 нт, SEQ ID NO:3), EcoRI (127 нт, SEQ ID NO:4) или PvuII (303 нт, SEQ ID NO:5) (каждый более длинный фрагмент целиком содержит в себе последовательности более коротких фрагментов, то есть 1-109 нуклеотиды во всех фрагментах одинаковы, пример 1), либо обрабатывают («+»), либо не обрабатывают («-») с помощью 2 пмоль Tth-ПНФазы при 65°С после отжига этих транскриптов, поодиночке или в присутствии олигонуклеотида R-38 (SEQ ID NO:8), комплементарного нуклеотидам 72-109. Для осуществления эксперимента используют условия и процедуры, изложенные в примерах 3 и 4.
С целью определения точной длины укороченных фрагментов используют транскрипты, [32Р]-меченные на 5'-конце, и продукты деградации разделяют с помощью электрофореза высокого разрешения в денатурирующих условиях, применяемого для секвенирования нуклеиновых кислот [55, 59]. Для приготовления 5'-меченых транскриптов, немеченые транскрипты (пример 1) обрабатывают фосфатазой с целью удаления 5'-трифосфата, а затем фосфорилируют, используя полинуклеотидкиназу и [γ-32Р]АТР [55]. После электрофореза гель экспонируют в течение ночи на экран MS и получают изображение радиоактивных полос, сканируя экран на фосфоримиджере Cyclone™ (Packard Instrument, США). Результаты приведены на фиг.4.
Часть А демонстрирует результаты, полученные с использованием олигонуклеотида R-38 (SEQ ID NO:8), химически синтезированного ЗАО "Синтол" (Россия), без каких-либо дополнительных обработок. Дорожки 1 и 2 содержат полноразмерные транскрипты SmaI и EcoRI, а дорожки 3 и 4 - обработанные Tth-ПНФазой транскрипты EcoRI и PvuII, соответственно.
Часть Б демонстрирует результаты, полученные с использованием олигонуклеотида R-38 с точным 5'-концом, полученного из более длинного олигонуклеотида с помощью эндонуклеазы рестрикции SmaI, как описано ранее [33]. Дорожка 1 содержит полноразмерный транскрипт SmaI, дорожка 5 - транскрипт EcoRI, частично деградированный слабой щелочью с целью генерации 1-нуклеотидной «лесенки» [59]. Дорожки 1-А содержат продукты деградации транскриптов SmaI, EcoRI и PvuII, соответственно, гибридизованных с олигонуклеотидом R-38, 5'-конец которого фосфорилирован, так как эндонуклеаза рестрикции расщепляет межнуклеотидную связь между фосфатом и 3'-гидроксилом [55]. Дорожки 6-8 содержат продукты деградации тех же транскриптов, гибридизованных с тем же олигонуклеотидом, дополнительно обработанным фосфатазой [55] для удаления 5'-фосфата.
Пример 8. Синтез LNA-модифицированных олигонуклеотидов
Модифицированные LNA олигонуклеотиды синтезируют по методике поставщика амидофосфитов - компании GlenResearch (США). Готовят 0,1 М растворы соответствующих амидофосфитов: для dA, dG, Т - в безводном ацетонитриле, для dC в растворе тетрагидрофуран/ацетонитрил 1:4. Время конденсации - 3 мин. Время окисления увеличивают в три раза по сравнению с условиями, рекомендуемыми для стандартных амидофосфитов (http://www.glenresearch.com/ProductFiles/10-2011.html).
Очистку олигонуклеотидов осуществляют с помощью денатурирующего гель-электрофореза в 15% полиакриламидном геле (пример 3) при комнатной температуре и напряжении 1 кВ. Продукт обнаруживают с помощью облучения геля УФ-светом с длиной волны 254 нм. Соответствующий целевому фрагмент элюируют из геля раствором 2М перхлората лития (в течение ночи при комнатной температуре) и осаждают ацетоном в течение 15 мин при -20°С, с последующим центрифугированием в течение 10 мин при 16000 об/мин в микроцентрифуге Eppendorf (Германия). Осадок растворяют в воде MilliQ (Millipore, США) и измеряют концентрацию олигонуклеотида по оптической плотности при 260 нм [55].
Пример 9. Защита РНК LNA-модифицированными олигонуклеотидами
Фиг.5 демонстрирует результат эксперимента, в котором транскрипт PvuII (303 нт, SEQ ID NO:5) либо обрабатывают («+»), либо не обрабатывают («-») с помощью 2 пмоль Tth-ПНФазы в течение 5 мин при 65°С после отжига в присутствии немодифицированного олигонуклеотида R-38 (SEQ ID NO:8) («н/м»), комплементарного нуклеотидам 72-109, либо в присутствии LNA-модифицированного олигонуклеотида R-38, в котором LNA-аналоги нуклеотидов (помечены надстрочной буквой «L») сгруппированы недалеко от 5'-конца: 5'-GGGCLTLALALCLALGTGCGGTAACACGCACTGGAGGTCCGAAA-3' (LNA-1) или рассредоточены по длине: 5'-GGGCLTAACLAGTGCLGGTAACLACGCLACTGGAGGTCLCGAAA-3' (LNA-2).
Для осуществления эксперимента используют условия и процедуры, изложенные в примерах 3 и 4. Продукты реакции разделяют в неденатурирующих условиях (пример 5).
Пример 10. Кинетика деградации Tth-ПНФазой незащищенной высокомолекулярной РНК
Фиг.6 демонстрирует результат эксперимента, в котором смесь 50 нг рибосомных РНК Е.coli (длиной 1542 и 2904 нт) и 20 нг мРНК маммаглобина длиной 486 нт (пример 1, SEQ ID NO:6) (дорожка 1) обрабатывают с помощью 2 пмоль Tth-ПНФазы в течение указанного времени при 65°С после отжига либо в отсутствие (дорожка 2), либо в присутствии LNA-модифицированного олигонуклеотида 5'-GAAAGAGAAGGTGTGGTTTGCAGCAATCCGTAGTTGG-P-3' (SEQ ID NO:10), комплементарного нуклеотидам 432-468, в котором LNA-аналоги нуклеотидов (помечены надстрочной буквой «L») рассредоточены по длине: 5'-GLAAALGAGLAAGLGTGLTGGLTTTGLCAGCAATCCGTAGTTGG-P-3' (LNA-1) или сгруппированы у 5'-конца: 5'-GLALALALGLALGLAAGGTGTGGTTTGCAGCAATCCGTAGTTGG-p-3' (LNA-2).
Для осуществления эксперимента используют условия и процедуры, изложенные в примерах 3 и 4. Продукты реакции разделяют в неденатурирующих условиях (пример 5).
Пример 11. Влияние обработки препарата РНК Tth-ПНФазой на специфичность ОТ-ПЦР
Фиг.7 демонстрирует результат эксперимента, в котором фрагмент мРНК теломеразы человека длиной 486 нт (пример 1, SEQ ID NO:7) (дорожки 1 и 4) либо суммарную РНК, выделенную из цельной крови здорового донора, как описано в примере 2 (дорожки 2 и 3), либо обрабатывают (дорожки 3 и 4), либо не обрабатывают (дорожки 1 и 2) с помощью 2 пмоль Tth-ПНФазы в течение 5 мин при 65°С после отжига этих РНК в присутствии LNA-модифицированного олигонуклеотида 5'-GLALGLCLCLALCLGLALALCTGTCGCATGTACGGCTGGAGGTCTGTC-3' (SEQ ID NO:11), комплементарного нуклеотидам 413-450 фрагмента мРНК теломеразы. Отжиг осуществляют как описано в примере 4, а реакцию фосфоролиза - как в примере 3. Затем ПНФазу инактивируют, для чего смесь инкубируют при 96°С в течение 2 мин. После обработки образцы РНК подвергают процедуре ОТ-ПЦР, которую осуществляют следующим образом.
Обратная транскрипция (ОТ). Реакционная смесь объемом 10 мкл содержит 50 мМ Tris-HCl (рН 8,3), 75 мМ KCl, 3 мМ MgCl2, 10 мМ дитиотрейтол, 0,5 мМ каждого из dATP, dGTP, dTTP и dCTP, 0,2 мкМ праймер 5'-CATGTACGGCTGGAGGTCTGTC-3' (SEQ ID NO:12), 500 нг поли(А), 2 ед. обратной транскриптазы M-MLV SuperScriptII (Invitrogen) и анализируемый образец РНК: мРНК hTERT (20 нг или ≈1011 молекул до обработки ПНФазой, образцы 1 и 4) или суммарную РНК, выделенную из 10 мкл цельной крови (≈100 нг до обработки ПНФазой, образцы 2 и 3). После инкубации в течение 60 мин при 50°С образцы пропускают через 200 мкл колонку Sephadex G-25 (насыщенный 10 мМ TrisHCI рН 9,0; 0,1 мМ ЭДТА).
ПЦР. Реакции осуществляют в ДНК-амплификаторе RapidCycler™ (Idaho Technology, США) в капиллярах объемом 10 мкл, содержащих 50 мМ Tris-HCl (рН 8,7), 2,5 мM MgCl2, 0,2 мМ каждого из dATP, dGTP, dTTP и dCTP, 0,2 мкМ каждого из праймеров [обратного (SEQ ID NO:12) и прямого 5'-TGTACTTTGTCAAGGTGGATGTG-3' (SEQ ID NO:13)], 10 мкг бычьего сывороточного альбумина (фракция V, Behringer Mannheim, Германия), 39 нг His6-модифицированной Taq ДНК-полимеразы [58] и часть полученного на стадии ОТ препарата кДНК, соответствующую 100 молекулам мРНК hTERT (образцы 1 и 4) или 5 мкл цельной крови (образцы 2 и 3). Препарат кДНК разводят в буфере 10 мМ Tris-HCl (рН 9,0), 0,1 мМ Na-EDTA, 0,1% полиэтиленгликоль 6000, 10 нг/мкл поли(А). Осуществляют 43 цикла ПЦР в следующем режиме: расплавление при 94°С в течение 15 с, отжиг при 68°С в течение 15 с, элонгация при 72°С в течение 1 мин.
Продукты ОТ-ПЦР разделяют в неденатурирующих условиях (пример 5) и окрашивают бромистым этидием. На фиг.7 показаны негативные изображения геля.
ПРОМЫШЛЕННАЯ ПРИМЕНИМОСТЬ
Осуществление изобретения в соответствии с настоящими описанием, примерами и формулой позволяет существенно повысить специфичность обратной транскрипции в случаях, когда анализируемый образец содержит большое количество посторонней (отличной от мишени) РНК. Предлагаемый способ не исключает, а дополняет известные в технике способы понижения уровня неспецифического синтеза ДНК при определении РНК-мишеней и может быть использован как наряду с ними, так и отдельно.
Предлагаемый способ может быть использован для повышения чувствительности и специфичности молекулярной диагностики, основанной на размножении РНК-мишеней, а также в любых других приложениях молекулярной биологии и биотехнологии, для которых повышение специфичности обратной транскрипции является важным фактором.
Источники информации















Изобретение относится к области молекулярной биологии и биохимии и может быть использовано в молекулярной диагностике, связанной с выявлением РНК-мишеней. Предложен способ повышения специфичности обратной транскрипции, включающий приготовление образца, предположительно содержащего хотя бы один вид подлежащей обратной транскрипции РНК (мишени) и хотя бы один вид РНК, отличной от мишени; обработку его полинуклеотидфосфорилазой Thermus thermophilus (Tth ПНФазой) при температуре 60-75°С и микромолярной концентрации магния, проводимую после отжига исследуемого образца с защитным олигонуклеотидом, располагающимся по 3'-сторону от участка, подлежащего обратной транскрипции, и образующим с мишенью гибрид, стабильный в условиях инкубации; и реакцию обратной транскрипции, которую проводят с полученной на стадии предварительной обработки реакционной смесью без ее дополнительной очистки. 4 з.п. ф-лы, 7 ил., 11 пр.
1. Способ повышения специфичности обратной транскрипции, включающий приготовление образца, предположительно содержащего целевую РНК-мишень и хотя бы один вид РНК, отличной от мишени, предварительную обработку указанного образца ферментом для снижения в нем содержания примесей нецелевых полинуклеотидов, осуществление обратной транскрипции всей РНК-мишени или ее части и анализ продуктов обратной транскрипции, отличающийся тем, что предварительная обработка образца предусматривает отжиг с мишень-специфичным защитным олигонуклеотидом, инкубацию с полинуклеотидфосфорилазой Thermus thermophilus (Tth ПНФазой) и инактивацию Tth ПНФазы, причем выбирают защитный олигонуклеотид, способный гибридизоваться с мишенью таким образом, что его 5'-конец находится не далее 8 нуклеотидов с 5'-стороны от 3'-конца защищаемого участка мишени, и образующий с мишенью гибрид, стабильный в условиях инкубации; инкубацию осуществляют при температуре 60-75°С и микромолярной (физиологической) концентрации магния, используя оптимальную для существенной деградации незащищенной и существенной сохранности защищенной РНК комбинацию концентрации фермента, концентрации фосфата и времени реакции, а обратную транскрипцию проводят с полученной на стадии предварительной обработки реакционной смесью без проведения ее дополнительной очистки.
2. Способ по п.1, отличающийся тем, что анализ продуктов обратной транскрипции включает их размножение с использованием бесклеточной ферментативной системы.
3. Способ по п.2, отличающийся тем, что в качестве бесклеточной ферментативной системы используют полимеразную цепную реакцию.
4. Способ по п.1, отличающийся тем, что защитный олигонуклеотид модифицируют с целью повышения стабильности его гибрида с РНК-мишенью.
5. Способ по п.4, отличающийся тем, что защитный олигонуклеотид модифицируют путем замены хотя бы одного из его нуклеотидов на LNA-аналог.
| BYRNE В | |||
| ЕТ AL., Nucl | |||
| Acids Res., 16(9), 4165, 1988 | |||
| HUANG Z | |||
| ЕТ AL., Biotechniques, 20(6), 10212-1020, 1996 | |||
| FALALEEVA M | |||
| ETAL., Abstracts of International Conference, Pushchno, June 2007 | |||
| VESTER B | |||
| and WENGEL J., Biochemistry, 43(42), 13233-13241, 2004 | |||
| KAUPINNEN S | |||
| ЕТ AL., Handb | |||
| Exp.Pharmacol., 173, 405-422, 2006. |

