Область техники, к которой относится изобретение
Настоящее изобретение относится к плазмиде, предназначенной для интеграции генов в микроорганизм, который принадлежит к роду Escherichia, но является отличным от штамма Escherichia coli К-12, способу интеграции генов в хромосому, рекомбинантному штамму, сконструированному согласно этому способу, и к способу получения представляющего практический интерес вещества с использованием рекомбинантного штамма.
Предпосылки создания изобретения
При использовании микроорганизма, сконструированного таким образом, чтобы в его клетку была интегрирована плазмида, экспрессирующая определенный ген, для получения представляющего интерес вещества часто возникают проблемы, связанные с осуществлением сверхэкспрессии гена и нестабильностью плазмиды. Эффективным методом решения этих проблем является модификация генов хромосомы. В течение продолжительного периода времени в качестве метода хромосомной инженерии для Escherichia coli применяли метод, основанный на использовании обработки мутагенами. Этот метод направлен на отбор мутантного штамма с требуемыми свойствами из полученных в результате неспецифического мутагенеза штаммов, и он является очень трудоемким. Кроме того, при этом практически невозможно осуществлять целенаправленные или рациональные манипуляции.
С другой стороны, известно, что P1-трансдукция с использованием фага Р1 является наиболее универсальным методом, позволяющим осуществлять целенаправленные и рациональные манипуляции с хромосомой Escherichia coli [N.D.Zinder и J.Lederberg, J.Bacteriol., 64, 679 (1952)].
Методы манипуляции с хромосомой, отличные от Р1-трансдукции, можно условно разделить на два типа.
К первому типу относится метод, который заключается во встраивании представляющего интерес гена в плазмиду, которая обладает способностью автономно реплицироваться в микроорганизмах, отличных от Escherichia coli, но не способна автономно реплицироваться в Escherichia coli, и трансформации Escherichia coli плазмидой с получением штамма, в котором представляющий интерес ген интегрирован в хромосому этого штамма с помощью метода гомологичной рекомбинации [A.Chen и д., J.BacterioL, 176, 1542 (1994)]. Однако данный метод имеет недостаток, заключающийся в том, что требуемый штамм с рекомбинантной хромосомой можно получать только с очень малой частотой, поскольку плазмида, которую создают с использованием в качестве клетки-хозяина микроорганизма, отличного от Escherichia coli, расщепляется рестриктазами в Escherichia coli.
Ко второму типу относится метод, который заключается в том, что применяют плазмиду, которая в нормальных условиях для роста обладает способностью автономно реплицироваться в штамме Escherichia coli К-12, но не способна автономно реплицироваться в определенных условиях, например, при высокой температуре, и представляющий интерес ген интегрируют в хромосому согласно методу гомологичной рекомбинации [Т.Hashimoto и М.Sekiguchi, J.Bacteriol., 127. 1561 (1976)].
В научной и промышленной сфере широко используют такие штаммы Escherichia coli, как К-12, В и W. Многие рекомбинантные методы генной инженерии основаны на применении штамма К-12.
Штамм Escherichia coli W можно использовать для получения имеющих практическое значение веществ, таких как аминокислоты и т.д. [S.Fukurawa и др., Appl. Microbiol. Biotechnol., 29, 253 (1988)], и он также находит широкое применение в процессах ферментации, поскольку он ассимилирует сахарозу и его можно успешно использовать в культуре клеток с высокой плотностью [I.E.Gleiser и S.Bauer, Biotechnol. Bioeng., 23, 1015 (1981)].
Вышеуказанные методы генной инженерии касаются использования штамма К-12 и до настоящего времени не имеется информации о использовании микроорганизмов, принадлежащих к роду Escherichia, но отличных от штамма К-12.
Описание изобретения
В основу настоящего изобретения была положена задача разработать новый способ хромосомной рекомбинации с использованием микроорганизмов, принадлежащих к роду Escherichia, но отличных от штамма К-12, с целью решения указанных выше проблем.
Объекты настоящего изобретения перечислены ниже в пунктах (1)-(14).
(1) Чувствительная к температуре плазмида, которая способна автономно реплицироваться в штамме Escherichia coli К-12 при 10-30°С, но при температуре 33°С или выше не способна автономно реплицироваться в штамме Escherichia coli К-12 или негомогенно распределяется при делении клеток штамма Escherichia coli К-12, вследствие чего она не обладает способностью стабильно сохраняться в клетках штамма Escherichia coli К-12 при указанных температурах, и которая не способна автономно реплицироваться в микроорганизме, принадлежащем к роду Escherichia, но отличном от штамма Escherichia coli К-12, или негомогенно распределяется при делении клеток указанного микроорганизма при любой температуре, вследствие чего она не обладает способностью стабильно сохраняться в клетках указанного микроорганизма.
(2) Плазмида по указанному выше п.(1), где микроорганизм, принадлежащий к роду Escherichia, но отличный от штамма Escherichia coli К-12, представляет собой штамм Escherichia coli W или Escherichia coli В.
(3) Плазмида по указанным выше п.(1) или п.(2), где плазмида представляет собой плазмиду, содержащую фрагмент ДНК, который обладает способностью подвергаться гомологичной рекомбинации с хромосомой Escherichia coli.
(4) Плазмида по любому из указанных выше п.п.(1)-(3), где плазмида содержится в штамме Escherichia coli DH5α/pMTS11910 (FERM ВР-6904) или в штамме Escherichia coli DH5α/pMTS11914 (FERM BP-6905).
(5) Плазмида, несущая один или несколько интегрированных в нее генов, которую получают путем интеграции любых указанных генов в плазмиду по любому из п.п.(1)-(4).
(6) Способ интеграции одного или нескольких генов, который заключается в том, что плазмиду по любому из указанных выше п.п.(1)-(5) встраивают в микроорганизм, принадлежащий к роду Escherichia, но отличный от штамма Escherichia coli К-12.
(7) Способ интеграции одного или нескольких генов по указанному выше п.(6), где микроорганизм, принадлежащий к роду Escherichia, но отличный от штамма Escherichia coli К-12, представляет собой штамм Escherichia coli W или Escherichia coli В.
(8) Способ интеграции одного или нескольких генов по указанному выше п.(6) или п.(7), где интеграция генов представляет собой такую интеграцию генов, при которой плазмиду интегрируют в хромосому.
(9) Способ интеграции одного или нескольких генов по любому из указанных выше п.п.(6)-(8), где интеграция одного или нескольких генов представляет собой такую интеграцию, при которой фрагмент ДНК плазмиды заменяют фрагментом ДНК хромосомы путем гомологичной рекомбинации.
(10) Трансформант, который может быть получен способом согласно любому из указанных выше п.п.(6)-(9).
(11) Трансформант по указанному выше п.(10), где трансформант представляет собой трансформант, выбранный из группы, включающей штаммы Escherichia coli DH5α/pMTS11910 (FERM BP-6904), Escherichia coli DH5α/pMTS11914 (FERM BP-6905), Escherichia coli WLA-131 (FERM BP-6902) и Escherichia coli WL-1133 (FERM BP-6903).
(12) Способ получения имеющего практическое значение вещества, заключающийся в том, что трансформант по указанному выше п.(10) или п.(11) культивируют в среде, в результате чего происходит образование и накопление в культуре имеющего практическое значение вещества, и выделяют имеющее практическое значение вещество из культуры.
(13) Способ по указанному выше п.(12), где имеющее практическое значение вещество выбирают из группы, включающей аминокислоты, органические кислоты, нуклеиновые кислоты, вещества, связанные с нуклеиновыми кислотами, сахара, липиды, витамины и пигменты, а также их производные.
(14) Способ получения имеющего практическое значение вещества по указанному выше п.(12), где имеющее практическое значение вещество представляет собой протеин.
[1] Мутагенная обработка плазмиды
В качестве плазмиды используют pMW119 (которую можно получить от фирмы Nippon Gene Co., Ltd.) или плазмиду, содержащую такие же ori-область и ген rep, что и плазмида pMW119. ori-область представляет собой область, содержащую сайт инициации репликации, а ген rep представляет собой ген, кодирующий репликазу.
Мутагенную обработку плазмиды осуществляют согласно известному методу с использованием гидроксиламина [G.O. Humphreys и др., Mol. Gen. Genet., 145, 101 (1976)] и т.д.
Если используют гидроксиламин, то плазмиду можно, например, подвергать мутации путем растворения приблизительно 10 мкг плазмиды в фосфатном буфере [50 ммолей/л NaH2PO4, 1 моль/л ЭДТК·2Nа, рН 6,0 (NaOH)], содержащем 0,4 моля/л гидрохлорида гидроксиламина, и выдерживания раствора при 75°С в течение 30-60 мин.
[2] Получение чувствительных к температуре плазмид
Плазмиду, обладающую чувствительностью к температуре, можно получать в штамме Escherichia coli К-12 известным методом [Т.Hashimoto и М.Sekiguchi, J.Bacteriol, 127, 1561 (1976)].
Более конкретно, штамм Escherichia coli К-12 трансформируют плазмидой, подвергнутой мутагенной обработке, получая штамм, обладающий устойчивостью к такому антибиотику, как ампициллин, при температурах 10-32°С. Хотя можно применять любой метод трансформации, позволяющий трансформировать штамм Escherichia coli К-12, предпочтительно применять электропорацию [W.J.Dower и др., Nucleic Acids Res., 16, 6127 (1988)], которая обладает высокой эффективностью в отношении трансформации.
Штамм Escherichia coli К-12, подвергнутый обработке с целью трансформации, распределяют на агаровой LB-среде [1,0% бакто-триптона (фирма Difco), 0,5% дрожжевого экстракта (фирма Difco), 1,0% NaCl], содержащей антибиотик, применяемый в качестве маркера, и культивируют при 10-32°С в течение 6-24 ч.
В качестве трансформантов отбирают штаммы, у которых при культивировании был выявлен рост. Трансформанты выращивают на агаровой среде, не содержащей никакого дополнительного антибиотика, при температуре 33°С или более высокой температуре, при которой трансформанты могут расти.
После культивирования штаммы, у которых выявлен рост, распределяют на агаровой среде, содержащей антибиотик, такой как ампициллин, и культивируют в течение 6-24 ч при температуре 33°С или более высокой температуре, при которой трансформанты могут расти. Штаммы, которые не могут расти в этих условиях, отделяют от указанной выше агаровой среды, не содержащей никакой дополнительный антибиотик, и культивируют еще раз на среде, не содержащей никакой дополнительный антибиотик. После культивирования из штаммов выделяют плазмиду с помощью стандартного метода. Плазмида представляет собой чувствительную к температуре плазмиду.
[3] Получение плазмид, которые не способны автономно реплицироваться в клетках микроорганизма, принадлежащего к роду Escherichia. но отличного от штамма Escherichia coli К-12
Плазмиду по настоящему изобретению, а именно плазмиду, которая не способна реплицироваться в клетках микроорганизма, принадлежащего к роду Escherichia, но отличного от штамма Escherichia coli К-12, можно отобрать из обладающих чувствительностью к температуре плазмид в штамме Escherichia coli К-12, которые получают согласно способу, описанному выше в п.(1).
В контексте настоящего описания понятие микроорганизмы, принадлежащие к роду Escherichia, но отличные от штамма Escherichia coli К-12, обозначает любые микроорганизмы, принадлежащие к роду Escherichia, но отличные от штамма Escherichia coli K-12. Примерами таких микроорганизмов являются такие принадлежащие к роду Escherichia штаммы микроорганизмов, как Escherichia coli W, Escherichia coli В, Escherichia coli C, Escherichia coli 15 и т.д., причем предпочтительными являются штаммы Escherichia coli W и Escherichia coli В.
Более предпочтительно для получения трансформантов трансформацию микроорганизма, принадлежащего к роду Escherichia, но отличного от штамма Escherichia coli К-12, осуществляют с использованием плазмиды, обладающей чувствительностью к температуре в штамме Escherichia coli К-12. Если трансформанты не способны расти при любой температуре при их культивировании в среде, содержащей антибиотик, то такая чувствительная к температуре плазмида является плазмидой, обладающей характеристиками плазмиды по настоящему изобретению.
[4] Интеграция гена в хромосому
Для интеграции в хромосому используют плазмиду, которая не способна реплицироваться в микроорганизме, принадлежащем к роду Escherichia, но отличном от штамма Escherichia coli К-12, при любой температуре, полученную согласно описанному выше способу (ниже в настоящем описании обозначена как плазмида по настоящему изобретению).
Сначала в плазмиде по настоящему изобретению клонируют ген. Ген может представлять собой любой ген, участвующий в производстве требуемого имеющего практическое значение вещества и имеющий последовательность, гомологичную последовательности представляющего интерес гена в хромосоме микроорганизма, принадлежащего к роду Escherichia, но отличного от штамма Escherichia coli К-12. Имеющее практическое значение вещество может представлять собой любое имеющее практическое значение вещество, если ген участвует в ее производстве. Например, в качестве имеющих практическое значение веществ следует отметить (но не ограничиваясь ими) аминокислоты, такие как лейцин и т.д., органические кислоты, такие как изолимонная кислота и т.д., нуклеиновые кислоты или вещества, связанные с нуклеиновыми кислотами, такие как флавинадениндинуклеотид и т.д., сахара, такие как фруктоза и т.д., липиды, такие как фосфолипид, гликолипид и т.д., витамины, такие как биотин и т.д., и пигменты, такие как каротин и т.д., их производные, протеины, такие как кодируемые геном ферменты и т.д.
Клонирование гена можно осуществлять с использованием штамма Escherichia coli К-12 согласно известному методу [Molecular Cloning, A Laboratory Manual, 2-е изд., Cold Spring Harbor Laboratory Press (1989)] (далее ссылка обозначена как Molecular Cloning, 2-е изд.). Кроме того, можно искусственно модифицировать нуклеотидную последовательность клонируемого гена согласно известному методу (Molecular Cloning, 2-е изд.). Для клонирования гена и модификации клонированного гена в качестве клетки-хозяина используют штамм Escherichia coli К-12 и культивирование осуществляют при температуре 32°С или ниже.
Сконструированную таким образом рекомбинантную плазмиду используют для трансформации микроорганизма, принадлежащего к роду Escherichia, но отличного от штамма Escherichia coli К-12, и штамм культивируют в среде, содержащей антибиотик, такой как ампициллин, с целью создания устойчивого к антибиотику штамма, получая тем самым штамм с рекомбинантной хромосомой, в который интегрирована плазмида, содержащая представляющий интерес ген. Для трансформации микроорганизма можно применять любые известные методы, такие как электропорация, метод, основанный на использовании хлорида кальция и т.д. (Molecular Cloning, 2-е изд.).
Также можно получать штамм, в котором клонированный в плазмиде ген заменен геном, встречающимся в естественных условиях в хромосоме микроорганизма, принадлежащего к роду Escherichia, но отличного от штамма Escherichia coli К-12.
Более конкретно, штамм, трансформированный плазмидой по настоящему изобретению, культивируют в среде без добавления антибиотика, такого как ампициллин. Для культивирования, а также последующего выращивания трансформантов можно применять либо твердую агаровую среду, либо жидкую, не содержащую агар среду.
Культивируемые клетки соответствующим образом разбавляют стерилизованным физиологическим раствором, распределяют на агаровой среде LB, не содержащей антибиотик, такой как ампициллин, и культивируют. Каждую из выращенных колоний распределяют на агаровой среде, содержащей антибиотик, такой как ампициллин, и штаммы, соответствующие штаммам, которые не росли на среде с добавлением ампициллина, выделяют из исходной среды, не содержащей антибиотик. Из отобранных штаммов можно выделить штамм, в котором ген, встроенный в плазмиду по настоящему изобретению, и ген, присутствующий в естественных условиях в хромосоме микроорганизма, принадлежащего к роду Escherichia, но отличного от штамма Escherichia coli К-12, заменены в результате гомологичной рекомбинации.
Штамм с рекомбинантной хромосомой, в котором плазмида, содержащая представляющий интерес ген, встроена в хромосому, или штамм, в котором ген, встречающийся в естественных условиях в микроорганизме, принадлежащем к роду Escherichia, но отличному от штамма Escherichia coli К-12, заменен клонированным в плазмиде геном, в настоящем описании обозначен как микроорганизм по настоящему изобретению.
С помощью микроорганизма по настоящему изобретению можно получать имеющие практическое значение вещества.
Согласно вышеописанному методу, если интеграцию плазмиды в хромосому создают таким образом, чтобы сайт интеграции находился в нуклеотидной последовательности гена-мишени, можно дезинтегрировать ген-мишень в результате встраивания новой последовательности в ген-мишень. Такая дезинтеграция гена, получаемая при использовании указанного метода, имеет преимущество, заключающееся в большей надежности по сравнению с инактивацией гена мутацией.
Для культивирования микроорганизма по настоящему изобретению можно использовать любые естественные среды и синтетические среды, а также среду, обеспечивающую эффективное культивирование микроорганизма по настоящему изобретению, содержащую источники углерода, источники азота, неорганические соли и т.д., которые могут ассимилироваться микроорганизмом по настоящему изобретению.
В качестве источников углерода можно применять любые источники углерода, которые могут ассимилироваться микроорганизмом по настоящему изобретению. Примеры приемлемых источников углерода включают углеводы, в том числе глюкозу, фруктозу, сахарозу, мелассу, крахмал и крахмальный гидролизат; органические кислоты, такие как уксусная кислота и пропионовая кислота; и спирты, такие как этанол и пропанол.
В качестве источников азота можно применять аммиак, аммонийные соли различных органических или неорганических кислот, такие как хлорид аммония, сульфат аммония, ацетат аммония и фосфат аммония, и другие азотсодержащие соединения, такие как пептон, мясной экстракт, дрожжевой экстракт, жидкость, полученная после замачивания зерен кукурузы до разбухания, казеиновый гидролизат, соевый жмых, гидролизат соевого жмыха и различные подвергнутые брожению клетки микроорганизмов и продукты их расщепления.
Примеры неорганических солей включают первичный кислый фосфат калия, вторичный кислоты фосфат калия, фосфат магния, сульфат магния, хлорид магния, сульфат железа, сульфат марганца, сульфат меди и карбонат кальция.
Культивирование осуществляют в аэробных условиях, например, встряхивая культуру или вращая погруженную культуру при аэрации.
Приемлемая для культивирования температура составляет 15-40°, а период культивирования, как правило, составляет от 16 ч до 7 дней. В процессе культивирования значение рН предпочтительно поддерживают на уровне 3,0-9,0. Регулирование рН осуществляют с помощью органической или неорганической кислоты, щелочного раствора, мочевины, карбоната кальция, аммиака и т.д.
При необходимости в процессе культивирования в среду можно добавлять антибиотики, такие как ампициллин, тетрациклин и т.д.
С помощью описанного выше метода культивирования можно накапливать в культуре имеющее практическое значение вещество. После завершения культивирования имеющее практическое значение вещество можно выделять из культуры путем удаления осадков, таких как клетки, и подвергать различным методам обработки, таким как ионообменная хроматография, конденсация, высаливание и т.д.
Если имеющее практическое значение вещество представляет собой протеин, то его можно выделять и очищать общепринятыми методами, предназначенными для выделения и очистки протеинов. Например, если протеин экспрессируется в клетках в растворимой форме, то после завершения культивирования клетки выделяют центрифугированием и суспендируют в водном буфере с последующим разрушением с помощью ультразвукового дезинтегратора, пресса Френча, гомогенизатора Мантона Гаулина, мельницы типа Dynomill или т.п., получая бесклеточный экстракт. Очищенный протеин можно выделять из супернатанта, полученного центрифугированием бесклеточного экстракта, используя индивидуально или в сочетании общепринятые методы выделения и очистки протеинов, такие как экстракция растворителем, высаливание сульфатом аммония и т.д., обессоливание, осаждение органическим растворителем, анаонообменная хроматография с использованием смол, таких как диэтиламиноэтил (ДЭАЭ)-Сефароза и DIAION НРА-75 (фирма Mitsubishi Kasei Corporation), катоонообменная хроматография с использованием смол, таких как S-Сефароза FF (фирма Pharmacia), гидрофобная хроматография с использованием смол, таких как бутил-Сефароза и фенил-Сефароза, гель-фильтрация с использованием молекулярного сита, аффинная хроматография, хроматофокусирование, электрофорез, такой как изоэлектрическое фокусирование или т.п.
Если протеин экспрессируется в клетках в виде внутриклеточного тельца, то клетки также выделяют и разрушают, а затем подвергают центрифугированию, получая в осадившейся фракции содержащее протеин внутриклеточное тельце. Выделенное содержащее протеин внутриклеточное тельце солюбилизируют с помощью денатурирующего протеин агента. Раствор солюбилизированного протеина разбавляют или подвергают диализу, уменьшая концентрацию денатурирующего протеин агента, находящегося в растворе солюбилизированного протеина, восстанавливая тем самым нормальную стерическую структуру протеина. Затем, с помощью аналогичных описанным выше процессов выделения и очистки можно получать очищенный препарат протеина.
Ниже изобретение пояснено с помощью примеров. Эти примеры не направлены на ограничение объема изобретения.
Краткое описание чертежей
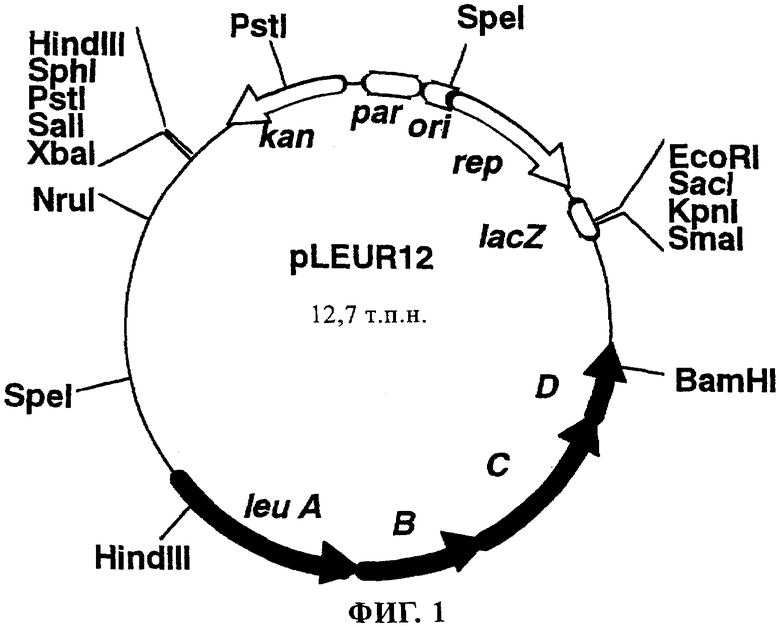
На фиг.1 в виде диаграммы представлена рестрикционная карта плазмиды pLEUR12, содержащей оперон leu, и положение каждого гена в опероне leu, предсказанное на основе информации о штамме Escherichia coli К-12.
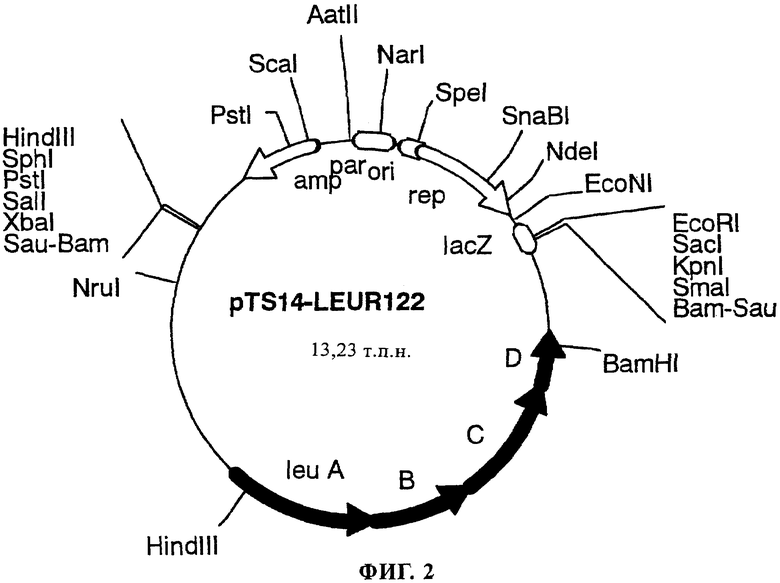
На фиг.2 в виде диаграммы представлена рестрикционная карта плазмиды pTS14-LEUR122, содержащей оперон leu, и положение каждого гена.
Предпочтительный вариант осуществления изобретения
Если не указано иное, то операции, приведенные в следующих примерах, можно осуществлять согласно методикам, описанным в Molecular Cloning, 2-е изд.
Пример 1
Получение плазмиды. которая не сохраняется в штамме Escherichia coli W
Типичный плазмидный вектор Escherichia coli pMW119 (продукт фирмы Nippon Gene Co., Ltd.) подвергали обработке мутагеном in vitro. Обработку мутагеном осуществляли согласно указанному ниже методу с использованием гидроксиламина, который описан у G.O.Humphreys и др., Mol. Gen. Genet., 145, 101 (1976).
Примерно 10 мкг полученной ультрацентрифугированием pMW119 растворяли в фосфатном буфере [50 ммолей/л NaH2PO4, 1 ммоль/л ЭДТК·2Na, рН 6,0 (NaOH)], содержащем 0,4 моля/л гидрохлорида гидроксиламина. Раствор выдерживали при 75°С в течение 40 мин. После этого обработанную ДНК осаждали, добавляя этанол, и растворяли в ТЕ-растворе [10 ммолей/л трис(гидроксиметил)метана, 1 ммоль/л ЭДТК·2Nа, рН 8,0 (HCl)].
С помощью раствора ДНК трансформировали штамм Escherichia coli DH5α (фирма Bethesda Research Laboratories). Штамм культивировали на агаровой среде LB [1% бакто-триптона (фирма Difco), 0,5% дрожжевого экстракта, 0,5% NaCl, 2% агара], дополненной 50 мг/л ампициллина, при 30°С и отбирали примерно 1700 выросших штаммов.
Обобранные штаммы пересевали на агаровую среду LB, не содержащую ампициллин, и культивировали при 42°С. Выросшие колонии вновь пересевали на агаровую среду LB, содержащую ампициллин, и культивировали при 42°С, и из исходного планшета отбирали 22 штамма, которые не могли расти на среде с ампициллином.
С использованием плазмиды, экстрагированной из отобранных штаммов, трансформировали штаммы Escherichia coli W (АТСС-11105) и Escherichia coli В (АТСС-11303) с помощью электропорации [William J. Dower и др., Nucleic Acids Research, 16, 6127, 1988]. В этом случае эффективность трансформации составляла 7×108 клеток/мкг pBR322.
Для каждого из штаммов W и В два типа плазмид, при применении которых получали трансформанты, обладающие устойчивостью к ампициллину, с частотой 1×10-7 или ниже по сравнению с частотой, с которой получали трансформанты при применении плазмиды pMW119, были обозначены соответственно как pMTS11910 и pMTS11914. Штамм Escherichia coli DH5α трансформировали с помощью pMTS11910 или pMTS11914. Полученные трансформанты, т.е. Escherichia coli DH5α/pMTS11910 и Escherichia coli DH5α/pMTS11914, в соответствии с Будапештским соглашением были депонированы в Международной коллекции организмов для процедуры патентования в Национальном институте передовой индустриальной науки и технологии(National Institute of Advanced Industrial Science and Technology, International Patent Organism Depositary: Tsukuba Central 6, 1-1-1-Higashi, Tsukuba, Ибараки, Япония) (ранее Агентство индустриальной науки и технологии Национального института биологии и технологии человека: Agency of Industrial Science and Technology, National Institute of Bioscience and Human-Technology: 1-1-1-Higashi, Tsukuba, Ибараки, Япония) 30 сентября 1999 г. под номерами FERM ВР-6904 и FERM ВР-6905 соответственно.
Пример 2
Конструирование чувствительной к температуре плазмиды, содержащей оперон leu
В качестве гена, применяемого для хромосомной генной интеграции, применяли оперон leu L-лейцина, продуцируемого штаммом Escherichia coli (FERM BP-4704) (опубликованная заявка на неисследованный японский патент (Japanese Published Unexamined Patent Application) 70879/96). Хромосомную ДНК штамма FERM BP-4704 экстрагировали согласно методу, описанному у Saito с соавторами [Н.Saito и К.Miura, Biochem. Biophys. Acta, 72, 619 (1963)], и хромосомную ДНК частично расщепляли обработкой рестриктазой Sau3AI. С другой стороны, pMW218 (фирма Nippon Gene Co., Ltd.) расщепляли обработкой рестриктазой BamHI и щелочной фосфатазой. Обработанные таким образом растворы ДНК смешивали и подвергали реакции лигирования с использованием ДНК-лигазы фага Т4.
Путем электропорации с использованием ДНК, подвергнутой обработке лигазой, трансформировали нуждающийся в присутствии L-лейцина штамм Escherichia coli CV437 [M.G.Davis и J.M.Calvo, J.Bacteriol., 129, 1078 (1977)]. В результате получали 66 штаммов, обладающих устойчивостью к канамицину и не нуждающихся в присутствии L-лейцина. Не нуждающиеся в присутствии L-лейцина штаммы получали путем отбора штаммов, которые могли расти при культивировании на минимальной среде М9, не содержащей L-лейцин.
Из этих штаммов произвольно отбирали 12 штаммов и экстрагировали плазмидную ДНК. С использованием плазмидной ДНК нуждающийся в присутствии L-лейцина штамм Escherichia coli CV524 [M.G.Davis и J.M.Calvo, J.Bacteriol., 129, 1078 (1977)] трансформировали электропорацией и плазмиду, из которой получали не нуждающиеся в присутствии L-лейцина штаммы, обозначили как pLEUR12. Поскольку штамм CV437 является дефектным в отношении leu А, а штамм CV524 - в отношении leu D, можно предположить, что оперон leu, который содержит по меньшей мере leu А и leu D, был встроен в pLEUR12. Кроме того, рестрикционная карта фрагмента ДНК, встроенного в полученную pLEUR12, совпадает с картой, соответствующей гену leu ABCD штамма Escherichia coli К-12, нуклеотидная последовательность которого описана ранее [Т.Ura и др., Nucleic Acids Research, 20, 3305 (1992)].
Предполагаемое строение pLEUR12 приведено на фиг.1.
Клонированный оперон leu встраивали в pMTS11914 с помощью следующих методов.
Сначала pLEUR12 расщепляли с помощью рестриктаз EcoRI и XbaI и расщепленные фрагменты ДНК подвергали электрофорезу на агарозном геле и из геля экстрагировали фрагмент длиной примерно 9 т.п.н., содержащий оперон leu.
Параллельно pMTS11914 расщепляли с помощью рестриктаз EcoRI и XbaI и дополнительно обрабатывали щелочной фосфатазой. Обработанные растворы ДНК смешивали и подвергали реакции лигирования с использованием ДНК-лигазы фага Т4. С помощью электропорации с использованием ДНК, подвергшейся обработке лигазой, трансформировали нуждающийся в присутствии L-лейцина штамм Escherichia coli C600 [Brenner S. и J.R.Beckwith, J. Mol. Biol., 139, 629 (1965]. В результате получали примерно 500 устойчивых к ампициллину штаммов. Из этих устойчивых к ампициллину штаммов с помощью указанного выше метода отбирали нуждающиеся в присутствии L-лейцина штаммы и культивировали при 42°С и отбирали штаммы, обладающие чувствительностью к ампициллину. Из отобранных штаммов экстрагировали плазмиду, оценивали схему расщепления плазмиды рестриктазами и плазмиду, в которую был встроен требуемый оперон leu, обозначили как pTS14-LEUR122 (фиг.2).
Пример 3
Интеграция pTS14-LEUR122 в хромосому штамма Escherichia coli W
Штамм Escherichia coli W 113-3 [АТСС-11105, J.BacterioL, 60, 17 (1950)] трансформировали pTS14-LEUR122 путем электропорации. С помощью метода биологического анализа устанавливали, обладают ли полученные в результате трансформации устойчивые к ампициллину штаммы способностью продуцировать L-лейцин или нет.
Более конкретно метод заключался в следующем: полученные устойчивые к ампициллину штаммы распределяли на минимальной агаровой среде М9 (5 г/л глюкозы, 6 г/л Na2HPO4, 3 г/л КН2PO4, 0,5 г/л NaCl, 1 г/л NH4Cl, 1 ммоль/л MgSO4, 0,1 ммоля/л CaCl2, 20 мг/л DL-метионина, 2% агара), содержащую описанный выше нуждающийся в присутствии L-лейцина штамм CV524 в концентрации приблизительно 1×107 клеток/мл, и культивировали при 37°С в течение 12 ч.
Вокруг штаммов, продуцирующих L-лейцин, наблюдался рост штамма CV524 и образовывалось мутное кольцо белого цвета (называемое ниже в настоящем описании как гало). Отбирали штаммы, которые образовывали гало, размер которых превышал средний размер образовавшихся гало, и культивировали при 42°С на агаровой среде LB, содержащей ампициллин. Из этих штаммов отбирали устойчивые к ампициллину штаммы в качестве штаммов, в хромосому которых интегрирована pTS14-LEUR122, и один из этих штаммов обозначили как штамм Escherichia coli WLA 131.
С помощью саузерн-гибридизации подтверждали, что pTS14-LEUR122 интегрирована путем гомологичной рекомбинации в хромосому штамма Escherichia coli WLA 131.
Штамм Escherichia coli WLA 131 в соответствии с Будапештским соглашением был депонирован в Международной коллекции организмов для процедуры патентования в Национальном институте передовой индустриальной науки и технологии (National Institute of Advanced Industrial Science and Technology, International Patent Organism Depositary: Tsukuba Central 6, 1-1-1-Higashi, Tsukuba, Ибараки, Япония) (ранее Агентство индустриальной науки и технологии Национального института биологии и технологии человека: Agency of Industrial Science and Technology, National Institute of Bioscience and Human-Technology: 1-1-1-Higashi, Tsukuba, Ибараки, Япония) 30 сентября 1999 г. под номером FERM ВР-6902.
Пример 4
Получение имеющего замены штамма из штамма, несущего интегрированную в хромосому плазмиду
Штаммом Escherichia coli WLA 131 инокулировали 5 мл среды LB и культивировали при встряхивании в течение 24 ч при 33°С. Полученной культурой (100 мкл) снова инокулировали 5 мл среды LB и культивировали при встряхивании в течение 24 ч при 33°С.
Культуру соответствующим образом разбавляли и наносили на агаровую среду LB.
Для отбора чувствительных к ампициллину штаммов выросшие колонии размножали на агаровой среде LB, содержащей или не содержащей ампициллин. Способность чувствительных к ампициллину штаммов продуцировать L-лейцин оценивали с помощью метода биологического анализа, описанного в примере 3, и один из полученных таким образом образующих гало штаммов обозначили как штамм Escherichia coli WLA 1133.
Штамм Escherichia coli WLA 1133 в соответствии с Будапештским соглашением был депонирован в Международной коллекции организмов для процедуры патентования в Национальном институте передовой индустриальной науки и технологии (National Institute of Advanced Industrial Science and Technology, International Patent Organism Depositary: Tsukuba Central 6, 1-1-1-Higashi, Tsukuba, Ибараки, Япония) (ранее Агентство индустриальной науки и технологии Национального института биологии и технологии человека: Agency of Industrial Science and Technology, National Institute of Bioscience and Human-Technology: 1-1-1-Higashi, Tsukuba, Ибараки, Япония) 30 сентября 1999 г. под номером FERM BP-6903.
Для подтверждения того, что ген, как и планировалось, встроен путем рекомбинации в хромосому штамма Escherichia coli WLA-1133, осуществляли анализ методом саузерн-гибридизации. Для этого pLEUR12 расщепляли с помощью HindIII и BamHI получая содержащий leu ABCD фрагмент длиной 4,4 т.п.н. Фрагмент подвергали модификации с помощью дигоксигенина и использовали в качестве зонда. Хромосомную ДНК экстрагировали из штаммов Escherichia coli W113-3, Escherichia coli WLA-131 и Escherichia coli WL-1133, разложенных с помощью SacII и KpnI, и подвергали электрофорезу на агарозном геле.
После электрофореза выделенную ДНК переносили на нитроцеллюлозную мембрану и осуществляли гибридизацию с зондом, после чего проводили окрашивание. В родительских штаммах Escherichia coli W113-3 и Escherichia coli WL-1133 был выявлен лишь фрагмент длиной 17 т.п.н., содержащий leu ABCD. В то же время в штамме Escherichia coli WLA-131 были выявлены два фрагмента длиной 11 т.п.н. и приблизительно 19 т.п.н.
pTS14-LEUR122, которую использовали для интеграции в хромосому, содержит один сайт KpnI в своем сайте множественного клонирования. Поэтому, если, как предполагалось, оперон leu в плазмиде интегрирован в хромосому путем гомологичной рекомбинации, то должны быть обнаружены два фрагмента длиной 19 т.п.н. и 11 т.п.н. Таким образом, можно утверждать, что штамм Escherichia coli WLA-131 содержит pTS14-LEUR122, интегрированную в оперон leu в его хромосоме, и что в штамме Escherichia coli WL-1133 в результате повторной гомологичной рекомбинации отсутствует интегрированная плазмида.
Это означает, что полученная из pMTS11914 область pTS14-LEUR122, интегрированная в хромосому, удалена из хромосомы и в хромосоме штамма Escherichia coli WL-1133 присутствует одна копия оперона leu.
Пример 5
Оценка производства аминокислот
Штаммами Escherichia coli W113-3, Escherichia coli WLA-131 и Escherichia coli WL-1133 соответственно инокулировали 20 мл затравочной среды (2% глюкозы, 1% пептона, 1% дрожжевого экстракта, 0,25% NaCl, pH 7,0), находящейся в колбе Эрленмейера объемом 250 мл, и культивировали при встряхивании в течение 16 ч при 30°С. Полученными затравочными культурами (по 2,5 мл каждой) инокулировали каждый раз соответственно 25 мл среды для получения продукта (3% глюкозы, 1,6% сульфата аммония, 0,1% первичного кислого фосфата калия, 0,2% жидкости, полученной после замачивания зерен кукурузы до разбухания, 150 мг/л DL-метионина, 4% тримагнийфосфата, 1% карбоната кальция, pH 7,0), находящейся в колбе Эрленмейера объемом 250 мл, и культивировали при встряхивании в течение 48 ч при 30°С. Накопление L-лейцина в культуре после завершения процесса культивирования оценивали количественно с помощью жидкостной хроматографии высокого разрешения.
Результаты приведены в Таблице.
Промышленная применимость
С использованием плазмиды, которая обладает способностью автономно реплицироваться в штамме Escherichia coli К-12, но не обладает способностью автономно реплицироваться в штаммах Escherichia, отличных от штамма Escherichia coli К-12 (микроорганизмы, принадлежащие к роду Escherichia, но отличные от штамма Escherichia coli К-12), можно интегрировать представляющие интерес гены в любую область хромосомы микроорганизмов, принадлежащих к роду Escherichia, но отличных от штамма Escherichia coli К-12, или эффективно модифицировать представляющие интерес гены, присутствующие в хромосоме.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИЙ СЕМЕЙСТВА ENTEROBACTERIACEAE | 2010 |
|
RU2460793C2 |
| СПОСОБ ПРОИЗВОДСТВА L-АМИНОКИСЛОТ | 2007 |
|
RU2422530C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ СЕМЕЙСТВА Enterobacteriaceae, ОБЛАДАЮЩЕЙ ПОВЫШЕННОЙ ЭКСПРЕССИЕЙ ГЕНОВ КАСКАДА ОБРАЗОВАНИЯ ФЛАГЕЛЛ И КЛЕТОЧНОЙ ПОДВИЖНОСТИ | 2012 |
|
RU2508404C1 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ СЕМЕЙСТВА Enterobacteriaceae | 2010 |
|
RU2501856C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ СЕМЕЙСТВА Enterobacteriaceae | 2010 |
|
RU2501858C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ СЕМЕЙСТВА Enterobacteriaceae | 2010 |
|
RU2497943C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ ESCHERICHIA | 2005 |
|
RU2311454C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ СЕМЕЙСТВА Enterobacteriaceae | 2010 |
|
RU2501857C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-треонина С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ Escherichia | 2006 |
|
RU2351646C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИЙ, ПРИНАДЛЕЖАЩИХ К РОДУ ESCHERICHIA | 2005 |
|
RU2304615C2 |
Изобретение относится к биотехнологии. Плазмидный вектор, способный к автономной репликации в штамме Escherichia coli К-12 при любой температуре. Содержит область ori с мутацией, ген rep из pMW119 и фрагмент ДНК. Фрагмент ДНК способен интегрироваться в хромосому микроорганизма, принадлежащего к Escherichia coli W или Escherichia coli В в результате гомологичной рекомбинации. Плазмиду встраивают в микроорганизм, принадлежащий к Escherichia coli W или Escherichia coli В с последующим получением аминокислоты. Штамм Escherichia coli DH5α/pMTS11910 (FERM ВР-6904) и штамм Escherichia coli DH5α/pMTS11914 (FERM BP-6905), предназначенные для хранения плазмидного вектора. Штамм Escherichia coli WLA-131 (FERM ВР-6902) и штамм Escherichia coli WL-1133 (FERM ВР-6903) - продуценты лейцина. Данное изобретение позволяет осуществлять сверхэкспрессию гена и получать стабильные плазмиды. 7 н. и 5 з.п. ф-лы, 2 ил., 1 табл.
| СПОСОБ НАСТРОЙКИ ЛИНЕЙНЫХ КОРРЕКТОРОВ В КАНАЛАХ ПРОВОДНОЙ СВЯЗИ | 0 |
|
SU191643A1 |
| ПЛАЗМИДА p74 , ОПРЕДЕЛЯЮЩАЯ СИНТЕЗ МИКРОЦИНА И ШТАММ БАКТЕРИЙ ESCHERICHIA COLI, ОБЛАДАЮЩИЙ АНТИБАКТЕРИАЛЬНОЙ АКТИВНОСТЬЮ | 1990 |
|
SU1819428A3 |

