Изобретение касается очищенного в значительной степени гелонина,
Основным требованием при создании лекарственного препарата для лечения любого заболевания является специфичность и эффективность действия. Различные имеющиеся лекарства для лечения рака не отвечают этим требованиям. Концепция направленной доставки токсических лекарственных препаратов к определенным опухолям стала предметом интенсивного изучения в последние несколько лет /Торпе /1985/ Biol. Clin. Applications 84: 475-512; под ред. Моллера /1982/ Immun. Rev. 62: 1-215/. Недавно, как моноклональные, так и поликлональные антитела, лектины, лимфокины и гормоны, опознающие специфические детерминанты на поверхности опухолевых клеток, стали применяться в качестве переносчиков для доставки токсических агентов в клетки, где последние могут проявлять свои цитотоксические свойства
/Блаттлер и др. /1985/ Biochemistry 24: 1517-1524;
Франкел и др. /1985/ I. Biol. Res. Modif. 4: 437-446;
Рейманн и др. /1988/ I. Clin. Invest. 82: 129-138;
Шварц и Васе /1988/ Endocrinology 122: 1695-1700;
Скотт и др. /1987/ I. Natl. Cancer Inst. 79: 1163-1172;
Сингх и др. /1989/ Biol. Chem. 264: 3089-3095;
Сринивасан и др. /1985/ FEBS Letters 192: 113;
Шварц и др. /1987/ Endocrinology 121: 1454-1460/.
Таким образом, продолжается изучение токсических частей молекул с агентами-переносчиками, включая радионуклиды /Чозе и др. /1967/ Br. Med. I 1: 90-96/, цитотоксические препараты, обычно используемые в хемиотерапии рака /Торп и Росс /1982/ Immun. Rev. 62: 119-157; Девегер и др. /1982/ Immun. Rev. 62: 29-45; Арнон и Села /1982/ Immun. Rev. 62: 5-27; Пимм и др. /1982/ Cancer Immun. Immunotherap. 12: 125-134; Ровланд и Акстон /1985/ Cancer Immun. Immunotherap. 19: 1-7/ и белки, выделенные из бактерий и растений, такие как дифтерия или рицин /Джансен и др. /1982/ Immun. Rev. 62: 185-216; Разо /1982/ Immun. Rev. 62: 93-117; Витета и др. /1982/ Immun. Rev. 62: 159-183; Нелвилле и Йоуле /1982/ Immun. Rev. 62: 47-73; Торпе и др. /1981/ Eur. I. Biochem. 116: 447-454/. Специфичная молекула конструируется путем замещения неспецифической В цепи на антителе или гормон.
Бактериальные и растительные токсины, такие как дифтерийный токсин /ДТ/, токсин A Pseudomonas aeruginosa абрин, рицин, токсин омелы белой, модессин и токсин Shigella являются мощными, губительно действующими на клетки агентами в связи с их способностью к нарушению важнейших клеточных функций. Например, ДТ и рицин ингибируют синтез белка в клетке путем инактивации 2-го фактора элогации и инактивации рибосомальных 60S субъединиц соответственно. /"Бактериальные токсины и клеточные мембраны" под ред. Ielajaszewicz & Wadstrom /1978/ Academic Press. с.291/. Это чрезвычайно сильные токсины, так как являются ферментами, действующими в большей степени каталитически, чем стехиометрически. Молекулы этих токсинов состоят из ферментативно активной цепи или фрагментов, обычно называемой "A" цепью или фрагментом, соединенной с одной или более полипептидными цепями или фрагментами, обычно называемыми В цепями или фрагментами, которые прикрепляют молекулу к клеточной поверхности и позволяют "A" цепи достичь своего места действия, т.е. цитозоля, и выполнить свою разрушающую функцию. Процесс поступления в цитозоль называется либо "интернализация", либо "интоксикация", либо "транслокация". Эти белковые токсины относятся к классу несущих две цепи, последние называются "A" и "B" цепями. "В" цепь обладает способностью связываться почти со всеми клетками, в то время как "A" цепь обладает цитотоксической активностью. Полагают, что "A" цепь должна вовремя отделяться от "В" цепи - обычно путем разрыва дисульфидной связи - для обеспечения функционирования "A" цепи. Вообще, эти природные токсины не обладают избирательностью к данному типу клеток или тканей, т. к. их "В" цепь узнает и связывается с рецепторами, имеющимися у различных клеток. Доступно ограниченное число токсинов, которые, сами по себе не обладают цитотоксичностью по отношению к различным клеткам. Применение определенных, встречающихся в природе одноцепочных молекул токсинов, которые сами не связываются с рецепторами, расположенными на поверхности клетки, и поэтому в норме не попадают в клетки, обеспечивает токсическими молекулами, которые сами по себе, относительно не токсичны для большинства, если не для всех клеток. Такие природные одноцепочечные токсины, известные к настоящему времени, включают /но не сводятся к/ антивирусный белок локоноса /Рамакришнан и Хоустон /1984/ Cancer Res. 44: 210-208/, сапонин / Торпе и др. /1985/ I. Natl. Cancer Inst. 75: 151-159/, и гелонин /Стирпе и др. /1980/ I. Biol. Chem. 255: 6947-6953/. Эти белки в свободной форме не токсичны для клеток, но могут ингибировать белковый синтез в том случае, если попадут внутрь клетки. Однако, доступность этих одноцепочечных токсинов в значительной степени очищенном виде ограничена из-за того, что их необходимо выделять из растений, в которых они содержаться в относительно низких количествах, и концентрация токсина в растениях зависит от условий роста растений и условий их сбора.
Гелонин - одноцепочечный полипептид, выделяемый из семян растения Gelonium multiforum, его молекулярный вес составляет приблизительно 28000 - 30000 кД. Гелонин является основным гликопротеином с изоэлектрической точкой приблизительно 8.15 и содержит остатки маннозы и глюкозамина /Фаласка и др. /1982/ Biochem I. 207: 505-509/. В противоположность другим растительным и бактериальным токсинам, этот белок сам по себе не токсичен для клеток, но доставленный в клетку через переносчик, он разрушает рибосомальные 60S субъединицы. Биологические данные in vivo и in vitro дают возможность полагать, что действие гелонина равно или превосходит другие растительные токсины. Действительно, результаты сравнения соединений гелонина in vitro и in vivo с соединениями других "A" цепей показывают, что гелонин обладает сходной силой действия, лучшей селективностью, лучшей избирательностью к опухолям и более значительным терапевтическим действием /Сиван и др. /1987/ Cancer Res. 47: 3169-3173/. Однако, наличие воспроизводимого, легко доступного гелонина из природных источников ограничено. К тому же, очистка гелонина из растительного сырья является трудной, а выход очень низок.
Показано, что гелонин, сам по себе, является абортийным средством для мыши и увеличивает зависящую от антител цитотоксичность /Йеунг и др. /1988/ Internatl. I. Peptide Protein Res. 31: 265-268/.
Некоторые исследователи использовали гелонин в качестве цитотоксического агента, химически связанного с моноклональными антителами или пептидными гормонами - высокоспецифичными клеточными лигандами. Однако химическая модификация гелонина и специфических к клеткам лигандов могут уменьшать эффективность и цитотоксическое действие самого гелонина. Более того, количества гелонина из природных источников варьируют в зависимости от условий сбора и роста растений, которые могут влиять на цитотоксическую активность гелонина. Возможность изготовлять синтетический токсин гелонин химически или используя рекомбинантную технологию обеспечивать обильный воспроизводимый источник токсина.
Данное изобретение обеспечивает получение очищенного в значительной степени гелонина, имеющего аминокислотную последовательность, представленную на фиг. 1 и SEQID No:1. Использование последовательности, представленной в настоящей заявке, дает возможность для получения очищенного в значительной степени гелонина методом рекомбинантной технологии, обеспечивающей также огромные количества токсина, которые прежде не могли быть получены из природных источников.
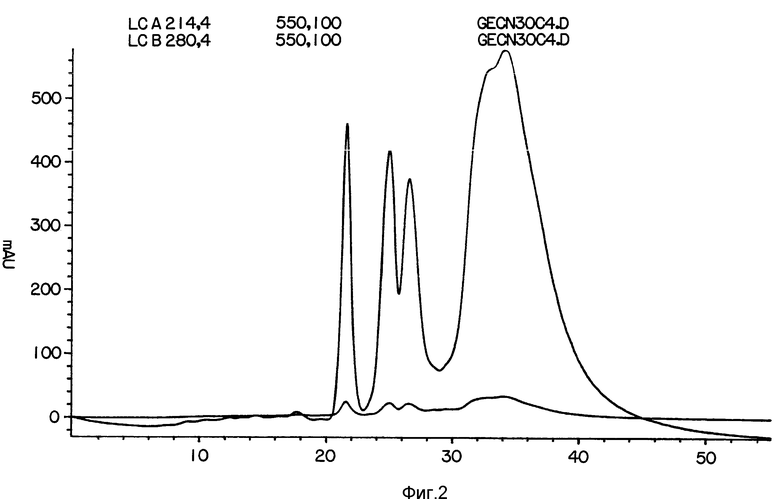
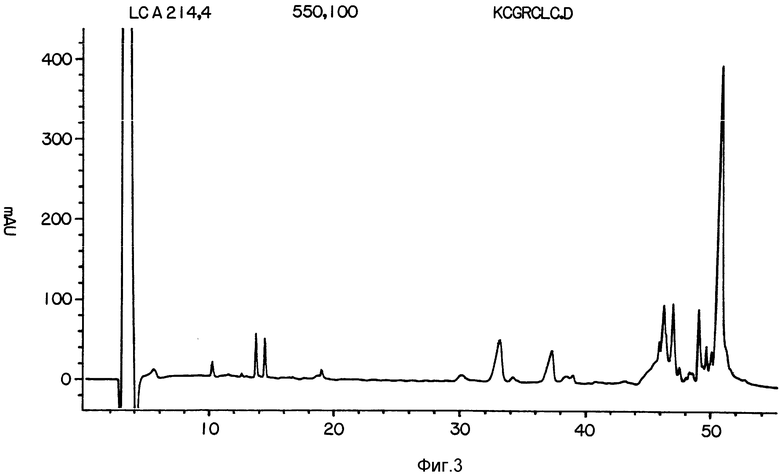
На фиг. 1 представлена аминокислотная последовательность гелонина; на фиг. 2 - HPLC профиль элюции BrCN-фрагментов; на фиг. 3 - HPLC профиль элюции гелонина; на фиг. 4 - протеазы Staphyloccus; на фиг. 5 - фрагментов, полученных при обработке гелонина гидроксиламином; на фиг. 6 - кривая гидрофобности гелонина; на фиг. 7 - трихосантина; на фиг. 8 - абрина; на фиг. 9 - рицина; на фиг. 10 - предшественника агглютинина.
Термин "очищенной в значительной степени" в применении к белку гелонину, описываемому в данной заявке, означает, что полипептид по существу очищен от других растительных белков, обычно связанных с природным гелонином, дает воспроизводимые электрофоретические и хроматографические результаты, профили элюции и токсическую активность. Термин "очищенный в значительной степени" не исключает искусственных или синтетических смесей белка гелонина с другими компонентами.
Гелонин выделяли из семян растения Gelonium multiforum методом, известным специалистам. Аминокислотную последовательность определяли модифицированным методом расщепления по Эдману.
Пробы гелонина вносили в реакционный резервуар с реверсионной фазой и проводили расщепление по Эдману. Было показано, что N-концы гелонина гетерогенны /половина молекул короче других на одну аминокислоту/. Такая гетерогенность затрудняла проведение более 40 циклов расщепления. Поэтому для определения дальнейшей последовательности аминокислот проводили ферментативное расщепление.
Серединные последовательности белков обычно получают перевариванием или расщеплением большой молекулы белка на меньшие куски, сочетая ферментативное и химическое расщепления. Когда природный гелонин инкубировали с различными протеолитическими ферментами, оказалось, что происходит неполное расщепление. Было показано, что это частично объясняется наличием дисульфидной связи в N-концевой части молекулы. В результате расщепления этой связи восстановлением и алкилированием иодуксусной кислотой образовался фрагмент, который был менее растворим, чем исходный материал при pH , необходимом для ферментативного переваривания. Сочетание переваривания нативного гелонина трипсином, лизинаминопептидазой /Lysc/, стафилококковой протеазой /V8/ и химотрипсином привело к получению пептидов, преимущественно C-концевой части молекулы. Это показывает, что N-концевая часть молекулы /из анализа N-конца до Асп-Ала-Про в районе 70 остатка/ не доступна для ферментативного переваривания.
Гелонин расщепляли бромцианом на 3 большие пептида. Аликвоты белка /0.2 мг/мл/ растворяли в 70% муравьиной кислоте. Кристалл бромциана добавляли к раствору и проводили реакцию в течение по крайней мере 18 ч. Затем раствор разводили водой и наносили на маленькую разделительную колонку. После нанесения пробы для элюции белковых фрагментов использовали градиент от 1 до 10% п-пропанола с 0.1% TFA. Профиль элюции представлен на фиг. 2.
При ферментативном переваривании всего белка или фрагментов, полученных обработкой бромцианом, получали перекрывающиеся пептиды. Ферментативное переваривание лизилэдопептидазой проводили в 0.1% SDS, 100 мМ трис, pH 8.0, протеазой из Staphylococcus Aureus в 0.1% SDS или трипсином в 0.1% Твине 20. Гелонин содержит 1 остаток цистеина на 49-ом месте. При восстановлении и карбоксилировании получали белок, который лучше очищался на HPLC с реверсионной фазой и был более чувствителен к ферментативному перевариванию. Поэтому ферментативную обработку проводили в основном в 0.1% SDS или 0.1% Твине. Затем на основе сравнения данных по расщеплению бромцианом и ферментативной обработки установили последовательность 160-ти C-концевых остатков. Оставшуюся неизвестной последовательность между 40-ым и 70-ым остатками определяли, сочетая химическую модификацию цистеина и иодуксусной кислотой и солюбилизацию алкилированного белка с помощью SDS. Гелонин, алкилированный RCM, инкубировали затем с избытком Лиз-с фермента при 37oC в течение коротких периодов времени /1-5 часов/.
Этот метод позволил получить новую последовательность, неполучавшуюся ранее. В этой новой последовательности присутствовала комбинация Асн-Гли. Эту аминокислотную комбинацию расщепляли химически, используя гидроксиламин.
Обработку гидроксиламином проводили, добавляя 100 мкг гелонина к свежеприготовленному гидроксиламину /2М/ в 0.2 М Трисе/ /pH 9.0/, содержащему 2 М NaCl, 1 мМ ЭДТА и 10% этанол. После 7-часовой инкубации при комнатной температуре реакционную смесь наносили на разделительную колонку. Затем колонку промывали 1% раствором TFA в воде и элюировали либо градиентом ацетонитрила, либо прямо разделяли смесь. При такой химической обработки получали большой гидрофобный пептид, содержащий последовательность примерно из 200 аминокислот, соединенных с Асп-Ала-Про при 70-ои остатке. Профиль элюции представлен на фиг. 3.
Оставшийся короткий кусок перекрывающейся последовательности, расположенный между 40 и 50 остатками, определяли перевариванием неалкилированного гелонина Lysc в SDS. Это привело к отделению большей части C-конца молекула. Затем смесь снова переваривали химтрипсином. Продукты такой обработки разделяли HPLC. Сиквенс-анализ последовательности большого пептида показал последовательность Сер-Тре-Лиз, начинающуюся примерно через 5 аминокислот от N-конца молекулы. Это было нужно, т.к. отделяло гетерогенную часть молекулы и давало доступ к длинной последовательности.
Белок гелонин состоит из 258 аминокислот, последовательность которых представлена на фиг.1. Аминокислотную последовательность гелонина сравнивали с другими известными последовательностями, имеющимися в банке сиквенсных данных /Генбанк, PIR, EMBL/, чтобы определить, имеет ли гелонин какие-либо последовательности, гомологичные таковым в других белках. Сравнение аминокислотной последовательности гелонина с другими белками с известными аминокислотными последовательностями показало, что гелонин имеет уникальную последовательность. Была найдена гомология определенных участков последовательности гелонина с участками других белков. Например, установлено 36.0% гомологии гелогина с альфатрихосантином из Trichosanthin Kirilowi, 33.8% гомологии с "A" цепью абрина из Indian Licquorice 35.2% гомологии с предшественником агглютинина из бобов Castor, 33.7% гомологии с рицином D, "A" цепью из бобов Castor и 27.3% гомологии с антивирусным белком /МАР/ из Mirabilis jalapa.
Гидрофобные профили, представленные на фиг. 6 - 10, показывают сходство гидрофобных областей трихосантина, рицина и других белков, ингибирующих рибосомы.
Профиль гидропатии структуры гелонина показывает гидрофобную область в участках 35-80 и 150-180. Эти области, вероятно, имеют прочную складчатую структуру. Сходный с этим гидрофобный участок также наблюдается в других токсинах /см. фиг. 6 - 10/, это дает возможность полагать, что активный центр фермента находится в этих складчатых участках. Следовательно, активный участок фермента не может быть найден в линейной части молекулы, и эти структуры, возможно, должны быть сложены определенным образом для формирования свойственного ему активного центра.
Используя кДНК гелонина можно синтезировать рекомбинантный гелонин. Можно специфично вводить в молекулу мутации чтобы снабдить рекомбинантный гелонин углеводными группами, которые могут изменять направление действия иммуноконъюгатов гелонина. Можно получать рекомбинантные молекулы гелонина с точными мутациями для получения большей токсической активности, чем у нативной молекулы, для более эффективного проникновения в цитозоль после прикрепления к поверхности клетки переносчиком, таким как моноклональное антитело или точно направленный лиганд, например, IL-2, EYF, IFN и т.д., для устойчивости к деградации лизосомами, получая, таким образом, более устойчивые и долго действующие токсины.
Можно также сконструировать рекомбинантные молекулы гелонина слиянием с другими функциональными молекулами, способными к уничтожению клеток, такими как обладающими ферментативной активностью, TNF, IPN-активностью, второй токсической активностью, например, активностью дифтерийного токсина /где названная "вторая активность" действует на иные, чем гелонин, биологические пути/, и таким образом создать "супертоксин" или токсин с мультифункциональным действием.
Белки, полученные слиянием с гелонином, могут быть сконструированы для переноса лекарств, таких как хемиотерапевтические, или изотопов для радиослежения или радиотерапии. Пептиды гелонина могут применяться в качестве абортивных средств, иммуносупрессоров, противораковых и антивирусных препаратов /таких как анти-HIV агент/.
В следующих примерах приводится детальное описание процесса выделения, характеристика и аминокислотная последовательность гелонина. Использовавшиеся экспериментальные методы детально описаны в приведенных ниже примерах. Эти примеры ни в какой степени не исчерпывают возможности данного изобретения.
Пример 1. Очистка и характеристика гелонина.
Гелонин выделяли из семян растения Gelonium в основном следуя ранее описанному методу /Стирпе и др. /1980/ multiforum 255: 6947-6953/. Вкратце: гелонин экстрагировали из семян гомогенизацией в забуференном солевом растворе /pH 7.4/. Супернатант концентрировали после диализа против 5 мМ Na-фосфата /pH 6.5/, далее проводили очистку ионообменной хроматографией, как описано ниже. Чистоту токсина гелонина оценивали по результатам жидкостной хроматографии под высоким давлением /HPLC/ и электрофореза в полиакриламидном геле в присутствии додецилсульфата Na /SDS-ПААГ/. Гелонин мигрировал одной полосой с молекулярным весом около 29-30000 дальтон.
Токсическую активность гелонина определяли так, как описано в примере 2 - по ингибированию белкового синтеза в бесклеточной системе.
Плоды Gelonium multiforum очищали и семена гомогенизировали с восьмью объемами 0.14 М NaCl, содержащего 5 мМ Na-фосфата /pH 7.4/. Гомогенат оставляли на ночь при 4oC при постоянном перемешивании, охлаждали на льду и центрифугировали при 35000g 20 мин. при 0oC. Супернатант отделяли, диализовали против 5 мМ Na-фосфата /pH 6.5/ и концентрировали на фильтре рм10. Пробы наносили на СМ-52 ионообменную колонку /20х15 см/, уравновешенную 5 мМ Na-фосфатом /pH 6,5/. Материал, связавшийся с ионообменником, элюировали 400 мл линейного градиента NaCl - от 0 до 0.3 М - со скоростью 25 мл/час при 4oC. Собирали фракции по 5 мл. Фракции промеряли на спектрофотометре при 280 нм. Гелонин элюировался примерно во фракциях 55-70 и был последним большим пиком элюата. Эти фракции собирали вместе, диализовали против 0.1 М NaCl в 0.1 М Na2HPO4 буфере /pH 7.4/. Затем пробы наносили на колонку с голубой сефарозой /24 2 см/, предварительно уравновешенной 0.1 М Na2HPO4 / 0.1 М NaCl буфером, колонку промывали 3-мя объемами буфера, равными объемами колонки, и элюировали 400 мл линейного градиента соли /от 0.1 М NaCl до 2 М NaCl/. За элюцией связанного материала следили, измеряя собранные с колонки фракции по методу Лоури. Фракции, содержащие единственный пик белка, собирали вместе и диализовали против PBS в течение ночи при 4oC. Гелонин был очищен более, чем до 97% по данным электрофореза в ПААГ, окрашенного серебром. Чистоту и молекулярный вес каждого препарата проверяли методом жидкостной хроматографии под высоким давлением на колонке с TSK 3000 гелем, уравновешенной 50 мМ Na-фосфатным буфером /pH 7.4/ и методом электрофореза в 15% полиакриламидном геле в присутствии додецилсульфата - Na /SDS-ПААГ/. Гелонин двигался единой полосой с молекулярным весом 29-30000 дальтон.
Пример 2. Проба на активность гелонина.
Активность гелонина определяли по ингибированию синтеза белка в бесклеточной системе. Измерение ингибирования синтеза белка в бесклеточной системе проводили последовательно добавляя к 50 мкл лизата ретикулоцитов кролика, размороженного непосредственно перед употреблением, перемешивая после каждой добавки, следующие компоненты: 0.5 мл 0.2 И трис - HCl /pH 7,8/, 8.9 мл этиленгликоля и 0.25 мл 1 М HCl.
20 мкл солево-аминокислотного раствора /SAEM/, состоящего из 0.375 М KCl, 10 мМ Mg /CH3CO2/2, 15 мМ глюкозы, 0,25-10 мМ аминокислот /за исключением лейцина/, 5 мМ АТФ, 1 мМ ГТФ, 50 мМ трис - HCl /pH 7,6/, 10 мкл креатинин-фосфат-креатинин фосфокиназы, 8 мкл /14С/ лейцина / Амершам, 348 м Ci мМ/, добавляли по 1.5 мкл растворов, содержащих различные концентрации гелонина. Пробы инкубировали 60 мин при 30oC. Включение 14C-лейцина определяли в аликвотах раствора путем преципитации синтезированного белка на фильтрах из стекловолокна с промывкой 10% ТХУ и ацетоном, и измеряли радиоактивность в β -счетчике, используя Aquasol сцинтилляционную жидкость. При таком способе определения удельная активность очищенного гелонина составляла 4 109 ед/мг белка. Единица активности гелонина - количество гелонина, на 50% ингибирующее включение 14-C-лейцина в белок в бесклеточной системе.
Пример 3. Определение аминокислотной последовательности гелонина.
Аминокислотную последовательность гелонина определяли методом расщепления по Эдману, используя автоматический аминокислотный анализатор, описанный в Европейской патентной заявке N EP-257735. Большие пептиды и нефрагментированный белок вносили в камеру анализатора с реверсионной фазой. Нежелательные буферные компоненты отмывали избытком воды. Белок или пробу пептида расщепляли по Эдману и экстрагированные ATZ производные аминокислот переводили в PTH-форму, действием 25% TFA в H2O при 65oC. Пробы PTH-производных индентифицировали аналитическим разделением в реверсивной фазе на Np 1090 колонке.
Для дальнейшей расшифровки аминокислотной последовательности белок обрабатывали различными протеолитическими ферментами и химическими агентами, затем пептиды очищали методом жидкостной хроматографии под высоким давлением. Было показано, что гелонин довольно устойчив к действию трипсина /разрыв после остатков дизина и аргинина/ и ацетилтрипсина/ разрывы только после остатков лизина/. Также было показано, что белок устойчив к 5% /W/W/ фермента. Устойчивость гелонина к протеолитическому ферменту трипсину не обусловлена недостатком участков, на которые действует трипсин, т.к. гелонин содержит 21 остаток лизина и 12 остатков аргинина. Полученные результаты указывают на то, что гелонин, по видимому, представляет собой жесткоупакованную молекулу, недоступную протеолитическим ферментам.
В связи с тем, что гелонин устойчив к расщеплению протеолитическими ферментами, исследовались фрагменты, полученные химическим расщеплением.
Пример 4. Расщепление гелонина бромцианом.
Гелонин, приготовленный как описано в примере 1, растворяли в 70% муравьиной кислоте. К раствору добавляли кристалл бромциана. Через, по меньшей мере, 18 ч раствор наносили либо на маленькую колонку /15 см х 5 см/ с реверсионной фазой /I. T. Bacer; 15 см C -1B связанная фаза Cat 11 7191-02/ или аналитическую / 4.6 х 100 мм/ колонку с реверсионной фазой. Элюировали градиентом от 1 до 70% н-пропанола с 1% TFA в воде, получено 5 пиков, показанных на фиг. 6. Каждый из пиков секвенировали и подвергали ферментативному перевариванию для расщепления серединных последовательностей. 1-ый пик сразу секвенировали и получили последовательность 38-ми остатков, начинающуюся с Фен /F/ и оканчивающуюся Глу /Е/. Эта последовательность была подтверждена данными масс-спектроскопии и перевариванием изолированных пептидов. 2-ой пик секвенировали и получили последовательность 47-ми остатков, начинающуюся с Вал /V/, и не прерывающуюся после Ала в 47-ом положении. Секвенирование 3-его пика дало ту же последовательность, что и 2-ой пик. SDS-гели пиков 2 и 3 также, как и переваривание Lys-c пиков 2 т 3 показали, что пик 3 также содержит C-концевой Br CN пептидов. При последующей трипсиновой обработке гелонина был получен пептид, который соединяет эти две Br CN-пептидные последовательности. Последовательность пептида, полученного обработкой трипсином, по результатам секвенирования сказалась следующей: TSGA NG MFSEAVELER . Оба пика 4 и 5 дали N-концевую последовательность GLDT. Эту последовательность переваривали Lys-c 1/8, для получения C-концевых пептидов.
Пример 5. Ферментативное переваривание гелонина, обработанного BrCN.
Пробы с целым белком или Br CN-фрагментом переваривали лизилэндопептидазой /Wako Chemical Dallas, TX/ в 0.1% SDS, 100 мМ трис pH 8,0 или протеазой Staphylococcus aureus в .1% SDS или трипсином /Sigma/ в 0.1% Твине 20. Смеси, полученные в результате переваривания, разделяли HPLC собранные пептиды секвенировали в аминокислотном анализаторе, используя метод расщепления по Эдману с газовой фазой.
Пример 6. Аминокислотная последовательность гелонина.
Общая последовательность, состоящая из 258 аминокислотных остатков была получена анализом BrCN -фрагментов, получаемых, как описано в примере 3. На фиг. 1 представлена аминокислотная последовательность гелонина. Вообще, гелонин состоит примерно из 285 аминокислотных остатков. Последовательность ДНК была получена методом дедукции, исходя из последовательности аминокислот.
Таким образом, для специалистов очевидно, что возможно провести много изменений и модификаций без отхода от существа или области изобретения, как изложено выше.





Использование: в медицине. Сущность изобретения: линейный пептид-гелонин, содержащий 258 аминокислотных остатка, выделен из Gelonium multiforum с использованием экстракции, диализа, ионообменной хроматографии, чистота оценивалась по результатам высокоэффективной жидкостной хроматографии и электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия (SDS-ПААГ), гелонин мигрировал одной полосой с молекулярной массой около 29 - 30000 дальтон. 10 ил.
Пептид-гелонин формулы, приведенной на с.
| Stipne | |||
| J.Biol | |||
| chem | |||
| Способ получения фтористых солей | 1914 |
|
SU1980A1 |
| Falaske et al | |||
| Biochem.j | |||
| Устройство для видения на расстоянии | 1915 |
|
SU1982A1 |
| al | |||
| International j | |||
| Peptide Profeine Res | |||
| Механическая топочная решетка с наклонными частью подвижными, частью неподвижными колосниковыми элементами | 1917 |
|
SU1988A1 |
| Якубке Х.-Д., Ешкайт Х | |||
| Аминокислоты | |||
| Пептиды | |||
| Белки | |||
| - М.: Мир, 1985, с.90 и 366. | |||

