Изобретение относится к термостабильному ферменту, который представляет собой ДНК-полимеразу, получаемую из Carboxydothermus hydrogenoformans.
Кроме того, настоящее изобретение относится к области молекулярной биологии и предлагает усовершенствованные способы репликации и амплификации последовательностей дезоксирибонуклеиновой (ДНК) и рибонуклеиновой (РНК) кислот. В предпочтительном осуществлении настоящее изобретение предлагает способ получения комплементарной ДНК-копии с РНК матрицы с помощью термореактивной ДНК-полимеразы. В другом аспекте в настоящем изобретении предложен способ амплификации сегмента ДНК из РНК- или ДНК-матрицы с использованием термостабильной ДНК-полимеразы (RT-ПЦР или ПЦР).
Термостабильные полимеразы (ЕС 2.7.7.7. ДНК нуклеотидилтрансфераза, ДНК-управляемая) были выделены из многочисленнных термофильных организмов (например, Kaledin et al., 1980, Biokimiya Vol.45, р.644-651; Kaledin et al. , 1981, Biokimiya Vol.46, p.1247-1254; Kaledin et al., 1982, Biokimiya Vol. 47, p. 1515-1521; Ruttiman et al., 1985, Eur. J. Biochem. Vol.149, р.41-46; Neuner et al., 1990, Arch. Microbiol. Vol.153, p.205-207).
Для некоторых организмов ген полимеразы был клонирован и экспрессирован (Lawyer et al., 1989, J. Biol. Chem. 264, р.6427-6437; Engelke et al., 1990, Anal. Biochem. 191, p.396-400; Lundberg et al., 1991, Gene 108, 1-6; Perler et al., 1992, Proc. Natl. Acad. Sci., USA 89, 5577).
Термофильные ДНК-полимеразы становятся все более важными инструментами для использования в области молекулярной биологии и наблюдается возрастающий интерес к обнаружению новых полимераз, которые обладали бы более подходящими свойствами и активностями для диагностического определения РНК и ДНК, клонирования генов и секвенирования ДНК. В настоящее время термофильные ДНК-полимеразы, которые в большинстве случаев используют для этих целей, являются полимеразами рода Thermus, подобно Tag полимеразе из T.aquaticus (Brock et al., 1969, J. Bacteriol. Vol.98, p.289-297).
Обратная транскрипция обычно осуществляется вирусными обратными транскриптпазами, подобными ферментам, выделенным из вируса милобластоза птиц или вируса лейкемии мышей Moloney, которые активны в присутствии ионов магния, но отличаются тем недостатком, что обладают активностью РНКазы Н, которая разрушает матричную РНК в процессе обратной транскрипции, и имеют температурный оптимум при 42oС или 37oС соответственно.
Описываются альтернативные способы, в которых используют обратнотранскриптазную активность ДНК-полимераз термофильных организмов, которые активны при более высоких температурах. Обратная транскрипция при более высоких температурах обладает тем преимуществом, что избегает образования вторичных структур РНК матрицы, которые могут привести к появлению продуктов преждевременного окончания транскрипции. Термостабильные ДНК-полимеразы с обратной транскриптазной активностью обычно выделяют из рода Thermus. Однако ДНК-полимеразы демонстрируют обратнотранскриптазную активность только в присутствии ионов магния. Эти условия реакции являются субоптимальными, так как присутствие ионов магния снижает точность транскрибирования матричной РНК ДНК-полимеразой.
Другой отличительной особенностью обычно используемых обратных транскриптаз является то, что они обладают 3'-5' энзонуклеазной активностью. Поэтому неправильно включенные нуклеотиды не могут быть удалены, и таким образом, кДНК копии с матричной РНК могут содержать значительное число мутаций.
Поэтому желательно создать обратную транскриптазу
- которая функционирует при более высоких температурах, чтобы избежать образования вторичных структур в матрице во избежание преждевременного окончания реакции и чтобы обеспечить получение кДНК без делеций,
- которая активна в присутствии ионов магния для получения кДНК из РНК-матриц с более высокой точностью, и
- которая содержит 3'-5' экзонуклеазу для удаления неправильно включенных нуклеотидов до продолжения синтеза ДНК и для получения продукта с низкой частотой мутаций.
Настоящее изобретение направлено на удовлетворение данных потребностей и обеспечивает термостабильную ДНК-полимеразу, активную при более высоких температурах, которая обладает обратнотранскриптазной активностью в присутствии ионов магния и которая обладает активностью 3'-5'-экзонуклеазы.
Целью данного изобретения является создание полимеразного фермента (ЕС 2.7.7.7. ), отличающегося тем, что он обладает обратнотранскриптазной активностью в присутствии ионов магния так же, как и в присутствии ионов марганца. С другой точки зрения, изобретение включает ДНК-полимеразу, выделенную из Carboxydothermus hidrogenoformans (Deutsche Sammlung von Microorganismen und Zellkulturen GmbH, Mascheroder Weg 1b, D-38124 Braunschweig, DSM 8979). В другом аспекте настоящее изобретение включает ДНК-полимеразу, обладающую обратнотранскриптазной активностью в присутствии ионов магния и практически в отсутствии ионов марганца. В еще одном аспекте настоящее изобретение включает ДНК-полимеразу с молекулярной массой от около 100 до 105 кДа по данным PFGE анализа in situ. В следующем аспекте настоящее изобретение включает обратную транскриптазу, которая является термостабильной. В следующем аспекте настоящее изобретение включает ДНК-полимеразу, обладающую 3'-5'-экзонуклеазной активностью. В следующем аспекте настоящее изобретение включает рекомбинантную последовательность ДНК, которая кодирует ДНК-полимеразную активность микроорганизма Carboxydothermus hidrogenoformans. В родственном аспекте эта последовательность ДНК представлена как Последовательность ИД 7. Во втором родственном аспекте настоящее изобретение включает рекомбинантную последовательность ДНК, которая кодирует практически аминокислотные остатки 1-831. В следующем аспекте изобретение включает рекомбинанатную ДНК-плазмиду, которая включает последовательность ДНК по изобретению, встроенную в плазмидные векторы и которую можно использовать для управления экспрессией термостабильной ДНК-полимеразы Carboxydothermus hidrogenoformans в клетке-хозяине, трансформированной плазмидой. В следующем аспекте изобретение включает рекомбинантный штамм, включающий вектор pDS56, содержащий ген ДНК-полимеразы Carboxydothermus hidrogenoformans и обозначенный pAR4. Штамм (BL21(DE3)pUBS520) E. coli, содержащий плазмиду pAR4, был депонирован в Deutsche Sammlung von Microorganismen und Zellkulturen GmbH, Mascheroder Weg 1b, D-38124 Braunschweig, DSM 11179) и обозначен AR96.
Что касается пептидной цепи, как состоящей из ряда аминокислот "существенно или эффективно" в соответствии со списком, не содержащим альтернатив, авторы включили в эту ссылку все варианты пептидной цепи, содержащей замены одной или более из аминокислот, таким образом, что вся структура и все функции белка, состоящего из этой пептидной цепи, существенно те же самые (или содержат незначительные отличия), что и у незамещенного варианта. Так, например, обычно можно поменять аланин и валин без существенных изменений в свойствах белка, особенно, если изменения сайта или сайтов находятся в положениях, которые не являются критическими для морфологии складчатого белка.
ДНК-полимераза является "термостабильной" в том смысле, что она стабильна при нагревании и преимущественно активна при более высоких температурах, особенно если высокие температуры используют для денатурации цепей ДНК. Более конкретно, термостабильные ДНК-полимеразы, по существу, не инактивируются при высоких температурах, применяемых в полимеразных цепных реакциях.
Термин "обратная транкриптаза" описывает класс полимераз, характеризующийся как РНК-зависимые ДНК-полимеразы. Все известные обратные транскриптазы требуют наличия праймера для синтеза ДНК транскрипта из матрицы РНК. Исторически сложилось, что обратные транскриптазы использовали, главным образом, для транскибирования мРНК в кДНК, которую затем клонировали в вектор для дальнейших манипуляций.
Остальные определения используют в соответствии с определениями, принятыми в данной области техники.
Carboxydothermus hydrogenoformans был выделен из горячего источника на Камчатке V. Svetlichny. Образец С. hydrogenoformans был депонирован в Deutsche Sammlung von Microorganismen und Zellkulturen GmbH (DSM) в соответствии с Будапештским соглашением и ему был присвоен регистрационный номер DSM 8979. Термостабильная полимераза, выделенная из Carboxydothermus hydrogenoformans, имеет молекулярную массу от 100 до 105 кДа и сохраняет более 60% исходной активности после нагревания при 95oС в течение 30 минут. Термостабильный фермент обладает 5'-3' полимеразной активностью, 3'-5' экзонуклеазной активностью и 5'-3'-экзонуклеазной активностью и обратнотранскриптазной активностью, которая является Mg++-зависимой. Полимераза по настоящему изобретению обладает обратнотранскриптазной активностью в присутствии ионов магния и, по существу, в отсутствие ионов марганца. Термостабильный фермент может быть нативным или рекомбинантным и может быть использован для синтеза первой и второй цепочки кДНК, при клонировании кДНК, секвенировании ДНК, мечении ДНК и для амплификации ДНК.
Для выделения нативного белка Carboxydothermus hydrogenoformans можно культивировать, используя любую подходящую методику, например методику, описанную Svetlichny et al. (1991) System. Appl. Microbiol. Vol.14, p.205-208. После того как клетки выросли, осуществляют один из предпочтительных способов выделения и очистки фермента, используя многостадийный процесс следующим образом:
Клетки оттаивают, суспендируют в буфере А (40 мМ Трис-HCl, рН 7,5, 0,1 мМ ЭДТА, 7 мМ 2-меркаптоэтанола, 0,4 М NaCl, 10 мМ Pefabloc) и изолируют, дважды пропуская через гомогенизатор Gaulin. Сырой экстракт осветляют центрифугированием, надосадочную жидкость диализуют против буфера В (40 мМ Трис-HCl, рН 7,5, 0,1 мМ ЭДТА, 7 мМ 2-меркаптоэтанола, 10% глицерина) и наносят на колонку, заполненную гепарин-сефарозой (Pharmacia). В каждом случае колонки уравновешивают исходным растворителем и после нанесения образца колонки промывают трехкратным их объемом данного растворителя. Элюирование из первой колонки проводят с линейным градиентом от 0 до 0,5 М NaCl в буфере В. Фракции, демонстрирующие полимеразную активность, собирают и добавляют сульфат аммония в конечной концентрации 20%. Данный раствор наносят на гидрофобную колонку, содержащую Butyl-TSK-Toyopearl (TosoHaas). В это время колонку элюируют по уменьшающемуся градиенту сульфата аммония 20%-0%. Фракцию, содержащую активность, диализуют и снова переносят в колонку, на этот раз с ДЭАЭ-сефарозой (Pharmacia), и элюируют по линейному градиенту 0-0,5 М NaCl в буфере В. Четвертая колонка содержит Tris-Acryl-Blue(Biosepra) и ее элюируют, как и в предыдущем случае. И, наконец, активные фракции диализуют против буфера С (20 мМ Трис-HCl, рН 7,5, 0,1 мМ ЭДТА, 7,0 мМ 2-меркаптоэтанола, 100 мМ NaCl, 50% глицерина).
Выделение рекомбинантной ДНК-полимеразы из Carboxydothermus hydrogenoformans может быть проведено в соответствии с тем же протоколом или в соответствии с другими обычно используемыми способами.
ДНК-полимеразную активность измеряли, вводя меченный дигидроксигенином дУТФ в синтезированную ДНК, и определение и количественный анализ включенного дигоксигенина проводили, по существу, как описано в  ; Sagner, G.; Kessler, С. and Schmitz, G., 1992, Biotechniques Vol.12, p.104-113.
; Sagner, G.; Kessler, С. and Schmitz, G., 1992, Biotechniques Vol.12, p.104-113.
Определение обратнотранскриптазной активности осуществляют практически, как описано для определения ДНК-полимеразной активности, за исключением того, что реакционная смесь состоит из компонентов, указанных в примере 3. In situ PAGE анализ полимеразной активности и обратнотранскриптазной активности проводили, по существу, в соответствии с методикой, описанной в Spanos А. и  ((1983) Methods in Enzymology 91, 263-277). Некоторые небольшие, но существенные модификации исходного способа состоят в том, что ренатурацию ДДС-денатурированных полипептидов проводят в присутствии ионов магния (3 мМ) и дАТФ (0,5-1 мкМ) для восстановления пространственной структуры (складчатости).
((1983) Methods in Enzymology 91, 263-277). Некоторые небольшие, но существенные модификации исходного способа состоят в том, что ренатурацию ДДС-денатурированных полипептидов проводят в присутствии ионов магния (3 мМ) и дАТФ (0,5-1 мкМ) для восстановления пространственной структуры (складчатости).
3'-5' экзонуклеазную активность обычно называют "корректорной" или "редактирующей" активностью ДНК полимераз. Она локализована в маленьком домене крупного фрагмента полимераз типа А. Эта активность удаляет неправильно спаренные нуклеотиды с 3'-конца затравочного конца ДНК в отсутствии нуклеозидтрифосфатов (Kornberg А. и Baker Т.А. (1992) DNA Replication W.H. Freemann & Company, New York). Данное нуклеазное действие подавляется дезоксинуклеозидтрифосфатазами, если они совместимы с матрицей и могут быть включены в полимер.
3'-5'-экзонуклеазную активность заявляемой ДНК-полимеразы можно измерить как деградацию или укорачивание меченного дигоксигенином 5'-олигонуклеотида, отожженного с матричной ДНК в отсутствие или в присутствии дезоксирибонуклеозидтрифосфатов или на фрагментах ДНК в отсутствие или в присутствии дезоксирибонуклеозидтрифосфатов.
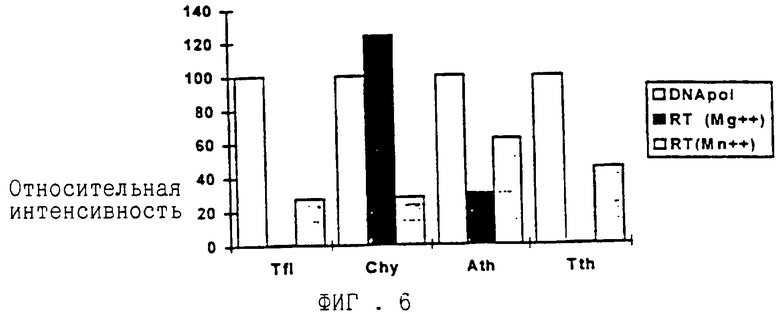
ДНК-полимераза Carboxydothermus hydrogenoformans является первой ДНК-полимеразой, выделенной из термофильных эубактерий, с более высокой активностью в присутствии ионов магния, нежели в присутствии ионов марганца, как представлено на Фиг.1. По сравнению с ДНК-полимеразной активностью обратнотранскриптазная активность в присутствии магния относительно высока. Это показано в сравнении с ДНК-полимеразами из T.filiformis A.thermophilum и наиболее часто используемой для обратной транскрипции ДНК-полимеразной Т. thermophilus - на Фиг. 6. Обратнотранскриптазная активность, зависящая от ионов магния, является преимуществом, так как ДНК-полимеразы синтезируют ДНК с более высокой точностью в присутствии ионов магния, нежели в присутствии ионов марганца (Beckman R. A. et al. (1985) Biochemistry 24, 5810-5817; Ricchetti M. и Buc Н. (1993) EMBO J. 12, 387-396). Низкая точность синтеза ДНК, по-видимому, приводит к получению мутантных копий исходной матрицы. Кроме того, ионы Mn2+ приводят к увеличению скорости деградации РНК, особенно при повышенных температурах, и это может вызвать синтез укороченных продуктов обратной транскрипции.
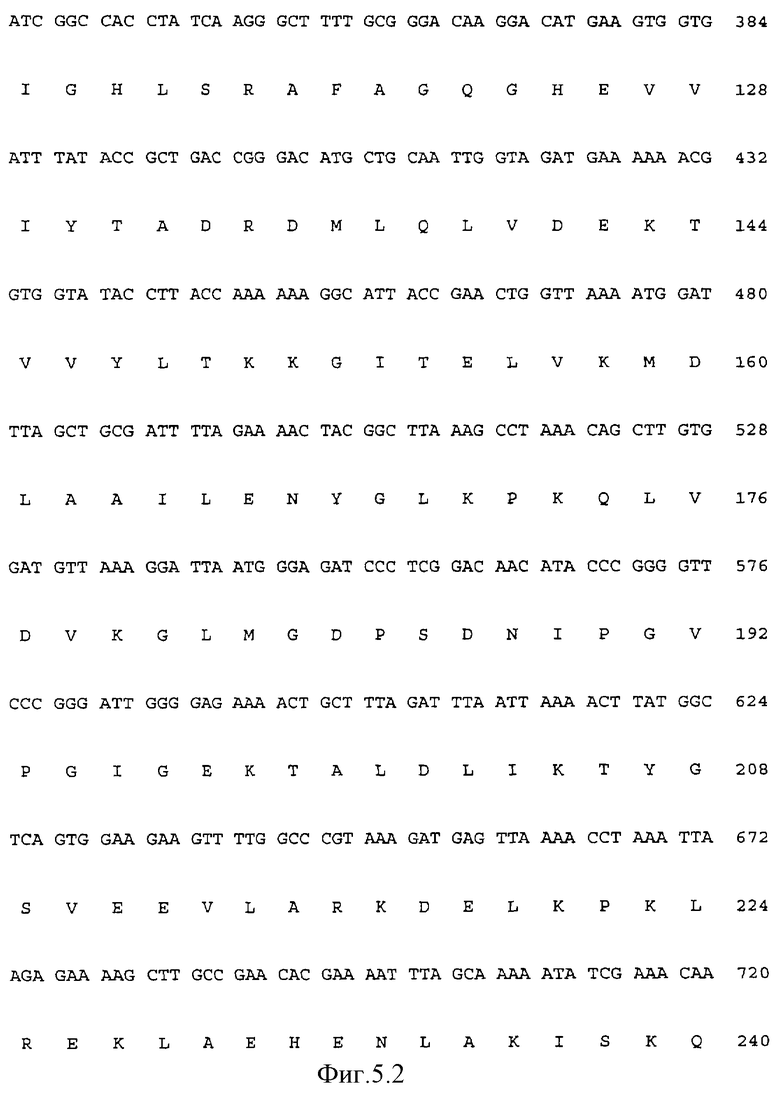
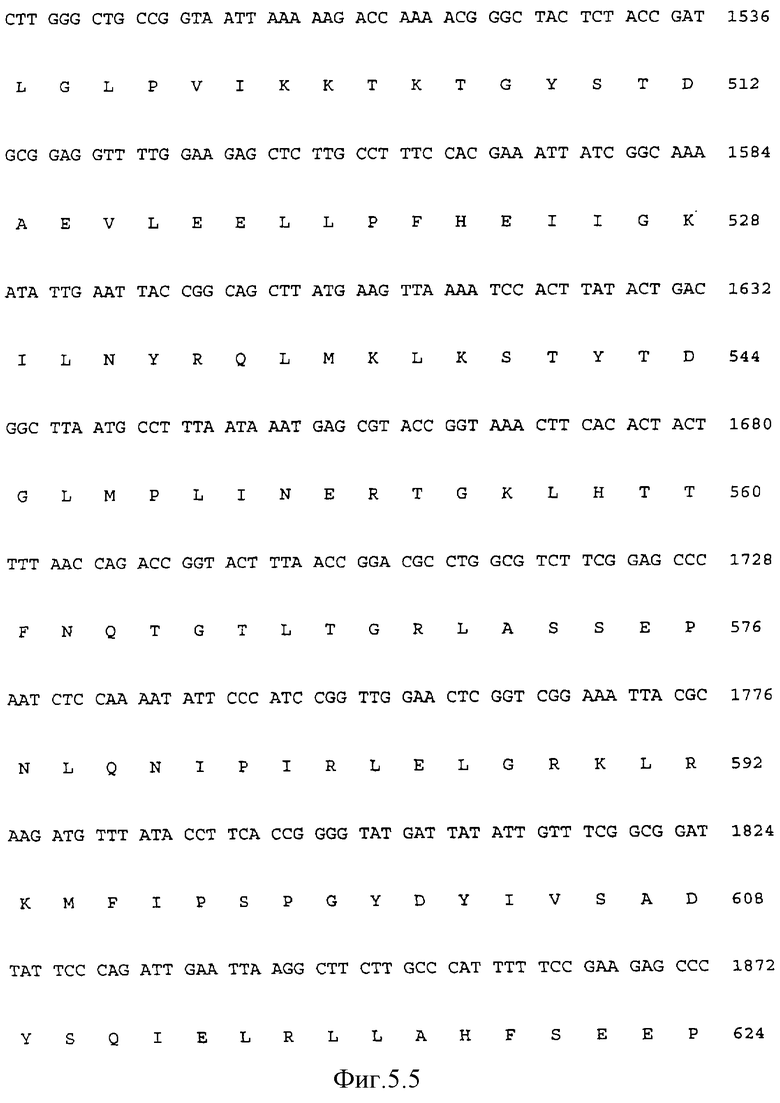
Последовательность ДНК (Последовательность ИД 7) полимеразы Carboxydothermus hydrogenoformans и производная аминокислотная последовательность фермента представлены на Фиг.5. Молекулярная масса, вычисленная для этой последовательности, составляет 94348 Да по данным электрофореза в ДДС-полиакриламидном геле, однако полимераза Carboxydothermus hydrogenoformans обладает электрофоретической подвижностью, более высокой, чем E.coli pol I (109 кДа), и более низкой подвижностью, чем Taq-полимераза (94 кДа) и фрагмент Кленова (76 кДа), как представлено на Фиг.2. Сравнивая миграционные свойства ДНК-полимераз Taq и E.coli со свойствами полимеразы Carboxydothermus hydrogenoformans, может быть вычислена молекулярная масса от 100 до 105 кДа. Так как полимераза Carboxydothermus hydrogenoformans, выделенная из нативного штамма, обладает теми же миграционными свойствами, что и рекомбинантный фермент, более "медленная" миграция в процессе гель-электрофореза с ДДС скорее является свойством фермента, чем артефактом клонирования. Возможным объяснением данного явления может быть то, что этот фермент, полученный из термофильного организма, обладает чрезвычайно стабильной структурой, которая сохраняет некоторую степень складчатости в стандартно используемых условиях денатурации.
Получение рекомбинантной формы ДНК-полимеразы Carboxydothermus hydrogenoformans обычно включает следующие стадии: хромосомную ДНК из Carboxydothermus hydrogenoformans выделяют, обрабатывая клетки детергентом, например ДДС, и протеиназой, например протеиназой К. Раствор экстрагируют добавлением фенола и хлороформа, и ДНК очищают, осаждая этанолом, ДНК растворяют в Трис/ЭДТА буфере, и ген, кодирующий ДНК полимеразу, специфично амплифицируют по методу ПЦР, используя два смешанных олигонуклеотида (праймер 1 и 2). Данные нуклеотиды, описанные последовательностью ИД 1 и последовательностью ИД 2, были сконструированы на базе консервативных участков ДНК-полимераз семейства А, что опубликовано Braithwaite D.K. и Ito J., 1993, Nucl. Acids Res. Vol. 21, p. 787-802. Специфично амплифицированный фрагмент лигируют в вектор, предпочтительно вектор pCRTMII (Invitrogen), и последовательность определяют по методу циклического секвенирования. Полное выделение кодирующего участка и фланкирующих последовательностей гена ДНК-полимеразы можно осуществить с помощью рестрикционной фрагментации ДНК Carboxydothermus hydrogenoformans другим ферментом рестрикции, как в первом цикле скрининга обратной ПЦР (Innis et al., (1990) PCR Protocols; Academic Press, Inc., p. 219-227). Это можно осуществить с помощью синтезированных олигонуклеотидных праймеров, связывающихся с внешними последовательностями ДНК части гена, но противоположно ориентированных. Данные олигонуклеотиды, описываемые последовательностями ИД 3 и ИД 4, были сконструированы на основе последовательностей, которые были определены в первой описанной выше ПЦР. В качестве матрицы используют ДНК Carboxydothermus hydrogenoformans, которую расщепляют рестрикционным расщеплением, и замыкают в цикл путем взаимодействия с Т4 ДНК-лигазой. Для выделения кодирующего участка целого полимеразного гена проводят еще одну ПЦР, используя праймеры, показанные в Последовательностях ИД 5 и 6, для амплификации полного гена ДНК полимеразы непосредственно из геномной ДНК и встраивания концов, совместимых с линеаризованным экспрессирующим вектором.
Последовательность ИД 1:
Праймер 1: 5'-CCN AAY YTN CAR AAY ATH-3'
Последовательность ИД 2:
Праймер 2: 5'-YTC RTC RTG NAC YTG-3'
Последовательность ИД 3:
Праймер 3: 5'-GGG GGA AGA CGC TAT ATT CCT GAG C-3'
Последовательность ИД 4:
Праймер 4: 5'-GAA GCC TTA ATT CAA TCT GGG AAT AAT C-3'
Последовательность ИД 5:
Праймер 5: 5'-CGA ATT CAA TCC ATG GGA AAA GTA GTC CTG GTG GAT-3'
Последовательность ИД 6:
Праймер 6: 5'-CGA ATT CAA GGA TCC TTA CTT CGC TTC ATA CCA GTT-3'
Ген оперативно связывают с соответствующими контрольными последовательностями для экспрессии либо в прокариотической, либо в эукариотической системе хозяин/вектор. Вектор, предпочтительно, кодирует все функции, необходимые для трансформации и сохранения в подходящем хозяине, и может кодировать избираемые маркеры и/или контрольные последовательности для экспрессии полимеразы. Активную рекомбинантную термостабильную полимеразу можно получить, трансформируя культуры хозяев либо непрерывно, либо после индуцирования экспрессии. Активную термостабильную полимеразу можно выделить либо из клеток хозяина, либо из культуральной среды, если белок секретируется через клеточную мембрану.
Также предпочтительно, чтобы экспрессия термостабильной полимеразы Carboxydothermus hydrogenoformans строго контролировалась бы в Е. coli во время клонирования и экспрессии. Векторы, применимые в практике настоящего изобретения, должны обеспечивать различные степени контролируемой экспрессии Carboxydothermus hydrogenoformans полимеразы, обеспечивая некоторые или все из следующих контрольных особенностей: (1) промоторы или сайты инициации транскрипции, либо непосредственно прилегающие к началу гена полимеразы, либо как гибридные белки, (2) операторы, которые можно использовать для "включения" или "выключения" экспрессии гена, (3) сайты связывания рибосом для улучшения трансляции и (4) сайты окончания транскрипции или трансляции для повышения стабильности. Подходящие векторы, применяемые для клонирования и экспрессии полимеразы Carboxydothermus hydrogenoformans включают, например, фаги и плазмиды. Примеры фагов включают лямбда gtl1 (Promega), лямбда Dash (Stratagene), лямбда ZapII (Stratagene). Примеры плазмид включают pBR322, pBTac2 (Boehringer Mannheim), pBluescript (Stratagene), pSP73 (Promega), рЕТ3А (Rosenberg, supra (1987) Gene 56:125-135), pASK75 (Biometra), pDS56 ( , Matile, H. and Garotta G. (1990) Immunological Methods. Letkovcs, I. and Pernis, В., eds.) и pET11C (Studier, F.W. (1990) Methods in Enzymology, 185:60-89). Использование плазмиды в соответствии с настоящим изобретением, как было показано, является выгодным, особенно это касается pDS56. Плазмиду pDS56, содержащую ген ДНК-полимеразы Carboxydothermus hydrogenoformans, далее обозначают pAR4.
, Matile, H. and Garotta G. (1990) Immunological Methods. Letkovcs, I. and Pernis, В., eds.) и pET11C (Studier, F.W. (1990) Methods in Enzymology, 185:60-89). Использование плазмиды в соответствии с настоящим изобретением, как было показано, является выгодным, особенно это касается pDS56. Плазмиду pDS56, содержащую ген ДНК-полимеразы Carboxydothermus hydrogenoformans, далее обозначают pAR4.
Существуют стандартные методики трансформации, инфицирования фагом и культивирования клеток (Maniatis et al. (1982) Molecular Cloning: A Laboratory Manual, Cold Spring Harbour Laboratory Press). Из многочисленных штаммов Е. coli, которые можно использовать для трансформации плазмидами, предпочтительные штаммы включают JM110 (ATCC 47013), LE392 pUBS520 (Maniatis et al. Supra; Brinkmann et al, (1989) Gene 85:109-114), JM101 (ATCC 33876), XL1 (Stratagene) и RR1 (ATCC 31343), BL21 (DE3) pUBS520 (Brinkman, U. et al. (1989) Gene 85,109-114) и BL21 (DE3) plysS (Studier, F.W. et al. (1990) Methods in Enzymology, см. ранее). Как было показано, выгодным в соответствии с настоящим изобретением является использование Е. coli штамма BL21 (DE3) pUBS520. Штамм BL21 (DE3) pUBS520 Е. coli, трансформированный плазмидой pAR4, обозначают затем как AR96 (DSM 11179). Штаммы XLl-Blue (Stratagene) и ER1458 (Raleigh, E.A. et al. (1988) Nucleic Acids Research 16:1563-1575) E. coli относятся к штаммам, которые можно использовать для фага лямбда, а Y1089 можно использовать для лямбда gtl1-лизогении. Трансформированные клетки предпочтительно культивируют при 37oС, а экспрессию клонированного гена индуцируют ИПТГ.
Выделение рекомбинантной ДНК-полимеразы можно осуществить стандартными методами. Выделение и очистку ДНК-полимеразы из экстракта Е. coli можно осуществить стандартными методами. Данные методы включают, например, методы, использующие растворимость, такие как осаждение солью и осаждение растворителем, методы, использующие разницу по молекулярной массе, такие как диализ, ультрафильтрация, гель-фильтрация и электрофорез в ДДС-полиакриламидном геле, методы, использующие различие в электрических зарядах, такие как ионообменная колоночная хроматография, методы, использующие специфичные взаимодействия, такие как афинная хроматография, методы, использующие различия в гидрофобности, такие как высокоэффективная жидкостная хроматография с обращенной фазой, и методы, использующие различия в изоэлектрических точках, такие как электрофорез с изоэлектрическим фокусированием.
Настоящее изобретение обеспечивает усовершенствованные способы эффективной транскрипции РНК и амплификации РНК или ДНК. Данные усовершенствования достигнуты в результате открытия и применения неизвестных ранее свойств термореактивных ДНК-полимераз.
Термостабильный фермент по данному изобретению можно использовать для любых целей, когда необходима или желательна такая ферментативная активность. В наиболее предпочтительном воплощении фермент катализирует реакцию амплификации нуклеиновой кислоты, известную как ПЦР. Данный процесс амплификации последовательностей нуклеиновых кислот описан и заявлен в Патенте США 4683202. Метод амплификации нуклеиновой кислоты посредством ПЦР включает амплификацию, по крайней мере, одной специфической последовательности нуклеиновой кислоты, содержащейся в нуклеиновой кислоте или в смеси нуклеиновых кислот, и получение двухцепочечной ДНК. Любая последовательность нуклеиновой кислоты в очищенной или в неочищенной форме может быть использована в качестве исходной нуклеиновой кислоты (нуклеиновых кислот) при условии, что она содержит или предположительно содержит желаемую специфическую последовательность нуклеиновой кислоты. Нуклеиновую кислоту, подлежащую амплификации, можно получить из любого источника, например из такой плазмиды, как pBR322, из клонированной ДНК или РНК, из природной ДНК или РНК из любого источника, включая бактерии, дрожжи, вирусы, органеллы и такие высшие организмы, как растения и животные, или из препаратов нуклеиновых кислот, полученных in vitro.
ДНК или РНК может быть выделена из крови, такого тканевого материала, как хорионические ворсинки или амниотические клетки, различными методами. Смотри, например, Maniatis Т. et al. (1982) Molecular Cloning: A Laboratory Manual (Cold. Spring Harbour Laboratory, Cold Spring Harbour, New York), pp. 280-281. Таким образом, в этом способе можно использовать, например, ДНК или РНК, включая матричную РНК, причем ДНК или РНК могут быть как одноцепочечными, так и двухцепочечными. Кроме того, можно использовать гибрид ДНК-РНК, который содержит по одной цепочке каждой из них.
Амплификацию последовательностей мишеней в ДНК или с РНК можно осуществить для доказательства присутствия конкретной последовательности в образце подлежащей анализу нуклеиновой кислоты или для клонирования специфического гена. Для этих целей очень удобно использовать ДНК-полимеразу из Carboxydothermus hydrogenoformans. Благодаря ее 3'-5' экзонуклеазной активности можно синтезировать продукты с повышенной точностью, как в случае с обратными транскриптазами на известном уровне техники.
ДНК-полимераза из Carboxydothermus hydrogenoformans может быть также использована для упрощения и усовершенствования способов определения молекул-мишеней РНК в образце. В данных способах ДНК-полимераза из Carboxydothermus hydrogenoformans может катализировать: (а) обратную транскрипцию, (b) синтез второй цепочки кДНК и, при желании, (с) амплификацию с помощью ПЦР. Использование ДНК-полимеразы из Carboxydothermus hydrogenoformans в описанных способах устраняет необходимые ранее требования проведения двух наборов условий инкубации, что было необходимо из-за использования различных ферментов на каждой стадии. Использовать ДНК-полимеразу из Carboxydothermus hydrogenoformans можно для осуществления обратной транскрипции РНК и амплификации получаемой комплементарной ДНК, что повышает специфичность и предусматривает меньше стадий, чем при известных ранее методиках клонирования РНК и в диагностических методиках.
Краткое описание рисунков
На ФИГ.1 показана относительная обратнотранскриптазная активность ДНК-полимеразы из Carboxydothermus hydrogenoformans в зависимости от солей магния и марганца.
На ФИГ.2 представлена фотография анализа ДНК-полимеразы, осуществленного in situ. Активность ДНК-полимеразы из Carboxydothermus hydrogenoformans и полимераз сравнения определяют in situ. Фракцию ДНК-полимеразы из С. hydrogenoformans и ферменты сравнения подвергали электрофорезу на ДДС-полиакриламидном геле, содержащем активированную ДНК тимуса теленка. После электрофореза ДДС удаляли, белки ренатурировали и инкубировали при 65oС в присутствии соли магния, дНТФ и меченого дигоксигенином дУТФ для обеспечения синтеза ДНК. Получали блот нуклеиновой кислоты на нейлоновой мембране и вновь синтезированную ДНК регистрировали с помощью хемилюминесцентной реакции.
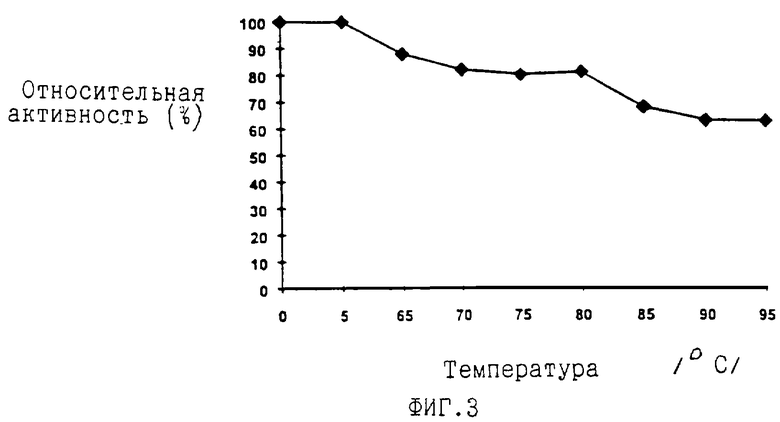
На ФИГ. 3 показана термостабильность ДНК полимеразы из Carboxydothermus hydrogenoformans. Аликвоты ДНК полимеразы инкубировали в течение 30 мин при температурах, указанных на фигуре, и затем определяли остаточную ферментативную активность.
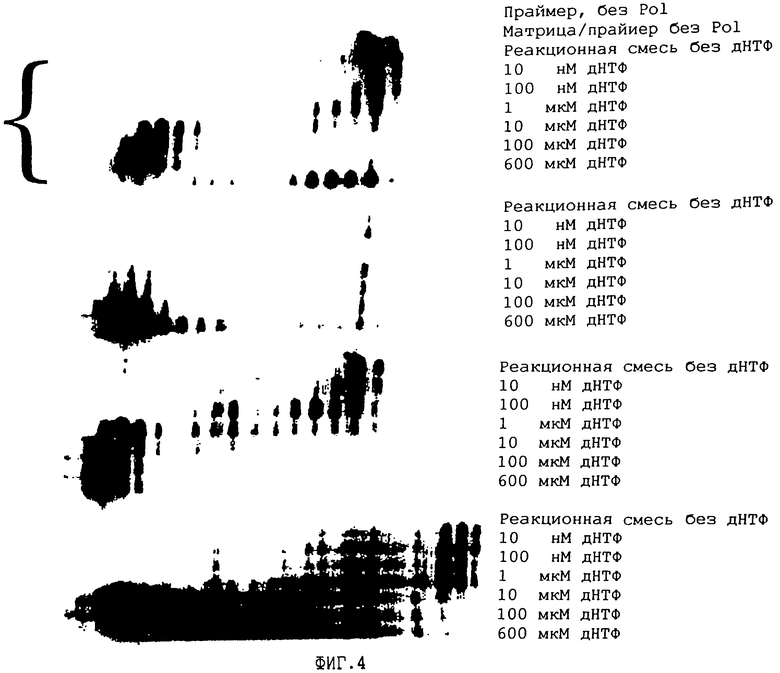
На ФИГ.4 представлен анализ 3'-5'-экзонуклеазной активности ДНК-полимеразы из Carboxydothermus hydrogenoformans в сравнении с ДНК-полимеразой из Thermus aquaticus и Pyrococcus woeseii. Анализ проводят в присутствии или в отсутствие дНТФ. 22-мepный праймер, меченый дигоксигенином по 5'-концу, отжигают с 34-мepнoй матричной ДНК, оставляя 5'-выступающий конец матричной ДНК длиной 12 п. н. ДНК-полимеразы из Carboxydothermus hydrogenoformans, Thermus aquaticus и Pyrococcus woeseii инкубировали с данным субстратом в присутствии магния с или без дНТФ. Продукты разделяли на секвенирующем геле, получали блот на нейлоновой мембране и регистрировали с помощью хемилюминесцентной реакции.
На ФИГ. 5 представлена последовательность ДНК гена полимеразы Carboxydothermus hydrogenoformans с Последовательностью ИД 7 и производная пептидная последовательность белка ДНК-полимеразы с Последовательностью ИД 8.
На ФИГ. 6 представлено сравнение обратнотранскриптазной активности термостабильных ДНК-полимераз Carboxydothermus hydrogenoformans, Anaerocellun thermophilum, Thermus filiformis (Pacific Enzymes) и Thermus thermophilus. Анализировали аналогичные количества в единицах ДНК-полимераз. Каждый фермент тестировали на предмет ДНК-полимеразной активности, обратнотранскриптазной активности в присутствии Mg++(5 мМ) и обратнотранскриптазной активности в присутствии Mn++ (1 мМ) в условиях реакции, оптимальных для конкретных ферментов. Синтез ДНК измеряли по включению меченых дигоксиленином нуклеотидов. Для сравнения отношения ДНК-полимеразной активности к обратнотранскриптазной активности оптическую плотность в относительных единицах (RLU, о. е. ), измеренную в анализе ДНК-полимеразы, принимают за 100. Оптическую плотность в о.е., измеренную в тестах определения обратнотранскриптазной активности, выражают как процент от полимеразной активности.
ПРИМЕР 1
Определение эндонуклеазной, экзонуклеазной и рибонуклеазной активностей
Отсутствие эндонуклеазной активности: 1 мкг плазмидной ДНК инкубируют в течение 4 часов с избытком очищенной ДНК-полимеразы в 50 мкл тестового буфера с нанесенным слоем парафинового масла при 72oС.
Отсутствие неспецифической экзонуклеазной активности: 1 мкг EcoRI/HindIII-фрагментов ДНК лямбда инкубируют в 100 мкл тестового буфера в отсутствие или в присутствии дНТФ (1 мМ конечной концентрации каждого) с избытком очищенной ДНК-полимеразы в течение 4 часов при 72oС.
Отсутствие рибонуклеазной активности: 3 мкг РНК MS2 инкубируют с избытком ДНК-полимеразы в 20 мкг тестового буфера в течение 4 часов при 72oС. РНК затем анализируют с помощью электрофореза в MOPS геле (Maniatis et al. (1982) Molecular Cloning: A Laboratory Manual, Cold Spring Harbour, New York).
ПРИМЕР 2
Определение ДНК-полимеразной активности
ДНК-полимеразную активность определяли по включению меченого дигоксигенином дУТФ в синтезированную ДНК с помощью регистрации и количественного определения дигоксигенина, по существу, в соответствии с методикой  ; Sagner, G.; Kessler, С. and Schmitz, G. (1992) Biotechniques 12, 104-113. Реакцию проводят в реакционном объеме 50 мкл, содержащем 1 или 2 мкл разбавленной (0,05 ЕД - 0,01 ЕД) ДНК-полимеразы и 50 мМ Трис-HCl, рН 8,5; 12,5 мМ (NH4)2SO4; 10 мМ KCl; 5 мМ MgCl2; 10 мМ 2-меркаптоэтанола; 33 мкМ дНТФ; 200 мкг/мл BSA; 12 мкг ДНК из тимуса теленка, активированной ДНКазой I, и 0,036 мкМ дигоксигенин-дУТФ.
; Sagner, G.; Kessler, С. and Schmitz, G. (1992) Biotechniques 12, 104-113. Реакцию проводят в реакционном объеме 50 мкл, содержащем 1 или 2 мкл разбавленной (0,05 ЕД - 0,01 ЕД) ДНК-полимеразы и 50 мМ Трис-HCl, рН 8,5; 12,5 мМ (NH4)2SO4; 10 мМ KCl; 5 мМ MgCl2; 10 мМ 2-меркаптоэтанола; 33 мкМ дНТФ; 200 мкг/мл BSA; 12 мкг ДНК из тимуса теленка, активированной ДНКазой I, и 0,036 мкМ дигоксигенин-дУТФ.
Образцы инкубируют в течение 30 мин при 72oС, реакцию останавливают, добавляя 2 мкл 0,5 ЭДТА, и пробирки помещают на лед. После добавления 8 мкл 5 М NaCl и 150 мкл этанола (предварительно охлажденного до -20oС) ДНК осаждают, инкубируя в течение 15 мин на льду, и осаждают центрифугированием в течение 10 мин при 13000 об/мин и 4oС. Осадок промывают 100 мкл 70% этанола (предварительно охлажденного до -20oС) и 0,2 М NaCl, снова центрифугируют и сушат под вакуумом.
Осадок растворяют в 50 мкл Трис-ЭДТА (10 мМ, 0,1 мМ; рН 7,5). 5 мкл образца вводят в лунку белого планшета для микротитрования с дном из нейлоновой мембраны (Pall Filtrationstechnic GmbH, Dreieich, FRG, продукт SM045BWP). ДНК фиксируют на мембране, прогревая в течение 10 мин при 70oС. Лунки, содержащие ДНК, заполняют 100 мкл профильтрованного через 0,45 мкм фильтр 1% блокирующего раствора (100 мМ малеиновой кислоты, 150 мМ NaCl, 1% (вес/об) казеина, рН 7,5). Все последующие стадии инкубации осуществляют при комнатной температуре. После инкубации в течение 2 мин раствор отсасывают через мембрану с подходящим вакуумным коллектором при давлении - 0,4 бар. После повторения стадии промывки лунки заполняют 100 мкл антидигоксигенин-АР. Fab-фрагментами в разбавлении 1: 10000 (Boehringer Mannheim, FRG, 1093274), разбавленными вышеуказанным блокирующим раствором. После инкубации в течение 2 мин и отсасывания эту стадию повторяют еще раз. Лунки дважды промывают под вакуумом 200 мкл промывочного буфера (100 мМ малеиновой кислоты, 150 мМ NaCl, 0,3% (об/об) TweenTM20 (Поли(оксиэтилен)n-сорбитанмонолаурат), рН 7,5). После дополнительных двух промывок под вакуумом, каждый раз по 200 мкл промывочного буфера 2 (10 мМ Трис-HCl, 100 мМ NaCl, 50 мМ MgCl2, pH 9,5), лунки инкубируют в течение 5 мин с 50 мкл CSPDTM (Boehringer Mannheim, 1655884) в разбавлении 1:100 в промывочном буфере 2, который служит хемилюминесцентным субстратом для щелочной фосфатазы. Раствор отсасывают через мембрану и после 10 мин инкубации при комнатной температуре количество о.е./с (относительных единиц в секунду) регистрируют с помощью люминометра, например MicroLumat LB 96P (EG&G Berthold, Wildbad, FRG).
С помощью серийных разбавлений Taq ДНК-полимеразы строят калибровочную кривую, в которой линейный участок служит стандартом для определения активности подлежащей анализу ДНК-полимеразы.
ПРИМЕР 3
Определение обратнотранскриптазной активности
Анализ осуществляют, по существу, как описано для определения ДНК-полимеразной активности, за исключением того, что реакционная смесь состоит из следующих компонентов: 1 мкг полидА-(дТ)15, 33 мкМ дТТФ, 0,36 мкм дигоксигенин-дУТФ, 200 мг/мл BSA, 10 мМ Трис-HCl, рН 8,5, 20 мМ KCl, 5 мМ MgCl2, 10 мМ ДТЭ и различных количеств ДНК-полимеразы. Инкубирование проводят при 50oС.
ПРИМЕР 4
Определение ДНК-полимеразной активности in situ
Анализ полимеразной и обратнотранскриптазной активностей с помощью PAGE in situ осуществляют, по существу, в соответствии с методикой, описанной Spanos А. и  , 1983, Methods in Enzymology, Vol.91, p.263-277. Некоторые небольшие, но существенные модификации исходной методики состоят в том, что ренатурацию ДДС-денатурированных полипептидов осуществляют в присутствии ионов магния (3 мМ) и дАТФ (0,5-1 мкМ) для способствования восстановлению пространственной структуры.
, 1983, Methods in Enzymology, Vol.91, p.263-277. Некоторые небольшие, но существенные модификации исходной методики состоят в том, что ренатурацию ДДС-денатурированных полипептидов осуществляют в присутствии ионов магния (3 мМ) и дАТФ (0,5-1 мкМ) для способствования восстановлению пространственной структуры.
Вкратце, методика состоит в следующем:
После выделения полипептидов либо из сырых клеточных экстрактов, либо из очищенных образцов на денатурирующих 8% полиакриламидных гелях (концентрирующий гель 5% акриламида, который содержит 150 мкг активированной ДНК тимуса теленка на мл объема геля) гели промывают четыре раза по 30 мин каждый при комнатной температуре при умеренном встряхивании в избытке ренатурирующего буфера (Трис-HCl 50 мМ, рН 8,3; ЭДТА 1 мМ; 2-меркаптоэтанол 3 мМ; KCl 50 мМ; глицерин 5-10%) для удаления ДДС. Затем гель инкубируют в течение ночи в том же самом буфере, включающем 3 мМ MgCl2 и 0,5-1 мкМ дАТФ при 4oС без перемешивания. Первые четыре промывки ренатурирущим буфером повторяют на следующий день. После удаления ДДС и ренатурации белков гель переносят в реакционную смесь, состоящую из Трис-HCl 50 мМ, рН 8,3; KCl 50 мМ; ДТТ 3 мМ; MgCl2 7 мМ; 12 мкМ дАТФ, дЦТФ, дГТФ (каждого), 8 мкМ дТТФ и 4 мкМ Диг-дУТФ, 10% (об/об) глицерина. Гели вначале инкубируют при встряхивании при комнатной температуре в течение 1 часа, а затем медленно нагревают поэтапно до 37oС, 45oС, 55oС, 65oС и 72oС. В каждом интервале температур синтез ДНК проводят в течение 60 мин.
После синтеза ДНК ее переносят либо с помощью контактного блоттинга, либо с помощью капиллярного блоттинга (15•SSC, Maniatis et al., см. ранее) на нейлоновые мембраны (Boehringer Mannheim, GmbH) и сшивают.
Вновь синтезированную Dig-меченую ДНК определяют способом, описанным в предыдущем разделе (определение ДНК-полимеразной активности).
Для определения молекулярной массы в тот же гель, но в различные полосы вводят маркерные полимеразы известной молекулярной массы (например, полимеразу Кленова, Pol I, Taq-полимеразу, Tth-полимеразу, HIV RT, M-MuLV RT).
Молекулярная масса заявленной ДНК-полимераэы, определенная данным способом, составляет 100-105 кДа.
ПРИМЕР 5
Определение 3'-5' экзонуклеазной активности
3'-5' экзонуклеазную активность обычно называют "корректорной" или "редактирующей" активностью ДНК-полимераз. Она локализована в маленьком домене крупного фрагмента полимераз типа А. Данной активностью удаляются нуклеотиды с 3'-конца затравочного конца ДНК в отсутствие нуклеозидтрифосфатов (Kornberg А. и Baker Т.А. (1992) DNA Replication W.H. Freemann & Company, New York). Данный эффект нуклеазы подавляется дезоксинуклеозидтрифосфатами, если они совместимы с матрицей и могут быть включены в полимер.
3'-5' экзонуклеазную активность заявленной ДНК-полимеразы можно измерить по деградации или укорачиванию 5'-меченого дигоксигенином олигонуклеотида, отожженного с матричной ДНК, в отсутствие или в присутствии дезоксирибонуклеозидтрифосфатов или на фрагментах ДНК в отсутствие или в присутствии дезоксирибонуклеозидтрифосфатов.
Деградация меченого дигоксигенином олигонуклеотида: Реакционная смесь практически та же, что и для определения ДНК-полимеразной активности (50 мМ Трис-HCl, рН 8,4; 12,5 мМ (NH4)2SO4; 10 мМ KCl; 5 мМ MgCl2; 10 мМ 2-меркаптоэтанола), за исключением того, что концентрацию дНТФ снижали до 12,5 мкМ, а активированную ДНК тимуса теленка заменяли 500 фМ праймера или смеси матрица/праймер.
Последовательность праймера:
Последовательность ИД 8:
Dig-GCATGGATCCCACTGCCCAGGG (5'-3'). Этот праймер отжигают с молекулами матрицы с различными 5'-выступающими концами праймера длиной 12 п.н. Образцы ДНК-полимеразы, обычно по 0,1 Ед, инкубируют в общем объеме 10 мкл в течение 30 мин при 72oС в термореакторе Perkin Elmer. Реакцию останавливают, добавляя равный объем формамидного буфера (98% формамида; 10 мМ ЭДТА; бромфеноловый синий и ксилолцианол), и денатурируют, нагревая в течение 10 мин при 95oС. Образцы быстро охлаждают на льду и вводят в секвенирующий гель (20% денатурирующий полиакриламид/мочевина). Электрофорез осуществляют при 60oС и 2000 В в течение 2,5 часов.
После выделения ДНК переносят на положительно заряженную нейлоновую мембрану (Boehringer Mannheim) с помощью контактного блоттинга в течение 30 мин. ДНК сшивают с мембраной, облучая УФ с энергией 120 мДж (Stratalinker, Stratagene). Мембрану блокируют блокирующим раствором (100 мМ малеиновой кислоты, 150 мМ NaCl, 1% (вес/об) казеина, рН доводят до 7,5, добавляя 1 М NaOH) при комнатной температуре в течение, по крайней мере, 30 мин. Меченую дигоксигенином праймерную ДНК определяют с помощью антидигоксигенин-AP. Fab-фрагментов (Boehringer Mannheim, FRG, 1093274) в разбавлении 1:1000 в блокирующем растворе (30 мин при комнатной температуре). Избыток несвязанных антител удаляют, промывая 3-4 раза (по 10-15 мин каждая стадия) промывочным буфером (100 мМ малеиновой кислоты, 150 мМ NaCl, 0,3% (об/об) TweenTM20 (поли(оксиэтилен)n-сорбитанмонолаурат), рН 7.5). Мембрану переносят в буфер, содержащий 10 мМ Трис-HCl, 100 мМ NaCl, рН 9,5, и промывают еще дважды по 10-15 мин при комнатной температуре. И наконец, мембраны замачивают в 1:1000 разбавленном растворе CDP-StarTM (Boehringer Mannheim). CDP-StarTM служит хемилюминесцентным субстратом для щелочной фосфатазы. Затем мембрану переносят на фильтровальную бумагу (Whatman 3MM) для удаления избытка жидкости, располагают между двумя прозрачными листами и экспонируют на рентгеновские пленки (Chemiluminescent Detection Film, Boehringer Mannheim) на 5-10 мин. 3'-5'-экзонуклеазную активность определяют по деградации или укорачиванию праймера по сравнению с контролем (без добавления полимеразы). В качестве негативного и позитивного контролей включают ДНК-полимеразы из Thermus aquaticus (не обладающая 3'-5'-экзонуклеазной активностью) и из Pyrococcus woeseii (обладающая 3'-5'-экзонуклеазной активностью).
Деградация фрагментов ДНК в присутствии или в отсутствие дезоксинуклеозидтрифосфатов: Серию разбавлений Chy-полимеразы инкубировали в течение 2 часов при 70oС с 1 мкг маркера III молекулярной массы ДНК (Boehringer Mannheim) в присутствии и в отсутствие дНТФ по 1 мМ каждого в 50 мкл следующего инкубационного буфера: 50 мМ Трис-HCl, рН 7,8; 10 мМ MgCl2; 7 мМ 2-меркаптоэтанола, с нанесенным поверх слоем парафина. Фрагменты ДНК разделяли на 1% агарозном геле, содержащем бромид этидия. В отсутствие дНТФ можно было обнаружить следы фрагментов ДНК либо отсутствие ДНК, тогда как в присутствии дНТФ фрагменты ДНК оставались целыми.
ПРИМЕР 6
Клонирование гена ДНК полимеразы Carboxydothermus hydrogenoformans
Получение хромосомной ДНК из Carboxydothermus hydrogenoformans.
0,8 г биомассы Carboxydothermus hydrogenoformans cycпендировали в 20 мл 1 М KCl и центрифугировали. Осадок снова суспендировали в 4,8 мл SET-буфера (150 мМ NaCl, 15 мМ ЭДТА, рН 8,0, 60 мМ Трис-HCl, рН 8,0, 50 мкг/мкл РНКазы А), после чего добавляют 1 мл 20% SDS и 50 мкл протеиназы К (10 мг/мл). Полученную смесь выдерживали при 37oС в течение 45 мин. После экстрагирования фенолом и хлороформом ДНК осаждали этанолом и растворяли в H2O. Таким образом получали около 4,1 мг ДНК.
Амплификация специфической ДНК с помощью ПЦР.
Для амплификации гена, кодирующего ДНК-полимеразу Carboxydothermus nydrogenoformans с помощью ПЦР, конструировали два смешанных олигонуклеотида (праймер 1 и праймер 2) на основе консервативных участков ДНК-полимераз семейства А, как опубликовано Braithwaite D.K. and Ito J. (1993) Nucl. Acids Res. 21, 787-802.
Последовательность ИД 1
Праймер 1: 5'-CCN AAY YTN CAR AAY ATH-3'
Последовательность ИД 2:
Праймер 2: 5'-YTC RTC RTG NAC YTG-3'
ПЦР-амплификацию проводили в 100 мкл буфера, содержащего 750 нг геномной ДНК из Carboxydothermus hydrogenoformans, 10 мМ Трис-HCl, рН 8,8, 2,5 мМ MgCl2, 50 мМ KCl, 200 мкМ дНТФ, 100 пмоль каждого праймера и 2,5 ЕД. Taq-полимеразы (Boehringer Mannheim, GmbH, FRG). Последовательность-мишень амплифицировали, вначале денатурируя при 95oС в течение 2 мин, а затем осуществляя 30 циклов при 95oС в течение 0,5 мин, 47oС в течение 1 мин, 72oС в течение 2 мин. Циклическую реакцию ПЦР осуществляли в термореакторе Perkin Elmer GenAmp 9600. Электрофорез в агарозном геле показал, что специфически амплифицировался фрагмент примерно 600 пар оснований. Этот фрагмент лигировали в вектор pCRTMII (Invitrogen), и последовательность определяют, используя циклосеквенирование. Аминокислотная последовательность, установленная на основании данной нуклеотидной последовательности, оказалась очень близка к последовательности для других известных ДНК-полимераз, так что праймеры 3 и 4 можно было сконструировать для обратной ПЦР.
Последовательность ИД 3
Праймер 3: 5'-GGG CGA AGA CGC TAT ATT CCT GAG C-3'
Последовательность ИД 4:
Праймер 4: 5'-GAA GCC TTA ATT CAA TCT GGG AAT AAT C-3'
Обратную ПЦР проводили, по существу, в соответствии с методикой, описанной Triglia Т. et al. (1988) Nucleic Acids Res. 16, 8186. 5 мкг геномной ДНК из Carboxydothermus hydrogenoformans расщепляли EcoRI в соответствии с инструкциями изготовителя (Boehringer Mannheim, GmbH) и обрабатывали равным объемом смеси фенол/хлороформ. Водную фазу удаляли и ДНК осаждали этанолом и собирали центрифугированием.
Для циклизации расщепленную ДНК разбавляли до концентрации 50 нг/мкл в лигирующем буфере (Boehringer Mannheim, GmbH, FRG). Реакцию лигирования проводили путем добавления Т4 ДНК-лигазы (Boehringer Mannheim GmbH, FRG) в концентрации 0,2 ед/мкл, и реакцию вели в течение 15 ч при 15oС. Затем лигированную ДНК осаждали этанолом и собирали центрифугированием.
ПЦР проводили в 50 мкл буфера, содержащего 50 мМ Трис-HCl, рН 9,2; 16 mm (NH4)2SO4; 2,25 мМ MgCl2; 2% (об/об) ДМСО, 0,1% (об/об) TweenTM20, 700 нг циклической ДНК, полученной, как указано выше, 50 пмоль каждого праймера, 500 мкМ дНТФ и 0,75 мкл смеси ферментов (Expand Long Template PCR System, Boehrinqer Mannheim, GmbH).
Условия протекания циклов были следующие:
1 • денатурирование матрицы в течение 2 мин при 92oС

Электрофорез на агарозном геле выявил специфически амплифицированный фрагмент ДНК длиной 7000 пар оснований. Фрагмент ДНК лигировали в вектор pCRTMII (Invitrogen) и секвенировали. Оказалось возможным сконструировать установленные на основании данной последовательности праймеры 5 и 6, кодирующие 5'- и 3'- концы участка полимеразы соответственно. Праймер 5 содержит NcoI-сайт, а праймер 6 содержит BamHI-сайт. ПЦР проводили в описанных выше условиях (условия обратной ПЦР), используя 750 нг геномной ДНК из Carboxydothermus hydrogenoformans в качестве матрицы.
Последовательность ИД 5:
Праймер 5: 5'-CGA ATT CAA TCC ATG GGA ААА GTA GTC CTG GTG GAT-3'
Последовательность ИД 6:
Праймер 6: 5'-CGA ATT CAA GGA ТСС ТТА СТТ CGC ТТС АТА ССА GTT-3'
Клонирование и экспрессия
Продукт ПЦР очищали с помощью электрофореза 20 мкл смеси ПЦР в 0,8% агарозном геле. Полосу 2,496 кДа участка, кодирующего полимеразу, выделяли из агарозы, экстрагируя фенолом. Затем ДНК обрабатывали хлороформом и осаждали этанолом. Осадок ресуспендировали и расщепляли NcoI и BamHI в соответствии с инструкциями изготовителя (Boehringer Mannheim, GmbH) для получения липких концов для направленного клонирования. ДНК лигировали в экспрессирующий вектор pDS56 ( , Matile H. и Garotta G. (1990) Immunological Methods, Letkovcs, I. и Pernis, В., eds.), который также расщепляли NcoI и BamHI. Продукты лигирования вводили в штамм BL21(DE3)pUBS520 E. coli (Brinkmann U. et al. (1989) Gene 85, 109-114) путем трансформации. Трансформанты культивировали на L-агаре, содержащем 100 мкг/мл ампициллина и 50 мкг/мл канамицина, чтобы обеспечить селекцию рекомбинантов. Колонии отбирали и выращивали в L-бульоне, содержащем 100 мкг/мл ампициллина и 50 мкг/мл канамицина, и плазмидную ДНК получали путем щелочного лизиса. Плазмиды скринировали на предмет включения вставок, расщепляя BamHI. Рекомбинанты, содержащие вставки, выращивали в L-бульоне, содержащем ампициллин и канамицин, и тестировали на предмет экспрессии термофильной ДНК-полимеразы, индуцируя экспотенциальный рост культур 1 мМ IPTG и анализируя термообработанные экстракты на активность ДНК-полимеразы, как описано выше (определение ДНК-полимеразной активности). Получали рекомбинанты, экспрессирующие ДНК-полимеразу из Carboxydothermus hydroqenoformans. Этот штамм обозначали Е. coli AR96 (DSM 11179) и плазмиду pAR4.
, Matile H. и Garotta G. (1990) Immunological Methods, Letkovcs, I. и Pernis, В., eds.), который также расщепляли NcoI и BamHI. Продукты лигирования вводили в штамм BL21(DE3)pUBS520 E. coli (Brinkmann U. et al. (1989) Gene 85, 109-114) путем трансформации. Трансформанты культивировали на L-агаре, содержащем 100 мкг/мл ампициллина и 50 мкг/мл канамицина, чтобы обеспечить селекцию рекомбинантов. Колонии отбирали и выращивали в L-бульоне, содержащем 100 мкг/мл ампициллина и 50 мкг/мл канамицина, и плазмидную ДНК получали путем щелочного лизиса. Плазмиды скринировали на предмет включения вставок, расщепляя BamHI. Рекомбинанты, содержащие вставки, выращивали в L-бульоне, содержащем ампициллин и канамицин, и тестировали на предмет экспрессии термофильной ДНК-полимеразы, индуцируя экспотенциальный рост культур 1 мМ IPTG и анализируя термообработанные экстракты на активность ДНК-полимеразы, как описано выше (определение ДНК-полимеразной активности). Получали рекомбинанты, экспрессирующие ДНК-полимеразу из Carboxydothermus hydroqenoformans. Этот штамм обозначали Е. coli AR96 (DSM 11179) и плазмиду pAR4.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ СИНТЕЗА кДНК С ИСПОЛЬЗОВАНИЕМ ТЕРМОСТАБИЛЬНОЙ ДНК-ПОЛИМЕРАЗЫ | 1997 |
|
RU2198222C2 |
| СПОСОБ АМПЛИФИКАЦИИ ДНК, СПОСОБ КЛОНИРОВАНИЯ ВТОРОЙ кДНК И СПОСОБ ОБРАТНОЙ ТРАНСКРИПЦИИ ДНК | 1997 |
|
RU2297454C2 |
| СПОСОБ АМПЛИФИКАЦИИ ДНК И КОМПОЗИЦИЯ ДЛЯ ЕГО ОСУЩЕСТВЛЕНИЯ | 2000 |
|
RU2260055C2 |
| ОЧИЩЕННАЯ ДНК-ПОЛИМЕРАЗА, РЕКОМБИНАНТНАЯ ДНК-ПОЛИМЕРАЗА, СПОСОБЫ ИХ ПОЛУЧЕНИЯ, ВЫДЕЛЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ ДНК И НАБОР ДЛЯ ПОЛИМЕРАЗНОЙ ЦЕПНОЙ РЕАКЦИИ | 1998 |
|
RU2221866C2 |
| ОЧИЩЕННАЯ ТЕРМОСТАБИЛЬНАЯ ДНК-ПОЛИМЕРАЗА ИЗ THERMOCOCCUS GORGONARIUS, ЕЕ ПОЛУЧЕНИЕ И ПРИМЕНЕНИЕ | 1997 |
|
RU2221867C2 |
| МОДИФИЦИРОВАННАЯ ТЕРМОСТАБИЛЬНАЯ ДНК-ПОЛИМЕРАЗА, СПОСОБ ЕЁ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЕ | 1997 |
|
RU2235773C2 |
| СУХАЯ СМЕСЬ ДЛЯ ПРИГОТОВЛЕНИЯ РЕАКЦИОННОЙ СМЕСИ ДЛЯ АМПЛИФИКАЦИИ НУКЛЕИНОВОЙ КИСЛОТЫ И СПОСОБ ЕЕ ПОЛУЧЕНИЯ | 2012 |
|
RU2535995C2 |
| ДИИЗОПРОПИЛФТОРФОСФАТАЗА, ЕЕ ПРИМЕНЕНИЕ И ПОЛУЧЕНИЕ | 1999 |
|
RU2235774C2 |
| КОМПОЗИЦИИ, СПОСОБЫ И НАБОРЫ ДЛЯ СИНТЕЗА И ОБНАРУЖЕНИЯ НУКЛЕИНОВЫХ КИСЛОТ | 2017 |
|
RU2757416C2 |
| СПОСОБ ОБНАРУЖЕНИЯ N-КОНЦЕВОГО ПРО-МНП | 2000 |
|
RU2218568C2 |

















Изобретение относится к биотехнологии. ДНК-полимераза получена из термофильной эубактерии Carboxydothermus hydrogenoformans или Е. coli BL21 (DE3) pUBS520pAR4 DSM 11179. ДНК-полимераза проявляет зависимую от ионов магния обратнотранскриптазную активность и 3'-5'-экзонуклеазную активность, имеет кажущуюся молекулярную массу около 100-105 кДа. Изобретение включает также рекомбинантные плазмиды и трансформированные клетки-хозяева, способные продуцировать фермент. Фрагмент выделенной ДНК с нуклеотидной последовательностью, приведенной в описании, кодирует термостабильную ДНК-полимеразу. Фрагмент ДНК получают из Carboxydothermus hydrogenoformans. Культивируют природный штамм Carboxydothermus hydrogenoformans или Е. coli BL21 (DE3) pUBS520pAR4 DSM 11179. Клетки штамма суспендируют в буфере, разрушают клетки. Очищают выделенную ДНК-полимеразу методом хроматографии. При амплификации, введении метки и проведении обратной транскрипции используют термостабильную ДНК-полимеразу. Изобретение обеспечивает получение ДНК-полимеразы, активной при более высоких температурах, обладающей 3'-5'-полимеразной активностью и обратнотранскриптазной активностью в присутствии ионов магния и отсутствии ионов марганца. 6 с. и 2 з.п. ф-лы, 12 ил.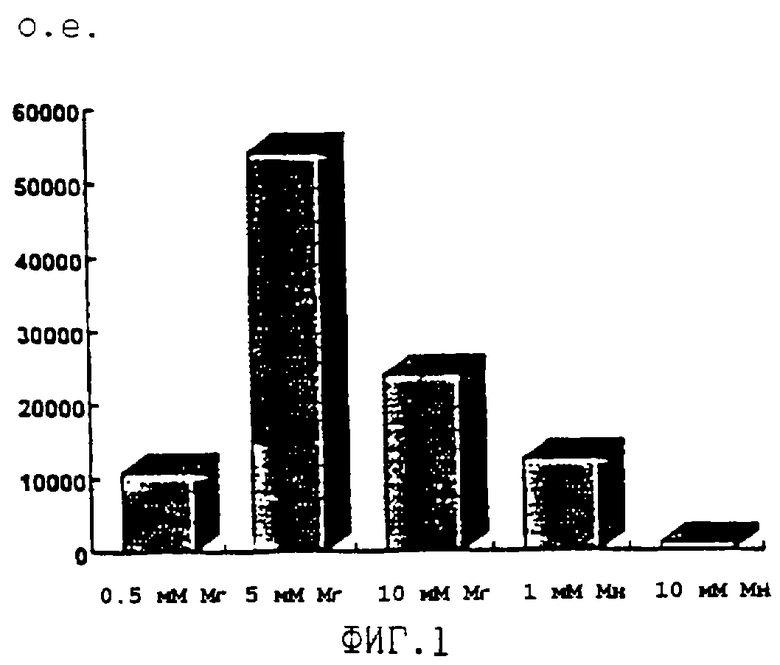
| WO 9203556 А, 05.03.1992 | |||
| WO 9610640 А, 11.04.1996 | |||
| PERLER F.B et al | |||
| Thermostable DNA polymerases Advances in protein Chemistry, 1996, v.48, p.377-435. |

