Изобретение относится к способу получения пировиноградной кислоты путем взаимодействия L-молочной кислоты с кислородом в водном растворе в присутствии биокатализаторов гликолат-оксидазы (оксидазы (S)-2-гидроксикислот (КФ 1.1.3.15) и каталазы (КФ 1.11.1.6).
Известны методы получения пировиноградной кислоты процессом сбраживания различных источников углерода (например, глюкозы, дрожжевых экстрактов и пептона). Однако при таких методах пировиноградная кислота обычно образуется с низкими выходами (в расчете на вводимый источник углерода) и с относительно низкой концентрацией ее как одного из компонентов сбраживаемой биомассы. Процесс разделения и выделения пировиноградной кислоты из такого сложного состава смеси продуктов брожения затруднен и дорогостоящ.
В патенте США N 4900668, выданном 13 февраля 1990 года, Cooper раскрывает способ получения пировиноградной кислоты микробиологическим окислением оптически чистой D(-)молочной кислоты. Хотя преимущество указанного способа по сравнению с другими известными методами проведения процесса ферментации состоит в том, что D-молочную кислоту не используют в качестве источника углерода для получения необходимой для реакции биомассы, он требует использования ростовой среды, содержащей второй источник углерода (например маннитол и кукурузный экстракт) как для продукции биомассы, так и ферментативного превращения D-молочной кислоты в пировиноградную кислоту. Кроме того, D-молочная кислота, по сравнению с L-молочной кислотой, менее распространена в природе и является гораздо более дорогостоящим материалом как для производства, так и для покупки.
Известен способ превращения L-молочной кислоты в пировиноградную кислоту с использованием в качестве биокатализатора L-лактат-оксидазы (L-лактат: O-оксидоредуктаза, некарбоксиллирующая, КФ 1.1.3.2) (B.A. Burdick and J.R. Schaeffer, Biotech. Lett. , том 9, стр. 253-258, 1987). L-лактат-оксидаза (выделенная из Pediococcus) катализирует окисление L-лактата при действии O2 с образованием комплекса пирувата с перекисью водорода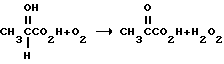
Для ингибирования окисления пирувата побочным продуктом, перекисью водорода, которая приводит к образованию ацетата и CO2, используют L-лактат-оксидазу, соиммибилизованную на одном носителе с каталазой (соотношение оксидазы к каталазе 1:281, М.ед./М.ед.). Окисление 49 мМ растворов L-лактата в 0,1 М фосфатном буфере при pH 7 приводило к 47% суммарному выходу пировиноградной кислоты (выделенную затем в виде производного 2,4-динитрофенилгидразона).
Гликолат-оксидаза (оксидаза (S)-2-гидроксикислот, КФ 1.1.3.15), фермент, обнаруженный, как известно, в листьях зеленых растений и клетках тканей млекопитающих, катализирует окисление гликолевой кислоты до глиоксалевой кислоты. Этот же фермент также катализирует окисление L-молочной кислоты до пировиноградной кислоты, причем с одновременной продукцией перекиси водорода. C. O. Clagget, N.E. Tolbert и R.H.Burris впервые сообщили в журнале J. Biol. Chem., вып. 178, стр. 977-987, 1949 данные об оксидазе α- гидроксикислот, выделенной из большого разнообразия лиственных зеленых растений, которая катализирует окисление гликолевой кислоты и, кроме того, проявляет специфичность относительно L-изомера молочной кислоты. При окислении 80 мМ раствора dl-лактата pH оптимум составлял 7,6, при этом не было идентифицировано или выделено никаких продуктов реакции окисления. N.E. Tolbert и др., J.Biol.Chem., вып. 181 стр. 905-914, 1949 использовали очищенную оксидазу α- гидроксикислот, выделенную из листьев табака, для окисления примерно 113 мМ раствора DL-молочной кислоты в фосфатном буфере при pH 8; в результате реакции было выделено некоторое количество пировиноградной кислоты, о котором не сообщалось, в виде 2,4-динитрофенилгидразона, а также обнаружено значительное количество двуокиси углерода. K.E. Richarson и N.E. Tolbert и др., опубликовали позднее в журнале J.Biol.Chem., вып. 236 стр. 1280-1284, 1961 данные о том, что эта оксидаза α- гидроксикислот, общеизвестна специалистам как оксидаза гликолевой кислоты (или иначе, гликолат-оксидаза).
I Zelitch и S. Ochoa опубликовали в журнале J.Biol.Chem., вып. 201 стр. 707-718, 1953 данные о том, что оксидаза гликолевой кислоты катализирует окисление L-молочной кислоты в присутствии молекулярного кислорода с образованием пировиноградной кислоты и перекиси водорода и что при отсутствии каталазы перекись водорода взаимодействует неферментативным путем с пируватом, образуя при этом ацетат, CO2 и воду. Флавинмононуклеотид (ФМН) был идентифицирован как кофактор, необходимый ферменту для осуществления реакции, и прибавление ФМН к водным растворам этого фермента значительно повышало устойчивость гликолат-оксидазы. В результате окисления 3,3 мМ растворов L-лактата в 50 мМ фосфатном буфере (pH 8,0) и в присутствии избытка вводимой каталазы получена реакционная масса, содержащая 3,3 мМ пировиноградной кислоты (определенное колориметрическим методом); не было выделено ни одного продукта.
Известен также метод окисление лактата до пирувата с использованием оксидазы L -α- гидроксикислот, выделенной из почки крысы M.Blanchard и др., J. Biol. Chem., вып. 163 стр. 137-144, 1946). При окислении 33 мМ раствора лактата в 0,167 М фосфатном буфере, pH 8,0 и в присутствии каталазы, вводимой в избытке, получен 79% выход пирувата (выделенного в виде производного 2,4-динитрофенилгидразона). В качестве дополнительных источников информации, относящихся к катализируемому растворимыми ферментами окислению молочной кислоты до пировиноградной кислоты, можно указать: J.C.Robinson и др., J. Biol. Chem., вып. 237 стр. 2001-2010, 1962 (оксидаза L -α- гидроксикислот из коркового вещества почки свиньи), D.W.Fry и K.E. Richardson, Biochim.Biophys. Acta, вып. 568, стр. 135-144, 1979 (оксидаза гликолевой кислоты из человеческой печени), M.E.Emes и K.H.Erismann, Int.J.Biochem., вып. 16, стр. 1373-1378, 1984 (L-гликолат-оксидаза из Lemna minor), H.S.Kim и J.D.Choi, Korean Biochem.J, вып. 20, стр. 350-356, 1987 (гликолат-оксидаза из листьев шпината).
Хотя специалисты хорошо знакомы с катализируемым ферментами окислением L-молочной кислоты в присутствии кислорода, высокая избирательность к пировиноградной кислоте была продемонстрирована только в одном эксперименте (I Zelitch и S. Ochoa), где концентрация L-лактата составляла 3,3 мМ; такая низкая концентрация L-лактата ограничивала концентрацию образующейся перекиси водорода, а присутствие избытка каталазы лимитировало реакцию перекиси водорода с пируватом до выхода ацетата и CO2. Выделение пирувата при такой низкой концентрацией (примерно 3,2 мМ) из водных растворов реакционной массы нецелесообразно на практике по экономическим соображениям.
Данное изобретение относится к способу получения пировиноградной кислоты (или ее соли, которая более подробно раскрывается далее в описании) путем окисления кислородом L-молочной кислоты (или ее соли) в водном растворе в присутствии комплексных катализаторов, состоящих из двух ферментов гликолат-оксидазы (оксидазы (S)-2-гидроксикислот (КФ 1.1.3.15) и каталазы (КФ 1.11.1.6). Таким образом, изобретение предлагает улучшенный способ получения пировиноградной кислоты, состоящий из стадий взаимодействия в водном растворе L-молочной кислоты с кислородом в присутствии ферментов гликолат-оксидазы и каталазы в течение времени, достаточном для конверсии L-молочной кислоты в пировиноградную кислоту с высокими ее выходами, в котором исходная концентрация L-молочной кислоты составляет примерно от 0,1 до 2,0 М и последующего выделения пировиноградной кислоты. Необходимо понимать, что для целей настоящего изобретения использование терминов для характеристики пировиноградной кислоты и L-молочной кислоты, особенно касающиеся водного раствора при pH от 6 до 10, относятся более конкретно к состоянию с высокой степенью диссоциации, где частично нейтрализованные кислоты в растворе преимущественно присутствуют в виде пируват-иона и L-лактат-иона соответственно. Пировиноградную кислоту можно использовать в качестве химического промежуточного соединения для получения продуктов тонкого органического синтеза, агрохимикатов и лекарственных веществ. Таким образом, в соответствии с целями настоящего изобретения следует понимать, что ссылка в описании изобретения на высокие выходы пировиноградной кислоты и ее выделение предназначена для включения в качестве варианта способ, в котором пировиноградную кислоту получают как промежуточный продукт, включая любое производное пировиноградной кислоты, образуемое соответственно с высоким выходом.
В соответствии с предлагаемым изобретением пировиноградную кислоту получают путем катализируемого ферментами окисления L-молочной кислоты, взятой при концентрациях до 0,5 М, и последующего выделения пирувата при 96% выходе конечного продукта (в виде натриевой соли, 98% чистоты). Использование высокой исходной концентрации L-молочной кислоты, как можно было ожидать, приводит к ингибированию активности гликолат-оксидазы в субстрате, которая обычно ограничивает скорость реакции и/или конечную концентрацию целевого продукта. Аналогичным образом, 0,5 М концентрация пировиноградной кислоты должна была привести к ингибированию активности фермента, который в свою очередь ограничивает концентрацию получаемого продукта.
Кроме того, в ходе создания настоящего изобретения установлено, что высокие выходы пирувата вплоть до 98-99% можно получить в незабуференных реакционных средах без какого-либо регулирования pH; во всех приводимых ранее примерах ферментативное окисление L-лактата осуществляли с использованием буфера, обычно фосфатного буфера.
Высокие выходы пирувата, полученного при отсутствии буфера, аналогичны или превышают его выходы при проведении реакции в присутствии буфера. Получение пирувата при осуществлении реакции в отсутствии буфера позволяет осуществлять выделение продукта непосредственно из реакционной массы; катализатор удаляют фильтрацией или центрифугированием, при этом остается водный раствор, содержащий пируват, который можно легко извлечь при удалении воды (например путем отгонки воды при пониженном давлении, методом лиофилизации и т.п.)
В объеме предлагаемого изобретения показано, что полученные методом генной инженерии биокатализаторы из целых клеток с регулируемой проницаемостью мембраны (например, микробные клетки-трансформанты, содержащие как гликолат-оксидазу, так и каталазу), можно использовать для окисления L-лактата до пирувата. Такие биокатализаторы из целых клеток имеют ряд преимуществ по сравнению с растворимыми ферментами, используемыми в качестве катализаторов в предлагаемом изобретении. Растворы, содержащие биокатализатор из целой клетки, можно барботировать кислородом или кислородсодержащим газом, что приводит к ускорению реакции окисления; барботирование же реакционной массы, содержащей растворимые ферменты, приводит к быстрой необратимой денатурации гликолат-оксидазы.
На заключительной стадии реакции такой биокатализатор из целой клетки можно легко извлечь фильтрованием или центрифугированием для повторного использования, в то время как растворимые ферменты нельзя отделять центрифугированием, а фильтрование приводит к значительной потере активности растворимой гликолат-оксидазы. Показатель восстановления ферментативной активности биокатализаторов из целых клеток чрезвычайно высок после каждого рецикла катализатора (примерно 95% и выше), в то время как активность, определенная в оставшейся растворимой гликолат-оксидазе после одной реакции, обычно варьирует от 40 до 60%. Использование гликолат-оксидазы и каталазы, прикрепленных или иммобилизованных на одной инертной подложке-носителе, имеет множество аналогичных преимуществ и по существу их следует рассматривать как вариант осуществления настоящего изобретения.
Сравнительные данные, полученные в результате окисления 0,50 М растворов L-лактата с использованием 1) растворимой гликолат-оксидазы (ГО) и растворимой каталазы, 2) биокатализатора из клетки-продуцента Hansenula polymorpha с регулированной проницаемостью и 3) биокатализатора из клетки-продуцента Pichia pastoris с регулируемой проницаемостью при одинаковых реакционных условиях, сведены в таблицу (см. примеры 5, 12 и 13, раскрывающие реакционные условия):
Из данных таблицы видно, что выход пирувата выше, а продуцирование побочного ацетата ниже при использовании обоих биокатализаторов из целой клетки с проницаемой мембраной по сравнению с растворимой гликолат-оксидазой и каталазой, используемых в качестве катализатора. При использовании биокатализатора H.polymorpha, взятого при такой же концентрации, которую в эксперименте с растворимыми ферментами используют для гликолат-оксидазы, образуется меньшее количество ацетата (продуцируемого в результате взаимодействия побочной перекиси водорода с пируватом), даже хотя концентрация внутриклеточной каталазы в реакционной массе составляет половину концентрации, используемой в реакции с растворимыми ферментами. Эти результаты и данные, приведенные в примерах, свидетельствуют о неожиданном увеличении продукции пирувата при использовании клеток-трансформантов с регулированной проницаемостью мембраны в качестве катализаторов.
Каталитическое окисление L-молочной кислоты или соответствующей ее соли целесообразно проводить путем взаимодействия L-молочной кислоты с источником молекулярного кислорода в присутствии биокатализатора, который катализирует реакцию окислительной конверсии L-молочной кислоты O2 в пировиноградную кислоту. В качестве такого катализатора используют фермент - гликолат-оксидазу, называемый также оксидазой гликолевой кислоты. Гликолат-оксидазу можно выделить из многочисленных источников, известных специалистам в данной области (которые упоминались выше). Гликолат-оксидазу необходимо использовать в эффективной концентрации в реакции окисления, обычно при концентрации примерно от 0,01 до 1000 МЕ/мл, предпочтительно от примерно 0,01 до примерно 10 МЕ/мл. МЕ (Международная единица активности) определена как количество фермента, которое катализирует трансформацию одного микромоля субстрата в минуту. Методику проведения анализа этого фермента описывают I.Zelitch S. Ochoa, J. Biol.Chem., вып. 201, стр. 707-718, 1953. Этот метод также используют для определения активности извлеченной или регенерированной для повторного использования гликолат-оксидазы.
Оптимальные результаты при использовании гликолат-оксидазы в качестве катализатора окислительной конверсии L-молочной кислоты в пирувиновую кислоту получают введением в реакционный раствор катализатора для разложения перекиси водорода, образующейся в процессе окисления. В качестве одного такого пероксид-разлагающего катализатора, который может действовать в комбинации с гликолат-оксидазой, используют каталазу (КФ 1.11.1.6). Каталаза катализирует разложение перекиси водорода на воду и молекулярный водород. Полагают, что каталаза повышает конечный выход пировиноградной кислоты в предлагаемом способе в результате ускорения реакции распада перекиси водорода, образующейся вместе с пировиноградной кислотой в катализированной гликолат-оксидазой реакции молочной кислоты с O2. Каталазу необходимо использовать при концентрации в интервале от 50 до 50000 МЕ/мл, предпочтительно 2000 - 15000 МЕ/мл. Концентрации каталазы и гликолат-оксидазы целесообразно поддерживать в вышеуказанных интервалах так, чтобы соотношение (определенное в МЕ активности для каждого фермента) каталазы к гликолат-оксидазе составляло по крайней мере 250:1.
Кроме растворимых ферментов, используемых в качестве катализаторов, были получены микробные клетки-трансформанты, которые экспрессируют гликолат-оксидазную активность, а также активность эндогенной каталазы, и продемонстрированы их использование в качестве микробного биокатализатора в пределах объема предлагаемого изобретения. Используемый в предлагаемом изобретении биокатализатор из микробной клетки представляет собой трансформант клеток Hansenula polymorpha (метилотрофных дрожжей). Несколько H. polymorpha трансформантов, имеющих достаточную гликолат-оксидазную активность, получено путем введения фрагмента ДНК, содержащего ген гликолат-оксидазы в экспрессирующий вектор при регуляции промотором форматдегидрогеназой (ФМД). Клетки H. polymorpha трансформировали этой векторной ДНК и проводили селекцию штамма, продуцирующего высокий уровень гликолат-оксидазы, обозначенного как штамм H. polymorpha G01 (депонированный в коллекции типовых культур ARS Patent Culture Collection при лаборатории Nothern Regional Research Laboratory, рег. номер: Y-21065, присвоенный USDA, Peoria, Illinois).
Биокатализаторы из клетки-трансформанта H. polymorpha получены путем выращивания вначале инокулята трансформанта H. polymorpha в 500 мл дрожжевой пептонодекстрозной среде (фирмы Difco), pH 4,4. Полученный инокулят затем вносили в ферментер с 10 л среды, включающей в свой состав 14 г азотистого основания без аминокислот с дрожжами, 50 г сульфата аммония и 100 г метанола, pH 5,0. Ферментер работал в течение 42,5 ч при 37oC, скорости перемешиванием биомассы 400 об/мин, постоянном значении pH 5,0, с 40% содержанием растворенного кислорода (с регулируемой подачей) и давлении воздуха 0,984 кг/см2. На заключительной стадии процесса ферментации прибавляли 1,0 кг глицерина, после чего урожай клеток собирали центрифугированием, замораживали в жидком азоте и хранили при температуре -80oC.
Второй биокатализатор на основе микробной клетки, используемый в предлагаемом изобретении, представляет собой трансформант клеток Pichia pastoris (метилотрофных дрожжей), который экспрессирует гликолат-оксидазу из шпината, а также эндогенную каталазу. Несколько клеток-трансформантов Pichia pastoris, имеющих достаточную гликолат-оксидазную активность, получено путем введения фрагмента ДНК, содержащего ген гликолат-оксидазы шпината, в экспрессирующий вектор Pichia pastoris (pHIL-D4) при регуляции промотором метанол-индуцируемой алкоголь-оксидазой I, в результате чего получили плазмиду pMP1. Штамм P. pastoris GTS115 (NRRL Y-15851) трансформировали плазмидой pMP1, а селекцию проводили с возможностью интеграции линеаризованной плазмиды pMP1 в хромосомный локус алкоголь-оксидазы I и заменой гена алкоголь-оксидазы геном гликолат-оксидазы. Затем проводили селекцию пула указанных трансформантов на максимальное количество число копий интегрированных генов из кассеты генной экспрессии. Выделенный высококопийный трансформант, обозначенный как штамм P. pastoris GTS115-MSP10 был депонирован как штамм NRRL Y-21001.
Клетки P. pastoris получены путем выращивания вначале инокулята в 100 мл среды из дрожжей и азотистого основания, содержащей 1% глицерин. Через 48 ч инкубирования при 30oC клеточную суспензию переносили в ферментер с 10 л среды, включающей в свой состав 134 г азотистого основания без аминокислот с дрожжами, 100 г глицерина и 20 г биотина. Ферментацию проводили при pH 5,0 (регулируемую NH4OH), температуре 30oC, скорости перемешиванием 200 об/мин, аэрации 5 л/мин, давлении воздуха 0,0352 кг/см2 и в присутствии растворенного кислорода, поддерживаемого на уровне по меньшей мере 50% насыщения. Сразу же после истощения глицерина клетки для экспрессии гликолат-оксидазы активировали инкубированием в той же самой среде, за исключением того, что вместо глицерина брали 50 г метанола. Во время индукции проводили ферментативный анализ для определения активности гликолат-оксидазы. Через 24 часовой цикл индуцирования клеточный урожай собирали и обрабатывали глицерином (1 кг). После этого клетки замораживали в жидком азоте и хранили при температуре -80oC.
Перед использованием в качестве катализатора для ускорения окислительной конверсии гликолевой кислоты в глиоксалевую кислоту клетки-трансформантов H. polymorha и Pichia pastoris необходимо активировать. Для получения клеток с достаточной гликолат-оксидазной активностью можно использовать большое разнообразие известных методов, чтобы увеличить клеточную проницаемость (см., например, Felix, Anal. Biochem, вып. 120, стр. 211-234, 1982). В соответствии со стандартной методикой суспензию, содержащую 10 мас.% мокрых клеток в 0,1% растворе (об/об) Тритона X-100 и 20 мМ фосфатного буфера (pH 7,0) перемешивают в течение 15 мин, затем обработанные клетки замораживают в жидком азоте, оттаивают и промывают в буферной системе 20 мМ фосфата и 0,1 мМ флавинмонуклеотид (pH 7,0). Второй способ увеличения клеточной проницаемости проводят перемешиванием суспензии, содержащей 10 мас.% мокрых клеток в 0,2% растворе (мас./об) бензалконийхлорида и 20 мМ фосфатного буфера (pH 7,0), перемешивают в течение 60 мин, затем обработанные клетки промывают в буферной системе 20 мМ фосфата и 0,1 мМ флавинмонуклеотид (pH 7,0). Сразу же после активации количество биокатализатора из целых клеток, добавляемое в реакционную массу, подбирают с возможностью обеспечения необходимого уровня гликолат-оксидазной и каталазной активности, о чем упоминалось ранее в описании для соответствующих растворимых ферментов. Восстановление активности глаколат-оксидазы и каталазы более чем 100% от их первоначальных значений является результатом увеличения проницаемости клеток в течение всего времени протекания реакционного процесса.
Анализ микробных клеток-трансформантов для определения активности глаколат-оксидазы проводили путем точного отмера примерно 5-10 мг мокрых клеток (блотированных на фильтровальной бумаге для удаления избыточной влаги) в 3-мл кварцевую кювету, снабженную магнитной мешалкой и содержащую 2,0 мл раствора, который представлял собой буферную систему, приготовленную из 0,12 мМ 2,6-дихлорфенолиндофенола (ДХФИФ) и 80 мМ Трис (трис(гидроксиметил) аминометана), pH 8,3. Кювету закрывали крышкой с резиновой прокладкой и затем раствор барботировали азотом в течение 5 мин для выделения свободного кислорода. После этого в кювету вводили с помощью шприца 40 мкл смешанного раствора из 1,0 М гликолевой кислоты и 1,0 М трис-буфера, (pH 8,3), и полученную смесь перемешивали с одновременным измерением изменения величины поглощения в зависимости от времени при 605 нм (ε = 22,000).
Анализ каталазной активности проводили путем точного отмера примерно 2-5 мг мокрых клеток (блотированных на фильтровальной бумаге для удаления избыточной влаги) в 3-мл кварцевую кювету, снабженную магнитной мешалкой и содержащую 2,0 мл дистиллированной воды раствора, к которой затем добавляли 1,0 мл 59 мМ перекиси водорода в 50 мм фосфатном буфере (pH 7,0) и измеряли изменения величины поглощения в зависимости от времени при 240 нм (ε = 39,4). Показатели гликолат-оксидазной и каталазной активности мокрых клеток H. polymorpha P. pastoris
(активированных, культивированных в различных средах варьировали от 20 до 120 М.ед. дихлорфенолиндофенола на 1 г мокрых клеток по гликолат-оксидазе и 30000 до 200000 М.ед. дихлорфенолиндофенола на 1 г мокрых клеток по эндогенной каталазе.
К необязательному, но полезному ингредиенту в реакционном растворе относится флавинмононуклеотид (ФМН), который обычно используют при концентрации до примерно 2,0 мМ, предпочтительно от примерно 0,01 до 0,2 мМ. Полагают, что ФМН повышает производительность гликолат-оксидазы, которая выражается как количество гликолевой кислоты, превращенной в глиоксалевую кислоту на единицу активности фермента. Следует понимать, что концентрация вводимого ФМН относится, кроме того, к любому ФМН, который используют с ферментом, поскольку ФМН часто добавляют к ферменту во время его приготовления. Информацию по структуре ФМН и методике его анализа можно найти у K. Yagai, Methods of Biochemical Analysis, vol. X, InterScience Publishes, New-York, стр. 319-355.
L-молочная кислота является коммерчески доступным материалом. В предлагаемой реакции окисления ее исходная концентрация варьирует в диапазоне от 0,10 до 2,0 М, предпочтительно от 0,25 до 1,0 М. В реакции этот реагент можно использовать в виде кислоты или совместимой ее соли, то есть соли, которая растворяется в воде и катион которой не мешает необходимой биоконверсии L-молочная кислоты в пировиноградную кислоту. Подходящие и совместимые солеобразующие катионные группы можно легко найти экспериментальным путем. В качестве примера таких солей можно указать соли щелочных, щелочноземельных металлов, соли аммония, замещенного аммония, фосфония и замещенного фосфония, L-молочную кислоту, продуцируемую в процессе ферментации можно использовать в качестве субстрата в виде раствора, отфильтрованного непосредственно из ферментера без ее очистки или выделения из сбраживаемой среды.
Конверсию L-молочной кислоты в пировиноградную кислоту удобно и предпочтительно проводить в водной среде. pH реакционной массы поддерживают в интервале 6-10, предпочтительно 7-9. В пределах этого интервала pH, для обеспечения требуемой pH раствора, можно задавать точное ее значение прибавлением любого совместимого, не мешающего реакции основания, включая (но не ограничивая объема) гидроокиси, карбонаты, бикарбонаты и фосфаты щелочных металлов. pH незабуференной реакционной смеси снижается примерно на 2 ед. pH по мере протекания реакции, поэтому часто целесообразно использовать для запуска реакции, достигшей пика максимальной ферментативной активности, диапазон pH примерно от 9,0-8,5, с последующим снижением pH в ходе реакции; конечная величина pH незабуференной реакционной массы меняется в диапазоне примерно от 6,7 до 7,5. Величину pH можно при желании регулировать путем прибавления отдельного любого не мешающего реакции неорганического или органического буфера, который имеет буферную емкость примерно pH 7,5, поскольку оптимальная активность фермента для окисления L-лактата приближается к этому значению; в случае использования подходящего буферного раствора начальная pH составляет 7,5. Следует иметь в виду, что L-молочная и пировиноградная кислоты сильно диссоциированы в воде, и что при pH 7-10 они в большей степени, если не практически полностью присутствуют в виде L-лактат- и пируват-ионов.
Молекулярный кислород (O2), используемый в качестве окислителя в процессе превращения L-молочной кислоты в пировиноградную кислоту, можно вводить в реакцию в виде газа путем интенсивного перемешивания культуральной жидкости на поверхности раздела между газовой и жидкой фазами или через проницаемую для кислорода мембрану. При использовании катализатора из целой клетки с увеличенной проницаемостью мембраны молекулярный кислород можно вводить барботированием кислорода воздуха или кислородсодержащего газа через реакционную смесь. В оптимальных условиях скорость реакции по меньшей мере регулируется частично скоростью, при которой O2 может раствориться в водной среде. Таким образом, хотя O2 можно вводить в реакционную массу в виде воздуха, возможно также использование кислорода в относительно чистой форме. Поскольку не установлено никакого верхнего граничного значения давления кислорода, то можно использовать давление до 50 атм, предпочтительно, чтобы верхнее граничное значение давления составляло примерно 15 атм. Перемешивание играет важное значение для ускорения процесса растворения молекулярного кислорода, необходимого для проведения реакции. Для этого можно использовать любой удобный метод, например механическое перемешивание. Перемешиванием с высокой степенью сдвига или перемешивание с образованием пены может привести к снижению активности растворимого фермента (ов), и поэтому в случае использования катализаторов из растворимых ферментов эти методы следует избегать.
Реакционная температура является важной переменной, поскольку она влияет на скорость реакции окисления и устойчивость ферментов. Обычно можно использовать температурный режим для проведения реакции до примерно 40oC без существенной потери каталитической активности, хотя предпочтительно использование температуры реакции в интервале примерно от 0oC до 15oC. Проведение реакции в предпочтительном температурном интервале обеспечивает максимальную активность восстановленного фермента на заключительной стадии проведения реакции. Не следует проводить реакцию при такой низкой температуре, которая вызвала бы замерзание водного раствора. Температуру реакционной массы можно регулировать традиционными методами, например, но не ограничивая объема, при использовании реактора с охладительной рубашкой, через которую пропускают жидкость с требуемой температурой. Реактор может быть выполнен из любого инертного материала относительно компонентов реакционной массы.
После завершения реакции катализаторы из растворимых ферментов можно удалить фильтрованием или центрифугированием и, при желании, использовать повторно. В альтернативном варианте растворимые биокатализаторы могут быть денатурированы и осаждены путем нагревания, например, до температуры 70oC в течение 5 мин и/или их можно оставить в реакционной массе, если их присутствие не мешает реакции. Проницаемые клеточные биокатализаторы можно отделить от реакционной массы для повторного использования методом центрифугирования или фильтрации. После отделения растворимых ферментов или катализаторов из микробных клеток, флавинмононуклеотид (ФМН) можно, при желании, удалить путем обработки раствора активированным углем. Целевой продукт (то есть пировиноградную кислоту и ее соли) можно затем извлечь непосредственно в виде раствора или полученный раствор можно упарить и выделить пировиноградную кислоту путем удаления оставшейся воды, например, путем отгонки при пониженном давлении, лиофилизацией (сушкой при низкой температуре) или любым другим общеизвестным в данной области методом.
В приводимых ниже примерах, которые служат для дополнительной иллюстрации предлагаемого изобретения, выходы пирувата и ацетата, и выход оставшегося L-лактата приведены в процентах в расчете от общего количества используемой вначале реакции L-молочной кислоты, за исключением особо оговоренных случаев. Анализ реакционной массы проводили методом высокоэффективной жидкостной хроматографии (ВЭЖХ) с использованием колонки Bio-Rad HPX-87H.
Пример 1. В 88,7 см3 стеклянный реакционный сосуд Fisher-Porter аэрозольного типа, оснащенный магнитным сердечником для перемешивания, загружают 10 мл водного раствора, содержащего L-лактат лития (0,75 М), ФМН (0,01 мМ), изомасляную кислоту (ВЭЖХ, внутренний стандарт, 0,100 М), буциновый буфер (0,788 М), гликолат-оксидазу из шпината (1,0 ед.ак/мл), м каталазу из Aspergillus niger (1400 ед.ак./мл), pH 8,9. Реакционный сосуд герметично закрывают, и реакционную массу охлаждают до температуры 5oC. После этого в сосуд закачивают кислород под давлением до 4,9 кг/см2, а затем выпускают до атмосферного давления пять раз при перемешивании. Затем в реакционный сосуд опять нагнетают кислород под давлением до 4,9 кг/см2, и реакционную массу перемешивают при температуре 5oC. Для регулирования процесса развития реакции отбирают аликвоты (0,10 мл) с помощью шприца через отверстие для взятия пробы (не снижая давление в сосуде) через определенные промежутки времени для анализа ВЭЖХ. Через 28,5 ч реакционного процесса выходы пирувата и ацетата по данным ВЭЖХ составляют 47,7 и 43,6% соответственно, и остаток лактата составляет 11,5%. Остаточная активность гликолат-оксидазы и каталазы составляет 40 и 100% соответственно от их исходной активности.
Пример 2. Повторяют методику примера 1, за исключением того, что используют 10 мл водного раствора, содержащего L-молочную кислоту (смесь 96% L-изомера, 4% D-изомера, 0,750 М), KH2PO4 (0,750 М), ФМН (0,01 мМ), изомасляную кислоту (ВЭЖХБ, внутренний стандарт, 0,100 М), гликолат-оксидазу из шпината (1,0 ед.ак/мл), и растворимую каталазу из Aspergillus niger (14000 ед.ак./мл), pH 8,1, температура 5oC. Через 48 ч реакционного процесса выходы пирувата и ацетата по данным ВЭЖХ составляют 79,6 и 3,8% соответственно и лактата остается еще в количестве 20,2%. Остаточная активность гликолат-оксидазы и каталазы составляет 22 и 100% соответственно от их исходной активности.
Пример 3. Повторяют методику примера 1, за исключением того, что используют 10 мл водного раствора, содержащего L-молочную кислоту (смесь 96% L-изомера, 4% D-изомера, 0,500 М), KH2PO4 (0,50 М), ФМН (0,01 мМ), изомасляную кислоту (ВЭЖХБ, внутренний стандарт, 0,100 М), гликолат-оксидазу из шпината (2,0 ед. ак/мл), и растворимую каталазу из Aspergillus niger (14000 ед.ак. /мл), pH 8,3, и реакцию проводят при температуре 5oC. Через 18 ч реакционного процесса выходы пирувата и ацетата по данным ВЭЖХ составляют 90,5 и 4,2% соответственно и остается лактат в количестве 6,4%. Остаточная активность гликолат-оксидазы и каталазы составляет 57 и 100% соответственно от их исходной активности.
Пример 4. Повторяют методику примера 1, за исключением того, что используют 10 мл водного раствора, содержащего L-лактат натрия (0,500 М), изомасляную кислоту (ВЭЖХБ, внутренний стандарт, 0,100 М), гликолат-оксидазу из шпината (2,0 ед.ак/мл), и растворимую каталазу из Aspergillus niger (20000 ед. ак./мл), pH 9,0 (регулируют при добавлении 50% NaOH), и реакцию проводят при температуре 15oC; в отсутствии буфера. Через 7 ч реакционного процесса выходы пирувата и ацетата по данным ВЭЖХ составляют 91,6 и 0,6% соответственно и остается лактат в количестве 7,1%. Остаточная активность гликолат-оксидазы и каталазы составляет 21 и 100% соответственно от их исходной активности.
Пример 5. Повторяют методику примера 1, за исключением того, что используют 10 мл водного раствора, содержащего L-лактат натрия (0,500 М), изомасляную кислоту (ВЭЖХБ, внутренний стандарт, 0,100 М), гликолат-оксидазу из шпината (6,0 ед.ак/мл), и растворимую каталазу из Aspergillus niger (10000 ед. ак./мл), pH 9,0 (регулируют при добавлении 50% NaOH), и реакцию проводят при температуре 15oC; в отсутствии буфера. Через 5 ч реакционного процесса выходы пирувата и ацетата по данным ВЭЖХ составляют 95,3 и 0,9% соответственно и остается лактат в количестве 4,5%. Остаточная активность гликолат-оксидазы и каталазы составляет 68 и 100% соответственно от их исходной активности.
Пример 6. В 88,7 см3 стеклянный реакционный сосуд Fisher-Porter аэрозольного типа, оснащенный магнитным сердечником для перемешивания, загружают 10 мл водного раствора, содержащего L-лактат лития (0,500 М), KH2PO4 (0,50 М), изомасляную кислоту (ВЭЖХБ, внутренний стандарт, 0,100 М) при pH 9,0 (регулируют при добавлении 50% NaOH), и реакционный раствор охлаждают до температуры 5oC. Затем в сосуд добавляют 0,75 клеток-трансформантов Pichia pastoris GS115-MSP10 (с гликолат-оксидазной активностью 65,2 М.ед, и каталазной активностью 101000 М. ед.), которые были обработаны 0,1% бензалконийхлоридом (фирмы Lonza Berquat, OJ-50) для увеличения их проницаемости, затем реакционный сосуд герметично закрывают и реакционную смесь охлаждают до температуры 5oC. Реакционный сосуд герметично закрывают. После этого в сосуд закачивают кислород под давлением до 4,9 кг/см2, а затем выпускают до атмосферного давления пять раз при перемешивании. Затем в реакционный сосуд опять нагнетают кислород под давлением до 4,9 кг/см2, и реакционную массу перемешивают при температуре 5oC. Для регулирования процесса развития реакции отбирают аликвоты (0,10 мл) с помощью шприца через отверстие для взятия пробы (не снижая давление в сосуде) через определенные промежутки времени для анализа ВЭЖХ. Через 5 ч реакции выходы пирувата и ацетата по данным ВЭЖХ составляют 97,6 и 2,55% соответственно и остается 0,3% лактата. Остаточная каталитическая активность клеток по гликолат-оксидазе и каталазе составляет 104 и 105% соответственно от их исходной активности.
Пример 7. Реакцию проводят по методике Примера 7 за исключением того, что бициновый буфер (0,5 М) используют вместо KH2PO4 (0,50 М). Через 5 ч реакционного процесса выходы пирувата и ацетата по данным ВЭЖХ составляют 93,1 и 6,3% соответственно и остается 0,4% лактата. Остаточная каталитическая активность клеток по гликолат-оксидазе и каталазе составляет 107 и 122% соответственно от их исходной активности.
Пример 8. Повторяют методику примера 6, за исключением того, что используют 10 мл водного раствора, содержащего L-лактат натрия (0,500 М) и изомасляную кислоту (ВЭЖХБ, внутренний стандарт, 0,100 М) при pH 9,0 (регулируют при добавлении 50% NaOH), к которому добавляют 0,75 г клеток-трансформантов Pichia pastoris GS115-MSP10 (с гликолат-оксидазной активностью 65,2 М.ед. и каталазной активностью 101000 М.ед.), предварительно обработанные 0,1% бензалконийхлоридом (фирмы Lonza Barquat, OJ-50) для увеличения их проницаемости; не добавляют никакого буфера. Через 5 ч реакции выходы пирувата и ацетата по данным ВЭЖХ составляют 99,0 и 0,7% соответственно и остается 0,4 лактата. Остаточная каталитическая активность клеток по гликолат-оксидазе и каталазе составляет 119 и 113% соответственно от их исходной активности.
Пример 9. Повторяют методику примера 6, за исключением того, что используют 10 мл водного раствора, содержащего L-лактат натрия (0,500 М) и изомасляную кислоту (ВЭЖХБ, внутренний стандарт, 0,100 М) при pH 9,0 (регулируют при добавлении 50% NaOH), к которому добавляют 0,35 г клеток-трансформантов Pichia pastoris GS115-MSP10 (с гликолат-оксидазной активностью 22,6 М.ед. и каталазной активностью 50000 М.ед.), предварительно обработанные 0,1% бензалконийхлоридом (фирмы Lonza Barquat, OJ-50) для увеличения их проницаемости; не добавляют никакого буфера. Через 8 ч реакции выходы пирувата и ацетата по данным ВЭЖХ составляют 97,4 и 2,3% соответственно и остается 0,4% лактата. Остаточная каталитическая активность клеток по гликолат-оксидазе и каталазе составляет 123 и 150% соответственно от их исходной активности.
Пример 10. Повторяют методику Примера 6, за исключением того, что используют 10 мл водного раствора, содержащего L-лактат натрия (0,500 М) и изомасляную кислоту (ВЭЖХБ, внутренний стандарт, 0,100 М) при pH 9,0 (регулируют при добавлении 50% NaOH), к которому добавляют 0,18 г клеток-трансформантов Pichia pastoris GS115-MSP10 (с гликолат-оксидазной активностью 11,3 М.ед. и каталазной активностью 25000 М.ед.), предварительно обработанные 0,1% бензалконийхлоридом (фирмы Lonza Barquat, OJ-50) для увеличения их проницаемости; не добавляют никакого буфера. Через 10 ч реакции выходы пирувата и ацетата по результата ВЭЖХ составляют 92,9 и 5,0% соответственно и остается 3,3% лактата. Остаточная каталитическая активность клеток по гликолат-оксидазе и каталазе составляет 121 и 228% соответственно от их исходной активности.
Пример 11. Повторяют методику примера 6, за исключением того, что используют 10 мл водного раствора, содержащего L-лактат натрия (1,00 М) и изомасляную кислоту (ВЭЖХБ, внутренний стандарт, 0,100 М) при pH 9,0 (регулируют при добавлении 50% NaOH), к которому добавляют 0,71 г клеток-трансформантов Pichia pastoris GS115-MSP10 (с гликолат-оксидазной активностью 45,9 М.ед. и каталазной активностью 100000 М.ед.), предварительно обработанные 0,1% бензалконийхлоридом (фирмы Lonza Barquat, OJ-50) для увеличения их проницаемости; не добавляют никакого буфера. Через 8 ч реакции выходы пирувата и ацетата по результатам ВЭЖХ составляют 89,1 и 8,4% соответственно и остается 1,3% лактата. Остаточная каталитическая активность клеток по гликолат-оксидазе и каталазе составляет 124 и 145% соответственно от их исходной активности.
Пример 12. Повторяют методику примера 6, за исключением того, что используют 10 мл водного раствора, содержащего L-лактат натрия (0,500 М) и изомасляную кислоту (ВЭЖХБ, внутренний стандарт, 0,100 М) при pH 9,0 (регулируют при добавлении 50% NaOH), к которому добавляют 0,66 г клеток-трансформантов Pichia pastoris GS115-MSP10 (с гликолат-оксидазной активностью 62,7 М.ед. и каталазной активностью 100000 М. ед. ), предварительно обработанные 20,1% бензалконийхлоридом (фирмы Lonza Barquat, OJ-50) для увеличения их проницаемости; не добавляют никакого буфера. Реакцию проводят при температуре 15oC и давлении кислорода 4,9 кг/см2. Через 3 ч реакции выходы пирувата и ацетата по результатам ВЭЖХ составляют 98,2 и 1,2% соответственно и остается 0,6% лактата. Остаточная каталитическая активность клеток по гликолат-оксидазе и каталазе составляет 124 и 130% соответственно от их исходной активности.
Пример 13. Повторяют методику примера 12, за исключением того, что используют 10 мл водного раствора, содержащего L-лактат натрия (0,50 М) и изомасляную кислоту (ВЭЖХЮ, внутренний стандарт, 0,100 М) при pH 9,0 (регулируют при добавлении 50% NaOH), к которому добавляют 1,04 г клеток-трансформантов Hansenula polymorpha GO1 (с гликолат-оксидазной активностью 64,7 М. ед. и каталазной активностью 50000 М.ед.), предварительно обработанные 0,1% бензалконийхлоридом (фирмы Lonza Barquat, OJ-50) для увеличения их проницаемости; не добавляют никакого буфера. Реакцию проводят при температуре 15oC и давлении кислорода 4,9 кг/см2. Через 2 ч реакции выходы пирувата и ацетата по результатам ВЭЖХ составляют 97,0 и 2,5% соответственно и остается 0,4% лактата. Остаточная каталитическая активность клеток по гликолат-оксидазе и каталазе составляет 99 и 155% соответственно от их исходной активности.
Пример 14. Реакцию, описанную в примере 13, повторяют при температуре 5oC и давлении кислорода 4,9 кг/см2. Через 4 ч реакции выходы пирувата и ацетата по результатам ВЭЖХ составляют 93,1 и 3,7% соответственно и остается 2,2% лактата. Остаточная каталитическая активность клеток по гликолат-оксидазе и каталазе составляет 66 и 180% соответственно от их исходной активности.
Пример 15. Реакцию, описанную в примере 13, повторяют при температуре 30oC и давлении кислорода 4,9 кг/см2. Через 3 ч реакции выходы пирувата и ацетата по результатам ВЭЖХ составляют 89,9 и 6,5% соответственно и остается 0,6% лактата. Остаточная каталитическая активность клеток по гликолат-оксидазе и каталазе составляет 45 и 140% соответственно от их исходной активности.
Пример 16. В 300-мл реактор EZE-Seal автоклавного типа, оснащенный лопастной мешалкой Dispersimax (фирмы Autoclave Engineers), загружают 100 мл раствора, содержащего L-лактат натрия (5,5 г, 0,50 М). Затем в реактор добавляют 6,70 г клеток-трансформантов Pichia pastoris GS115-MSP10 (с гликолат-оксидазной активностью 670 М.ед, и каталазной активностью 1177000 М.ед. ), которые были обработаны 0,1% бензалконийхлоридом (фирмы Lonza Barquat, OJ-50) для увеличения их проницаемости, и затем полученную реакционную массу доводят до pH 9,0 при добавлении 50% NaOH и охлаждают до температуры 5oC. Реактор продувают кислородом воздуха, после чего реакционную массу перемешивают при скорости мешалки 750 об/мин, при этом барботируют через массу кислород под действием турбинной мешалки при температуре 5oC и давлении кислорода 2,8 кг/см2. Реакционный процесс регулируют путем отбора 0,40 мл аликвотного количества реакционной массы через определенные промежутки времени, фильтрованием отобранной аликвоты через фильтровальную установку Millipore Ultrafree-MC 10,000 NMWL, и фильтрат анализируют методом ВЭЖХ с использованием 0,10 М раствора изомасляной кислоты, который прибавляют к образцу в качестве внутреннего стандарта. Через 3 ч реакции выходы пирувата и ацетата по результатам ВЭЖХ составляют 99,2 и 1,4% соответственно и 0,6% остатка лактата. Остаточная каталитическая активность клеток по гликолат-оксидазе и каталазе составляет 107 и 106% от их исходной активности соответственно.
Реакционную массу центрифугируют для отделения катализатора из активированных клеток, и полученную надосадочную жидкость фильтруют через 0,2 мм нейлоновый фильтр. pH полученного фильтрата доводят до 4,6 путем добавления 1,0 N HCl, а затем раствор замораживают. После удаления воды лиофилизацией получают 5,20 г натриевой соли пировиноградной кислоты (выход конечного продукта 96%, 98% чистота пирувата натрия по результатам ВЖЭХ).
Пример 17. Культуральную жидкость, содержащую 109,9 г/л лактата аммония (смесь 97,8% L-лактата и 2,2% D-лактата), 0,8 г/л ацетата и 2,8 г/л мальтозы центрифугируют для удаления корпускулярного вещества с последующей фильтрацией через 0,45 мм фильтр. Содержание лактата аммония в полученном растворе составляет 1,10 М (118,6 г/л, определенная анализом методом ВЖЭХ). В 300-мл реактор EZE-Seal автоклавного типа, оснащенный лопастной мешалкой Dispersimax (фирмы Autoclave Engineers), загружают 45 мл 1,10 М отфильтрованной культуральной жидкости, а затем добавляют к ней 55 мл дистиллированной воды с получением 100 мл водного раствора, содержащего 0,50 М лактата аммония. Затем в реактор добавляют 6,70 г клеток-трансформантов регенерированного штамма Pichia pastoris GS115-MSP10 (с гликолат-оксидазной активностью 666 М.ед, и каталазной активностью 1107000 М.ед.), которые были обработаны 0,1% бензалконийхлоридом (фирмы Lonza Barquat, OJ-50) для увеличения их проницаемости, и затем полученную реакционную массу доводят до pH 7,5 при добавлении 50% NaOH и охлаждают до температуры 5oC. Реактор продувают кислородом воздуха, после чего реакционную массу перемешивают при скорости мешалки 750 об/мин, при этом барботируют через массу кислород под действием турбинной мешалки при температуре 5oC и давлении кислорода 2,8 кг/см2. Реакционный процесс регулируют путем отбора 0,40 мл аликвотного количества реакционной массы через определенные промежутки времени, фильтрованием отобранной аликвоты через фильтровальную установку Millipore Ultrafree-MC 10,000 NMWL, и фильтрат анализируют методом ВЭЖХ с использованием 0,10 М раствора изомасляной кислоты, который прибавляют к образцу в качестве внутреннего стандарта. Через 3 ч реакции выходы пирувата и ацетата по результатам ВЭЖХ составляют 94,1 (96,2% в расчете на L-лактат) и 2,8% соответственно и остаток лактата составляет 2,5%.
Несмотря на проиллюстрированные и описанные выше конкретные варианты осуществления изобретения, следует понимать, что настоящее изобретение не ограничивается изложенными вариантами или отдельными их деталями, и что возможны различные изменения, не выходящие за пределы нижеследующей формулы изобретения.

Способ включает ферментативную реакцию L-молочной кислоты с кислородом в водном растворе в присутствии биокатализаторов - гликолат-оксидазы (оксидазы (S)-2-гидроксикислот) и каталазы. В качестве биокатализаторов используют растворимые ферменты или микробные клетки-трансформанты, в частности Pichia pastoris NRRL У-21001 или Hansenula polymorpha NRRL У-21065. Растворимые ферменты используют при активности 0,01-1000 МЕ/мл (гликолат-оксидаза) и 50-5000 МЕ/мл (каталаза). Процесс осуществляют при рН 7-9. Предпочтительно используют гликолат-оксидазу с активностью 0,1-10 МЕ/мл, а каталазу с активностью 2000-15000 МЕ/мл при соотношении МЕ/мл каталазы и гликолат-оксидазы по меньшей мере 250:1. Технический результат реализации способа - высокие выходы целевого продукта (96%) с высокой частотой (в виде натриевой соли 98% чистоты). 4 з.п.ф-лы, 1 табл.
3. Способ по п.1, отличающийся тем, что ферментные катализаторы используют в виде микробных клеток-трансформантов штамма Pichia pastories GS 115-MSP 10(NRRL Y-21001) или штамма Hansenula polymorpha GOI (NRRL Y-21065).
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Zelitch et al | |||
| "Oxidation and reduction of glycolic and dlyoxylic acids in plants", J.Biol | |||
| Chem., v.201, 1953, p.707-718 | |||
| N | |||
| Tolbest et al."Products of the oxidation of glycolic acid and l-lactic acid by enzymes from tobacco leaves", J.Biol | |||
| Chem, v.181, 1949, p.905-914 | |||
| B.A.Burdick, Y.R SCHAEFFER | |||
| Biotech | |||
| Lett., v.9, 1987, p.253-258 | |||
| US 4900668 C, 13.02.90. | |||

