Изобретение относится к новому белку - человеческому фактору стволовых клеток (ФСК, SCP), ДНК - последовательностям, кодирующим этот белок, его применению в терапии, в частности при оплодотворении in vitro, а также к фармацевтическим композициям, содержащим такой белок.
Успешная имплантация эмбриона требует правильного развития доимплантационного эмбриона, приводящего в результате к вызревшей бластоцисте, которая пригодна для имплантации в рецептивный эндометрий. Собрано значительное количество данных, которые подтверждают предположение, что растворимые факторы роста, если они секретированы эпителием матки, непосредственно воздействуют на эмбрион, регулируя этот процесс (Pampfer, S., et al, Bioessays, 13: 535-540 (1991); Tartakousky, B., and Ben Yair, E., Development Biology, 146: 345-352 (1991); Anderson, E.D. J.Cellular Biochem., 53; 280-287 (1993); и Schultz, G.A. and Hevner S., Mutat. Res., 296; 17-31 (1992)).
Кроме того, показано, что развивающиеся эмбрионы продуцируют ряд цитокинов, которые могут действовать на эндометрий как аутокрины (autocrine) и влияют на его восприимчивость. Примеры указанных факторов роста, которые продуцируются человеческими эмбрионами, включают ИЛ-1, ИЛ-6, КСФ-1 и ФHO-α (Zolti et al, Fertil. Steril., 56 (1991) 265-272, и Witkin et al, J.Reprod. Immunol., 19 (1991) 85-93).
Показано, что ФНО-α присутствует в культуральной среде человеческих эмбрионов до стадии морула, но не в среде бластоцисты (Lachappelle et al, Human Reproduction, 8: 1032-1038 (1993)). Продуцирование цитокинов эмбрионами может, следовательно, регулироваться стадий-специфическим образом.
Данные о возможном прямом действии цитокинов на эмбрионы первоначально были получены при экспериментах с мышами, когда было показано, что многие цитокины влияют на развитие доимплантационных эмбрионов in vitro.
γ-Интерферон и КСФ-1 при физиологических концентрациях ингибируют развитие многих эмбрионов до стадии бластоцисты (Hill et al, J. Iimmunol., 139 (1987) 2250-2254). Также показано, что ФНО-α обладает менее заметным действием. Хотя ФНО-α не оказывает явного действия на скорости образования бластоцисты, этот фактор специфически ингибирует пролиферацию клеток, вносящих вклад во внутреннюю массу клетки (ICM), что дает, в результате, бластоцисты с уменьшенной ICM (Pamplfer et al, Endocrinology, 134; 206-212 (1994)).
Другие факторы роста также имеют специфическое влияние на ICM клеток. Например, инсулиноподобные факторы роста 1 и 2 стимулируют пролиферацию ICM, в то время как фактор ингибирования лейкемии (ФИЛ, LIP) ингибирует их дифференцировку (Harvey et al, Mol. Reprod. Dev., 31 (1992) 195-199).
На мышиных моделях наблюдали, что эмбрионы, культивированные in vitro, отстают в развитии при сравнении с контролем in vivo и обнаруживается меньший коэффициент беременности после трансплантации эмбриона (Bowman, P., and McLaren, A. , J.Embriol. Exp. Morphol., 24; 203-207 (1970)). Таким образом, лучшее понимание роли факторов роста при развитии может привести к улучшению условий культивирования in vitro и к усилению эффективности программ оплодотворения человека in vitro (IVF).
Фактор стволовых клеток (ФСК, SCF) является фактором роста, отнесенным по структуре к КСФ-1, и действует через тирозинкиназный рецептор с-набора (с-kit). В костном мозге ФСК и КСФ-1 действуют синергически, чтобы промотировать пролиферацию и дифференцировку стволовых клеток в макрофаговых колониях. В EP-A-0423980 описывается полинуклеотидная последовательность человеческого ФСК и обсуждается возможное применение ФСК при условиях, требующих стимуляции клеточной пролиферации, в частности, клеток крови.
На мышах показано, что с-набор экспрессируется в течение всего времени доимплантационного развития (Arceci et al (1992)). Заявители теперь обнаружили, что то же справедливо для человеческих эмбрионов. На некоторых стадиях человеческие эмбрионы также экспрессируют мРНК ФСК, причем предполагается, что этот фактор роста может действовать наподобие аутокрина. Это противоположно случаю с мышами, когда экспрессия ФСК не обнаруживается в доимплантационных эмбрионах (Arceci et al (1992)).
Первичный транскрипт ФСК состоит из восьми экзонов (Martin, F.H. et al, Cell, 63: 203-211 (1990), и в этой работе также описывается вариантная форма ФСК. Также описан сплайс-вариант ФСК, который появляется вследствие потери экзона 6 (Flanagan et al, Cell, 63:1025-1035 (1991)).
Теперь также обнаружен иной, новый сплайс-вариант, который появляется вследствие включения между экзонами 3 и 4 нового экзона, состоящего из 155 пар оснований. Это приводит также к сдвигу рамки генетического кода, и кодирует разновидность ФСК, содержащую 33 новых аминокислоты после экзона 3, перед окончанием при внутрирамочном стоп-кодоне, который теперь находится в экзоне 4 вследствие сдвига в рамке считывания.
Таким образом, настоящее изобретение относится к ФСК, который включает следующую С-концевую последовательность:
Glu Ile Cys Ser Leu Leu Ile Gly Leu Thr Ala Tyr Lys Glu Leu Ser Leu Pro Lys Arg Lys Glu Thr Cys Arg Ala Ile Gln His Pro Arg Lys Asp Ile Gln His Pro Arg Lys Asp
или последовательность, которая, по существу, гомологична указанной последовательности.
Предпочтительно, новый ФСК изобретения содержит первые 39 аминокислот первичного ФСК (без включения какой-либо сигнальной последовательности), за которыми следует вышеупомянутые новые 33 аминокислоты. В одном из вариантов осуществления изобретения новый ФСК изобретения имеет последовательность, в позициях 1-39 гомологичную, по существу, последовательности, показанной на фиг. 2.
По содержанию аминокислот последовательность белка может рассматриваться, по существу, как гомологичная последовательности другого белка, если значительное число составляющих аминокислот обнаруживают гомологию. Гомологичными могут быть, по меньшей мере 40%, 50%, 60%, 70%, 80%, 90%, 95% или даже 99% аминокислот, при предпочтении в порядке возрастания.
Таким образом, механизм альтернативного сплайсинга может привести, в результате, к продуцированию нового ФСК в человеческих эмбрионах. Следовательно, новый ФСК по изобретению может применяться при обработке доимплантационных эмбрионов для обеспечения правильной дифференцировки и развития перед имплантацией субъекту.
Кроме того, настоящее изобретение также относится к ДНК-последовательности, кодирующей белок изобретения, и указанная последовательность включает последовательность, по существу, гомологичную последовательности
GAA ATC TGT TCA TTG TTG ATA GGG CTG ACG GCC TAT AAG GAA TTA TCA CTC CCT AAA AGG AAA GAA ACT TGC AGA GCA ATT CAG CAT CCA AGG AAA GAC TGA
и включает все другие нуклеотидные последовательности, которые вследствие вырождения генетического кода также кодируют данную аминокислотную последовательность, или которые гомологичны, по существу, такой последовательности.
Последовательности, обладающие существенной гомологией, могут рассматриваться как последовательности, которые будут гибридизироваться с нуклеотидной последовательностью, показанной на фиг. 2, в строгих условиях (например, при 35-65oC в солевом растворе около 0,9 M).
Конструкции ДНК, содержащие ДНК-последовательности изобретения, составляют другой аспект настоящего изобретения.
Как описано здесь, белок изобретения полезен при обработке эмбрионов для обеспечения правильного развития перед имплантацией. Показано, что ФСК действует посредством связывания с с-набором трансмембранных рецепторов. Кроме того, заявители показали, что человеческие эмбрионы экспрессируют с-набор на протяжении большинства стадий развития доимплантационного эмбриона.
Таким образом, в своих других аспектах, настоящее изобретение относится
а) к способу обеспечения правильного развития доимплантационного эмбриона, который включает стадию введения ФСК настоящего изобретения доимплантационному эмбриону (и, предпочтительно, человеческому эмбриону); и
б) к способу обеспечения правильного развития человеческого доимплантационного эмбриона, который включает стадию введения ФСК человеческому доимплантационному эмбриону. При этом способе используемый ФСК может находиться в любой встречающейся в природе форме, включая ранее описанные варианты (Martin et al, цит. выше, и Flangan et al, цит. выше), а также новый вариант, описанный здесь.
Кроме того, настоящее изобретение также относится к применению ФСК при производстве лекарственного средства для применения при обеспечении правильного развития доимплантационных человеческих эмбрионов. И снова, любая форма ФСК может быть использована для получения подходящего лекарственного средства.
Лекарственное средство представляет собой, преимущественно, форму фармацевтической композиции, содержащей белок изобретения вместе с одним или несколькими фармацевтически приемлемыми носителями и/или эксципиентами. Такие фармацевтические композиции образуют еще один аспект настоящего изобретения. Такие фармацевтические композиции представляют один из методов, с помощью которых ФСК можно использовать в описанных выше способах.
Теперь настоящее изобретение будет описываться с помощью приведенных ниже примеров, которые не следует рассматривать как ограничивающие каким-либо образом настоящее изобретение. Примеры отсылают к фигурам, краткое описание которых приводится ниже.
Фиг. 1 представляет последовательность нового экзона и предсказанную аминокислотную последовательность.
На фиг. 2 приводится последовательность человеческого ФСК.
На фиг. 3 показан агарозный гель с продуктами "гнездной" ОТ-ПЦР (обратная транскриптаза - полимеразная цепная реакция) амплификации на РНК из человеческих эмбрионов. Каждая полоса показывает продукты амплификации с затравками, специфическими для различных кДНК-мишеней. Амплифицированные кДНК из различных эмбрионов загружают на каждую дорожку. Дорожки помечают в соответствии с обозначениями кДНК в таблице (см. ниже). Дополнительными образцами являются: дорожка p - трофобласт первого триместра; дорожка q-кДНК из 200 клеток BeWO; дорожка r - 10 нг человеческой геномной ДНК; и дорожка s - без внесения кДНК, как негативный контроль. Маркером молекулярной массы ДНК является лэддер (ladder) 123 пар оснований, загруженных в дорожку 1. Размеры ожидаемых продуктов ПЦР показаны в парах оснований (п.о.).
Таблица
кДНК человеческих эмбрионов и контроли, название стадии развития
a 2 клетка
b 3 клетка
c 4 клетка
d 6 клетка
e 8 клетка
f морула
g бластоциста
h культуральный супернатант для a-g
j три объединенные бластоцисты
k культуральный супернатант для j
l 2х6 клеток и 1х8 клеток
m культуральный супернатант для l
n 1х4 клеток и 1х6 клеток
o культуральный супернатант для n;
образцы a-h от одного донора.
На фиг. 4 показаны затравки, использованные для ОТ-ПЦР, внешняя пара A и B, внутренняя пара C и D.
Пример 1
Эмбриональная культура и экстракция РНК
При этом исследовании используют криоконсервированные человеческие эмбрионы, которые оплодотворены как часть программы IVF. Эти эмбрионы были даны родителями для исследовательских целей, и это исследование проводилось согласно требованиям Комиссии по эмбриологии и оплодотворению человека и местного Комитета по этике. Замороженные эмбрионы оттаивают и культивируют в сбалансированной по солям среде Эрла с добавлением 0,4% человеческого сывороточного альбумина (Armour Pharma icals UK) до требуемой стадии развития, затем быстро замораживают в жидком азоте в 5 мкл культуральной жидкости (и, таким образом, лизируют кристаллами льда). Идентичный объем культурального супернатанта замораживают как контрольный образец. Все остальные скопившиеся клетки удаляют в ходе обычной переработки.
Всю РНК из трофобласта первого триместра выделяют методом Chomsczynski и Sacchi, Anal. Biochem., 162; 156-159 (1987), при котором замороженную ткань гомогенизируют в 5 мл буферного раствора, содержащего 4 М тиоцианата гуанидиния (Gibko BRL Livingston, Шотландия), 25 мМ цитрата натрия, pH 7,0, 0,5% саркозила и 0,1 М 2-меркаптоэтанола. Лизат подкисляют, добавляя 0,5 мл 2 М ацетата натрия, pH 4, и экстрагируют фенолхлороформом с использованием 5 мл буферного раствора, насыщенного фенолом, и 1 мл смеси хлороформа с изоамиловым спиртом (49: 1, о/о). Суспензию помещают на лед на 15 минут и центрифугируют при 10000 g в течение 20 минут при 4oC. Водную фазу, содержащую РНК, преципитируют, промывают дважды в 70% этаноле, сушат и ресуспендируют в ТЭ (10 мМ трис-HCl, pH 7,4, и 1 мМ ЭДТК). Концентрацию РНК определяют спектрофотометрически при 260 нм.
РНК получают из эмбрионов отдельного человека с использованием протокола пропорционального уменьшения, основанного на вышеописанной процедуре. Чтобы способствовать преципитации РНК, на стадии гомогенизации добавляют 100 мкг тРНК дрожжевого носителя (Gibco BRL, Livingston, Шотландия). Остальные детали такие же, какие описаны выше, за исключением того, что все объемы в 50 раз меньше, и всю процедуру выполняют в 400-мкл пробирках Эппендорфа.
Пример 2
Полимеразная цепная реакция с обратной транскриптазой (ОТ-ПЦР)
Синтезируют кДНК из половины общего количества РНК из каждого эмбриона, используя обратную транскриптазу AMV (Super RT, HT Biotech, Кэмбридж, Соед. Кор.). Праймируют 305 мкг РНК олиго-dT (Pharmacia), в соответствии с инструкциями изготовителей, в течение 60 минут при 42oC. ПЦР-амплификацию препаратов кДНК осуществляют, как описано ранее (Snarkey, A.et al, Molecular Endocrinol., 6: 1235-1241 (1992)), с термоячейкой для (Hybaid) омнигенной ДНК в конечном объеме 30 мкл с использованием 1 E ДНК-полимеразы Taq (Cetus, Emeryville, CA) и 10 мкМ каждой из пар наружных затравок (см. фиг. 4) в рекомендованном изготовителем буфере. Используют следующий циклический профиль, 30 с при 95oC, 30 с при XoC, 30 с при 72oC в течение 30 циклов, где X является температурой отжига для каждой пары цитокиновых затравок, как показано ниже.
Наружные затравки (oC)
ФСК - 54
Гис-тРНК - 52
с-набор - 56
Внутренние затравки (oC)
ФСК - 54
Гис-тРНК - 59
с-набор - 56
Олигонуклеотидные затравки
Олигонуклеотидные затравки для ФСК, с-набора и гистидил-тРНК-синтетазы синтезируют на синтезаторе ДНК PS250 Cruachem. Последовательности затравок конструируют из опубликованных нуклеотидных последовательностей (см. фиг. 4) таким образом, что амплификация любой контаминирующей геномной ДНК приведет в результате к продуктам разного размера из образцов кДНК.
Из-за небольшого количества материала используют две пары затравок для каждой кДНК-мишени, по протоколу гнездной ПЦР. Одну тридцатую кДНК-продуктов амплифицируют с использованием Amplitaq (Cetus) в рекомендованном изготовителями буфере. После 30 циклов ПЦР с использованием наружной пары затравок одну пятидесятую реакционной смеси первого цикла переносят в другую пробирку, содержащую внутреннюю пару затравок, и подвергают амплификации еще на протяжении 30 циклов. В качестве негативного контроля равный объем культуральной жидкости, в которой выращивают эмбрионы, экстрагируют и подвергают ОТ-ПЦР по той же схеме. Кроме того, в качестве позитивного контроля экстрагируют 200 клеток клеточной линии BeWo (ЕСАСС N 86082803).
Затравки, используемые при этом исследовании, показаны на фиг. 4 вместе с размером ожидаемого продукта. Идентичность каждого продукта подтверждают путем клонирования и секвенирования, как описано ранее (Sharkey et al, Mol. Endocrinol. (1992)). Чтобы убедиться, что обнаруженный продукт является результатом амплификации кДНК, а не контаминирующей геномной ДНК, выбирают затравки, чтобы перекрыть интрон-экзонные стыки. Также 10 нг геномной ДНК подвергают ПЦР в то же время, что и кДНК, чтобы проверить отсутствие продукта ожидаемого размера, образованного из геномной ДНК.
Результаты
Метод ОТ-ПЦР применяют ко всей РНК, экстрагированной из человеческих эмбрионов, полученных путем оплодотворения in vitro. Эмбрионы выращивают до соответствующей стадии, затем быстро замораживают, и экстрагируют всю РНК. Чтобы получить поддающийся обнаружению продукт ОТ-ПЦР из всей РНК, экстрагированной из одного эмбриона, используют протокол гнездной ПЦР, по которому кДНК подвергают двум сериям ПЦР-амплификации с наружной парой затравок, а затем с внутренней парой. Затравки основаны на опубликованных кДНК-последовательностях, и сконструированы для перекрытия интрон-экзонных стыков, чтобы амплификация контаминирующей геномной ДНК могла быть легко отделена от продуктов кДНК.
Сначала кДНК от каждого эмбриона проверяют с праймерами для гистидил-тРНК-синтетазы (гистPC), чтобы подтвердить успешное выделение РНК и обратную транскрипцию. Используемые затравки дают начало неустойчивым продуктам более 400 п. о. из геномной ДНК и 110 п.о. из кДНК, произошедшей от мРНК гистPC. Транскрипты для гистPC обнаруживаются в мРНК из эмбрионов на всех стадиях развития, так же как и в отпадающей оболочке матки и в хориокарциномной клеточной линии BeWo, используемых в качестве позитивного контроля (фиг. 3, дорожки p и q, соответственно). Продукт не обнаруживается в равном объеме супернатанта культуры эмбрионов, экстрагированного и подвергнутого ОТ-ПЦР по той же схеме, что указывает на отсутствие контаминации культуры посторонней кДНК или РНК.
Примеры подобного ОТ-ПЦР анализа с затравками для ФСК и с-набора приведены на фиг. 3. Запасы кДНК обратно транскрибируют из каждого образца РНК на два отдельных события, и ПЦР-анализы повторяют дважды на каждой исходной кДНК. Результаты показаны на фиг. 3, на котором выделены образцы экспрессии с-набора и ФСК во время доимплантационного развития. Идентичность фрагментов ПЦР правильного размера подтверждают секвенированием клонированных продуктов ПЦР. В случае, когда обнаруживаются продукты нового размера, их также клонируют и секвенируют.
Для ФСК предсказанный фрагмент составляет 966 п.о. Однако, оказывается, что транскрипты ФСК показывают стадий-специфические различия в размерах. После клонирования и секвенирования оказывается, что из-за события альтернативного сплайсинга, который встраивает новый экзон между экзонами 3 и 4, появляется новый продукт. Предсказанная последовательность нового транскрипта показана на фиг. 1. Картина нового сплайсинга также включает сдвиг рамки считывания, давая в итоге 33 новых аминокислоты перед внутрирамочным стоп-кодоном в экзоне 4.
При подобном анализе с использованием затравок, специфических для с-набора, рецептор для ФСК показывает, что с-набор экспрессируется на большинстве стадий развития человеческого доимплантационного эмбриона. Это наводит на мысль, что эмбрион обладает способностью отвечать на ФСК на протяжении этого периода.
Обсуждение
Показано, что многие факторы роста влияют на развитие выращиваемых доимплантационных эмбрионов млекопитающих (для обзора см. Anderson, E.D., J. Cellular Biochem., 53; 280-287 (1993), и Schultz, G.A. and Hevner S., Mutat. Res., 296; 17-31 (1992).
Однако имеются убедительные данные о различиях от образца к образцу в экспрессии рецепторов факторов роста при доимплантационном развитии. Например, мРНК ЭФР (эпидермальный фактор роста) экспрессируется в эмбрионе свиньи, но не обнаружен на какой-либо стадии с мышиных доимплантационных эмбрионах (Vaughan et al, Development, 116; 663-669 (1992); Rapolee et al. Science, 241; 1823-1825 (1988): Watson, A. J. et al, Biol. Reprod., 50; 725-733 (1994)).
Следовательно, полезность этих исследований для поиска в направлении факторов, регулирующих развитие человеческого доимплантационного эмбриона, ограничена. Кроме того, специфические факторы роста и рецепторы, исследованные в указанных работах, часто выбирались на данный случай (ад хок). Как по моральным, так и по практическим соображениям, такой подход не является подходящим для применения в случае человеческих эмбрионов. Поэтому заявители использовали метод "гнездной" ОТ-ПЦР, который позволил произвести скрининг для экспрессии фактора роста и рецепторных мРНК в доимплантационных эмбрионах отдельного человека. Этот способ широко применяется в последние несколько лет для других видов, так как он надежен, чувствителен и экономичен при работе с эмбриональным материалом.
ОТ-ПЦР с затравками для гистидил-тРНК-синтетазы применяли на образцах кДНК для подтверждения того, что кДНК успешно получена из образца РНК каждого эмбриона. Специфическая для этого обязательного гена кДНК успешно обнаружена в образцах кДНК, полученных даже из отдельного 2-клеточного эмбриона, что указывает, что метод достаточно чувствителен для этой стадии.
ФСК экспрессирован на 2-клеточной стадии и затем снова появлялся на 6-клеточной стадии. Это совместимо с материнской экспрессией с последующей реэкспрессией из генома эмбриона на 6-клеточной стадии (Braude, P., et al, Nature, 332; 459-461 (1988)). Оказалось, что транскрипты ФСК обнаруживают стадия-зависимые различия в величине транскрипта. Они обнаружены после клонирования и секвенирования вследствие альтернативного сплайсинга первичного транскрипта. Два из этих вариантов подобны вариантам, опубликованным ранее (Martin et al, цит. выше, и Sharkey et al, Mol. Endocrinol., 6: 1235-1241 (1992)), и один вариант представляет новую форму, которая предсказывает образец ФСК с 33 новыми аминокислотами у карбоксильного конца. Теперь известно несколько вариантов ФСК, некоторые из которых являются мембраносвязанными и биоактивными. Разновидности, экспрессированные доимплантационным эмбрионом, включают образцы, о которых известно, что они биоактивны, и показано, что различные формы ФСК могут действовать через с-набор, экспрессированный эмбрионом, и могут влиять на развитие эмбриона в этот период.
Список последовательностей приведен в конце описания.











Изобретение касается нового белка - человеческого фактора стволовых клеток (ФСК). ДНК кодирует ФСК, имеющий аминокислотную последовательность, приведенную для положений 1-39 на фиг. 2. Начиная с положения 40 ФСК включает следующую С-концевую аминокислотную последовательность Glu Ilе Cys Ser Leu Leu Ile Cly Leu Thr Ala Tyr Lys Glu Leu Ser Leu Pro Lys Arg Lys Glu Thr Cys Arg Ala Ile Gln His Pro Arg Lys Asp. ФСК обеспечивает правильное развитие доимплантационных эмбрионов при обработке им эмбрионов. 2 с. и 1 з.п. ф-лы, 4 ил., 1 табл.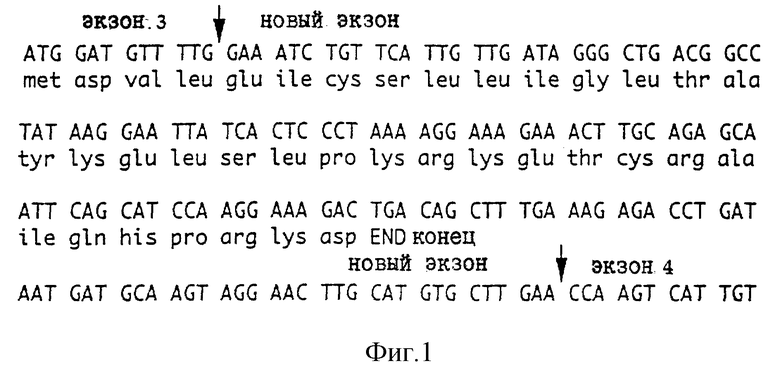
Glu Ile Cys Ser Leu Leu Ile Gly Leu Thr Ala Tyr Lys Glu Leu Ser Leu Pro Les Arg Lys Glu Thr Cys Arg Ala Ile Gln His Pro Arg Lys Asp
2. ДНК по п.1, отличающаяся тем, что включает следующую последовательность:
GAA ATC TGT TCA TTG TTG ATA GGG CTG ACG GCC TAT AAG GAA TTA TCA CTC CCT AAA AGG AAA GAA ACT TGC AGA GCA ATT CAG CAT CCA AGG AAA GAC TGA.
| Способ получения последовательности ДНК, содержащей фрагмент, кодирующий человеческий проаполипопротеин А-1 | 1988 |
|
SU1834904A3 |
| КРИОГЕННЫЙ СОСУД | 1972 |
|
SU423980A1 |
