ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Изобретение относится к области биотехнологии. Оно описывает новый антагонист интерлейкина-1 (IL-1), активный в отношении как IL-1a, так и IL-1В, новую последовательность DNA, кодирующую антагонист IL-1, и способ получения антагониста IL-1 методикой рекомбинантной DNA. Оно также описывает профилактическое, терапевтическое и диагностическое применение такого нового антагониста IL-1 при патологиях, зависящих от образования IL-1.
ПРЕДПОСЫЛКИ К СОЗДАНИЮ ИЗОБРЕТЕНИЯ
Существуют два различных гена, кодирующих интерлейкин-1 (IL-1), названные IL-1а и IL-1В, которые кодируют белок IL-1а и IL-1В, соответственно.
Интерлейкины IL-1а и IL-1В являются плейотропными цитокинами, которые, хотя их последовательности и обнаруживают недостаточную аналогию, оказывают множество сходных воздействий на различные ткани и влияют на многие патологии человека, в частности на иммунный ответ организма и на воспалительные процессы.
Оба белка имеют молекулярную массу около 17,5 кД, и они предварительно синтезируются как молекула предшественника большего размера, имеющая молекулярную массу около 31 кД.
IL-1'ы являются сильными воспалительными и пирогенными цитокинами, которые обычно имеют полезные эффекты, но могут также иметь эффекты, вредные для здоровья организма.
Они могут, например, принимать участие в патогенезе симптомов аутоиммунных патологий, подобных красной волчанке, в частности, они участвуют как медиаторы в провоцировании поражения тканей, как, например, при ревматоидном артрите.
Многие биологические эффекты IL-1 подобны тем, которые могут наблюдаться в случае сепсиса. Последние исследования продемонстрировали, что внутривенное введение IL-1 в дозах от 1 до 10 нг/кг вызывает лихорадочное состояние, сонливость, отсутствие аппетита, общую миалгию, артралгию и сильную головную боль.
Так как IL-1 имеет плейотропные биологические свойства, многие из которых отрицательно воздействуют на организм, мощные эффекты IL-1 должны находиться под четким физиологическим контролем.
Синтез IL-1 ингибируется противовоспалительными цитокинами, простагландинами и глюкокортикоидами, и наличие множества уровней ингибироваиия IL-1 указывает на необходимость точного регулирования этого медиатора.
IL-1 является единственным цитокином, для рецептора которого до настоящего момента описан антагонист-полипептид, третьим известным на сегодня компонентом семейства IL-1 является антагонист рецептора IL-1 (IL-1rа).
Все три компонента (IL-1a, IL-1В и IL-1ra) распознаются и связываются с одним и тем же рецептором на поверхности клетки (IL-1R); связывание IL-1a и IL-1В с IL-1R передает сигнал, a IL-ra - нет.
Существует два типа рецепторов IL-1, названные IL-1RI и IL-1RII. IL-1ra является полипептидом, который связывает IL-1RI и обладает меньшим сродством к IL-1RII без какой-либо агонистической активности.
Образование IL-1ra вызывается IgG, цитокинами и бактериальными продуктами в различных клеточных типах, включая мононуклеарные фагоциты, полиморфонуклеарные клетки (ПМН) и фибробласты.
До сих пор идентифицированы и клонированы две молекулярные формы IL-1ra: 1) секретированный IL-1ra (sIL-1ra) содержит классическую лидерную последовательность из 25 аминокислот, дающую зрелый белок из 152 аминокислот; 2) внутриклеточный IL-1ra (icIL-1ra) испытывает недостаток в лидерной последовательности, а это предопределяет то, что белок остается внутриклеточным.
sIL-1a и icIL-1ra образуются из одного и того же гена. Транскрипты icIL-1ra берут начало от сайта альтернативной инициации и от сплайсинга первого альтернативного экзона во внутреннем акцепторном участке сплайсинга, размещенном в первом экзоне sIL-1ra. Прогнозируемые белки, таким образом, идентичны за исключением их NН2-концов, где первые 21 аминокислота sIL-1ra замещены четырьмя аминокислотами в icIL-1ra.
Экспрессия транскриптов, кодирующих sIL-1ra и icIL-1ra, регулируется по-разному. Биологическая значимость icIL-1ra еще не выяснена.
Подразумевается, что IL-1 принимает участие в патогенезе многих болезней, поэтому очевидна необходимость иметь доступные медикаменты, пригодные для ограничения вредных для здоровья эффектов IL-1.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Цель настоящего изобретения - обеспечить антагонист IL-1 активный в отношении как IL-1a, так и IL-1B и их комбинации.
Следующая цель настоящего изобретения - обеспечить DNA последовательность, кодирующую антагонист IL-1, и способ получения такого нового антагониста методикой рекомбинантной DNA.
Другая цель настоящего изобретения - обеспечить антагонист в практически очищенной форме, чтобы он был пригоден для применения в фармацевтических композициях, действенных против патологий, которые требуют ингибирования IL- 1.
Дальнейшие цели и преимущества изобретения станут очевидными в следующем описании.
КРАТКОЕ ОПИСАНИЕ ФИГУР И СПИСКА ПОСЛЕДОВАТЕЛЬНОСТЕЙ
Фиг. 1 описывает последовательность DNA и последовательность белка для части вне общей, icIL-1raII (SEQ ID NO:8 и SEQ ID NO:9) в сравнении с тем же для sIL-1ra (icIL-1raI; SEQ ID NO:6) и sIL-1ra (SEQ ID NO:4 и SEQ ID NO:5), а также описывает DNA последовательность и кодируемый ею белок для общей части IL-1ra (SEQ ID NO:13 и SEQ ID NO:14).
Фиг. 2 (A, B, C, C) описывает RT-PCR анализ экспрессии icIL-1raII в различных типах клеток.
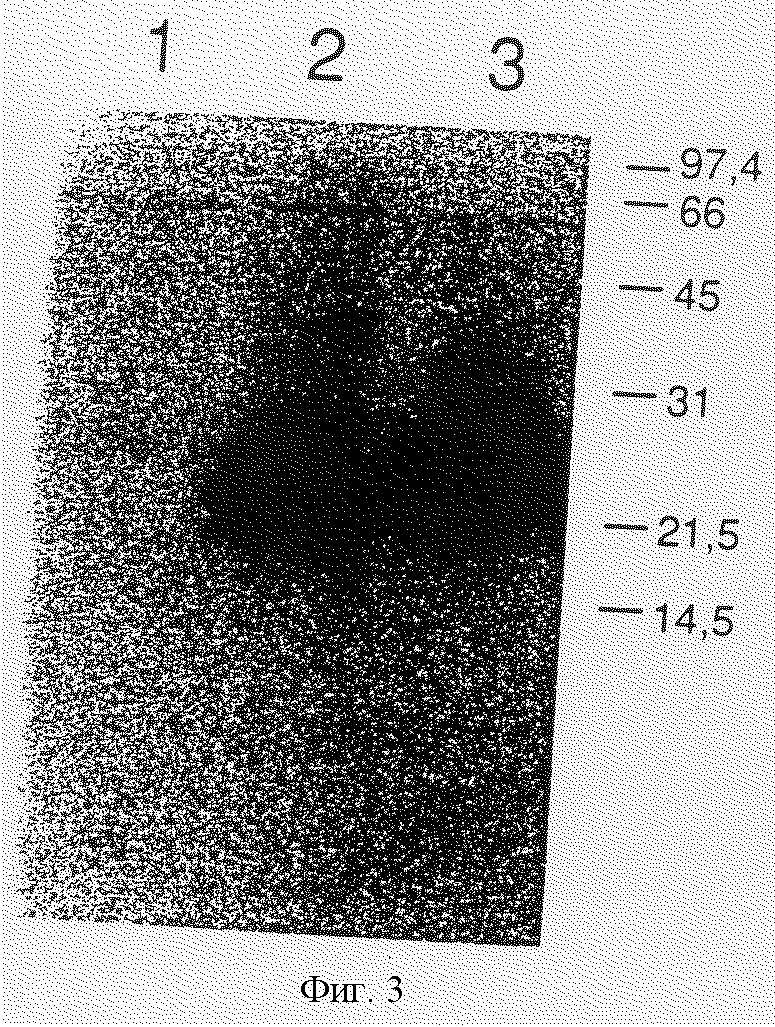
Фиг. 3 описывает вестерн-блоттинг рекомбинантного icIL-1raII.
Фиг. 4 (A, B) описывает влияния icIL-1raII на индуцированную IL-1 экспрессию Е-селектина в эндотелиальных клетках.
SEQ ID NO:1 представляет последовательность олигонуклеотида, названного IRА5, для применения в RT-PCR.
SEQ ID NO:2 представляет последовательность олигонуклеотида, соответствующую нуклеотидам 69-70 В-актина cDNA, для применения в RT-PCR.
SEQ ID NO:3 представляет последовательность обратного олигонуклеотида, комплементарного нуклеотидам 430-449, для применения в RT-PCR.
SEQ ID NO:4 представляет DNA последовательность, кодирующую sIL-1ra, для части вне общей.
SEQ ID NO:5 представляет аминокислотную последовательность sIL-1rа для части вне общей.
SEQ ID NO:6 представляет DNA последовательность, кодирующую три аминокислоты icIL-1raI, для части вне общей.
SEQ ID NO:7 представляет три аминокислоты icIL-1raI для части вне общей.
SEQ ID NO:8 представляет DNA последовательность, кодирующую icIL-1raII, для части вне общей.
SEQ ID NO: 9 представляет аминокислотную последовательность icIL-1raII для части вне общей.
SEQ ID NO:10 представляет DNA последовательность IL-1ra, для части вне общей. Что касается вопросов, относящихся к программе "Patеntin ЕРО", для получения последовательностей G нуклеотид присоединяют в первом положении последовательности для того, чтобы обеспечить кодирование первой аминокислоты Glu и с тем, чтобы избежать образования терминирующего кодона на внутренней стороне последовательности.
SEQ ID NO:11 представляет аминокислотную последовательность IL-1rа для общей части.
SEQ ID NO:12 представляет последовательность из 21 аминокислоты, которая представляет фрагмент icIL-1raII не общий с другими IL-1ra.
SEQ ID NO: 13 представляет DNA последовательность, кодирующую полный icIL-1raII.
SEQ ID NO: 14 представляет аминокислотную последовательность полного icIL-1raII.
ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Этот новый антагонист IL-1 получен путем инсерции в рамку DNA, кодирующей icIL-1rа, новой последовательности из 63 пар оснований (bр) между первым icIL-1ra специфичным экзоном и внутренним акцепторным участком первого экзона sIL-1ra.
Путем RT-PCR экспериментов авторы этого изобретения обнаружили, что этот новый транскрипт экспрессируется в активированных моноцитах и фибробластах и в полиморфонуклеарных клетках (ПМН).
Экспрессия в клетках COS показала, что этот новый антагонист является в наибольшей степени внутриклеточным и имеет молекулярную массу (MW) приблизительно 25 кД по SDS-PAGE.
Новый рекомбинантный антагонист обнаруживает способность ингибировать IL-1.
В настоящей заявке, для ясности и простоты, известный в настоящее время icIL-1ra обозначен как icIl-1ra типа I (icIL-1raI), в то время как новый антагонист, описанный здесь и являющийся предметом настоящего изобретения, определен как icIL-1ra типа II (icIL-1raII).
Примерами патологий, при которых новый антагонист в соответствии с настоящим изобретением может быть с успехом использован для профилактики, терапии или диагностики, являются ревматоидный артрит, септический шок, острый миеломоноцитарный лейкоз, иммунологическая реакция трансплантат против хозяина, синдром приобретенного иммунодефицита (СПИД), язвенный колит и все аутоиммунные заболевания вообще.
Воплощением изобретения является введение фармакологически активного количества icIL-1raII людям, имеющим высокий риск развития патологий, требующих ингибирования IL-1. или людям, у которых уже проявляются патологии подобные сепсису.
Примером вышеуказанной категории являются пациенты, ожидающие хирургической операции.
Любой путь введения, совместимый с действующим началом, может быть использован, но особенно предпочтительным является парентеральное введение, так как позволяет получить за короткое время системные эффекты.
По этой причине предпочтительно введение внутривенного болюса непосредственно до, во время или после хирургической операции. Дозу icIL-1raII, которая должна быть введена, подбирают на основе медицинских показаний в соответствии с возрастом, массой и индивидуальной реакцией пациента.
Дозировка может быть между 0,05 и 30 мг/кг массы тела, а предпочтительная дозировка - между 0,1 и 10 мг/кг массы тела.
Фармацевтическая композиция для парентерального применения может быть приготовлена в форме для вливания, содержащей действующее начало и подходящий носитель. Носители для парентерального введения хорошо известны на практике и содержат, например, воду, солевой раствор, раствор Рингера и декстрозу.
Носитель может содержать меньшие количества наполнителей для того, чтобы обеспечить стабильность и изотоничность раствора.
Приготовление указанных растворов может быть осуществлено в соответствии с обычными приемами, а предпочтительное содержание icIL-1raII должно быть между 1 мг/мл и 10 мг/мл.
Кроме того, примерами патологий, при которых новый антагонист в соответствии с настоящим изобретением может быть с успехом применен в целях профилактики, терапии и диагностики, являются ревматоидный артрит, септический шок, острый миеломоноцитарный лейкоз, иммунологическая реакция трансплантат против хозяина, синдром приобретенного иммунодефицита (СПИД), язвенный колит и все аутоиммунные заболевания вообще.
Настоящее изобретение описано со ссылкой на специфические его воплощения, но содержание описания охватывает все модификации и замещения, которые могут быть произведены специалистом в этой области, не выходя за рамки содержания и целей пунктов формулы изобретения.
В следующей части будут описаны некоторые способы осуществления изобретения, хотя могут быть использованы и эквивалентные материалы и способы. Поэтому следующие примеры являются чисто иллюстративными и не ограничивающими изобретения.
ПРИМЕР 1
Клонирование и характеризация icIL-1raII
МАТЕРИАЛЫ И СПОСОБЫ
Реагенты
Для культивирования и отделения клеток используют следующие коммерчески доступные реагенты: апирогенный солевой раствор и дистиллированную воду для клинического применения; среду RPMI 1640; среду DMEM; среду M199; L-глутамин; Percoll, Ficoll-Hipaque; взятую в асептических условиях сыворотку телячьих эмбрионов; ростовую добавку эндотелиальных клеток (ECGS), полученную из бычьего мозга; гепарин.
Все реагенты содержат менее 0,125 EU/мл эндотоксина, что проверено способом анализа с использованием лизата амебоцитов Limulus.
Клетки
ПМН и моноциты человека, циркулирующие в кровеносной системе, выделяют из периферической крови здоровых доноров центрифугированием в ступенчатом (46% для моноцитов и 62% для ПМН) градиенте изоосмотического (285 mOsm) Percoll, как описано Colotta F., Peri G., Villa SA., Mantovani A., Rapid killing of actinimycin D treated tumour cells by human mononuclear cells. J. Immunol. 132:936, 1984. Клетки выделяют на границе раздела, дважды промывают в солевом растворе и ресуспендируют в среде.
Процент выделяемых ПМН и моноцитов превышает 90%, а чистота - 98%, что определяют морфологическим изучением окрашенных центрифугированных в цитоцентрифуге клеток. Для ПМН и моноцитов обычно применяют среду для культивирования клеток RPMI 1640 с 2 мМ L-глутамина и 10% FCS.
Эндотелиальные клетки человека (ЕС) получают из пупочных вен и культивируют, как подробно описано в литературе (Allavena P., Paganin С., Martin-Padura I., Peri G., Gaboli M., Dejana E., Marchisio P.C., Mantovani A., Molecules and structures involved in the adgesion of natural killer cells to vascular endotelium, J. Exp. Med.. 173:439, 1991).
Обычно применяют конфлюэнтные клетки 2-го-5-го пассажей, поддерживаемые в среде M199 с 10% FCS с добавлением ECGS (50 мкг/мл) и гепарина (100 мкг/мл).
Клетки COS культивируют в среде DMEM с 10% FCS, а клетки фибробластов 8387 - в среде RPMI 1640 с 10% FCS.
После соответствующей обработки клетки исследуют на mRNA IL-1ra или белок IL-1ra. как описано ниже.
RT-PCR
Тотальную RNA экстрагируют способом с использованием изотиоцианата гуанидина с незначительными модификациями.
RT-PCR проводят как описано Colotta F., Polеntarutti N., Sironi M., Mantovani A., J. Biol. Chem,. 267:18278, 1992.
Коротко говоря, проводят обратную транкрипцию 1 мкг тотальной RNA в обратно транскриптазном буфере (5 мМ MgCl2, 50 мМ КСl, 10 мМ трис-HCl; рН 8,3) с 2,5 мМ случайных гексамеров, 1 мМ каждого дезоксинуклеотидтрифосфата, 1 ед/мл ингибитора RNазы и 2,5 ед/мл транскриптазы вируса лейкоза мышей moloney (Perkin Elmer Cetus, Norwalk, CT).
Образцы инкубируют в течение 10 мин при 25oС, а затем в течение 45 мин при 42oС. Потом в реакцию cDNA добавляют специфичную пару праймеров, разработанных для амплификации cDNAs, кодирующих icIL-1raI или icIL-1raII, а также В-актин человека в качестве внутреннего контроля.
Амплификацию проводят в 2 мМ MgCl2, 50 мМ КСl, 0,2 M каждого дезоксинуклеотидтрифосфата, 2,5 единицы/100 мл полимеразы Taq (Perkin Elmer Cetus) и 4 мг/мл каждого специфичного праймера (см. ниже). Амплификацию (30 циклов) ведут в автоматизированном термальном циклере (Perkin Elmer Cetus) при 95oС, при 55oС и при 75oС по 1,5 мин для каждого условия.
Амплифицированные продукты проводят через агарозный гель, окрашенный 1% бромидом этидия вместе с стандартами молекулярной массы (Boehringer Mannheim, Mannheim, Germany).
Олигонуклеотиды синтезируют фосфороамидитным способом. Последовательности нуклеотидов, применяемые для селективной амплификации icIL-1ra идентичны описанным Haskill S. et al., Nati Acad., USA, 88:3681, 1991.
В частности, авторы используют олигонуклеотиды GM397 (описаны здесь как IRA 1) и GM368 (IRA 4).
Для амплификации icIL-1raII авторы используют IRA 4 и IRA 5 (SEQ ID NO: l), которые специфично распознают дополнительный экзон, который, как описано здесь, входит в последовательность icIL-1raII.
Для амплификации В-актина описан "прямой" олигонуклеотид в SEQ ID NO:2, соответствующий нуклеотидам 60-79 cDNA В-актина.
"Обратный" олигонуклеотид, комплементарный нуклеотидам 430-449, представлен SEQ ID NO: 3. Продукты амплификации субклонируют (ТА Cloning System, Invitrogen, San Diego, CA) и секвенируют способом терминации дидезоксицепи.
Экспрессия продуктов IcIL-1ra в клетках COS
cDNAs, содержащие 32 пары оснований 5'-нетранслируемого участка, полную открытую рамку считывания и 6 пар оснований (включающих стоп-кодон) 3'- нетранслируемого участка как icIL-1raI, так и icIL-1raII, получают RT-PCR с олигонуклеотидами IRA 4 и IRA 5, как подробно описано выше, и затем лигируют в экспрессирующий вектор pSF5. Достоверность обратной транскрипции и амплификации проверяют секвенированием.
Плазмиды, содержащие cDNA в корректной ориентации, очищают в градиенте CsCl, а затем трансфицируют ими клетки COS способом осаждения кальция, как описано Sambrook J. et al.. Cold Spring Harbor Laboratory Press, 1989.
Через два дня супернатанты культуры и лизаты обработанных ультразвуком клеток исследуют ELISA или иммуноблоттингом, как подробно описано ниже. Пустую (нетрансфицированную) плазмиду используют в качестве контроля.
Идентификация иммунореактивного IL-1ra
Используют коммерческий тест ELISA (Amersham, Buckinamshire, UK.), который идентифицирует как sIL-1ra, так и icIL-1ra. Для вестерн-блоттинга (анализа Western blot) используют поликлональные антисыворотки двух кроликов и одной козы.
Образцы лизатов и супернатанты клеток COS подвергают электрофорезу 12.5% SDS-PAGE, а затем блоттингу на нитроцеллюлозном фильтре (Stratagene, La Jolla, CA. USA).
Инкубацию с первичными и вторичными антителами осуществляют согласно стандартным методикам. Первичные антитела представлены поликлональными антителами кролика против IL-1ra.
Вторичные антитела представлены фракцией козьего иммуноглобулина против кролика, связанной с пероксидазой хрена (Amersham). Бэнды фракции иммунореактивного белка обнаруживают способом на основе хемилюминесценции (ECL Detection, Amersham), осуществляемым согласно рекомендациям производителя.
Индуцируемая IL-1 экспрессия Е-селектина на ЕС
Конфлюэнтные ЕС, культивированные в 96-луночных планшетах (Falcon), инкубируют в течение 30 минут с количеством лизата трансфицированных клеток COS (см. выше), которое соответствует 25 до 100 нг рекомбинантного IL-1ra (icIL-1raI или icIL-1raII), что определяют специфическим анализом ELISA (Amersham).
В качестве контроля параллельно используют равное количество лизата COS, полученного из "обманно" трансфицированных клеток. Затем ЕС обрабатывают в течение 6 часов 0,1-1 нг/мл рекомбинантного IL-1B человека. Детекцию экспрессии Е-селектина осуществляют анализом ELISA на адгезионных ЕС с моноклональным антителом BB1G-E2 против Е-селектина в качестве первичного антитела и антисывороткой кроликов против мышиного Ig, конъюгированной с пероксидазой хрена, в качестве вторичного антитела. O.D. (оптическую плотность) образцов определяют детекцией планшетов на спектрофотометре (Flow) при длине волны 405.
РЕЗУЛЬТАТЫ
Идентификация icIL-1raII
Специфические олигонуклеотидные праймеры разрабатывают (указаны как IRA 1 и IRA 4 на фиг. 1) для получения полной кодирующей последовательности icIL-1ra (Фиг.1) с помощью RT-PCR. Амплифицированные продукты из ПМН человека субклонируют и секвенируют.
Дополнительно к ранее известной последовательности icIL-1ra изобретатель выделяет ряд клонов, последовательности которых идентичны опубликованной кодирующей последовательности icIL-1ra при важном исключении, которое составляет дополнительная последовательность из 63 пар оснований между нуклеотидами 132 и 133 последовательности icIL-1ra. С помощью описанных экзон-интронных связей icIL-1ra дополнительную последовательность вводят между первым безлидерным экзоном icIL-1ra и внутренним акцепторным сайтом первого экзона sIL-1ra (фиг.1).
На фиг. 1 представлена рассчитанная аминокислотная последовательность. Новый белок (далее называемый icIL-1ra типа II) имеет первые три аминокислоты на NH2-конце как у классического icIL-1ra (icIL-1ra типа I), за которыми следует новая последовательность из 21 аминокислоты. Остальные части у двух белков идентичны.
Любопытно, что связывание с внутренним акцепторным сайтом первого экзона sIL-1ra всегда дает как у sIL-1ra, так и у icIL-1raI и icIL-1raII, остаток одной и той же аминокислоты, например глутаминовой кислоты (фиг.1).
Наиболее удивительной характеристикой вставленной аминокислотной дополнительной последовательности является наличие семи остатков глицина, шесть из которых являются последовательными. Остатки глицина фланкированы с обеих сторон остатками глутаминовой кислоты. icIL-lraII состоит из 180 аминокислот.
В целом гидрофильный образец icIL-1raII близок к таковому icIL-1raI, но не имеет гидрофобного лидерного пептида на NH2-конце.
Экспрессия icIL-raII
Для идентификации транскриптов icIL-1raII проводят анализ RT-PCR с парой специфически созданных олигонуклеотидов (IRA 5 и IRA 4, фиг.1) с ожидаемым амплифицированным продуктом из 33 пар оснований.
Как показано на фиг.2, транскрипты, кодирующие icIL-1raII, определяют в РМА-, IL-1- и TNF-активированных фибробластах. Слабый, но поддающийся определению бэнд отмечен в моноцитах, обработанных LPS.
ПМН, как обработанные, так и необработанные (фиг.2), также имеют очень слабый бэнд ожидаемого размера.
Специфичность амплифицированных продуктов, показанная на Фиг.2, подтверждают субклонированием и секвенированием.
Экспрессия рекомбинантного icIL-1raII
Клетки COS трансфицируют последовательностью DNA, кодирующей icIL-1raII, и, для сравнения, последовательностью, кодирующей icIL-1raI. Затем клеточные лизаты и супернатанты исследуют вестрен-блоттингом.
Поликлональные антисыворотки, используемые в этих экспериментах, одинаково хорошо распознают icIL-1raII и icIL-1raI (фиг.3). Большинство, если не все, icIL-1raII и icIL-1raI обнаруживают в клеточных лизатах.
Рекомбинантный icIL-1raI мигрирует как предоминантный бэнд 22 кД, тогда как icIL-1raII имеет массу приблизительно 25 кД.
Ингибированне активности EL-1B рекомбинантным icIL-1raII
Рекомбинантный icIL-1raII исследуют на активность ингибирования IL-1. Для этой цели авторы выбирают индуцированную IL-1 экспрессию Е-селектина на эндотелиальных клетках, поскольку этот анализ является чувствительным (определяемая индукция 100 пг/мл IL-1 или менее) и быстрым (6 часов инкубации с IL-1).
Лизаты "обманно" трансфицированных клеток COS не приводят к значительному снижению активности IL-1.
icIL-1raII не обладает агонистической активностью.
Как показано на фиг.4, рекомбинантный icIL-1raII ингибирует доза-зависимым образом активность IL-1.
Эти данные доказывают, что icIL-1raII действительно является ингибитором IL-1.
Очистка белка
Очистка белка по изобретению осуществляется любым из методов, известных для этой цели, то есть при помощи любой общепринятой процедуры, включающей экстракцию, осаждение, хроматографию, электрофорез, или подобные методы. Дальнейшая процедура очистки, которая может использоваться для предпочтительной очистки белка по изобретению - афинная хроматография с использованием моноклональных антител, которые связывают целевой белок и которые получены и иммобилизованы на гелевой матрице, находящейся в колонке. Неочищенные препараты, содержащие рекомбинантный белок, пропускают через колонку. Белок будет связываться в колонке специфическим антителом, в то время как примеси будут проходить через колонку. После промывки белок элюируется из геля за счет изменения рН или ионной силы.
Фармацевтические композиции
Лекарственное средство предпочтительно представлено в форме фармацевтической композиции, включающей белок по изобретению вместе с одним или более фармацевтически приемлемыми носителями и/или наполнителями. Такие фармацевтические композиции формируют так же, как дальнейший аспект настоящего изобретения. Белок изобретения должен вводится в эффективном количестве.
"Эффективное количество" относится к количеству активных ингредиентов, который является достаточным действием для курса лечения и тяжести заболеваний, описанных выше, проводя к снижению тяжести или ремиссии такой патологии. Эффективное количество будет зависеть от пути введения и состояния пациента.
"Фармацевтически приемлемый" понимается как охватывающий любой носитель, который не влияет на эффективность биологического действия активного ингредиента, и который не является токсичным по отношению к объекту, которому он вводится. Например, для парентерального введения, вышеупомянутые активные ингредиенты могут быть приготовлены в виде единичной дозировочной формы для инъекции в таких носителях, как солевой раствор, растворы декстрозы, сывороточного альбумина и раствор Рингера.
Помимо фармацевтически приемлемого носителя, композиции по изобретению могут также включать незначительные количества добавок, таких как стабилизаторы, наполнители, буферы и консерванты.
Введение такой фармацевтической композиции может быть внутривенным, внутримышечным или подкожным. Другие пути введения, которые могут обеспечить желательные уровни соответствующих ингредиентов в крови, также входят в объем притязаний настоящего изобретения.
Дозировка вводимого терапевтического соединения определяется в соответствии с клинической практикой и будет варьироваться в зависимости от таких факторов, как возраст пациента, рост, вес, пол, предыдущего лечения и общемедицинского состояния. Обычно, желательно вводить пациенту начальную дозу от приблизительно 50 мкг/кг веса до приблизительно 30 мг/кг веса пациента, ежедневно или несколько (например, 3) раз в неделю, хотя может вводиться и более низкая или более высокая доза. В частности, доза определяется фармакокинетикой клиренса вводимого белка, с использованием стандартных фармакокинетиких правил, известным в данной области. Способы изготовления таких дозировок известны или являются очевидными для специалистов, см., например, Gennaro. A, R, Ed. , Remington's pharmaceutical sciences, 18th ed.. Mack, Easton Pa. (1990), то есть концентрация активного вещества настоящего изобретения в терапевтических составах не критична, хотя обычно варьирует от приблизительно 1 мкг/мл до приблизительно 20 мг/мл.
ОБСУЖДЕНИЕ
Изобретатели описывают новую молекулярную форму icIL-1rа. Новую молекулу получают инсерцией 63 пар оснований между первым безлидерным экзоном icIL-1rа и внутренним акцепторным сайтом первого экзона sIL-1rа.
Поскольку полученный белок частично идентичен классическому icIL-1ra, за исключением дополнительной последовательности из 21 аминокислоты, расположенной на NH2-конце молекулы, изобретатели предполагают назвать эту новую форму icIL-1ra типа II, считая классическую последовательность icIL-1ra как icIL-1rа типа I.
Эксперименты с использованием RT-PCR показывают, что транскрипты icIL-1raII индуцируются в моноцитах и фибробластах. Рекомбинантный icIL-1raII, экспрессирующийся в клетках COS, имеет среднюю мол. массу приблизительно 25 кД и активность ингибирования IL-1, сравнимую с активностью, проявляемой icIL-1raI при экспрессии в тех же экспериментальных условиях.
Транскрипты, кодирующие icIL-1rа и sIL-1ra, получают с одного и того же гена, используя различный сплайсинг. icIL-1ra получают с помощью альтернативного начала транскрипции экзона, введенного во внутренний акцепторный сайт первого экзона, содержащего лидерную последовательность sIL-1rа.
Результаты, полученные изобретателями, предлагают новую организацию гена IL-1rа, при которой дополнительный экзон расположен между первым экзоном классического icIL-1ra и sIL-1ra, соответственно. При использовании этого нового экзона получают полипептиднуто молекулу, которая при отсутствии сигнального пептида отличается по N-концу от icIL-1raI инсерцией 21 аминокислоты, сохраняя при этом способность ингибировать IL-1.
Использование альтернативного сплайсинга для создания различных молекул IL-1ra представляется регулируемым в высокой степени. Транскрипты icIL-1raII индуцируют с помощью IL-1, TNF и форболовых эфиров в фибробластах и с помощью LPS - в моноцитах. Обнаружено, что в фибробластах форболовые эфиры селективно индуцируют транскрипты icIL-1ra, тогда как IL-1 и TNF индуцируют mRNAs как sIL-1ra, так и icIL-1ra. В моноцитах IL-13, увеличивающий транскрипцию как sIL-1rа, так и icIL-1raI, не индуцирует icIL-1raII.
Наконец, ПМН, в которых sIL-1ra и icIL-1ra конститутивно экспрессируются и индуцируются, экспрссируют очень мало транскриптов, как установлено RT-PCR. Более того, эти данные показывают, что механизмы, индуцирующие с помощью различного сплайсинга образование трех форм IL-1ra, регулируются различным образом в ответ на внешние сигналы.
Аминокислотная последовательность описанной здесь дополнительной последовательности удивительна тем, что она содержит семь остатков глицина, шесть из которых расположены последовательно.
Богатые глицином последовательности присутствуют в молекулах с различными биологическими активностями, включая предсердный рецептор натрийуретического клиренса, гомеобокс гена НОХ11, промежуточные нити кератинов и ядерные белки, участвующие в связывании цетромера или сплайсинге RNA.
Однако, за исключением остатков глицина, очевидная гомология аминокислотных последовательностей, фланкирующих богатые глицином участки, не обнаружена между этими белками и icIL-1raII.
Система IL-1 демонстрирует необычайный уровень сложности, представленный двумя агонистами, двумя рецепторами, один из которых является ингибитором IL-1, и антагонистом рецептора, для которого могут существовать, по меньшей мере, три различные молекулярные формы, если принять во внимание полученные результаты.
Хотя биологическое значение внутриклеточных форм IL-1rа требует четкого установления, приведенные здесь данные указывают, что альтернативным сплайсингом можно получить две различные формы icIL-1ra с различными N-концами в ответ на выбранные внешние стимулы.
Наличие множественных и сложных уровней контроля IL-1 указывает на абсолютное требование жесткого физиологического контроля воспалительного потенциала этого цитокина.
ОПИСАНИЕ ФИГУР
Фигура 1
Последовательность DNA и рассчитанная последовательность белка icIL-1raII в сравнении с классическим icIL-1ra (icIL-1raI) и sIL-1ra.
В верхней части фиг. 1 показаны последовательности DNA и белка, специфично представленные в sIL-1ra, icIL-1raI и icIL-1raII. В нижней части фиг. 1 показана последовательность, общая для трех форм IL-1rа.
Полные последовательности каждой молекулы таким образом получают связыванием каждой специфичной части с общей последовательностью. Для ясности, последовательность DNA icIL-1ra начинается с нуклеотида 91 опубликованной 5'-нетранслируемой последовательности, и описаны только 6 пар оснований 3'-нетранслируемой последовательности.
Общая последовательность IL-1ra начинается с внутреннего акцепторного сайта, расположенного в первом экзоне sIL-1ra, соответствующем нуклеотиду 133 полной последовательности icIL-lraI и нуклеотиду 88 полной последовательности sIL-1rа.
Стрелки указывают прямой (IRA 1 и IRA 5) и обратный (IRA 4) олигонуклеотиды, используемые для анализа RT-PCR, как описано в тексте. Олигонуклеотид IRA 5 распознает только DNA icIL-1raII.
Фигура 2
Анализ RT-PCR экспрессии icIL-1raII в клетках различных типов
Проводят обратную транскрипцию RNAs фибробластов 8387 (панель А), моноцитов (В) и ПМН (С). Продукты каждой реакции синтеза DNA затем разделяют на два образца, один из которых амплифицируют с олигонуклеотидами IRA 5 (прямой) и IRA 4 (обратный) с целью детекции транскриптов icIL-1raII, а другой амплифицируют с В-актин-специфичными олигонуклеотидами (см. Раздел Материалы и Способы).
Затем амплифицированные продукты исследуют в агарозном геле, окрашенном бромидом этидия. Амплифицированные продукты, соответствующие В-актину, приведены слева от стандарта, а амплифицированные продукты, соответствующие icIL-1raII (справа), указаны стрелкой. Специфичность этих бэндов подтверждают субклонированием и секвенированием.
Фигура 3
Вестрен-блоттинг рекомбинантного icIL-1raII
Клеточные лизаты из клеток COS, трансфицированных DNAs, которые кодируют icIL-1raI (2) или icIL-1raII (3) или "пустым" вектором, который не содержит такой DNA (1), исследуют иммуноблоттингом с поликлональными антителами кролика против icIL-1rа. Стандарты молекулярной массы указаны.
Фигура 4
Эффекты icIL-1raII на индуцированную DL-1 экспрессию Е-селектина на эндотелиальных клетках
Эндотелиальные клетки обрабатывают 0,1 или 1 нг/мл IL-1B человека с или без 25-100 нг/мл icIL-1raII или эквивалентными количествами лизатов клеток COS, полученных из клеток, которые "обманно" трансфицированы с помощью пустого вектора, как подробно описано в разделе Материалы и Способы.
Через 6 часов инкубации эндотелиальные клетки исследуют на экспрессию Е-селектина тестом ELISA, который проводят на адгезионных клетках.
Приведенные данные являются процентами индуцированной IL-1 экспрессии Е-селектина относительно контроля.















Изобретение относится к области молекулярной биологии и генной инженерии и может быть использовано в медицине. Охарактеризована последовательность ДНК, кодирующая новый антагонист рецептора интерлейкина-1 (icIL-1raII). Путем встраивания этой последовательности в векторную плазмиду pSF5 и трансформации сконструированным экспрессирующим вектором COS-клеток получена экспрессия рекомбинантного полипептида с мол.массой около 25 кД, который обладает ингибирующей активностью в отношении IL-Ia и IL-1В, сопоставимой с активностью внутриклеточной формы антагониста рецептора интерлейкина -1 icIL-1ra. Описано применение нового антагониста рецептора IL-1 в качестве активного начала в составе фармкомпозиции, используемой при патологиях, связанных с продуцированием IL-1. 6 с. и 2 з.п.ф-лы, 4 ил.
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Огнетушитель | 0 |
|
SU91A1 |
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| S.HASKILL et al | |||
| Proc | |||
| Nat.Acad.Sci | |||
| USA, v.88, 1991, p.3681-3685 | |||
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| N.F.SANDER et al | |||
| Proc | |||
| Nat.Acad.Sci | |||
| USA, v.85, 1988, p.2929-2933 | |||
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |
| С.Н | |||
| ЩЕЛКУНОВ | |||
| Конструирование гибридных молекул ДНК | |||
| - Новосибирск: Наука, 1987, с.164. | |||

