ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
[0001] По настоящей заявке испрашивается приоритет предварительной заявки США № 62/720,234, поданной 21 августа 2018 г., содержание которой включено в настоящее описание во всей своей полноте посредством ссылки.
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
[0002] Настоящая заявка содержит список последовательностей, представленный в формате ASCII через EFS-Web, который полностью включен в настоящее описание посредством ссылки. Копия ASCII, созданная 21 августа 2019 года в виде файла SequenceListing.txt размером 15,6 КБ.
УРОВЕНЬ ТЕХНИКИ
[0003] Описание всех публикаций, патентов, публикаций заявок на патент и книг, упомянутых в настоящем описании, полностью включены посредством ссылки в настоящую заявку для более полного описания области техники, к которой относится настоящее изобретение.
[0004] Т-клеточный белок-3, содержащий домены иммуноглобулина и муцина (Tim-3), также известный как клеточный рецептор 2 вируса гепатита А (HAVCR2), представляет собой трансмембранный белок типа I и является ингибирующим рецептором, который экспрессируется на Т-клетках, продуцирующих IFN-g, Foxp3+ Treg клетках и клетках врожденного иммунитета, таких как макрофаги и дендритные клетки. Tim-3 может подавлять иммунные ответы при взаимодействии со своими лигандами. Tim-3 содержит N-концевой иммуноглобулин-подобный домен (IgV), муциновый домен, содержащий сайты O-связанного гликозилирования, домен “стебель” (stalk), расположенный между муцином и трансмембранным доменом с сайтами для N-связанных сахаров, трансмембранный домен и цитоплазматический хвост (Monney 2002). Авторы изобретения ранее сообщали о том, что домен IgV Tim-3 является функциональным доменом, который может связывать углеводы (Cao 2007). Домен IgV Tim-3 содержит две неканонические дисульфидные связи и уникальный карман (щель) CC'-FG, который является характерной структурой, идентифицируемой во всех белках семейства Tim, но не присутствует в других членах суперсемейства иммуноглобулинов (Cao 2007). Также имеются сообщения о том, что адаптивная устойчивость к терапевтической блокаде PD-1 связана с активацией альтернативных иммунных контрольных точек, включая Tim-3 (Koyama 2016). В других сообщениях говорится о том, что помимо углеводов (Cao 2007; Wilker 2007), лигандами Tim-3 также являются другие молекулы, включая галектин-9 (Zhu 2005), фосфатидилсерин (DeKruyff 2010), HMGB1 (высокоподвижный блок группы 1) (Chiba 2012) и LILRB2 (лейкоцитарный иммуноглобулиноподобный рецептор В2) (публикация РСТ № WO16/111947).
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
[0005] Настоящее изобретение относится к антителу или его антигенсвязывающему фрагменту, которое связывается с доменом IgV TIM-3 человека (Т-клеточного белка-3, содержащего домены иммуноглобулина и муцина) или с IgV муцин-стебель TIM-3 человека, содержащее:
а) тяжелую цепь, содержащую одну или более из:
GYSFTGYTIN (SEQ ID NO: 1) (определяющую комплементарность область (CDR) 1 (CDR1));
LFNPYNGGTT (SEQ ID NO:2) (CDR2);
ARRYYGYDAMDY (SEQ ID NO:3) (CDR3);
или
GFNIKDYYMH (SEQ ID NO:7) (CDR1);
WIDPENDNTIY (SEQ ID NO:8) (CDR2);
ARDFGYVAWLVY (SEQ ID NO:9) (CDR3); и
b) легкую цепь, содержащую одну или более из:
KSSQSVLYSSNQKNHLA (SEQ ID NO:4) (CDR1);
WASTRES (SEQ ID NO:5) (CDR2);
HQYLSSYT (SEQ ID NO:6) (CDR3);
или
KASQNVDTAVA (SEQ ID NO:10) (CDR1);
SASNRYT (SEQ ID NO:11) (CDR2);
QQYSSYPT (SEQ ID NO:12) (CDR3).
[0006] Настоящее изобретение относится к антителу или его антигенсвязывающему фрагменту, которое связывается с доменом IgV TIM-3 человека или с IgV муцин-стебель TIM-3 человека, содержащее:
тяжелую цепь, содержащую
GYSFTGYTIN (SEQ ID NO:1) (CDR1);
LFNPYNGGTT (SEQ ID NO:2) (CDR2);
ARRYYGYDAMDY (SEQ ID NO:3) (CDR3); и
легкую цепь, содержащую
KSSQSVLYSSNQKNHLA (SEQ ID NO:4) (CDR1);
WASTRES (SEQ ID NO:5) (CDR2);
HQYLSSYT (SEQ ID NO:6) (CDR3).
[0007] Настоящее изобретение относится к антителу или его антигенсвязывающему фрагменту, которое связывается с доменом IgV TIM-3 человека или с IgV муцин-стебель TIM-3 человека, содержащее:
тяжелую цепь, содержащую
GFNIKDYYMH (SEQ ID NO:7) (CDR1);
WIDPENDNTIY (SEQ ID NO:8) (CDR2);
ARDFGYVAWLVY (SEQ ID NO:9) (CDR3); и
легкую цепь, содержащую
KASQNVDTAVA (SEQ ID NO:10) (CDR1);
SASNRYT (SEQ ID NO:11) (CDR2);
QQYSSYPT (SEQ ID NO:12) (CDR3).
[0008] Настоящее изобретение относится к выделенному антителу, которое связывается с доменом IgV TIM-3 человека или IgV муцин-стебель TIM-3 человека, со сродством KD=10,0 нМ или сильнее.
[0009] Раскрыта нуклеиновая кислота, кодирующая тяжелую цепь антитела, содержащую одну или более из:
GYSFTGYTIN (SEQ ID NO:1) (CDR1);
LFNPYNGGTT (SEQ ID NO:2) (CDR2);
ARRYYGYDAMDY (SEQ ID NO:3) (CDR3).
[0010] Раскрыта нуклеиновая кислота, кодирующая тяжелую цепь антитела, содержащую одну или более из:
GFNIKDYYMH (SEQ ID NO:7) (CDR1);
WIDPENDNTIY (SEQ ID NO:8) (CDR2);
ARDFGYVAWLVY (SEQ ID NO:9) (CDR3).
[0011] Раскрыта нуклеиновая кислота, кодирующая легкую цепь антитела, содержащую одну или более из:
KSSQSVLYSSNQKNHLA (SEQ ID NO:4) (CDR1);
WASTRES (SEQ ID NO:5) (CDR2);
HQYLSSYT (SEQ ID NO:6) (CDR3).
[0012] Раскрыта нуклеиновая кислота, кодирующая легкую цепь антитела, содержащую одну или более из:
KASQNVDTAVA (SEQ ID NO:10) (CDR1);
SASNRYT (SEQ ID NO:11) (CDR2)
QQYSSYPT (SEQ ID NO:12) (CDR3).
[0013] Раскрыта клетка-хозяин, содержащая одну или более нуклеиновых кислот, представленные в настоящем описании.
[0014] В настоящем описании раскрыто антитело или его фрагмент, связанное или конъюгированное с терапевтическим агентом.
[0015] Раскрыт способ ингибирования TIM-3 человека у индивида, включающий введение представленного в настоящем описании антитела или его фрагмента в количестве, эффективном для ингибирования TIM-3 человека.
[0016] Раскрыт способ ингибирования опосредованной Tim-3 супрессии Т-клеток у индивида, включающий введение представленного в настоящем описании антитела или его фрагмента в количестве, эффективном для ингибирования опосредованной Tim-3 супрессии Т-клеток.
[0017] Раскрыт способ лечения злокачественного заболевания у индивида, включающий введение представленного в настоящем описании антитела или его фрагмента в количестве, эффективном для лечения злокачественного заболевания у индивида.
[0018] Раскрыт способ детектирования у индивида клетки, положительной по TIM-3 человека, включающий введение представленного в настоящем описании антитела или его фрагмента, несущего конъюгированный с ним детектируемый маркер, в количестве, эффективном для мечения клетки, положительной по TIM-3 человека, и затем обнаружение метки у индивида, таким образом детектируя у индивида клетку, положительную по TIM-3 человека.
[0019] Предлагается выделенная молекула нуклеиновой кислоты, кодирующая представленное в настоящем описании антитело или его фрагмент. В одном из вариантов осуществления нуклеиновая кислота представляет собой ДНК. В одном из вариантов осуществления нуклеиновая кислота представляет собой кДНК. В одном из вариантов осуществления нуклеиновая кислота представляет собой РНК.
[0020] В некоторых вариантах осуществления изобретения антигенсвязывающий фрагмент антитела представляет собой фрагмент антитела, связывающий домен IgV Tim-3. В некоторых вариантах осуществления изобретения антигенсвязывающий фрагмент антитела представляет собой фрагмент антитела, связывающий домен IgV TIM-3 человека.
[0021] Раскрыт вектор, кодирующий молекулу нуклеиновой кислоты, раскрытую в настоящем описании. Раскрыта клетка-хозяин, содержащая раскрытую в настоящем описании молекулу нуклеиновой кислоты или раскрытый в настоящем описании вектор.
[0022] Раскрыт способ получения антитела к домену IgV Tim-3 или его фрагмента, связывающего домен IgV Tim-3, включающий культивирование клетки-хозяина, раскрытой в настоящем описании, в условиях, при которых антитело к домену IgV Tim-3 или его фрагмент, связывающий домен Tim-3 IgV, продуцируется клеткой-хозяином.
[0023] Раскрыта фармацевтическая композиция, содержащая представленное в настоящем описании антитело к домену IgV Tim-3 или его фрагмент, связывающий домен IgV Tim-3, и фармацевтически приемлемый наполнитель.
[0024] Раскрыт способ снижения активности Tim-3 у нуждающегося в этом индивида, включающий введение указанному индивиду терапевтически эффективного количества представленного в настоящем описании антитела к домену IgV Tim-3 или его фрагмента, связывающего домен IgV Tim-3.
КРАТКОЕ ОПИСАНИЕ РИСУНКОВ
[0025] Фиг. 1. Результаты ELISA показывают, что 50B5 связывается с белком Tim-3-IgV и всей внеклеточной частью белка Tim-3 (IgV-муцин-стебель), а не с контрольным белком человеческого IgG.
[0026] Фиг. 2. Результаты FACS показывают, что 50B5 связывается с клеточными линиями, экспрессирующими человеческий Tim-3, а не мышиный Tim-3. 50B5 (незаштрихованные гистограммы) или контрольный мышиный изотип IgG1 (заштрихованные гистограммы)
[0027] Фиг. 3. Кинетические параметры для 50B5, определенные методом поверхностного плазмонного резонанса.
[0028] Фиг. 4A-4B. 4A: в анализе реакции смешанной культуры лимфоцитов подвергнутые делению CD4 и CD8 Т-клетки, но не клетки CD4 и CD8 T-клетки, которые не были подвергнуты делению, экспрессировали Tim-3 на 4 день. 4B: в анализе реакции смешанной культуры лимфоцитов mAb 50B5 приводило к существенному увеличению продуцирования IFN-g в CD4 и CD8 T-клетках на 4-й день; N=8; **P<0,01
[0029] Фиг. 5. Результаты FACS показывают, что 15B4 связывается с линией клеток, экспрессирующих человеческий Tim-3. 15B4 (незаштрихованные гистограммы) или контрольный мышиный изотип IgG1 (заштрихованные гистограммы).
[0030] Фиг. 6. Кинетические параметры, полученные методом поверхностного плазмонного резонанса.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0031] Раскрыто антитело или его антигенсвязывающий фрагмент, которое связывается с доменом IgV TIM-3 человека (Т-клеточного белка-3, содержащего домены иммуноглобулина и муцина) или IgV муцин-стебель TIM-3 человека, содержащее:
а) тяжелую цепь, содержащую одну или более из:
GYSFTGYTIN (SEQ ID NO:1) (CDR1);
LFNPYNGGTT (SEQ ID NO:2) (CDR2);
ARRYYGYDAMDY (SEQ ID NO:3) (CDR3);
или
GFNIKDYYMH (SEQ ID NO:7) (CDR1);
WIDPENDNTIY (SEQ ID NO:8) (CDR2);
ARDFGYVAWLVY (SEQ ID NO:9) (CDR3); и
b) легкую цепь, содержащую одну или более из:
KSSQSVLYSSNQKNHLA (SEQ ID NO:4) (CDR1);
WASTRES (SEQ ID NO:5) (CDR2);
HQYLSSYT (SEQ ID NO:6) (CDR3);
или
KASQNVDTAVA (SEQ ID NO:10) (CDR1);
SASNRYT (SEQ ID NO:11) (CDR2);
QQYSSYPT (SEQ ID NO:12) (CDR3).
[0032] Раскрыто антитело или его антигенсвязывающий фрагмент, которое связывается с доменом IgV TIM-3 человека или IgV муцин-стебель TIM-3 человека, содержащее:
тяжелую цепь, содержащую
GYSFTGYTIN (SEQ ID NO:1) (CDR1);
LFNPYNGGTT (SEQ ID NO:2) (CDR2);
ARRYYGYDAMDY (SEQ ID NO:3) (CDR3); и
легкую цепь, содержащую
KSSQSVLYSSNQKNHLA (SEQ ID NO:4) (CDR1);
WASTRES (SEQ ID NO:5) (CDR2);
HQYLSSYT (SEQ ID NO:6) (CDR3).
[0033] Раскрыто антитело или его антигенсвязывающий фрагмент, которое связывается с доменом IgV TIM-3 человека или IgV муцин-стебель TIM-3 человека, и содержит:
тяжелую цепь, содержащую
GFNIKDYYMH (SEQ ID NO:7) (CDR1);
WIDPENDNTIY (SEQ ID NO:8) (CDR2);
ARDFGYVAWLVY (SEQ ID NO:9) (CDR3); и
легкую цепь, содержащую
KASQNVDTAVA (SEQ ID NO:10) (CDR1);
SASNRYT (SEQ ID NO:11) (CDR2);
QQYSSYPT (SEQ ID NO:12) (CDR3).
[0034] В некоторых вариантах осуществления изобретения каркасные области легкой цепи и тяжелой цепи являются каркасными областями человека или идентичны ей на 85% или более.
[0035] В некоторых вариантах осуществления изобретения каркасные области легкой цепи и тяжелой цепи представляют собой человеческие каркасные области. Предлагается выделенное антитело или его антигенсвязывающий фрагмент, которое связывается с доменом IgV TIM-3 человека или IgV муцин-стебель TIM-3 человека, со сродством KD=10,0 нМ или сильнее.
[0036] В некоторых вариантах осуществления изобретения антитело связывается с доменом IgV TIM-3 человека или IgV муцин-стебель TIM-3 человека, со сродством KD=1,0 нМ или сильнее. В некоторых вариантах осуществления изобретения антитело связывается с доменом IgV TIM-3 человека или IgV муцин-стебель TIM-3 человека, со сродством KD=0,5 нМ или сильнее.
[0037] В некоторых вариантах осуществления изобретения антитело не связывается с мышиным Tim-3.
[0038] В некоторых вариантах осуществления изобретения антитело ингибирует связывание TIM-3 человека с фосфатидилсерином, экспрессируемом на обработанных дексаметазоном Т-клетках Jurkat.
[0039] В некоторых вариантах осуществления изобретения выделенное антитело или его антигенсвязывающий фрагмент имеет Fc-область с человеческой последовательностью.
[0040] В некоторых вариантах осуществления изобретения выделенное антитело или его антигенсвязывающий фрагмент содержит тяжелую цепь, содержащую одну или более из:
GYSFTGYTIN (SEQ ID NO:1) (CDR1)
LFNPYNGGTT (SEQ ID NO:2) (CDR2)
ARRYYGYDAMDY (SEQ ID NO:3) (CDR3).
[0041] В некоторых вариантах осуществления изобретения выделенное антитело или его антигенсвязывающий фрагмент содержит легкую цепь, содержащую одну или более:
KSSQSVLYSSNQKNHLA (SEQ ID NO:4) (CDR1)
WASTRES (SEQ ID NO:5) (CDR2)
HQYLSSYT (SEQ ID NO:6) (CDR3).
[0042] В некоторых вариантах осуществления изобретения выделенное антитело или его антигенсвязывающий фрагмент содержит тяжелую цепь, содержащую одну или более:
GFNIKDYYMH (SEQ ID NO:7) (CDR1)
WIDPENDNTIY (SEQ ID NO:8) (CDR2)
ARDFGYVAWLVY (SEQ ID NO:9) (CDR3).
[0043] В некоторых вариантах осуществления изобретения выделенное антитело или его антигенсвязывающий фрагмент содержит легкую цепь, содержащую одну или более:
KASQNVDTAVA (SEQ ID NO:10) (CDR1)
SASNRYT (SEQ ID NO:11) (CDR2)
QQYSSYPT (SEQ ID NO:12) (CDR3).
[0044] В некоторых вариантах осуществления изобретения выделенное антитело или его антигенсвязывающий фрагмент является химерным или гуманизированным.
[0045] В некоторых вариантах осуществления изобретения выделенное антитело или его антигенсвязывающий фрагмент выбрано из группы, состоящей из моноклонального антитела, scFv, Fab фрагмента, Fab' фрагмента и F(ab)' фрагмента. Следует отметить, что хотя scFv не является фрагментом антитела в буквальном смысле, а скорее представляет собой слитый белок, в контексте настоящего описания фрагмент антитела включает scFv, если не указано иное.
[0046] Раскрыта нуклеиновая кислота, кодирующая тяжелую цепь антитела, которая содержит одну или более из:
GYSFTGYTIN (SEQ ID NO:1) (CDR1);
LFNPYNGGTT (SEQ ID NO:2) (CDR2);
ARRYYGYDAMDY (SEQ ID NO:3) (CDR3).
[0047] Раскрыта нуклеиновая кислота, кодирующая тяжелую цепь антитела, которая содержит одну или более из:
GFNIKDYYMH (SEQ ID NO:7) (CDR1);
WIDPENDNTIY (SEQ ID NO:8) (CDR2);
ARDFGYVAWLVY (SEQ ID NO:9) (CDR3).
[0048] Раскрыта нуклеиновая кислота, кодирующая легкую цепь антитела, которая содержит одну или более из:
KSSQSVLYSSNQKNHLA (SEQ ID NO:4) (CDR1);
WASTRES (SEQ ID NO:5) (CDR2);
HQYLSSYT (SEQ ID NO:6) (CDR3).
[0049] Раскрыта нуклеиновая кислота, кодирующая легкую цепь антитела, которая содержит одну или более из:
KASQNVDTAVA (SEQ ID NO:10) (CDR1);
SASNRYT (SEQ ID NO:11) (CDR2)
QQYSSYPT (SEQ ID NO:12) (CDR3).
[0050] Раскрыта клетка-хозяин, содержащая одну или более нуклеиновых кислот, раскрытых в настоящем описании.
[0051] Представленное в настоящем описании антитело или его фрагмент связано или конъюгировано с терапевтическим агентом.
[0052] В некоторых вариантах осуществления изобретения терапевтический агент представляет собой цитотоксическое лекарственное вещество, радиоактивный изотоп, иммуномодулятор или второе антитело.
[0053] Раскрыт способ ингибирования TIM-3 человека у индивида, включающий введение определенного представленного в настоящем описании антитела или его фрагмента, или его фрагмента, связывающего человеческий Tim3, в количестве, эффективном для ингибирования TIM-3 человека.
[0054] В некоторых вариантах осуществления изобретения индивид болен раком.
[0055] Раскрыт способ ингибирования опосредованной Tim-3 супрессии Т-клеток у индивида, включающий введение представленного в настоящем описании антитела или его фрагмента, или его фрагмента, связывающего Tim3 человека, в количестве эффективном для ингибирования опосредованной Tim-3 суппрессии Т-клеток.
[0056] Раскрыт способ лечения злокачественного заболевания у индивида, включающий введение представленного в настоящем описании антитела или его фрагмента в количестве, эффективном для лечения злокачественного заболевания у индивида.
[0057] В некоторых вариантах осуществления изобретения злокачественное заболевание представляет собой Tim-3-положительный злокачественное заболевание человека.
[0058] Раскрыт способ обнаружения у индивида клетки, положительной по TIM-3 человека, включающий введение представленного в настоящем описании антитела или его фрагмента, несущего конъюгированный с ним детектируемый маркер, в количестве, эффективном для мечения клетки, положительной по TIM-3 человека, и затем обнаружение метки у индивида, таким образом детектируя у индивида клетку, положительную по TIM-3 человека.
[0059] В некоторых вариантах осуществления изобретения метку детектируют путем визуализации.
[0060] В некоторых вариантах осуществления изобретения клетка представляет собой раковую клетку.
[0061] В некоторых вариантах осуществления изобретения злокачественное заболевание представляет собой гематологическое злокачественное новообразование. В некоторых вариантах осуществления изобретения злокачественное заболевание представляет собой рак легких, желудка, головы или шеи, шванному, меланому или фолликулярную B-клеточную неходжкинскую лимфому.
[0062] В некоторых вариантах осуществления изобретения злокачественное заболевание включает солидную опухоль.
[0063] В некоторых вариантах осуществления изобретения индивид получает анти-PD-1 или анти-PD-L1, или анти-CTLA4 терапию.
[0064] В некоторых вариантах осуществления изобретения антитело к домену IgV Tim-3 или его фрагмент, связывающий домен IgV Tim-3, содержит (i) каркасную область VH, содержащую последовательность каркасной области человеческой зародышевой линии IGHV1-2*02, IGHV1-2*04, IGHV1-2*05, IGHV1-18*04, IGHV1-69-2*01, IGHV1-46*01, IGHD5-12*01, IGHD5-24*01, IGHD6-25*01, IGHJ3*01, IGHJ4*01, IGHJ4*03, IGHJ6*01, IGHJ6*02 и/или (ii) каркасную область VL, содержащую последовательность каркасной области человеческой зародышевой линии IGKV1-13*02, IGKV1-27*01, IGKV3-7*02, IGKV4-1*01, IGKV1D-13*02, IGKV3D-7*01, IGKJ1*01, IGKJ2*01, IGKJ4*01, IGKJ4*02.
[0065] В некоторых вариантах осуществления изобретения антитело к домену IgV Tim-3 или его фрагмент, связывающий домен IgV Tim-3, связывается с доменом IgV Tim-3 со сродством связывания (KD) от примерно 1×10-9 М до примерно 1×10-12 М.
[0066] В некоторых вариантах осуществления изобретения антитело к домену IgV Tim-3 или его фрагмент, связывающий домен IgV Tim-3, представляет собой моноклональное антитело.
[0067] В некоторых вариантах осуществления изобретения антитело к домену IgV Tim-3 или его фрагмент, связывающий домен IgV Tim-3, представляет собой рекомбинантное антитело.
[0068] В некоторых вариантах осуществления изобретения антитело к домену IgV Tim-3 или его фрагмент, связывающий домен IgV Tim-3, имеет человеческую каркасную область.
[0069] В некоторых вариантах осуществления изобретения антитело к домену IgV Tim-3 или его фрагмент, связывающий домен IgV Tim-3, имеет человеческую константную область.
[0070] В некоторых вариантах осуществления изобретения раскрыто антитело к домену IgV Tim-3.
[0071] В некоторых вариантах осуществления изобретения предлагается фрагмент антитела, связывающий домен IgV Tim-3.
[0072] В некоторых вариантах осуществления изобретения фрагмент, связывающий домен IgV Tim-3, представляет собой Fab, F(ab)2 или scFv.
[0073] Предлагается выделенная молекула нуклеиновой кислоты, кодирующая представленное в настоящем описании антитело к домену IgV Tim-3 или его фрагмент, связывающий домен IgV Tim-3. В одном из вариантов осуществления нуклеиновая кислота представляет собой ДНК. В одном из вариантов осуществления нуклеиновая кислота представляет собой кДНК. В одном из вариантов осуществления нуклеиновая кислота представляет собой РНК.
[0074] Предлагается вектор, кодирующий молекулу нуклеиновой кислоты, раскрытую в настоящем описании. Раскрыта клетка-хозяин, содержащая раскрытую в настоящем описании молекулу нуклеиновой кислоты или раскрытый в настоящем описании вектор.
[0075] Раскрыт способ получения антитела к домену IgV Tim-3 или его фрагмента, связывающего домен IgV Tim-3, включающий культивирование клетки-хозяина, раскрытой в настоящем описании, в условиях, в которых клетка-хозяин продуцирует антитело к домену IgV Tim-3 или фрагмент, связывающий домен IgV Tim-3.
[0076] Предлагается композиция, содержащая представленное в настоящем описании антитело к домену IgV Tim-3 или его фрагмент, связывающий домен IgV Tim-3, и носитель. В одном из вариантов осуществления композиция представляет собой фармацевтическую композицию, и носитель представляет собой фармацевтический носитель. Также предлагается фармацевтическая композиция, содержащая представленное в настоящем описании антитело или его связывающий фрагмент, и фармацевтически приемлемый наполнитель.
[0077] В контексте настоящего описания, термин «антитело» относится к интактному антителу, т.е. с полными Fc и Fv областями. «Фрагмент» относится к любой части антитела или частям антитела, связанным вместе, таким как, без ограничения, Fab, F(ab)2, одноцепочечный Fv (scFv), причем эта часть меньше, чем целое антитело, но является антигенсвязывающей частью и конкурирует с интактным антителом, фрагментом которого оно является, за специфическое связывание. В этом случае антиген представляет собой человеческий домен IgV Tim-3.
[0078] Такие фрагменты могут быть получены, например, путем расщепления интактного антитела или рекомбинантными способами. Для общего сведения см. Fundamental Immunology, Ch. 7 (Paul, W., ed., 2nd ed. Raven Press, N.Y. (1989), полностью включенную в настоящее описание посредством ссылки). Антигенсвязывающие фрагменты могут быть получены методами рекомбинантной ДНК или ферментативным или химическим расщеплением интактных антител или методами молекулярной биологии. В некоторых вариантах осуществления изобретения фрагмент представляет собой Fab, Fab', F(ab')2, Fd, Fv, фрагмент определяющей комплементарность области (CDR), одноцепочечное антитело (scFv), (вариабельный домен легкой цепи (VL) и вариабельный домен тяжелой цепи (VH), связанные пептидным линкером). В одном из вариантов осуществления scFv содержит каркасную последовательность вариабельного домена, имеющую последовательность, идентичную FR1, FR2, FR3 или FR4 человеческого вариабельного домена. В одном из вариантов осуществления scFv содержит линкерный пептид длиной от 5 до 30 аминокислотных остатков. В одном из вариантов осуществления scFv содержит линкерный пептид, содержащий один или более остатков глицина, серина и треонина.
[0079] В одном из вариантов осуществления линкер scFv имеет длину 10-25 аминокислот. В одном из вариантов осуществления пептидный линкер содержит остатки глицина, серина и/или треонина (см., например, Bird 1988 и Huston 1988, оба из которых полностью включены в настоящее описание посредством ссылки) или полипептид, который содержит по меньшей мере часть антитела, достаточную для связывания антигена, специфического к IgV домену Tim-3, на полипептиде, включая диатело. Вариабельные домены как зрелой легкой, так и тяжелой цепей содержат, в направлении от N-конца к C-концу, области FR1, CDR1, FR2, CDR2, FR3, CDR3 и FR4. Отнесение аминокислот к каждому домену осуществляется в соответствии с определениями, описанными Kabat, Sequences of Proteins of Immunological Interest (National Institutes of Health, Bethesda, MD (1987 и 1991)), Chothia & Lesk 1987 или Chothia 1989, каждый из этих документов полностью включен в настоящее описание посредством ссылки. В контексте настоящего описания термин «полипептид» включает нативные или искусственные белки, фрагменты белков и аналоги полипептидов белковой последовательности. Полипептид может быть мономерным или полимерным. В контексте настоящего описания Fd-фрагмент означает фрагмент антитела, который состоит из доменов VH и CH1; Fv-фрагмент состоит из доменов V1 и VH одного плеча антитела; и dAb фрагмент (Ward 1989, полностью включен в настоящее описание посредством ссылки) состоит из домена VH. В некоторых вариантах осуществления фрагменты имеют длину по меньшей мере 5, 6, 8 или 10 аминокислот. В других вариантах осуществления фрагменты имеют длину по меньшей мере 14, по меньшей мере 20, по меньшей мере 50 или по меньшей мере 70, 80, 90, 100, 150 или 200 аминокислот.
[0080] Термин «моноклональное антитело», используемый в настоящем описании, относится к антителу, члену популяции по существу гомогенных антител, т.е. отдельные антитела, составляющие популяцию, идентичны, за исключением возможных мутаций, например, встречающихся в природе мутаций, которые могут присутствовать в незначительных количествах. Таким образом, модификатор «моноклональное» характеризует антитело, как не являющееся смесью отдельных антител. В некоторых вариантах осуществления такое моноклональное антитело обычно включает антитело, содержащее полипептидную последовательность, которая связывается с антителом, содержащим домен IgV TIM-3 человека. В отличие от препаратов поликлональных антител, которые обычно включают разные антитела, направленные против разных детерминант (эпитопов), каждое моноклональное антитело в препарате моноклональных антител направлено против одной детерминанты антигена. Помимо специфичности, преимущество препаратов моноклональных антител заключается в том, что они обычно не загрязнены другими иммуноглобулинами. Таким образом, идентифицированное моноклональное антитело может быть получено методами, не связанными с получением гибридом, например подходящими рекомбинантными средствами после идентификации его последовательности.
[0081] В варианте осуществления изобретений, раскрытых в настоящем описании, антитело выделяют. В контексте настоящего описания термин «выделенное антитело» относится к антителу, которое в силу своего происхождения или источника происхождения имеет одну, две, три или четыре из следующих характеристик: (1) не связано с компонентами, сопровождающими его и связанными с ним в естественных условиях в естественном состоянии, (2) не содержит других белков того же вида, (3) экспрессируется клеткой другого вида и (4) не встречается в природе без вмешательства человека.
[0082] В одном из вариантов осуществления антитело является гуманизированным. «Гуманизированные» формы нечеловеческих (например, мышиных) антител представляют собой химерные антитела, которые содержат минимальную последовательность, полученную из нечеловеческого иммуноглобулина. В одном из вариантов осуществления гуманизированное антитело представляет собой человеческий иммуноглобулин (реципиентное антитело), в котором остатки гипервариабельной области (HVR) (или CDR) реципиента заменены остатками с требуемой специфичностью, сродством и/или способностью из HVR (или CDR) не относящегося к человеку вида (донорное антитело), такого как мышь, крыса, кролик или не относящийся к человеку примат. В одном из вариантов осуществления антитело имеет 1, 2, 3, 4, 5 или все 6 CDR1-3 как тяжелой, так и легкой цепей мышиных антител, раскрытых в настоящем описании (mAb 15B4 и mAb 50B5). В предпочтительном варианте осуществления остатки каркасной области (FR) мышиного mAb заменены соответствующими остатками каркасной области вариабельного домена (FR) человеческого иммуноглобулина. Они могут быть дополнительно модифицированы в некоторых вариантах осуществления для дальнейшего улучшения характеристик антител. Кроме того, в конкретном варианте осуществления гуманизированные антитела могут содержать остатки, которые отсутствуют в антителе-реципиенте или в антителе-доноре. В одном из вариантов осуществлении гуманизированные антитела не содержат остатки, которые не обнаружены в антителе-реципиенте или в антителе-доноре. Обычно, гуманизированное антитело содержит по существу все, по меньшей мере один, и обычно два вариабельных домена, в которых все, или в некоторых вариантах осуществления по существу все, гипервариабельные петли соответствуют петлям иммуноглобулина, который не является человеческим иммуноглобулином (например, mAb 15B4 и mAb 50B5, раскрытые в настоящем описании), и все, или в некоторых вариантах осуществления по существу все, FR происходят из последовательности человеческого иммуноглобулина. Гуманизированное антитело также необязательно может содержать по меньшей мере часть константной области (Fc) иммуноглобулина, обычно человеческого иммуноглобулина. См., например, Jones 1986; Riechmann 1988; Presta 1992; Vaswani & Hamilton 1998; Harris 1995; Hurle & Gross 1994; и патенты США №№ 6,982,321 и 7,087,409; 6,982,321 и 7,087,409, содержание каждого из которых полностью включено в настоящее описание посредством ссылки. В одном из вариантов осуществления, в котором гуманизированные антитела содержат остатки, которые не обнаружены в реципиентном антителе или в донорном антителе, Fc-области антител модифицированы, как описано в WO 99/58572, содержание которого включено в настоящее описание во всей своей полноте посредством ссылки.
[0083] Методы гуманизации моноклональных антител хорошо известны и описаны, например, в патентах США №№ 4,816,567; 5,807,715; 5,866,692; 6,331,415; 5,530,101; 5,693,761; 5,693,762; 5,585,089; и 6,180,370, содержание каждого из которых включено в настоящее описание во всей своей полноте посредством ссылки. Описано несколько «гуманизированных» молекул антител, содержащих антигенсвязывающий участок, полученный из нечеловеческого иммуноглобулина, включая антитела, имеющие V-области или модифицированные V-области грызунов и связанные с ними определяющие комплементарность области (CDR), слитые с человеческими константными доменами. См., например, Winter 1991, Lobuglio 1989, Shaw 1987 и Brown 1987, содержание каждой из которых включено в настоящее описание во всей полноте посредством ссылки. В других источниках описаны гипервариабельные области или CDR грызунов, трансплантированные в человеческую поддерживающую каркасную область (FR) перед слиянием с подходящим константным доменом человеческого антитела. См., например, Riechmann 1988, Verhoeyen 1988 и Jones 1986, содержание каждой из которых включено в настоящее описание во всей полноте посредством ссылки. В другой ссылке описаны CDR грызунов, поддерживаемые рекомбинантно венированными каркасными областями грызунов, см. публикацию Европейского патента № 0519596 (полностью включена посредством ссылки). Эти «гуманизированные» молекулы предназначены для минимизации нежелательного иммунологического ответа на молекулы античеловеческих антител грызунов, который ограничивает продолжительность и эффективность терапевтического применения этих фрагментов у людей-реципиентов. Константная область антитела может быть сконструирована таким образом, чтобы она была иммунологически инертной (например, не запускала лизис комплемента). См., например, публикацию PCT № WO99/58572; заявку на патент Великобритании № 9809951.8. Другие методы гуманизации антител, которые также можно использовать, раскрыты в Daugherty 1991; патентах США № 6,180,377; 6,054,297; 5,997,867; 5,866,692; 6210671; и 6,350,861; и публикации PCT № WO01/27160, каждый из этих документов полностью включен в настоящее описание посредством ссылки.
[0084] Другие формы гуманизированных антител имеют одну или более, или все CDR (CDR L1, CDR L2, CDR L3, CDR H1, CDR H2 или CDR H3), которые изменены относительно исходного антитела и которые также упоминаются как одна или более CDR, «происходящие из» одной или более CDR исходного антитела. В одном из вариантов осуществления антитело по изобретению представляет собой гуманизированное mAb 15B4 и mAb 50B5 с одной или более, или всеми CDR (CDR L1, CDR L2, CDR L3, CDR H1, CDR H2 или CDR H3), которые изменены относительно исходного антитела. В одном из вариантов осуществления антитело по изобретению представляет собой гуманизированные mAb 15B4 и mAb 50B5, в которых ни одна из CDR (CDR L1, CDR L2, CDR L3, CDR H1, CDR H2 или CDR H3) не изменена относительно исходного антитела.
[0085] В некоторых вариантах осуществления настоящего изобретения антитела или фрагменты могут быть получены рекомбинантным способом, такие как, антитела, экспрессируемые рекомбинантным вектором экспрессии, трансфицированным в клетку-хозяина, антитела, выделенные из рекомбинантной комбинаторной библиотеки человеческих антител, антитела, выделенные из животного (например, мыши), которое является трансгенным по генам человеческого иммуноглобулина.
[0086] В одном из вариантов осуществления антитело к домену IgV Tim-3, представленное в настоящем описании, способно специфически связываться или специфически связывается с доменом IgV TIM-3 человека. В контексте настоящего описания термин «способен специфически связываться» или «специфически связывается» относится к свойству антитела или фрагмента связываться с (конкретным) антигеном с константой диссоциации, составляющей <1 мкМ, предпочтительно <1 нМ и наиболее предпочтительно <10 пМ. В одном из вариантов осуществления Kd антитела (или фрагмента) к домену IgV Tim-3 лучше, чем 10,0 нМ. В одном из вариантов осуществления Kd антитела (или фрагмента) к IgV-домену Tim-3 лучше, чем 1,0 нМ. В одном из вариантов осуществления Kd антитела (или фрагмента) к IgV-домену Tim-3 лучше, чем 0,5 нМ. В одном из вариантов осуществления Kd антитела (или фрагмента) к домену IgV Tim-3 составляет 0,1 нМ или сильнее.
[0087] Термин «Kd», используемый в настоящем описании, означает константу диссоциации взаимодействия антитело-антиген. Одним из способов определения Kd или сродства связывания антител с доменом IgV Tim-3 является измерение сродства связывания монофункциональных Fab-фрагментов антитела. (Константа сродства является обратной величиной константы диссоциации). Для получения монофункциональных Fab-фрагментов антитело (например, IgG) можно расщеплять папаином или экспрессировать рекомбинантно. Сродство фрагмента антитела к домену IgV TIM-3 человека может быть определено, например, методом поверхностного плазмонного резонанса (система поверхностного плазмонного резонанса (SPR) BIAcore3000ТМ, BlAcore Inc., Piscataway N.J.). Чипы CM5 можно активировать с помощью гидрохлорида N-этил-N'-(3-диметиламинопропил)карбодиинида (EDC) и N-гидроксисукцинимида (NHS) в соответствии с инструкциями поставщика. Антиген может быть разбавлен в 10 мМ ацетате натрия, pH 4,0, и введен через активированный чип в концентрации 0,005 мг/мл. Путем изменения времени прохождения через отдельные каналы чипа, можно получить два диапазона плотности антигена: 100-200 единиц ответа (RU) для детального изучения кинетических характеристик и 500-600 RU для скрининговых анализов. Серийные разведения (0,1-10-кратное расчетное значение Kd) очищенных образцов Fab вводят в течение 1 мин со скоростью 100 мкл/мин, и дают время для диссоциации до 2 ч. Концентрации Fab-белков определяют с помощью ELISA и/или SDS-PAGE-электрофореза с использованием Fab с известной концентрацией (определенной аминокислотным анализом) в качестве стандарта. Кинетические скорости ассоциации (kon) и диссоциации (koff) получают одновременно путем подгонки данных к модели 1:1 связывания Ленгмюра (см. Karlsson et al. “Kinetic and concentration analysis using BIA technology, Methods: A Companion to Methods Enzymol 6:99-110 (1994), содержание которого включено в настоящее описание во всей своей полноте) с помощью программного обеспечения для оценки BIA. Значения константы равновесной диссоциации (Kd) рассчитывают, как koff/kon. Этот протокол подходит для использования при определении сродства связывания антитела или фрагмента с любым антигеном. Также можно использовать другие протоколы, известные в данной области, например ELISA.
[0088] Эпитоп, который «специфически связывается» с антителом или полипептидом, является термином, хорошо известным в данной области, и способы определения такого специфического или предпочтительного связывания также хорошо известны в данной области. Говорят, что молекулярный объект проявляет «специфическое связывание» или «предпочтительное связывание», если он реагирует или связывается чаще, быстрее, с большей продолжительностью и/или с большим сродством с конкретной клеткой или веществом по сравнению с альтернативными клетками или веществами. Антитело «специфически связывается» или «предпочтительно связывается» с мишенью, если оно связывается с ней более высоким сродством, авидностью, легче и/или с большей продолжительностью, чем оно связывается с другими веществами. Например, антитело, которое специфически или предпочтительно связывается с доменом IgV TIM-3 человека, представляет собой антитело, которое связывается с этим эпитопом с большим сродством, авидностью, более легко и/или с большей продолжительностью, чем оно связывается с другими эпитопами Tim-3 или эпитопами, не относящимися к Tim-3. Из этого определения также понятно, что, например, антитело (или фрагмент, или эпитоп), которое специфически или предпочтительно связывается с первой мишенью, может или не может специфически или предпочтительно связываться со второй мишенью. По существу, «специфическое связывание» или «предпочтительное связывание» не обязательно требует (хотя может включать) исключительного связывания.
[0089] Термин «конкурировать», используемый в настоящем описании в отношении антитела, означает, что первое антитело или его антигенсвязывающая часть связывается с эпитопом способом, в достаточной степени аналогичным способу связывания второго антитела или его антигенсвязывающей части, таким образом, что результат связывания первого антитела с родственным ему эпитопом детектируемо снижается в присутствии второго антитела по сравнению со связыванием первого антитела в отсутствие второго антитела. Альтернативно, связывание второго антитела со своим эпитопом также может, но необязательно, заметно снижаться в присутствии первого антитела. Иными словами, первое антитело может ингибировать связывание второго антитела с его эпитопом, при этом второе антитело не ингибирует связывание первого антитела с соответствующим ему эпитопом. Однако, если каждое антитело детектируемо ингибирует связывание другого антитела с родственным ему эпитопом или лигандом, в той же, большей или меньшей степени, то говорят, что антитела «перекрестно конкурируют» друг с другом за связывание соответствующего им эпитопа(ов). Настоящее изобретение охватывает как конкурирующие, так и перекрестно-конкурирующие антитела. Независимо от механизма такой конкуренции или перекрестной конкуренции (например, наличия стерических препятствий, конформационные изменения или связывание с общим эпитопом или его частью), специалист в данной области поймет, основываясь на положениях, изложенных в настоящем описании, что такие конкурирующие и/или перекрестно конкурирующие антитела охвачены и могут быть полезны для способов, раскрытых в настоящем описании.
[0090] В зависимости от аминокислотных последовательностей константных доменов тяжелых цепей антитела (иммуноглобулины) могут быть отнесены к разным классам. Антитело или фрагмент может быть, например, любым из антител IgG, IgD, IgE, IgA или IgM или их фрагментов, соответственно. В одном из вариантов осуществления антитело представляет собой иммуноглобулин G. В одном из вариантов осуществления фрагмент антитела представляет собой фрагмент иммуноглобулина G. В одном из вариантов осуществления антитело представляет собой IgG1, IgG2, IgG2a, IgG2b, IgG3 или IgG4. В одном из вариантов осуществления антитело содержит последовательности из человеческого IgG1, человеческого IgG2, человеческого IgG2a, человеческого IgG2b, человеческого IgG3 или человеческого IgG4. Также можно использовать комбинацию любого из этих подтипов антител. Одним из соображений при выборе типа используемого антитела является требуемый период полувыведения антитела из сыворотки. Например, период полувыведения из сыворотки обычно составляет 23 дня для IgG, 6 дней для IgA, 5 дней для IgM, 3 дня для IgD и 2 дня для IgE. (Abbas AK, Lichtman AH, Pober JS. Cellular and Molecular Immunology, 4th edition, W.B. Saunders Co., Philadelphia, 2000, включенное в настоящее описание во всей своей полноте посредством ссылки).
[0091] «Вариабельная область» или «вариабельный домен» антитела относится к аминоконцевым доменам тяжелой или легкой цепи антитела. Вариабельный домен тяжелой цепи может быть обозначен как «VH». Вариабельный домен легкой цепи может быть обозначен как «VL». Эти домены обычно являются наиболее вариабельными частями антитела и содержат антигенсвязывающие участки. Термин «вариабельный» относится к тому факту, что последовательности определенных частей вариабельных доменов антител сильно различаются и используются при связывании и определяют специфичность каждого конкретного антитела к своему конкретному антигену. Однако вариабельность неравномерно распределена по вариабельным доменам антител. Она сконцентрирована в трех сегментах, называемых гипервариабельными областями (HVR) (или CDR), как в вариабельных доменах легкой цепи, так и в вариабельных доменах тяжелой цепи. Более высококонсервативные части вариабельных доменов называются каркасными областями (FR). Каждый из вариабельных доменов нативных тяжелых и легких цепей содержит четыре области FR, в большинстве случаев имеющих конфигурацию бета-листов, соединенных тремя CDR, которые образуют петли, соединяющие и, в некоторых случаях, образующие часть структуры бета-листов. CDR в каждой цепи удерживаются вместе в непосредственной близости от областей FR и вместе с CDR другой цепи вносят вклад в образование антигенсвязывающего участка антител (см. Kabat et al., Sequences of Proteins of Immunological Interest, Fifth Edition, National Institute of Health, Bethesda, Md. (1991)). Константные домены не участвуют непосредственно в связывании антитела с антигеном, но проявляют различные эффекторные функции, такие как участие антитела в антителозависимой клеточной токсичности.
[0092] «Легкие цепи» антител (иммуноглобулинов) позвоночных любого вида могут быть отнесены к одному из двух четко различаемых типов, называемых каппа (κ) и лямбда (λ), исходя из аминокислотных последовательностей их константных доменов.
[0093] «Каркасные» или «FR» остатки представляют собой остатки вариабельного домена, которые отличаются от остатков HVR, как определено в настоящем описании.
[0094] Термин «гипервариабельная область» или «HVR», используемый в настоящем описании, относится к областям вариабельного домена антитела, последовательности которых являются гипервариабельными и/или образуют структурно определяемые петли. Обычно антитела содержат шесть HVR: три в VH (H1, H2, H3) и три в VL (L1, L2, L3). В нативных антителах H3 и L3 демонстрируют наибольшее разнообразие из шести HVR, и, как полагают, в частности, H3 играет уникальную роль в придании антителам высокой специфичности. См., например, Xu et al., Immunity 13:37-45 (2000); Johnson and Wu, in Methods in Molecular Biology 248:1-25 (Lo, ed., Human Press, Totowa, N.J., 2003). Действительно, встречающиеся в природе антитела верблюда, состоящие только из тяжелой цепи, являются функциональными и стабильными в отсутствие легкой цепи. (см., например, Hamers-Casterman 1993; Sheriff & Constantine 1996). Имеется несколько описаний HVR, которые включены в настоящую заявку. Области, определяющие комплементарность (CDR) по Кабат, основаны на вариабельности последовательностей и являются наиболее часто используемыми (Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, Md. (1991), полностью включенный в настоящее описание посредством ссылки). Каждая тяжелая и легкая цепь содержит CDR 1, 2 и 3. Чотиа, напротив, ссылается на расположение структурных петель (Chothia & Lesk 1987). HVR по AbM представляют собой компромисс между HVR по Кабату и структурными петлями по Чотиа и используются в программном обеспечении Oxford Molecular's AbM для моделирования антител. «Контактные» HVR основаны на анализе имеющихся сложных кристаллических структур. HVR могут включать «расширенные HVR»: 24-36 или 24-34 (L1), 46-56 или 50-56 (L2) и 89-97 или 89-96 (L3) в VL и 26-35 (H1), 50-65 или 49-65 (H2) и 93-102, 94-102 или 95-102 (H3) в VH. Остатки вариабельных доменов пронумерованы по Кабат и др., см. выше, для каждого из этих определений.
[0095] В контексте настоящего описания термин «Fc-область» используется для определения C-концевой области тяжелой цепи иммуноглобулина, включая Fc-области с нативной последовательностью и вариантные Fc-области. Хотя границы Fc-области тяжелой цепи иммуноглобулина могут меняться, Fc-область тяжелой цепи человеческого IgG обычно определяют как простирающуюся от аминокислотного остатка в положении Cys226 или Pro230 до карбоксильного конца. С-концевой лизин Fc-области может быть удален, например, во время продуцирования или очистки антитела или путем рекомбинантной инженерии нуклеиновой кислоты, кодирующей тяжелую цепь антитела. Соответственно, используемое в настоящей заявке интактное антитело может быть антителом с С-концевым лизином или без него.
[0096] Композиции или фармацевтические композиции, содержащие антитела, scFv или фрагменты антител, раскрытые в настоящем описании, предпочтительно содержат стабилизаторы для предотвращения потери активности или структурной целостности белка в результате денатурации, окисления или агрегации во время хранения и транспортировки до использования. Композиции или фармацевтические композиции могут содержать одну или более комбинаций солей, поверхностно-активных веществ, агентов pH и регулирующих тоничность агентов, таких как сахара, которые позволяют решить проблемы, связанные с агрегацией. Когда композиция или фармацевтическая композиция по настоящему изобретению используется в виде инъекции, предпочтительно, чтобы значение pH находилось в диапазоне приблизительно нейтральных значений pH, также полезно минимизировать уровни поверхностно-активных веществ, чтобы избежать образования пузырьков в составе, оказывающие пагубное воздействие при инъекции индивиду. В одном из вариантов осуществления композиция или фармацевтическая композиция находится в жидкой форме, позволяющей стабильно поддерживать высокие концентрации биоактивного антитела в растворе, и подходит для введения путем ингаляции или парентерального введения. В одном из вариантов осуществления композиция или фармацевтическая композиция подходит для внутривенной, внутримышечной, внутрибрюшинной, внутрикожной и/или подкожной инъекции. В одном из вариантов осуществления композиция или фармацевтическая композиция находится в жидкой форме, позволяющей минимизировать риск образования пузырьков и возникновения анафилактоидных реакций. В одном из вариантов осуществления композиция или фармацевтическая композиция является изотоничной. В одном из вариантов осуществления композиция или фармацевтическая композиция имеет pH в диапазоне от 6,8 до 7,4.
[0097] В одном из вариантов осуществления ScFv или фрагменты антител, раскрытые в настоящем описании, лиофилизируют и/или лиофилизируют и восстанавливают для применения.
[0098] Примеры фармацевтически приемлемых носителей включают, без ограничения, забуференный фосфатом физиологический раствор, стерильную воду (включая воду для инъекций USP), эмульсии, такие как эмульсия масло/вода, и различные типы смачивающих агентов. Предпочтительными разбавителями для аэрозольного или парентерального введения являются забуференный фосфатом физиологический раствор или нормальный (0,9%) физиологический раствор, например 0,9% раствор хлорида натрия, USP. Композиции, содержащие такие носители, готовят хорошо известными традиционными методами (см., например, Remington's Pharmaceutical Sciences, 18-е издание, A. Gennaro, ed., Mack Publishing Co., Easton, PA, 1990; и Remington, The Science and Practice of Pharmacy 20th Ed. Mack Publishing, 2000, содержание каждого из которых включено в настоящее описание во всей своей полноте). В неограничивающих примерах такие композиции могут включать одно или более из: двухосновного фосфата натрия, хлорида калия, одноосновного фосфата калия, полисорбата 80 (например, 2-[2-[3,5-бис(2-гидроксиэтокси)оксолан-2-ил]-2-(2-гидроксиэтокси)этокси]этил (E)-октадек-9-еноат), дигидрат динатрия эдетата, сахарозы, моногидрата одноосновного фосфата натрия и дигидрата двухосновного фосфата натрия.
[0099] Антитела или фрагменты антител или композиции или фармацевтические композиции, раскрытые в настоящем описании, также могут быть лиофилизированы или представлены в любых подходящих формах, включая, без ограничения, растворы для инъекций или растворы для ингаляции, гелевые формы и таблетированные формы.
[0100] В контексте настоящего описания термин «Fc-домен» используется для определения С-концевой области тяжелой цепи иммуноглобулина, включая Fc-области с нативной последовательностью и вариантные Fc-области. Хотя границы Fc-домена тяжелой цепи иммуноглобулина могут меняться, Fc-домен тяжелой цепи человеческого IgG обычно определяют как простирающийся от аминокислотного остатка в положении Cys226 или Pro230 до его карбоксильного конца. С-концевой лизин Fc-домена можно удалить, например, путем рекомбинантной инженерии кодирующей его нуклеиновой кислоты. В некоторых вариантах осуществления изобретения антитело содержит Fc-домен. В одном из вариантов осуществления Fc-домен имеет такую же последовательность, что и Fc-домен человеческого IgG1, или имеет 99% или более сходства с последовательностью с Fc-домена человеческого IgG1. В одном из вариантов осуществления Fc-домен имеет такую же последовательность, что и Fc-домен человеческого IgG2, или имеет 99% или более сходства с последовательностью с Fc-домена человеческого IgG2. В одном из вариантов осуществления Fc-домен имеет такую же последовательность, что и Fc-домен человеческого IgG3, или имеет 99% или более сходства с последовательностью с Fc-домена человеческого IgG3. В одном из вариантов осуществления Fc-домен имеет такую же последовательность, что и Fc-домен человеческого IgG4, или имеет 99% или более сходства с последовательностью с Fc-домена человеческого IgG4. В одном варианте осуществления Fc-домен не мутирован. В одном из вариантов осуществления Fc-домен мутирован на границе доменов CH2-CH3 для увеличения сродства IgG к FcRn при кислом, но не нейтральном pH (Dall'Acqua 2006; Yeung 2009). В одном из вариантов осуществления Fc-домен имеет ту же последовательность, что и Fc-домен человеческого IgG1.
[0101] В некоторых вариантах осуществления изобретение включает модификации вариабельных областей, раскрытых в настоящем описании. Например, изобретение включает антитела, содержащие функционально эквивалентные вариабельные области и CDR, которые существенно не влияют на свойства антител, а также варианты, которые обладают повышенной или пониженной активностью и/или сродством. Например, аминокислотная последовательность может быть мутирована для получения антитела с требуемым сродством связывания с IgV TIM-3 человека. Модификация полипептидов является обычной практикой в данной области, и в настоящем описании нет необходимости в ее подробном описании. Примеры модифицированных полипептидов включают полипептиды с консервативными заменами аминокислотных остатков, одну или более делеций или добавлений аминокислот, которые существенно не изменяют функциональную активность или которые обеспечивают созревание (увеличение) аффинности полипептида к его лиганду, или использование химических аналогов.
[0102] Вставки аминокислотной последовательности включают амино- и/или карбоксиконцевые слияния длиной от одного остатка до полипептидов, содержащих сто или более остатков, а также вставки одного или более аминокислотных остатков внутри последовательности. Примеры концевых вставок включают антитело с N-концевым метионильным остатком или антитело, слитое с эпитопной меткой. Другие инсерционные варианты молекулы антитела включают слияние с N- или С-концом антитела фермента или полипептида, что увеличивает период полувыведения антитела из кровотока.
[0103] Варианты с замещением имеют по меньшей мере один удаленный аминокислотный остаток в молекуле антитела и вставленный на его место другой остаток. Участки, представляющие наибольший интерес с точки зрения замещающего мутагенеза, включают гипервариабельные области, но также рассматриваются изменения в каркасе. Консервативные замены показаны в таблице 1 под заголовком «консервативные замены». Если такие замены приводят к изменению биологической активности, тогда могут быть введены более существенные изменения, приведенные в Таблице 1 в колонке, названной «примерными заменами», или дополнительно описанные ниже со ссылкой на классы аминокислот, и продукты подвергнуты скринингу.
Таблица 1. Аминокислотные замены
[0104] Существенные модификации биологических свойств антитела достигаются путем выбора замен, которые значительно отличаются по своему влиянию на поддержание (а) структуры полипептидного остова в области замены, например, конформации в виде β-слоя или спирали, (b) заряда или гидрофобности молекулы в целевом участке, или (c) величины боковой цепи. Встречающиеся в природе остатки делятся на группы на основе общих свойств боковой цепи:
(1) Неполярные: норлейцин, Met, Ala, Val, Leu, Ile;
(2) Полярные без заряда: Cys, Ser, Thr, Asn, Gln;
(3) Кислые (отрицательно заряженные): Asp, Glu;
(4) Основные (положительно заряженные): Lys, Arg;
(5) Остатки, влияющие на ориентацию цепи: Gly, Pro; и
(6) Ароматические: Trp, Tyr, Phe, His.
[0105] Неконсервативные замены осуществляют путем замены члена одного из этих классов на другой класс.
[0106] Одним из типов возможных замен, например, может быть замена одного или более цистеинов в антителе, которые могут быть химически реактивными, на другой остаток, такой как, без ограничения, аланин или серин. Например, возможна замена на неканонический цистеин. Замена может быть выполнена в CDR или каркасной области вариабельного домена или в константной области антитела. В некоторых вариантах осуществления цистеин является каноническим. Любой остаток цистеина, не участвующий в поддержании правильной конформации антитела, также может быть заменен, обычно серином, для улучшения окислительной стабильности молекулы и предотвращения аберрантной перекрестной сшивки. И наоборот, для улучшения стабильности антитела, особенно если антитело представляет собой фрагмент антитела, такой как Fv-фрагмент, к антителу может быть добавлена цистеиновая связь(и).
[0107] Антитела также могут быть модифицированы, например, в вариабельных доменах тяжелой и/или легкой цепи, например, для изменения связывающей способности антитела. Изменения в вариабельной области могут изменять сродство связывания и/или специфичность. В некоторых вариантах осуществления в домен CDR вводят не более одной-пяти консервативных аминокислотных замен. В других вариантах осуществления в домен CDR вводят не более одной-трех консервативных аминокислотных замен. Например, для увеличения или уменьшения KD антитела к домену IgV TIM-3 человека, для увеличения или уменьшения koff или для изменения специфичности связывания антитела может быть введена мутация в одну или более областей CDR. Методы сайт-направленного мутагенеза хорошо известны в данной области.
[0108] Для увеличения периода полувыведения антитела к домену IgV TIM-3 человека также возможна модификация или мутация в каркасной области или константной области антитела. См., например, публикацию РСТ № WO 00/09560. Мутация в каркасной или константной области также может быть введена для изменения иммуногенности антитела, для получения участка ковалентного или нековалентного связывания с другой молекулой или для изменения таких свойств, как фиксация комплемента, связывание FcR и антитело-зависимая клеточно-опосредованная цитотоксичность. Согласно изобретению, одно антитело может иметь мутации в любой одной или более CDR или каркасных областях вариабельного домена или в константной области.
[0109] В одном из вариантов осуществления антитело по настоящему изобретению получают рекомбинантным способом. В одном из вариантов осуществления слитый белок продуцируется в эукариотической системе экспрессии.
[0110] В одном из вариантов осуществления слитый белок, продуцируемый в эукариотической системе экспрессии, включает гликозилирование по остатку в Fc-участке, соответствующему Asn297.
[0111] В одном из вариантов осуществления композиция или фармацевтическая композиция, содержащая антитело или его антигенсвязывающий фрагмент, раскрытые в настоящем описании, являются по существу чистыми в отношении антитела или его антигенсвязывающего фрагмента. Композиция или фармацевтическая композиция, содержащая антитело или его антигенсвязывающий фрагмент, раскрытые в настоящем описании, являются «по существу чистыми» по отношению к антителу или фрагменту, если по меньшей мере от 60% до 75% образца композиции или фармацевтической композиции содержит один вид антитела или его антигенсвязывающего фрагмента. По существу чистая композиция или фармацевтическая композиция, содержащая антитело или его антигенсвязывающий фрагмент, раскрытые в настоящем описании, может содержать, применительно к антителу или его антигенсвязывающему фрагменту, 60%, 70%, 80% или 90% антитела или антигенсвязывающего фрагмента одного вида, обычно примерно 95% и предпочтительно более 99%. Чистоту или гомогенность можно проверить несколькими способами, хорошо известными в данной области, такими как электрофорез в полиакриламидном геле или ВЭЖХ.
[0112] В одном из вариантов осуществления человеческий Tim3 имеет следующую последовательность:
MFSHLPFDCVLLLLLLLLTRSSEVEYRAEVGQNAYLPCFYTPAAPGNLVPVCWGKGACPVFECGNVVLRTDERDVNYWTSRYWLNGDFRKGDVSLTIENVTLADSGIYCCRIQIPGIMNDEKFNLKLVIKPAKVTPAPTRQRDFTAAFPRMLTTRGHGPAETQTLGSLPDINLTQISTLANELRDSRLANDLRDSGATIRIGIYIGAGICAGLALALIFGALIFKWYSHSKEKIQNLSLISLANLPPSGLANAVAEGIRSEENIYTIEENVYEVEEPNEYYCYVSSRQQPSQPLGCRFAMP (SEQ ID NO:13)
[0113] В одном из вариантов осуществления последовательность домена IgV антитела содержит:
SEVEYRAEVGQNAYLPCFYTPAAPGNLVPVCWGKGACPVFECGNVVLRTDERDVNYWTSRYWLNGDFRKGDVSLTIENVTLADSGIYCCRIQIPGIMNDEKFNLKLVIKPA (SEQ ID NO:22):
[0114] «И/или» в контексте настоящего описания, например, в случае варианта A и/или варианта B, охватывает отдельные варианты осуществления (i) варианта A, (ii) варианта B и (iii) варианта A плюс варианта B.
[0115] Все комбинации различных элементов, раскрытых в настоящем описании, входят в объем изобретения, если не указано иное или если иное в явном виде не противоречит контексту.
[0116] Настоящее изобретение можно лучше понять из подробно описанных экспериментов, которые приведены ниже.
ПРИМЕРЫ
[0117] Получение mAb, специфически связывающихся с доменом IgV TIM-3 человека: слитый белок Tim-3 IgV-Ig получали путем слияния кодирующей области IgV TIM-3 человека (S22-A132) с меткой Fc человеческого IgG1 плазмиды pMT/BiP, как описано ранее (Zhao 2013). Слитый белок экспрессировали в системе S2 и затем очищали. Мышей иммунизировали слитым белком Tim-3 IgV-Ig, и гибридомы получали стандартными методами из спленоцитов, слитых с клетками миеломы NSO.
[0118] Характеристика mAb 50B5: Создавали mAb 50B5, которое представляло собой IgG1 с каппа-цепью. В анализе ELISA оно связывалось как с белком Tim-3-IgV, так и белком всей внеклеточной части Tim-3 (IgV-муцин-стебель), но не с белком IgG человека, (фиг. 1). Подтверждали, что 50B5 связывается с человеческим Tim-3, но не с мышиным Tim-3, экспрессируемыми в линиях клеток в FACS (фиг. 2).
[0119] Методом поверхностного плазменного резонанса определили, что сродство связывания mAb 50B5 с человеческим белком Tim-3 составляет KD=0,13 нМ (фиг. 3). Для определения блокирующей функции mAb 50B5 проводили анализ связывания, и обнаружили, что mAb 50B5 блокирует связывание TIM-3 человека с фосфатидилсерином, экспрессируемым в Т-клетках Jurkat, обработанных дексаметазоном. Поскольку Tim-3 является ингибирующим рецептором, который экспрессируется на Т-клетках, продуцирующих IFN-g, для тестирования антагонистической активности mAb 50B5 использовали реакцию смешанной культуры лимфоцитов. Зрелые дендритные клетки, которые были дифференцированы из моноцитов, выделенных из PBMC одного донора, инкубировали с очищенными Т-клетками из PBMC, полученными от другого донора, в течение четырех дней. Были обнаружены, что на своей поверхности Tim-3 экспрессировали претерпевшие деление CD4 Т-клетки и претерпевшие деление CD8 Т-клетки, но не CD4 и CD8 Т-клетки, которые не претерпели деление (фиг. 4A). В реакции смешанной культуры лимфоцитов добавляли mAb 50B5 или контрольный мышиный IgG1, и было обнаружено, что mAb 50B5 увеличивало продуцирование IFN-g более чем в три раза по сравнению с контрольным IgG1 (фиг. 4B).
[0120] Вместе эти результаты демонстрируют, что mAb 50B5 является антагонистическим антителом к TIM-3 человека. Наконец, секвенировали mAb 50B5 гибридомы, и было обнаружено, что mAb имеет уникальные последовательности VH и VL.
[0121] 50B5 Тяжелая цепь: последовательность ДНК (414 п.н.)
Лидерная последовательность-FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4
(Лидерная последовательность выделена жирным шрифтом, CDR1-3 подчеркнуты):
ATGGGATGGAGCTGGATCTTTCTCTTCCTCCTGTCAGGAACTGCAGGTGTCCACTCTGA
GGTCCAGCTGCAACAGTCTGGACCTGAGCTGGTGAAGCCTGGAGCTTCAATGAAGATAT
CCTGCAAGGCTTCTGGTTACTCATTCACTGGCTACACCATAAACTGGGTGAAGCAGAGC
CATGGAAAGAACCTTGAGTGGATTGGACTTTTTAATCCTTACAATGGTGGTACTACCTT
CAACCAGAAGTTCAAGGGCAAGGCCACATTAACTGTTGACAAGTCATCCAGCACAGCCT
ACATGGAGCTCCTCAGTCTGACATCTGAGGACTCTGCAGTCTATTACTGTGCAAGACGA
TACTACGGCTACGATGCTATGGACTACTGGGGTCAAGGAACCTCAGTCACCGTCTCCTCA
(SEQ ID NO: 14)
[0122] Тяжелая цепь 50B5: аминокислотная последовательность (138 аа)
Лидерная последовательность-FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4
(Лидерная последовательность выделена жирным шрифтом, CDR1-3 подчеркнуты):
MGWSWIFLFLLSGTAGVHSEVQLQQSGPELVKPGASMKISCKASGYSFTGYTINWVKQS
HGKNLEWIGLFNPYNGGTTFNQKFKGKATLTVDKSSSTAYMELLSLTSEDSAVYYCARR
YYGYDAMDYWGQGTSVTVSS
(SEQ ID NO: 15)
[0123] Легкая цепь 50B5: последовательность ДНК (396 п.н.)
Лидерная последовательность-FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4
(Лидерная последовательность выделена жирным шрифтом, CDR1-3 подчеркнуты):
ATGGAATCACAGACTCAGGTCTTCCTCTCCCTGCTGCTCTGGGTATCTGGTACCTGTGG
GAACATTATGATGACACAGTCGCCATCATCTCTGGCTGTGTCTGCAGGAGAAAAGGTCA
CTATGAGCTGTAAGTCCAGTCAAAGTGTTTTATACAGTTCAAATCAGAAGAACCACTTG
GCCTGGTACCAGCAGAAACCAGGGCAGTCTCCTAAACTGCTAATCTACTGGGCATCCAC
TAGGGAATCTGGTGTCCCTGATCGCTTCACAGGCAGTGGATCTGGGACAGATTTTACTC
TTACCATCAGCAGTGTACAAGCTGAAGACCTGGCAGTTTATTACTGTCATCAATACCTC
TCCTCGTACACGTTCGGAGGGGGGACCAAGCTGGAAATTAAG
(SEQ ID NO: 16)
[0124] Легкая цепь 50B5: аминокислотная последовательность (132 аминокислотных остатка)
Лидерная последовательность-FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4
(Лидерная последовательность выделена жирным шрифтом, CDR1-3 подчеркнуты):
MESQTQVFLSLLLWVSGTCGNIMMTQSPSSLAVSAGEKVTMSCKSSQSVLYSSNQKNHL
AWYQQKPGQSPKLLIYWASTRESGVPDRFTGSGSGTDFTLTISSVQAEDLAVYYCHQYL
SSYTFGGGTKLEIK
(SEQ ID NO: 17)
[0125] Характеристика mAb 15B4: Другое mAb, 15B4, получали по той же методике, как описано выше. mAb 15B4 представляет собой IgG1 с каппа-цепью и связывается с человеческим Tim-3, экспрессируемым в линиях клеток в FACS (фиг. 5). Методом поверхностного плазменного резонанса определили, что сродство связывания mAb 15B4 с человеческим белком Tim-3 составляет KD=0,32 нМ (фиг. 6). Для определения блокирующей функции mAb 50B5 проводили анализ связывания, и обнаружили, что 15B4 блокирует связывание TIM-3 человека с фосфатидилсерином, экспрессируемым в Т-клетках Jurkat, обработанных дексаметазоном. Наконец, секвенировали 15B4 гибридомы, и было обнаружено, что mAb имеет уникальные последовательности VH и VL:
[0126] Тяжелая цепь 15B4: последовательность ДНК (414 п.н.):
Лидерная последовательность-FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4
(Лидерная последовательность выделена жирным шрифтом, CDR1-3 подчеркнуты):
ATGGAATGGAGCTGGGTCTTTCTCTTCCTGATGGCAGTGGTTACAGGGGTCAATTCAGA
GGTTCAGCTGCAGCAGTCTGGGGCTGAGGTTGTGAGGCCAGGGGCCTTAGTCAAGTTGT
CCTGCAAAGCTTCTGGCTTCAACATTAAAGACTACTATATGCACTGGGTGAGGCAGAGG
CCTGAACAGGGCCTGGAGTGGATTGGATGGATTGATCCTGAGAATGACAATACTATATA
TGACCCGAAGTTCCAGGACAGGGCCAGTATAACAGCAGACACATCCTCCAACACAGCCT
ACCTGCAGCTCAGCAGCCTGACATCTGAGGACACTGCCGTCTATTACTGTGCTAGGGAC
TTCGGCTACGTAGCCTGGCTTGTTTACTGGGGCCAAGGGACTCTGGTCACTGTCTCTGC
A
(SEQ ID NO: 18).
[0127] Тяжелая цепь 15B4: Аминокислотная последовательность (138 аминокислотных остатков):
Лидерная последовательность-FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4
(Лидерная последовательность выделена жирным шрифтом, CDR1-3 подчеркнуты):
MEWSWVFLFLMAVVTGVNSEVQLQQSGAEVVRPGALVKLSCKASGFNIKDYYMHWVRQR
PEQGLEWIGWIDPENDNTIYDPKFQDRASITADTSSNTAYLQLSSLTSEDTAVYYCARD
FGYVAWLVYWGQGTLVTVSA
(SEQ ID NO: 19).
[0128] Легкая цепь 15B4: последовательность ДНК (378 п.н.):
Лидерная последовательность-FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4
(Лидерная последовательность выделена жирным шрифтом, CDR1-3 подчеркнуты):
ATGTCACAGTCTCAGGTCTTTGTATTCGCGTTTCTCTGGTTGTCTGGTGTTGATGGAGA
CATTGTGATGACCCAGTCTCAAGAATTCATGTCCACATCAGTAGGAGACAGGGTCAGCA
TCACCTGCAAGGCCAGTCAGAATGTGGATACTGCTGTAGCCTGGTATCAACAGAAACCA
GGACAATCTCCTAAACTACTGATTTACTCGGCATCCAATCGGTACACTGGAGTCCCTGA
TCGCTTCACAGGCACTGGATCTGGGACAGATTTCACTCTCACCATCAACAATATGCAGT
CTGAAGACCTGGCAGATTATTTCTGCCAGCAATATAGCAGCTATCCCACGTTCGGAGGG
AGGACCAAGCTGGAAATAAAACGG
(SEQ ID NO: 20).
[0129] Легкая цепь 15B4: Аминокислотная последовательность (126 аминокислотных остатков):
Лидерная последовательность-FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4
(Лидерная последовательность выделена жирным шрифтом, CDR1-3 подчеркнуты):
MSQSQVFVFAFLWLSGVDGDIVMTQSQEFMSTSVGDRVSITCKASQNVDTAVAWYQQKP
GQSPKLLIYSASNRYTGVPDRFTGTGSGTDFTLTINNMQSEDLADYFCQQYSSYPTFGG
RTKLEIKR
(SEQ ID NO: 21).
Источники информации
Bird et al. Science 242(4877):423-426 (1988)
Brown et al. Cancer Res 47(13):3577-3583 (1987)
Cao et al. Immunity 26(3):311-321 (2007)
Chiba et al. Nat Immunol 13(9):832-842 (2012)
Chothia & Lesk J Mol Biol 196(4):901-917 (1987)
Chothia et al. Nature 342(6252):877-883 (1989)
Dall’Acqua et al. J Biol Chem 281(33):23514-23524 (2006)
Daugherty et al. Nucleic Acids Res 19(9):2471-2476 (1991)
DeKruyff et al. J Immunol 184(4):1918-1930 (2010)
Hamers-Casterman et al. Nature 363(6428):446-448 (1993)
Harris Biochem Soc Trans 23(4):1035-1038 (1995)
Hurle & Gross Curr Opin Biotechnol 5(4):428-433 (1994)
Huston et al. Proc Natl Acad Sci USA 85(16):5879-5883 (1988)
Jones et al. Nature 321(6069):522-525 (1986)
Koyama et al. Nat Commun 7:10501 (2016)
Lobuglio et al. Proc Natl Acad Sci USA 86(11):4220-4224 (1989)
Monney et al. Nature 415(6871):536-541 (2002)
Presta Curr Opin Biotechnol 3(4):394-398 (1992)
Riechmann et al. Nature 332(6162):323-327 (1988)
Shaw et al. J Immunol 138(12):4534-4538 (1987)
Sheriff & Constantine Nat Struct Biol 3(9):733-736 (1996)
Vaswani & Hamilton Ann Allergy Asthma Immunol 81(2):105-115 (1998)
Verhoeyen et al. Science 239(4847):1534-1536 (1988)
Ward et al. Nature 341(6242):544-546 (1989)
Wilker et al. Int Immunol 19(6):763-773 (2007)
Winter et al. Nature 349(6307):293-299 (1991)
Yeung et al. J Immunol 182:7663-7671 (2009)
Zhao et al. Proc Natl Acad Sci USA 110(24):9879-9884 (2013)
Zhu et al. Nat Immunol 6(12):1245-1252 (2005).
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> АЛЬБЕРТ ЭЙНШТЕЙН КОЛЛЕДЖ ОФ МЕДСИН
ЦЗАН, Синсин
<120> МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА К TIM-3 ЧЕЛОВЕКА
<130> 129807-8004.WO00
<150> US 62/720,234
<151> 2018-08-21
<160> 22
<170> PatentIn version 3.5
<210> 1
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> 50B5 HCDR1
<400> 1
Gly Tyr Ser Phe Thr Gly Tyr Thr Ile Asn
1 5 10
<210> 2
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> 50B5 HCDR2
<400> 2
Leu Phe Asn Pro Tyr Asn Gly Gly Thr Thr
1 5 10
<210> 3
<211> 12
<212> Белок
<213> Искусственная последовательность
<220>
<223> 50B5 HCDR3
<400> 3
Ala Arg Arg Tyr Tyr Gly Tyr Asp Ala Met Asp Tyr
1 5 10
<210> 4
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> 50B5 LCDR1
<400> 4
Lys Ser Ser Gln Ser Val Leu Tyr Ser Ser Asn Gln Lys Asn His Leu
1 5 10 15
Ala
<210> 5
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> 50B5 LCDR2
<400> 5
Trp Ala Ser Thr Arg Glu Ser
1 5
<210> 6
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> 50B5 LCDR3
<400> 6
His Gln Tyr Leu Ser Ser Tyr Thr
1 5
<210> 7
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> 15B4 HCDR1
<400> 7
Gly Phe Asn Ile Lys Asp Tyr Tyr Met His
1 5 10
<210> 8
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> 15B4 HCDR2
<400> 8
Trp Ile Asp Pro Glu Asn Asp Asn Thr Ile Tyr
1 5 10
<210> 9
<211> 12
<212> Белок
<213> Искусственная последовательность
<220>
<223> 15B4 HCDR3
<400> 9
Ala Arg Asp Phe Gly Tyr Val Ala Trp Leu Val Tyr
1 5 10
<210> 10
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> 15B4 LCDR1
<400> 10
Lys Ala Ser Gln Asn Val Asp Thr Ala Val Ala
1 5 10
<210> 11
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> 15B4 LCDR2
<400> 11
Ser Ala Ser Asn Arg Tyr Thr
1 5
<210> 12
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> LCDR3
<400> 12
Gln Gln Tyr Ser Ser Tyr Pro Thr
1 5
<210> 13
<211> 301
<212> Белок
<213> Homo sapiens
<400> 13
Met Phe Ser His Leu Pro Phe Asp Cys Val Leu Leu Leu Leu Leu Leu
1 5 10 15
Leu Leu Thr Arg Ser Ser Glu Val Glu Tyr Arg Ala Glu Val Gly Gln
20 25 30
Asn Ala Tyr Leu Pro Cys Phe Tyr Thr Pro Ala Ala Pro Gly Asn Leu
35 40 45
Val Pro Val Cys Trp Gly Lys Gly Ala Cys Pro Val Phe Glu Cys Gly
50 55 60
Asn Val Val Leu Arg Thr Asp Glu Arg Asp Val Asn Tyr Trp Thr Ser
65 70 75 80
Arg Tyr Trp Leu Asn Gly Asp Phe Arg Lys Gly Asp Val Ser Leu Thr
85 90 95
Ile Glu Asn Val Thr Leu Ala Asp Ser Gly Ile Tyr Cys Cys Arg Ile
100 105 110
Gln Ile Pro Gly Ile Met Asn Asp Glu Lys Phe Asn Leu Lys Leu Val
115 120 125
Ile Lys Pro Ala Lys Val Thr Pro Ala Pro Thr Arg Gln Arg Asp Phe
130 135 140
Thr Ala Ala Phe Pro Arg Met Leu Thr Thr Arg Gly His Gly Pro Ala
145 150 155 160
Glu Thr Gln Thr Leu Gly Ser Leu Pro Asp Ile Asn Leu Thr Gln Ile
165 170 175
Ser Thr Leu Ala Asn Glu Leu Arg Asp Ser Arg Leu Ala Asn Asp Leu
180 185 190
Arg Asp Ser Gly Ala Thr Ile Arg Ile Gly Ile Tyr Ile Gly Ala Gly
195 200 205
Ile Cys Ala Gly Leu Ala Leu Ala Leu Ile Phe Gly Ala Leu Ile Phe
210 215 220
Lys Trp Tyr Ser His Ser Lys Glu Lys Ile Gln Asn Leu Ser Leu Ile
225 230 235 240
Ser Leu Ala Asn Leu Pro Pro Ser Gly Leu Ala Asn Ala Val Ala Glu
245 250 255
Gly Ile Arg Ser Glu Glu Asn Ile Tyr Thr Ile Glu Glu Asn Val Tyr
260 265 270
Glu Val Glu Glu Pro Asn Glu Tyr Tyr Cys Tyr Val Ser Ser Arg Gln
275 280 285
Gln Pro Ser Gln Pro Leu Gly Cys Arg Phe Ala Met Pro
290 295 300
<210> 14
<211> 414
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 50B5 тяжелая цепь
<220>
<221> прочий_признак
<222> (1)..(57)
<223> Лидерная последовательность
<220>
<221> прочий_признак
<222> (133)..(162)
<223> HCDR1
<220>
<221> прочий_признак
<222> (205)..(234)
<223> HCDR2
<220>
<221> прочий_признак
<222> (346)..(381)
<223> HCDR3
<400> 14
atgggatgga gctggatctt tctcttcctc ctgtcaggaa ctgcaggtgt ccactctgag 60
gtccagctgc aacagtctgg acctgagctg gtgaagcctg gagcttcaat gaagatatcc 120
tgcaaggctt ctggttactc attcactggc tacaccataa actgggtgaa gcagagccat 180
ggaaagaacc ttgagtggat tggacttttt aatccttaca atggtggtac taccttcaac 240
cagaagttca agggcaaggc cacattaact gttgacaagt catccagcac agcctacatg 300
gagctcctca gtctgacatc tgaggactct gcagtctatt actgtgcaag acgatactac 360
ggctacgatg ctatggacta ctggggtcaa ggaacctcag tcaccgtctc ctca 414
<210> 15
<211> 138
<212> Белок
<213> Искусственная последовательность
<220>
<223> 50B5 тяжелая цепь
<220>
<221> ПРОЧИЙ_ПРИЗНАК
<222> (1)..(19)
<223> Лидерная последовательность
<220>
<221> ПРОЧИЙ_ПРИЗНАК
<222> (45)..(54)
<223> HCDR1
<220>
<221> ПРОЧИЙ_ПРИЗНАК
<222> (69)..(78)
<223> HCDR2
<220>
<221> ПРОЧИЙ_ПРИЗНАК
<222> (116)..(127)
<223> HCDR3
<400> 15
Met Gly Trp Ser Trp Ile Phe Leu Phe Leu Leu Ser Gly Thr Ala Gly
1 5 10 15
Val His Ser Glu Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Val Lys
20 25 30
Pro Gly Ala Ser Met Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ser Phe
35 40 45
Thr Gly Tyr Thr Ile Asn Trp Val Lys Gln Ser His Gly Lys Asn Leu
50 55 60
Glu Trp Ile Gly Leu Phe Asn Pro Tyr Asn Gly Gly Thr Thr Phe Asn
65 70 75 80
Gln Lys Phe Lys Gly Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser
85 90 95
Thr Ala Tyr Met Glu Leu Leu Ser Leu Thr Ser Glu Asp Ser Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Arg Tyr Tyr Gly Tyr Asp Ala Met Asp Tyr Trp
115 120 125
Gly Gln Gly Thr Ser Val Thr Val Ser Ser
130 135
<210> 16
<211> 396
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 50B5 легкая цепь
<220>
<221> прочий_признак
<222> (1)..(60)
<223> Лидерная последовательность
<220>
<221> прочий_признак
<222> (130)..(180)
<223> LCDR1
<220>
<221> прочий_признак
<222> (226)..(246)
<223> LCDR2
<220>
<221> прочий_признак
<222> (343)..(366)
<223> LCDR3
<400> 16
atggaatcac agactcaggt cttcctctcc ctgctgctct gggtatctgg tacctgtggg 60
aacattatga tgacacagtc gccatcatct ctggctgtgt ctgcaggaga aaaggtcact 120
atgagctgta agtccagtca aagtgtttta tacagttcaa atcagaagaa ccacttggcc 180
tggtaccagc agaaaccagg gcagtctcct aaactgctaa tctactgggc atccactagg 240
gaatctggtg tccctgatcg cttcacaggc agtggatctg ggacagattt tactcttacc 300
atcagcagtg tacaagctga agacctggca gtttattact gtcatcaata cctctcctcg 360
tacacgttcg gaggggggac caagctggaa attaag 396
<210> 17
<211> 132
<212> Белок
<213> Искусственная последовательность
<220>
<223> 50B5 легкая цепь
<220>
<221> ПРОЧИЙ_ПРИЗНАК
<222> (1)..(20)
<223> Лидерная последовательность
<220>
<221> ПРОЧИЙ_ПРИЗНАК
<222> (44)..(60)
<223> LCDR1
<220>
<221> ПРОЧИЙ_ПРИЗНАК
<222> (76)..(82)
<223> LCDR2
<220>
<221> ПРОЧИЙ_ПРИЗНАК
<222> (115)..(122)
<223> LCDR3
<400> 17
Met Glu Ser Gln Thr Gln Val Phe Leu Ser Leu Leu Leu Trp Val Ser
1 5 10 15
Gly Thr Cys Gly Asn Ile Met Met Thr Gln Ser Pro Ser Ser Leu Ala
20 25 30
Val Ser Ala Gly Glu Lys Val Thr Met Ser Cys Lys Ser Ser Gln Ser
35 40 45
Val Leu Tyr Ser Ser Asn Gln Lys Asn His Leu Ala Trp Tyr Gln Gln
50 55 60
Lys Pro Gly Gln Ser Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg
65 70 75 80
Glu Ser Gly Val Pro Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp
85 90 95
Phe Thr Leu Thr Ile Ser Ser Val Gln Ala Glu Asp Leu Ala Val Tyr
100 105 110
Tyr Cys His Gln Tyr Leu Ser Ser Tyr Thr Phe Gly Gly Gly Thr Lys
115 120 125
Leu Glu Ile Lys
130
<210> 18
<211> 414
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 15B4 тяжелая цепь
<220>
<221> прочий_признак
<222> (1)..(57)
<223> Лидерная последовательность
<220>
<221> прочий_признак
<222> (133)..(162)
<223> HCDR1
<220>
<221> прочий_признак
<222> (208)..(237)
<223> HCDR2
<220>
<221> прочий_признак
<222> (346)..(381)
<223> HCDR3
<400> 18
atggaatgga gctgggtctt tctcttcctg atggcagtgg ttacaggggt caattcagag 60
gttcagctgc agcagtctgg ggctgaggtt gtgaggccag gggccttagt caagttgtcc 120
tgcaaagctt ctggcttcaa cattaaagac tactatatgc actgggtgag gcagaggcct 180
gaacagggcc tggagtggat tggatggatt gatcctgaga atgacaatac tatatatgac 240
ccgaagttcc aggacagggc cagtataaca gcagacacat cctccaacac agcctacctg 300
cagctcagca gcctgacatc tgaggacact gccgtctatt actgtgctag ggacttcggc 360
tacgtagcct ggcttgttta ctggggccaa gggactctgg tcactgtctc tgca 414
<210> 19
<211> 138
<212> Белок
<213> Искусственная последовательность
<220>
<223> 15B4 тяжелая цепь
<220>
<221> ПРОЧИЙ_ПРИЗНАК
<222> (1)..(19)
<223> Лидерная последовательность
<220>
<221> ПРОЧИЙ_ПРИЗНАК
<222> (45)..(54)
<223> HCDR1
<220>
<221> ПРОЧИЙ_ПРИЗНАК
<222> (69)..(79)
<223> HCDR2
<220>
<221> ПРОЧИЙ_ПРИЗНАК
<222> (116)..(127)
<223> HCDR3
<400> 19
Met Glu Trp Ser Trp Val Phe Leu Phe Leu Met Ala Val Val Thr Gly
1 5 10 15
Val Asn Ser Glu Val Gln Leu Gln Gln Ser Gly Ala Glu Val Val Arg
20 25 30
Pro Gly Ala Leu Val Lys Leu Ser Cys Lys Ala Ser Gly Phe Asn Ile
35 40 45
Lys Asp Tyr Tyr Met His Trp Val Arg Gln Arg Pro Glu Gln Gly Leu
50 55 60
Glu Trp Ile Gly Trp Ile Asp Pro Glu Asn Asp Asn Thr Ile Tyr Asp
65 70 75 80
Pro Lys Phe Gln Asp Arg Ala Ser Ile Thr Ala Asp Thr Ser Ser Asn
85 90 95
Thr Ala Tyr Leu Gln Leu Ser Ser Leu Thr Ser Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Asp Phe Gly Tyr Val Ala Trp Leu Val Tyr Trp
115 120 125
Gly Gln Gly Thr Leu Val Thr Val Ser Ala
130 135
<210> 20
<211> 378
<212> ДНК
<213> Искусственная последовательность
<220>
<223> 15B4 легкая цепь
<220>
<221> прочий_признак
<222> (1)..(57)
<223> Лидерная последовательность
<220>
<221> прочий_признак
<222> (127)..(159)
<223> LCDR1
<220>
<221> прочий_признак
<222> (205)..(225)
<223> LCDR2
<220>
<221> прочий_признак
<222> (322)..(345)
<223> LCDR3
<400> 20
atgtcacagt ctcaggtctt tgtattcgcg tttctctggt tgtctggtgt tgatggagac 60
attgtgatga cccagtctca agaattcatg tccacatcag taggagacag ggtcagcatc 120
acctgcaagg ccagtcagaa tgtggatact gctgtagcct ggtatcaaca gaaaccagga 180
caatctccta aactactgat ttactcggca tccaatcggt acactggagt ccctgatcgc 240
ttcacaggca ctggatctgg gacagatttc actctcacca tcaacaatat gcagtctgaa 300
gacctggcag attatttctg ccagcaatat agcagctatc ccacgttcgg agggaggacc 360
aagctggaaa taaaacgg 378
<210> 21
<211> 126
<212> Белок
<213> Искусственная последовательность
<220>
<223> 15B4 легкая цепь
<220>
<221> ПРОЧИЙ_ПРИЗНАК
<222> (1)..(19)
<223> Лидерная последовательность
<220>
<221> ПРОЧИЙ_ПРИЗНАК
<222> (43)..(53)
<223> LCDR1
<220>
<221> ПРОЧИЙ_ПРИЗНАК
<222> (69)..(75)
<223> LCDR2
<220>
<221> ПРОЧИЙ_ПРИЗНАК
<222> (107)..(115)
<223> LCDR3
<400> 21
Met Ser Gln Ser Gln Val Phe Val Phe Ala Phe Leu Trp Leu Ser Gly
1 5 10 15
Val Asp Gly Asp Ile Val Met Thr Gln Ser Gln Glu Phe Met Ser Thr
20 25 30
Ser Val Gly Asp Arg Val Ser Ile Thr Cys Lys Ala Ser Gln Asn Val
35 40 45
Asp Thr Ala Val Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Lys
50 55 60
Leu Leu Ile Tyr Ser Ala Ser Asn Arg Tyr Thr Gly Val Pro Asp Arg
65 70 75 80
Phe Thr Gly Thr Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Asn
85 90 95
Met Gln Ser Glu Asp Leu Ala Asp Tyr Phe Cys Gln Gln Tyr Ser Ser
100 105 110
Tyr Pro Thr Phe Gly Gly Arg Thr Lys Leu Glu Ile Lys Arg
115 120 125
<210> 22
<211> 111
<212> Белок
<213> Искусственная последовательность
<220>
<223> IgV домен
<400> 22
Ser Glu Val Glu Tyr Arg Ala Glu Val Gly Gln Asn Ala Tyr Leu Pro
1 5 10 15
Cys Phe Tyr Thr Pro Ala Ala Pro Gly Asn Leu Val Pro Val Cys Trp
20 25 30
Gly Lys Gly Ala Cys Pro Val Phe Glu Cys Gly Asn Val Val Leu Arg
35 40 45
Thr Asp Glu Arg Asp Val Asn Tyr Trp Thr Ser Arg Tyr Trp Leu Asn
50 55 60
Gly Asp Phe Arg Lys Gly Asp Val Ser Leu Thr Ile Glu Asn Val Thr
65 70 75 80
Leu Ala Asp Ser Gly Ile Tyr Cys Cys Arg Ile Gln Ile Pro Gly Ile
85 90 95
Met Asn Asp Glu Lys Phe Asn Leu Lys Leu Val Ile Lys Pro Ala
100 105 110
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| КОМБИНИРОВАННЫЕ ЛЕКАРСТВЕННЫЕ СРЕДСТВА, НАЦЕЛЕННЫЕ НА PD-1, TIM-3 И LAG-3 | 2018 |
|
RU2795232C2 |
| АНТИТЕЛА, КОТОРЫЕ СВЯЗЫВАЮТСЯ С СОРТИЛИНОМ И ПОДАВЛЯЮТ СВЯЗЫВАНИЕ ПРОГРАНУЛИНА | 2016 |
|
RU2735639C2 |
| УСОВЕРШЕНСТВОВАННЫЕ ПРОКОАГУЛЯНТНЫЕ АНТИТЕЛА | 2019 |
|
RU2810748C2 |
| АНТИТЕЛА К PD-1 СОБАК | 2014 |
|
RU2732604C2 |
| АНТИТЕЛА ПРОТИВ VISTA И ИХ ПРИМЕНЕНИЕ | 2018 |
|
RU2750723C1 |
| АНТИТЕЛА, НАПРАВЛЕННЫЕ ПРОТИВ ЛЕЙКОТОКСИНОВ STAPHYLOCOCCUS AUREUS | 2019 |
|
RU2805969C2 |
| КОМБИНАЦИИ АНТИТЕЛ К STAPHYLOCOCCUS AUREUS | 2019 |
|
RU2804815C2 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕ SIGLEC15, И ВАРИАНТЫ ИХ ПРИМЕНЕНИЯ | 2021 |
|
RU2839071C1 |
| АНТИТЕЛА К PD-1 | 2015 |
|
RU2715628C2 |
| Белки, связывающие NKG2D, CD16 и опухолеассоциированный антиген | 2018 |
|
RU2816716C2 |
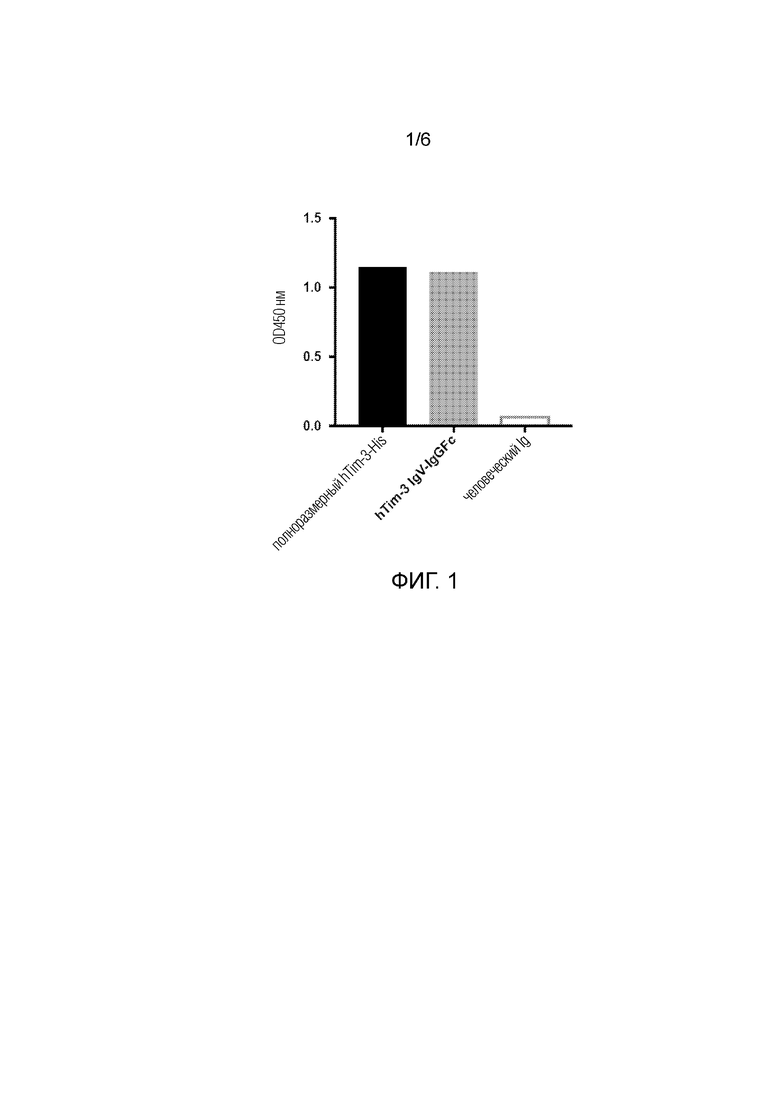
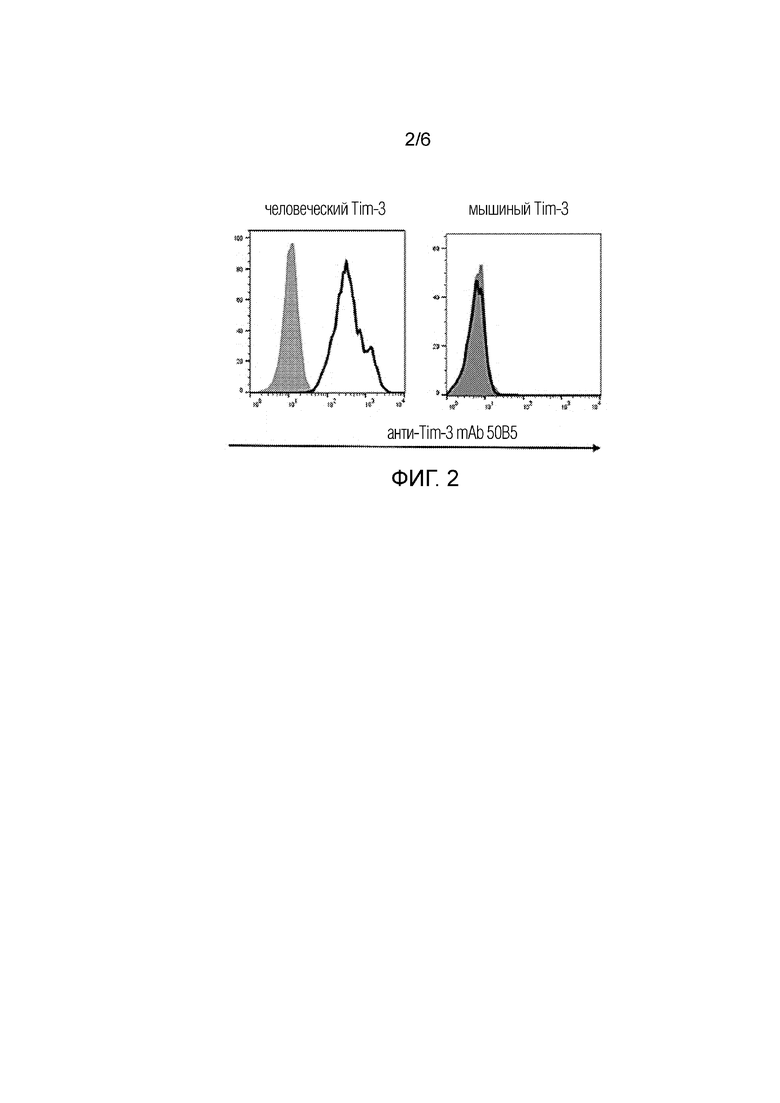
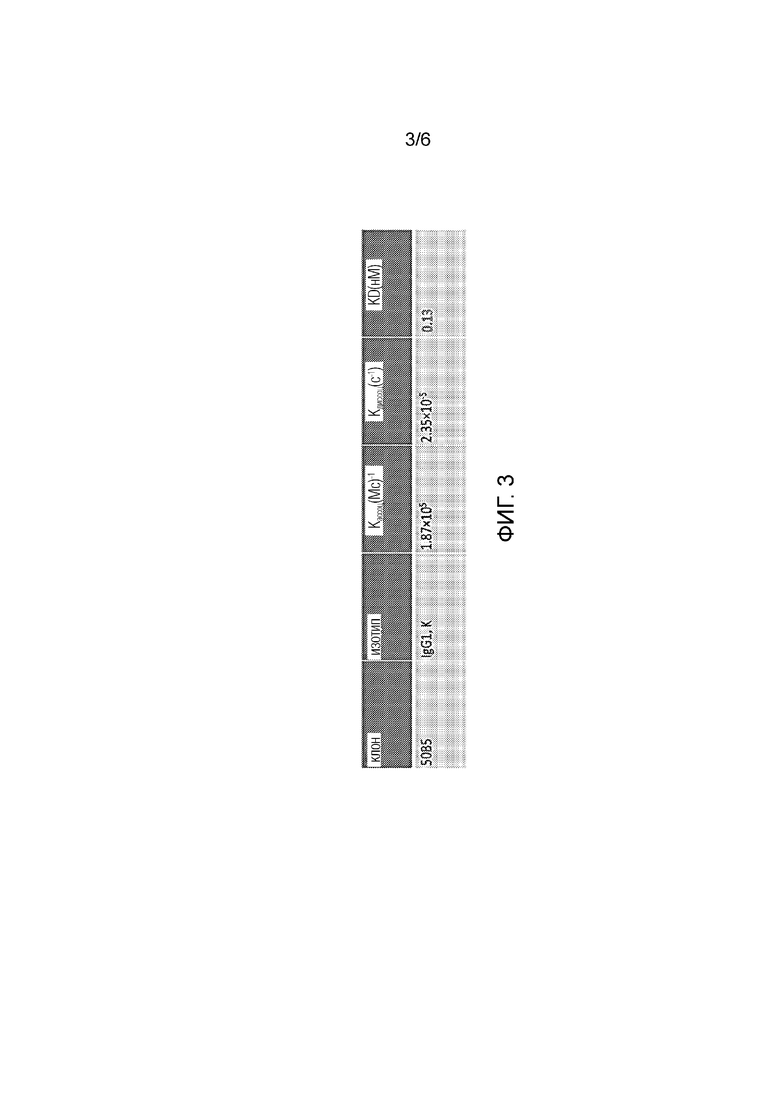
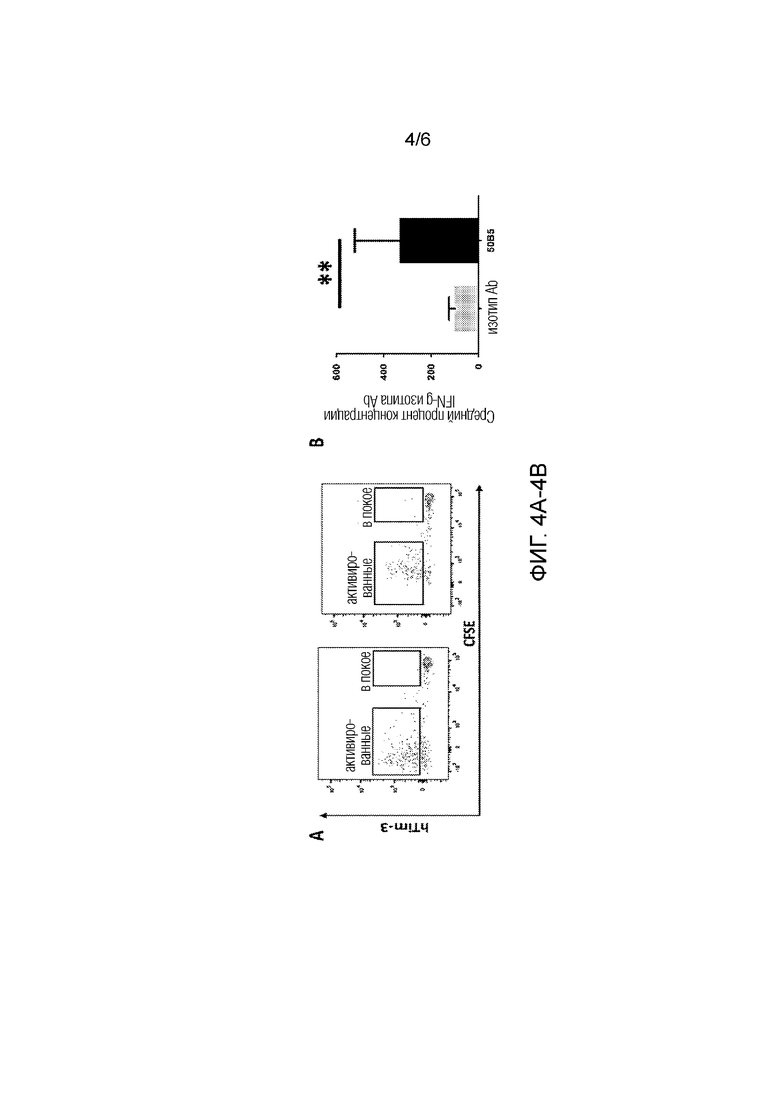

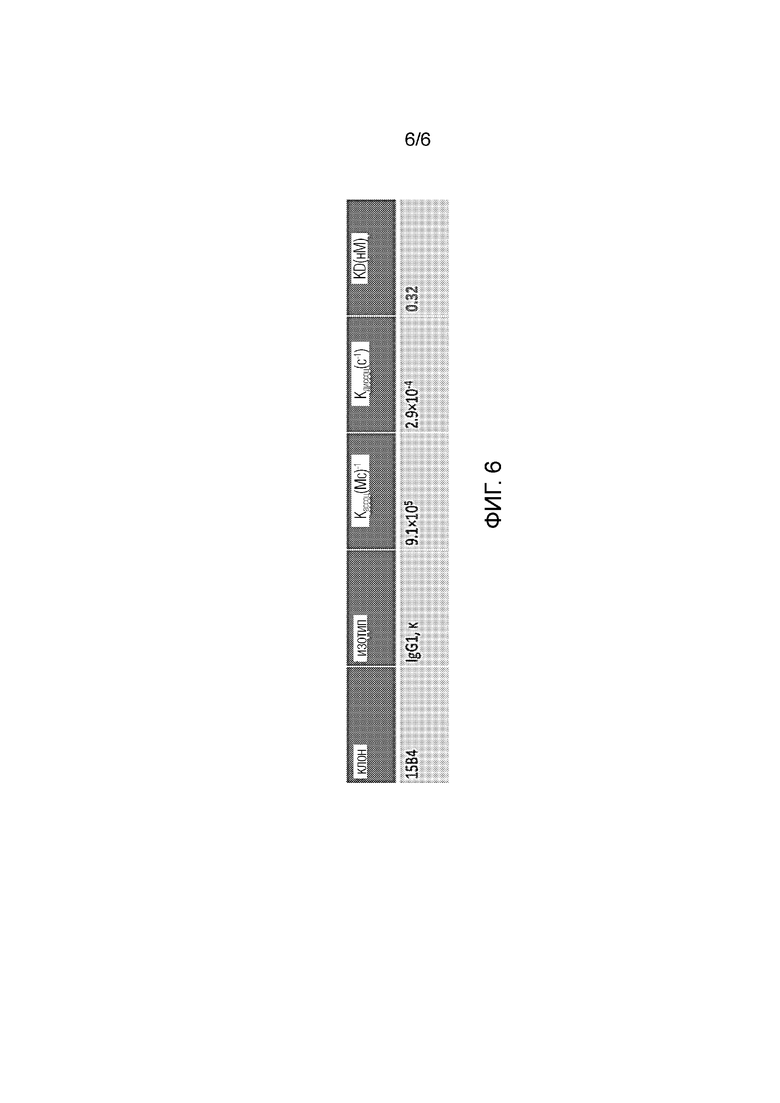
Изобретение относится к области биотехнологии, конкретно к получению антител к домену IgV TIM-3 человека, и может быть использовано в медицине для ингибирования TIM-3 человека, ингибирования Tim-3-опосредованной супрессии Т-клеток у индивида и лечения Tim-3-положительного злокачественного заболевания, а также для детектирования клетки, положительной по TIM-3 человека. Изобретение обеспечивает получение антител, которые специфически связываются с доменом IgV TIM-3 человека или IgV муцин-стебель TIM-3 человека и содержат тяжелую цепь с GYSFTGYTIN(CDR1), LFNPYNGGTT (CDR2) и ARRYYGYDAMDY (CDR3); а также легкую цепь с KSSQSVLYSSNQKNHLA (CDR1); WASTRES (CDR2) и HQYLSSYT (CDR3). 7 н. и 15 з.п. ф-лы, 6 ил., 1 табл., 1 пр.
1. Антитело или его антигенсвязывающий фрагмент, которые специфически связываются с доменом IgV TIM-3 человека или IgV муцин-стебель TIM-3 человека, содержащие:
тяжелую цепь, содержащую
GYSFTGYTIN (SEQ ID NO:1) (CDR1);
LFNPYNGGTT (SEQ ID NO:2) (CDR2);
ARRYYGYDAMDY (SEQ ID NO:3) (CDR3); и
легкую цепь, содержащую
KSSQSVLYSSNQKNHLA (SEQ ID NO:4) (CDR1);
WASTRES (SEQ ID NO:5) (CDR2);
HQYLSSYT (SEQ ID NO:6) (CDR3).
2. Антитело или его антигенсвязывающий фрагмент по п.1, где каркасные области легкой цепи и тяжелой цепи являются каркасными областями человека или на 85% идентичны им.
3. Антитело или его антигенсвязывающий фрагмент по п.1 или 2, где каркасные области легкой цепи и тяжелой цепи являются каркасными областями человека.
4. Выделенное антитело или его антигенсвязывающий фрагмент по любому из пп.1-3, которые специфически связываются с доменом IgV TIM-3 человека или IgV муцин-стебель TIM-3 человека со сродством KD=10,0 нМ или сильнее.
5. Выделенное антитело или его антигенсвязывающий фрагмент по п.4, которые специфически связываются с доменом IgV TIM-3 человека или IgV муцин-стебель TIM-3 человека со сродством KD=0,5 нМ или сильнее.
6. Антитело или его антигенсвязывающий фрагмент по любому из пп.1-5, которые имеют Fc-область с человеческой последовательностью.
7. Антитело или его антигенсвязывающий фрагмент по любому из пп.1-6, которые ингибируют связывание TIM-3 человека с фосфатидилсерином, экспрессируемым на обработанных дексаметазоном Т-клетках Jurkat.
8. Антитело или его антигенсвязывающий фрагмент по любому из пп.1-7, где антитело или его антигенсвязывающий фрагмент являются химерными или гуманизированными.
9. Антитело или его антигенсвязывающий фрагмент по любому из пп.1-8, где антитело или его антигенсвязывающий фрагмент выбраны из группы, состоящей из моноклонального антитела, scFv, Fab-фрагмента, Fab' фрагмента и F(ab)' фрагмента.
10. Нуклеиновая кислота, кодирующая антитело или его антигенсвязывающий фрагмент по любому из пп.1-9.
11. Клетка-хозяин, продуцирующая антитело или его антигенсвязывающий фрагмент по любому из пп.1-9, где клетка содержит нуклеиновую кислоту по п.10.
12. Конъюгат антитела для лечения Tim-3-положительного злокачественного заболевания, где конъюгат содержит антитело или его антигенсвязывающий фрагмент по любому из пп.1-9, связанное или конъюгированное с терапевтическим агентом.
13. Конъюгат по п.12, в котором терапевтический агент представляет собой цитотоксическое лекарственное вещество, радиоактивный изотоп, иммуномодулятор или второе антитело.
14. Способ ингибирования TIM-3 человека или ингибирования Tim-3-опосредованной супрессии Т-клеток у индивида, включающий введение эффективного количества антитела или его антигенсвязывающего фрагмента по любому из пп.1-9.
15. Способ по п.14, где индивид болен раком.
16. Способ лечения Tim-3-положительного злокачественного заболевания у человека, включающий введение антитела или его антигенсвязывающего фрагмента по любому из пп.1-9 в эффективном количестве.
17. Способ детектирования клетки, положительной по TIM-3 человека, у индивида, включающий введение антитела или его антигенсвязывающего фрагмента по любому из пп.1-9, содержащего детектируемый конъюгированный с ним маркер, в количестве, эффективном для мечения клетки, положительной по TIM-3 человека, и затем обнаружение метки у индивида, таким образом детектируя у индивида клетку, положительную по TIM-3 человека.
18. Способ по п.17, в котором метку детектируют путем визуализации.
19. Способ по п.17, в котором клетка представляет собой раковую клетку.
20. Способ по пп.15, 16 или 19, в котором злокачественное заболевание является гематологической злокачественной опухолью.
21. Способ по пп.15, 16 или 19, в котором злокачественное заболевание включает солидную опухоль.
22. Способ по пп.15, 16, 17 или 19-21, в котором индивид получает анти-PD-1 или анти-PD-L1, или анти-CTLA4 терапию.
| US 2017088616 А1, 30.03.2017, EP 2534178 A1, 19.12.2012, WO 2008148884 A1, 11.12.2008, WO 2017153955 A1, 14.09.2017, US 9321833 B2, 26/04/2016, WO 2008061013 A2, 22.05.2008, RU 2006114694 A, 10.11.2007, "СЛОВАРЬ БИОТЕХНОЛОГИЧЕСКИХ ТЕРМИНОВ" под ред | |||
| д.б.н | |||
| В.З | |||
| Тарантула, Москва, 2005, стр | |||
| Прибор для нагревания перетягиваемых бандажей подвижного состава | 1917 |
|
SU15A1 |
| et al., Affinity improvement of single | |||

