Изобретение относится к новым плазмидам, более конкретно, к челночным векторным плазмидам, способным к репликации как в клетках Escherichia coli, так и в клетках цианобактерий. Плазмиды содержат фрагмент ДНК, включающий ori A - участок плазмиды рBA1, полученный из цианобактерий Anacystis nidulans.
Цианобактерии Anacystis nidulans способны синтезировать органические вещества из воды, углекислого газа и небольшого количества неорганических солей, и, таким образом, способны к аутотрофному тосту. Далее A.nidulans является одноклеточным организмом и образует колонии на агаровой среде. В частности, штамм R2 способен воспринимать чужеродную ДНК и, таким образом, подлежит трансформации с помощью известных методов.
Разработаны различные клонирующие векторы на основе эндогенных плазмид штамма R2 A. nidulans (например, pUH24 и pUH25) (C.A.M.J.J. von der Hondel et al. Proc. Natl. Acad. Sci. USA 77(3); 1570-1574 (1980); C. J. Kuhlemeier et al. , Mol. Gen. Genet. 184:249-254 (1981); L. A. Sherman and P. van de Putte, J. Bacteriol. 150(1):410-413 (1982); S. Gendel et al., J. Bacteriol. 156(1): 148-154 (1983); S. S. Golden and L. A. Sherman, J. Bacteriol. 155(3) : 966-972 (1983); C. J. Kuhlemeier et al., Plasmid 10:156-163 (1983); C. J. Kuhlemeier et al. , Gene 31; 109-116 (1984); D. E. Laudenbach et al., Mol. Gen. Genet. 199: 300-305 (1985); M. Y. Gruber et al., Curr. Microbiol. 15: 265-268 (1987), etc.),
Указанные векторы использовались, например, для введения чужеродных генов и клонирования генов.
Примерами гетерогенных белков, экспрессируемых с использованием штамма R2 могут служить карбоновая ангидраза человека и Iac.lQ репрессорный белок Escherichia coli I. F. D. Price and M.R.Badger, Plant Physiol. 91:505-513 (1989), α- амилаза Bacillus amyloliquefaciens A50 [I.V.Elanskaya and Morzunoba, Mol. Gene Microbiol.Virusol. 0(9):7-1 (1989), инсектицидный белок B. sphaericus 1593M [N. Tandeau de Marsac et al., Mol.Gen.Genet. 209; 396-398 (1987), β- галактозидаза Escherichia coli [D.] Scanlan et al.. Gene 90:43-49 (1990), M. R.Schaefer and S.S.Golden, J.Bacteriol. 171(7):3973-3981 (1989), Mn-супероксиддисмутаза Escherichia coli [M.Y.Gruber et al., Proc.Natl.Acad. Sci. USA 87:2608-2612, cIts репрессорный белок λ- фага [D.Friedberg and J. Seiiffers, Mol. Gen. Genet. 203: 505-510 (1986), дисатураза цианобактерий Synechosystis PCC 6803 штамм (des A) [H.Wada et al.. Nature 347:200-203 (1990)] и т.д.
С другой стороны, проводились исследования с клетками A.nidulans 6301, при использовании эндогенных плазмидных (pBA1) векторов (челночные векторы Escherichia coli), таких, как pBAS 18 [12kb, K.Shinozaki et al., Gene 19: 221-225 (1982)/ и pBAS 5 [14kb., Kazuo Shinozaki "Shokubutsu Idenshi Sosa Gijutsu" (Plant Gene Manipulation Techniques), под редакцией Hikoyuki Yamaguchi, страницы 98-110, опубликовано CMC]. Однако любой из указанных векторов затруднительно использовать при манипулировании генами. Это связано, например, с тем, что размер гена, который желательно встроить в вектор обычно составляет около 10 кб и более, а вектор имеет только небольшое число сайтов, распознаваемых рестрикционными эндонуклеазами, пригодных для клонирования. Кроме того, штамм 6301 с трудом захватывает ДНК по сравнению со штаммов R2, и в результате частота его трансформации ниже, чем у штамма R2. Для повышения частоты трансформации штамма 6301 H.Daniell et al., [Proc.Natl. Acad. Sci. USA 83:2546-2550 (1986)] предложено обрабатывать клетки ЭДТА-лизоцимом, чтобы увеличить их проницаемость и облегчить введение ДНК. Действительно, авторам удалось увеличить частоту трансформации штамма 6301 с помощью указанного способа, однако он чрезвычайно трудоемок и, кроме того, клетки повреждаются при обработке ЭДТА-лизоцимом.
Таким образом, задача изобретения заключается в конструировании плазмид, повышающих частоту трансформации штамма A.nidulans 6301 при упрощении методик генетических манипуляций. Плазмиды сконструированы на основе эндогенной плазмиды pBA1 штамма 6301 путем сокращения (насколько это возможно) составных элементов данной плазмиды, кроме участка ori, и объединения ее с фрагментом ДНК, который содержит ori плазмиды Col/El Escherichia coli (участок оri E). При этом ген верхнего участка (rop) (этот участок также называют rom-участком, но в настоящем описании он обозначен как rop-участок) удаляют, а затем плазмиду лигируют с сайтом множественного клонирования плазмиды pUC18. Ближайшие аналоги изобретения не обнаружены.
Таким образом, согласно изобретению заявлены следующие плазмиды.
Челночная векторная плазмида, способная к репликации в клетках Escherichia coli и Anacystis nidulans, содержит следующие конструктивные элементы:
область начала репликации (оri A) размером 2,8 - 8,0 т.п.о, полученную из плазмиды pBA1, содержащейся в клетках цианобактерий Anacystis nidulans 6301;
сайт множественного клонирования;
область начала репликации (оri E), из которой удален ген, определяющий rop-функцию, полученную из col E1 плазмиды Escherichia coli.
При этом область ori E может дополнительно содержать ген устойчивости к ампициллину.
Челночная векторная плазмида по п. 1, отличающаяся тем, что ori E дополнительно содержит ген устойчивости к ампициллину.
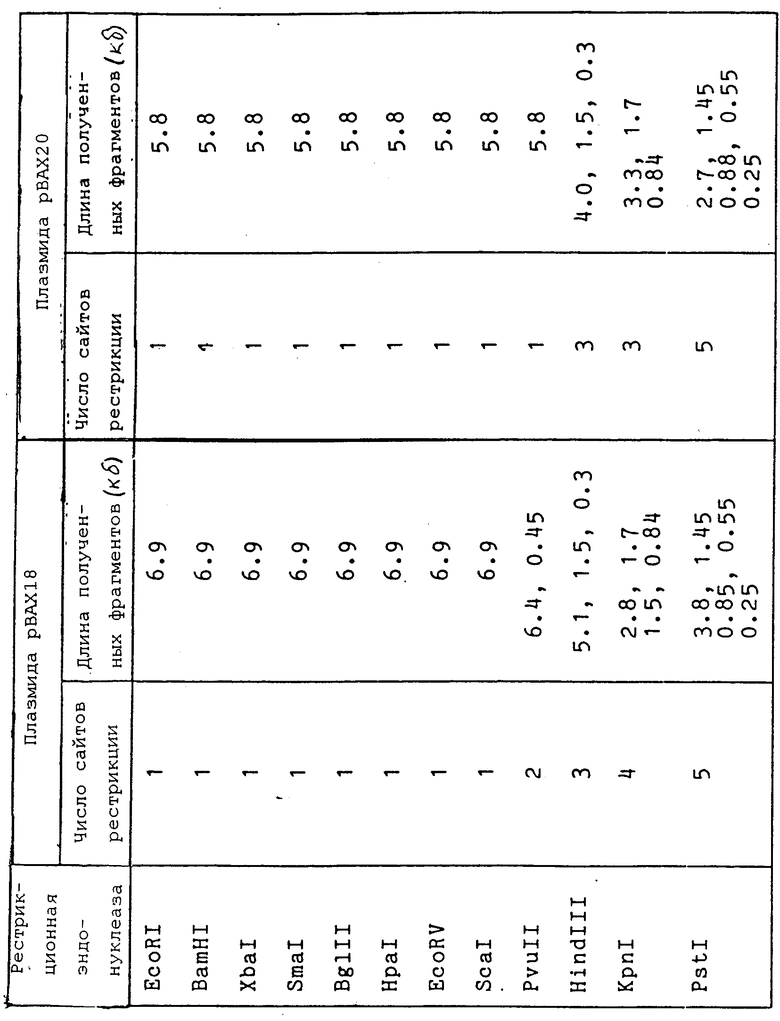
Челночная векторная плазмида pBAX18 размером около 6,9 т.п.о., имеющая рестрикционную карту, приведенную на фиг. 1, содержит следующие конструктивные элементы:
область начала репликации (ori A) размером 2,8 - 8,0 т.п.о., полученную из плазмиды pBA1, содержащейся в клетках цианобактерий Anacystis nidulans 6301;
сайт множественного клонирования;
область начала репликации (ori E), из которой удален ген, определяющий rop-функцию, размером 1,5 - 4,1 т.п.о., полученную из плазмиды pBR322, содержащейся в клетках Escherichia coli;
ген устойчивости к ампициллину в составе ori E.
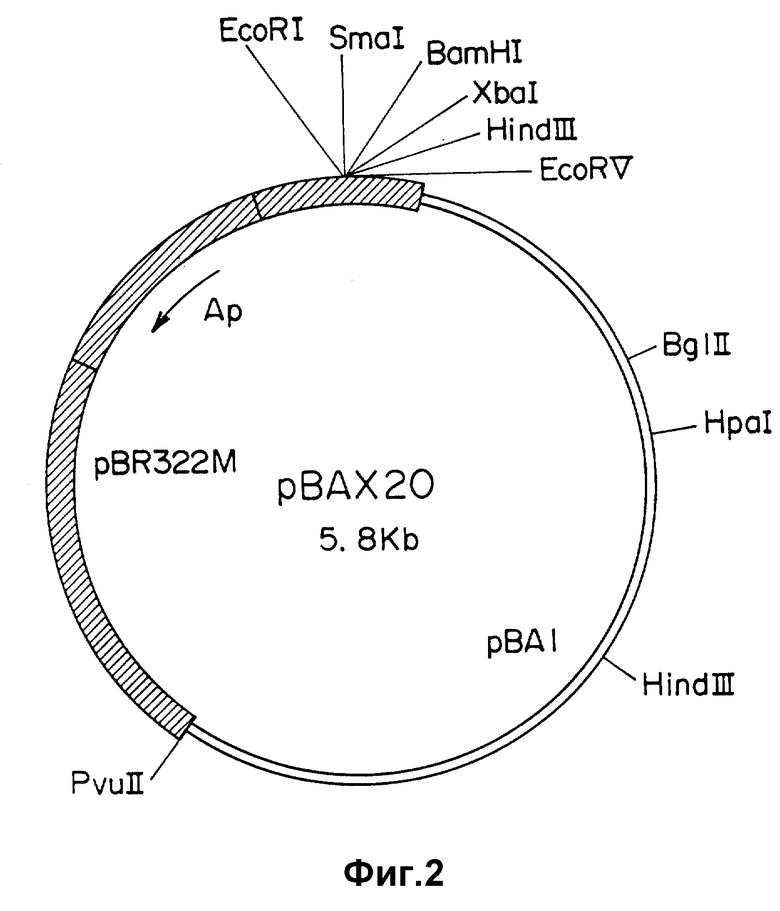
Челночная векторная плазмида pBAX20 размером около 5,8 т.п.о., имеющая рестрикционную карту, приведенную на фиг. 2, содержит следующие конструктивные элементы:
область начала репликации (ori A) размером 2,8-8,0 т.п.о., полученную из плазмиды pBA1, содержащейся в клетках цианобактерий Anacystis nidulans 6301;
сайт множественного клонирования;
область начала репликации (ori E), из которой удален ген, определяющий rop-функцию, размером 1,5-4,1 т.п.о., полученную из плазмиды pBR 322, содержащейся в клетках Escherichia coli;
ген устойчивости к ампициллину в составе ori E.
На фиг. 1 приведена эндонуклеазная карта рестрикции плазмиды pBAX18; на фиг. 2 - эндонуклеазная карта рестрикции плазмиды pBAX20; на фиг. 3 - схема получения плазмид pBAX18 и pBAX20.
Сведения, подтверждающие возможность осуществления изобретения.
Область начала репликации оri A представляет собой генный фрагмент, полученный из плазмиды pBA1 с молекулярной массой (5,04 ± 0,26) • 106 (согласно электронномикроскопическому анализу) или 5,2 • 106 (согласно электрофорезу в агарозном геле) [K.Shinozaki et al., Gene, 9:221-224 (1982)], которая является эндогенной плазмидой цианобактерий Anacystis nidulans 6301 (Synechococcus PPC 6301, АТСС 27144, штамм UTEX 625), принадлежащих к роду Cloococcus (сине-зеленые водоросли). Размер ori A, как правило, находится в пределах от 2,8 до 8,0 т.п.о., предпочтительно от 2,8 до 4,4 т.п.о. Конкретными примерами oriA могут служить фрагмент ДНК, имеющий размер примерно 4,4 т.п.о., полученный путем расщепления плазмиды pBA1 с помощью эндонуклеаз рестрикции XhoI и BamHI и фрагмент ДНК, имеющий размер около 3,3 т.п.о., полученный путем расщепления плазмиды pBA1 с помощью PvuII и BamHI, и т.д.
Сайт множественного клонирования (СМК) представляет собой последовательность (которую также называют полилинкером), имеющую различные сайты эндонуклеаз рестрикции. Примером СМК является Col E1 плазмиды pUS, используемой в качестве клонирующего вектора Escherichia coli, скажем, фрагмент ДНК размером 55 пар оснований, имеющий сайты для следующих эндонуклеаз рестрикции : EcoRI, SacI, KpnI, SmaI, XmaI, BamHI, XbaI, SalI, AccI, HincII, PstI, SphI и HindIII. Кроме того, могут использоваться СМК, полученный из плазмид pUS 12, pUC 13, pUC 19 и т.д., а также из фагового вектора M 13 Escherichia coli. СМК могут быть также синтезированы химическим путем.
Область начала репликации ori E, из которой удален ген, определяющий rop-функцию, получают из col E1 плазмиды E.coli. Примером ori E может служить фрагмент ДНК, выделенный из плазмиды pBR 322 с помощью эндонуклеаз рестрикции PvuII, Ava I, BamHI, Eco 47111, EcoRV, PstI и т.д., размер которого составляет от 1,5 до 4,1 т.п.о., предпочтительно от 1,5 до 2,5 т.п.о. Можно использовать и ori E, содержащие фрагменты ДНК, выделенные из плазмид pUC 18, и т.п.
Кроме того, ori E может содержать ген устойчивости к лекарственным препаратам, например, ген устойчивости к ампициллину (Ampr), который используется в качестве генетического маркера.
Таким образом, плазмиды, заявленные согласно настоящему изобретению, содержат три необходимых элемента, т. е. ori A, CMK и ori E. Кроме того, плазмиды могут содержать фрагменты ДНК, несущие другую генетическую информацию, например фрагмент ДНК, соответствующий гену устойчивости к ампициллину, гену устойчивости к хлорамфениколу и т.д. Типичными признаками таких плазмид являются две плазмиды, обозначенные авторами настоящего изобретения как pBAX18 и pBAX20, которые содержат ori A, CMK и ori E и имеют мол. массу примерно 4,48 мегадальтон (около 6,9 тыс. оснований) и 3,76 мегадальтон (около 5,8 тыс. оснований) соответственно.
Молекулярную массу плазмид определяют электрофорезом в агарозном геле.
Количество сайтов для эндонуклеаз рестрикции в плазмидах pBAX18 и pBAX20 с указанием длины образующихся при расщеплении фрагментов приведено в таблице.
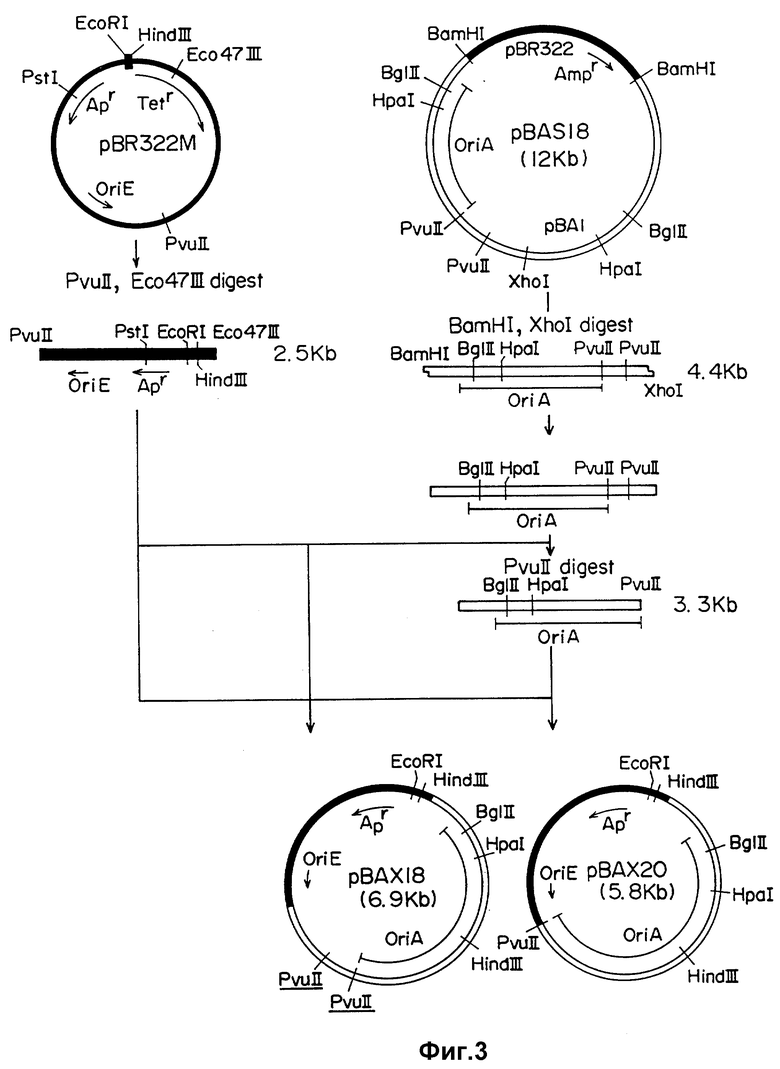
Плазмиды pBAX18 и pBAX20 получают следующим образом.
Ori A - содержащий фрагмент ДНК плазмиды pBA1, выделенной из Anacystic nidulans, получают путем клонирования челночного вектора pBAS 18 в Anacystic nidulans и Escherichia coli [K.Shinozaki et al., Gene 19:221-224 (1982)] известным способом (T. Maniatis et al., Молекулярное клонирование, Лабораторное руководство, опубликовано лабораторией Cold Spring Harbor Laboratory). Далее фрагменты ДНК из клонированной плазмиды pBAS18 получают, как описано T.Maniatis et al. (Молекулярное клонирование) при использовании эндонуклеаз рестрикции BamHI и XhoI или BamHI и PvuII. Указанные ori A - содержащие фрагменты ДНК, имеют размер около 4,1 т.п.о. и 3,3 т.п.о. соответственно. При необходимости выступающий 5'-конец этих фрагментов ДНК затупляют с помощью ДНК-полимеразы T4.
С другой стороны, CMK плазмиды pUC получают путем вырезания его из плазмиды pUC18 с помощью эндонуклеаз рестрикции EcoRI и HindIII.
Далее используют плазмиду pBR 322, типичную Col EI плазмиду в качестве источника фрагмента ДНК, содержащего участок ori E, из которого удален ген, содержащий rop-функцию.
Полученный таким образом CMK объединяют с плазмидой pBR 322, расщепленной эндонуклеазами рестрикции EcoRI и HindIII, и с помощью ДНК-лигазы T4 получают плазмиду pBR 322M, в которую интегрирован CMK вышеуказанный. Затем фрагмент ДНК длиной около 2,5 т.п.о., полученный при расщеплении плазмиды pBR 322M рестрикционными эндонуклеазами Eco 47111 и PvuII и содержащий как ori E, так и CMK участок объединяют с ori A, выделенной, как указано выше, с помощью ДНК-лигазы T4 с получением плазмид pBAX18 и pBAX20.
Рестрикционная карта плазмид pBAX18 и pBAX20 приведена на фиг. 1 и 2, а способ их получения схематически отражен на фиг. 3 и более подробно описан ниже.
Таким образом, заявленные плазмиды представляют собой челночные векторы, имеющие оri E, обеспечивающую их автономную репликацию в клетках Escherichia coli, и оri A, обеспечивающую их автономную репликацию в клетках Anacystis nidulans. Поэтому данные плазмиды можно получить в большом количестве, используя бактерии Escherichia coli, растущие с высокой скоростью. Затем данными плазмидами можно эффективно трансформировать клетки цианобактерий, например Anacystis nidulans.
Далее для вставки чужеродных генов может быть использован CMK, содержащий сайты эндонуклеаз рестрикции EcoRI, SmaI, BamHI, XbaI, ScaI и EcoRV.
Далее размер заявленных плазмид можно уменьшить примерно наполовину (от 6,9 до 5,8 т.п.о) по сравнению с плазмидой pBAS 18 (12 т.п.о.), поскольку участки ДНК, отличные от оri A плазмиды pBAl, из заявленных плазмид удалены. Следовательно, в заявленные плазмиды можно встроить чужеродный ген, имеющий больший размер по сравнению с тем чужеродным геном, который можно ввести в плазмиду pBAS18.
Трансформацию клеток хозяина данными плазмидами можно осуществлять известными способами (D.A.Lightfool et al., J.General Microbiol. 134:1509-1514 (1988)).
При этом частота трансформации Anacystis nidulans 6301 может быть примерно в 1000 раз выше, чем при трансформации данного штамма с помощью плазмиды pBAS18. Поэтому, используя заявленные плазмиды, можно даже при встраивании чужеродного гена легко отобрать клетки, содержащие плазмиду с нужным геном.
В качестве структурных генов, которые могут быть интегрированы в заявленные плазмиды, можно указать структурные гены Anacystis nidulans других цианобактерий, структурные гены бактерий других видов, структурные гены высших животных и растений, и т.д. Кроме того, в качестве структурных генов можно использовать химически синтезированные гены. Примером таких генов является химически синтезированный ген супероксиддисмутазы человека [h-SOD, h-SOD-Ala6 (выложенная открытая заявка Японии N 156884/1990) и другие.
Пример 1.
(1) Ведение сайта множественного клонирования (выделенного из плазмиды pUC18) в плазмиду pBR322.
А) Расщепление EcoRI - Hind III и обработка pBR 322 щелочной фосфатазой.
К 20 мкл (10 мкг) раствора ДНК плазмиды pBR 322 добавляют 40 мкл буфера 5 x Hind III (50 мМ/л трис-HCl (pH 7,5), 35 мМ/л MgCl2, 300 мМ NaCl), 80 единиц (10 мкл) Hind III (Takara Shizo Co., Ltd) и 130 мкл стерилизованной воды, и смесь инкубируют при 37oC в течение 2 ч. После реакции к полученному раствору добавляют 40 мкл 5xEcoRI буфера (500 мМ/л Трис-HCl (pH 7,5), 35 мМ/л MgCl2, 250 мМ/л NaCl, 35 мМ/л 2-меркаптоэтанола, 0,05% бычьего сывороточного альбумина), 120 единиц (10 мкл) EcoRI (Takara Shuzo Co., Ltd) и 50 мкл стерилизованной воды, и смесь подвергают реакции при 37oC в течение 2 ч. После реакции смесь обрабатывают фенолхлороформом и осаждают этанолом, и полученную ДНК собирают и растворяют в 100 мкл 0,1 моль/л трис-HCl (pH 8,0).
К этому раствору добавляют 10 мкл щелочной фосфатазы (Takara Shuzo Co., Ltd) в виде раствора (1 ед/10 мкл щелочной фосфатазы, 10 мМ/л трис-HCl (pH 7,5), 50 мМ/л NaCl, 1 мМ/л ZnSO4), и смесь инкубируют при 37oC в течение 1 ч. После окончания реакции добавляют еще 10 мкл раствора щелочной фосфатазы, и смесь инкубируют при 65oC в течение 30 мин. Полученный раствор обрабатывают фенол-хлороформом, затем осаждают этанолом, и собирают ДНК.
Б) Выделение сайта множественно клонирования (EcoRI-Hind III) из плазмиды pUC18.
Подготавливают две эппендорфовские пробирки, каждая из которых содержит 30 мкл (20 мкг) раствора ДНК pUC 18 с добавлением 20 мкл 10 x K буфера (200 мМ/л трис-HCl (pH 8,5), 100 мМ/л MgCl2, 10 мМ/л DTT, 100 мМ/л KCl), 80 единиц (10 мкл) Hind III и 140 мкл стерилизованной воды, и каждую пробирку инкубируют при 37oC в течение 3 ч. После реакции смесь обрабатывают фенолхлороформом и осаждают этанолом, собирают ДНК и растворяют ее в 112,5 мкл стерилизованной воды. К каждому из полученных растворов добавляют 30 мкл 5 • EcoRI буфера и 190 единиц (7,5 мкл) EcoRI и полученную смесь инкубируют при 37oC в течение 3 ч. ДНК осаждают этанолом, собирают и подвергают электрофорезу в 1,5%-ном агарозном геле для выделения нужного фрагмента ДНК (примерно 50 пар оснований). ДНК выделяют из геля методом электроэлюции, обрабатывают фенолхлороформом и очищают осаждением этанолом.
В) Лигирование плазмиды pBR 322 (расщепленной EcoRI - Hind III) с CMK.
К 0,2 мкг (1 мкл) ДНК pBR 322 (обработка Eco RI - Hind III и щелочной фосфатазой) и 0,2 мкг (1 мкл) ДНК множественного сайта клонирования добавляют 1 мкл TE (10 мМ/л трис-HCl (pH 8,0), 1 мМ/л ЭДТА) и 24 мкл раствора для лигирования фирмы Takara, набор А, и смесь тщательно перемешивают. К полученному раствору добавляют 3 мкл раствора для лигирования Takara, набор Б и смесь инкубируют при 16oC в течение 4 ч.
С) Клонирование плазмиды pBR 322M.
К 3 мкл (40 нг) полученного раствора для лигирования добавляют 200 мкл клеточной суспензии E.coli HB 101, обработанной 50 мМ/л CaCl2, и смесь слабо перемешивают. Затем ее инкубируют в ледяной воде в течение 30 мин, а далее при 42oC в течение 2 мин, чтобы клетки приобрели способность захватывать ДНК. К полученной суспензии добавляют 1,8 мл жидкой среды 2 YT (16 г/л триптона, 10 г/л дрожжевого экстракта, 5 г/л NaCl), и смесь встряхивают при 37oC в течение 1 ч, а затем помещают на агаровую среду LB (10 г/л триптона, 8 г/л NaCl, 5 г/л дрожжевого экстракта), содержащую 50 мкг/мл ампициллина. Плазмиды выделяют из полученных колоний, анализируют их карты расщепления эндонуклеазами рестрикции, чтобы отобрать колонии, содержащие нужную плазмиду (pBR 322M). Отобранные колонии культивируют в 200 мл жидкой среды 2YT, содержащей 100 мкг/мл ампициллина и плазмидную ДНК получают в большом количестве щелочным методом при использовании SDS.
(2) Выделение PvuII-Eco 47111 фрагмента (2550 пар оснований).
Подготавливают три эппендорфовских пробирки, в каждую из которых вносят 10 мкг (10 мкл) ДНК плазмиды pBR 322M, полученной, как указано на стадии (1), и затем добавляют 20 мкл 10 x M буфера (100 мМ/л трис-HCl (pH 7,5), 100 мМ/л MgCl2, 10 мМ/л DTT, 500 мМ/л NaCl). 120 единиц (10 мкл) PvuII (Takara Shuzo Co., Ltd) и стерилизованной воды до общего объема смеси 200 мкл. Пробирки инкубируют при 37oC в течение 3 ч. После этого смеси обрабатывают фенолхлороформом и собирают ДНК путем осаждения этанолом и растворяют в 174-мкл порциях стерилизованной воды. К полученным растворам добавляют 20-мкл порции 10 x H буфера и аликвоты, содержащие 24 единицы Eco47111 (Takara Shuzo Co., Ltd). Затем смеси инкубируют при 37oC в течение 3 ч. ДНК выделяют осаждением этанолом и нужный фрагмент ДНК (2550 пар оснований) отделяют от остальной ДНК с помощью электрофореза в 1,5%-ном агарозном геле. Выделенный фрагмент ДНК очищают, используя систему Geneclean, и помещают его в 50 мкл раствора 0,1 моль трис-HCl (pH 8,0). К указанному раствору добавляют 5 мкл раствора щелочной фосфатазы и далее смесь инкубируют при 37oC в течение 1 ч. После этого добавляют 5 мкл раствора щелочной фосфатазы, и смесь инкубируют при 65oC в течение 30 мин. После окончании реакции смесь обрабатывают фенолхлороформом и осаждают ДНК этанолом. ДНК собирают и растворяют в 20 мкл TE-буфера.
(3) Выделение точки начала репликации pBAS18 в A.nidulans.
В клетки E.coli HB 101 встраивают челночный вектор pBAS 18 (K.Shinozaki et al. , Gene 19:221-224 (1982)) Escherichia coli и A.nidulans, причем эндогенную плазмиду (pBA 1, расщепленную BamHI) штамма 6301 A.nidulans вводят в сайт BamHI плазмиды pBR322. Полученный штамм культивируют в жидкой среде LB, содержащей 50 мкг/мл ампициллина, и векторную плазмиду выделяют в большом количестве, используя SDS - щелочной метод. К 14 мкг (20 мкл) полученной ДНК pBAS 18 добавляют 20 мкл 10 • K буфера, 100 единиц (10 мкл) BamHI (Takara Shuzo Co., Ltd) и стерилизованной воды до конечного объема 200 мкл в эппендорфовской пробирке. Приготавливают три эппендорфовские пробирки, содержащие те же компоненты, как указано выше, и пробирки инкубируют при 30oC в течение 3 ч. После окончания реакции ДНК выделяют осаждением этанолом и нужный фрагмент ДНК (pBA 1, около 8,0 т.п.о.) отделяют от остальной ДНК с помощью гель-электрофореза в 1%-ном агарозном геле. Затем фрагмент очищают с помощью системы Geneclean. К 2 мкг (5 мкл) выделенного и очищенного фрагмента pBA 1 (расщепленного BamHI) добавляют 5 мкл 10 x K буфера, 24 единицы (2 мкл) XhoI (Takara Shuzo Co., Ltd) и 38 мкл стерилизованной воды, и смесь инкубируют при 37oC в течение 3 ч. После окончания реакции смесь обрабатывают фенолхлороформом и ДНК осаждают этанолом. Оба концевых участка полученного фрагмента, расщепленного pBAI и BamHI - Xho 1, затупляют при использовании набора Takara Blanting.
(4) Конструирование "миниатюризованной" E.coli - A.nidulans челночной векторной плазмиды pBAX 18 (6,9 т.п.о.).
К 40 нг (2 мкл) ДНК с тупыми концами и 200 нг (4 мкл) PvuII - Eco 47111 - фрагмента ДНК добавляют 48 мкл раствора для лигирования, набор А, фирма Takara, после чего тщательно перемешивают. Затем добавляют 6 мкл раствора Б, и инкубируют смесь при 16oC в течение 4 ч. Штамм HB 101 E.coli трансформируют с помощью указанного раствора и переносят клетки на агаровую среду LB, содержащую 50 мкг/мл ампициллина и 1,5% агара, для получения колоний. Плазмиды выделяют из полученных колоний, анализируют их карты расщепления эндонуклеазами рестрикции, чтобы отобрать колонии, содержащие нужную плазмиду pBAX18. Отобранные колонии культивируют в 45-мл аликвотах жидкой среды 2 YT, содержащей 100 мкг/мл ампициллина, и плазмидную ДНК получают щелочным методом при использовании SDS.
(5) Конструирование pBAX20 (около 5,8 тыс. оснований).
К 1 мкг (4 мкл) BamHI-XhoI (затупленному фрагменту ДНК, полученному на стадии (3), добавляют 2 мкл 10 • M буфера, 1 мкл (12 единиц) PvuII и 13 мкл стерилизованной воды и инкубируют смесь при 37oC в течение 3 ч. К 2 мкл (100 нг) полученного реакционного раствора добавляют 100 нг (2 мкл) PvuII-Eco 47 - фрагмента ДНК, полученного на стадии (2), 16 мкл раствора для лигирования, набор А, фирма Takara. Затем после тщательного перемешивания добавляют раствор Б, и осуществляют инкубирование при 16oC в течение 2 ч. После окончания реакции штамм HB 101 E.coli трансформируют с помощью указанного раствора и переносят клетки на агаровую среду LB, содержащую 50 мкг/мл ампициллина и 1,5% агара для получения колоний. Колонии, содержащие нужную плазмиду (pBAX20), отделяют от остальных колоний и культивируют в 200 мл жидкой среды 2 YT, содержащей 100 мкг/мл ампициллина, и плазмидную ДНК получают щелочным методом при использовании SDS.
Пример 2.
Трансформация A.nidulans 6301.
Клетки, выращенные в 100 мл жидкой среды BG-11 в течение 1-5 дней, собирают центрифугированием при 8000 оборотах в минуту в течение 5 мин и суспендируют в 10 мл свежей жидкой среды (108-109 клеток/мл). Аликвоты полученной клеточной суспензии объемом 1 мл помещают в полиэтиленовые пробирки (Falcon 2059), и затем в них добавляют плазмидную ДНК, полученную, как указано в примере 1 в концентрации 1 мкг (количество pBAS 19 составляет 10 мкг). Пробирки закрывают алюминиевой фольгой и инкубируют в течение ночи при 30oC. Затем алюминиевую фольгу удаляют и продолжают культивирование при 30oC на свету (1000-2000 лк). Клеточные суспензии разбавляют в 5 раз (суспензии с pBAS 18 не разбавляют) и помещают в количестве от 100 до 500 мкл на агаровую среду BG-11, содержащую 1 мкг/мл ампициллина, 1 мМ/л тиосульфата натрия и 1,5% агара. Затем культуры на чашках выращивают на свету (2000-3000 лк) в течение 4-10 дней.
Частота трансформации полученных колоний составляет 10-3 - 10-4 клеток/мкг плазмидной ДНК при использовании заявленных плазмид. Эта величина в 1000 раз превышает частоту трансформации (10-7 клеток/мкг плазмидной ДНК) при использовании плазмиды pBAS 18.
Плазмиды предназначены для репликации в клетках Escherichia coli и цианобактерий. Плазмиды содержат следующие конструктивные элементы: область начала репликации (ori A), множественный сайт клонирования, oбласть начала репликации (ori E), из которой удален ген, определяющий rop-функцию. Плазмида pBAXI8 имеет размер 6,9 т.п.о., а рВАХ20 - 5,8 т.п.о. Cконструированные плазмиды позволяют повысить частоту трансформации штамма Anacystis nidulans 6301 при упрощении методик генетических манипуляций. 3 с. и 1 з.п. ф-лы, 1 табл., 3 ил.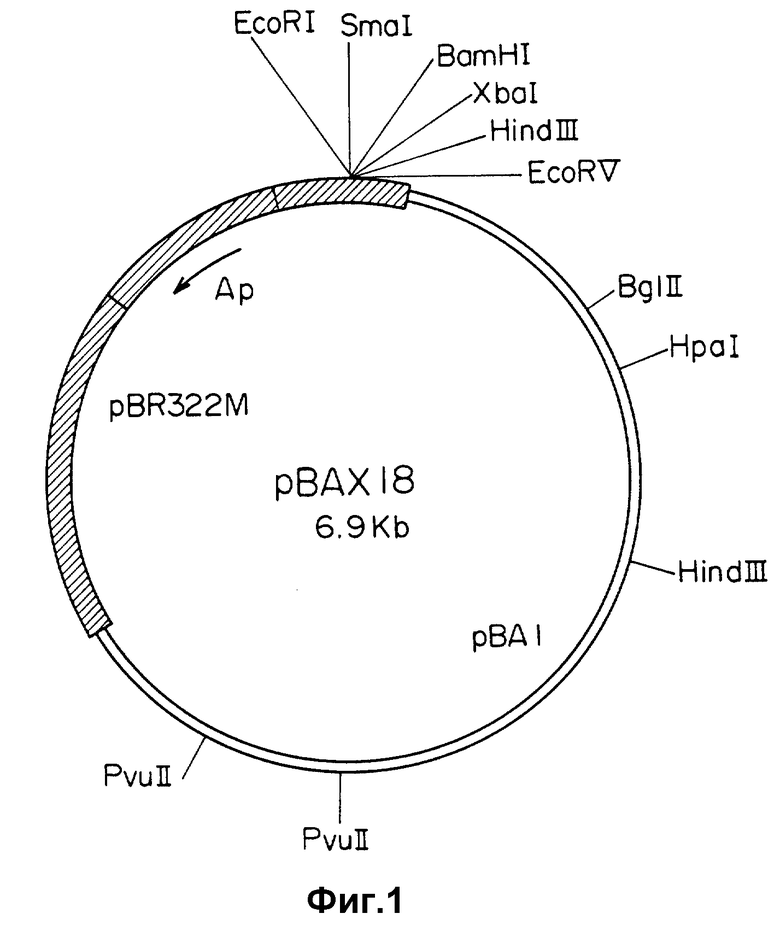
| K.Shinozaki et al | |||
| РАЗБОРНЫЙ ПЛАСТИНЧАТЫЙ РАДИАТОР | 1925 |
|
SU6301A1 |
| Gene, uol | |||
| Способ изготовления электрических сопротивлений посредством осаждения слоя проводника на поверхности изолятора | 1921 |
|
SU19A1 |

