Изобретение относится к продуцированию определенных защитных антигенов с применением технологии рекомбинантной ДНК.
Паразиты вызывают широкий спектр заболеваний как у человека, так и у домашних животных. До настоящего времени для лечения этих заболеваний применялись химиотерапевтические методы, однако в последнее время, совсем недавно начали применяться иммунологические методы. Несмотря на то, что паразиты на большинстве стадий своего жизненного цикла представляют собой относительно крупные биологические объекты, на которые воздействует клеточная иммунная защита системы животного-хозяина, они могут проявлять чувствительность к антителам, которые действуют таким образом, что вызывают инактивацию жизненно важных функций паразита. В частности, была показана возможность иммунизировать организм животного-хозяина антигенами, связанными на поверхности кишки паразита, так что генерируемые при этом антитела связываются с антигенами на такой внутренней мембране, когда жидкости тела, содержащие антитела, проглатываются паразитами. Такие антигены могут быть названы "скрытыми антигенами", поскольку они не повышают естественный иммунитет против паразитов.
Мы обнаружили, что особой эффективностью обладает "скрытый антиген", представляющий собой гельминтоцидный антиген H110D, выделенный из Halmonchus contortus, который был описан в WO 088/00835 и 90/11086. В работе WO 93/23542 мы описали дальнейшие наши результаты, связанные с тем, что H110D представляет собой мембраносвязанную аминопептидазу из кишки гельминта. Считается, что этот фермент необходим для превращения белковых питательных компонентов в аминокислоты для поглощения на кишки, при этом его инактивация под действием антител к H110D вызывает гибель паразита от нарушения поглощения питательных веществ. Далее мы обнаружили, что большое число паразитов демонстрирует чувствительность к применяемой в отношении них одной и той же стратегии на основе "скрытого антигена", в том смысле, что ферменты, связанные на мембране кишки, являются необходимыми для поглощения и дальнейшего метаболизма белковых компонентов. Такие ферменты включают аспартилпротеазы (как описано в PCT/GB 93 01521) и тиолпротеазы, в частности катепсин. Эти ферменты локализованы на кишке большого числа паразитов, таких как гельминты, в том числе, различные виды семейств Halmonchus, Ostertagia, Trichostroogylus, Nematodirus, Dictyocaulus, Cooperia, Ascaris, Dirofilaria, Trichuris, Strongylus и Fasciola; и виды членистоногих, особенно представители класса паукообразных и насекомых, и в особенности эктопаразиты, такие как кровососущие насекомые (в том числе представители двух групп насекомых, как с неполным, так и с полным циклом превращения), и двукрылые, такие, как журчалки (Lucilia), миязные мухи (myiais jlies), слепняки, вши, клещи, блохи, овечьи рунцы и клопы.
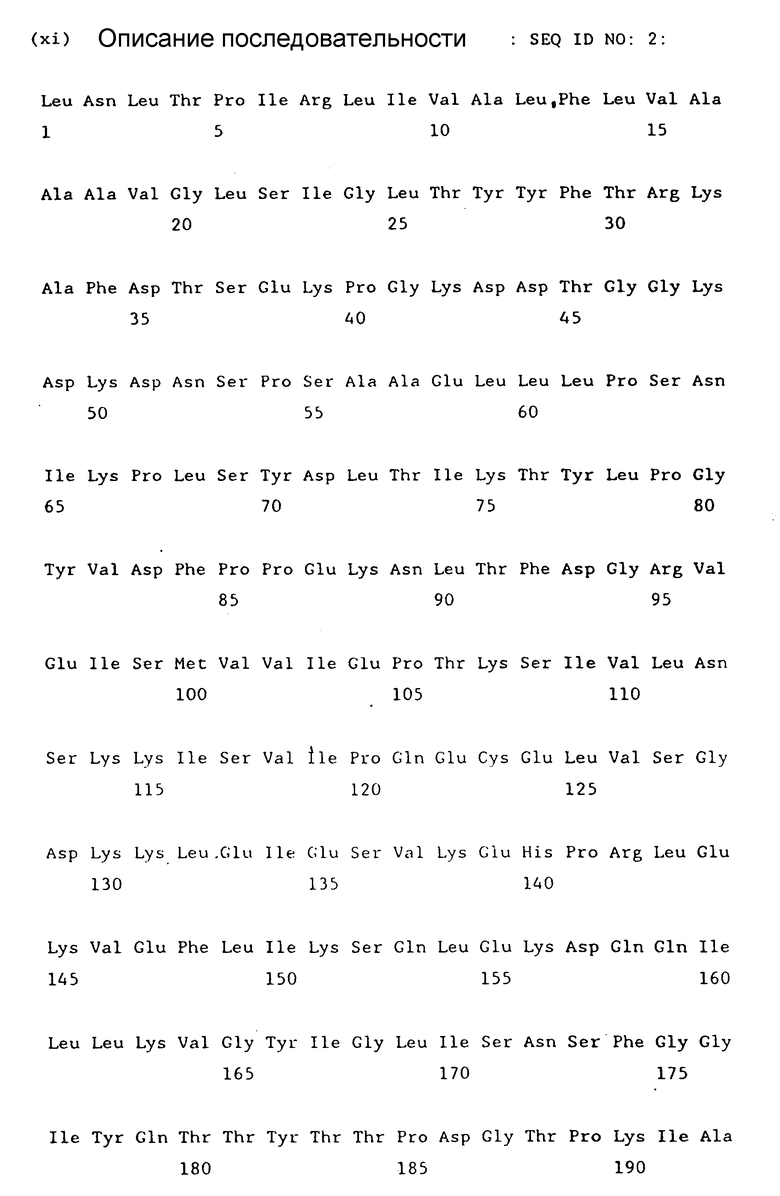
В приведенной выше работе WO 93/23542 также описывается продуцирование фрагментов антигена H110D с применением технологии рекомбинантной ДНК. В этой работе материал H110D был получен в результате экспрессии в E.coli с использованием pGEX вектора, инъецируемого в овец, что приводило к повышению уровня антител к H110D. В дальнейшей работе мы использовали бакуловирусы в клетках насекомых (Sf 9 клетки), при этом удалось осуществить экспрессию клопа гена H110D размером 3,5 rb. Нуклеотидная последовательность его приведена на фиг. 1 (seg. 1D NO:1). Соответствующий продукт трансляции показан на фиг. 2 (seg. 1D NO:2).
Однако для продуцирования вакцин с целью их использования на животных предпочтительно проводить экспрессию антигена в клетках млекопитающих. Такая система характеризуется лучшими показателями репродукции нативной формы и защитных эпитопов антигена, поскольку система экспрессии эукариотов позволяет осуществлять значительно более близкий к нужному процесс гликозилирования, образования дисульфидных связей и других пост-трансляционных модификаций, нежели это имеет место в E.coli, в которой образуется в результате экспрессии нерастворимый белок требующий складчатого реструктурирования, при этом указанная система E. coli характеризуется низким уровнем репродукции нативной формы.
Кроме того, гликозилирование в системе млекопитающих, по-видимому, не индуцирует иммунную реакцию, отвлекающую ответную функцию антипротеинов, а именно эта ситуация имеет место в материале, полученном с применением клеточной линии насекомых, которая объясняется различным характером гликозилирования. Для защиты человека и домашних животных предпочтительно в этой связи использовать клеточные линии фибробластов или миеломных клеток человека или животных, таких, как Hela - клеточная линия человека; ВНК - почечные клетки детенышей хомячков; VERO и COS - клеточная линия почек обезьян; FR3T3 - крысиные фибробласты Фишера; NIH3T3 - клеточная линия фибробластов мышей; C1271 - клеточная линия опухоли молочной железы мышей; CV - 1 фибробласты почек Африканской зеленой обезьяны; 3T6 - эмбриональные фибробласты мышей; L - клетки - клеточная линия мышей; CHO - клеточная линия яичников китайского хомячка; NSO NSI, SP2 и другие клеточные миеломные линии мышей и клеточные линии миеломы крыс, такие, как YB 2/0 и Y3.
Поскольку продуцируемые скрытые антигены жизненно важны для процесса поглощения и усвоения паразитом питательных веществ, ферменты и другие функциональные белки, имеющие одинаковую активность, являются общими для большого количества доступных для изучения эукариотных клеточных линий и они не только мешают селекции клопов, продуцирующих нужный антиген, но и создают также дополнительные трудности при выделении искомого антигена из клеточных продуктов и дальнейшей очистке. Кроме того, выбранная клеточная линия хозяина должна быть непременно генетически близкой с точки зрения белковой последовательности к тому животному, которое служит объектом защиты, так что контаминация нужного чужеродного скрытого антигена таким эндогенным антигеном скорее всего приведет к нежелательной аутоиммунной реакции хозяина. Вдобавок к этому, значительно усложняются контрольные исследования качества экспрессируемого "скрытого антигена".
Изобретение основано на той концепции, что рекомбинантная ДНК осуществляет экспрессию искомого чужеродного энзиматически активного "скрытого антигена" в трансформированной клеточной линии хозяина-млекопитающего, которая в тех случаях, когда оба фермента либо связаны с клеточной мембраной, либо оба локализованы в цитоплазме, что одинаково мешает их физико-химическому разделению, по существу свободна от эндогенных антигенов, имеющих те же самые энзиматические функции, что и чужеродный "скрытый антиген".
В соответствии с настоящим изобретением, мы предлагаем способ экспрессии фермента-антигена, который в естественном состоянии представляет собой фермент, связанный с мембраной кишки паразита, или его фрагмент, обладающий сходной энзиматической и/или антигенной активностью, при этом в хозяйские клетки вводится вектор, адаптированный к экспрессии упомянутого фермента-антигена или его фрагмента, характерным для этого случая является тот факт, что перед трансформацией хозяйские клетки по существу не содержат эндогенных ферментов, выполняющих (а) ту же самую функцию и характеризующихся (б) наличием такой же интеграции с клеточной мембраной или отсутствием такой интеграции, как и у упомянутого фермента-антигена паразита или его фрагмента.
Таким образом, если хозяйская клетка содержит в цитоплазме эндогенный фермент, такой как аминопептидаза, и поскольку чужеродный фермент экспрессируется с транс-мембранной последовательностью и локализуется на мембране хозяйской клетки, не представляет трудностей проведение разделения эндогенного и чужеродного ферментов посредством процесса, приводящего к отделению мембранных фрагментов, т.е. посредством центрифугирования. С другой стороны, чужеродный антиген может быть модифицирован с целью продуцирования фрагмента, в котором отсутствует участок, необходимый для связывания с мембраной, и если мембрана хозяйской клетки несет эндогенный фермент, имеющий ту же самую активность, что и чужеродный фрагмент, представляется также возможным воздействовать на разделение посредством удаления материала клеточной мембраны; таким образом, кодирующая последовательность для трансмембранного региона фрагмента паразита может быть замещена в экспрессируемом гене сигнальной последовательностью, которая оказывает воздействие на секрецию.
Особый интерес предъявляют антигены аминопептидазы гельминтов, такие как приведенный выше H110D и его фрагменты. Такие антигены и их фрагменты могут экспрессироваться в широком диапазоне клеток млекопитающих с использованием соответствующих векторов. Для экспрессии антигена H110D H. contortus, который демонстрирует преимущественно аминопептидазную активность А-типа или М-типа и способен по этой причине расщеплять метиониновую и лейциновую пептидные связи, мы показали возможность использования COS-I клеток в соответствии с настоящим изобретением, поскольку у них отсутствует значимая аминопептидазная активность А-типа и М-типа. Похоже, что они обладают аминопептидазной активностью, расщепляющей аланиновые пептидные связи, которая в слабой степени ассоциирована с клеточной мембраной, так что в этом случае не возникает проблем при разделении эндогенного фермента и экспрессированного фермента H110D, который локализуется на клеточной мембране.
Было показано, что протеолитические ферменты, такие, как трипсин, расщепляют на мембранный паразитарный антиген H110D с продуцированием растворимого H110D (H11S). Такие ферменты могут использоваться для выборочного расщепления чужеродного "скрытого антигена" и отделения его таким образом от эндогенного фермента со сходной активностью. Подходящие клеточные линии для использования в соответствии с настоящим изобретением могут быть либо отобраны из существующих штаммов посредством скрининга по признаку наружной энзиматической активности и/или по признаку локализации любого релевантного фермента либо на клеточной мембране, либо в цитоплазме. В последнем случае связь с клеточной мембраной может быть установлена посредством экстрагирования лизированных клеток вначале таким детергентом, как твин, который не экстрагирует интегральные мембранные ферменты, а затем, например, тритоном, который высвобождает такие ферменты.
Представляется также возможным создать клеточную линию млекопитающих с низким содержанием интересующего фермента. Один из таких способов может включать следующие процедуры: повышают уровень антител к тому ферменту млекопитающих, который должен быть удален. Затем клеточные линии подвергают модификации - облучением или химическими реагентами - с целью индукции точечных мутаций. Клетки культивируют в среде, подходящим образом подобранной, так чтобы компенсировать потерю энзиматической активности. После этого клетки обрабатывают мечеными флюоресцентными антителами и пропускают через лазерный анализатор (сортировщик) клеток по интенсивности флюоресценции (FACS). Клетки, не обладающие флюоресценцией, клонируют, из них повторно отбирают клетки, лишенные упомянутого фермента, при этом осуществляют постоянный мониторинг стабильности такой потери.
Клетки могут быть также модифицированы посредством делеции в гене или рациональной (направленной) техники внесения мутаций для удаления или мутирования гена, ответственного за синтез релевантного эндогенного фермента.
В технике хорошо известны векторы, подходящие для введения их в различные классы клеточных линий млекопитающих. В общем, они должны включать промотор и/или усилитель, оперативно связанный с геном, ответственным за экспрессию фермента-антигена или его фрагмента. Таким образом, в частности, фрагмент гена для H110D размером 3,5 kb может быть связан в рамке считывания с соответствующим промотором. Подходящие промоторы включают ранний или поздний промотор SV-40, т.е. PSVL вектор, промотор цитомегаловируса (CMV), промотор металлотионеина I мышей и терминальную последовательность вируса опухоли молочной железы мышей. Предпочтительно, чтобы вектор включал подходящий маркер, такой, как ген, кодирующий дигидрофолатредуктазу или ген, кодирующий глутаминсинтетазу. Векторы указанных типов описаны в работах WO 86/05807, WO 87/04462, WO 89/01036 и WO 89/10404.
Уровень трансфекции хозяйских клеток может быть усилен в рамках стандартных технологий, использующих, например, фосфат кальция, ДЭАЭ-декстран, полибрен, метод слияния протопластов, липосомы, направленную микроинъекцию, бомбардирование гена или электропорацию. Последний метод предпочтителен, при этом способы трансфекции клеточных линий, используеющих электропорацию в литературе (Andreason G. L. и Evans G.A., Introduction and Expression of DNA molecules in encaryotic cells by electroporation, Biotechnigues, 6, 65, 1980). В общем случае, линейная ДНК вводится в клетки легче, чем кольцевая ДНК. Антиген H110D демонстрирует лейцинаминопептидазную (М-типа) и метионинаминопептидазную активность (А-типа) и в целом предпочтительно, чтобы хозяйские клетки не содержали хотя бы эти два типа аминопептидазной активности. Следующие примеры даны только лишь с иллюстративной целью. В этих примерах представлено следующее:
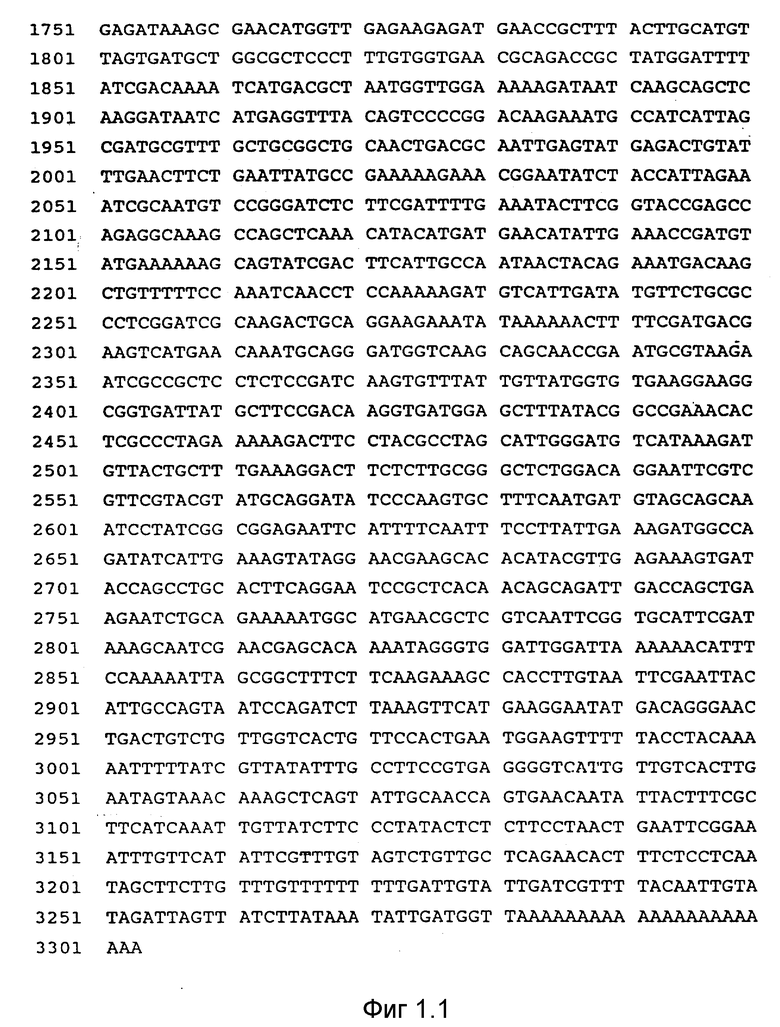
на фиг. 1 и фиг.1.1 показана последовательность ДНК размером 3,5 kb для полимеразной цепной реакции (ППР), клон 2 (последовательность ID NO:1);
на фиг. 2 показан аминокислотный состав продукта трансляции клона 2 размером 3,5 kb для полимеразной цепной реакции (последовательность ID NO:2).
Пример 1. Определение ферментативной активности в COS-I клетках.
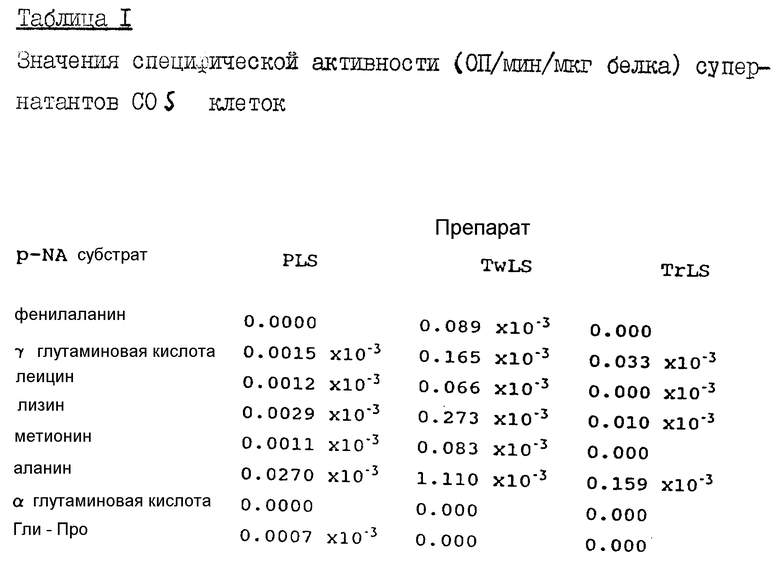
COS-I клетки были получены из Университета в Сурее (Surrey University) в 5 мл ростовой среды CUMD (DMEM). Клетки были разделены на две 200-мл колбы для выращивания культуры, при этом каждая содержала 11 мл среды, в которой клетки росли вплоть до слипания. COS клетки слипаются, так что с поверхности колбы их можно удалить аспирацией, используя для этого стеклянную пастеровскую пипетку с 10 мл PBS буфера. Когда суспензия будет содержать все клетки, ее переносят в обычную пробирку и замораживают при температуре -20oC. Замороженные клетки оттаивают и замораживают несколько раз в жидком азоте для разрушения клеток, после чего их откручивают для отделения PBS супернатанта (PLS). Для последующих экстракций используют 500 мкл PBS с 0,1% Твина и PBS с 2% Тритона с получением соответственно TwLS и TrLS супернатантов. Все супернатанты концентрировали до объема 200 мкл с использованием Миллипоровских микроконцентраторов. Экстракты на основе только PBS содержат ферменты, которые находились в свободном состоянии в цитоплазме, экстракты на основе Твина содержат ферменты, которые были слабо связаны с клеточной мембраной, а экстракты на основе Тритона содержат белки, интегрированные с мембраной.
Супернатанты исследовали против 25 мМ: пНА (pNA) (п-нитроанилидных) субстратов, представленных фенилаланином, гамма-глутаминовой кислотой, лейцином, лизином, метионином, аланином, альфа-глутаминовой кислотой, Гли-Про и аспарагиновой кислотой, в HEPES бикарбонатном буфере при pH 7,0. После 30 минут инкубации при 37oC не было обнаружено явной ферментативной активности, для оценки которой срок инкубации был увеличен до 18 часов. Были подсчитаны значения удельной активности, результаты представлены в таблице 1.
Наибольшая активность была получена в экстракте на основании Твина, при этом самая высокая активность была показана для аланинового pHA субстрата, некоторая активность была также отмечена относительно фенилаланинового, на основе гамма-глутаминой кислоты, лейцинового, лизинового и метионинового pHA субстратов. PLS и TrLS обладали невысокой акивностью, по-видимому, некоторым уровнем активности гамма-ГТФ и остаточной активности лизиновой AP и аланиновой AP.
10 мкл каждого супернатанта COS-I клеток добавляют к 25 мМ п-нитроанилидного субстрата в 250 мкл бикарбонатного HEPES буфера при pH 7,0 и инкубируют при 37oC в течение 18 часов.
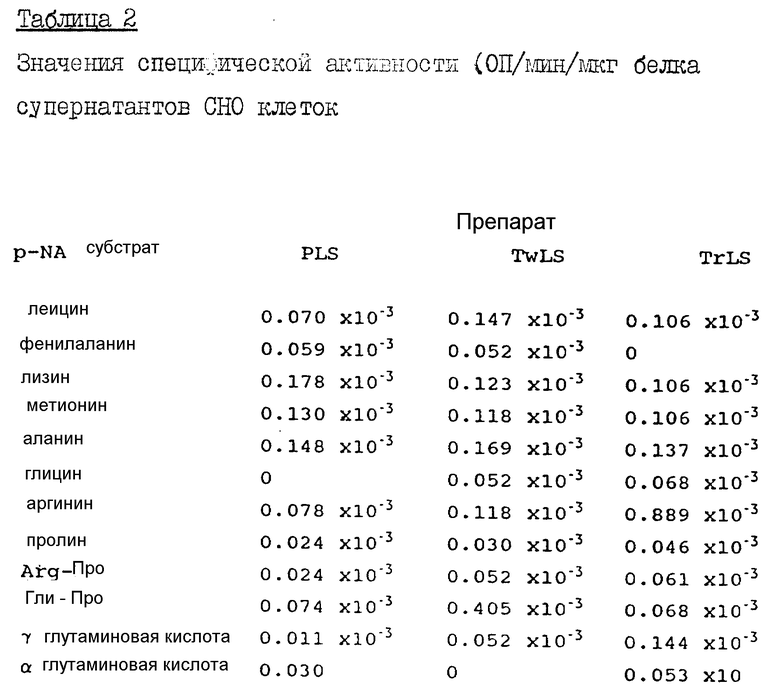
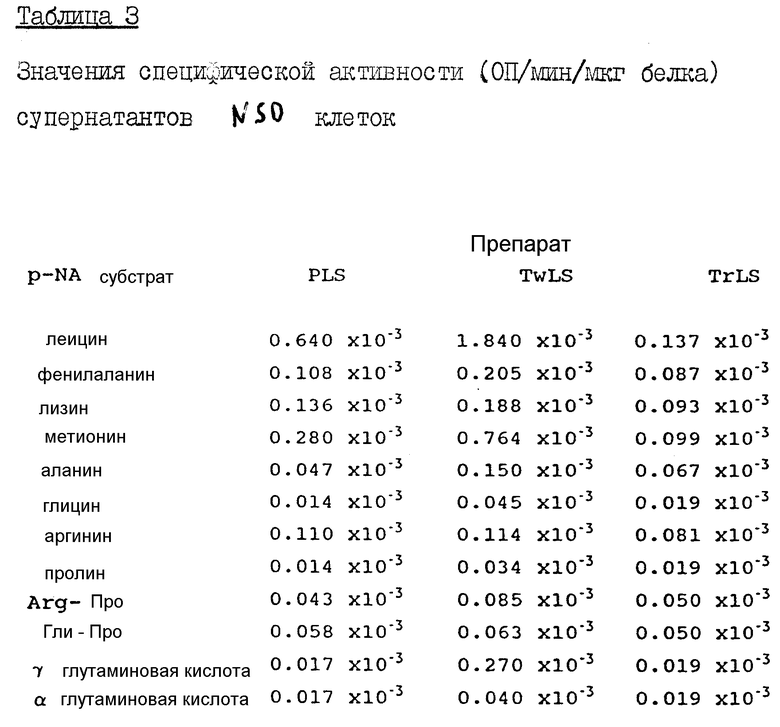
Определение ферментативной активности в клетках яичников китайского хомячка (CHO) и NSO клетках.
Экстракты культивированных CHO и NSO клеток готовят способом, аналогичным тому, что был описан выше в разделе, посвященном COS-I клеткам. Указанные экстракты анализировали против следующих паранитроаналидных субстратов: аланина, аргинина, глицина, альфа-глутаминовой кислоты, гамма-глутаминовой кислоты, лейцина, лизина, метианина, фенилаланина, пролина и Гли-про - все субстраты берутся в количестве 25 мМ в бикарбонатном HEPES буфере. Инкубация проводится при 37oC в течение 30 минут, после чего записываются конечные значения ОП, на основе которых вычисляются удельные активности фермента.
Из табл. 2 видно, что экстракты растворимых и мембраносвязанных компонентов CHO клеток имеют низкий уровень энзиматической активности, за исключением случая определения активности в TwLS против Гли-Про субстрата. А TrLS экстракт демонстрирует низкую активность во всех случаях, за исключением использования аргининового субстрата. Последняя активность отличается от аминопептидазной активности H110D.
Было показано, что как PLS, так и TwLS экстракты NSO клеток обладают аминопептидазной активностью, особенно высока она против лейцина и метионина (табл. 3). В отличие от них, TrLS экстракт имеет низкую активность, что позволяет предположить, что процесс очистки, включающий экстракцию тритоном, которая и применялась в случае с H110D, приводит к достижению очень низкого уровня загрязнения такого экстракта эндогенной энзиматической активностью
Сравнительные примеры.
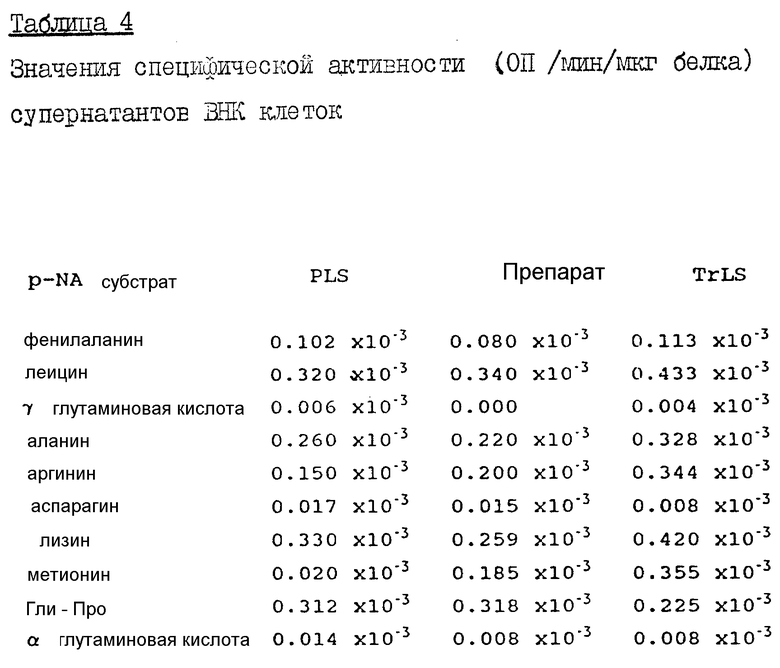
ВНИК клетки получают с помощью такого же метода, что и COS-I клетки, описанные в примере 1, правда, в большем количестве (2 х 1 л круглые бутыли со слипшимися клетками в 100 мл среды). Супернатанты исследовали против: фенилаланинового, лейцинового, на основе гамма-глутаминовой кислоты, аланинового, аргининового, на основе аспарагиновой кислоты, лизинового, метионинового, Гли-Про и на основе альфа-глутаминовой кислоты пНА субстартов при pH 7,0. Тестирование проводили в условиях инкубации при 37oC в течение 30 минут с регистрацией конечного значения ОП.
Экстракты ВНК клеток содержат значительную энзиматическую активность, которая обнаруживается во всех супернатантах (табл. 4). Незначительная активность отмечается лишь к таким субстратам, как гамма-глутаминовая кислота, аспарагиновая кислота или альфа-глутаминовая кислота. Для всех супернатантов была отмечена высокая активность к лизиновому пНА субстрату (которая была наибольшей в PLS супернатанте), лейцину, аланину и Гли-Про. Активность относительно метионинового пНА субстрата, которая была незначительной в PLS супернатанте, была минимальной в TrLS супернатанте. Видно, что такие ВНК клетки не подходят для экспресии H110D, поскольку высокий уровень аминопептидазной активности А-типа и М-типа был найден как в цитоплазме, так и в связанной с мембраной интегрированной форме.
К 10 мкл каждого супернатанта ВНК клеток добавляют 25 мМ п-нитроанилидного субстрата в 250 мкл бикарбонатного HEPES буфера при pH 7,0 и инкубируют полученную смесь при pH 7,0 в течение 30 минут при 37oC.
Пример 2. Клонирование HIIOD последовательности вектора экспрессии млекопитающих.
ДНК, которую подвергают клонированию, и которая представляет собой клон 2 ПЦР (полимеразной цепной реакции) гена H11OD размером 3,5 kb была описана в работе WO 93/23542. Последовательность ДНК (Последовательность ID NO: 1) такой вставки), полученной в ходе полимеразной цепной реакции (ПЦР), показана на фиг. 1, а аминокислотная последовательность продукта трансляции (Последовательность ID NO: 2) на фиг. 2. Эта ДНК была вырезана из вектора pT7Blue-T вектора (Novagene) посредством расщепления BamH1 и клонирования на BamH1 сайте множественно клонируемого сайта вектора pSPT18 (Берингер Маннгейм) с получением клона pSPT18-3,5-2. Было проведено частичное расщепление с помощью BamH1 клона pSPT18-3,5-2, а полученная при этом линейная ДНК была дважды подвергнута очистке с помощью гель-электрофореза. "Липкие" концы были "затуплены" посредством введения дНТФ с помощью фермента Кленова (большой фрагмент ДНК-полимеразы), а с помощью Ncol линкера, содержащего АТГ (Берингер Мангейм Кат N 1171160), тупые концы были соединены с получением плазмиды. Клоны подвергли скринингу с применением рестрикционного анализа по признаку наличия указанного линкера на 5' конце вставки размером 2,5 kb, что давало внутрирамочный сайт инициации АТГ под контролем промотора T7. Модифицированная вставка размером 3,5 kb из одного такого клона (клон pST18-3,5-2 N 44) была вырезана и затем субклонирована в векторе экспрессии млекопитающих pRC/CMV с использованием следующей стратегии:
1. ДНК клона pSPT18 (T7-3,5-2 N 44) расщепляют с помощью рестрикционной эндонуклеазы Smal.
2. Notl линкер соединяют с этим модифицированным сайтом Smal, что дает возможность появлению клонов с Notl сайтом на 5' конце вставки, который предшествует Ncol линкеру, содержащему сайт старта считывания АТГ.
3. Очищенную ДНК из подходящего клона расщепляют с помощью как Notl, так и Xbal с высвобождением вставки размером 3,5 kb.
К инкубационной среде добавляют фермент Pvul для того, чтобы разрезать пополам ДНК вектора pSPT18, что необходимо, поскольку близкие размеры вектора и вставки затрудняют процесс очистки.
4. Вставку размером 3,5 чистят в 0,6-0,7% агарозном геле.
5. Вектор экспрессии млекопитающих pRC/CMV расщепляют с помощью Notl и Xbal, а полосу, соответствующую линейной плазмиде, чистят в агарозном геле.
6. Проводят лигирование вставки размером 3,5 kb и переведенного в линейную форму вектора.
7. Отбирают соответствующие клоны, которые имеют размер 3,5 kb при расщеплении с помощью Notl и Xbal. Эти клоны обозначают как pRC/CMV-3,5-2.
Трансфекция COS-1 клеток млекопитающих.
ДНК вектора экспрессии млекопитающих со вставленным в него клоном антигена HIIOD, обозначенным как pRC/CMV-3,5-2, подвергают высокой степени очистки путем центрифугирования в градиенте хлористого цезия. (Sambrook J., Fritch E.F. и Maniatis T. Molecular Cloning, A Laboratory Manual, Second edition, Cold Spring Harbar Press, 1989).
Временной экспрессии HIIOD можно достичь используя эту очищенную ДНК клона pRC/CMV-3,5-2 для трансфекции COS-1 клеток (полученных от ECACC, Портон). Трансфекцию проводят с использованием ДЭАЭ-декстрана (Cullen B.R., Use of Eucaryotic expression technology in the functional analysis of cloned genes, Metods in Enzymology: Guide to molecular cloning techniques, Eds S.L. Berger и A.K. Kimmal, Academic Press, 1987, pp 684-704). Клетки культивируют на среде Игла в модификации Дульбекко (DMEM) [Гибко БРЛ (Gibco BRL)], содержащей 10% эмбриональную сыворотку теленка (FCS) [Гибко БРЛ (Gibco BRL)] и через 48-72 часа с момента начала инкубации анализируют продукт экспрессии.
Трансфекция CHO клеток млекопитающих (клеток яичника китайского хомячка).
Векторную ДНК вводят в CHO клетки (полученные из ECACC, Портон) с использованием кальцийфосфатного метода (такого, в частности, как метод, описанный в работе Cullen B.R Use of eucaryotic compression technology in the functional analysis of cloned genes. Methods in Enzymology: Guide to Molecular Cloning Techniques, Eds S.L. Berger и A.R. Kimmel, Academic Press, 187, pp. 684-704). Трансформированные клетки культивируют в DMEM среде, содержащей 10% эмбриональной сыворотки теленка (Гибко БРЛ). Генетицин-устойчивая линия (G418) представляет собой единственную трансформированную клеточную линию, в отличие от нетрансформированных клеток, хорошо растущую, клетки выращивают в культуре до концентрации 800 мкг/мл. Затем трансформированные клетки клонируют с помощью метода ограниченных разведений на микротитровальных плато.
Анализ клеток млекопитающих, подвергнутых трансфекции.
Трансформированные и нетрансформированные CHO клетки могут быть перенесены для роста на покровное стекло с целью проведения иммунофлюоресцентного анализа. Клеткам позволяют вырасти до образования небольших колоний, фиксируют их метанолом и обрабатывают овечьей антисывороткой к HIIOD и затем флюоресцентным красителем (т. е. FTCC), конъюгированным с иммуноглобулином овечьей антисыворотки. Для поиска положительных колоний используют флюоресцентный микроскоп.
Клетки в трансформированных клеточных линиях разрушают в RIPA буфере (150 мМ хлорида натрия, 1% Нонидета П40 [Nonidet P40], 0,5% дезоксихолата, 0,1% додецилсульфата натрия, 50 мМ Трис-HCl, pH 8,0) при удалении от массы клеток ростовой среды и затем посредством осторожного растирания клеток в течение 5 минут в RIPA буфере. Затем клетки переносят в пробирки для микроцентрифугирования и откручивают их в микроцентрифуге при полной скорости в течение 15 минут с получением свежего лизата, который переносят в чистую пробирку. Аликвоты этого лизата, содержащие 2•105 клеток, подвергают электрофорезу в SDS полиакриламидном геле (SDS-PAGE), а белки из геля переносят на нитроцеллюлозную мембрану посредством вестерн-блоттинга. Мембрану обрабатывают, возможно, с включением периодатной стадии обработки, и анализируют ее против антисывороток, полученных к различным формам HIIOD антигена. Периодатная обработка разрушает углеводы эпитопа; углеводы эпитопов млекопитающих могут значительно отличаться от нативных углеводов гельминтов. Фрагменты, полученные при вестер-блоттинге трансформированных клеток, показывают наличие белка, распознаваемого антисыворотками, специфичными к H11OD.
Экстракты трансформированных и нетрансформированных клеток готовят как описано в примере 1, а лизаты, полученные с использованием RIPA, анализируют на наличие энзиматической активности в точности так, как было описано в примере 1. Трансформированные клетки, содержащие HIIOD, демонстрируют более высокий уровень аминопептидазной активности, чем нетрансформированные клетки.
Наличие введенной векторной ДНК, которая содержит клон 2 ПЦР размером 3,5 kb, устойчивой к генетицину клеточной линии CHO, определяют с помощью саузерн-блоттинга в препаратах ДНК, полученных из этих клеточных линий. ДНК экстрагируют из клеток с помощью известного метода (Sambrook, J., Fritsch, E. F. и Maniatis, T. Molecular Cloning, A Laboratory Manual, Second Ed. Cold Spring Harbor Press, 1989). 10-20 мкг ДНК расщепляют с помощью рестрикционных эндонуклеаз и затем проводят электрофоретическое разделение в агарозном геле, перенося ДНК на мембрану с помощью саузерн-блоттинга (Southern, E. Detection of specific sequences among DNA fragments separated by gel electrophoreses, J. Mol. Biol., 1975, 98, p. 503). Эту мембрану гибридизуют с пробой, кодирующей клон 2 ПЦР размером 3,5 kb, которую после тщательного промывания подвергают ауторадиографии. В трансформированных клетках обнаруживают полосы, специфичные для клона 2 ПЦР размером 3,5 kb.
Экспрессия клона 2 ПЦР размером 3,5 kb на РНК уровне в трансформированных клетках млекопитающих определяется с помощью нозерн-анализа РНК, выделенной из этих клеток. РНК экстрагируют из клеток с использованием РНКзола (RNAzol) [Цинна Биотекс, Техас (Cinna (Biotecx, Texas)], снабженного инструкциями производителя, упомянутую РНК в количестве до 20 мкг разгоняют в агарозном геле, после чего переносят на мембрану с помощью нозерн-блоттинга (Sambrook, J., Fritsch, E.F. и Maniatis, T. Molecular Cloning, A Laboratory Manual, Second Ed. Cold Spring Harbor Press, 1989). Эту мембрану затем гибридизируют с пробой, кодирующей клон 2 ПЦР размером 3,5 kb, которую после тщательного промывания исследуют методом ауторадиографии. В трансформированных клетках отмечаются специфические гибридизационные полосы, соответствующие клону 2 ПЦР размером 3,5 kb.












Изобретение относится к биотехнологии и генной инженерии. Способ позволяет проводить экспрессию антигенов, интегрированных с мембраной аминопептидаз гельминтов. Проводят трансфекцию хозяйских клеток млекопитающих вектором. Вектор адаптирован для экспрессии аминопептидазного антигена Н110D или его фрагмента. Перед трансфекцией хозяйские клетки освобождают от эндогенных ферментов, обладающих той же самой функцией и такой же интеграцией с клеточной мембраной или ее отсутствием, как и аминопептидазный антиген Н110D паразита или его фрагмент. Хозяйская клетка представляет собой СOS-1 или СНO клетки. Аминопептидазный антиген Н110D или его фрагмент экспрессируются интегрированными с клеточной мембраной хозяйской клетки и/или в виде растворимого цитоплазматического фермента. 4 з.п. ф-лы, 2 ил., 4 табл.
(а) той же самой функцией и
(b) такой же интеграцией с клеточной мембраной или ее отсутствием, как и аминопептидазный антиген Н11ОД паразита или его фрагмент.
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| WO 9011086 A, 1990 | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Домовый номерной фонарь, служащий одновременно для указания названия улицы и номера дома и для освещения прилежащего участка улицы | 1917 |
|
SU93A1 |
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| WO 8800835 A, 1988 | |||
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |
| Установка для вспучивания шлакового расплава | 1975 |
|
SU524834A1 |
| Кипятильник для воды | 1921 |
|
SU5A1 |
| Методы генетической инженерии Т.Маниатис и др | |||
| Молекулярное клонирование | |||
| - М.: Мир, 1984, с | |||
| Способ получения гидроцеллюлозы | 1920 |
|
SU359A1 |
