В основном изобретение касается альфа (α)- интерфероновых композиций. В большей степени изобретение относится к улучшенной альфа-интерфероновой композиции, характеризующейся высокой специфической активностью и чистотой, превышающей 95%, и которая, будучи применена на людях, отличается соответствующей редукцией побочных эффектов, обычно ассоциированных с такого рода композициями. Кроме того, изобретение относится к крупномасштабному получению данной композиции с помощью лейкоцитов периферической крови человека
Альфа-интерфероны человека - это семейство более чем пятнадцати белков с антивирусной, антиростовой и иммунорегуляторной активностью [Pestka et al., Ann. Rev. Biochem. , 56: 727 (1987)]. Терапевтическая активность альфа-интерферонов человека показана для раковых заболеваний и вирусных инфекций. Например, рекомбинантные интерфероны (ИНФ α- 2a, ИНФ α- 2b, ИНФ α- 2c), интерфероны, полученные с помощью клеточных линий (ИНФ α- n1) и лейкоцитов (ИФН α- n3) в настоящее время используются для терапии заболеваний, вызываемых Condyloma acuminata, и гепатитов (Weck et. al., Am. J. Med., 85 (Suppl 2A): 159 (1988); Korenman et al., Annal. Intern. Med; 114 : 629 (1991); Friedman - Kien et al., JAMA, 259 : 533 (1988)) для регрессии некоторых злокачественных новообразований [Baron et al., JAMA, 266 : 1375 (1991)], лечения связанной со СПИДОМ саркомы Капоши [Physicians Desk Reference, 47 th edit., eds. Medical Economics Data, Montvale, NJ, p.2194 и 2006 (1993)] и считаются перспективными препаратами для лечения синдрома приобретенного иммунодефицита (СПИД) как самостоятельно, так и в сочетании с другими противовирусными агентами [Hirsch, Am. J. Med., 85 (Suppl. 2A) : 182 (1988)].
Рекомбинантные интерфероны могут быть получены с помощью генно-инженерных технологий, например при крупномасштабном культивировании трансформированных Escherichia coli [Gocddel et al., Nature, 287 : 411 (1980); Streuli et al. , Science, 209 : 1343 (1980)]. Однако генно-инженерные интерфероны представлены белками только одной определенной формы, которая не подвергается посттрансляционной модификации и не идентична природным интерферонам, что может ограничивать биологическую активность соответствующей композиции. Например, препараты Intron® A (ИНФ α- 2b) [Schering Plough], и Roferon® A (ИНФ α- 2a) [ Hoffman - La Roche] представляют собой исключительно α-2- форму интерферона.
Интерфероны полученные с помощью природных источников, таких как лимфобластоидная клеточная линия человека Namalwa [Mizrahi, Meth. Enzymol., 78 : 54 (1981); Phillips et al., Meth. Enzymol., 119 : 35 (1986)] и лейкоциты периферической крови человека [Mogensen et al., Pharmacol. Ther. Part C, 1 : 369 (1977); Cantell et al., Methods Enzymol., 78 : 29 (1981); Horowitz, Methods Enzymol., 119 : 39 (1986)] представлены многими формами, каждая из которых имеет определенную структуру и биологическую активность.
Такие "природные" интерфероны по мнению некоторых исследователей способны обеспечить потенциально более лучший терапевтический эффект по сравнению с рекомбинантными белками. Например, природный альфа-интерферон может быть использован в четырехкратно меньшей дозе по сравнению с рекомбинантными для лечения заболеваний, вызываемых Condyloma [см., например, Physicians Desk Reference как указано выше, стр. 1879 и 2194]. Наиболее значительным преимуществом природного лейкоцитарного интерферона как терапевтического агента является его низкая иммуногенность при введении больным, нуждающимся в таком лечении. Показано, что у больных, получающих указанные выше рекомбинантные интерфероны и лимфобластоидный интерферон Wellferon® ИФН α- n1 [Burroughs Wellcome] образуются нейтрализующие антитела к интерферону [Loket et al., Heratology, 12 : 126 (1990). Jacobs et al., J, Biol. Resp. Mod., 7 : 447 (1988). Weck et al. , J. Interferon Res., 1 (Suppl):37 (1989)]. Однако у больных, получающих лейкоцитарный интерферон (ИНФ α- n3 или частично очищенный интерферон Кэнтелла), в сыворотке крови не выявляются антитела к интерферону [Von Wussow et al., Lancet, 2 : 635 (1987); Liao et al., J. Infect. Dis. , 165 : 757 - 760 (1992)]. Образование нейтрализующих антител к интерферону способно блокировать терапевтический эффект данных препаратов и поэтому может быть существенным фактором клинического применения интерферонов. Однако показано, что многие больные, резистентные к рекомбинантному α- интерферону, отвечают на лечение природным α- интерфероном.
Опубликованные методы получения α- интерферона из лейкоцитов периферической крови человека неэффективны и очень дороги [Mogensen et al., (1977); Cantell et al. , (1981); Horowitz, (1986), все работы указаны выше]. Метод получения природного альфа-интерферона с помощью лейкоцитов, описанный Кэнтеллом, приводит к получению относительно низкого титра интерферона (60000 ЦПДЕд/ мл) при сравнительно высокой клеточной плотности (1•1-7 клеток/мл).
Известные методы очистки природного альфа-интерферона тоже имеют свои ограничения. Например, в результате использования последовательного метода преципитации достигается только частичная очистка лейкоцитарного интерферона человека при его получении в больших объемах (Cantell et al., Merh. Enzymol. , 78:499 (1981)). Полученный препарат обладает 1%-ной чистотой и состоит из многих типов интерфероновых белков.
Berg et al.[Meth.Enzymol., 78 : 487 (1981)] описали способ очистки лейкоцитарного интерферона человека с помощью моноклональных антител, конъюгированных с Сефарозой 4B. И наконец Horowitz (1986, см. выше) опубликован метод получения и очистки лейкоцитарного интерферона человека в больших объемах при использовании в качестве источника данного соединения лейкоцитов периферической крови. Исследователь применял способ преципитации Кэнтелла или аффинную хроматографию на основе антител NK2.
В настоящее время количество препаратов α- интерферона, который используется для лечения различных состояний, постоянно возрастает. Однако все препараты обладают многочисленными побочными эффектами. К ним относятся такие простудные реакции, как лихорадка, изменение формулы крови, желудочно-кишечные расстройства, например рвота и диарея, нарушение работы почек, легких, аллергические реакции, такие как спазм бронхов, анафилаксия или кожные высыпания, а также выпадение волос.
И хотя некоторые побочные эффекты сами по себе не очень серьезны, они могут вызвать существенные негативные реакции у больных, которые вынуждены принимать значительные дозы препаратов интерферона в течение длительного промежутка времени. Например, в некоторых случаях, скажем при лечении ассоциированной со СПИДом саркомой Капоши или симптоматического СПИДа, эффективная доза интерферона вызывает побочные эффекты, которые опаснее, чем проявление самого заболевания на определенной стадии. В клинических исследованиях по этим показателям больные были вынуждены отказаться от лечения из-за возникших побочных эффектов, несмотря на продолжительный положительный эффект.
Таким образом, остается потребность в разработке улучшенных α- интерфероновых композиций, которые характеризуются очень низкой токсичностью и высокой чистотой, а также вызывают минимальные побочные эффекты при соответствующей терапии.
Один аспект настоящего изобретения относится к практически чистым α- интерфероновым композициям, содержащим природную смесь α- интерферонов человека различных типов и подтипов. Более того, эта композиция характеризуется очень высокой специфической активностью, около 4•108 Ед/мг. Существенно, что такая композиция отличается способностью оказывать терапевтический эффект в отношении заболеваний человека при значительной редукции побочных эффектов, в норме ассоциированных с α- интерферонами.
Другим предметом изобретения является фармакологическая композиция, содержащая в качестве одного активного ингредиента эффективное количество α- интерфероновой композиции, как описано выше, в сочетании с фармакологически приемлемым носителем. Фармакологическая композиция может содержать α- интерфероновую композицию в комплексе с каким-либо другим подходящим фармакологическим агентом, полезным для лечения данного заболевания или нарушения.
Другой аспект изобретения заключается в разработке нового метода получения описанной выше α- интерфероновой композиции с помощью лейкоцитов периферической крови. Метод отличается некоторыми оптимизированными подходами к индукции и очистке интерферона, которые позволяют эффективно и с минимальными затратами получать большие количества указанной композиции.
Еще одним аспектом настоящего изобретения является использование очищенной α- интерфероновой композиции, описанной выше, для лечения различных заболеваний или нарушений иммунной системы, терапии рака и/или вирусных инфекций. Эти методы лечения включают назначение заявленных фармакологических композиций как самостоятельно, так и в сочетании с другими лекарствами, соответствующим больным.
Другие аспекты и преимущества настоящего изобретения описаны ниже при более детальном изложении существа изобретения и предпочтительных вариантах его использования.
Фиг. 1 определяет титр интерферона и продуктивность лейкоцитов как функцию плотности клеток. Продуктивность выражается в интерфероновых ИРМА-единицах на 106 лейкоцитов (ИРМА - иммунорадиометрический анализ).
Фиг. 2 определяет титр интерферона как функцию индукции культурального объема 6-литровых плоскодонных флаконов.
Фиг. 3. Показан типичный профиль элюции ИФН α- n3a с хроматографической колонки, заполненной сефарозой.
Фиг. 4A. Показан типичный профиль элюции ИФН α- n3a с полупрепаративной C4 - колонки (10х250 мм), полученный методом обратно-фазовой жидкостной хроматографии высокого разрешения (RP - HPLC от анг reverse - phase high performance liquid chromatography). Можно различить пики 1a, 1b и 2 - 6. На фиг. 4B для сравнения показан контрольный "беспиковый" профиль элюции, полученный тем же методом.
Фиг. 5 иллюстрирует взаимосвязь между биологической активностью и гидрофобностью белков в RP-HPLC -пиках.
Фиг. 6. Показана окрашенная Кумасси синим электрофореграмма, полученная при осуществлении электрофореза ИФН α- n3a в полиакриламидном геле в присутствии додецилсульфата натрия (SDS - PAGE) в невосстанавливающих условиях. Колонка A: положение предварительно окрашенных низкомолекулярных маркеров (Biorad). Колонка B: положение ИФН α- n3a (не фракционированного). Колонка C: RP - HPLC - пик 1a. Колонка D: RP - HPLC - пик 1b. Колонка E: RP - HPLC - пик 2. Колонка F: RP - HPLC - писк 3. Колонка G: RP - HPLC - пик 4. Колонка H: RP - HPLC - пик 5. Колонка I: RP - HPLC - пик 6.
Фиг. 7. Показана окрашенная Кумасси синим электрофореграмма ИФН α- n3a, полученная при осуществлении SPS-PAGE, в восстанавливающих условиях. Обозначения колонок те же, что и на фиг. 6.
Фиг. 8. Полученные с помощью иммуноокрашивания результаты Вестерн-блоттинга ИФН α- n3a в невосстанавливающих условиях. Обозначения колонок те же, что и на фиг. 6.
Фиг. 9. Полученные с помощью иммуноокрашивания результаты Вестерн-блоттинга ИФН α- n3a в восстанавливающих условиях. Обозначения колонок те же, что и на фиг. 6.
Настоящее изобретение относится к улучшенной α- интерфероновой композиции, которая состоит из ИФН α- n3a, способу получения данной композиции и фармацевтической композиции, содержащей ИФН α- n3a-композицию для лечения заболеваний, поддающихся терапии с помощью α- интерферона.
Композиция.
α- Интерфероновая композиция, согласно настоящему изобретению представляет собой очищенную смесь природных α- интерферонов человека различных типов и субъединиц. Как подробно описано ниже, эта композиция отличается от других продуктов на основе природного α- интерферона, например ИФН α- n3a, более чем в два раза высокой антивирусной специфической активностью, по крайней мере в пять раз меньшей загрязненностью и пирогенной активностью.
Совокупность белков ИФН α- n3a содержит протеины различных аминокислотных последовательностей, которые могут быть определены на основе опубликованной нуклеотидной последовательности гена интерферона. Более точно, композиция на основе ИФН α- n3a состоит из смеси по крайней мере шести типов белков α- интерферона человека. На основании N-терминальной последовательности, описанной ниже, по крайней мере один тип интерферона выбирается из совокупности α 2b, α2c или их комбинации. По крайней мере второй тип выбирается из совокупности α 4a, α 4b и/или α 16 или их комбинации. Соответственно, третий из шести типов интерферон выбирается из α 7a, α 7b и/или α 7c или их комбинации, четвертый - из совокупности α 8a, α/ 8b и/или α 8c или их комбинации. Пятый тип представляет собой ИФН α- 10a и наконец шестой выбирается из совокупности α 17a, α 17b, α 17c, α 17d, α 21a и/или α 21b или их композиции. Последовательности всех указанных интерферонов могут быть определены на основе различных баз данных, в том числе базы данных фирмы Intell Genetics, Inc ( Genbank® database, Intell Genetics, Inc., Mountain, View, Ca).
Смесь α- n3a интерфероновых белков не содержит ИФН- α 14, ИФН α -2a и ИФН - α 1.
Описанные выше типы и подтипы α- интерферона, содержащиеся в заявленной композиции, могут быть охарактеризованы с помощью обратно-фазовой жидкостной хроматографии высокого разрешения (RP - HPLC), которая разделяет типы интерферона с соответствии с их относительной гидрофобностью, как детально описано в примере 3. Композиция на основе ИФН α- n3a имеет гидрофобность, определимую на RP-HPLC - колонке путем элюции 40 - 60%-ным ацетонитрилом. N- и C-терминальные последовательности указанных выше типов и подтипов α- интерферона, составляющих заявленную композицию, могут быть определены, как детально описано в примере 6 (разделы B и D).
Кроме того, заявленная ИФН α- n3a - композиция характеризуется по своей активности в различных биологических системах. Как подробно раскрывается в примере 4A, антивирусная активность (см., например, Linnette et al., Cancer Therapy and Control, 1:109 - 120 (1990)) тестируется на клетках человека, быка и кролика/ Антивирусная специфическая активность ИФН α- n3a, определенная на клетках быка и человека, составляет (1-10•108 Ед/мг и в целом равна или превышает 2•108 Ед/мг). Единица активности интерферона в данном исследовании определяется как обратная величина разведения в 50%-ной конечной точке и корректируется в соответствии с контрольным стандартам Национального института здравоохранения (Международный референс-препарат лейкоцитарного интерферона, 6a 23-902-530). Средняя специфическая антивирусная активность ИФН α- n3a на клетках человека HEp-2 составляет около (4 • 10)8 Ед/мг.
Показано, что ИФН- α 2, представленный подтипами α 2b и α 2c, обнаруживается в пиках 1а и 1б, и на его долю приходится около 50% общей белковой массы α- -n3a. Однако при раздельном исследовании пиков (см. табл. 4) данный тип интерферона отвечает только за 25% всей антивирусной активности заявленной композиции. Более того, как детально показано в примере 8, ИФН α-n3a в 10-100 раз более эффективен, чем эквивалентное количество рекомбинантного интерферона α 2, при определении in vitro их специфической активности по отношению к вирусу СПИДа на моноцитах.
Кроме того, биологическая активность интерферона определялась и по его антипролиферативному действию на клетки человека, как описано в примере 4Б [(см., например, Gillis et al., J. Immunol. 12 : 2027 (1978)]. Специфическая активность составляла (1-10) • 108 Ед/мг и в целом была равна или превышала 1 • 108 Ед/мг. Средняя специфическая антипролиферативная активность на клетках человека линии Daudi составляла около 1,5 • 108 Ед/мг. Единица активности интерферона в данном исследовании определялась как обратная величина разведения в 50%-ной конечной точке.
Биологическая активность (≥ 99%) заявленной композиции, определяемая в антивирусном исследовании, может быть нейтрализована поликлональной антисывороткой овцы (полученной к очищенному ИФН α- n3), специфической к лейкоцитарному интерферону человека, при осуществлении стандартного нейтрализующего теста [Y. Kawada, Meth. Enzymology, 119 : 558-573 (1986)].
Дополнительно в пирогенном тесте, который проводили на кроликах (Пирогеновый тест. USP XXII/NF XVII, Фармакопейная Конвенция США, с. 1515 (1990)) увеличение средней температуры тела кроликов на 0,2oC свидетельствует о чрезвычайно низкой пирогенной активности заявленной композиции. Действительно, такой результат по крайней мере в пять раз меньше, чем определяемый для ИФН α- n3.
Кроме того, были определены и физические свойства заявленной композиции на основе ИФН α- n3a. Например, она отличается значительной чистотой, а именно состоит более чем на 95% из α- интерфероновых белков человека. Менее чем 5% составляют примеси, такие как α- интерфероновые белки нечеловеческой природы, нуклеиновые кислоты или ферменты, липиды и тому подобное. Точнее, заявленная композиция на 98 или более процентов состоит из α- интерфероновых белков человека, а наиболее предпочтительно - примерно на 99%.
Чистота композиции определяется электрофорезом в полиакриламидном геле в присутствии додецилсуфата натрия (SDS - PAGE) и Вестерн-блоттингом, как детально изложено в примере 5. В соответствии с методом получения, описанном ниже, чистота композиции увеличивается в 800 раз в сравнении с первой стадией очистки (аффинная хроматография с помощью антител N K2) и примерно в два раза по сравнению со второй стадией очистки с помощью суперозы. В целом, общая степень чистоты ИФН α- n3a примерно в 12 000 раз выше по сравнению с сырым продуктом, полученным из лейкоцитов, как детально описано ниже.
Анализ композиции с помощью Вестерн-блоттинга (фиг. 8 и 9) в невосстанавливающих и восстанавливающих условиях соответственно показывает, что все белковые полосы идентифицируются как интерфероны человека с уровнем загрязнения нефракционированного препарата около 1% или менее. Последующее фракционирование с помощью RP - HPL C приводит к тому, что какое-либо загрязнение не определяется ни в одном пике.
Двумерный анализ в геле нефракционированного ИФН α- n3a показал, что во всех образцах выявляются многочисленные пятна, распознаваемые моноклональным антителом LIT-1 (см. пример 5, часть A), свидетельствуя о том, что эти пятна - интерфероновые белки. Согласно методу SDS-PAGE, примерная мол. масса композиции составляет 16-27 кДа. Кроме того, композиция характеризуется изоэлектрической точкой в диапазоне 5,0 - 7,5, более точно в диапазоне 5,5 - 6,5. Количество "пятен" и мол.масса белков каждого пика приведены в примере 5, табл. 8.
Биохимические свойства интерфероновой композиции согласно настоящему изобретению, например определяемые методом аминокислотного анализа и амино- и карбокситерминального секвенирования, детально описаны в примере 6. Амино- и карбокситерминальное секвенирование показывает, что интерференовая композиция содержит интактные молекулы интерферона, которые состоят примерно из 165-166 аминокислотных остатков на молекулу. Кроме того, в композиции обнаружены две дисульфидные связи.
Далее, композиция характеризуется содержанием углеводов, например сахаров, нейтральных сахаров и сиаловых кислот. Результаты таких исследований тоже подробно описаны в примере 6B. Предложенная композиция отличается O-связанным углеводным гликозилированием, причем среди углеводов различают галактозамин (GalN), глюкозамин (GlcN), галактозу, глюкозу, маннозу и сиаловую кислоту. В среднем, композиция содержит ≤ 6 моль сахаров на моль интерфероновых белков, точнее 1-3 моль сахара/моль интерферона. Гликозилирование ИФН α- n3a должно происходить по другому сайту, нежели остаток Asn, который отвечает за N-связанное гликозилирование, поскольку ИФН α- n3a не содержит ИФН - α 14, единственного типа α- интерфероновых белков, отличающегося потенциальной возможностью N-гликозилирования по остатку Asn. Сайт O-связанного гликозилирования определяют путем ферментативного расщепления в процессе аминокислотного секвенирования. Результаты исследования показывают, что гликозилирование происходит по остатку треонина в положении 106 (пример 6B).
Другое биохимическое свойство заявленной ИФН α- n3a-композиции включает в себя наличие связанных сайтов для моноклональных антител N K 2 (Celltech, U.K.). Согласно сообщениям антитела N K2 распознают только одну подгруппу α- интерферона, а именно α 2 (a/b/c), α 8 (a/b/c), α 10a, α 17 (a/b/c/d), α 7 (a/b/c), α 6 и α 21 (a/b). Они не распознают интерфероны α 1, α 5, α 10b, α 14 и α- омега 1. Кроме того, ИФН α- n3a-композиция имеет pH-стабильность в интервале 2-9 и стабильна к нагреванию при 56oC в течение двух и более часов.
Заявленные композиции на основе ИФН α- n3a существенно отличаются от других α- интерферонов как природных, так и рекомбинантных, которые в настоящее время поступают на рынок и используются для клинических нужд. Наиболее существенное отличие - это гораздо меньшая токсичность ИФН α- n3a по сравнению с известными α-/ интерфероновыми композициями. В коммерческой и специальной литературе, посвященной ИФН α- n3, отмечается, что природные α- ИФН, полученные с помощью лейкоцитов периферической крови, при тестировании на большом количестве больных обладают характерными для α- интерферонов побочными эффектами.
Другие различия между заявленной композицией и известными природными α- интерферонами показаны выше. В сравнении с известным препаратом на основе ИФН α- n3, который используется в клинических исследованиях, новая ИФН α- n3a-композиция имеет в два раза большую специфическую антивирусную активность, равную примерно 4 • 108 Ед/мг. Заявленная композиция имеет уровень загрязнения около 2% по сравнению с 5-10% известных продуктов. Более того, заявленная композиция имеет очень низкую пирогенную активность, около 0,2oC, в сравнении с 1oC, что характерно для известных препаратов ИФН α- n3. Кроме того, различия затрагивают и метод получения заявленных и известных продуктов.
Различия в токсичности между заявленным и рекомбинантным интерферонами еще более значимы. В дополнении к тому обстоятельству, что данная композиция содержит смесь α- интерферонов в сравнении с одним типом белка рекомбинантного продукта, природный α- интерферон не получают с помощью бактерий как рекомбинантный и поэтому он не загрязнен бактериальными белками, которые у человека могут вызывать нежелательные побочные эффекты.
Для того, чтобы убедиться в отсутствии побочных эффектов заявленной согласно настоящему изобретению композиции, в примере 7 детально описаны результаты экспериментов по определению токсичности данной композиции, которые проводились в соответствии с методикой, утвержденной ВОЗ и описанной W.B. Abrams et al. , Marcel Dekkev, Inc., chap 10, pp. 195-216 (1984). Такие исследования токсичности, проведенные на больных, получавших заявленную композицию в качестве терапевтического средства, практически не обнаружили у нее побочных эффектов.
В действительности, в табл. 16 показаны различия в побочных эффектах, которыми обладают известные продукты на основе α- ИФН, в сравнении с результатами клинических испытаний композиции, заявленной согласно настоящему изобретению. Эти данные показывают, что новая композиция практически не обладает побочными эффектами, что является большим преимуществом для ее применения на больных в течение продолжительного промежутка времени. Более того, могут быть применены и более высокие дозы заявленной композиции без риска побочных эффектов в сравнении с известными α- ИФН композициями.
Авторам в настоящее время неизвестно, почему заявленная композиция обнаруживает такую непредвиденно высокую степень редукции побочных эффектов. Не желая быть связанными теорией, авторы считают, что причины значительного снижения токсичности и побочного действия данной композиции и вследствие этого ее большей терапевтической эффективности, могут быть основаны на том, что различные типы α- интерферона человека, составляющие композицию, связываются более, чем с одним эпитопом или рецептором на клетках больного, которому назначена данная композиция. Кроме того, одной из причин неожиданных свойств заявленной композиции может быть очень низкое содержание в ней примесей.
Далее, поскольку специфическая активность композиции в антивирусных и антипролиферативных исследованиях выше, чем для других α- интерферонов, для лечения больных могут использоваться меньшие дозы полученного интерферона и эти малые дозы могут быть меньше, чем токсическая доза композиции, если она вообще обладает токсическим эффектом. Кроме того, посттрансляционное гликозилирование белков композиции может способствовать дезагрегации, приводя к ослаблению побочного действия. Процессинг, в то же время, может уменьшить оксидацию и химическую модификацию аминокислотных остатков α- интрефероновых белков. Углеводы α- интерфероновых белков надежно защищаются в процессе получения заявленной композиции и поэтому сходны с природными α- интерферонами человеческого организма. Указанный процесс может приводить к тому, что в заявленной композиции главным образом преобладают активные α- интерфероновые мономеры и образуется небольшое количество неактивных α- интерфероновых олигомеров.
2. Способ получения композиции.
В целом способ получения α- интерфероновой композиции согласно настоящему изобретению включает как процесс индукции, так и процесс чистки. Процесс индукции детально описан в примере 1, а процедура очистки - в примере 2.
В процессе индукции ИФН α- n3a образуется индуцированной вирусом Сендай суспензией лейкоцитов периферической крови (ЛПК) человека. Основными факторами, которые значительным образом влияют на количество α- n3a интерферона, который образуется ЛПК, являются следующие: плотность клеток в процессе индукции, концентрация бикарбоната натрия в индукционной среде, концентрация сыворотки крови человека, свободной от гамма-глобулинов, концентрация и тип праймера, кинетика индукции, объем культур во флаконах, скорость перемешивания культур, температура индукции, состав среды, а также концентрация и свойства вируса Сендай.
В соответствии с указанным процессом индукции кровь забирают у здоровых доноров и получают ЛПК, собирая лейкоцитарную пленку и лизируя эритроциты с помощью хлорида аммония, предпочтительно 0,83%-ной концентрации. Затем лейкоциты суспендируют в среде индукции до получения желаемой плотности клеток.
Плотность клеток варьирует в интервале (1-10) • 106 кл/мл. Относительная эффективность продукции α- интерферона может быть выражена как количество данного белка, которое образуется в расчете на клетку при определенной плотности клеток. Авторы определили, что при возрастающей продукции α- интерферона в соответствии с увеличением плотности клеток оптимальная продукция α- интерферона ЛПК человека происходит при клеточной плотности 4 • 106 кл/мл. Продуктивность может быть определена путем иммунорадиометрического анализа (ИРМА) [Gelltech Ltd. ] или по цитопатическому действию (ЦПД), как описано ниже. В этих исследованиях титры интерферона определяются в соответствии со стандартом и выражаются в Ед/мл (для ИРМА) и МЕд/мл (международные единицы на 1 мл для исследования ЦПД). В то время как максимальный титр интерферона (около 50000 ИРМА Ед/мл, что эквивалентно ~100000 ЦПД Ед/мл) достигается при плотности клеток в процессе индукции 1 • 107 лейкоцитов/мл, наиболее высокая продукция интерферона (около 43000 ИРМА Ед/мл, что эквивалентно ~86000 ЦПД Ед/мл) отмечается при плотности 4 • 106 кл/мл. Таким образом, продуктивность клеток при плотности 107 кл/мл составляет примерно половину от таковой при клеточной плотности 4 • 106 кл/мл (cм., например, фиг. 1).
Среда индукции представляет собой минимальную необходимую среду (МЕМ) желаемой концентрации 0,5Х - 1,5Х, содержащую солевой раствор Эрла, pH 7,4, дополненный L-глутамином, не необходимыми аминокислотами, трицином (4,46 мг/мл) и неомидин сульфатом (24 мкг/мл). Текущие данные показывают, что наиболее высокий индукционный титр наблюдается при концентрации МЕМ 0,85Х - 1,15Х, которая имеет осмолярность, максимально приближенную к осмолярности физиологических жидкостей организма.
Кроме того, необходимо, чтобы среда индукции содержала сыворотку крови человека, лишенную IgG, для обеспечения продукции α- интерферона. Эффективная концентрация сыворотки составляет 0,1 - 1,5 мг/мл, более предпочтительно, 0,1 - 1,0 мг/мл. Преимущественно используется концентрация 0,4 мг/мл.
Более высокая концентрация сыворотки неэффективна для продукции α- интерферона, возможно из-за присутствия ингибирующих факторов. Оптимальная концентрация сыворотки, найденная при осуществлении описанного здесь процесса индукции, намного ниже, чем рекомендуемая концентрация, равная 2,4 мг/мл, согласно методу Кэнтелла. Использование более низкой концентрации сыворотки во время индукции существенно уменьшает загрязненность α- интерферонов сывороточными белками во время нисходящих процессов их очистки.
Процесс индукции оптимизируется добавлением некоторых других компонентов в среду. Желательно, чтобы индукционная среда содержала высокую концентрацию бикарбоната натрия (NaHCO3) в противоположность известным методам, которые предусматривают использование этого вещества в низкой концентрации или не предусматривают вообще. Увеличение концентрации NaHCO3 в МЕМ оказывает значительный эффект на выход интерферона (например, в 3-4 раза). Предпочтительная концентрация NaHCO3 составляет около 0,3 - 2,5 мг/мл, более предпочтительная - 1,7 - 2,2 мг/мл. Концентрация NaHCO3, составляющая 2,2 мг/мл, по крайней мере в два раза увеличивает титр интерферона, полученного в соответствии с настоящим изобретением.
Другим фактором, лимитирующим продукцию интерферона ЛПК, могут быть необходимые питательные компоненты, например редкие минералы, витамины или жирные кислоты. Добавление некоторых витаминов в индукционные культуры согласно настоящему изобретению способствует увеличению продукции интерферона. Добавление витаминов C и B3 (ниацина) по отдельности в индукционные культуры увеличивает выход интерферона на 20-30% в сравнении с культурами, не получавшими указанных витаминов. Эффективная концентрация витамина C для увеличения образования интерферона составляет 0,0002 - 2 мг/мл. Более высокие концентрации витамина C (> 2 мг/мл) ингибируют образование интерферона ЛПК. Эффективная концентрация витамина B3 для увеличения выхода интерферона составляет 0,5 - 1000 мкг/мл. Другие соединения, например сульфат железа, масляная кислота, токоферолфосфат, токоферол, глюкоза и холестерол, не оказывают воздействия на продукцию α- интерферона или ингибируют ее.
Настоящее изобретение относится и к способу получения интерферона в больших объемах. Объем индукционной среды во флаконах может варьировать от 150 мл до 6 л или более. Объем индукционной культуры может быть увеличен до полной вместимости флаконов (или уменьшен), не влияя на выход интерферона. Более того, поскольку флаконы могут быть заполнены полностью, способ согласно настоящему изобретению позволяет увеличить выход интерферона при минимальном количестве флаконов, таким образом уменьшая стоимость процесса и расход посуды. Крупномасштабная индукция в объемах до нескольких тысяч литров может быть достигнута при использовании многослойных больших флаконов или биореакторов в описанных согласно настоящему изобретению условиях индукции.
Сырой или очищенный лейкоцитарный α- интерферон затем добавляют в качестве праймера к суспендированным в индукционной среде ЛПК. При использовании в качестве праймера сырого и очищенного природного лейкоцитарного α- интерферона индуцируется образование сходных количеств продукта лейкоцитами. Лейкоциты могут быть праймированы в количестве 1 - 100000 МЕд/мд, предпочтительно около 10 - 100 МЕд/мл сырого α- интерферонового супернатанта или очищенного α- интерферона. Под "сырым интерфероном" понимают неочищенный продукт, полученный излагаемым здесь способом индукции. Под "очищенным интерфероном" понимают ИФН α- n3a, полученный излагаемыми здесь методами индукции и очистки.
Понятно, что в качестве праймеров могут быть использованы другие интерфероны, например β- ИФН, γ- ИФН и, возможно, другие цитокины. Например, природный γ- ИФН человека в концентрации 100 - 1000 МЕд/мл может стимулировать продукцию α- интерферона, но приводит к более низкому выходу продукта, чем при использовании в качестве праймеров сырого или очищенного природного α- интерферона.
Найдено, что оптимальное время начала праймирования примерно составляет 2-3 ч до внесения вируса Сендай при 36oC. Изменение этого времени существенным образом изменяет количество интерферона, секретируемого лейкоцитами. Праймирование лейкоцитов менее, чем за один час до внесения вируса, снижает выход интерферона, более чем за 3 часа - вообще не приводит к каким-либо ощутимым результатам. Предпочтительно, на стадии праймирования лейкоциты в количестве 107 кл/мл обрабатывают 20 МЕд/мл сырого интерферона примерно за 3 часа до внесения вируса Сендай.
После праймирования в суспензию лейкоцитов добавляют вирус Сендай, важнейший индуктор интерферонов. Выход интерферона существенным образом варьирует в зависимости от различий в препаратах вируса. Например, это может быть обусловлено присутствием различных количеств дефектных интерферирующих частиц вируса Сендай в некоторых препаратах. Высокие количества этих частиц могут оказать ингибирующее воздействие на выход интерферона.
В целом, адсорбция вируса Сейдай на лейкоцитах происходит наиболее эффективно при высокой плотности клеток, указанной выше, и определенной концентрации вируса. Оптимальная концентрация вируса, необходимая для максимально эффективной индукции интерферона, должна быть определена эмпирически и обычно составляет около 50 - 500 гемаглютинирующих (ГА) единиц/мл (конечная концентрация в тканевой культуральной жидкости). Более предпочтительно, когда концентрация вируса составляет 50 - 250 ГА единиц/мл.
Продукция интерферона может быть максимизирована применением метода абсорбции/разведения, при котором концентрированные клеточные культуры разбавляются. Согласно данному методу вирус Сендай в конечной концентрации около 375 ГА единиц/мл адсорбируется на лейкоцитах при плотности клеток 107 кл/мл примерно в течение 1 ч и 36oC. Предпочтительно, культуры разводят в 2,5 раза индукционной средой, содержащей сыворотку без γ- глобулинов, до конечной плотности клеток 4•106 кл/мл и концентрации вируса Сендай около 150 ГА Ед/мл. При любых условиях обнаружено, что метод адсорбции/разведения примерно на 10% увеличивает титр интерферона в сравнении с тем процессом, когда данный метод не используется. Среда разведения может быть комнатной температуры или предварительно нагрета до 36oC и может содержать либо не содержать праймеры.
Индукцию необязательно проводить при использовании метода адсорбции/разведения. В этом случае лейкоциты при определенной клеточной плотности ((1-10) • 106 кл/мл) могут быть праймированы сырым интерфероном примерно за 3 часа до внесения вируса Сендай в установленных концентрациях. Затем культуры инкубируют в течение 15-16 ч и собирают интерферон аналогичным образом.
Температура инкубации лейкоцитарных культур при осуществлении заявленного способа получения α- интерферона составляет 35 - 37oC. Однако оптимальная температура инкубации составляет 36oC. Согласно методу Кэнтелла оптимальная температура инкубации для обеспечения продукции интерферона составляет 37,5oC [Mogense et al., (1977); Cantell et al., Methods Enzymology., (1981) и Horrowitz (1986), все работы указаны выше]. Тем не менее, авторы обнаружили, что температура менее 37oC приводит к увеличению выхода интерферона. Результаты индукции в тройных 6-литровых флаконах при плотности клеток (4-5) • 106 кл/мл, инкубированных при 35, 36 или 37oC во время процесса индукции, показали, что инкубация при 36oC на 20-50% увеличивает выход интерферона, по сравнению с температурой 35 или 37oC.
Скорость перемешивания индукционных культур с помощью магнитной мешалки тоже является важным фактором для поддержания оптимальной аэрации и необходимого перемешивания компонентов. При скорости вращения магнитной мешалки 72 rpm из 500-мл культуральных флаконов, содержащих 4 • 106 кл/мл, получают 17 000 ИРМА ЕД/мл интерферона. При увеличении скорости перемешивания до 170-250 rpm количество образующегося интерферона существенно увеличивается до 30000 ИРМА Ед/мл. Таким образом, скорость перемешивания между 100 и 300 rpm, предпочтительно около 170 rpm, необходима для обеспечения максимальной продукции интерферона в процессе индукции.
Общее время инкубации после добавления вируса Сендай составляет 1-48 ч, предпочтительно 15-22 ч. После соответствующего времени инкубации при 36oC суспензию центрифугируют примерно при 2500 rpm для удаления клеток и дебриса и собирают сырой α- интерферон.
В табл. 1 приведены суммарные различия заявленного способа индукции α- интерферона (А) и описанного Кэнтелллом [Cantell et al., (1981), см. выше] (Б).
Таблица 1.
Среда индукции
А. Среда Игла МЕМ с добавлением солей Эрла, L-глутамина, не необходимых аминокислот, 4,6 мг/мл трицина, 24 мкг/мл неомицина при pH 7,4.
Б. Среда Игла МЕМ без фосфата с добавлением 3 мг/мл трицина, 24 мкг/м неомицина при pH 7,4.
NaHCO3, сыворотка крови человека без γ- глобулинов, добавки.
А. 0,1 - 2,5 мг/мл NaHCO3, 0,1 - 1,5 мг/мл сыворотки и витамин C или витамин B (ниацин).
Б. Только 2,4 мг/мл сыворотки.
Объем флаконов.
А. 150 мл, 500 мл, 2 л и 6 л. Объем культуры может составлять 5 - 100% вместимости флаконов.
Б. 2 л или 6 л. Объем культуры составляет менее 50% вместимости флаконов.
Плотность клеток.
А. 107 кл/мл на стадии праймирования и адсорбции вируса с последующим разбавлением через 1 ч после добавления вируса до 4 • 106 кл/мл. Или 4 • 106 кл/мл на стадии праймирования и адсорбции вируса и в течение ночной инкубации.
Б. 107 кл/мл.
Праймер и условия праймирования.
А. Сырой супернатант α- интерферона или очищенный α- интерферон в количестве 10 - 50 МЕд/мл; праймирование 3 ч при 36oC.
Б. Сырой супернатант α- интерферона в количестве 100 - 200 МЕд/мл, праймирование 2 ч при 37,5oC.
Вирус Сендай.
А. 125 - 500 ГАЕд/мл на стадии адсорбции вируса с последующим разведением до 50 - 200 ГА Ед/мл.
Б. 100 - 150 ГА Ед/мл.
Скорость перемешивания и условия инкубации.
А. 130 - 250 rpm; 15 - 20 ч при 36oC.
Б. Не указано; 17 ч при 37,5oC.
После индукции титр интерферона определяют с помощью ИРМА [Celltech, Berkshire, UK] или в исследовании цитопатического действия (ИПД) вируса везикулярного стоматита (ВВС) (штамм Индиана АТСС # VR-158) на эпидермальные клетки человека НЕр-2 (АТСС CCL 23) согласно опубликованной методике [Linette et al. , Cancer Therapy and Control, 1 : 109 - 120 (1990)]. Титр интерферона, определяемый с помощью ИРМА, выражают в ИРМА Ед/мл или Ед/мл, а титр, полученный методом ЦПД, - в МЕд/мл (международные единицы/мл) в сравнении с лейкоцитарным стандартом Национального Института здравоохранения США (G2 23-902-530). В методе ИРМА в качестве детекторных антител используют радиоактивно-меченные моноклональные антитела N КА. Основываясь на специфическом узнавании эпитопов интерферона N КА-антителами, ИРМА-титр бывает значительно ниже, чем титр, определяемый по ЦПД. Обычно ИРМА-титр сырого интерферона после индукции составляет примерно половину ЦПД-титра.
Очистка сырого интерферона.
Сырой интерферон очищают с помощью методики, подробно описанной в примере 2. Кратко, методика состоит в следующем. Во-первых, лейкоцитарные культуры, содержащие сырой интерферон после индукции, центрифугируют примерно при 2900 g в течение 15 - 20 мин или фильтруют через соответствующие патронные фильтрационные системы. Эти системы состоят, например, из престерилизованного фильтра Polygard-CR высокой эффективности, патронных фильтров с размерами пор 0,1 мкм, последовательно соединенных с фильтрами Polysep-TP с размерами пор 0,5 мкм (Millipore Corporation, Bedford, MA).
Полученный в результате указанных процедур сырой α- интерфероновый супернатант затем концентрируют в 10 - 100 раз, в основном с помощью фильтрационных систем тангенциального потока. Предпочтительно концентрирование в 50 раз. Сконцентрированный супернатант затем можно отцентрифугировать примерно при 9000 g в течение 30 мин. Однако, если фильтрация осуществляется методом коллекции, дополнительного центрифугирования не требуется.
Полученный на данной стадии сырой сконцентрированный α- интерферон подвергают первой колоночной хроматографии (аффинная колонка с N К2-антителами). С колонки препарат элюируют при pH 2 после интенсивного отмывания. В результате антительной аффинной хроматографии удаляются сывороточные белки человека и другие загрязнители. Интерферон очищается в 8000 раз и восстанавливается примерно 80 - 90% ИРМА-активности интерферона. Чистота препарата на этой стадии обычно превышает 90%.
Элюированный α- интерферон подвергают кислотной инкубации в течение 5 дней при 4oC, затем нейтрализации, концентрированию и гель-фильтрации на хроматографической колонке (колонка с суперозой 12), в результате приводящей к получению заявленной согласно настоящему изобретению композиции. Гель-фильтрация удаляет высоко- и низкомолекулярные примеси, такие как мышиный IgG, вымываемый с N К2-колонки, IgG человека, олигомеры интерферона и продукты его деградации.
Обычный проифль элюции с гель-фильтрационной хроматографической колонки показан на фиг. 3, где основной пик поглощения соответствует элюату α- интерферона. В целом, после осуществления аффинной и гель-фильтрационной хроматографии восстанавливается до 60 - 70% ИРМА-активности интерферона, а степень очистки препарата повышается примерно в 12000 раз. Окончательная чистота препарата интерферона составляет около 99% при определении и Вестерн-блоттингом, противовирусная специфическая активность оценивается в 4 • 108 МЕд/мг (см. пример 4А).
3. Фармацевтическая композиция и методы ее применения
Полученный согласно настоящему изобретению очищенный α-n3а-интерферон может быть использован для лечения различных заболеваний иммунной системы, рака или вирусных инфекций. К этим заболеваниям относятся, но не ограничиваются приведенным перечнем, следующие: болезнь, вызываемая Condyloma acuminata, гепатит B, гепатит C, волосатоклеточный лейкоз, СПИД, саркома Капоши, синдром хронической усталости, генитальный герпес, генитальные бородавки, цервикальная дисплазия, цервикальная карцинома и вагинальная кондилома. Кроме того, полученный согласно изобретению препарат может быть использован для лечения и других заболеваний, чувствительных к α- интерферону.
Заявленные фармацевтические композиции, содержащие эффективное количество α- интерферона, полученного согласно настоящему изобретению, в комплексе с фармацевтически приемлемым носителем могут быть применены для лечения больных, чье состояние поддается терапии с помощью α- интерферона.
Таким образом, терапевтические и фармацевтические композиции, заявленные согласно настоящему изобретению, содержат терапевтически эффективное количество ИФН α- n3a в смеси с фармацевтически приемлемым носителем. Фармацевтические композиции, имеющие антивирусную, антираковую или иммуномодулирующую активность, могут быть использованы в соответствующей форме, например в виде растворов, сиропов, эмульсий, препаратов для инъекций, таблеток, капсул, суппозиториев или препаратов для наружного применения.
Подходящие носители хорошо известны специалистам, работающим в фармакологии (см. например Практическую фармакологию Ремингтона). В качестве примеров могут служит стерильный физиологический раствор и сахара, такие как ксилотол, глицерол, лактоза и сукроза. К другим носителям относятся фосфат кальция, желатин, декстрин, агар, целлюлоза, гидроксиэтилцеллюлоза (для препаратов наружного применения), нефтяной гель, полиэтиленгликоль, пектин, арахисовое масло, оливковое масло, сезамовое масло, сквален и вода.
Дополнительно, носитель или разбавитель может относиться к растворяющемуся с течением времени материалу, например глицеролмоностеарату, или глицеролдистеарату, или их смеси. Кроме этого, подходящие химические стабилизаторы могут быть использованы для повышения стабильности фармацевтического препарата. Соответствующие химические стабилизаторы хорошо известны специалистам в данной области исследований и включают, например, лимонную кислоту и другие агенты доведения pH, хелатирующие или секвестрирующие агенты и антиоксиданты.
Состав фармацевтической композиции, содержащей ИФН α- n3a, может относиться к единичной дозе и быть приготовлен любым подходящим способом. Альтернативно, композиция может быть приготовлена в лекарственной форме, из которой активное начало медленно высвобождается in vivo, как известно из уровня техники. Все методы получения композиции подразумевают соединение активного ингредиента с носителем, который может состоять из одного или более вспомогательных ингредиентов.
Количество очищенного продукта, которое будет эффективно для лечения определенного заболевания или состояния, зависит от его природы и может быть определено стандартными клиническими методами. Согласно настоящему изобретению желательно определить in vitro цитотоксичность подлежащей лечению опухоли, а затем и на подходящей модельной системе на животных, прежде чем тестировать и использовать интерферон для терапии человека.
Методы введения препарата включают, но не ограничиваются указанными, следующие: внутрикожный, внутримышечный, интраперитонеальный, внутривенный, подкожный, оральный, интраназальный и наружный.
Более того, может возникнуть необходимость применения заявленной фармацевтической композиции локально на каком-либо участке, нуждающемся в лечении. Это может быть достигнуто методом локальной инфузии во время хирургической операции, путем инъекции, при использовании катетеров или имплантатов (пористых, непористых, состоящих из желатиноподобного материала, в том числе мембран, например сиаластичных, или волокон). Методы локальной терапии не ограничиваются перечисленными.
Изобретение также относится к фармацевтическим композициям на основе ИФН α-/ n3a, которые применяют в составе липосом, микрочастиц или микрокапсул. В различных вариантах изобретения использование таких лекарственных форм может оказаться полезным для обеспечения постепенного высвобождения составляющих композиции. В особом случае желательно использовать липосомы, связанные с антителами к специфическим опухолевым антигенам (например, к поверхностным клеточным антигенам нейробластомы или SCLC) [Leonetti et al., Proc. Natl. Acad. Sci. USA, 87: 2448 - 2451 (1990); Renneisen et al., J. Biol. Chem., 265: 16337 - 16342 (1990)].
Полученный согласно настоящему изобретению α- интерферон также может быть использован в соответствии с методами и в составе композиций, заявленных авторами, как самостоятельным образом, так и в комплексе с другими терапевтическими или диагностическими агентами, полезными для направленной или дополнительной терапии некоторых типов раковых заболеваний, иммунных нарушений или вирусных инфекций. Ожидается, что ИФН α- n3a может быть использован в комбинации с другими агентами, например антиметаболитами, алкилирующими агентами, алкалоидами барвинка, антинеопластическими антибиотиками, производными платины, заменителями мочевины, адренокортикотропными стероидами, цитокинами, интерлейкинами, АЗТ, ddI, ddc, другими интерферонами, другими антивирусными или противоопухолевыми агентами или антителами.
Режим приема и дозировка эффективного количества ИФН α- n3a, применяемого для лечения описанных выше заболеваний, будет определяться наблюдающим врачом при учете различных факторов, которые изменяют действие лекарств, например, таких как природа заболевания, вес тела, пол, питание больного, тяжесть опухоли, продолжительность приема препарата и другие клинические факторы. Доза заявленной композиции, используемой для лечения специфических болезненных состояний, описанных выше, может варьировать в зависимости от характера болезни и ее стадии. Например, при бородавках ожидаемая доза композиции составляет 0,25 млн МЕд/бородавку ИФН α- n3 или менее того. Для лечения СПИДа или других заболеваний, требующих более жесткой терапии, подходящая доза может быть существенно меньше чем 30 - 36 МЕд, что установлено для принятого к использованию рекомбинантного α- интерферона. Более того, ожидаемая доза интерферона α- n3 для лечения таких состояний составляет примерно 1 - 15 млн. Ед.
В целом, препарат можно применять 3 раза в неделю. Другие дозы и схемы приема препарата могут быть определены одним из известных методов специалистом в данной области исследований.
Помимо терапии описанных выше заболеваний человека указанные методы и композиции согласно настоящему изобретению могут быть использованы для ветеринарных целей, в частности для лечения рака, иммунных заболеваний или вирусных инфекций лошадей, свиней, рогатого скота, собак, кошек и домашней птицы. Для лечения этих заболеваний могут быть использованы те же самые дозы интерферона, которые были указаны ранее, когда речь шла о терапии заболеваний человека.
Следующие примеры приведены лишь в целях иллюстрации и ни в каком аспекте не ограничивают заявленного изобретения.
Пример 1. Метод получения ИФН α- n3a, стадия индукции.
Для получения лейкоцитарной пленки используют кровь из специальных центров (утвержденных FDA), содержащую 500 единиц ЛПК человека. Эритроциты лизируют с помощью хлорида аммония (0,83%) согласно методике Кэнтелла с соавт. (Cantell et al., 1981, см. выше).
Обработанную хлоридом аммония лейкоцитарную пленку ресуспендируют в IX среде Игла МЕМ, содержащей соли Игла (Gibco 410 - 1500), L-глутамин, не необходимые аминокислоты, 4,46 мг/мл Трицина (Aldtich), pH 7,4, 2,2 мг/мл бикарбоната натрия (Fisher), 24 мкг/мл неомицинсульфата и 0,4 мг/мл сыворотки крови человека без γ- глобулинов (NABI).
Лейкоциты (107 кл/мл) суспендируют в среде 1 ч МЕМ и в качестве праймера добавляют 20 Ед/мл сырого α- интерферона. Сырой α- интерферон - это продукт, получаемый в результате процесса индукции, как здесь описано, но не подвергаемый очистке и обработке HCl до pH 2 в течение пяти дней с целью инактивации каких-либо посторонних агентов или потенциальных вирусных контаминантов.
Полученную суспензию инкубируют в 6-литровых стерильных стеклянных флаконах. Лейкоциты праймируют в течение 3 ч при 36oC, а затем добавляют вирус Сендай (штамм Кэнтелла, полученный из SPA FAS Storr, CT) до конечной концентрации 150 ГА Ед/мл.
После одного часа инкубации для прикрепления вируса лейкоциты разводят до 4 • 106 кл/мл (в 2,5 раза) той же средой, содержащей сыворотку крови человека без γ- глобулинов, бикарбонат натрия, но не содержащей праймера. Затем культуры инкубируют 15 ч при 36oC, удаляют клетки и дебрис центрифугированием при 2 500 об/мин (центрифуга Beckman модель J6-B) и определяют титр интерферона с помощью ИРМА или метода ЦПД, как указано выше.
Пример 2. Метод получения ИФН α- n3a, стадия очистки.
Все процедуры очистки проводят при 2 - 8oC, если не указано другой.
А. Сбор/концентрирование сырого интерферона.
После инкубации в течение 15 - 20 ч лейкоциты удаляют из индукционных культур, полученных как описано в примере 1, центрифугированием примерно при 2,900 g в течение 15 - 20 мин. Супернатант (сырой раствор интерферона) сохраняют, а лейкоциты отбрасывают. Если это не осуществляют немедленно, то супернатант хранят в стерилизованных контейнерах при 4oC.
Сырой раствор интерферона концентрируют в 50 раз, используя фильтрующую систему с тангенциальным потоком, через которую проходят соединения с мол. массой 10000 Да и менее. Концентрированный интерферон центрифугируют примерно при 9000 g около 30 мин и затем хранят при -70oC.
Б. Аффинная очистка.
1. Приготовление хроматографической колонки.
Данный метод очистки предусматривает использование моноклональных антител, специфичных к α- интерферонам человека, например антител N К2, производимых фирмой Celltech Limited (Slough, Englan). Моноклональные антитела соединяют с активированной CNBr Сефарозой 4B (например Reselute NK2) и хранят при 4oC. Размер аффинной колонки зависит от связывающей емкости аффинного геля для интерферона, которую определяют для каждого препарата. Аффинные колонки готовят, наполняя соответствующим количеством сефарозного геля, связанного с антителами, подходящие стеклянные колонки. Колонку с моноклональными антителами промывают примерно пятью объемами (имеется в виду объем колонки) фосфатного буферного раствора и затем тремя объемами раствора, содержащего 0,1 М лимонной кислоты и 0,3 М хлорида натрия при pH 2. После этого pH колонки нейтрализуют до 7,4, промывая ее тремя объемами фосфатного буферного раствора (PBS). Описанный цикл может быть повторен еще раза два.
2. Приготовление концентрированного сырого ИФН.
Концентрированный сырой интерферон очищают центрифугированием примерно при 17700 g в течение 60 мин и затем фильтруют через подходящий фильтровальный патрон с размерами пор 0,22 или 0,45 мкм до нанесения на колонку с моноклональными антителами.
Аффинная хроматография.
Примерно 150 млн единиц сырого отфильтрованного интерферона наносят на 1 мл аффинного геля. Количество единиц интерферона определяют путем ИРМА (Celltech Ltd.). Колонку промывают примерно 1,5 объемами фосфатного буферного раствора, а затем 10 объемами 20 мМ фосфатного буфера (pH 7,4), содержащего 50% (по объему) этиленгликоля и 1,5 М хлорида натрия. Окончательно колонку промывают 10 - 30 объемами фосфатного буферного раствора. Интерферон элюируют с аффинной колонки раствором, содержащем 0,1 М лимонной кислоты и 0,3 М хлорида натрия (pH 2).
Восстановление аффинной колонки.
Колонку с моноклональными антителами отмывают 3 - 5 объемами фосфатного буфферного раствора пока pH элюата не станет нейтральным. Для сохранения колонки ее промывают 3 - 5 объемами фосфатного буферного раствора, содержащего 0,1% азида натрия.
В. Кислотная инкубация и нейтрализация.
Элюируемый с аффинной колонки раствор интерферона (pH ~2) инкубируют при 4oC минимум в течение 5 дней. Эта стадия кислотной инкубации необходима для инактивации любых возможных посторонних агентов, таких как ВИЧ-1. После пяти дней инкубации pH раствора доводят до 7,4 - ,0 Трис-HCCl (гидроксиметиламинометан). Интерфероновые белки в растворе сконцентрированы примерно до 1 - 3 мг/мл.
Г. Гель-фильтрация.
Для гель-фильтрации используют препаративную гранулированную сефарозу 12 (Pharmacia, Piscataway, NJ). Концентрированный интерферон в количестве 5% от объема колонки наносят на колонку и проводят элюцию фосфатным буферным раствором. Все фракции, содержащие основной пик с интерфероном, отбирают в стерильных условиях. Очищенный интерферон пропускают через фильтр с размером пор 0,2 мкм или менее и хранят при -70oC.
Д. Результаты.
Результаты обычной процедуры очистки показаны в табл. 2, где приведены данные об этапах очистки, общем объеме и массе белков, общей активности, которую определяют как число единиц интерферона, снятых с колонки, умноженное на 109, выходе интерферона в процентах (общее число единиц ИРМА, возвращаемое на каждой стадии), специфической активности, измеряемой в миллионах единиц на 1 мг, и степени очистки. Активность интерферона определяют с помощью иммунорадиометрического анализа (Celltech, Ltd), в котором в качестве детектирующих антител используют радиоактивно-меченные антитела N К2.
Пример 3. Обратно фазовая жидкостная хроматография высокого разрешения (RP-HPLC).
В этом примере показывается, как различные формы α- интерферона в композиции разделяются в соответствии с их относительной гидрофобностью с помощью RP-HPLC [Janssen et al., J. Chromatographic. Sci., 22: 234 (1984), Stone et al., J. Chrom., 359; 203 (1986)]. Разделение достигается постепенным увеличением концентрации ацетонитрила. Менее гидрофобные молекулы интерферона элюируются в ранних пиках, а более гидрофобные молекулы - позднее.
Типичный профиль RP-HPLC-элюции получают с полупрепаративной колонки (см. фиг. 4), как описано ниже. Около 1-2 мг очищенного интерферона α -n3a фракционируют на полупрепаративной обратно-фазовой HPLC-колонке Vydac C4 (10 • 250 мм). Градиент элюции, используемый для полупрепаративных RP-HPLC-колонок C4 приведен в табл. 3. Градиент линейный и создается следующими буферами: А : 90% H2O/10% ACN/0,1% TFA (объем/объем/вес Б : 90% ACN/10% H2O/0,1% TFA (объем/объем/вес).
Очищенный интерферон фракционируют в виде семи пиков: 1a, 1b, 2, 3, 4, 5 и 6. Первый пик подразделяется на два частично перекрывающихся пика, т.е. 1a и 1b. Эти два пика во всех анализах характеризуют отдельно. Белки каждого пика выделяют отдельно.
В табл. 4 приведены относительные пропорции каждого пика в типичном RP-HPLC-профиле элюции.
Как показано выше, пики 2 и 4 содержат небольшое количество белка. Пик 2 в дальнейшем не был исследован на аминокислотный состав и содержание углеводов. Пик 4 в дальнейшем не был охарактеризован на содержание аминосахаров с помощью HCCl-гилролиза.
После лиофилизации, интерферон в каждом пике был восстановлен 25 мМ Трис-HCCl-буфером при 7,0. Восстановленный материал затем собирают со всех колонок для дальнейшего исследования.
Пример 4. Биологические исследования.
Антивирусная активность.
Исследование антивирусной активности осуществляют при использовании трех различных клеточных линий: 1) линии НЕр-2 клеток человека [АТСС CCL 23], 2) клеток быка MDBK [АТСС CRL 6071] и 3) клеток кролика RK-13 [АТСС CCL 37]. Интерферон последовательно разводят в два раза в 96-луночных платах, затем добавляют клетки в количестве 30000 кл/лунку. После инкубации в течение ночи клетки инфицируют вирусом визикулярного стоматита (BBC, штамм Индиана АТСС # VR-158), затем платы инкубируют в течение ночи еще раз. Цитопатическое действие (ЦПД) определяют с помощью микроскопа на фоне контроля вируса, контроля клеток и клеток, обработанных препаратом стандартного интерферона. Клетки окрашивают кристаллическим фиолетовым, когда в лунках, содержащих препарат стандартного интерферона, обнаруживается выраженное ЦПД. Для всех образцов 50%-ное ЦПД определяют визуально, а титр интерферона вычисляют в сравнении с лабораторными стандартами, которые были предварительно стандартизированы против референс-интерферона, рекомендованного Национальным институтом здравоохранения (Ga 23-902-530). Результаты исследований приведены в табл. 5.
Б. Исследования по ингибированию пролиферации.
Исследования по ингибированию пролиферации проводили на лимфобластоидных клетках человека линии Daudi. В 96-луночных платах готовили последовательные пятикратные разведения интерферонов (100 мкл на лунку) и затем к ним добавляли 104 клеток линии Daudi на лунку (в объеме 100 мкл). После 40-часовой инкубации клетки обрабатывали 1,5 мкК/лунку (в объеме 25 мкл) 3H-тимидина в течение 7 ч. Поглощение тимидина определяли, собирая и отмывая клетки водой на фильтрах из стеклянного волокна и затем снимая показания поглощенной радиоактивности с помощью сцинцилляторного счетчика. Титры интерферона рассчитывали и корректировали в соответствии с лабораторными стандартами.
В. Результаты исследований.
Антивирусная и антипролиферативная активность нефракционированного ИФН α n3a и в составе RP-HPLC-пиков приведены в табл. 5.
Специфическую биологическую активность определяют как число биологических единиц интерферона на 1 мг общей белковой фракции. Результаты, приведенные в табл. 5, показывают, что специфическая ЦПД-активность на человеческих клетках HEp-2 и клетках быка МДВК в каждом пике примерно одинаково. Специфическая антипролиферативная активность на клетках Daudi примерно в 2 раза меньше, чем антивирусная активность в каждом пике. При исследовании ЦДП-активности на клетках почки кролика (RK-13) обнаружено очень незначительное биологическое действие. Специфическая активность на клетках RK-13 по крайней мере в 100-1000 раз меньше, чем на клетках человека или быка. Интересно, что интерферон в составе третьего пика обладает наибольшей специфической активностью на клетках RK-13.
Кроме того, сведения, приведенные в табл.5, показывают, что интерферон в составе первых пиков элюции, таких как 1a и 1b, обнаруживает более низкую специфическую активность на клетках HEp-2, МДВК и Daudi. В данных исследованиях в пиках 1a и 1b выявляется только оокло 25%-ной всей тотальной антивирусной активности композиции ИФН α- n3a. Последний шестой пик элюции обнаруживает наиболее высокую специфическую активность. Действительно, при построении графика зависимости специфической активности интерферона от процентного содержания ацетонитрила в градиенте элюции (фиг. 5) обнаруживается прямая связь между увеличением специфической активности и относительным увеличением гидрофобности α- интерфероновых белков. Чем большей гидрофобностью обладает интерферон, тем большую специфическую активность он проявляет. Можно предположить, что более гидрофобные подтипы интерферона способны связываться и интерфероновыми рецепторами с более высокой аффинностью или инициировать клеточные процессы более эффективно и вследствие этого обладать более высокой специфической активностью.
Пример 5. Физические свойства ИФН α- n3a.
Интерфероновые белки в составе семи пиков, фракционированных методом обратно-фазовой жидкостной хроматографии высокого разрешения (RP-HPLC), характеризуют с помощью SDS-PAGE.
A. Одномерный SDS-PAGE.
1. Методы.
Одномерный SDS-PAGE осуществляют по методике, сходной с той, что описана Лэммли [Laemmli, Nature, 277 : 680 (1970)] . ИФН α- n3a анализируют в 14,5%-ном полиакриламидном геле как в восстанавливающих, так и в невосстанавливающих условиях. Полосы белка выявляют визуально путем окрашивания Кумасса синим. Вестерн-блоттинг дубликатов SDS - геля окрашивают с помощью мышиных моноклональных антител LIT-1, специфичных по отношению к α- ИФН человека и исследуют, как описано Towbin et al., Proc. Natl. Sci (USA) 76: 4350 (1979) и Haid et al., Meth. Enzymol, 96 : 192 (1983).
2. Результаты.
Данные по анализу профилей элюции с помощью SDS-PAGE в невосстанавливающих и восстанавливающих условиях суммированы в табл. 6 и 7 соответственно. Кроме того, на фиг. 6 и 7 показаны соответствующие SDS-PAGE - профили, полученные в невосстанавливающих и восстанавливающих условиях, окрашенные Кумасси синим.
Данные, приведенные в табл. 6 и 7, показывают гетерогенность (наличие более одной белковой полосы) большинства пиков. При использовании описанного метода исследований только в восстанавливающих условиях пик 6, по-видимому, содержит одну белковую полосу. Пики 1a и 1b содержат белки сходных мол. масс при исследовании как в восстанавливающих, так и в не восстанавливающих условиях. Кроме того, эти два пика, 1a и 1b, содержат один и тот же основной тип интерферона, ИФН α- 2b, как показывает частичное аминокислотное секвенирование.
Относительное количество полос интерфероновых белков в каждом пике определяют с помощью лазерной денситометрии. Относительную мол. массу рассчитывают в сравнении с маркерами мол. массы.
Результаты Вестерн-блоттинга приведены на фиг. 8 и 9 для невосстанавливающих и восстанавливающих условий соответственно. Полученные данные свидетельствуют о том, что каждая белковая полоса, выявляемая Кумасси синим, распознается моноклональными антителами LIT-1. Это показывает, что все белковые полосы в каждом пике идентифицируются как α- интерфероны человека. Уровень загрязнений нефракционированного интерферона составляет около 1%. После фракционирования с помощью RP - HPLC ни в одном из пиков этот низкий уровень загрязненности не обнаруживается.
Свойства интерферона продолжают исследовать с помощью двумерного анализа в геле.
Б. Двумерный SDS-PAGE.
1. Методы.
Двумерный гель-электрофорез включает в себя изоэлектрическое фокусирование в первом направлении и SDS-PAGE во втором направлении. В первом исследовании 50 - 10 мкг интерферона из каждого RP - HPLC-пика наносят на гель, состоящий из акриламида и мочевины и залитый в стеклянные трубки (2 • 180 мм). Затем белки разгоняют при постоянном напряжении 400 В в течение 16 ч, а после при 800 В в течение 1 ч. Для изоэлектрофокусирования используют амфолины (Millipore) с диапазоном pH 3 - 10. Гель из трубок выдавливают, уравновешивают в буфере для образцов, содержащем SDS, и наносят на пластину 15%-ного SDS-полиакриламидного геля для анализа во втором направлении. SDS-PAGE осуществляют в трис-глициновом буфере при pH 8,3 и постоянной силе тока 35  Белки в гелях, исследованных с помощью двумерного SDS-PAGE, выявляют визуально путем окрашивания серебром. Интерфероновые белки идентифицируют иммуноокрашиванием с помощью моноклональных антител LIT-1.
Белки в гелях, исследованных с помощью двумерного SDS-PAGE, выявляют визуально путем окрашивания серебром. Интерфероновые белки идентифицируют иммуноокрашиванием с помощью моноклональных антител LIT-1.
2. Результаты.
Результаты двумерного гель-электрофореза нефракционированного ИФН α- n3a и белков каждого RP-HPLC C-пика показывают, что во всех образцах обнаруживаются многочисленные "пятна". В образцах нефракционированного ИФН выявляется примерно 30 пятен. Все пятна распознаются моноклональными антителами LIT-1. Это свидетельствуют о том, что эти пятна представляют собой интерфероновые белки. Число пятен и мол. масса белков в каждом пике суммированы в табл. 8. Изоэлектрические точки всех белков ИФН α- n3a находятся в интервале 5,0 - 7,5.
Результаты исследований, полученные с помощью двумерного SDS-PAGE для пиков 1a и 1b, очень схожи. Обнаружены незначительные различия в размере молекул и заряде. Пики могут быть элюированы по отдельности с помощью RP-HPLC благодаря некоторым, не связанным с зарядом модификациям белков. Результаты исследований остальных пиков с помощью двумерного SDS-PAGE существенно различаются между собой. Профиль каждого пика имеет свои индивидуальные особенности. В каждом пике обнаруживаются многочисленные пятна, свидетельствуя о гетерогенности популяции белков. Эта гетерогенность может быть обусловлена различиями в трансляции и/или пост-трансляционными модификациями белков.
Пример 6. Биохимические свойства ИФН α- n3a.
A. Аминокислотный состав.
Интерфероновые белки гидролизуют с помощью 6н. HCCl при 104oC в течение 24 ч в нагревающем устройстве Pico-Tag. Гидролизаты высушивают под вакуумом и обрабатывают 10%-ным фенилизотиоцианатом (ФИТЦ) в растворе, содержащем 70% этанола, 10% воды и 10% триэтиламина. Полученные в результате описанной процедуры ФТЦ-аминокислоты разделяют путем жидкостной хроматографии высокого разрешения на клоне Pico-Tag C18. Элюцию ФТЦ-аминокислот осуществляют при использовании 0-60%-ного линейного градиента ацетонитрила в воде, содержащей 140 мМ ацетата натрия (pH 6,4) и 0,05% триэтиламина. Поглощение ФТЦ-аминокислот измеряют при 269 нм. Данные поглощения, полученные в каждом исследовании, выражают в числовой форме и хранят в микрокомпьютере. Цистеиновый и триптофановый аминокислотные остатки не модифицируются ФИТЦ в данной реакции и поэтому не дают сигнала поглощения. Данные, заложенные в компьютере, затем анализируют на состав и количественное содержание аминокислот, используя программное обеспечение фирмы Water's Expert.
Результаты исследований аминокислотного состава α- интерферона и отдельных RP-HPLC-пиков представлены в табл. 9. Данные таблицы показывают, что в целом аминокислотный состав индивидуальных пиков сходен с таковым нефракционированного интерферона и теми типами интерферона, которые распознаются моноклональными антителами NK2. При сравнении состава каждого пика с теоретическим составом специфических подтипов α- интерферона, идентифицированных с помощью N-терминального секвенирования, выявлялось совпадение в пределах одной аминокислоты. Эти данные строго подтверждают идентичность основных подтипов интерферона, выявляемых N-терминальным секвенированием.
Б. N-терминальное аминокислотное секвенирование.
Процедуру N-терминального аминоксилотного секвенирования осуществляют методами, сходными с известными [Edman, Acta. Chem. Scant. 4: 283 (1950); Edman et al., Euc. J. Biochem., 1: 80 (1967)].
Примерно 500 пмоль белка каждого пика наносят на прециклизованный фильтр и секвенируют в течение 30-35 циклов. Процедуру проводят при использовании секвенатора Ahhlied Biosystems ABI - 470A, оснащенного последовательным фенилгидантионным аминокислотным анализатором. Методика севенирования основана на процессе деградации, разработанном Эдманом.
N-терминальное секвенирование по Эдману высокоэффективно в целях идентификации и количественного определения остатков аминокислот в относительно коротких пептидных последовательностях (то есть, менее 20). Однако для длинных последовательностей (более 30 аминокислот) идентификация аминокислотных остатков, близких с C-концу белка или полипептида, становится недостоверной. Такое увеличение неточности анализа обусловлено повторным выходом, который намного меньше 95% и поэтому приводит к кумулятивной потере сигнала в каждом дополнительном цикле.
По этой причине белки в пиках 1a и 1b расщепляют гидролизом по метиониновому остатку с помощью CNBr для подтверждения их принадлежности к подтипам ИФН- α 2. Расщепление осуществляют путем растворения белков ИФН- α 2 в 70%-ной муравьиной кислоте и последующего добавления CNBr до концентрации 1M, и затем инкубируя смесь при 4oC в течение 24 ч. Реакцию расщепления останавливают скоростной лиофилизацией под вакуумом, что удаляет из реакционной пробирки все растворители, включая CNBr. Высушенные CNBr-фрагменты белков растворяют в 10%-ной трифторуксусной кислоте (TFA) и наносят на колонку C18 (0,2 • 25 см, Phenomenex). Элюцию фрагментов осуществляют в многоступенчатом градиенте 0,1% TFA/H2O (растворитель А) и 0,1% TFA/ацетонитрил (раствор Б) при скорости потока 0,2 мл/мин. Многоступенчатые линейные градиенты используют, как описано ниже: 1) 0-40%-ный растворитель Б в течение 60 мин; 2) 40-60%-ный растворитель Б в течение 50 мин; 3) 60-100%-ный растворитель Б в течение 13 мин и 4) постоянно 100%-ный растворитель Б в течение дополнительных 5 мин. Элюируемые фрагменты выявляют при постоянном наблюдении величины поглощения при 214 нм. Распавшиеся фрагменты секвенируют, как описано выше.
Расщепление приводит к образованию шести фрагментов, которые распадаются при осуществлении RP - HPLC. Пик, элюируемый в интервале 75-80 мин, секвенируют и в нем обнаруживаются две последовательности. Одна последовательность может быть линеаризована с CNBr-фрагментом NK2, состоящим из аминокислот в положениях 22-59 ИФН α- 2b, в то время как вторая последовательность может быть линеаризована с CNBr-фрагментом N 5, состоящим из аминокислот в положениях 112-148 ИФН- α 2b.
3. Результаты.
N-терминальные аминокислотные последовательности нефракционированного ИФН α- n3a и фракционированных RP-HPLC-пиков представлены в табл. 10. Результаты исследований показывают, что нефракционированный α- ИФН так же, как индивидуальные пики, содержит многочисленные последовательности. Все они могут быть идентифицированы как определенные типы α- интерфероновых белков человека. Основные последовательности каждого пика показаны в табл. 11.
Как свидетельствуют результаты N-терминального секвенирования, обнаружено 18 потенциально возможных подтипов ИФН α- n3a. Выявленные последовательности соответствуют подтипам интерферона, распознаваемым моноклональными антителами NK2, как показано фирмой Celltech, Ltd. ИФН- α 2b и α 8 (a/b/c) - это основные подтипы, выявляемые в ИФН α- n3a. ИФН - α 2b преимущественно обнаруживается в пиках 1a и 1b и в небольших количествах - в пике 2 ИФН α- 2c в следовых количествах выявляется в пике 1a. ИФН - α 8 (a/b/c) преимущественно обнаруживается в пиках 4 и 6.
Другие подтипы интерферона, которые присутствуют в меньших количествах, - это интерфероны α 4 (a/b) и/или α 16, выявляемые в пике 2. Подтип α 10a обнаруживается в пике 3, α 16 в следовых количествах - в пике 2. Подтипы α 17 (a/b/c) и α 21 (a/b) преимущественно обнаруживаются в пике 5 и в следовых количествах - в пике 4. Интерфероны α7 (a/b/c) преимущественно выявляются в пике 5. Поскольку аминокислотный анализ осуществляется в течение 30-45 циклов, трудно различить некоторые подтипы интерферона, которые в этих 30-35 циклах имеют идентичные последовательности. Опубликованные данные по N-терминальному секвенированию всех подтипов α- ИФН человека показывают, что ИФН - α 21 (a/b) и α 17 (a/b/c) имеют сходные N-терминальные последовательности до аминокислоты в положении 33), α 4 (a/b) и α 16 - тоже до аминокислоты в положении 33, а ИФН-α/ 2b сравним с α 2c. Таким образом, некоторые минорные подтипы ИФН не могли быть выявлены из-за ограничений аминокислотного анализа.
Небольшие различия, которые обнаруживаются в аминокислотных последовательностях индивидуальных интерфероновых белков, могут быть обусловлены различиями белковых молекул подтипов α- ИФН или аллельностью генов. Последнее может привести к появлению сходных белков с аминокислотными модификациями в определенном участке.
Полученный интерфероновый продукт далее анализируют на содержание подтипов ИФН - α 2. Как сообщалось предварительно, известны три подтипа ИФН - α 2, а именно ИФН - α 2a, ИФН α- 2b ИФН - α 2c.
Различия в N-терминальных последовательностях этих трех подтипов ИФН - α 2 выявляются в циклах 23 и 34 и представлены в табл. 12.
Данные по секвенированию интактных белков и CNBr-фрагмента # 2 для пиков 1a и 1b до 35 цикла (табл. 9) показывают, что ИФН -α 2b - это основной компонент, обнаруживаемый в RP - HPLC-пиках 1a и 1b. Аминокислоты, высвобождаемые в циклах 23 и 34, представляют собой аргинин (Arg) и гистидин (His) соответственно, что согласуется с известными данными о последовательности ИФН α- 2b. Не обнаружено заметного высвобождение лизина (Lys) в цикле 23, свидетельствуя о том, что ИФН -α 2a не выявляется в пиках 1a и 1b или в каком-либо другом RP - HPLC-пике ИФН α- n3a.
Полученные результаты верифицируют пептидным картированием и N-терминальным секвенированием белковых последовательностей, высвобождаемых в результате расщепления CNBr. Более того, для выявления ИФН -α 2c CNBr-фрагмент, расположенный между Arg в положении 22 и Met в положении 60, изолируют и расщепляют трипсином. Полученные пептиды разделяют путем RP - HPLC и секвенируют. Количественный анализ последовательностей показывает, что более чем 90% ИФН-α 2 в ИФН α- n3a представляет собой ИФН -α 2b. ИФН -α 2c может содержаться в полученном препарате, однако его концентрация ниже определяемого уровня.
B. Содержание углеводов.
Образец интерферонового белка (150 мкг) диализуют в соотношении 2 log против 0,1 М уксусной кислоты и его объем уменьшается до 300 мкл. Сконцентрированный образец разделяют на три равные аликвоты и упаривают досуха. Полученные образцы затем гидролизуют следующим образом: 1) в 2М трифторуксусной кислоты (TFA) при 100oC в течение 4 ч для определения нейтральных сахаров, 2) в 6 н. HCCl при 100oC в течение 4 ч для определения аминосахаров или 3) в 0,1 н. HCCl при 80oC в течение 1 ч для определения сиаловых кислот. Гидролизаты упаривают досуха, добавляют к воде, которая предназначена для использования в HPLC, и быстро прогревают до 80-100oC для облегчения растворения.
Растворы моносахаридов, а именно углеводные стандарты, содержащие 2,5 мкг/мл глюкозы, галактозы, глюкозамина, галактозамина, фукозы и маннозы, приготавливают на HPLC-воде. Для разделения моносахаридов используют систему Dionex® HPLC, оснащенную анионообменной колонкой CarboPac® HA1. Пульсовой амперометрический детектор уравновешивают 19 мМ NaOH при скорости потока 1 мл/мин и затем постколоночным стабилизатором (post column stabiilzer) 0,5 M NaOH при скорости потока 0,5 мл/мин в течение 2 ч 90 мкл стандарта или образца вводят в систему посредством автоматического инъектора и сахара элюируют в равновесных условиях с помощью 19 мМ NaOH.
Стандартный раствор сиаловых кислот в концентрации 2,5 мкг/мл готовят на HPLC-воде. Сиаловые кислоты элюируют при использовании той же самой системы Dionex® HPLC, как описано выше. Пульсовой амперометрический детектор уравновешивают 250 мМ NaOH. 90 мкл стандарта или образца гидролизата вводят в систему и элюируют в равновесных условиях с помощью 250 мМ NaOH.
Содержание углеводов вычисляют как отношение молей сахара к молям α- интерферонового белка.
Результаты исследований по содержанию углеводов для аминосахаров, нейтральных сахаров и сиаловых кислот приведены в табл. 13.
Для нефракционированного ИФН α- n3a так же, как и для всех индивидуальных пиков, содержание углеводов составляет 1-3 моль сахара на моль интерферона. Пики 1a и 1b содержат наибольшее количество углеводов по сравнению с пиками поздней элюции. Пик 6, иными словами ИФН -α 8 (a/b/c) содержит наименьшее количество углеводов. В поздних пиках с 3 по 6 не определяется содержание маннозы или сиаловых кислот. Это обстоятельство связано с гидрофобностью белков. Низкое содержание углеводов приводит к их более высокой гидрофобности и поэтому такие белки элюируются на поздних этапах RP - HPLC. Для белков второго пика данные не получены из-за низкого содержания в них исследуемого материала.
В другом эксперименте для определения сайта гликозилирования примерно 100 мкг белка пиков 1a и 1b денатурируют, восстанавливают и алкилируют. Затем препарат переваривают с помощью эндопептидазы V8, расщепляющей белки в месте расположения глутаминовой кислоты, при соотношении 1:100 (энзим:субстрат). Полученные V8-фрагменты разделяют с помощью HPLC и собирают фракции, которые затем гидролизуют для высвобождения углеводов и анализируют на их содержание. Основное количество углеводов содержат фракции 52-53. Соответственно эти фракции подвергают N-терминальному секвенированию.
Секвенирование позволяет установить, что на N-терминальном участке пептида ИФН -α 2 приблизительно в положении 97-110 расположена последовательность AXVIQGVGVXETP [SEQ ID N 0:1], где первый X возможно соответствует C, а второй - T. Эту последовательность получают расщеплением ИФН α- 2b по сайту E96 с помощью эндопептидазы V8. Потеря сигнала, соответствующего T в положении 106, в то время как T в положении 108 явным образом обнаруживается, свидетельствует о том, что T в положении 106 может быть модифицирован в результате O-связывания углеводов. Таким образом, треонин 106 идентифицируется как сайт гликозилирования ИФН -α 2 (b/c).
Г. Карбокситерминальные аминокислотные последовательности.
Белки пиков 1a и 1b анализируют методом секвенирования для определения карбокситерминальных последовательностей при использовании карбоксипептидазы P. Карбоксипептидаза P гидролизует белки более быстро и практически не отдает предпочтение какой-либо аминокислоте. Интерференовые белки пиков 2-6 не анализируют из-за низкого содержания исследуемого материала.
Образец белка (10-30 мкг) диализуют против 50 мМ ацетата натрия при pH 5,5. Карбоксипептидазу P добавляют в соотношении энзим:протеин 1:100 (по весу). Полученную смесь инкубируют при 37oC и отбирают аликвоты через 0,5, 1,2, 5, 10 и 60 мин. Каждую процедуру переваривания и анализа повторяют по меньшей мере дважды.
Высвобождаемые на каждом временном промежутке аминокислоты обрабатывают O-фатальдегидом (OPA) в растворе, содержащем 2% SDS/0,4 М бората лития, в течение 30 с. Производные аминокислот разделяют на обратно-фазовой колонке Pico-Tag C18, используя метаноловый градиент против водного буфера, содержащего ацетат натрия и тетрагидрофуран. Идентификацию и количественное определение высвобождаемых аминокислот проводят в сравнении с HPLC-профилем OPA-производных стандартных аминокислот.
Результаты C-терминального секвенирования ИФН α- n3a, RP - HPLC-пиков и подтипов α- ИФН суммированы в табл. 14.
Последовательности идентифицируют на основе порядка появления и расположения отдельных аминокислотных остатков. Например, глутаминовая кислота появляется и располагается раньше, чем другие и, следовательно, она является первым аминокислотным остатком C-конца. В других циклах последовательности выявляются по аналогичному принципу. В случае нефракционированного α- ИФН, основная C-терминальная последовательность определяется как Leu-Arg-(Ser/Arg)-Lys-Glu [SEQ ID NO:2].
Для сравнения теоретически ожидаемые последовательности подтипов α- ИФН тоже представлены в табл. 14. Результаты исследований показывают, что основная C-концевая последовательность для пиков 1a и 1b так же, как и для нефракционированного интерферона, сходна с таковой ИФН -α 2b. Это свидетельствует о том, что нефракционированный интерферон и белки пиков 1a и 1b содержат интактную карбокситерминальную последовательность. Используемый здесь метод C-терминального секвенирования значительно менее чувствителен, чем N-терминальное секвенирование. Какие-либо минимальные различия в последовательностях могут быть и не обнаружены. Например, C-терминальная последовательность ИФН α- 8 (Leu-Lys-Ser-Lys-Glu [SEQ ID NO:3] не выявляется в присутствии основной C-терминальной последовательности ИФН -α 2b (Leu-Arg-Ser-Lys-Glu) [SEQ ID NO:4] при том, что эти две последовательности различаются только в цикле -4. Присутствие минорных интерфероновых компонентов с редуцированными C-концами не выявляется, если содержание этих компонентов составляет менее 20%.
Пример 7. Стадия 1 исследований токсичности.
Первую стадию исследований токсичности ИФН α- n3a проводят на 20 добровольцах с асимптоматической ВИЧ-инфекцией, в крови которых содержание CD4+ T-клеток составляет ≥400/мм3. Композиция ИФН α- n3a до соответствующего дозированного разведения содержит 5 МЕд/мл ИФН α- n3a в 3,3 мг фенола, 1 мг альбумина человека, PBS, pH 7, 8,0 мг/мл NaCl, 1,74 мг/мл двухосновного фосфата натрия, 0,2 мг/мл моноосновного фосфата калия и 0,2 мг/мл хлорида калия.
Эту композицию применяют подкожно 3 раза в неделю (понедельник, среда и пятница) в течение 3-6 месяцев в следующих дозах: 5 человек по 1 млн. МЕд/дозу, 10 человек по 2,5 млн. МЕд/дозу в течение первой недели и затем 5 млн. МЕд/дозу и 5-и больным вводят их максимально толерантную дозу. Больных наблюдают в клинике в течение первой недели проведения лекарственной терапии.
Результаты исследований приведены ниже в табл. 15. ИФН α- n3a обладает хорошей толерантностью при дозах ≤17,5 млн. Ед. По-существу, не наблюдалось лихорадки или простудоподобных симптомов, признаков желудочно-кишечных заболеваний, а также кожных высыпаний. Лабораторные исследования общего количества лейкоцитов, гранулоцитов и тромбоцитов показали результаты, лежащие в пределах нормы, в соответствии с требованиями ВОЗ. У большинства больных обнаруживались дополнительные маркеры α-интерфероновой активности (увеличенная экспрессия антигенов главного комплекса гистосовместимости класса 1 на поверхности лейкоцитов крови, индуцирования α- ИФН экспрессия генов).
Частота возникновения побочных эффектов и аномалии лабораторных исследований, о которых сообщалось в отношении рекомбинантного α- интерферона, не наблюдались для эквивалентных доз заявленной согласно настоящему изобретению композиции. Эта необходимая безопасность и толерогенность в сочетании с убедительным доказательством in vitro антивирусной активности менее очищенного продукта, ранее используемого в клинических исследованиях и описанного как ИФН α- n3 [см. Alferon A, вкладыш, прилагаемый к упаковке, Interferon Sciences, Inc., New Gersey], свидетельствует о возможности использования в терапии заявленной композиции как эффективного и в достаточной степени безопасного антивирусного средства.
Для того, чтобы показать поразительные преимущества заявленной композиции (ее чрезвычайно низкую токсичность) над другими известными α- интерфероновыми композициями, результаты экспериментов по исследованию токсичности в сравнении со многими другими описанными препаратами ИФН -α (природными, рекомбинантными и полученными с помощью клеток) приведены в табл. 16. Для облегчения сравнения ИФН α- n3a сравнивают с несколькими α- ИФН, так же как и с β- и γ- интерферонами, описанными по отдельности в ниже приведенных источниках информации. В табл. 16 номера ссылок соответствуют следующему списку:
1. G.M. Scott et al., British Medical Journal, 282: 1345 (1981).
2. S.L.Sacks et al., Antimicrobial Agents & Chemotherapy, 21: 9 (1982).
3. J.H.Schiller et al., J. Clinical Investigation, 86: 1211 (1990).
4. S. Ingimarsson et al., J. Infection Disease, 140: 560 (1979).
5. H.C.Lane et al., Annals of Internal Medicine, 112: 805 (199).
6. R.Kurzrock et al., J. Clinical Oncology, 4: 1101 (1986).
7. R.J.Spiegel, Clinical Overview of alfa-ИФН, page 627 (1987).
8. T.Taguchi, Cancer, 57: 1705 (1986).
9. R.M.Bukowski et al., Cancer Research, 51: 836 (1991).
10. H.G.Klingemann et al., Blood, 78: 3306 (1991).
11. Roferon® A, PDR 2006 (1993).
12. Intron® A PDR 2194 (1993).
13. R.J. Wells et al., J. Interferon Res., 8: 309 (1988).
14. G.J.Jones et al., Canser, 57: 1709 (1986).
15. Alferon® N Injection, Package Insert.
Пример 8. Активность интерферона α- п3а в отношении ВИЧ-1.
Известно, что интерфероны как in vivo, так и in vitro обладают антивирусной активностью. Типы ИФН α- n3a тестируют на антивирусную активность в серии экспериментов с вирусом иммунодефицита человека типа 1 (ВИЧ-1). Относительную активность ИФН α- n3a сравнивают с активностью двух рекомбинантных форм альфа-интерферона (ИФН α-/ 2a и ИФН α- 2b).
A. Клеточная культура и исследования обратной транскриптазы.
Клетки, используемые в этих исследованиях, представляют собой нормальные моноциты человека, полученные из фракции мононуклеарных периферических клеток, серонегативных в отношении ВИЧ и вируса гепатит B доноров. Клетки получают в результате забора лейкоцитов, очистки и подсчета мононуклеаров из обогащенной лейкоцитами фракции клеток крови с помощью проточной центрифуги, оснащенной элютриатором. В клеточной суспензии обнаруживается более 98% моноцитов, которые культивируют в виде прикрепленного монослоя (1,5 • 105 кл/мл) в среде ДМЕМ (Gibco, Grand Island, N J) с добавлением 10%-ной инактивированной нагреванием сыворотки крови человека, содержащей альбумин, 50 мкг/мл гентамицина и 1000 Ед/мл высокоочищенного рекомбинантного макрофагального колониестимулирующего фактора (M-CSF, FAP-809, Cetus Corp., Emeryville, CA). Во всех компонентах культуральной среды при тестировании не обнаружена контаминация эндотоксинами.
Прикрепившиеся моноциты культивируют в течение 7 дней в отсутствии интерферона до внесения тропного по отношению к этим клеткам ВИЧ-1 штамма АДА. Инфицированные клетки инкубируют до четырех недель в отсутствии или при добавлении интерферона. Культуральную среду наполовину заменяют каждые 2 - 3 дня.
Интерферон, используемый в этих экспериментах, имеет различное происхождение и, следовательно, его предварительно титруют против других вирусов, а именно вируса везикулярного стоматита (BBC), методом ЦПД с помощью клеток MDBK. При этом для описываемых исследований титр интерферона в антивирусных единицах должен соответствовать титру интерферона в единицах на мл культуры.
Когда ИФН добавляют в одно время с вирусом или непосредственно перед ним, обнаруживается дозозависимое ингибирование продукции ВИЧ под влиянием интерферона.
В указанных экспериментах для количественного определения уровня вирусной экспрессии используется обратная транскриптаза ВИЧ-1 из реплицирующихся клеточных культур. Жидкие пробы культуральной среды собирают в различные временные промежутки и затем добавляют в реакционную смесь, состоящую из 0,05%-ного Нонидета P-40 (Sigma Chemical Co), 10 мкл/мл поли(A), 0,25 ЕД/мл одного (dT (Pharmacia, Piscataway, NJ), 5 мМ дитиотреитола (Pharmacia), 150 мМ KCCl, 15 мМ MgCl2 и 3H-dTTP (2 Ки/ммоль Amersham Corp., Arlington) в трис-HCCl-буфере (pH 7,9) и оставляют на 24 ч при 37oC. Поглощенную радиоактивность преципитируют холодной трихлоруксусной кислотой, отмывают и собирают с автоматического харвестера клеток (Skatron Inc., Sterling, VA на стеклянные фильтровальные диски. Уровень радиоактивности измеряют на жидкостном сцинтилляторном счетчике.
Б. Сравнительное действие интерферонов на ВИЧ-1 ADA, которым инфицированы культуры моноцитов.
Дозависимые эффекты ИФН α- n3a исследуют in vitro на ВИЧ-1-инфицированных моноцитах в сравнении с двумя другими известными препаратами, а именно рекомбинантным ИФН α- 2a ( Roferon® A, Hoffman-La Roche, Nutley) и рекомбинантным ИФН α- 2b ( Intron® A, Schering Plough, Kenilworth, NJ). Возрастающие дозы интерферона (0 - 500 Ед/мл) добавляют к моноцитам, инфицированным ВИЧ-1ADA (множественность заражения составляет 0,01) и инкубируют в течение 12 дней. Результаты исследований представлены в табл. 17. Они представляют собой усредненные данные трех экспериментов и показывают, что от дозы к дозе ИФН α- n3a увеличивает свою ингибирующую активность в отношении ВИЧ-1-инфицированных моноцитов по сравнению с двумя другими препаратами рекомбинантного интерферона.
Эти эксперименты повторяли много раз для получения достоверных результатов. Полученные данные суммированы в табл. 18.
Приведенные результаты свидетельствуют о том, что ИФН α- n3a в 10 раз более эффективен, чем ИФН α- 2b, и примерно в 100 раз более эффективен, чем ИФН α- 2a, в данных исследованиях. Это наблюдается даже после приведения относительной концентрации интерферонов каждого препарата к тому же самому титру, который был определен в исследованиях ЦПД, как описано выше.
В. Исследование активности ИФН α- n3a в отношении ВИЧ-1.
В качестве препаратов ИФН α- n3a, обладающих повышенной активностью в отношении инцифированных ВИЧ-1 культур моноцитов, используют фракционированные пики ИФН α- n3a, полученные с помощью обратно-фазовой HPLC. ИФН α- n3a фракционируют на RP-HPLC-колонке, как описано выше, и каждый пик, содержащий активность ИФН, титруют на антивирусную активность (по отношению к BBC) в исследовании ЦПД на клетках MDBK для приведения к тому же самому титру, прежде чем используют в описываемых экспериментах. После стандартизации ЦПД-титра исследуют пики (a + b), 2, 3, 4, 5 и 6. В каждом исследовании используют 100 МЕд/мл ИФН активности при множественности заражения ВИЧ-1 ≤ 0,02. Полученные результаты представлены в табл. 19.
Приведенные результаты показывают, что основная активность в отношении ВИЧ-1 ассоциирована с типами ИФН, которые содержатся в RP-HPLC-пиках 2 и 6, причем пики 3, 4, 5 имеют достаточно сильную но все-таки меньшую активность. Интересно и неожиданно обнаружение того факта, что типы ИФН в пике 1 имеют наименьшую активность в отношении ретровирусов среди всех типов ИФН α- n3a. В этом пике типы интерферона относятся к ИФН α- 2 (b/c). Этим объясняется низкая активность в отношении ВИЧ-1 как ИФН α- 2a, так и ИФН α- 2b, что описано выше.
В то время как здесь подробно описаны некоторые возможные пути осуществления изобретения, специалистам в данной области исследования предельно ясно, что могут существовать многочисленные варианты и модификации предложенных технических решений. Таким образом, предложенное изобретение не ограничивается каким-либо детально описанным вариантом его осуществления и определяется не только приведенной ниже совокупностью существенных признаков.
Описание последовательностей
(1) Основная информация:
(i) Заявитель:
Testa, Douglas
Liao, Mei-June
Ferencz-Biro, Katalin
Rashidbaigi, Abbas
Di Paola, Mario
Padhye, Manisha
(ii) Название изобретения: Улучшенные альфа-интерфероновые композиции и способ их получения с помощью лейкоцитов периферической крови человека
(iii) Количество последовательностей : 22
(iv) Адрес для переписки:
(А) Адресат: Howson and Howson
(Б) Улица: Spring House Corporate Center P.O. Box 457
(В) Город: Спрингхаус
(Г) Штат: Пенсильвания
(Д) Страна: США
(Е) Почтовый код: 19477
(v) Форма для компьютерной обработки:
(А) Носитель информации: гибкий диск
(Б) Компьютер: IBM-совместимый
(В) Операционная система: PC-DOS/MS-DOS
(Г) Программа: Patent In Release 1, версия 1,25
(vi) Текущие данные заявки:
(А) Номер заявки:
(Б) Дата подачи заявки:
(В) Классификация:
(vii) Предшествующие данные заявки:
(А) Номер заявки: VS 07/835,030
(Б) Дата подачи заявки: 10.02.92
(viii) Информация о патентном поверенном:
(А) Имя: Bak, Mary E.
(Б) Регистрационный номер: 31,215
(В) Номер для ссылки: NPO92USA
(ix) Информация о каналах связи:
(A) Телефон: 215-540-9206
(Б) Телефакс: 215-540-5818
(2) Информация о последовательности SEQ ID NO: 1:
(i) Характеристика последовательности:
(А) Длина: 13 аминокислот
(Б) Тип: аминокислотная последовательность
(В) Количество цепей: неизвестно
(Г) Топология: неизвестна
(ii) Молекулярная природа: протеин
(ix) Особенности:
(А) Ключевое слово: участок
(Б) Расположение: 2
(В) Другая информация: (note = эта аминокислота может быть Cys)
(ix) Особенности:
(А) Ключевое слово: участок
(Б) Расположение: 10
(В) Другая информация: (note = эта аминокислота может быть Thr)
(xi) Структура последовательности SEQ ID NO 1:
(2) Информация о последовательности SEQ ID NO 02:
(i) Характеристика последовательности:
(А) Длина: 5 аминокислот
(Б) Тип: аминокислотная последовательность
(В) Количество цепей: неизвестно
(Г) Топология: неизвестна
(ii) Молекулярная природа: протеин
(ix) Особенности:
(А) Ключевое слово: участок
(Б) Расположение: 3
(В) Другая информация: (note = эта аминокислота может быть Arg)
(xi) Структура последовательности SEQ ID NO 2:
(2) Информация о последовательности SEQ ID NO 3:
(i) Характеристика последовательности:
(А) Длина: 5 аминокислот
(Б) Тип: аминокислотная последовательность
(В) Количество цепей: неизвестно
(Г) Топология: линейная
(ii) Молекулярная природа: протеин
(xi) Структура последовательности SEQ ID NO 3:
(2) Информация о последовательности SEQ ID NO 4:
(i) Характеристика последовательности:
(А) Длина: 5 аминокислот
(Б) Тип: аминокислотная последовательность
(В) Количество цепей: неизвестно
(Г) Топология: неизвестна
(ii) Молекулярная природа: протеин
(xi) Структура последовательности SEQ ID NO 4:
(2) Информация о последовательности SEQ ID NO 5:
(i) Характеристика последовательности:
(А) Длина: 34 аминокислоты
(Б) Тип: аминокислотная последовательность
(В) Количество цепей: неизвестно
(Г) Топология: неизвестна
(ii) Молекулярная природа: протеин
(ix) Особенности:
(А) Ключевое слово: участок
(Б) Расположение: 1
(В) Другая информация: (note = эта аминокислота может быть Cys)
(ix) Особенности:
(А) Ключевое слово: участок
(Б) Расположение: 11
(В) Другая информация: (note = эта аминокислота может быть Ser)
(ix) Особенности:
(А) Ключевое слово: участок
(Б) Расположение: 14
(В) Другая информация: (note = эта аминокислота может быть Thr)
(ix) Особенности:
(А) Ключевое слово: участок
(Б) Расположение: 16
(В) Другая информация: (note = эта аминокислота также может быть Met)
(ix) Особенности:
(А) Ключевое слово: участок
(Б) Расположение: 19
(В) Другая информация: (note = эта аминокислота также может быть Gly)
(ix) Особенности:
(А) Ключевое слово: участок
(Б) Расположение: 22
(В) Другая информация: (note = эта аминокислота также может быть Gly)
(ix) Особенности:
(А) Ключевое слово: участок
(Б) Расположение: 27
(В) Другая информация: (note = эта аминокислота также может быть Pro)
(ix) Особенности:
(А) Ключевое слово: участок
(Б) расположение: 29
(В) Другая информация: (note = эта аминокислота также может быть Cys)
(xi) Структура последовательности SEQ ID NO 5:
(2) Информация о последовательности SEQ ID NO6:
(i) Характеристика последовательности:
(А) Длина: 34 аминокислоты
(Б) Тип: аминокислотная последовательность
(В) Количество цепей: неизвестно
(Г) Топология: неизвестна
(ii) Молекулярный тип: протеин
(xi) Структура последовательности SEQ ID NO6:
(2) Информация о последовательности SEQ ID NO 7:
(ii) Характеристика последовательности:
(А) Длина: 34 аминокислоты
(Б) Тип: аминокислотная последовательность
(В) Количество цепей: неизвестно
(Г) Топология: неизвестна
(ii) Молекулярный тип: протеин
(xi) Структура последовательности SEQ ID NO 7: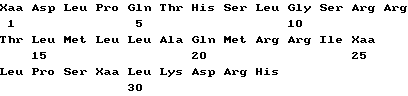
(2) Информация о последовательности SEQ ID NO 8:
(ii) Характеристика последовательности:
(А) Длина: 34 аминокислоты
(В) Тип: аминокислотная последовательность
(В) Количество цепей: неизвестно
(Г) Топология: неизвестна
(ii) Молекулярный тип: протеин
(ix) Особенности:
(А) Ключевое слово: участок
(Б) Расположение: 11
(Е) Другая информация: (note = эта аминокислота может также быть Ser)
(ix) Особенности:
(А) Ключевое слово: участок
(Б) Расположение: 22
(В) Другая информация: (note = эта аминокислота может также быть Arg)
(xi) Структура последовательности SEQ ID NO 8: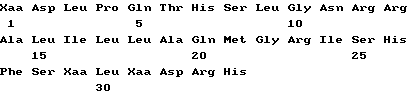
(2) Информация о последовательности SEQ ID NO 9:
(i) Характеристика последовательности:
(А) Длина: 34 аминокислоты
(Б) Тип: аминокислотная последовательность
(В) Количество цепей: неизвестно
(Г) Топология: неизвестна
(ii) Молекулярный тип: протеин
(xi) Структура последовательности SEQ ID NO 9: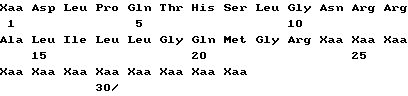
(2) Информация о последовательности SEQ ID NO 10:
(i) Характеристика последовательности:
(А) Длина: 34 аминокислоты
(Б) Тип: аминокислотная последовательность
(В) Количество цепей: неизвестно
(Г) Топология: неизвестна
(ii) Молекулярный тип: протеин
(xi) Особенности:
(А) Ключевое слово: участок
(Б) Расположение: 22
(В) Другая информация: (note = эта аминокислота может быть Gly
(xi) Структура последовательности SEQ ID NO 10: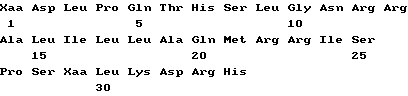
(2) Информация о последовательности SEQ ID NO 11:
(i) Характеристика последовательности:
(А) Длина: 34 аминокислоты
(Б) Тип: аминокислотная последовательность
(В) Количество цепей: неизвестно
(Г) Топология: неизвестна
(ix) Особенности:
(А) Ключевое слово: участок
(Б) Расположение: 10
(В) Другая информация: (note = эта аминокислота может также быть Arg
(xi) Структура последовательности SEQ ID NO 11:
(2) Информация о последовательности SEQ ID NO 12:
(i) Характеристика последовательности:
(А) Длина: 34 аминокислоты
(Б) Тип: аминокислотная последовательность
(В) Количество цепей: неизвестно
(Г) Топология: неизвестна
(ii) Молекулярный тип: протеин
(xi) Структура последовательности SEQ ID NO 12:
(2) Информация о последовательности SEQ ID NO 13:
(i) Характеристика последовательности:
(А) Длина: 5 аминокислот
(Б) Тип: аминокислотная последовательность
(В) Количество цепей: неизвестно
(Г) Топология: неизвестна
(ii) Молекулярный тип: протеин
(xi) Структура последовательности SEQ ID NO 13:
(А) Информация о последовательности SEQ ID NO 14:
(i) Характеристика последовательности:
(А) Длина: 5 аминокислот
(Б) Тип: аминокислотная последовательность
(В) Количество цепей: неизвестно
(Г) Топология: неизвестна
(ii) Молекулярный тип: протеин
(xi) Структура последовательности SEQ ID NO 14:
(2) Информация о последовательности SEQ ID NO 15:
(i) Характеристика последовательности:
(А) Длина: 5 аминокислот
(Б) Тип: аминокислотная последовательность
(В) Количество цепей: неизвестно
(Г) Топология: неизвестна
(ii) Молекулярный тип: протеин
(xi) Структура последовательности SEQ ID NO 15:
(2) Информация о последовательности SEQ ID NO 16:
(i) Характеристика последовательности:
(А) Длина: 5 аминокислот
(Б) Тип: аминокислотная последовательность
(В) Количество цепей: неизвестно
(Г) Топология: неизвестна
(ii) Молекулярный тип: протеин
(xi) Структура последовательности SEQ ID NO 16:
(2) Информация о последовательности SEQ ID NO 17:
(i) Характеристика последовательности:
(А) Длина: 5 аминокислот
(Б) Тип: аминокислотная последовательность
(В) Количество цепей: неизвестно
(Г) Топология: неизвестна
(ii) Молекулярный тип: протеин
(xi) Структура последовательности SEQ ID NO 17:
(2) Информация о последовательности SEQ ID NO 18:
(i) Характеристика последовательности:
(А) Длина: 5 аминокислот
(Б) Тип: аминокислотная последовательность
(В) Количество цепей: неизвестно
(Г) Топология: неизвестна
(ii) Молекулярный тип: протеин
(xi) Структура последовательности SEQ ID NO 18:
(2) Информация о последовательности SEQ ID NO 19:
(i) Характеристика последовательности:
(А) Длина: 5 аминокислот
(Б) Тип: аминокислотная последовательность
(В) Количество цепей: неизвестно
(Г) Топология: неизвестна
(ii) Молекулярный тип: протеин
(xi) Структура последовательности SEQ ID NO 19:
(2) Информация о последовательности SEQ ID NO 20:
(i) Характеристика последовательности:
(А) Длина: 5 аминокислот
(Б) Тип: аминокислотная последовательность
(В) Количество цепей: неизвестно
(Г) Топология: неизвестна
(ii) Молекулярный тип: протеин
(ix) Особенности:
(А) Ключевое слово: участок
(Б) Расположение: 3
(В) Другая информация: (note = эта аминокислота также может быть Arg)
(xi) Структура последовательности SEQ ID NO 20:
(2) Информация о последовательности SEQ ID NO 21:
(i) Характеристика последовательности:
(А) Длина: 5 аминокислот
(Б) Тип: аминокислотная последовательность
(В) Количество цепей: неизвестно
(Г) Топология: неизвестна
(ii) Молекулярный тип: протеин
(xi) Структура последовательности SEQ ID NO 21:
(2) Информация о последовательности SEQ ID NO 22:
(i) Характеристика последовательности:
(А) Длина: 5 аминокислот
(Б) Тип: аминокислотная последовательность
(В) Количество цепей: неизвестно
(Г) Топология: неизвестна
(ii) Молекулярный тип: протеин
(xi) Структура последовательности SEQ ID NO 22: (
(
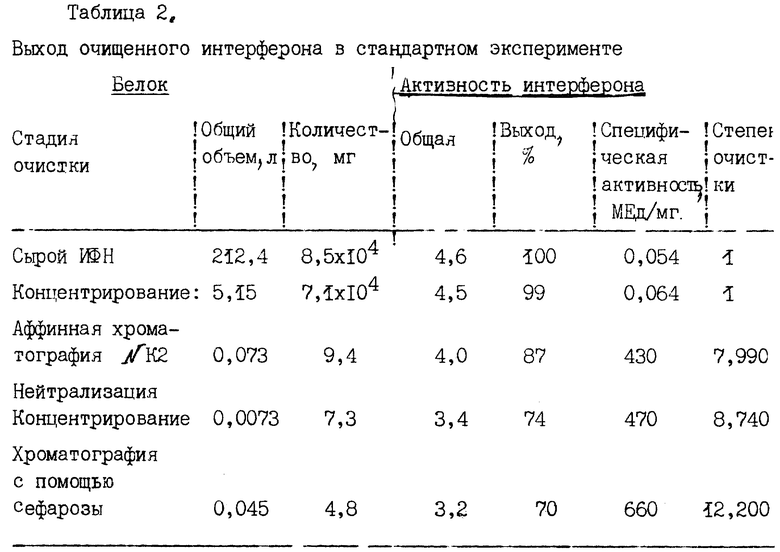
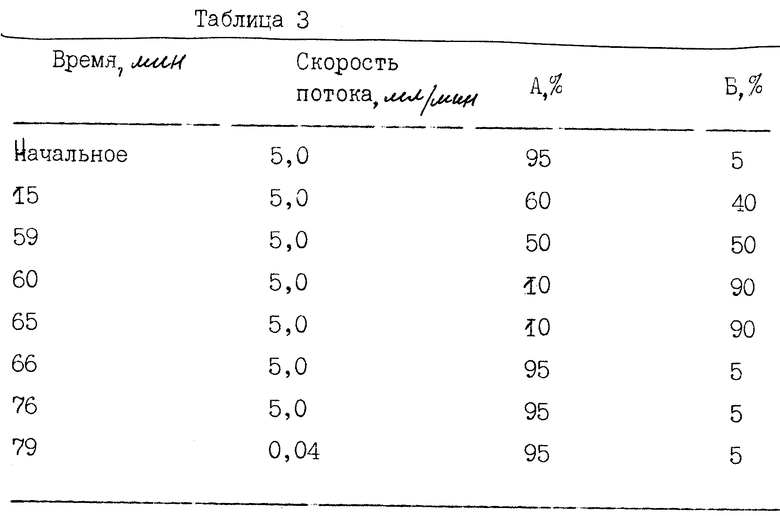

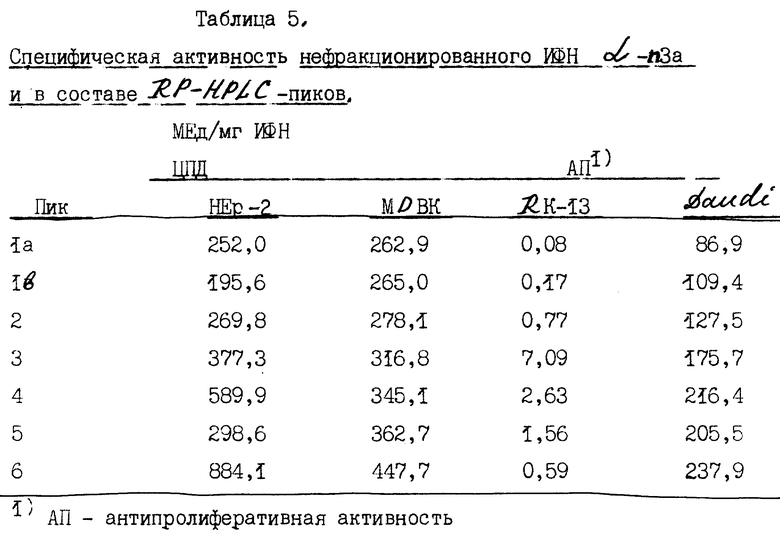
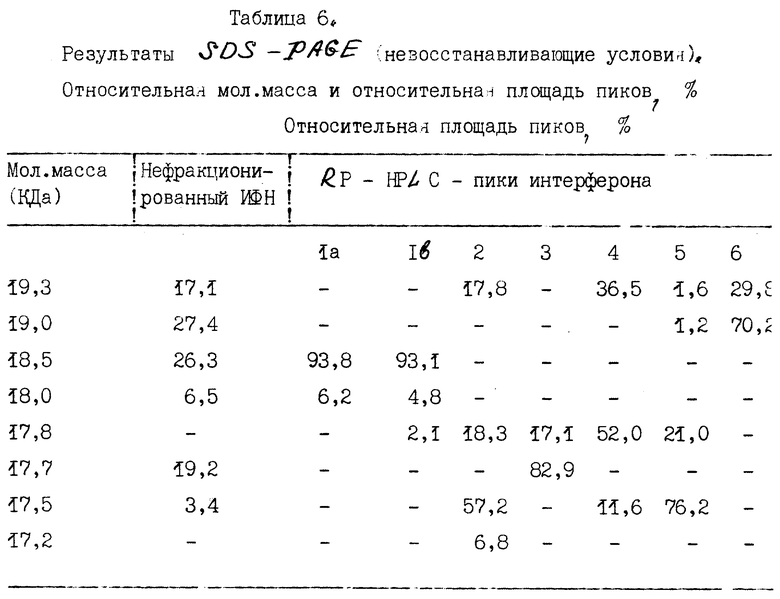
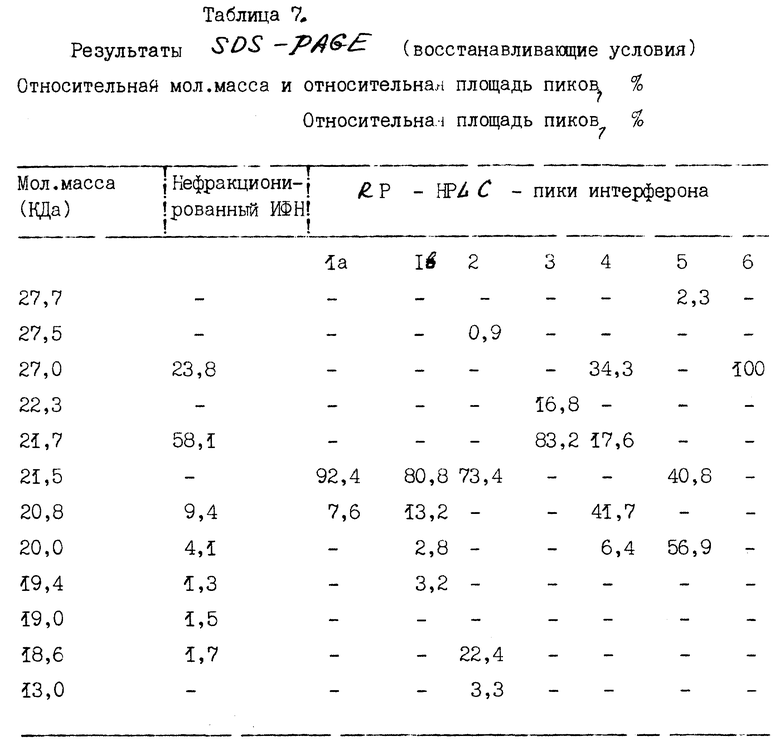

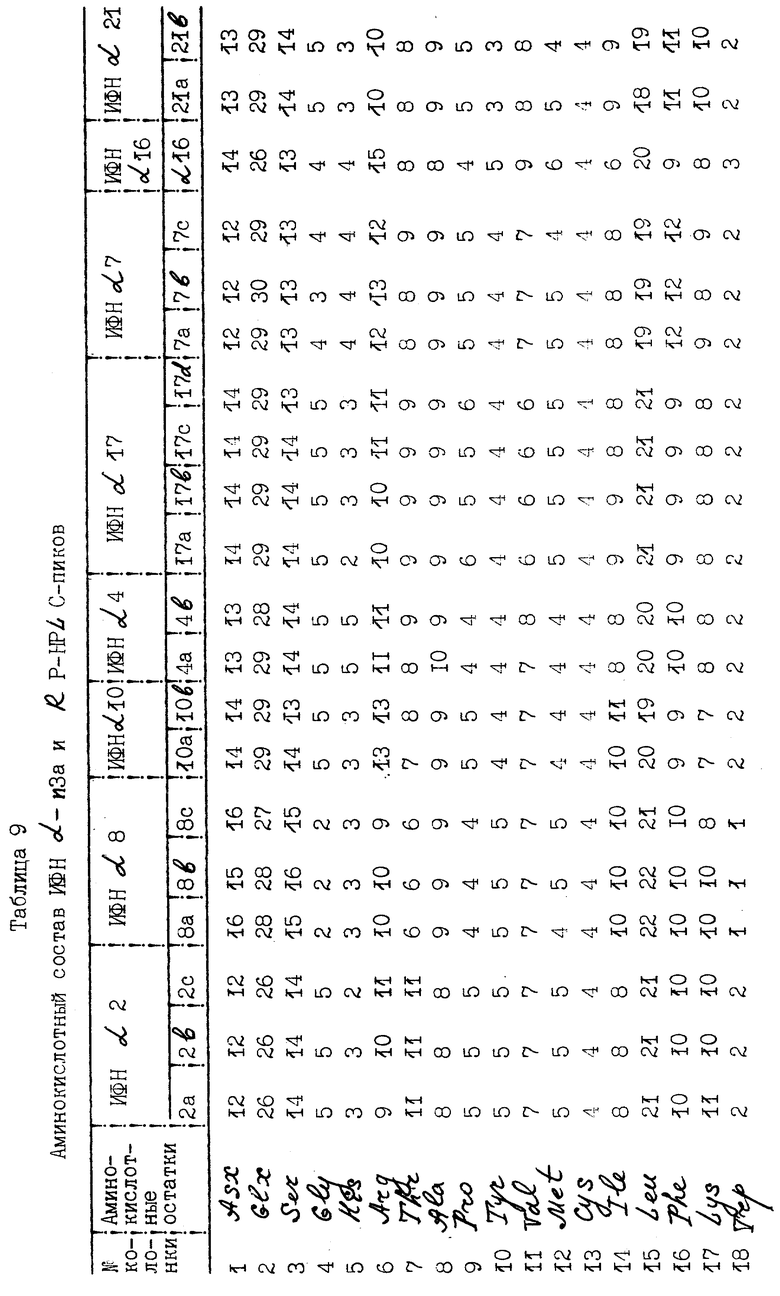
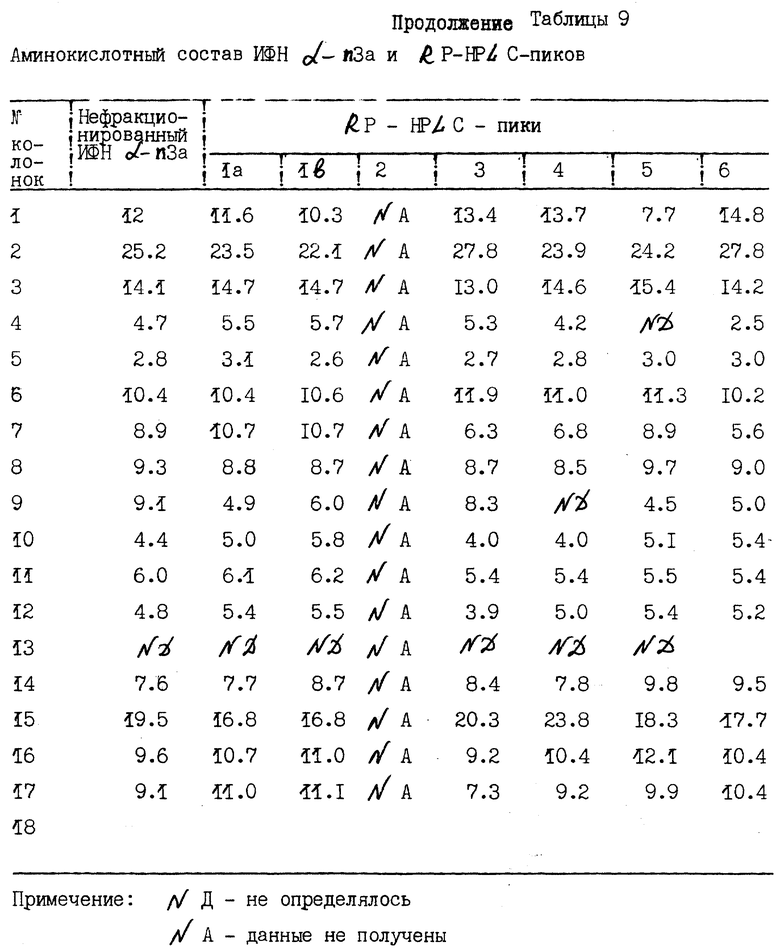
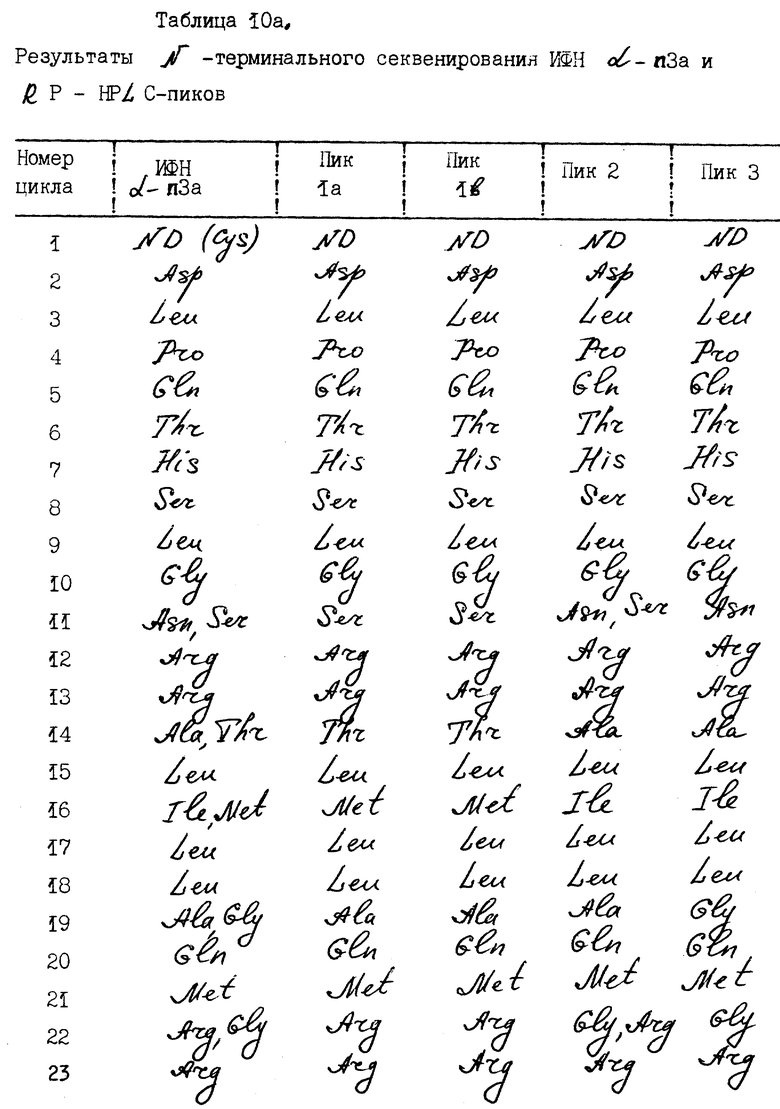
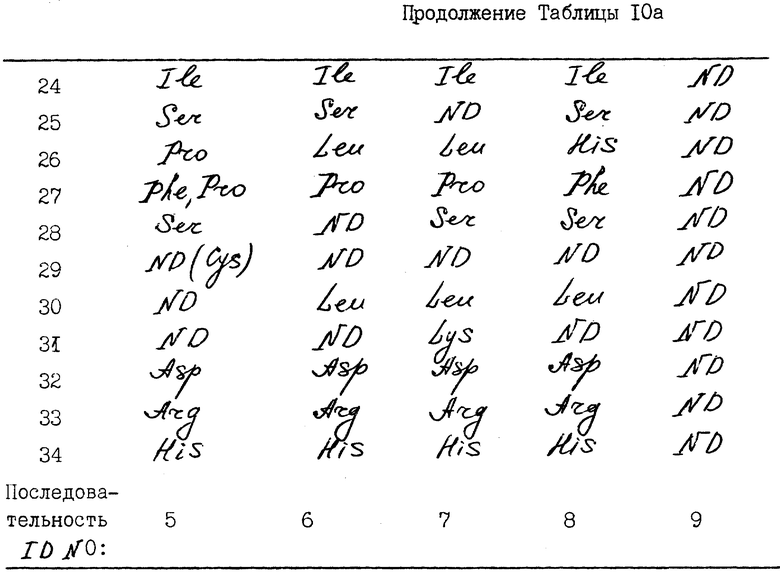
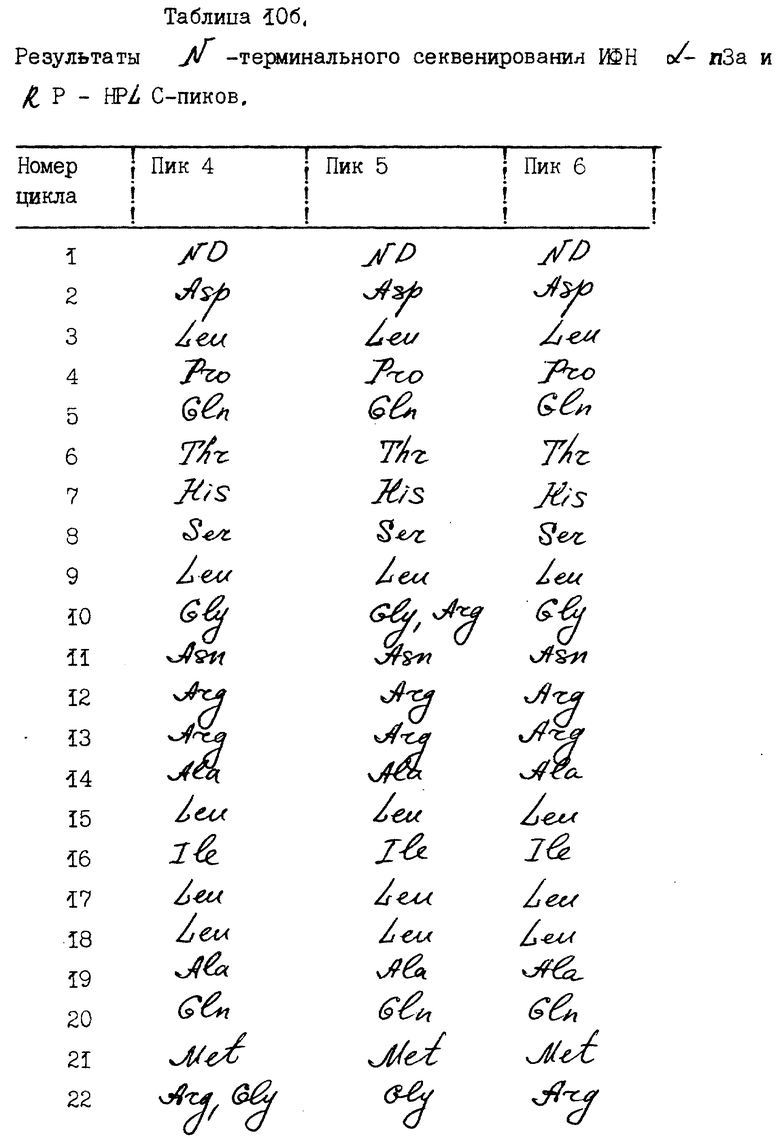
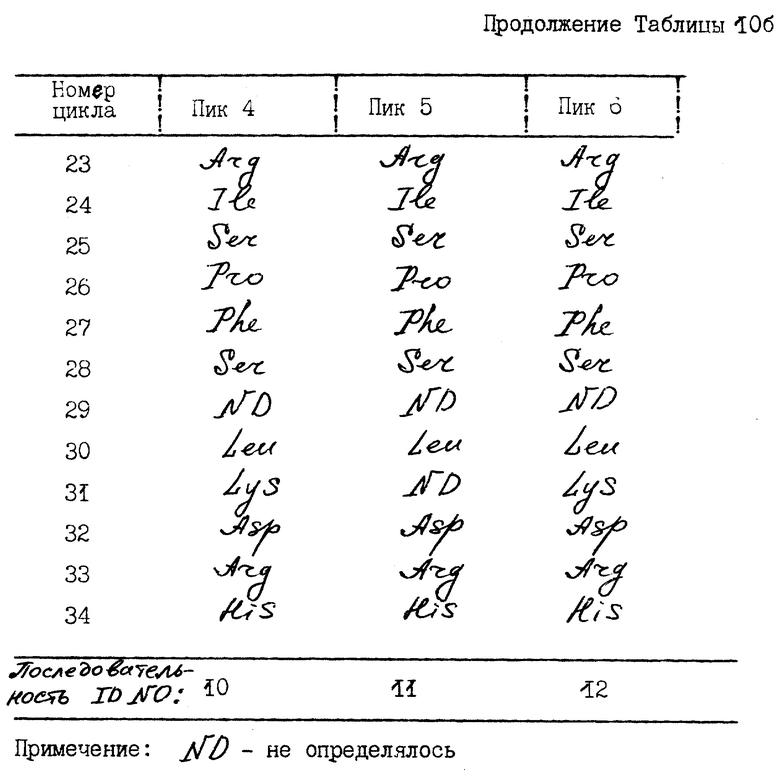


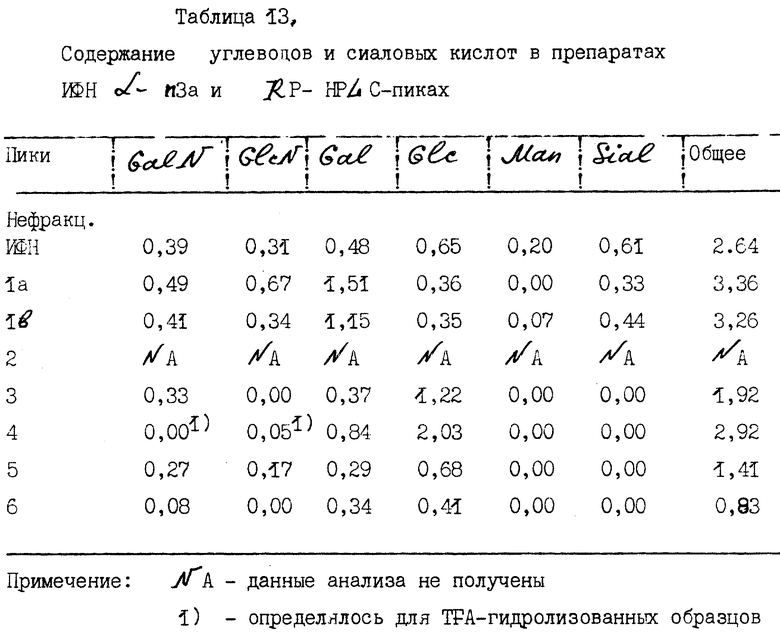
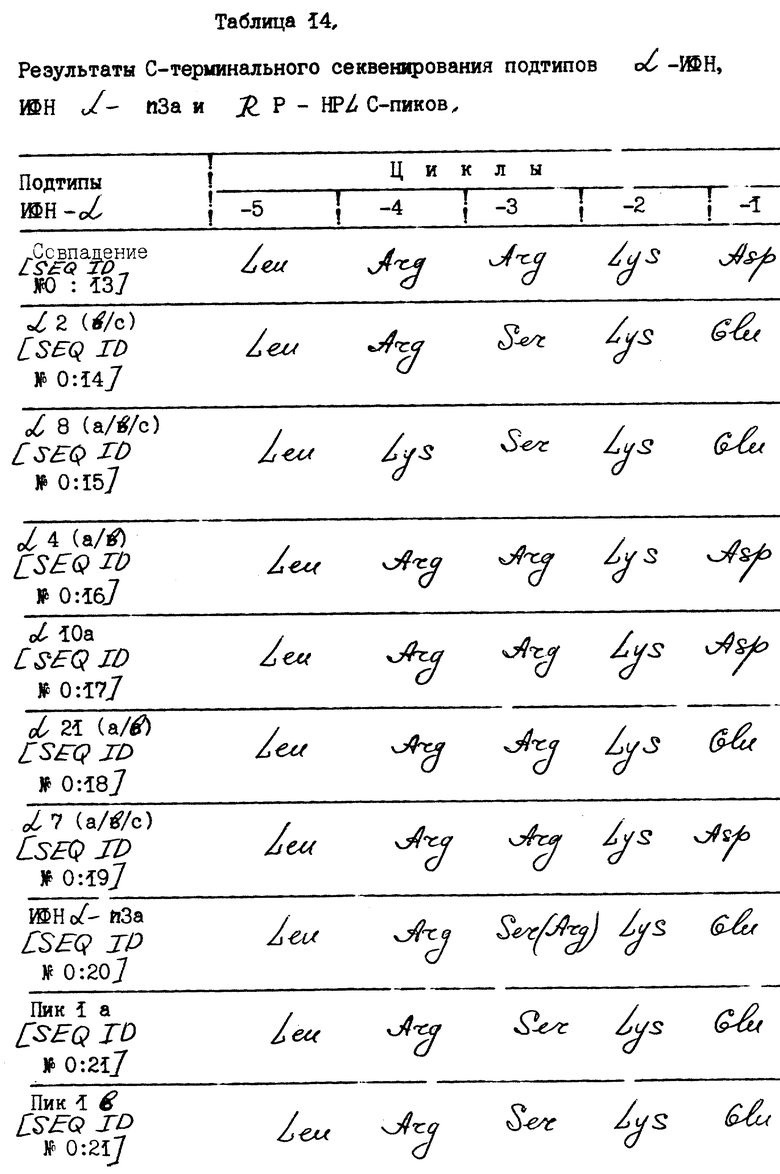
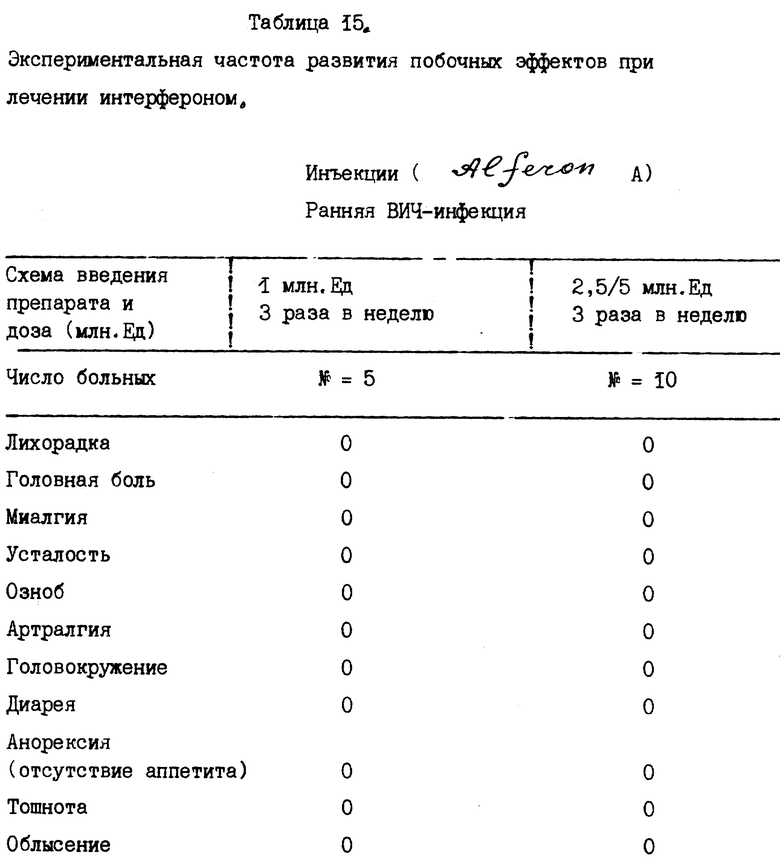
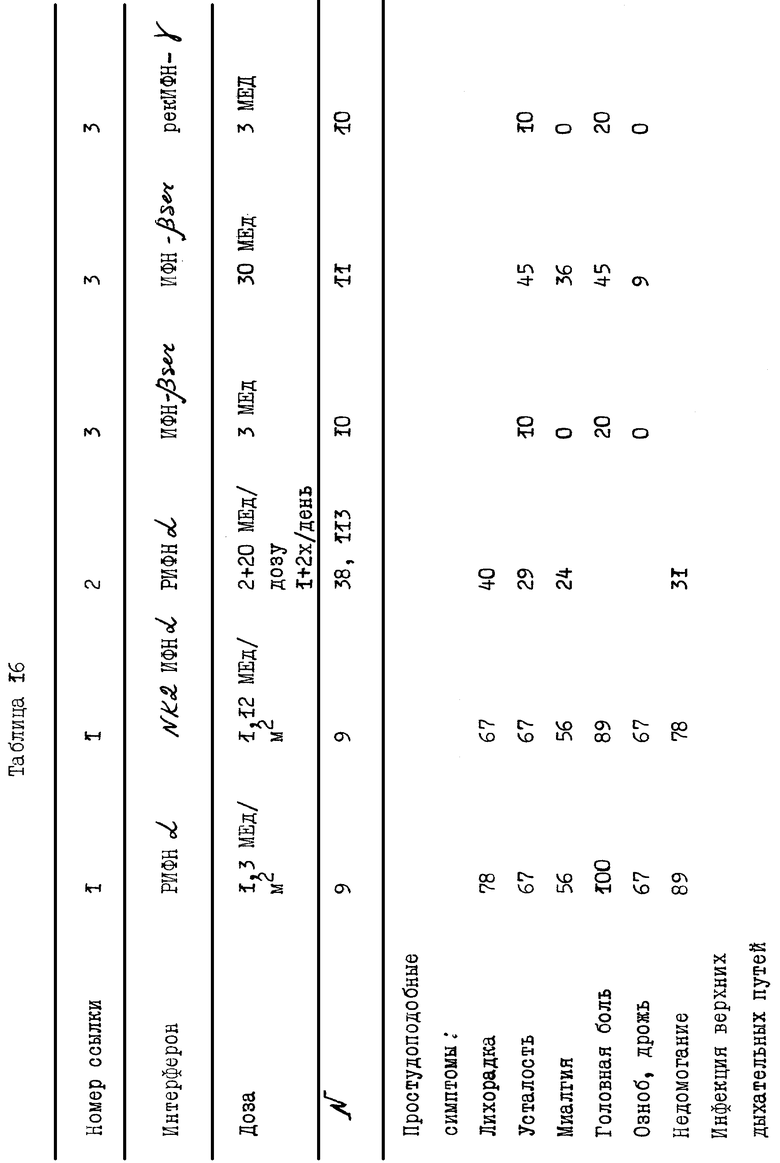














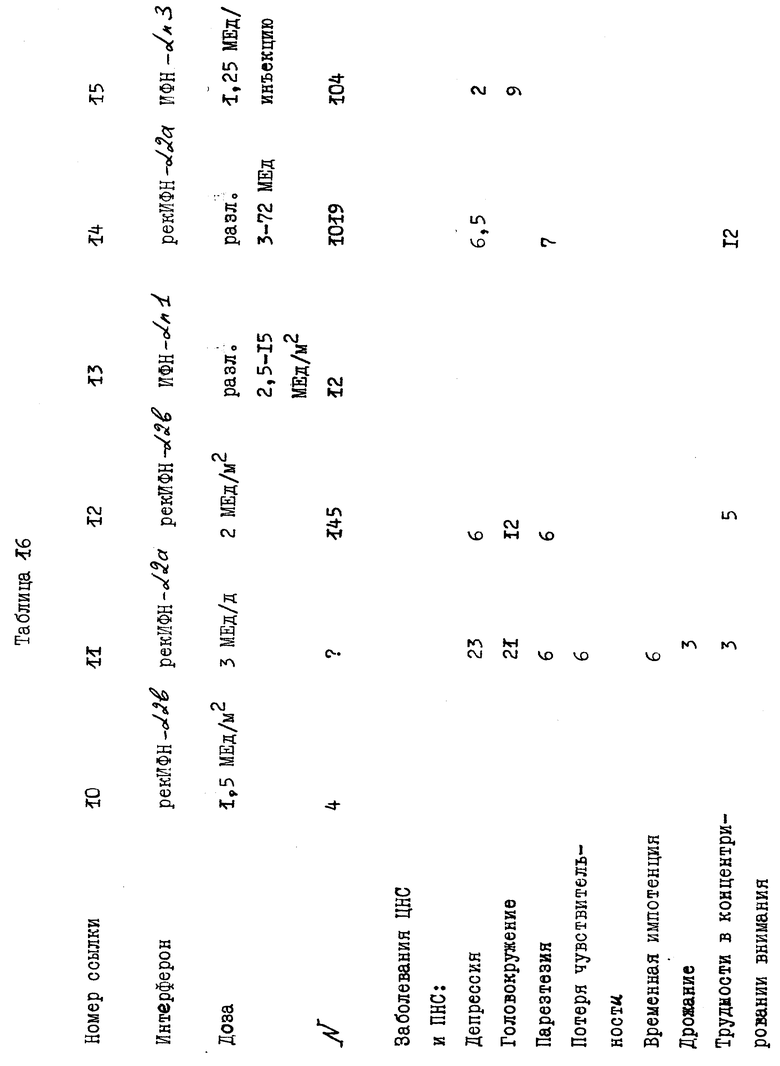





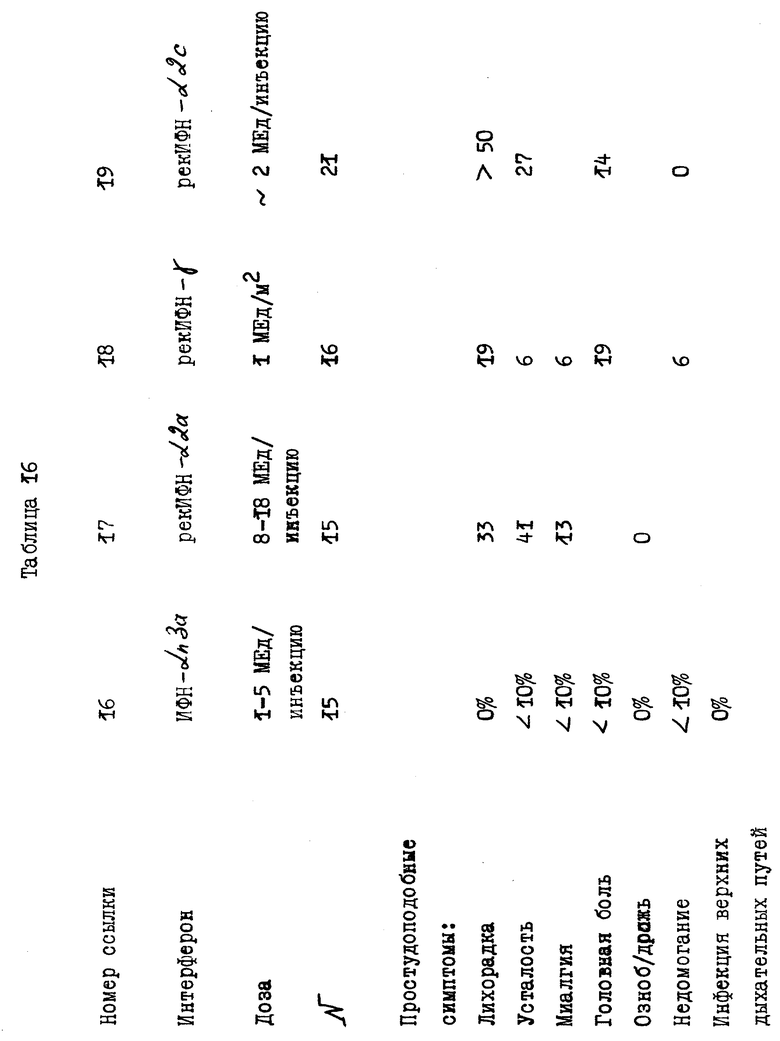






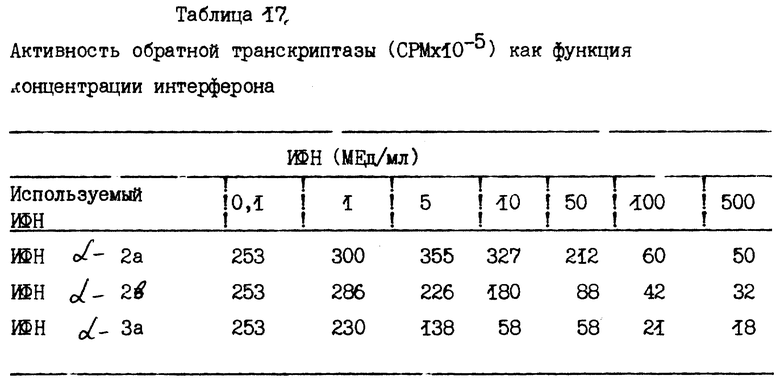


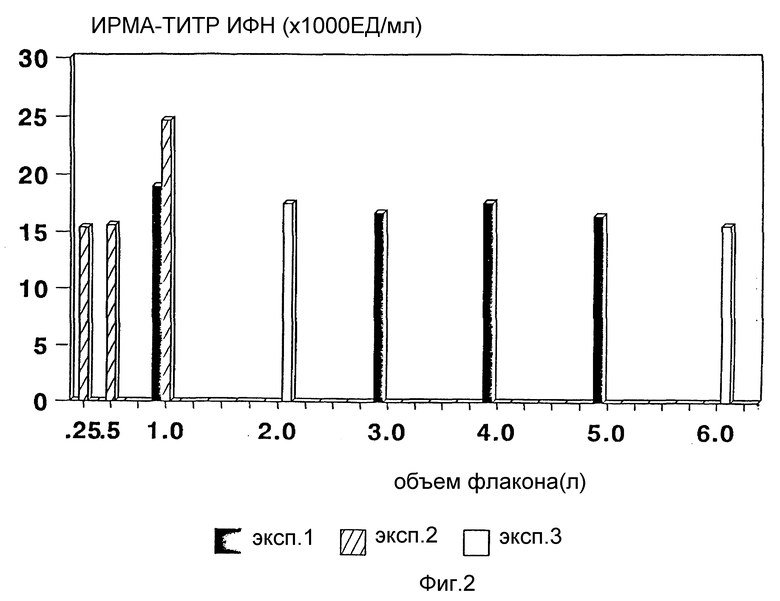
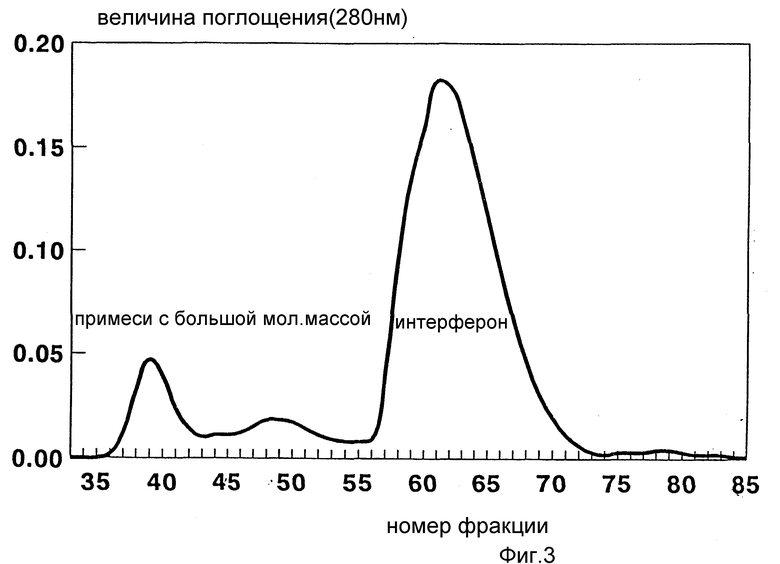
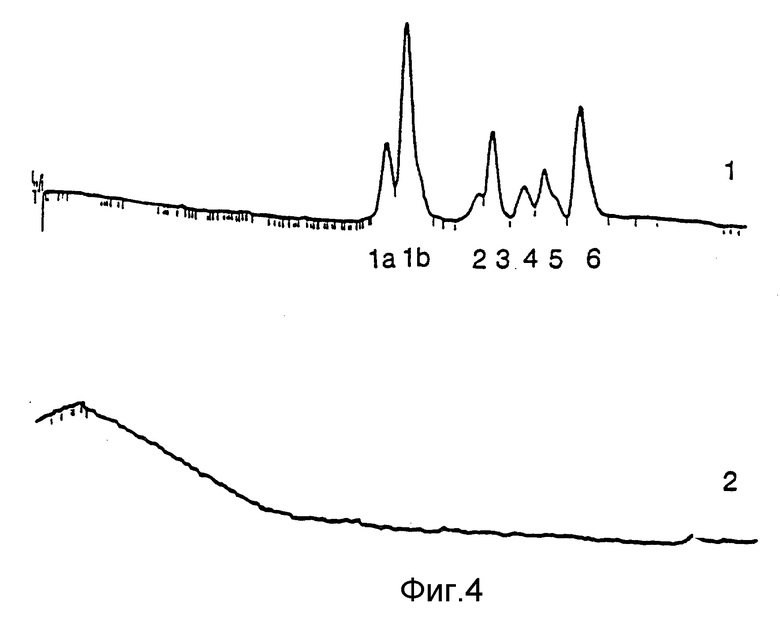
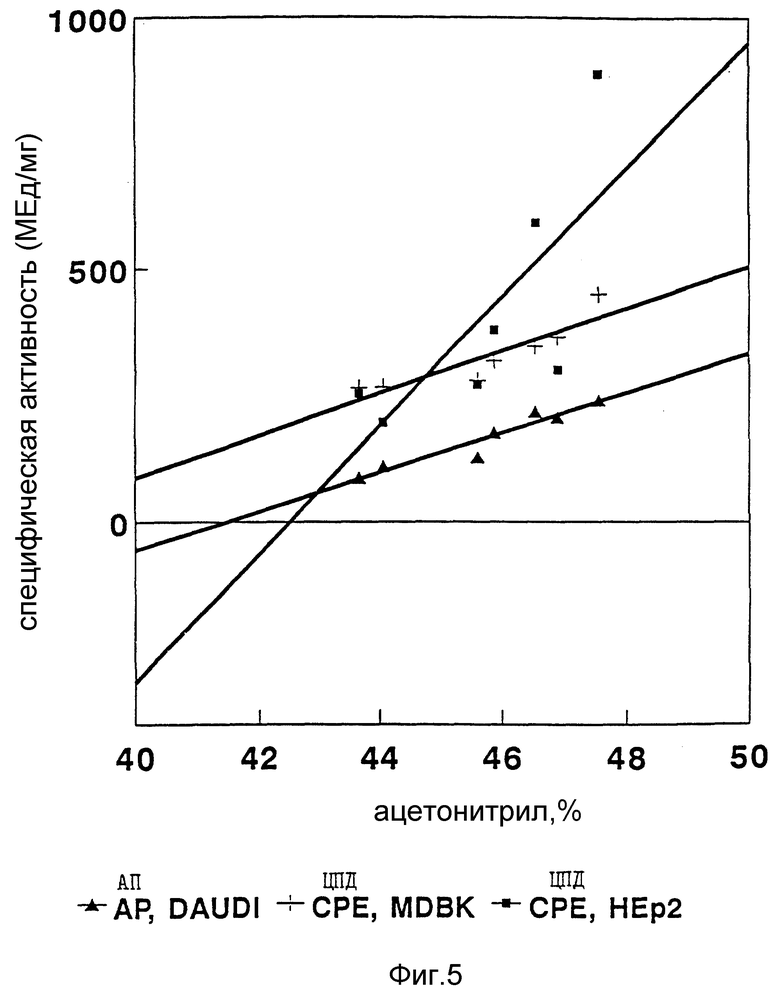
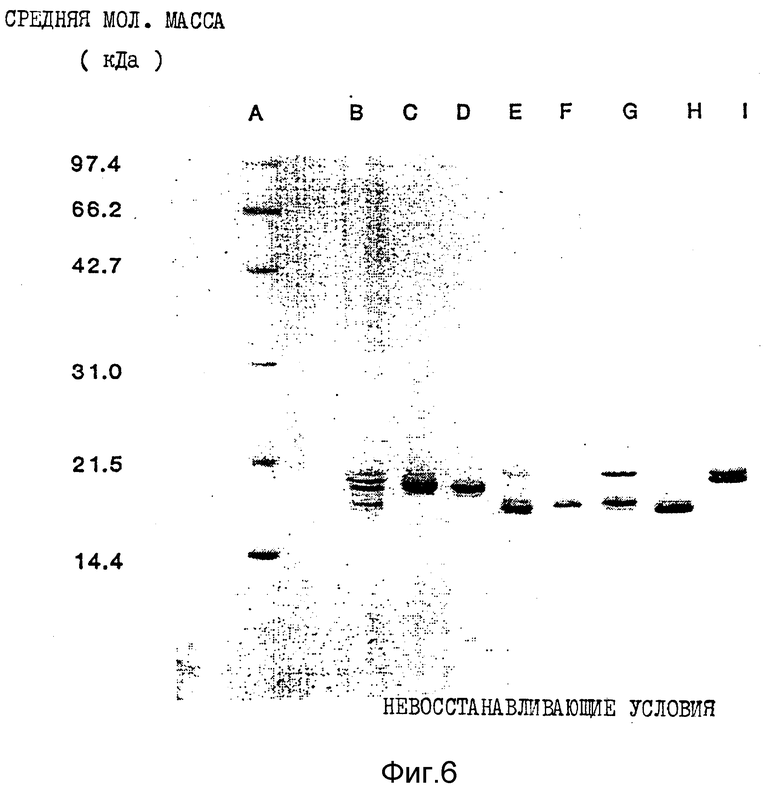
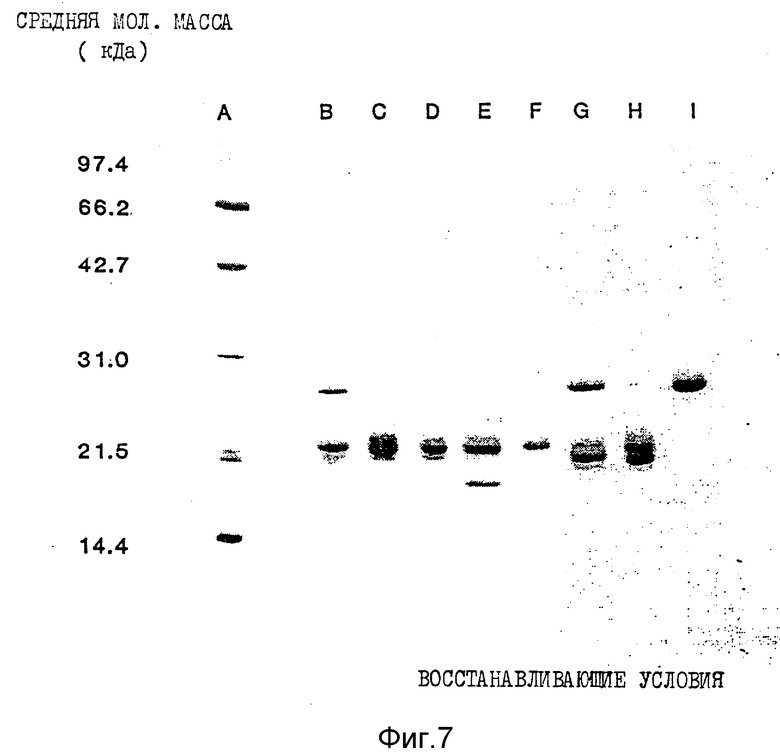
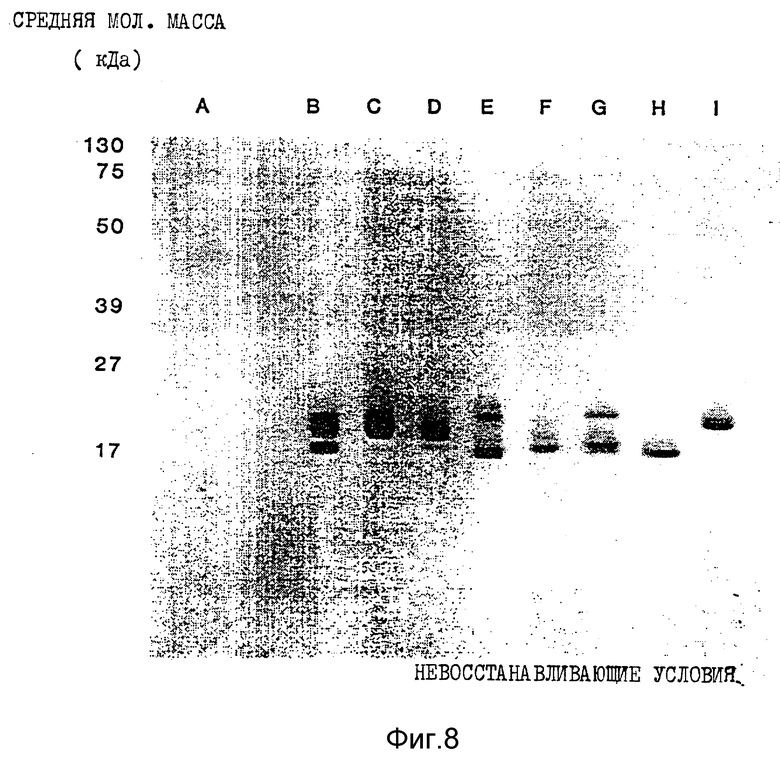
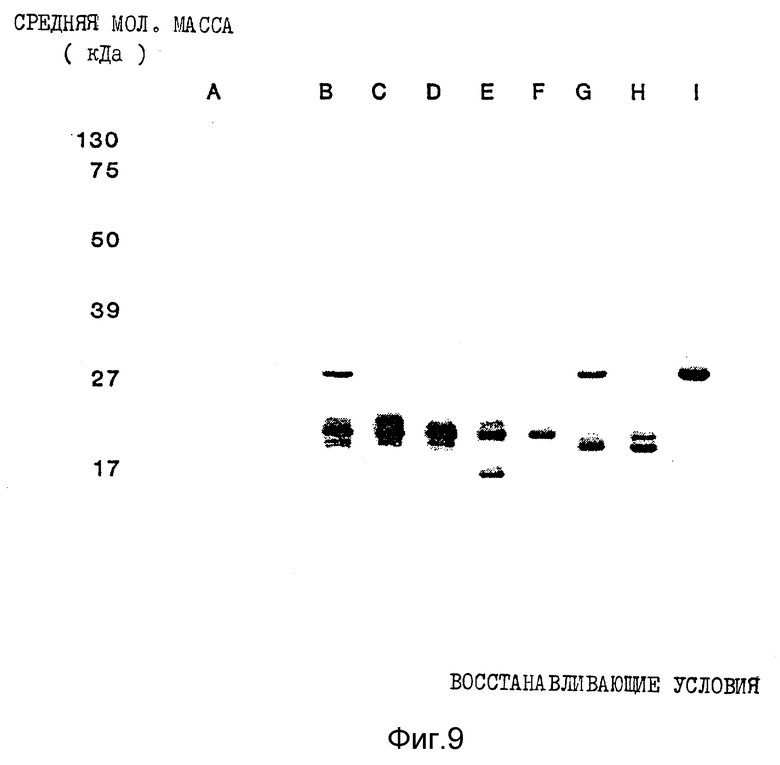
Изобретение относится к новой улучшенной композиции альфа-интерферона, характеризующейся специфической активностью, чистотой свыше 95%, и которая при использовании для человека характеризуется значительным уменьшением побочных эффектов, обычно связанных с композициями альфа-интерферона. Описан также способ получения данной композиции из лейкоцитов периферийной крови человека. Данная композиция альфа-интерферона может использоваться терапевтически при лечении раковых заболеваний, заболеваний иммунной системы и/или вирусной этиологии. Технический результат изобретения сводится к расширению арсенала средств нового поколения, обладающих противоопухолевой и иммуномодулирующей активностью. 3 с. и 6 з.п. ф-лы, 19 табл., 9 ил.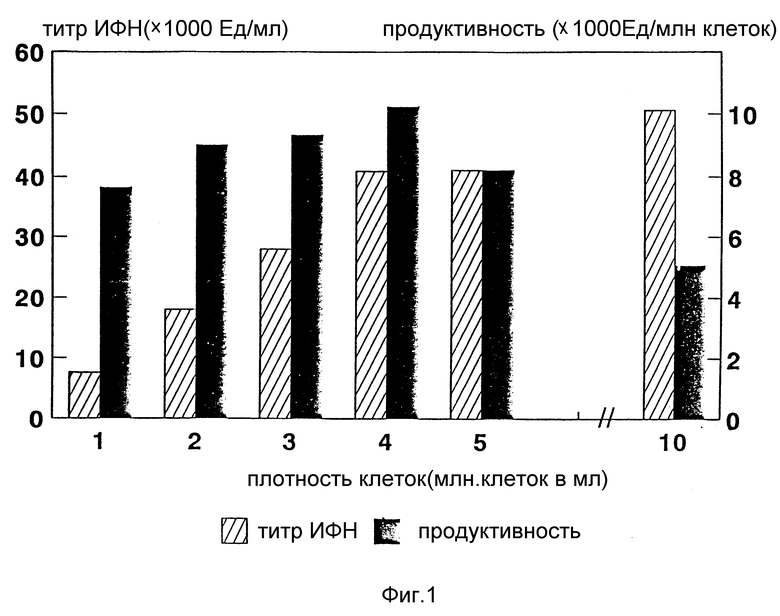
| ЭКРАН С ПЕРЕМЕЩАЮЩЕЮСЯ ПОВЕРХНОСТЬЮ | 1927 |
|
SU9702A1 |
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |
| Машковский М.Д | |||
| Лекарственные средства | |||
| Прибор для охлаждения жидкостей в зимнее время | 1921 |
|
SU1994A1 |
