Эта заявка является частичным продолжением заявки на патент США в с серийным номером 07/748562, зарегистрированной 22 августа 1991 г.
Настоящее изобретение относится, в основном, к материалам и методам, применимым при предупреждении (профилактике) и лечении инфекции, вызываемой вирусом иммунодефицита человека (ВИЧ-I). Более конкретно, изобретение относится к моноклональным антителам, пригодным для пассивной иммунизации животных, восприимчивых к/или инфицированных ВИЧ-I, особенно людей.
Инфекционный процесс, вызванный ВИЧ-I in vivo, недавно был темой обзорной статьи Me Cune, Cell, 64, pp 351-363 (1991). Кратко, ВИЧ-I инфицирует ряд последовательных клеточных поколений, таких как Т-клетки, моноциты/макрофаги и нейроны, которые обладают выраженным рецептором CD4, поскольку подавляющее большинство клеток CD4 + в организме являются "отдыхающими" или находятся в состоянии покоя и делятся только в ответ на специфические сигналы, в результате инфекции ВИЧ-I CD4 + - клетки включают транскрипционно неактивный вирус. Стимуляция иммунной системы инфицированных животных, включая активную иммунизацию, может приводить к поликлональной активации и сигналу для остальных клеток CD4 +, перейти в фазу клеточного цикла. Реплицирующиеся клетки затем активно продуцируют вирусные частицы, вызывая распространение инфекции. Принимая во внимание этот отрицательный эффект стимуляции иммунной системы животных, инфицированных ВИЧ-I, возможно, что наиболее эффективный метод предупреждения или лечения инфекции ВИЧ-I это пассивная иммунизация, которая заключается во введении антител анти-ВИЧ-I восприимчивым или инфицированным животным.
Jackson et al., Lancet, 2, pp. 647-652 (1988) сообщает, что однократное введение антител анти-ВИЧ-I в виде плазмы больным людям, страдающим развитым синдромом приобретенного иммунодефицита (СПИД, синдром прогрессирующей деградации иммунной системы, связанной с инфекцией ВИЧ-I), временно приводит к снижению симптомов, преходящему увеличению числа Т-лимфоцитов, снижению частоты инфекций, вызванных условно-патогенными микроорганизмами, и снижению частоты, с которой ВИЧ-I мог бы быть выделен из плазмы или лимфоцитов больных. Смотрите также, Karpas et al. , Proc. Natl. Acad. Sci. USA 85, pp. 9234-9237 (1988). Кроме того, Emini et al., Nature 355, pp. 728-730 (1992) сообщает, что введение антител, специфически реагирующих с ВИЧ-I, шимпанзе перед заражением ВИЧ-I приводит к тому, что у животного не проявляются признаки вирусной инфекции, эти исследования показывают, что антитела, способные к нейтрализации ВИЧ-I, могут быть полезны при профилактике лечении инфекции ВИЧ-I.
Главный гликопротеин внешней оболочки ВИЧ-I, gp 120, связывается с клеточным рецептором CD 4 и облегчает внедрение вируса. Несколько эпитопов гликопротеина могут быть связаны с получением нейтрализующих антител. Но et al., Science 239, pp. 1021-1023 (1988) сообщает, что аминокислоты 254-274 из gp. 120 создают основу поликлональной антисыворотки, способной к группоспецифической нейтрализации трех различных штаммов ВИЧ-I. Структурозависимые эпитопы, эпитопы, не состоящие из первичных последовательностей аминокислот на gp 120, также включены в выработку антител, которые нейтрализуют разные штаммы вируса, согласно Haigwood et al., Vaccins 90, pp. 313-320 (1990) и Ho et al., J. Virol 65(1), pp. 489-493 (1991). Так называемая "главная нейтрализующая детерминанта"/ (PND) gp 120 ВИЧ-I локализована на "V3 петле" gp 120. Смотрите Putney et al., Science, 234, pp. 1392-1395 (1986), Rusche et al., Proc. Natl. Acad. Sci. USA, 85, pp. 3198- 3202 (1988); Goudsmit et al., Proc. Natl. Acad. Sci, USA 85, pp. 4478-4482 (1988); Palker et al., Proc. Natl. Acad. Sci, USA 85, pp. 1932-1936 (1988); Holley et al., Proc. Natl. Acad. Sci, USA 85, pp. 6800-6804 (1991). V3 петля состоит из гипервариабельной области, которая образуется с помощью дисульфидной связи между цистеиновыми остатками, ограничивающими область. V3 - петля ВИЧ-Iмн, например, образуется дисульфидной связью между цистеиновыми остатками в положениях 302 и 336 gp 120.
Рекомбинантные и синтетические протеиновые фрагменты, включая последовательности аминокислотных остатков петли V3 из различных изолятов ВИЧ, как сообщалось, вызывают образование изолят- и типоспецифичных нейтрализующих антител у грызунов, согласно Lasky et al., Science, 233, pp. 209-212 (1986) Palker et al., Supra; Matsushita et al., J. Virol 62, pp. 2107-2114, (1988); Javakerianet at al, Proc. Natl. Acad. Sci, USA 86, pp. 6788-6772 (1989). Более недавние исследования [Putney et al., supra and La Rosa et al., Science 249, pp. 932-935 (1990) показали, что структура V3- петли с β - поворотом - это сайт, распознаваемый изолятспецифичными антителами. Scott et al., Proc. Natl. Acad. Sci. USA, 87, pp. 8597-8601 (1990) сообщили, что PND может также индуцировать образование типоспецифичных антител у людей. Гипервариабельность PND может отвечать за типоспецифическую нейтрализующую активность, генерируемую эпитопом.
Несколько исследований позволяют предположить, что антитела, полученные против рекомбинантного gp 120, очищенного gp 120 или синтетических пептидов, могут нейтрализовать различные изоляты ВИЧ-I. Javaherian et al., Science 250, pp. 1590- 1593 (1990) и Weiss et al., Nature 324, pp. 572-575 (1986), каждый, описывают нейтрализацию как МN, так и Шв изолятов поликлональной сывороткой, полученной от кроликов, предпочтительно иммунизированных пептидом, соответствующим PND MN- изолятов, и рекомбинантным gp 120, полученным из Шв изолята. Смотрите также, Haynes et al., US Letters Patent 5 019 387.
A Herblom et al., AIDS, 4, pp. 953-960 (1990) описывает препараты моноклональных антител, которые нейтрализуют Шв и одиннадцать первичных изолятов ВИЧ-I. Смотрите также PCT Published Patent Application N WO 91/11198. Гомология штаммов первичных изолятов ВИЧ-I не определена, однако, и эти одиннадцать изолятов могут также быть Шв Durda et al., AIDS Res. Hum. Retro V. 6, pp. 1115-1123 (1990) сообщают о моноклональных антителах, которые блокируют образование синцитиев клетками, инфицированными как MN, так и Шв, но не нейтрализовали инфекционность MN, что установлено с помощью "иммунореакции связывания ЛАВ", исследование, которое имеет целью обеспечить результаты, которые коррелируют с активностью обратной транскриптазы. Патентная заявка N WO 90/15078 Scott et al., по Договору о патентной кооперации, опубликованная 13 декабря 1990 г. описывает моноклональные антитела, которые подавляют образование синцития клетками, инфицированными вакцинным вирусом, экспрессирующих РND изолятов МN или "MN - подобных". Не продемонстрировано ни одно из антител с подтвержденным "широким нейтрализующим" действием с помощью стандартных исследований на обратную транскриптазу, p24 или МТ-2, которые нейтрализуют многочисленные штаммы живых ВИЧ-I. Смотрите также PCT Published Patent Application Nos. WO 88/09181 и WO 91/09625 и Lion et al., J. Immunol. , 143 (12), pp. 3967 - 3975 (1989).
Предшествующие публикации показывают, что моноклональные антитела, реагирующие с PND ВИЧ-I, полученные к настоящему времени, проявляют различные уровни групповой реактивности, но не могут иметь широкое нейтрализующее действие. Различная типо- и группоспецифическая реактивность, установленная в этих исследованиях, может быть связана как с аминокислотными последовательностями, так и со структурой области петли gp 120.
Несколько исследований позволяют предположить, что рецептор CD4 не может представлять только клеточный рецептор, ответственный за вирусную инвазию. Результаты этих исследований подтверждают возможность того, что применение ранее описанных антител, которые блокируют инфицирование клеток CD4 у пациента, может обеспечить только ограниченную защиту против инфекции ВИЧ-I Cheng-Mayer et al., Proc. Natl. Acad. Sci, USA, 84, pp. 3526- 3530 (1987), сообщают об инфицировании глиальных клеток ВИЧ-I, включающем другой рецептор, а не молекулу СD4. Кроме того, Takeda et al., Science 242, pp. 580-583 (1988) показали, что комплексы антитело /ВИЧ-I могут инфицировать моноциты путем эндоцитоза, опосредуемого рецептором, и усиливать репликацию вируса. Сходное усиление инфекции, связанное с антителами, было описано у Halsted et al., Nature 265, pp. 739-741, (1977), Peiris et al., Nature 289, pp. 189-191 (1981); Schlesinger et al., J. Immunol. 127, pp. 659-665 (1981).
Предшествующая работа показала, что определенные вирусы животных инактивируются комплементом, особенно CIq через антителозависимый механизм. Смотрите Weiss в Molecular Biology of Tumor Viruses, RNA Tumor Viruses, Weis et al., Eds, Cold Spriny Harbor Laboratory, New Yerk, pp. 1218-1220 (1982); Welsh et al., Virology, 74, pp. 432-440 (1976); Bartholomew et al., J. Exp. Med. 147, pp. 844-853 (1978); Cooper et al., J. Exp. Med 144, pp. 970-984 (1976); Sherwin et al., Jint. J. Cancer 21, pp. 6-11 (1978). В то время как Banapour et al., Virology., 152, pp. 268-271 (1986) описывает, что препараты непрогретой сыворотки не действуют на плотность ВИЧ-I и его способность инфицировать мононуклеарные клетки периферической крови, Spear et al., J. Virol., 64 (12) pp. 5869-5873 (1990) сообщает, что ВИЧ-I, обработанный комбинацией комплемента и цельной сывороткой от пациентов с сероположительной реакцией на ВИЧ-I, проявляет сниженную инфекционность.
Таким образом, сохраняется потребность в новых препаратах моноклональных антител (включая, например, антитела мышиные, "гуманизированные" и иммунологически активные фрагменты антител), которые обладают специфической иммунореактивностью с ВИЧ-I. В идеале такие антитела должны характеризоваться способностью осуществлять нейтрализацию многочисленных штаммов ВИЧ-I (например Шв и МN), что определяется стандартными методами исследований на обратную транскриптазу, p24, МТ-2 и образование синцития, используя подходящие культивируемые клетки-хозяева (например, клетки H9). С точки зрения предполагаемого использования при пассивной иммунизации инфицированных и неинфицированных пациентов, такие моноклональные антитела должны оптимально быть способны к участию в ( т.е. опосредовании) комплементзависимого виролиза частиц ВИЧ-I и антителозависимого цитолиза клеток, инфицированных ВИЧ-I.
Краткое изложение изобретения
Настоящее изобретение представляет моноклональные антитела, которые специфически реактивны с соответствующими участками протеинов gp 120 или gp 160 ВИЧ-I, включающих аминокислотную последовательность глицин-пролин-глицин-аргинин (Г-П-Г-Р), показанную в ПОСЛ ИД N 1, и характеризуются способностью нейтрализовать инфицирование клеток H9 в культуре живыми штаммами ВИЧ-I МN и Шв, что определяли исследованиями на обратную транскриптазу, p24, МТ-2 и образование синцития. Продукты этого изобретения могут быть, кроме того, охарактеризованы по их способности опосредовать комплементзависимый виролиз частиц ВИЧ-I и/или антителозависимую клеточную цитотоксичность для клеток, инфицированных ВИЧ-I.
Моноклональные антитела по настоящему изобретению, предпочтительно антитела IgG, особенно подходят для использования при анти-ВИЧ-I лечения животных, особенно людей, восприимчивых к/или инфицированных ВИЧ-I. Иммунологически эффективные количества моноклональных антител вводятся пациенту, инфицированному ВИЧ-I или при риске инфицирования вирусом, чтобы получить пассивный иммунитет к ВИЧ-I инфекции, и предпочтительно, чтобы осуществить комплементзависимый виролиз частиц ВИЧ-I и/или клеточную цитотоксичность, зависящую от антител, для клеток пациента, инфицированных ВИЧ-I.
Химерные или "очеловеченные" антитела (включая CDR - трансплантированные антитела), фрагменты антител и особенно биспецифические антитела, основанные на заявляемых моноклональных антителах, включены в рассмотрение настоящего изобретения так же, как и рекомбинантные связанные с антителами продукты, продуцируемые прокариотическими и эукариотическими клетками.
Например, фрагменты антител, такие как Fab и F(ab)2 фрагменты, могут быть получены в культуре клеток-хозяев, таких как Е. coli дрожжи, клетки насекомых и клетки млекопитающих с определением информации о структуре (последовательности) для разных областей антител этого изобретения, информация о последовательности для различных областей антител делает возможным также получение CDR - трансплантированных антител. Кроме того, химерные антитела (например, мышино-человеческие антитела) могут быть получены при использовании трансформированных клеток миеломы мышей или гибридомных клеток, а биспецифические антитела могут также быть получены с помощью гибридных гибридомных клеток.
В рассмотрение настоящего изобретения входит также использование при анти-ВИЧ-I лечении комбинации продуктов настоящего изобретения и других иммунологических агентов и/или химических терапевтических агентов. Потенциальные агенты для комбинированного применения включают комплемент, антитела, которые связываются с различными нейтрализующими и ненейтрализующими областями протеинов ВИЧ-I, и химические агенты, такие как AZT.
В качестве пояснения к дальнейшему детальному описанию моноклональные антитела настоящего изобретения получали путем иммунизации подходящего хозяина живым ВИЧ-I, таким образом получая gp 120 в его природной конформации.
Специфически иллюстрируют нестоящее изобретение мышиные моноклональные антитела (обозначаемые NМ-01), продуцируемые линией гибридомных клеток HB 10726, которая была получена на хранение American Type Culture Collection, Rockville, Maryland, 9 апреля 1991 г. и обозначена А.Т.С.С. поступление N 0. HB 10726.
Многочисленные аспекты и преимущества настоящего изобретения будут очевидны при рассмотрении иллюстрирующих примеров и описаний применения настоящего изобретения в его последующем детальном описании, справа по чертежам представлена, причем:
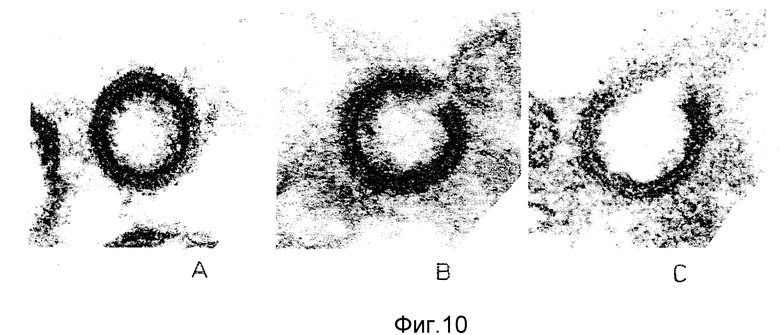
ФИГУРА 1 является сложный ауторадиограммой неинфицированных H9 клеток, протеинов HIV-IMN и HIV - IIIIB, подвергнутых иммуноблоттингу с моноклональными антителами этого изобретения и имунной сывороткой от сероположительных больных СПИД.
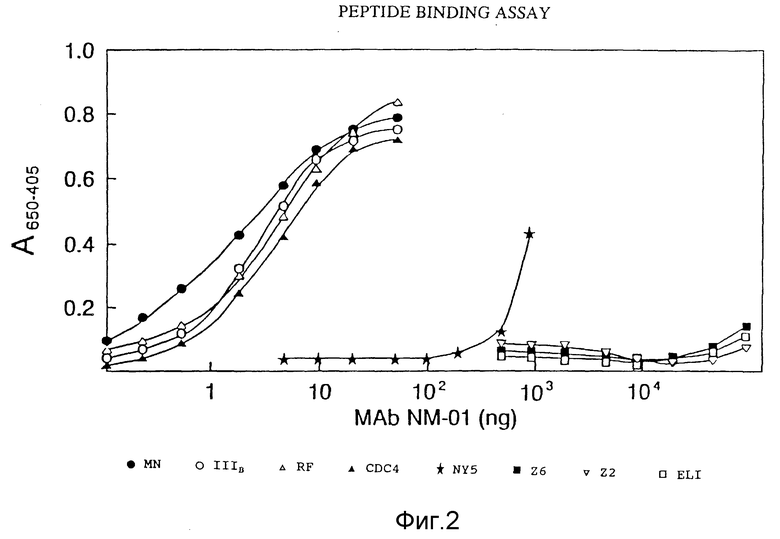
ФИГУРА 2 графически представляет результаты определения иммунореактивности антител этого изобретения с пептидами, соответствующими области петли V3 различных штаммов ВИЧ-I.
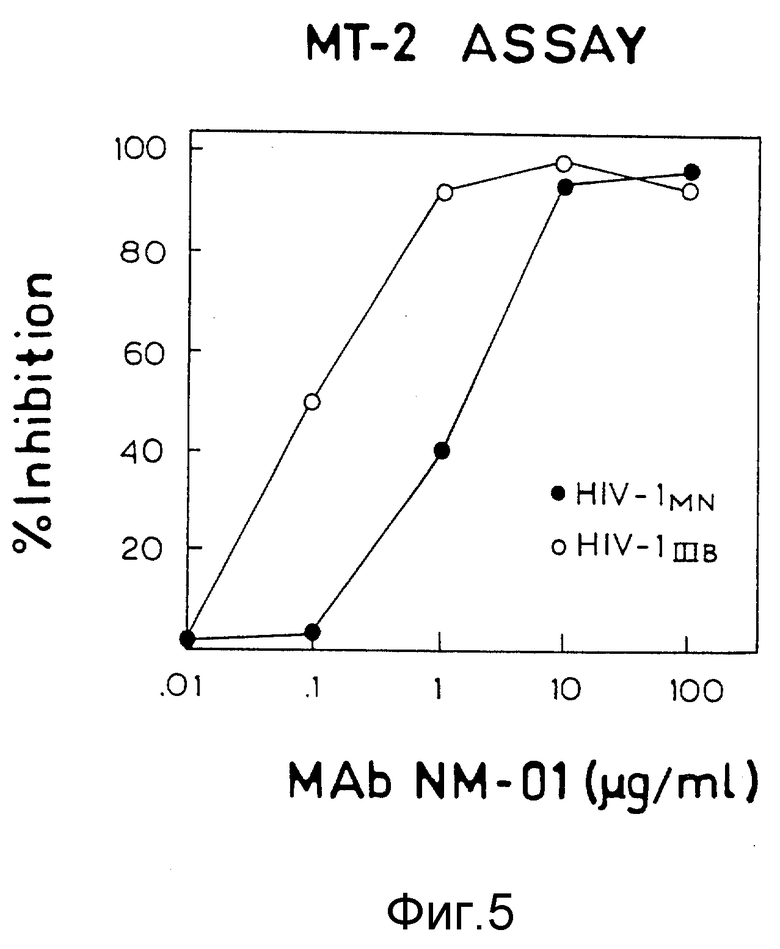
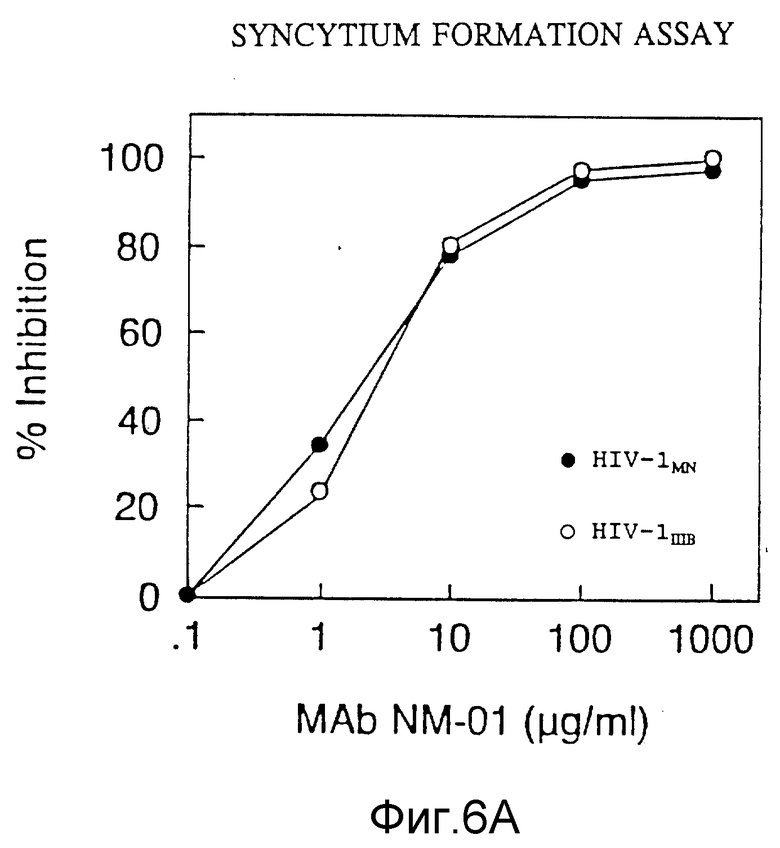
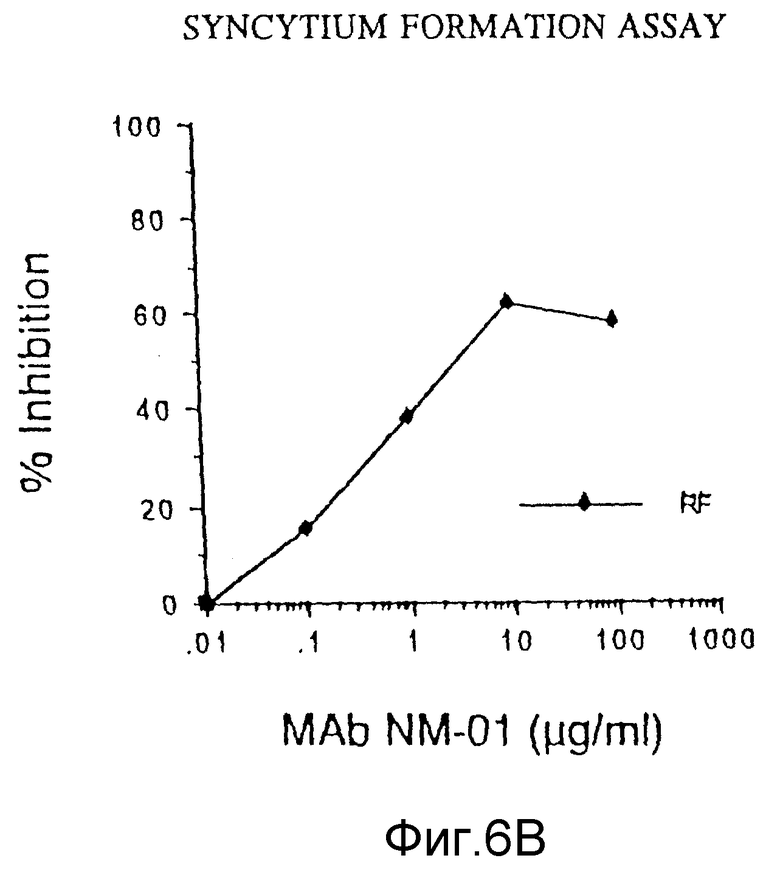
ФИГУРЫ с 3A по 3C, 4,5 и 6A по 6B графически представляют результаты определений обратной транскриптазы, p24, МТ-2 и образования синцития, соответственно, и способности моноклональных антител этого изобретения нейтрализовать инфекцию клеток H9 живыми штаммами ВИЧ-I.
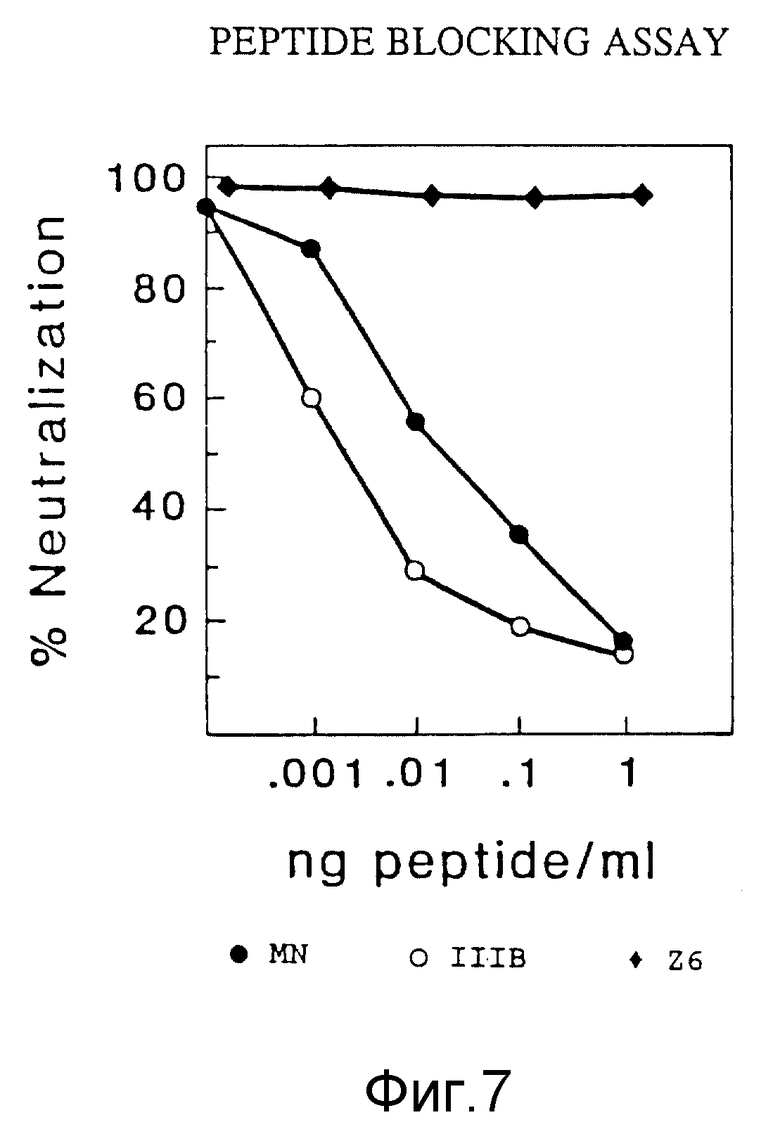
ФИГУРА 7 графически показывает результаты исследования по определению пептидной блокады нейтрализации инфекционности для антител данного изобретения.
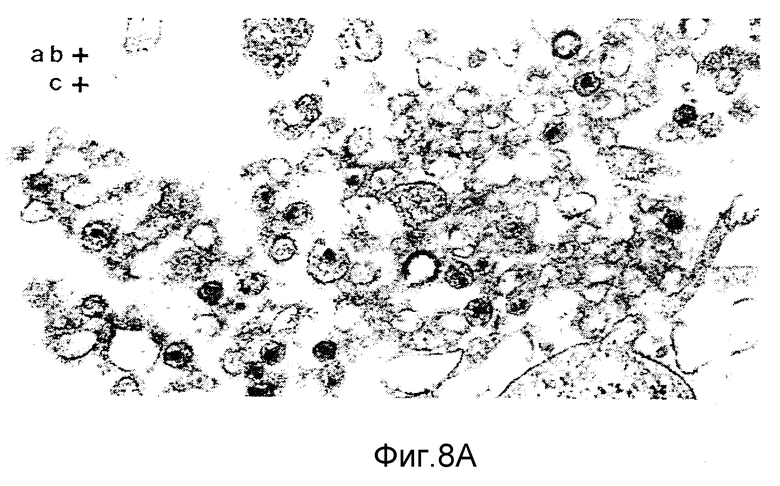
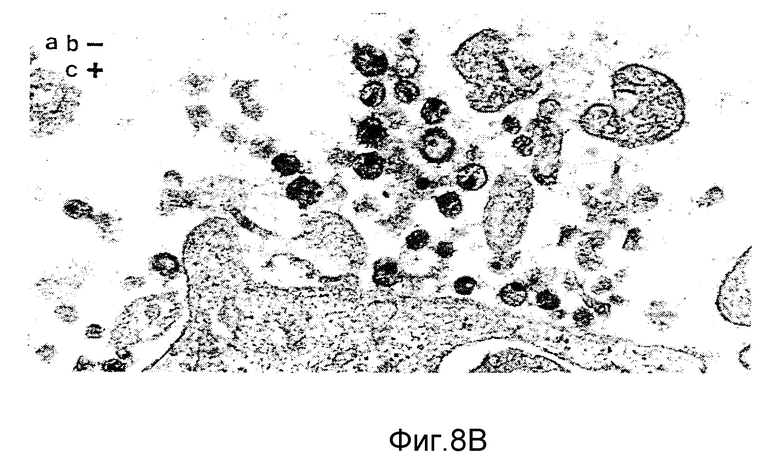
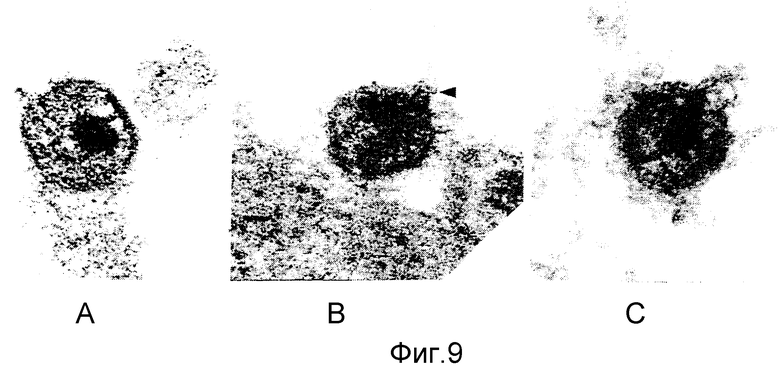
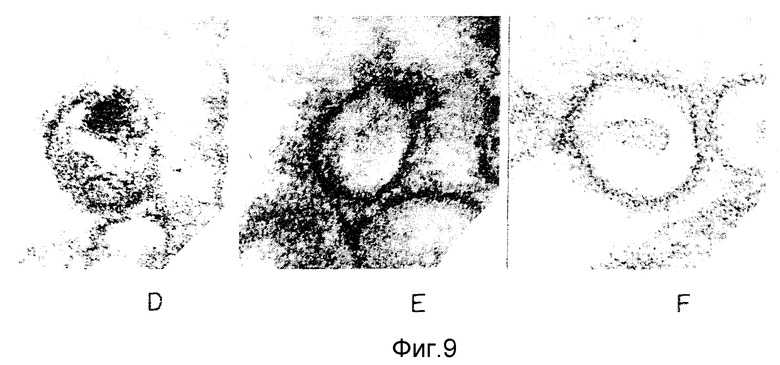
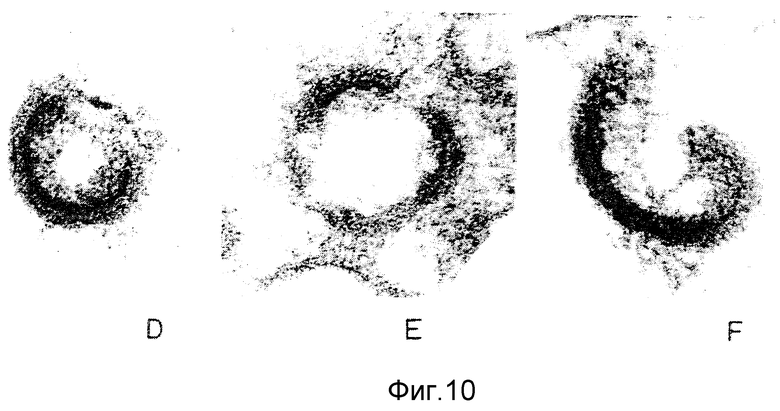
ФИГУРЫ с 8A по B, 9A по F и с 10A по F это электронные микрофотографии частиц ВИЧ-I, которые были обработаны комбинацией моноклональных антител данного изобретения и комплемента.
ФИГУРЫ 11 и 12 показывают аминокислотные последовательностей различных областей легких и тяжелых цепей, соответственно, моноклональных антител данного изобретения, NМ-01, так же, как аминокислотные последовательности легких и тяжелых цепей разных анти-ВИЧ-I моноклональных антител.
ПРИМЕРЫ
Следующие примеры иллюстрируют применение изобретения в получении линии гибридомных клеток НВ 10726, выделение из культуры моноклональных антител, иммунореактивных с gр 120 ВИЧ-I (или его предшественником gр 160) протеинами так же, как и с пептидами, включающими аминокислотную последовательность Г-П-Г-Р, показанную в ПОСЛ ИД N 1, характеристику таких моноклональных антител.
Более конкретно, пример 1 показывает получение гибридомных клеток линии HB 10726 и выделение из них моноклональных антител NCM-01. В примере 2 говорится о картировании вирусного эпитопа, распознаваемого антителом NМ-01. Пример 3 описывает характеристику реактивности моноклональных антител с различными изолятами ВИЧ-I. В примере 4 рассказывается об определении способности антител NM-01 нейтрализовать инфицирование клеток H9 различными штаммами живых ВИЧ-I, что продемонстрировано исследованиями на обратную транскриптазу и p24. В примере 5 говорится о дальнейшем определении способности антител нейтрализовать инфекционность живых изолятов ВИЧ-I, что демонстрируется исследованиями на МТ-2 и образование синцития. Пример 6 относится к пептидной блокаде нейтрализации инфекционности ВИЧ-I моноклональными антителами NМ-01. Пример 7 описывает анализ способности моноклональных антител NМ-01 опосредовать комплементзависимый лизис ВИЧ-I. Пример 8 относится к определению действия комбинации моноклональных антител NМ-01 и комплемента на инфекционность ВИЧ-I в отношении чувствительных клеток в культуре. Пример 9 описывает ДНК и производные аминокислотные последовательности тяжелых и легких цепей различных областей моноклонального антитела NM-01, а также относится к получению химерных/"очеловеченных" вариантов моноклональных антител NМ-01.
Пример 1
Линия гибридомных клеток HB 10726 была получена с использованием стандартных иммунологических методов, такие как описаны у Oi и Herzenberg, Selected Methods Cell Immunology, pp. 351-372 (1979) и Codding, J. Immunol. Meth. 39, pp, 285-308 (1980) и подробно изложены ниже.
Очистка живых ВИЧ-IMN
300 мл инфицированной ВИЧ-IMN культуры клеток H9 собирали и центрифугировали при 1500 об/мин в течение 5 минут при 4oC до образования плотного осадка клеток. Содержащий вирус супернатант отбирался и сохранялся, тогда как осадок снова центрифугировался при 2100 об/мин в течение 20 мин. Второй супернатент собирался и соединялся с первым, и супернатант ультрацентрифугировался в роторе SW 27 при 25 000 об/мин в течение 90 минут при 4oC до образования плотного осадка вирусных частиц. Полученный супернатант отбрасывался, осадок вирусных частиц ресуспендировался в примере 10 мл буфера ТНЭ (100 мМ NaCl, 10 мМ Трис-HCl, pH 7,7, 1 мм ЭДТА). Для ультрацентрифугирования подготавливали пробирку, содержащую в нижнем слое 10 мл 50% сахарозной ТНЭ, в среднем слое 10 мл 25% сахарозного ТНЭ и в верхнем слое 10 мл образца, содержащего вирусы, и центрифугировали в ультрацентрифуге при 25000 об/мин при 4oC в течение 90 минут. Вирус осаждался в виде белой полосы между слоями растворов сахарозы в ТНЭ и собирался с помощью пастеровской пипетки. 20 мл ТНЭ/15 мМ ЭДТА (100 мМ NaCl, 10 мМ Трис-HCl, pH 7,7, 15 мм ЭДТА) добавляли к вирусам и образец с вирусами снова центрифугировали при 25 000 об/мин при 4oC в течение 90 минут. Полученный в результате осадок содержал очищенный живой ВИЧ-IMN.
Иммунизация и получение гибридомы
100 мкг живого ВИЧ-IMN использовали для иммунизации каждой из трех двухмесячных BaLb/c -мышей путем внутрибрюшинной инъекции. Каждая мышь повторно иммунизировалась через 3 недели 30 мкг вируса и опять еще через 3 недели 100 мкг вирусного препарата. Мышей забивали через 3 дня после второй повторной иммунизации и линии гибридомных клеток получали путем слияния спленоцитов с P3-X63-Ag8 - V1 клетками (АТСС. Номер поступления CRL 1597). Линии гиридомных клеток получали также из селезенок мышей, иммунизированных хронически инфицированными клетками H9 (10 мышей), остро инфицированными клетками H9 (9 мышей) и мембранами инфицированных клеток H9 (3 мыши). Хронически инфицированные клетки H9 - это клетки после 2-3 недельной инъекции, имеющие показатели обратной транскриптазы (ОТ) от 100000 до 150000 имп/мин, тогда как остро инфицированные клетки H9 - это клетки после 10-12 дней инфекции, имеющие показатели ОТ от 200000 имп/мин до 250000 имп/мин.
Линии гибридомных клеток были получены следующим методом. Смесь клеток селезенки от иммунизированных мышей центрифугировали при 800 д в течение 5 минут. Супернатант отсасывали с осадка клеток и добавляли к осадку 1 мл теплого (37oC) 50% PEG-1500 на 108 клеток в течение 1 минуты (добавлять 0,25 мл, взболтать слегка тонким концом пипетки в течение 15 секунд и повторить). Смесь взбалтывали дополнительно в течение минуты тем же самым концом пипетки, не разбивая комочки клеток. 1 мл "неполной среды" [PPMI 1640 (JPH Biosciences) с добавлением 25 мМ HEPES (Sigma Co), 10000 ед/мл пенициллина и 10000 мг/мл стрептомицина] добавляли затем в течение 1 минуты таким образом (0,25 мл каждые 15 секунд) и еще 1 мл добавляли в течение следующей минуты.
Затем подмешивали 7 мл неполной среды в течение 2-3 минут (1 мл каждые 20 секунд), получая в результате суспензию мелких клеточных комочков. Окончательная суспензия центрифугировалась при 500 g при позиции 5 на клинической центрифуге в течение 5 минут, и супернатант удалялся. Осадок ресуспендировался путем вращения пробирки (не взбалтывая или пипетируя раствор вверх и вниз) в "полной среде" ["неполная среда", как описано выше, дополненная 15% эмбриональной телячьей сыворотки (ЭТС)] до концентрации 2 •106 клеток на мл среды. 0,1 мл этой суспензии (общее число клеток 2•105) помещали в ячейку 96-ячеечной платы, платы инкубировали при 37oC в атмосфере с 7% CO2. День слияния считался днем 0.
Селекция и первоначальное выявление гибридом в среде ГАТ
Через 24 часа после слияния (день 1) в каждую ячейку добавляли 0,1 мл среды ГАТ (10-4М гипоксантина, 5•10-7 М аминоптерина и 1,6•10-5М тимидина). В дни 2, 3, 5, 8, 11, 14, 17 и 21 из каждой ячейки удаляли 0,1 мл среды и заменяли 0,1 мл свежей среды ГАТ. В дни с 2 по 5 ячейки, как казалось, содержали только нежизнеспособные клетки. Гибридомы начали появляться между днями 5 и 10. Гибридомы были легко различимы в виде колоний высокопреломляющих клеток, окруженных клеточными обломками.
Выявление гибридом
Для выявления гибридомных супернатантов использовано несколько проб. Гибридомы, секретирующие антитела, реактивные к ВИЧ-I, первоначально идентифицировали с помощью скренинговых (сортирующих) мембран, получаемых из неинфицированных или инфицированных МN клеток H9 путем ELISA с супернатантами гибридомных культур. За этим первоначальным отбором следовала проверка с помощью иммунофлюоресцентного и радиоиммунного исследований, чтобы дополнить данные ELISA данными по связыванию антител с живыми инфицированными клетками.
Клеточные мембраны для ELISA получали из инфицированных и неинфицированных клеток H9. Клетки суспендировали в растворе 250 мМ сахарозы в 10 мМ Трис-HCl буфере с pH 7,4, содержащем 1 мМ ЭДТА. Суспензия гомогенизировалась в гомогенизаторе Дунса, помещенном в ледяную баню, до тех пор, когда переставали определяться живые клетки с помощью исключающей пробы с трипановым синим. Смесь центрифугировалась в течение 2 минут при 50 g. Полученный осадок повторно гомогенизировался и центрифугировался. Два супернатанта объединялись и центрифугировались при 20000 в течение 20 минут. Осадок снова гомогенизировался в том же буфере и центрифугировался в течение 20 минут, и осадок ресуспендировали в 7 мл первоначального 250 мМ сахарозно-ЭДТА буфера. Этот раствор затем наслаивался поверх раствора 2 М сахарозы в 10 мМ трис-HCl буфере, содержащего 1 мМ ЭДТА, и центрифугировали в течение 1 часа при 80000 g. В результате получается рыхлая белая поверхность раздела, которую собирали и ресуспендировали в 250 мМ сахарозном буфере. Содержание протеина определяли по методу BCA (Pierce Chemical Company). Суспензия делилась на образцы и хранилась при -70oC.
Для ELISA клеточные мембраны добавляли в количестве 400 н г на ячейку в 96-ячеичные платы и высушивали в течение ночи при 25oM. Платы промывались 0,5% Тритон-X®/фосфатнобуферным физиологическим раствором (ФБР), блокировались 5% эмбриональной бычьей сывороткой (ЭБС) /ФБР и отзывались опять. Гибридомный супернатант (40 мкл) разбавлялся 50 мкл ФБР и добавлялся в ячейки с выдерживанием в течение ночи при 4oC. После отмывания добавляли кроличий антимышиный 1 (H + ), конъюгированный с пероксидазой хрена (ПOX) (Zymed) в ячейки и оставляли на 2 часа при 25oC. Ячейки отмывали 0,5% Тритон-X®/ФБР и затем инкубировали в присутствии АБTС (Bio-Rad субстратный комплект) в течение 20 минут перед определением оптической плотность при 405 и 650 нм.
Супернатанты гибридом, полученных из клеток селезенки мышей, иммунизированных хронически инфицированными клетками и остро инфицированными клетками, давали положительные реакции с мембраной как инфицированной, так и неинфицированной клетки в ELISA, что свидетельствует об отсутствии ВИЧ-1- специфичности у антител, продуцируемых гибридомами. Из 1039 гибридом, полученных из клеток селезенки мышей, иммунизированных мембранами инфицированных клеток, 5 их супернатантов сильно реагировали с мембранами инфицированных клеток и очень слабо реагировали с мембранами неинфицированных клеток. Вестерн-блоттинг выполняли с супернатантами этих линий гибридомных клеток и определили, что три из моноклональных антител давали полосы с ВИЧ-1 p55, одно - полосу с ВИЧ-I p55 и p24 и остальные не давали полосы в Вестерн-блот (данные не показаны).
1187 гибридом были получены из клеток селезенки мышей, иммунизированных живым ВИЧ-IMN. Четыре линии гибридомных клеток были отобраны для дальнейшего отбора по результатам ELISA, показывающим, что антитела четырех супернатантов сильно реагировали с мембраной инфицированных клеток. Результаты ELISA представлены в таблице 1 в виде отношений значений, полученных для мембран инфицированных клеток в сравнении с мембранами неинфицированных клеток.
Супернатанты четырех гибридом подвергали клонированию с ограниченным разбавлением и анализировали с помощью радиоиммунного исследования (РИА). Кроличьи антимышиные IgG, меченные 125I (RαM IgG - 125I), очищали на колонке с сефадексом G - 50 (NeN - DuPont). Неинфицированные клетки H9 или клетки H9 (7,5•105 клеток в 150 мкмл), инфицированные ВИЧ-IMN, помещали в 15 мл пробирки. 50 мкл супернатанта от каждой гибридомы добавляли в каждую из пробирок, содержащих неинфицированные и инфицированные клетки, и смеси инкубировали в течение ночи при 4oC. Клетки отмывали 2 раза 2 мл ФБР/50% Твин-20 с вращением пробирки между промываниями. Добавляли 50 мкл RαM IgG-125I (750000 имп/мин) в ФБР/5% ЭБС и смесь снова инкубировали в течение ночи при 4oC. После инкубации клетки отмывали 3 раза ФБР/50% Твин-20. 100 мкл ФБР/5% Тритон-X добавляли, чтобы дезинфицировать клетки и 100 мкл 1 М NaOH добавляли, чтобы улучшить перенос метки в кюветы сцинтилляционного счетчика. Образцы просчитывали, и результаты РИА представлены в таблице 1, ниже в виде отношений значений имп/мин-1, полученных для инфицированных клеток в сравнении со значениями имп/мин-1 для неинфицированных клеток.
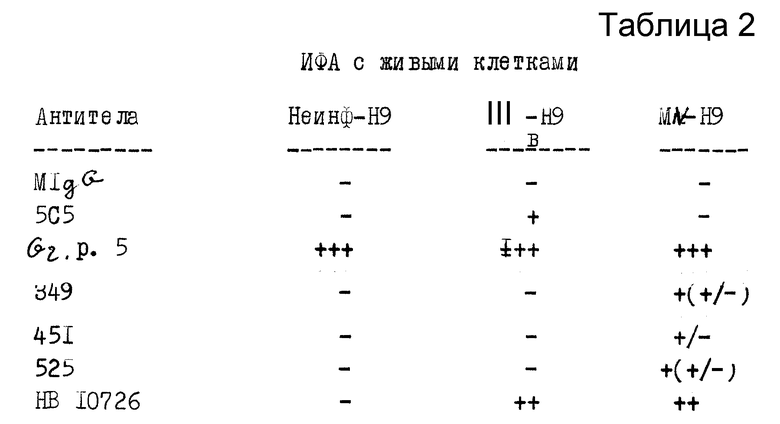
Далее, четыре линии гибридомных клеток исследовали с помощью иммунофлюоресценции (ИФА) 2 мл взвеси или неинфицированных, или инфицированных ВИЧ-I клеток H9 (примерно 1•106 клеток в мл) помещали в 10 мл стерильную центрифужную пробирку с 10 мл ФБР (без Ca++ или Mg++). Клетки отмывали один раз 10 мл ФБР путем наполнения пробирки, взбалтывания вращением, центрифугирования при 100 об/мин в течение 5 минут и отсасывания всей жидкости, кроме примерно 100 мкл супернатанта, оставляя "молочную" клеточную суспензию. При работе под колпаком с ламинарным потоком 51 мм 10-ячеичные предметные стекла (Cell Line Association) покрывали клеточной суспензией, заполняя каждую ячейку и удаляя излишки суспензии концом пипетки. Покрытые предметные стекла подсушивали на воздухе и фиксировали метанолом при комнатной температуре в течение 10 минут.
Определяли реактивность препаратов неинфицированных и инфицированных клеток на предметных стеклах с неразбавленными супернатантами каждой из четырех гибридом и с их разведениями с титром 1:50 (супернатант разбавляли 0,02%-ным снятым молоком). 15 мкл неразбавленного или разбавленного супернатанта наносили на каждую ячейку предметного стекла. Стекла инкубировали при 37oC в течение 30 минут и погружали в ФБР с взбалтыванием на 5 минут. Предметные стекла затем быстро прополаскивали в дистиллированной воде и высушивали под колпаком в ламинарном потоке воздуха. На каждую ячейку добавляли 16 мкл козлиных-против-мышиных IgG (H +L) F(ab)2 фрагмента (Cuppel Biomedical), разбавленных 1:80 0,02%-ным снятым молоком. Стекла снова инкубировали при 37oC в течение 30 минут и затем погружали в ФБР. Предметные стекла прополаскивали в 0,01%-ном растворе Эваснс-синего в ФБР в течение 5 секунд и 2 раза споласкивали дистиллированной водой. Стекла просматривали на иммунофлюоресценцию, и результаты анализа представлены в таблице 2, причем мышиный IgG (MIgG) 5C5 антитела (анти-Шв) и супернатант гр. 5 (из гибридомы, полученной из селезенок мышей, иммунизированных мембранами инфицированных клеток, антиинфицированными и неинфицированными клетками) являются контрольными антителами.
Гибридомная клеточная линия HB 10726 была выбрана как наиболее обещающая в отношении антител на основе данных РИА и иммунофлюоресценции. Эта клеточная линия не имела наивысшего показателя связывания в ELISA, но т.к. РИА и результаты иммунофлюоресценции представляют связывание с живыми инфицированными клетками, тогда как ELISA представляет связывание с высушенными клеточными мембранами, данные РИА более важны. Клеточная линия субклонировалась дважды, и моноклональные антитела, которые она продуцирует, были обозначены NM-01. Эту клеточную линию вводили мышам внутрибрюшинно обычным способом и Mab NM-01 концентрировали из асцитной жидкости путем очистки на протеин А аффинной колонке (Pierce). Изотип антител NM-01 был определи как IgG2, с помощью типоспецифических антисывороток (Bio-Rad). Антитела (1,8 мг/мл) разбавляли средой RPM 1640 с 15% ФБР и использовали в следующих примерах.
Пример 2
Чтобы охарактеризовать вирусный эпитоп, распознаваемый моноклональным антителом MN-01, антитела сначала подвергали вестерн-блот анализу на реактивность с очищенными протеинами вирионов МN и Шв и затем ELISA на реактивность с перекрывающими пептидами, соответствующими аминокислотной последовательности области петли 3 ВИЧ-I gp 120.
Вестерн-блот анализ
Вирионы МN и Шв очищали из супернатантов культур инфицированных клеток H9 и разрушали в 1,3% SDS/3% β - меркаптоэтаноле и затем подвергали электрофорезу в 0,1% SDS/10% полиакриламидном геле. После переноса протеинов на нитроцеллюлозную бумагу полоски выдерживали в течение ночи с MAb NM-01 в блокирующем буфере (0,02 М Трис-HCl, pH 7,4, 0,1 M NaCl, 0,05% нормальной козлиной сыворотки и 5% обезжиренного сухого молока) при 4oC и затем отмывали в 0,02 М Трис-HCl, pH 7,4, 0,1 NaCl и 0,3% Твин. Полоски затем инкубировали с биотинилированными козлиным против мышиного антигана IgG (Zymed) в течение 2 часа, отмывали и подвергали взаимодействию с 125I - стрептавидином (Amersham, Arlington. Heights, IL) в течение еще одного часа при 4oC. MAb реактивность NM-01 определяли с помощью ауторадиографии.
Результаты ауторадиографии представлены на фигуре 1, причем: полосы геля 1 и 3 содержали мембраны неинфицированных клеток H9, полоса 4 содержала мембраны клеток H9, инфицированных ВИЧ-IMN; полосы 2 и 5 содержали вирус ВИЧ-IMN, и полосы 6 и 7 содержали ВИЧШв вирус. Антитела NM-01 реагировали с протеинами в полосах 1, 2 и 6, тогда как сыворотка сероположительных больных реагировала с протеинами в полосах 3-5 и 7.
Моноклональные антитела NM-01 проявляли реактивность с протеинами вирусов NM и Шв, имеющими молекулярный вес примерно 120 кD, но не реагируют с любыми другими вирусными антигенами, демонстрируя таким образом, что антитела распознают эпитоп gp 120.
Картирование эпитопа с помощью ELISA
Чтобы идентифицировать специфический эпитоп gp 120, распознаваемый антителом NМ-01, определяли реактивность антител с перекрывающими пептидами, соответствующими области V3 петли gp 120 с помощью ELISA. Пептиды, синтезированные с помощью Maltiple Peptide System, San Diego, CA, соответствовали аминокислотам 302-316, 312-326 и 322-336 gp 120 ВИЧ-IMN.
Три пептида (250 нг/50 мкл 0,1 М боратного буфера, pH 8,0 на ячейку) инкубировали в течение ночи при 37oC в платах Immulon (Dynatech). Платы промывали ФБР и блокировали ФБР /1% Твин /1% бычьего сывороточного альбумина (БСА) в течение 1 часа при комнатной температуре. Блокирующий агент удаляли и в платы добавляли различные количества антител NM-01 или мышиного IgG(MIgG), разведенных в 100 мкл среды HT. Антителам давали возможность прореагировать в течение 2 часов при комнатной температуре, платы промывали затем водопроводной водой 10 раз. В ФБР /0,05% Твин /5% БСА вводили конъюгированные с HPP кроличьи против мышиного антигена вторичные антитела, разбавленные 1: 1000, и 100 мкл этого раствора добавляли в каждую ячейку. Платы инкубировали в течение 1 часа при комнатной температуре и затем промывали водопроводной водой 10 раз. Добавляли субстрат АБТС (Bio-Rad) на 20 минут и платы анализировали при 650 нм. Посл. IDNOS2-4 показывают аминокислотные последовательности пептидов, и таблица 3 показывает результаты исследования с использованием перекрывающих пептидов, причем антитела MIgG и НТ среда были отрицательными контролями.
В то время как отсутствовала определяемая реактивность на фоне MAb NM-01 с пептидами, соответствующими аминокислотам 302-316 или 322-336 петли V3, связывание антител с пептидами, представляющими аминокислоты 312-326, было видно невооруженным глазом. Контрольные антитела, мышиный IgG не связывали пептидов.
Пример 3
Демонстрация того, что моноклональные антитела NM-01 связываются с областью петли V3 gp 120 ВИЧ-IMN, побудила на дальнейшие исследования по степени этой реактивности с другими изоляторами ВИЧ-I. Определяли реактивность антител с пептидами, соответствующими области петли V3, изолятов Шв, RF, CDC4, NY/5, Z6, Z2 и ELI ВИЧ-I с помощью ELISA. Аминокислотные последовательности пептидов показаны ниже в таблице 4 и в списке последовательностей, как Посл ИД NN 5-12 соответственно.
Определение пептидного связывания
Пептиды (250 нг/0,1 М боратного буфера, pH 8,0, синтезированные American Biotechnologies Cambridge, NA, инкубировали в течение ночи при 4oC в платах Immylon 2 (Dymatech). Платы промывали ФБР, блокировали 0,1% Твин /0,1% БСА/ФБР в течение 2 часов при 25oC и затем инкубировали с MAb NM-01 в течение 1 часа при 37oC. После промывания водопроводной водой платы инкубировали с кроличьими против мышиного антигена вторичными антителами, конъюгированными с ПХ, и затем с субстратом (Bio-Rad) в течение 20 минут. Реактивность определяли по оптической плотности при 650-405 нм. Результаты исследования представлены на фигуре 2.
Моноклональные антитела NM-01 реагировали с пептидами петли изолятов МN (закрашенный кружок), Шв (незакрашенный кружок), P (незакрашенный треугольник) и CDC4 (закрашенный треугольник). Связывание антител с пептидами Шв, RF и CDC4 было сравнимо с таковым с пептидом МN. Антитела показывали также меньшее сродство к пептиду N Y/5 (звездочка). Моноклональные антитела NМ-01 к тому же ложно реактивы с RF-подобным пептидом, показанным в Посл ИД N 13. Напротив, была слабая или никакой реактивности с пептидами петли изолятов Z6 (закрашенный квадрат), Z2 (перевернутый незакрашенный треугольник) ELI (незакрашенный квадрат), эти результаты показывают, что моноклональные антитела NM-01 распознают, в частности, эпитоп петли gp 120 многочисленных изолятов ВИЧ-I, имеющих аминокислотную последовательность, показанную в Посл ИД N 1, G-P-G-R.
Другие моноклональные антитела против gp120 ВИЧ-I, моноклональные антитела ВАТ123, описаны у Liou et al. как реактивные с МN-подобными и Шв-подобными пептидами петли V3 и нереактивные с RF- подобным пептидом (фигура 5А), эта описанная реактивность отлична от таковой моноклональных антител NM-01, приведенной в предыдущем абзаце. В то же время оба моноклональных антитела NM-01 и BAT123 одинаково хорошо связываются с пептидом Шв, примерно пятидесятикратное увеличение концентрации ВАТ123 необходимо, чтобы получить связывание с пептидом MN, сходное со связыванием с NМ-01. Кроме того, NМ-01 реактивны с RF-подобным пептидом, показанным в таблице 4 и Посл ИД N 7, тогда как ВАТ 123 не связывается с RF-подобным пептидом Посл ИД N 13, даже при концентрациях антител 10000 мкг/мл.
Пример 4
Моноклональные антитела NM-01 испытывали на способность нейтрализовать инфекцию клеток H9 живыми штаммами ВИЧ-I, МN, Шв и RF, что определяли по исследованию активности обратной транскриптазы, а при инициировании штаммами ВИЧ-I МN и Шв определяли с помощью пробы на p24.
Исследования на обратную транскриптазу и p24
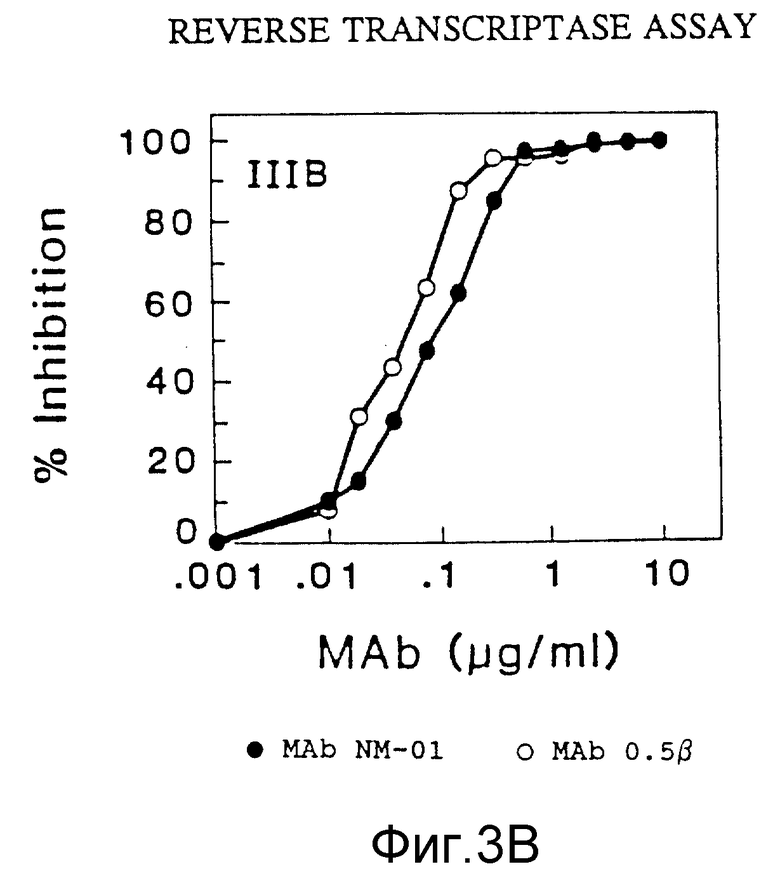
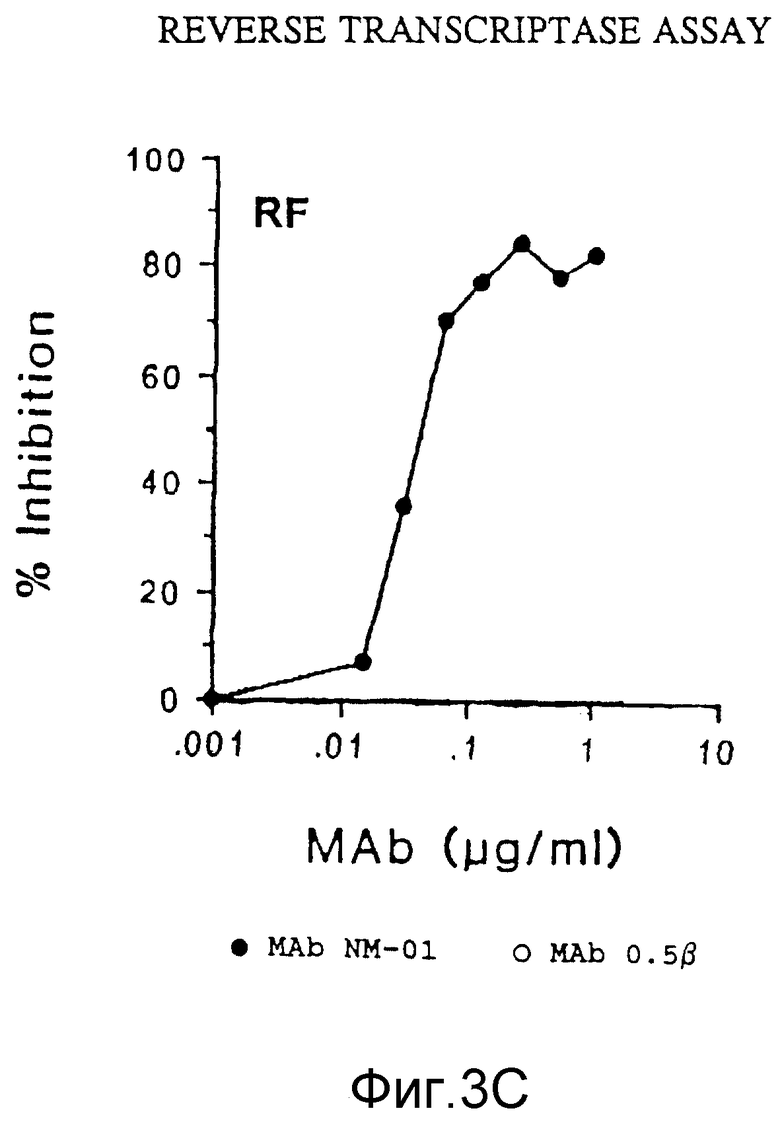
Разведения MAb NM-01 инкубировали с 40 TCLD50 c МN или 100 TCLD50 Шв живых вирусов в 96-ячеичных платах в течение 1,5 часов при 37oC. MAb 0,5 β (AIDS Research and Reference Reagent Program Catalog, National Institute of Allergy и Infections Diseases) использован в качестве как положительного, так и отрицательного контроля в исследованиях ОТ; они связываются с gp 120 ВИЧ-IIIIв . Затем добавляли клетки H9 (2,5 • 104) в каждую ячейку и платы инкубировали еще в течение часа при 37oC. Суспензию клеток H9 затем разводили в RPMI 640 /15% ФБР и инкубировали в 24-ячеичной плате при 37oC. Продукцию вируса определяли по исследованию на обратную транскриптазу (ОТ), выполняемому на 7 день, как описано у Poiesz et al., Proc Natl. Acad. Sci. USA, 77, рр 7415-7419 (1980) и пробой на p24, выполняемой на 5 день (Du Pont HIV-1 п. 24 Core Profile ELISA). Результаты этих двух исследований представлены на фигурах 3A, 3B и 4 соответственно.
Моноклональные антитела NM-01 (закрашенные кружочки в фиг. 3A) полностью нейтрализовали инфекционность живого вируса МN, как выявило исследование на ОТ, в концентрациях 10-100 мкг/мл. Кроме того, использование антител в концентрации < 1 мкг/мл приводило к 50%-ному подавлению вирусной инфекционности (ID50). Эти данные контрастировали с отсутствием определяемой нейтрализации с MAb 0,5β (незакрашенные кружочки, фиг. 3А). MAb NM-01 также нейтрализовали живой вирус Шв при ID50 примерно 0,1 мкг/1 мл (фигура 3В). MAb 0,5β нейтрализовали Шв немного более эффективно, чем МАb NM-01 (фигура 3B). Сходные результаты были получены для ВИЧ-I МN и Шв при определении p24 (фигура 4). При исследовании по обратной транскриптазе моноклональные антитела NM-01 также подавляли живые вирусы RF при ID50 около 0,05 мкг/мл (фигура 3C).
Эти данные показывают, что моноклональные антитела NМ-01 нейтрализуют инфекционность по крайней мере трех различных штаммов ВИЧ-I.
Пример 5
Исследование нейтрализации инфекционности живых вирусов ВИЧ-I, продемонстрированное пробами на обратную транскриптазу и p24, было расширено изучением действия моноклональных антител NM-01 при испытании с МТ-2 с использованием живых вирусов МN и Шв и на образование синцития с использованием живых вирусов МN, Шв и RF.
Проба с МТ-2
Проба с МТ-2 выполнялась, как описано у Richman, AIDS Research and Reference Reagent Program, Courier N 90-01, pp. 6-9 (1990) с некоторыми модификациями. Живые вирусы MN и Шв инкубировали с разведениями MAb NM-01 в течение 1,5 часов при 4oC в 96-ячеечных платах. Клетки МТ-2 (8•105) добавляли в ячейки, и платы инкубировали в течение 3 дней при 37oC. Чтобы оценить жизнеспособность клеток, затем определяли снижение (восстановление) MTT красителя, по Mosmann, J. Immunol. Meth, 65, pp. 55-63 (1983) и Panwels et al., J. Virol. Meth. , 20, pp. 55-63 (1983). Результаты пробы МТ-2 подтверждают результаты определений по обратной транскриптазе и P24 и представлены на фигуре 5, причем незакрашенные кружочки отмечают значения для Шв (100 TCID50), а закрашенные кружочки отмечают значения для MN (40 TCID50).
Моноклональные антитела NM-01 нейтрализовали инфекционность живых изолятов MN и Шв при ID50 в концентрации 2,0 и 0,1 мкг/мл соответственно.
Проба на образование синцития
Проба подавления связыванием была модификацией пробы, описанной ранее у Johnson and Wolker, Eds, Techniques in HIV-1 Research, Stockton Press, New-York, pp. 92-97 (1990). Вкратце, хронически инфицированные MN или Шв вирусом клетки H9 инкубировали с разведениями MAb NM-01 в течение 1 часа при 37oC. Клетки C8166 добавляли затем в каждую ячейку и инкубировали 2 часа при 37oC. Синцитии, более чем три лимфоцитных клетки, в диаметре подсчитывали и сравнивали с синцитиями, полученными для контрольных инфицированных клеток H9, обработанных в отсутствие антител. Результаты пробы образования синцития также подтверждают результаты исследований по ОТ и p24 и представлены на фигуре 6A, причем незакрашенные кружочки отмечают значения для Шв (100 TCID50) и закрашенные кружочки отмечают значения для MN (40 TCID50). Моноклональные антитела NM-01 подавляли образование синцития инфицированными MN клетками H9 при ID50 2 мкг/мл и инфицированными Шв клетками H9 при ID50 3 мкг/мл.
Соответствующие результаты подавления образования синцития для моноклональных антител ВАТ 123 представлены в таблице 3 WO 88/09181. Тогда как 25 мкг моноклональных антител NM-01 подавляет образование синцития клетками, инфицированными MN, на примерно 85%, 25 мкг ВАТ 123, как сообщалось, подавляют на 51%, и тогда как 25 мкг NМ-01 подавляют образование синцития инфицированными Шв клетками на примере 85%, ВАТ123, как сообщалось, подавляли на 77,8%. 25 мкг BAT123 так же, как сообщалось, подавляли образование синцития клетками, инфицированными RF на 51%. Моноклональные антитела NM-01 также подавляют образование синцития клетками, инфицированными RF (фигура 6B). В исследовании, описанном в предыдущем абзаце, концентрация 25 мкг NМ-01 подавляет около 59% образования синцития клетками, инфицированными RF. Моноклональные антитела NM-01 подавляли образование синцития инфицированными RF клетками при ID50 4 мкг/мл.
Объединенные результаты исследований по обратной транскриптазе, р. 24, МТ-2 и образования синцития примеров 4 и 5 показывают, что моноклональные антитела NM-01 нейтрализуют связывание и инфекционность различных штаммов ВИЧ-I при концентрациях менее чем 10 мкг/мл.
Пример 6
Чтобы подтвердить, что MAb NM-01 блокирует инфекционность MN и Шв ВИЧ-I путем связывания с частью петли V3 gp 120, были испытаны пептиды петли V3 на способность блокировать нейтрализацию антителами.
MAb NM-01 инкубировали с различными концентрациями пептидов, соответствующих петлям V3 штаммов MN, Шв и Z6 (последовательности пептидов даны в таблице 4) в течение 30 минут при 37oC перед добавлением 100 NCID50 живого вируса Шв. Затем добавляли клетки H9 на час, и активность ОТ определяли после подращивания клеток в полной среде в течение 7 дней, как описано в примере 4. Результаты исследования представлены на фигуре 7.
В то время как MAb NM-01 полностью нейтрализовали инфекционность Шв при наинизших концентрациях пептида, этот эффект прогрессивно блокировался при преинкубации с повышающимися концентрациями MN - (закрашенный кружочек) и Шв - (незакрашенный кружочек) пептидов петли. Не было определяемого эффекта со сходными концентрациями пептида, соответствующего петле штамма Z6 (закрашенный ромб), которая не имеет последовательности аминокислот, распознаваемой MAb NM-01. Эти результаты показывают, что моноклональные антитела NM-01 блокируют и ифекционность ВИЧ-1 путем реагирования со специфическим участком области V3 gp 120.
Пример 7
Проведены также исследования чтобы определить могут ли моноклональные антитела NM-01 активировать метаболизм комплемента и усиленно разрушать варионы ВИЧ-1.
В качестве источника комплемента была использована кроличья сыворотка.
Лизис ВИЧ-I с помощью MAb NМ-01 и комплемента
Клетки H9, инфицированные штаммом Шв ВИЧ-I, отмывали в среде на цитотоксичность (Cedarlane Lab. Ltd.). Клетки ресуспендировали в среде на цитотоксичность в отсутствие или с присутствием 40 мкг/мл MAb NM-01. После инкубации в течение 2 часов при 4oC добавляли кроличий комплемент (low-tox-MA; Cedarlane Lab. Ltd) в разведении 1:6. Клеточную суспензию инкубировали при 4oC в течение 20 минут и затем при 37oC в течение 45 минут. Клетки дважды фиксировали 2%-ным глютаральдегидом /0,1 М фосфатным буфером и 1% тетроксидом осмия /0,1 М фосфатным буфером. После помещения в эпоксидную смолу делались тонкие срезы и дважды окрашивались уранил-ацетатом и нитратом свинца. Фигуры 8,9 и 10 являются типичными электронными микрофотографиями этих тонких срезов.
Одна кроличья сыворотка (фигура 8B) и один MAb N М-01 не имели заметного действия на морфологию ВИЧ-I. Экспозиция ВИЧ-I с MAb NM-01 вместе с комплементом была связана с появлением многочисленных вирусных частиц с разрушенными оболочками и потерей электронной плотности сердцевины (фигура 8A). Типичные препараты показывают примерно 90% разрушенных вирионов, большинство из которых имели потерю внутреннего содержимого. Остальные 10% вирионов были интактными или имели частично разрушенные оболочки, более высокое увеличение выявило, что разрушение ВИЧ-I проявляется в виде прямого лизиса, как показано на сериях микрофотографий лизиса зрелых и неполных вирусных частиц на фигурах с 9А по F и с 10A по F соответственно.
Пример 8
Затем было определено действие комбинации моноклональных антител NМ-01 и комплемента на инфекционность ВИЧ-1.
Определение инфицирующей дозы для культуры ткани
Клетки H9, инфицированные ВИЧ-IIIIв , отмывали дважды в среде на цитотоксичность (Cedarlane Lab. Ltd.) и затем ресуспендировали в среде на цитотоксичность, содержащей 2 мкг/мл MAb NM-01 или контрольные IgG2. После инкубации при 4oC в течение 2 часов образцы разделяли на равные части и добавляли или кроличью сыворотку с комплементом, или кроличью сыворотку, инактивированную прогреванием (Cedarlane Lab. Ltd.) в разведении 1:6. Клетки инкубировали при 4oC в течение 20 минут и затем при 37oC в течение 45 минут отмывали средой, ресуспендировали в 50% ФБР /среде RPMI 1640 и взбалтывали. Супернатант или вирусный изолят разводили в 10 раз и затем серийно в 2 раза перед добавлением 25 мкл к клеткам H9 (1•105/25 мкл). После инкубации в течение 3 часов при 37oC выдерживаемые клетки разводили 10% ФБР /средой RPMI 1640 и выдерживали при 37oC. Вирусную инъекцию определяли через 6 дней путем определения обратной транскриптазы. Инфицирующую дозу для тканевой культуры для 50% образцов клеток H9 (TCID50) определяли по разведению, которое показывало инфекцию в 50% случаев. В таблице 5 представлены результаты этих экспериментов.
Тогда как одни антитела NM-01 способны нейтрализовать инфекционность ВИЧIIIв, воздействие вместе моноклональных антител NМ-01 и комплемента снижало инфекционность более чем в 10 раз. Эти результаты показывают, что экспозиция ВИЧ-1 с MAb NM-01 и комплементом вместе связана со значительным снижением вирусной инфекционности и дополнительно подтверждает роль MAb NM-01-опосредуемого комплемент-зависимого виролиза для терапии ВИЧ-1 инфекции.
Пример 9
Вариабельные области тяжелых и легких цепей моноклональных антител NМ-01 клонировали путем PCR и секвенировали. ДНК и производные аминокислотные последовательности тяжелых и легких цепей вариабельных областей NM-01 представлены в Посл. ИД NN 14 и 15 и Посл. ИД NN 16 и 17 соответственно. Нуклеотиды 1-21 и 334-363 Посл. ИД N 16 соответствуют праймерам (инициаторам)PCR, использованным для амплификации последовательностей легких цепей NМ-01 и нуклеотидов 1-27 и 385-402, Посл ИД N 14 соответствуют праймерам, использованным для амплификации последовательностей тяжелых цепей NM-01.
Первые 120 остатков аминокислотных последовательностей вариабельных областей тяжелых и легких цепей NM-01 также представлены на фигурах 11 и 12, соответственно, причем включенные аминокислоты являются участками, определяющими комплементарность (CDRs) антител, которые определяют специфичность связывания антитела.
На фигурах аминокислотную последовательность каждой тяжелой и легкой цепи сравнили с соответствующей аминокислотной последовательностью вариабельных областей моноклональных антител ВАТ-123, как сообщено Lion et al., выше вариабельный участок тяжелой цепи NM-01 отличен от такового ВАТ-123 по сорока четырем аминокислотам от общего числа сто двадцать. Вариабельные участки легкой цепи двух антител отличаются по двадцати четырем аминокислотам. Существенно, что три CDRs в тяжелой цепи (V-H) молекулы NM-01 отличны от последовательности ВАТ-123 на 44-50%, тогда как последовательности трех CDRs легких цепей (V-L) отличаются на 22-45%. Следовательно, из анализа структур NM-01 и BAT-123 ясно, что это два разных антитела.
Основываясь на информации о последовательности ДНК, представленной в Посл. ИД N: 14 и 16, по методу Tempest et al., Bio/TECHNOLOGY, 9, 266-271, (1991) и Riechmann et al., Nature, 322, 323-327 (1988), может быть получен "очеловеченный" вариант антител NМ-01. Резюмируя, последовательности ДНК, кодирующие шесть CDRs легких и тяжелых цепей моноклональных антител NМ-01, могут быть введены в структуры ДНК, кодирующие соответствующие человеческие структурные участки тяжелых и легких цепей. Полученные в результате структуры, кодирующие вариабельные области тяжелой и легкой цепей в целом, экспрессируются вместе в клетке с последовательностями ДНК, кодирующими константные области у человека, чтобы получить очеловеченные антитела со специфической связываемостью моноклональных антител NM-01.
Несмотря на то, что настоящее изобретение описано, с точки зрения предпочтительных путей использования, для специалистов понято, что могут иметь место изменения и усовершенствования. Поэтому, имеется в виду, что пункты прилагаемой заявки включают все такие равносильные изменения, которые входят в сферу действия этого изобретения, как заявлено.
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
(2) ИНФОРМАЦИЯ ПОСЛ ИД N: 1
(1) ХАРАКТЕРИСТИКИ ПОСЛЕДОВАТЕЛЬНОСТИ:
(A) ДЛИНА: 4 аминокислоты
(B) ТИП: аминокислотная
(C) ТОПОЛОГИЯ: линейная
(II) ТИП МОЛЕКУЛЫ: пептид
(XI) ОПИСАНИЕ ПОСЛЕДОВАТЕЛЬНОСТИ: ПОСЛ ИД N: 1 :
Gly Pro Gly Ary
1
(2) ИНФОРМАЦИЯ О ПОСЛ ИД N: 2 :
(1) ХАРАКТЕРИСТИКИ ПОСЛЕДОВАТЕЛЬНОСТИ:
(A) ДЛИНА: 15 аминокислот
(B) ТИП: аминокислотная
(D) ТОПОЛОГИЯ: линейная
(II) ТИП МОЛЕКУЛЫ: пептид
(XI) ОПИСАНИЕ ПОСЛЕДОВАТЕЛЬНОСТИ: ПОСЛ ИД N: 2 :
Cys thr Arg Asn Trp Asn Lys Arg Lys Arg Ile His Ile Gly
1 5 10
(2) ИНФОРМАЦИЯ О ПОСЛ ИД N 3:
(1) ХАРАКТЕРИСТИКИ ПОСЛЕДОВАТЕЛЬНОСТИ:
(A) ДЛИНА: 15 аминокислот
(B) ТИП: аминокислотная
(D)) ТОПОЛОГИЯ: линейная
(II) ТИП МОЛЕКУЛЫ: пептид
(XI) ОПИСАНИЕ ПОСЛЕДОВАТЕЛЬНОСТИ: ПОСЛ ИД N 3:
Arg Ile His Ile Cly Pro Gly Arg Aba Phe Tyr Thr Thr Thr Lys Asn
1 5 10 15
(2) ИНФОРМАЦИЯ О ПОСЛ ИД N 4:
(I) ХАРАКТЕРИСТИКИ ПОСЛЕДОВАТЕЛЬНОСТЕЙ:
(A) ДЛИНА: 15 аминокислот
(B) ТИП : аминокислотная
(C) ТОПОЛОГИЯ: линейная
(D) ТИП МОЛЕКУЛЫ: пептид
(XI) ОПИСАНИЕ ПОСЛЕДОВАТЕЛЬНОСТИ: ПОСЛ ИД: N 4
Tyr Thr Thr Lys Asn Ile Ile Thr Ile Arg Gln Ala His Cys
1 5 10 15
(2) ИНФОРМАЦИЯ О ПОСЛ ИД N 5:
(I) ХАРАКТЕРИСТИКИ ПОСЛЕДОВАТЕЛЬНОСТИ:
(A) ДЛИНА: 15 аминокислот
(B) ТИП: аминокислотная
(D) ТОПОЛОГИЯ: линейная
(II) ТИП МОЛЕКУЛЫ: пептид
(XI) ОПИСАНИЕ ПОСЛЕДОВАТЕЛЬНОСТИ: ПОСЛ ИД N 5:
Arg Ile His Ile Cly Pro Gly Arg Aba Phe Tyr Thr Thr Lys Asn
1 5 10 15
(2) ИНФОРМАЦИЯ О ПОСЛ ИД N 6:
(1) ХАРАКТЕРИСТИКИ ПОСЛЕДОВАТЕЛЬНОСТИ:
(A) ДЛИНА: 14 аминокислот
(B) ТИП: аминокислотная
(D) ТОПОЛОГИЯ: линейная
(II) ТИП МОЛЕКУЛЫ: пептид
(XI) ОПИСАНИЕ ПОСЛЕДОВАТЕЛЬНОСТИ: ПОСЛ ИД N 6 :
Ile Arg Ile Gly Pro Gly Arg Ala Phe Val Thr Ale Gly Lys
1 5 10
(2) ИНФОРМАЦИЯ О ПОСЛ ИД N: 7 :
(1) ХАРАКТЕРИСТИКИ ПОСЛЕДОВАТЕЛЬНОСТИ:
(А) ДЛИНА: 19 аминокислот
(В) ТИП: аминокислотная
(D) ТОПОЛОГИЯ: линейная
(II) ТИП МОЛЕКУЛЫ: пептид
(XI) ОПИСАНИЕ ПОСЛЕДОВАТЕЛЬНОСТИ: ПОСЛ ИД N: 7 :
Cys Asn Thr Arg Lys Ser Ile Lys Gly Pro Gly Arg Val Ile Tyr Ala
1 5 10 15
Thr Gly Glu
(2) ИНФОРМАЦИЯ О ПОСЛ ИД N 8:
(1) ХАРАКТЕРИСТИКИ ПОСЛЕДОВАТЕЛЬНОСТИ:
(А) ДЛИНА: 20 аминокислот
(В) ТИП: аминокислотная
(D) ТОПОЛОГИЯ: линейная
(II) ТИП МОЛЕКУЛЫ: пептид
(XI) ОПИСАНИЕ ПОСЛЕДОВАТЕЛЬНОСТИ: ПОСЛ ИД N 8:
Cys His Thr Arg Lys Arg Val Thr Leu Gly Pro Gly Arg Val Trp Tyr
1 5 10 15
Thr Thr Gly Glu
20
(2) ИНФОРМАЦИЯ О ПОСЛ ИД N 9:
(1) ХАРАКТЕРИСТИКИ ПОСЛЕДОВАТЕЛЬНОСТИ:
(А) ДЛИНА: 20 аминокислот
(В) ТИП: аминокислотная
(D) ТОПОЛОГИЯ: линейная
(II) ТИП МОЛЕКУЛЫ: пептид
(XI) ОПИСАНИЕ ПОСЛЕДОВАТЕЛЬНОСТИ: ПОСЛ ИД N: 9 :
Cys Asn Thr Lys Lys Gly Ile Ala Ile Gly Pro Gly Arg Thr Leu Tyr
1 5 10 15
Ala Arg Glu Lys
20
(2) ИНФОРМАЦИЯ О ПОСЛ ИД N 10:
(1) ХАРАКТЕРИСТИКИ ПОСЛЕДОВАТЕЛЬНОСТИ:
(А) ДЛИНА: 23 аминокислот
(В) ТИП: аминокислотная
(D) ТОПОЛОГИЯ: линейная
(II) ТИП МОЛЕКУЛЫ: пептид
(XI) ОПИСАНИЕ ПОСЛЕДОВАТЕЛЬНОСТИ: ПОСЛ ИД N 10:
Cys Asn Thr Arg Gln Ser Thr Pro Ile Gly Leu Gly Gln Ala Leu
1 5 10 15
Tyr Thr Arg Gly Arg Thr Lys
20
(2) ИНФОРМАЦИЯ О ПОСЛ ИД N 11:
(1) ХАРАКТЕРИСТИКИ ПОСЛЕДОВАТЕЛЬНОСТИ:
(А) ДЛИНА: 22 аминокислот
(В) ТИП: аминокислотная
(D) ТОПОЛОГИЯ: линейная
(II) ТИП МОЛЕКУЛЫ: пептид
(XI) ОПИСАНИЕ ПОСЛЕДОВАТЕЛЬНОСТИ: ПОСЛ ИД N 11:
Cys Asn Ile Arg Gln Arg Thr Ser Ile Gly Leu Gly Gln Ala Leu
1 5 10 15
Tyr Thr Thr Lys Thr Arg Ser
20
(2) ИНФОРМАЦИЯ О ПОСЛ ИД N 12:
(1) ХАРАКТЕРИСТИКИ ПОСЛЕДОВАТЕЛЬНОСТИ:
(А) ДЛИНА: 22 аминокислот
(В) ТИП: аминокислотная
(D) ТОПОЛОГИЯ: линейная
(II) ТИП МОЛЕКУЛЫ: пептид
(XI) ОПИСАНИЕ ПОСЛЕДОВАТЕЛЬНОСТИ: ПОСЛ ИД N: 12
Cys Asn Thr Arg Gln Arg Thr Pro Ile Gly Leu Gly Gln Ser Leu
1 5 10 15
Tyr Thr Thr Arg Ser Arg Ser
20
(2) ИНФОРМАЦИЯ О ПОСЛ ИД N 13:
(1) ХАРАКТЕРИСТИКИ ПОСЛЕДОВАТЕЛЬНОСТИ:
(А) ДЛИНА: 15 аминокислот
(В) ТИП: аминокислотная
(D) ТОПОЛОГИЯ: линейная
(II) ТИП МОЛЕКУЛЫ: пептид
(XI) ОПИСАНИЕ ПОСЛЕДОВАТЕЛЬНОСТИ: ПОСЛ ИД N 13:
Arg Ile Thr Lys Gly Pro Gly Arg Val Ile Val Ala Thr Gly Gln
1 5 10 15
(2) ИНФОРМАЦИЯ О ПОСЛ ИД N 14 (см. в конце описания):
(1) ХАРАКТЕРИСТИКИ ПОСЛЕДОВАТЕЛЬНОСТИ:
(A) ДЛИНА: 402 пары оснований
(B) ТИП: нуклеиновая кислота
(C) ЦЕПЬ: одинарная
(D) ТОПОЛОГИЯ: линейная
(II) ТИП МОЛЕКУЛЫ: ДНК
(IX) ОСОБЕННОСТИ:
(A) НАЗВАНИЕ/КЛЮЧ: CDS
(B) МЕСТОНАХОЖДЕНИЕ: 1..402
(2) ИНФОРМАЦИЯ О ПОСЛ ИД N 15 (см. в конце описания):
(1) ХАРАКТЕРИСТИКИ ПОСЛЕДОВАТЕЛЬНОСТИ:
(A) ДЛИНА: 134 аминокислоты
(B) ТИП: аминокислотная
(D) ТОПОЛОГИЯ: линейная
(II) ТИП МОЛЕКУЛЫ: протеин
(2) ИНФОРМАЦИЯ О ПОСЛ ИД N 16 (см. в конце описания):
(1) ХАРАКТЕРИСТИКИ ПОСЛЕДОВАТЕЛЬНОСТИ:
(A) ДЛИНА: 363 пары оснований
(B) ТИП: нуклеиновая кислота
(C) ЦЕПЬ: одинарная
(D) ТОПОЛОГИЯ: линейная
(II) ТИП МОЛЕКУЛЫ: ДНК
(IX) ОСОБЕННОСТИ:
(A) НАЗВАНИЕ/КЛЮЧ: CDS
(B) МЕСТОНАХОЖДЕНИЕ: 1..363
(2) ИНФОРМАЦИЯ О ПОСЛ ИД N 17 (см. в конце описания):
(1) ХАРАКТЕРИСТИКИ ПОСЛЕДОВАТЕЛЬНОСТИ:
(A) ДЛИНА: 121 аминокислота
(B) ТИП: аминокислотная
(D) ТОПОЛОГИЯ: линейная
(II) ТИП МОЛЕКУЛЫ: протеинП











Изобретение представляет моноклональное антитело NMO1, которое специфически иммуннореактивно с протеином ВИЧ-1 gp l20 иди gp l60 и характеризуется способностью нейтрализовать in vitro инфицирование клеток Н9 живыни штаммами ВИЧ-1. Антитела NMO1 выделяются гибридомной клеточной линией АТСС N НВ 10726. Фармацевтические композиции этого изобретения будут применяться при лечении людей, инфицированных ВНЧ-l, c помощью пассивной иммунизации. 4 с. п. ф-лы, 12 ил., 5 табл.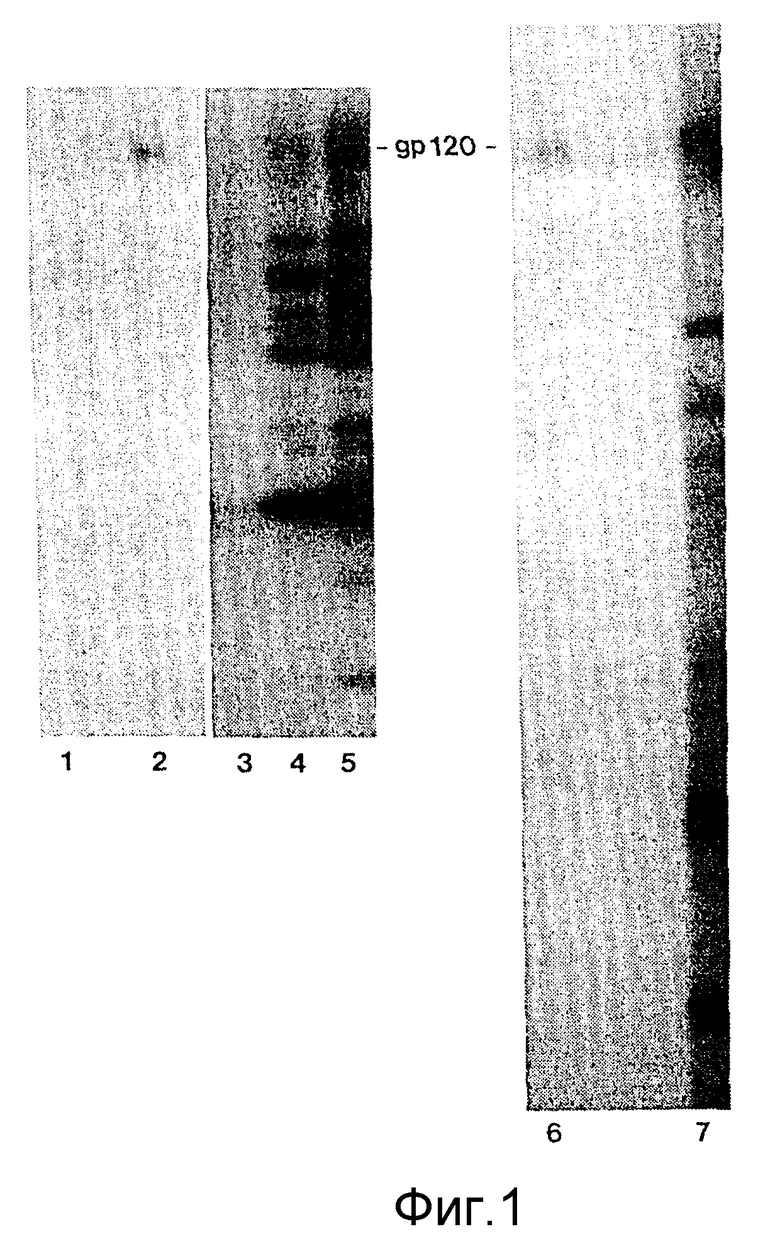



4. Фрагмент мышиного моноклонального антитела NM01, представляющий области, определяющие комплементарность (CDR) легкой цепи (V - L) указанного мышиного моноклонального антитела NM01, причем легкая цепь (V - L) содержит аминокислотную последовательность, где CDR имеют последовательности, заключенные в рамки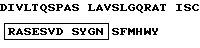
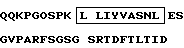

| WO 8809181, 01.12.88 | |||
| WO 9119797 A1, 26.12.91 | |||
| RU 2070926 C1, 27.12.96. |
