Изобретение относится к открытию новой адгезионной молекулы, обозначенной ICAM-3, которая вовлечена в процесс, за счет которого популяция лейкоцитов распознает и прикрепляется к клеточному субстрату. ICAM-3 является посредником в клеточных взаимодействиях с другими лимфоцитами, макрофагами и нейтрофилами в участках воспалений и участках иммунных реакций.
Далее изобретение относится к использованию ICAM-3 отдельно или в сочетании с ICAM-1 и/или ICAM-2, для ингибирования межклеточной адгезии клеток гранулоцитов, лимфоцитов или макрофагов. Использование таких молекул обеспечивает способы лечения специфических и неспецифических воспалений.
Изобретение относится к терапевтическим и профилактическим способам супрессии инфицирования лейкоцитов HIV и, особенно, HIV-1 у индивидуумов, подверженных HIV или инфицированных HIV за счет введения ICAM-3 одной или в сочетании с ICAM-2. Оно предлагает лечение таких заболеваний, как AIDS (СПИД - синдром приобретенного иммунодефицита), которые вызываются вирусом HIV.
Изобретение относится к терапевтическим способам супрессии миграции инфицированных HIV-1 клеток из циркуляторной системы за счет использования ICAM-3, одной или в сочетании с ICAM-1 и/или ICAM-2. Поэтому оно предлагает лечение таких заболеваний, как AIDS (СПИД-синдром приобретенного иммунодефицита), которое вызывается вирусом HIV.
Изобретение относится также к способам супрессии гибели Т-клеток и образования "синцитий" у индивидуумов, инфицированных HIV, за счет использования ICAM-3, одной или в сочетании с ICAM-1 и/или ICAM-2. Поэтому оно обеспечивает лечение таких заболеваний, как AIDS (СПИД-синдром приобретенного иммунодефицита), которое вызывается вирусом HIV.
Изобретение относится к использованию ICAM-3, одной или в сочетании с ICAM-1 и/или ICAM-2, при лечении астмы.
Изобретение дополнительно относится к молекулам, способным связывать ICAM-3 (здесь и далее анти-ICAM-3). Связывание анти-ICAM-3 молекул с ICAM-3 предназначено для модуляции биологических функций, связанных с ICAM-3. Связующие молекулы настоящего изобретения могут быть антителами, пептидами или углеводородами, которые способны связываться с ICAM-3. Такие связующие молекулы пригодны для модулирования биологических функций ICAM-3.
Изобретение относится также к использованию анти-ICAM-3, одной или в сочетании с анти-ICAM-1 и/или анти-ICAM-2, или ингибирования межклеточной адгезии клеток гранулоцитов, лимфоцитов или макрофагов. Использование таких молекул предлагает способ лечения специфических и неспецифических воспалений.
Изобретение относится также к терапевтическим и профилактическим способам супрессии инфицирования лейкоцитов HIV и, особенно, HIV-1 у индивидуумов, которые подвержены HIV или инфицированы HIV, за счет ведения анти-ICAM-3, одной или в сочетании с анти-ICAM-1 и/или анти-ICAM-2. Поэтому настоящее изобретение предлагает лечение таких заболеваний, как AIDS (СПИД-синдром приобретенного иммунодефицита), которые вызываются вирусом HIV.
Изобретение относится к терапевтическим способам супрессии миграции HIV-1 инфицированных клеток из циркуляторной системы, используя анти-ICAM-3 агент, один или в сочетании с анти-ICAM-1 и/или анти-ICAM-2 агентами. Поэтому оно предлагает лечение таких заболеваний, как AIDS (СПИД-синдром приобретенного иммунодефицита), который вызывается вирусом HIV-1.
Далее изобретение относится к использованию анти-ICAM-3 агента, одного или в сочетании с анти-ICAM-1 и/или анти-ICAM-2, при лечении астмы.
A. Прикрепление лейкоцитов и функции.
Лейкоциты должны быть способы присоединяться к клеточному субстрату или соответствующей защиты хозяина против чужеродного вторжения бактерий или вирусов - см. Eisen. H.W. (в Microbiology, 3d Ed., Harper and Row, Philodelphia, PA. PA, 1980, pp. 290-295 и 381-418) для обзора этих функций. Лейкоциты должны быть способны прикрепляться к эндотелиальным клеткам таким образом, чтобы они могли мигрировать из циркуляторной системы к участкам воспаления. Более того, они должны прикрепляться к представляющим антиген клеткам таким образом, чтобы могла осуществляться нормальная специфическая иммунная реакция, и, наконец, они должны прикрепляться к соответствующим мишеневым клеткам таким образом, чтобы мог происходить лизис инфицированных вирусом или опухолевых клеток.
Недавно молекулы поверхности лейкоцитов, вовлеченные в вышеуказанные функции присоединения, были идентифицированы с использованием гибридомной технологии. Короче, моноклональные антитела, направленные против T-клеток человека (Davignon, D, et al. , Proc.Natt.Acad. Sci., USA, 78: 4535-5439 (1981) и клеток селезенки мышей (Springer, T.et al., Eur., J. Imunol. 9: 310-306 (1979) были идентифицированы, известно, что они связаны с лейкоцитными поверхностями и ингибируют присоединения описанных ранее родственных функций (Springer, T. et al., Fed. Proc., 44: 2660-2663 (1985)). Молекулы, идентицифированные такими антителами, были названы Mac-1 и Антигеном-1, связанным с лимфоцитной функцией (LFA-1). Mac-1 является гетеродимером, который найден на макрофагах, гранулоцитах и крупных гранулярных лимфоцитах. LFA-1 является гетеродимером, найденным на большинстве лимфоцитов (Springer, F.A. et al. , Immunol. Rev., 68: 111-135 (1982)). Эти две молекулы плюс третья молекула, p. 150-95 (которая имеет распределение ткани, аналогичное Mac-1) играют роль в клеточной адгезии (Keizer, G.D., et al., Eur. J. Immunol., 15: 1142-1147 (1985)).
Было обнаружено, что вышеуказанные молекулы лейкоцитов являются членами родственного семейства гликопротеинов (Sanches - Madrid, F. et al., J. Exper. Med. 158: 1785-1803; Keizer. G.D.et al., Eur. J. Immunol. 15: 1142-1147 (1985)), называемого "CD-18 семейством" гликопротеинов. Это семейство гликопротеинов состоит из гетеродимеров, содержащих одну альфа-цепь и одну бета-цепь. Хотя альфа-цепь каждого из антигенов отличается от другой, было обнаружено, что бета-цепи являются высококонсервативными (Sanches-Madrid. F., et al., J.Exper.Med, 15: 1785-1803 (1983)). Бета-цепь гликопротеинового семейства (иногда называемого как "CD-18"), как было найдено, имеет молекулярный вес 95 кд, тогда как альфа-цепи имеют молекулярный вес от 150 мд до 180 кд (Springer. T., Fed.Proc., 44: 2660-2663 (1985)). Хотя альфа-субъединицы протеинов мембраны не обладают высокой гомологичностью, присущей бета-субъединицам, тщательный анализ альфа-субъединиц гликопротеинов показал, что между ними имеется существенное сходство. Обзор аналогий между альфа- и бета-субъединицами LFA-1 родственных гликопротеинов представлен Sanches-Madrid, F, et al. (J. Exper. Med., 158: 586-602 (1983); J.Exper.Med., 158: 1785-1803 (1983)).
Была идентифицирована группа индивидуумов, которые оказались неспособными экспрессировать нормальные количества ни одного из адгезионных протеинов этого семейства на поверхности клеток их лейкоцитов (Anderson, D.C., et al., Fed. Proc.44: 2671-2677 (1985); Anderson, D.C., et al., J.Infect. Dis., 152: 668-689 (1985)). Эти индивидуумы страдают от "болезни дефицита адгезивности лейкоцитов ("LAD") (Anderson, D.C., et al., Fed.Proc., 44: 2671-2677 (1985); Anderson, D.C., et al., J.Infect.Dis., 152: 668-689 (1985)). Характерные особенности больных LAD включают язвы с некротическими мягкими тканями, недостаточное образование гноя и плохое заживление ран, а также ненормальные, зависящие от адгезии, функции лейкоцитов in vitro и подверженность хроническим и повторным бактериальным инфекциям. Гранулоциты таких LAD пациентов ведут себя несколько дефективно in vitro по сравнению с нормальными образцами в присутствии анти-CD18 моноклональных антител. То есть они не способы осуществлять такие функции, связанные с адгезией, как аггрегация или прикрепление к эндотелиальным клеткам. Однако более важным наблюдением является, то что эти пациенты не способны вырабатывать нормальную воспалительную реакцию из-за неспособности их гранулоцитов прикрепляться к клеточным субстратам. Наиболее значительным является тот факт, что гранулоциты этих LAD пациентов не способны достичь участков воспаления, таких как кожные инфекции, из-за их неспособности прикрепляться к эндотелиальным клеткам в кровеносных сосудах вблизи воспаленных участков. Такое прикрепление является необходимым шагом для транссудации.
Лимфоциты этих пациентов демонстрируют in vitro дефекты, аналогичные нормальным лимфоцитам, у которых CD-18 семейство молекул было антагонизировано антителами. Более того, эти индивидуумы не способны вырабатывать нормальную иммунную реакцию из-за неспособности их клеток прикрепляться к клеточным субстратам (Anderson, D.C., Fed.Proc., 44: 2671-2677 (1985); Anderson. D.C., et al., J.Infect. Dis., 152: 668-689 (1986)). Эти данные показывают, что иммунные реакции ослабляются, если лимфоциты не способны нормальным образом прикрепляться из-за недостаточной функции адгезионных молекул CD-18 семейства.
Таким образом, способность лейкоцитов поддерживать здоровье и жизнеспособность животных требует, чтобы они были способны прикрепляться к другим клеткам (например, эндотелиальным клеткам). Как было показано, такое прикрепление требует контакта клетка-клетка, который включает специфические рецепторы молекул, имеющиеся на клеточной поверхности лейкоцитов. Эти рецепторы дают возможность лейкоцитам прикрепляться к другим лейкоцитам к эндотелиальным клеткам и к другим несосудистым клеткам. Было найдено, что клеточные поверхности рецепторных молекул, LFA-1, Mac-1 и p15095 очень похожи друг на друга. У тех людей, лейкоциты которых не имеют на поверхности клеток рецепторных молекул, наблюдаются хронические и повторные инфекции, а также другие клинические симптомы, включая дефектные реакции антител.
Кроме того, так как адгезия лейкоцитов включена в процесс, в результате которого идентифицируются и отторгаются чужеродные ткани, понимание этого процесса имеет существенное значение в области трансплантации таких твердых органов, как почка, трансплантации таких нетвердых органов, как костный мозг, при трансплантации тканей, при аллергии и в онкологии.
B. Инфицированные HIV
HIV инфекция является причиной AIDS. Описаны множество вариантов HIV: двумя основными являются HIV-1 и HIV-2. В Северной Америке и Европе приобладает HIV-1, а HIV-2 преобладает только в Африке. Эти вирусы имеют аналогичные структуры и кодируют протеины с аналогичными функциями. Предполагают, что инфекция происходит в результате связывания протеина вируса (именуемого "gp120") с рецепторной молекулой (именуемой "CD4"), присутствующей на поверхности Т4 ("Т хелпер") лимфоцитов (Schnittman, S.M., et. al., J. Immunol. 141: 4181-4186 (1988), включено сюда по ссылке). После связывания с этим рецептором вирус проникает в клетку и реплицируется, и в этом процессе убивает Т клетки. Таким образом, разрушение Т4 популяции у индивидуумов и является прямым результатом HIV инфекции.
Разрушение Т клеток приводит к ослаблению способности инфицированного пациента бороться против инфекций. Хотя у индивидуумов, пораженных AIDS, часто развиваются раковые заболевания, связь между этими раковыми заболевания и HIV инфекцией в большинстве случае неопределена.
Хотя сам факт репликации вируса HIV является летальным для инфицированных клеток, обычно такая репликация детектируется лишь в небольшой части Т4 клеток инфицированного индивидуума. Полученные недавно результаты позволяют предположить, что наблюдается гораздо большая виремия, нежели предполагавшаяся ранее, и что инфицирование Т-клеток часто достигает 1%.
Некоторые направления исследований разрабатывают другие механизма, за счет которых HIV вирус осуществляет деструкцию Т4 популяции.
Помимо HIV репликации, HIV инфицированные клетки могут быть разрушены за счет действия цитотоксичных, киллерных клеток. Киллерные клетки обычно присутствуют у людей и служат для контроля за хозяином и разрушения любых чужеродных клеток (как, например, при переливании неподходящей крови или при трансплантации органов и т.д.) при их проявлении. После инфицирования HIV клетки Т4 выдвигают молекулы gp120 на свою клеточную поверхность. Киллерные клетки распознают такие Т4 клетки как чужеродные (а не нативные) и соответственно осуществляют их разрушение.
HIV инфекция может также привести к разрушению неинфицированных, здоровых клеток. Инфицированные клетки могут секретировать gp 120 протеины в кровеносную систему. Свободные gp 120 молекулы могут затем связаться с CD4 рецепторами здоровых, неинфицированных клеток. Такое связывание заставляет эти клетки принимать вид HIV инфицированных клеток. Цитотоксичные, киллерные клетки распознают gp 120, связанные с неинфицированными Т4 клетками, принимают их за чужеродные и осуществляют деструкцию клеток.
Дополнительным механизмом и одним из наиболее интересных для настоящего изобретения, по которому HIV является причиной гибели Т4 клеток, является механизм, осуществляемый за счет образования "синцитий". "Синцитий" является гигантской многоядерной клеткой, образующейся при слиянии такого количества, как несколько сотен Т4 клеток. Инфицирование HIV вызывает у инфицированных клеток способность сливаться с другими Т4 клетками, независимо от того, являются ли они инфицированными HIV или здоровыми клетками. Такие синцитии не могут функционировать и вскоре гибнут. Эта гибель сопровождается деструкцией как HIV инфицированных, так и HIV неинфицированных Т клеток. Этот процесс представляет собой интерес для настоящего изобретения, так как включает непосредственный контакт клетка-клетка Т4 клеток. Способность HIV инфицированных клеток образовывать синцитии указывает на то, что такие клетки приобретают средства для слияния со здоровыми клетками.
HIV инфекция и особенно HIV-1 инфекция, по-видимому, влияют на экспрессию клеточной поверхности интегринов лейкоцитов и реакции клеточной адгезии, осуществляемые этими гетеродимерами (Pitit, A.J., et. al., J. Clin. Jnvest, 79: 188 (1987); Hildreth, G.E.K., et al., Sciense, 244: 1075 (1989); Valentin, A., et al., G. Immunology, 144: 934-937 (1990); Rossen, P.D., et al. , Trans Assoc. American. Physicians, 102: 117-130 (1989), причем все ссылки включены сюда по ссылке). После инфицирования возрастает гомотипическая аггрегация U937 клеток, а также экспрессия клеточной поверхностью CD18 и CD11b (Petit, A.J., et al., J. Chin, Jnvest., 79: 188 (1987)). Инфицированные HIV клетки U937 прикрепляются к IL-1 стимулированному эндотелию гораздо чаще, нежели неинфицированные U937 клетки; такое поведение можно подавить, обрабатывая инфицированные клетки анти-CD18 и/или анти-CD11а моноклональными антителами или обрабатывая эндотелиальные субстраты анти-ICAM-1 антителами (Rossen. P. D. , et al. , Trans Assoc. American Physicians, 102: 117-130 (1989)). Моноклональные антитела к CD18 или CD11, как было обнаружено, также способны ингибировать образование синцитий, включающих фитогемагглютинином (PHA)-стимулированные лимфобластоидные клетки и конструктивно инфицированные CD4-негативные Т клетки (Hildreth, I. E. K. , et al., Sciense, 224: 1075 (1989)). Как было показано, обработка инфицированных вирусом клеток только анти-CD18 или анти-CD11а моноклональными антителами оказывает слабое воздействие на образование синцитий, что предполагает, что эти антитела принципиально защищают неинфицированные мишеневые клетки от инфекции (Hildreth, I. E.K. et al., Science, 244: 1075 (1989); Valent, H.A., et al. J. Immunology, 144: 934-937 (1990)). Valentin et al., (Valent H.A., et al., J. Immunology, 144: 934-937 (1990) недавно подтвердил эти наблюдения, продемонстрировав то, что моноклональные антитела, специфические для CD18, ингибируют синцитии, образующиеся в случае, когда непрерывные Т-клеточные линии культивируют совместно с HIV-1 инфицированными U937 клетками.
Хотя механизм, за счет которого моноклональные антитела, специфические для CD18 или CD11а, защищают подверженные клетки от слияния с HIV инфицированными клетками, остается неизвестным и это исследование вовсе не обязательно для целей настоящего изобретения, исследования с радиомеченными gp 120 позволяют предположить, что гетеродимеры, содержащие CD18, не предоставляют сайтов связывания для вируса (Valentin, A., et al., J. Immunology, 144: 934-937 (1990)). Таким образом, HIV инфекция включает взаимодействие клетка-клетка и/или взаимодействие вирус-клетка, которые имитируют такие взаимодействия, как клетка-клетка. Взаимодействие типа клетка-клета могут привести к транспорту клеток, не содержащих вируса, или инфицированных вирусом клеток через эндотелиальные барьеры. Взаимодействия типа вирус-клетка, которые имитируют взаимодействия клетка-клетка, могут облегчить или обеспечить присоединение свободного вируса к здоровым клеткам и/или инфицировать здоровые клетки.
Таким образом, настоящее изобретение возникло, частично из наблюдения того, что HIV инфекция, и особенно HIV-1 инфекция, приводит к повышению экспрессии CD11а/CD18 гетеродимера и связывающего его лиганда. Такое повышение экспрессии отличается тем, что оно повышает способность HIV-инфицированных Т-клеток прикрепляться или аггрегироваться друг с другом (то есть претерпевают "гомотипическую аггрегацию"). Так как такая гомотипическая аггрегация не наблюдается между покоящимися нормальными лейкоцитами, это означает, что экспрессия CD11/CD18 рецепторов и/или лигандов, таких как ICAM-1, необходима для такой аггрегации. LFA-1 должен связаться с ICAM-1 для того, чтобы могла произойти гомотипическая аггрегация. Как здесь было указано, ICAM-1 является единственным членом семейства CAM молекул, которые с высокой степенью экспрессируются на покоящихся Т-клетках. Только анти-ICAM-3 антитела способны блокировать адгезию Т-клеток к LFA-1, если Т-клетки не "активированы". Поэтому анти-ICAM-3 антитела можно использовать для супрессии аггрегации Т-клеток.
Дополнительно анти-CAM-3 антитела можно использовать для блокирования процессов адгезии инфицированных Т-клеток, которые позволяют осуществлять передачу HIV-1 с инфицированных клеток на здоровые клетки индивидуума и также обеспечивают или облегчают инфицирование здоровых клеток свободным вирусом C.
Миграция клеток, инфицированных HIV.
Миграция и распространение лейкоцитов является важным фактором для защиты индивидуумов от последствий болезней. Однако эти процессы являются также ответственными за миграцию и распространение инфицированных вирусом лейкоцитов. Наибольший интерес представляет миграция и распространение лейкоцитов, инфицированных HIV. Миграция таких клеток приводит к образованию экстраваскулярного фокуса и может вызвать опухоли и другие нарушения.
Гистологические исследования поврежденных органов выявляют фокальные экстраваскулярные мононуклеарные клеточные инфильтраты. Попытки идентифицировать инфицированные вирусом клетки в таких инфильтратах в центральной нервной системе выявили наличие HIV-1 инфицированных клеток. Эти исследования показали, что вирус HIV-1 находится преимущественно в моноцитах и микрофагах и других клетках этого направления (R.T. Johnson, et al., FASEB, 2: 2970 (1988); M. H. Stoler et al., J. Amer. Med. Assn., 256: 2370 (1986): S Jartner et al., Science, 233: 215 (1986)).
Механизмы, которые стимулируют образование эстраваскулярных инфильтратов HIV-1 инфицированных моноцитоидных клеток, ранее не были определены. Эти механизмы могут включать либо транспорт клеток, свободных от вируса, либо транспорт вируса через эндотелиальные барьеры внутри цитоплазмы инфицированных моноядерных клеток.
Так как инфицирование HIV-1 стимулирует экспрессию клеточной поверхностью молекул, которые облегчают прикрепление лейкоцитов к эндотелиальным клеткам сосудов и трансляцию лейкоцитов из крови к участкам экстраваскулярных тканей (C.W. Smith et al., J. Clin. Invect., 82: 1746 (1988), включено по ссылке), было предложено использовать антитела, которые ингибируют клеточную миграцию, для предотвращения распространения HIV-инфицированных клеток (WO 90/13316).
Д. Астма: Клинические характеристики.
Астма является гетерогенными семейством заболеваний. Она характеризуется гипер-реактивностью трахеобронхов на стимулы (Kay, A.B. Allergy and Inflammation, Academic Pess. N. Y. (1987), которая включена сюда по ссылке). Клинически астма проявляется сильным сужением трахеобронхов, густыми выделениями, спазмами, одышкой (dyspnea), кашлем и насморком. Хотя относительный вклад каждого из этих состояний неизвестен, общим результатом является возрастание сопротивления воздушных путей, гиперинфляция легких и грудной клетки и ненормальное распределение вентиляции и попытка крови в легких. Заболевание выражается в эпизодических периодах острых симптомов, прерывающих спокойные периоды. Острые приступы приводят к гипоксии и могут быть фатальными. Приблизительно 3% всего населения мира страдает от астмы.
Были описаны два типа астмы: аллергическая астма и идеосинкразическая астма. Аллергическая астма обычно связана с наследственными аллергическими заболеваниями, такими как риниты, крапивница, экзема и т.д. Эти условия характеризуются положительными реакциями покраснения и появления волдырей на подкожные инъекции витающих в воздухе антигенов (таких как пыльца, загрязнители окружающей среды или среды обитания и т.д.) и повышением уровней Ig в сыворотке. Развитие аллергической астмы, по-видимому, тесно связано с наличием IgH антител у многих пациентов. Больные астмой, у которых не обнаруживаются вышеуказанные признаки, рассматриваются как больные идиосинкразической астмой.
Считают, что аллергическая астма зависит от IgH реакции, контролируемой T и B лимфоцитами и активируется взаимодействием существующих в воздухе антигенов со связанными с мастоцитами заранее образовавшиеся IgE молекулами. Отпор антигенов должен происходить при концентрациях, достаточных для доведения IgE продуцирования в течение достаточно продолжительного промежутка времени для сенсибилизации индивидуумов. Будучи однажды сенсибилизированным, больным астмой может демонстрировать симптомы в ответ на чрезвычайно низкие уровни антигена.
Симптомы астмы могут усиливаться за счет присутствия и количества антигенов-триггеров, факторов окружающей среды, мест обитания, физических выделений и эмоциональных стрессов.
Астму можно лечить метилксантинами (такими как теофиллин), бета-адренергическими агонистами (такими как катехоламины, резорцинолы, салигенины и эфедрин), глюкокортикоидами (такими как гидрокортизон), ингибиторами дегрануляции мастоцитов (например, такими хромонами, как хромолиннатрий) и анти-холинэргиками (например, атропином).
Считают, что астма включает приток эозинофилов ("эозинофилия") в ткани легких (Frigas, E. J. Allergy Clin. Immunol., 77: 527-537 (1986), включено по ссылке).
Взгляд на иммунологические основы астмы был брошен в результате исследования бронхеоальвеолярного лаважа (Godard. P, et al., J. Allergy Clin. Immunol. , 70: 88 (1982)) и исследования тканей гладкой респираторной мускулатуры со снятым эпителием (Flavaham, N.A. et al., J. Appl. Physiol., 58: 834 (1985); Barnes, P. I. et al., Br. J. Pharmacol., 86: 685 (1985)). Хотя эти исследования и не привели к выяснению механизма, лежащего в основе иммунологии астмы, они привели к развитию общепринятой гипотезы, относящейся к иммунологической этиологии заболевания (см. Frigas. E. et al., J. Allergy Clin. Immunol., 77: 527-537 (1986).
Критериями патологии астмы являются массивная инфильтрация легочной паренхимы эозинофилами и разрушение функций мерцательного эпителия.
"Эозинофильная гипотеза" предполагает, что эозинофиты прикреплены к бронхам для нейтрализации вредных медиаторов, высвобождаемых тучными клетками легких (мастоцитами). В соответствии с этой гипотезой эозинофилы притягиваются к бронхам, где они дегранулируют с высвобождением цитотоксических молекул. После дегрануляции эозинофилы высвобождают такие энзимы, как гистамин, арилсульфатазу и фосфолипазу D, которые энзиматически нейтрализуют вредные медиаторы мастоцитов. Однако эти молекулы промотируют также деструкцию мерцательного аппарата, что предотвращает очищение от бронхиальной секции и вносит вклад в повреждения легких, характерные для астмы.
Так как астма включает миграцию клеток, было предложено использовать антитела, которые ингибируют такую миграцию для уменьшения действия аллергенов на субъектах (WO 90/10453).
Изобретение основано на открытии новой клеточной адгезионной молекулы, названной Межклеточной адгезионной молекулой-3 (ICAM-3). Настоящее изобретение дополнительно относится к функциональным производным ICAM-3, анти-ICAM-3 антителам, фрагментам указанных антител, гуманизированным анти-ICAM-3 антителам и к другим молекулам, способным связываться с ICAM-3 и ингибировать биологические функции ICAM-3.
Кроме того, настоящее изобретение включает диагностическое и терапевтическое использование для всех вышеуказанных молекул.
Более конкретно, настоящее изобретение включает ICAM-3 или его функциональное производное, практически не содержащие природных примесей.
В настоящем изобретении предложен способ получения рекомбинантных или синтетических ДНК молекул, способных кодировать или экспрессировать ICAM-3 или его функциональные производные.
Дополнительно настоящее изобретение предлагает антитела и особенно моноклональные антитела, способные связываться с молекулами, выбранными из группы, состоящей из ICAM-3 и функциональных производных ICAM-3.
Изобретение включает способы получения целевых гибридомных клеток, которые продуцируют антитела, которые способны связываться с ICAM-3 или их функциональными производными, включающие стадии:
(a) иммунизации животного иммуногеном, выбранным из группы, состоящей из: практически чистых ICAM-3; клеток, экспрессирующих ICAM-3; мембраны клеток, экспрессирующих ICAM-3; ICAM-3, связанных с носителем; пептидных фрагментов ICAM-3; или пептидных фрагментов ICAM-3, связанных с носителем;
(b) слияния клеток селезенки, выделенных из указанного животного, с миеломной клеточной линией;
(c) предоставления возможности слитым селезеночным и миеломным клеткам образовывать антитела, способные секретировать гибридомные клетки,
(d) скринирования гибридомных клеток на предмет гибридомных клеток, способных продуцировать анти-ICAM-3 антитела.
В изобретении предложен также способ модулирования осуществляемых ICAM-3 биологических функций клеток, причем указанный способ включает введение субъекту, нуждающемуся в таком лечении, эффективного количества ICAM-3 модулирующего агента, причем указанным ICAM-3 модулирующий агент выбирают из группы, состоящей из антитела, способного связываться с ICAM-3; фрагмента указанного антитела; причем указанный фрагмент способен связываться с ICAM-3; ICAM-3; функционального производного ICAM-3; и неиммуноглобулинового антагониста ICAM-3, отличного от ICAM-1, ICAM-2 или членов CD-18 семейства молекул.
В настоящем способе предложен также способ лечения специфических воспалений у людей и других млекопитающих, отличающийся тем, что указанный способ включает введение нуждающемуся в лечении субъекту такого количества противовоспалительного агента, которого достаточно для подавления воспаления, где противовоспалительный агент выбирают из группы, состоящей из антител, способных связываться с ICAM-3; фрагмента указанного антитела; причем указанный фрагмент способен связываться с ICAM-3; практически чистого ICAM-3; функционального производного ICAM-3; или неиммуноглобулинового антагониста ICAM-3, причем указанное воспаление является результатом трансплантации твердого органа (например, почки), трансплантации нетвердого органа (например, костного мозга) или пересадки тканей.
В изобретении предложен также способ лечения неспецифических воспалений у людей и других млекопитающих, отличающийся тем, что указанный способ включает введение нуждающемуся в лечении субъекту такого количества противовоспалительного агента, которого достаточно для подавления воспаления; причем противовоспалительный агент выбирают из группы, состоящей из: антител, способных связываться с ICAM-3; фрагментов указанных антител, причем указанный фрагмент способен связываться с ICAM-3; практически чистого ICAM-3; функциональных производных ICAM-3, или неиммуноглобулинового антагониста ICAM-3.
Далее изобретение включает способ лечения воспалений, где воспаление связано с болезненными состояниями, выбранными из группы, состоящей из синдрома расстройства дыхательной системы у взрослых, синдрома множественного поражения органов вторичного после сепсиса, кровотечения или травмы; поражений при переливании тканей миокарда или других тканей: острых гломерулонефритов; реактивных артритов; дерматозов с острыми воспалительными компонентами; острых пурулентных менингитов или других воспалительных заболеваний центральной нервной системы, например ударов, ожогов, гемодиализа, лейкафереза, язвенных колитов, болезни Крона, некротирующих энтероколитов, синдромов, связанных с трансфузией гранулоцитов, и токсичности, вызванной цитокинами.
Изобретение включает также способ подавления метастазов кроветворных опухолевых клеток, клеток, использующих члены CD-18 (особенно LFA-1) для миграции, где указанный способ включает введение пациенту, нуждающемуся в лечении, такого количества агента, которого достаточно для подавления метастазов; причем указанный агент выбирают из группы, состоящей из антител, способных связываться с ICAM-3; токсин-производных указанных антител; фрагментов указанного антитела, причем указанный фрагмент способен связываться с ICAM-3; токсин-производного указанного фрагмента; практически чистого ICAM-3; токсин-производного ICAM-3; функционального производного ICAM-3, токсин-производного ICAM-3; или неиммуноглобулинового антагониста ICAM-3, отличного от члена CD-18 семейства молекул.
Изобретение включает также способ подавления роста ICAM-3 экспрессирующих опухолевых клеток, причем способ включает введение пациенту, нуждающемуся в таком лечении, такого количества агента, которого достаточно для подавления роста, причем указанный агент выбирают из группы, состоящей из антител, способных связываться с ICAM-3; токсин-производного указанного антитела; фрагментов указанного антитела, причем указанный фрагмент способен связываться с ICAM-3; токсин-производного указанного фрагмента; токсин-производного члена семейства CD-18 молекул; или токсин-производного функционального производного членов семейства молекул CD-18.
В изобретении предложен также способ детектирования наличия клеток, экспрессирующих ICAM-3, причем указанный способ включает:
(a) инкубирование клеток или экстракта клеток в присутствии молекул нуклеиновой кислоты, причем молекулы нуклеиновой кислоты способны гибридизоваться в ICAM-3 MRNA; и
(b) определение наличия гибридизации молекул нуклеиновой кислоты с комплементарными нуклеиновой кислоте молекулами, присутствующими в указанных клетках или в указанном экстракте указанных клеток.
В изобретении предложен способ определения наличия ICAM-3 в образцах биологических жидкостей, причем указанный способ включает:
(a) инкубирование указанного образца с антителами или их фрагментами, которые способны связываться с ICAM-3; и
(b) детектирование наличия или отсутствия связывания антитела указанным образом.
В изобретении предложена также фармацевтическая композиция, содержащая:
(a) агент, выбранный из группы, состоящей из: антитела, способного связываться с ICAM-3; фрагмента указанного антитела, причем указанный фрагмент способен связываться с ICAM-3; практически чистого ICAM-3; функционального производного ICAM-3 или неиммуноглобулинового антагониста ICAM-3, отличного от членов CD-18 семейства молекул иммуносуппрессивного агента.
Фиг. 1. Адгезия клеточных линий к очищенным LFA-1.
Адгезия клеточной линии (A) и лимфоцитов (B) к очищенным LFA-1 объясняется ICAM-1, ICAM-2 и ICAM-3. Связывание BCECF меченых клеток на LFA-1 - покрытых микротитровальных ячейках в присутствии блокирующих MAb специфических для LFA-1, ICAM-1, ICAM-2 и ICAM-3. Контрольные ячейки были покрыты без LFA-1.
Фиг. 2. Цитометрический анализ в потоке клеточной экспрессии ICAM-1, ICAM-2 и ICAM-3.
(A) Лимфобластоидные клеточные линии были мечены насыщающими количествами либо MAb RR1 (1) анти-ICAM-1, MAb CB-IC2/1 (анти-ICAM-2), MAb CBR-IC3/1 (анти-ICAM-3), либо несвязывающим контролем MAb X63 (тонкие линии), и затем с последующим FITC-антимышиным иммуноглобулином.
(B) Покоящиеся человеческие лейкоциты и трехдневные PHA-активированные лимфоциты анализировали на предмет ICAM экспрессий, как указано ранее.
Фиг. 3. Иммуноосаждение ICAM-3.
(A) Лизаты 125I-меченых клеток имммуноосажденные любым из несвязывающихся контрольных X63 MAb CB-IC3/1 (анти-ICAM-3).
(B) 125I-меченые SKW3 клеточные лизаты, обработанные (+) или (-)N-гликаназой и иммуноосажденные любым из несвязывающихся контрольных MAb X63, MAb W6/32 (анти-HLA-A, B, C), MAb RR1/1 (анти-CAM-1), MAb CBR-IC2/1 (анти-CAM-2), либо MAb CB-ICB (анти-ICAM-3).
Фиг. 4. Иммуноосаждение ICAM-3 из различных источников.
CAM-3 иммуноосаждают из 125I-меченых лимфоцитных и нейтрофильных лизатов, используя MAb-соединенную сефарозу. Иммуноосаждение ICAM-3 разрешают, используя электрофорез на полиакриламидном геле. Остальные MAb используют для иммуноосаждения других молекул клеточной поверхности. Человеческий Ig сефаразы (контрольные полоски), CBR-IC2/1 (ICAM-2), CBR-IC3/1 (ICAM-3), TS 2/4 (LFA-1), LM2/1 (MAC-1).
Фиг. 5. Влияние PMA на SKW3, связывание с ICAM-1 и CAM-3.
Способность SKW3 клеток связываться с очищенными ICAM-1 и ICAM-3 и влияние PMA стимуляции этого связывания тестируют описанным ранее способом (Dustin et al. , Nature, 341:619 91989); Marlin et al., Cell, 51: 813-819 1987)). Один из представительных экспериментов представлен, а прямоугольники ошибок показывают стандартное отклонение (SD).
Фиг. 6. Антитела, блокирующие PMA стимулированную SKW-3 клеточную адгезию.
Были тестированы различные антитела по их способности блокировать PMA стимулированные SKW-3 клетки связывания с очищенными ICAM-1 или ICAM-3, как было указано ранее (Dustin et al., Nature, 341:619 (1989); Marlin et al., Cell, 51: 813-819 (1987). Один представительный эксперимент представлен, а прямоугольники ошибок показывают SD.
Фиг. 7. Связывание COS клеточных трансфектантов с очищенными ICAM.
COS клетки трансфектируют LFA-1 вектором экспрессии, как было указано ранее (de Fougerolles et al., J.Exp.Med., 175: 185-190 (1992)). Эти трансфектированные клетки тестируют на предмет их способности связываться с иммобилизованными ICAM-1 и ICAM-3 в присутствии (или без) анти-LFA-1 антител (TS1/22). Представлен один представительный пример, а прямоугольники ошибок указывают SD.
Фиг. 8. Температурная зависимость PMA стимулированного SKW-3 связывания с ICAM.
PMA стимулированную SKW3 клеточную способность связываться с очищенным ICAM-1 и ICAM-3 тестируют при 4, 16, 22 и 37oC, как указано ранее (Marlin et al. , Cell, 51: 813-819 (1987). Анализ проведен в присутствии или без LFA-1 MAb (TS1/22). Представлен один представительный пример, а прямоугольник ошибок указывает SD.
Фиг. 9. Блокирование антителами PHA-стимулированного деления T клеток.
Тестировали различные антитела по их способности блокировать PHA стимулированное деление T клеток, как указано ранее (Krensky et al., J. Immunol., 131:616 (1983)).
Фиг. 10. Влияние анти-ICAM антител на смешанные лимфоцитные реакции.
Различные антитела тестировали по их способности блокировать смешанные лимфоцитные реакции, как указано ранее (Krensky et al., J. Immunol., 131: 611-616 (1983)).
Фиг. 11. Газохроматографический анализ пептидных фрагментов ICAM-3.
ICAM-3 очищают и переваривают Lys-C. Пептидные фрагменты разделяют, используя высокоэффективную жидкостную хроматографию.
Фиг. 12. Диаграммное представление клона 11.2.
Схематическое представление клона 11.2. Показаны положения NK-10 и NK-17 пептидов, различные ДНК-последовательности, полученные таким образом, и трансмембранные участки.
Фиг. 13. Последовательное сравнение NK-IT примированной последовательности с ICAM-1 человека.
GAP программу использовали для идентификации гомологичности последовательности NK-10 примированной последовательности и протеина ICAM-1 человека.
Фиг. 14. Сравнение последовательности RM13 примированной последовательности с ICAM-1 человека.
GAP программа была использована для идентификации гомологичности последовательности для RM13 примированной последовательности и ICAM-1 человека.
Фиг. 15. Сравнение последовательностей RM13 примированной последовательности с ICAM-2 человека.
GAP программу используют для идентификации гомологичности последовательности для RM13 примированной последовательности и ICAM-2 человека.
Фиг. 16. Сравнение последовательностей Т7 примированной последовательности с ICAM-1 человека.
GAP программу использовали для идентификации гомологичности последовательности T7 примированной последовательности и ICAM-1 человека.
Фиг. 17. Сравнение последовательностей T7 примированной последовательности с ICAM-2 человека.
GAP программу использовали для идентификации гомологичности последовательностей T7 примированной последовательности и ICAM-2 человека.
Фиг. 18. Частичная последовательность ICAM-3.
ДНК последовательность клона 11.2 определяют, используя стандартную процедуру.
A. Последовательности, полученные с 5 клона ICAM-3. Эти последовательности получают, примируя клон 11.2 T7 праймером.
B. Последовательности, полученные внутри ICAM-3 гена. Эти последовательности были получены при примировании клона 11.2 NK-10 зондом.
C. Последовательности, полученные с 3 конца ICAM-3. Эти последовательности были получены при примировании клона 11.2 обратным RM13 праймером.
Изобретение основано на открытии ранее неидентифицированного связывающего лиганда с LFA-1. Такие молекулы, как молекулы семейства CD-18, которые включены в процесс клеточной адгезии, называются далее "адгезионными молекулами".
I. LFA-1
Лейкоцитная адгезионная молекула LFA-1 осуществляет взаимодействия широкого круга лимфоцитов, моноцитов, нативных киллерных клеток и гранулоцитов с другими клетками в иммунных реакциях и при воспалительных процессах (Springer, T.A. et al., Ann. Rev. Immunol., 5: 223-252 (1987)).
A-1 является рецептором для ICAM-1, ICAM-2 и вновь идентифицированного ICAM-3 (раскрытого здесь). Эти молекулы поверхностей, в основном, экспрессируются на некоторых тканях и индуцируются на других в воспалительных процессах (Marlin, D. et al., Cell, 51: 813-819 (1987); Dustin. M.L. et al., Immunol, 137: 245-254 (1986); Dustein, M. L., et al., Immunol. Today 9: 213-215 (1988); патентная заявка США N 07/017440, поданная 26 февраля 1987 г. , и патентная заявка США N 07/250446, поданная 28 сентября 1988 г. (обе включены сюда по ссылке).
LFA-1 функции как в антиген-специфических, так и в антигенезависимых T цитотоксических, T хелперных, природных киллерах, гранулоцитах и моноцитах взаимодействует с другими типами клеток (Springer, T.A., et al., Ann Rew. Immunol. , 5: 223-252 (1987); Kishimoto, T.K., et al., Adv. Immunol. (1988 в печати)).
II. ICAM-1
ICAM-1 представляет собой однонитевой гликопротеин, меняющийся по массе на различных типах клеток от 76 до 114 кД, и является членом Ig суперсемейства с пятью C-подобными доменами (Dustin, M.L, et al., Immunol. Today, 9: 213-15 (1988); Stanton, D.E, et al., Cell 52: 925-933 (1988); Simmous, D et al., Nature. 331: 624-627 (1988)). ICAM-1 сильно индуцируется цитокинами, включая IFN-g, TNF и IL-1 на широком круге типов клеток (Dustin, M.L, et al. , Immunol. Today, 9: 213-215 (1988)). Индуцирование ICAM-1 на эпителиальных клетках, эндотелиальных клетках и фибробластах вызывает зависимую адгезию лимфоцитов (Dustin, M.L. et al. J. Immunol., 137: 245-254 (1986); Dustin, M. L. et al., J. Cell Biol., 107: 321-331 (1988); Dustin, M.L. et al., J. Exp. Med 167: 1323-1340 (1988)). Адгезия блокируется за счет предварительной обработки лимофитов LFA-1 MAb или предварительной обработки других клеток ICAM-1 MAb (Dustin. M.L. et al., J. Immunol, 137: 245-254 (1986); Dustin. M.L. et al. J. Cell Biol, 107: 231-331 (1988); Dustin. M.L. et al., J. Exp. Med., 167: 1323-1340 (1988)). Идентичные результаты с очищенным ICAM-1 в искусственных мембранах или чашках Петри демонстрируют, что LFA-1 и ICAM-1 являются рецепторами друг для друга (Marlin, S.D. et al., Cell, 51: 813-819 (1987); Makgoba, M.W., et al., Nature, 331: 86-88 (1988)). Для ясности здесь их будем называть "рецепторы" и "лиганды" соответственно. Дальнейшие описания ICAM-1 приведены в патентных заявках США N 07/045963, 07/115798; 07/155943, 07/189815 или 07/250446 (все эти заявки включены сюда по ссылке).
III. ICAM- 2
Другие LFA-1 лиганды, отличные от ICAM-1, были постулированы (Rothlein, R, et al., J. Immunol, 137: 1270-1274 (1986); Makgoba M.W., et al., Eur. J. Immynol, 18: 637-640 (1988); Dustin, M.L. et al., J. Cell Biol., 107: 321-331 (1988)). Второй идентифицированный LFA-1 лиганд обозначен "ICAM-2".
ICAM-2 отличается от ICAM-1 клеточным распределением и отсутствием индукции цитокина. ICAM-2 является интегральным мембранным протеином с двумя Ig - подобными доменами, тогда как ICAM-1 имеет 5 Ig - подобных доменов (Staunton, D.E, et al., Cell, 52: 925-933 (1988); Simmons, D, et al., Nature 331: 624-627 (1988)). Замечательно, что ICAM-2 гораздо более тесно связан с двумя наибольшими N-терминальными доменами ICAM-1 (34% идентичности), нежели ICAM-1 или ICAM-2, близкими другим членам суперсемейства Ig, что демонстрирует субсемейство Ig - подобных лигандов, которые связывают тот же рецептор семейства интегринов. Следующие описание ICAM-2 приводится в патентной заявке США N 07/454294 (включенной сюда по ссылке).
IV. ICAM-3
Изобретение относится к открытию третьего LFA-1 лиганда, обозначенного "ICAM-3".
Развитие MAb до ICAM-2 позволило проанализировать несколько LFA-1 - зависимых ICAM-1 - независимых явлений и предположить, что существует третий лиганд для LFI-1. Связывание нескольких типов клеток, таких как эпителиальные и эндотелиальные клетки, с очищенным LFA-1 можно полностью блокировать сочетанием ICAM-1 и ICAM-2 MAb, тогда как ICAM-1, ICAM-2 независимые пути адгезии к LFA-1 существовали для многих лимфоидных клеточных линий, включая T клеточную лимфомную линию клеток, SKW3 (фиг. 1).
Для характеристики такого хода адгезии Mab довели до SKW3 и, вместе с анти-ICAM-1 и анти-ICAM-2 Mab, скринировали на предмет способности ингибировать, связывать эту клеточную линию с очищенным LFA-1. Один MAb, CBR-IC3/1, выбирают, так как он полностью ингибирует этот новый путь адгезии (фиг. 1). SKW3 адгезия с очищенным LFA-1 только слегка ингибируется комбинацией блокирующих анти-ICAM-1 и анти-ICAM-2 MAb. Если анти-ICAM-3 MAb, CB-IC3/1, добавляют отдельно, она значительно ингибирует адгезию и это ингибирование оказывается полным, если объединена с блокирующей ICAM-1 MAb. Так адгезия SKW3 к очищенным LFA-1 в значительной степени осуществляется за счет ICAM-3, а также частично за счет ICAM- 2. При адгезии и LFA-1 каждая из четырех клеточных линий использует три ICAM в различной степени, на что указывают различные картины MAb ингибирования. Адгезия B лимфобластоидной клеточной линии, Y, происходит главным образом за счет ICAM-1 и за счет небольшого вклада за счет ICAM-2 и ICAM-3. Другая B лимфобластоидная клеточная линия, SLA, использует как ICAM-1, так и ICAM-3, так как адгезия не ингибирует ни ICAM-1, ни ICAM-3 MAb отдельно и почти полностью этими MAb вместе. Тимомная клеточная линия, Ju, использует как ICAM-2, так и ICAM-3 пути адгезии с небольшим вкладом ICAM-1. Адгезия как покоящихся T лимфоцитов, так и активированных фитогемагглютинином (РНА), также была исследована (фиг. 1B). Как показано ранее, покоящиеся лимфоциты эффективно связываются с очищенными LFA-1 (Dustin et al., Nature, 341: 619-624 (1989)) и, таким образом, было найдено, что такое связывание в значительной степени зависит от ICAM-3. После активации PHA ICAM-1 экспрессию индуцируют и адгезия к LFA-1 наблюдается главным образом за счет ICAM-1 и частично за счет ICAM-3. ICAM-2 компонента адгезии как в покоящихся, так и в активированных T клетках была детектируема, хотя и маскировалась наличием ICAM-1 и ICAM-3. Существует значительный излишек в использовании ICAM, так как для каждой клетки MAb, к по крайней мере двум ICAM, необходимы для достижения существенного ингибирования. Заметным исключением является адгезия покоящихся лимфоцитов к LFA-1, что в значительной степени происходит за счет ICAM-3. Во всех случаях комбинация всех трех анти-ICAM-MAb исключает связывание с LFA-1 до уровней, сравнимых с теми, которые наблюдаются в присутствии анти-LFA-1 MAb.
Относительное сродство трех ICAM к LFA-1 можно исследовать, сравнивая их вклады в связывание LFA-1, как определено ранее для их экспрессии клеточной поверхностью по данным иммунофлуоресценции цитометрии в потоке (фиг. 2). Сравнение поверхностной экспрессии CAM и их вклад в LFA-1 адгезию показывает, что ICAM-1 обладает высоким сродством к LFA-1 и что ICAM-2 и ICAM-3 обладают аналогичным, но более низким сродством. Так, например, Gurkat клетки экспрессируют ICAM-2 и ICAM-3 в аналогичных количествах и каждый вносит вклад в связывание с LFA-1. Там, где ICAM-2 экспрессия больше, чем экспрессия ICAM-3, как, например, для клеток Y, путь ICAM-2 LFA-1 адгезии превалирует над ICAM- 3. Напротив, SLA и SKW3 экспрессируют в 3-5 раз больше ICAM-3 нежели ICAM-2 и, что не удивительно, ICAM-3 адгезию преимущественно по сравнению с ICAM-2 направлением. Для покоящихся T лимфоцитов, где ICAM-3 хорошо экспрессируется, а ICAM-1 отсутствует, адгезия к LFA-1 осуществляется главным образом за счет ICAM-3. Если ICAM-1 хорошо экспрессируется, а также в случае с SLA, JY и PHA-активированными T клетками, это представляет основной путь адгезии. ***
Характер распределения ICAM-3 отличается от ICAM-1 и ICAM-2 несколькими путями. В отличие от ICAM-1 и ICAM-2, ICAM-3 не экспрессирует ни на копящихся, ни на стимулированных поверхностях эндотелия (не показано). Это находится в согласии с тем фактом, что LFA-A-1-зависимое связывание клеток как с покоящимися, так и со стимулированными клетками эндотелия оказывается ICAM-1 и ICAM-2-зависимым явлением. ICAM-3 ограничен линией кроветворения и в значительных количествах экспрессируется на лимфоидных и моноцитных клеточных линиях за редким исключением. Однако во всех случаях экспрессия ICAM-3 на клеточных линиях координируется с LFA-1-зависимой, ICAM-1, ICAM-2-независимым характером адгезии. Клеточные линии (HUVEC, Razi), связывающие LFA-1 исключительно за счет ICAM-1 и ICAM-2, не демонстрируют CAM-3 экспрессии, хотя клеточные линии (JY, U 937, Sup T), которые демонстрируют слабое связывание за счет третьего типа адгезии, обладают соответственно слабой ICAM-3 поверхностной экспрессией (данные не представлены).
ICAM-3 заметно отличается от ICAM-1 и ICAM-2 по экспрессии лейкоцитов (фиг. 2B). ICAM-3 экспрессируется с высокими уровнями на покоящихся лимфоцитах, моноцитах и нейтрофилах, тогда как ICAM-1 и ICAM-2 экспрессируются в гораздо меньшей степени или вовсе отсутствуют. Для сравнения, ICAM-1 и ICAM-2 слабо экспрессировались на моноцитах и только ICAM-2 присутствовали на покоящихся лимфоцитах. Ни ICAM-1, ни ICAM-2 не были экспрессированы на нейтрофилах. После активации лимфоцитов PHA ICAM-3 экспрессия повышается в 2-3 раза, тогда как экспрессия ICAM-1 заметно снижается (Dustin et al., J. Imnugnol. 137: 245-254 (1986)) (фиг. 2В).
Иммуноосажденные ICAM-3 из различных 125I - меченых клеточных линий выявляют резкую линию 124000 Мr в восстанавливающих условиях лишь с легким повышением подвижности в невосстанавливающих условиях (фиг. 3А). Обработка N-гликаназой приводит к уменьшению ICAM-3 полосы до Мr 87000, что указывает на то, что ICAM-3 подобно ICAM-1 и ICAM-2 является сильно гликозилированным протеином (фиг. 3В). Биохимические характеристики, типы экспрессии и функциональные характеристики ICAM-3 отличаются от ранее описанных адгезионных молекул, включая хоминг-рецептор человека LAM-1 (Tedder et al., J. Immynol. 144: 532-540 (1990)), индуцируемую эндотелиальную адгезионную молекулу VACAM-1 (Rice et al., J. Exp. Med. 171: 1369-1374 (1990); Carlos et al, Blood в печати (1990); Osborn et al., Cell 59: 1203-1211 (1989)) и VLA семейство матричных рецепторов (Hemler, M. E. Clin Rw. Immunol., 8:365-400 (1990)), и не было обнаружено MAb с аналогичным распределением клеток в четырех работах по данным о лейкоцитах (Gilks. W.R., et al., Leykocyte, Typing Data Base IV, Oxford. University - Ress., Oxford, England, (1990)).
Существование трех LFA-1 лигандов предполагает специализацию для различных аспектов LFA-1 - зависимых взаимодействий лейкоцитов ICAM-1 в основном экспрессируется на эндотелиальных и многих эпителиальных типах клеток и сильно индуцируется при воспалениях и иммунных заболеваниях, когда он регулирует клеточную локализацию и облегчает распознавание специфических антигенов (Wawryk et al., Immunol. Rw, 108: 135-161 (1989)). Так как ICAM-2 является преимущественно LFA-1 лигандом на покоящихся эндотелиальных клетках, его тип адгезии может иметь важные последствия для нормальной рециркуляции LFA-1 - несущих лимфоцитах за счет тканей эндотелия (Hamann et.al., J. Immunol, 140: 693-699 (1988); Mackay et al., J. Exp. Med., 171: 810-817 (1990); Pals et al., J. Immunol. 140: 1851-1853 (1988)) и Nuroi et al., Hum. Path. 19: 753-759 (1988)). В настоящее время функции ICAM-3 на нейтрофилах остаются неясными, так как гомотипичная аггрегация нейтрофилов по-видимому в первую очередь зависит от Mac-1, независимо от присутствия LFA-1 (Anderson et al., J. Immunol. 137: 15-27 (1986); Patarroyo et al., Scant. J. Immunol. 22: 619-631 (1985)). Обнаружение того факта, что адгезия покоящихся Т лимфоцитов к LFA-1 происходит прежде всего за счет ICAM-3 наряду с тем фактом, что CAM-3 гораздо лучше экспрессируется, нежели другие LFA-1 лиганды на моноцитах и покоящихся лимфоцитах, играет важную роль в инициировании иммунной реакции. Действительно адгезия Т клеток к антиген-представляющим клеткам требует LFA-1: ICAM взаимодействий (Draunsfield et., al., Imm. Rw. 114: 29-44 (1990); Makgoba et al., Immunol. Today, 10: 417-422 (1989)) и, как таковая, ICAM-3 может играет роль, особенно на покоящихся Т лимфоцитах, где ни ICAM-1, ни ICAM-2 не экспрессируются достаточно хорошо. Действительно, роль, предложенная для LFA-1 лиганда (лигандов), отличается от роли ICAM-1 как в аллогенных, так и в автологусных смешанных лимфоцитных реакциях (Bagnasco et al. , Cell Immunol, 128: 362-369 (1990)). Более того, ICAM-3, как можно предвидеть, играет важную роль в антиген-специфических взаимодействиях между Т и В лимфоцитами, где одна из этих клеток еще не активирована. ICAM-3 может также играть роль в лизисе некоторых мишеней Т клетками, то есть зависеть от LFA-1, но не ICAM-1 (Makgoba et al., Eur. J. Immunol. 18: 637-640 (1988)).
Существование множества ICAM играет важную роль для клинического лечения MAb до ICAM или ICAM аналогами. ICAM-1 MAb является эффективным in vivo для продления службы почечных (Cosinni et. , al., J. Immunol, 155: 4604-4612 (1990)) и сердечных (Flavin et., al., Transplant. Proc., 28: 533-534 (1991) аллотрансплантатов.
ICAM-3 MAb ингибирует ряд определенных иммунных реакций in vivo, так как они ингибируют LFA-1 - зависящие адгезивные взаимодействия отличающихся субнаборов типов клеток Y.
V. кДНК клонирование ICAM-3.
Для клонирования гена ICAM-3 можно использовать любую из множества процедур. Один из таких способов включает анализ шаттл-векторной библиотеки кДНК вставок (полученных из клеток, экспрессирующих ICAM-3) на присутствие вставки, которая содержит ICAM-3 ген. Такой анализ можно провести, трансфектируя клетки вектором, а затем анализируя на экспрессию ICAM-3.
кДНК предпочтительно идентифицируют, используя модифицированный способ Aruffo и Seed (Seed et al., Proc. Natl. Acad. Sci США 84: 3365-3369 (1987)) для идентификации лигандов адгезионных молекул. В этом способе кДНК библиотеку получают из клеток, которые экспрессируют ICAM-3 (таких как SLA, Jurkat или SKW 3 лимфобластоидных клеточных линий). Предпочтительно получать кДНК библиотеку из Т-клеток. Эту библиотеку используют для трансфекации клеток, которые нормально не экспрессируют ICAM-3 (например, Cos или HeLa клетки). Трансфектированные клетки вводят в чашки Петри, которые до этого были покрыты либо LFA-1, либо анти- ICAM-3 антителами. Трансфектированные клетки, содержащие ICAM-3 кодирующие последовательности, которые экспрессируют этот лиганд на их поверхностях, будут прикрепляться к LFA-1 или анти-ICAM-3 антителам на поверхности чашек Петри. Неприкрепившиеся клетки смывают, а прикрепившиеся клетки удаляют затем из чашек Петри и культивируют. Рекомбинантные ICAM-3 экспрессирующие последовательности в этих клетках затем удаляют и секвенируют.
Если для покрытия пластин Петри используют LFA-1, в них добавляют анти-ICAM-1 и анти-ICAM-2 специфические антитела для предотвращения адгезии ICAM-1 или ICAM-2 экспрессирующих клеток. Таким образом, адгезию ICAM-1+ или ICAM-2+ трансфактантов к покрытым LFA-1 пластинам можно ингибировать такими антителами, как RRJ/1 и анти-ICAM-1 MAb и CBR-IC2/2, и анти-ICAM-2 MAb. Связывание ICAM-3 трансфектированных клеток с LFA-1 покрытыми чашками Петри ингибируется EДТА и анти-LFA-1 MAb, но не ингибируется анти-ICAM-1 или анти-ICAM-2 MAb. Поэтому ICAM-1 или ICAM-2 экспрессирующие клетки не способны к адгезии на чашках Петри и поэтому в большинстве своем смываются вместе со всеми другими неадгезивными клетками. Это увеличивает содержание клеток, экспрессирующих ICAM-3.
Другим альтернативным способом получения последовательностей генов, кодирующих ICAM-3, является способ, хорошо известный специалистам, который легко адаптировать для получения целевого гена.
Одним из таких способов получения генной последовательности кодирующей ICAM-3 является использование олигонуклеотидного зонда для скринирования кДНК или геномной ДНК библиотеки. В этом способе ICAM-3 протеин очищают предпочтительно используя иммуноочищающие процедуры, известные специалистам, и терминальную аминокислотную последовательность определяют, используя один из известных специалистам способов. В другом варианте ICAM-3 энзимативно очищают так, что очищают пептиды Lys-C или трипсином, и определяют аминокислотную последовательность одного из внутренних фрагментов.
После того, как частичная аминокислотная последовательность определена, либо олигонуклеотидный зонд создают на основе кодона, предпочтительно предоставляемого организмом, либо дегенеративный зонд создают на основании всех возможных комбинаций кодонов или комбинаций предпочтительных кодонов, гомологичных с известными последовательностями ICAM-1 или ICAM-2 ДНК, и дегенеративный кодон используют для конструирования зондов. Затем такой зонд используют для экранирования либо геномной, либо кДНК библиотеки для последовательностей, которые гибридизуются с зондом.
Описанные выше методики либо их аналоги успешно обеспечивают клонирование генов альдегиддегидрогеназы человека (Asu, L.C. et.al., Proc. Natl. Acad. Sci США 82: 2771-3775 (1985)), фибронектина (Suruki, S.et al., Eur. Mol. Biol. Organ J. 4: 2519-2524 (1995)), ген рецептора эстрогена человека (Walter, P. et al., Proc. Natl. Acad. Sci, USA 82: 7889-7893 (1985)) и плазминогенный активатор типа тканей (Pennica, D et al. Nature 301: 214-221 (1983)).
В еще одном альтернативном способе клонирования ICAM-3 гена приготавливают экспрессионную библиотеку путем клонирования геномной ДНК или более предпочтительно кДНК из клеток, которые экспрессируют ICAM-3. Затем эту библиотеку скринируют на предмет членов, способных экспрессировать протеин, который связывается с анти-ICAM-3 антителами.
Клонирование ICAM-3 гена, полученного за счет использования любого из описанных способов, можно операбельно связать с вектором экспрессии и ввести в бактериальные или эукариотные клетки для продуцирования CAM-3 протеина. Способы таких манипуляций раскрыты у Maniatis, T. et al. ранее и хорошо известны специалистам.
В альтернативном способе, в котором используют PCR для клонирования в ICAM-3 ген, сделали предположение, что ICAM-3 гомологичен ICAM-1 и/или ICAM-2. Дегенеративные олигонуклеотидные праймеры, основанные на последовательностях заключенных между ICAM-1 и ICAM-2, используют для амплификации полимеразной цепной реакцией кДНК или мРНК из клеток, которые, как известно, экспрессируют ICAM-3 протеин, таких как SKW3 или миндалины (de Sougerolles et al. , J. Exp.Med 175: 185 - 190 (199)). Клоны секвенируют и те, которые отличаются от ICAM-1 и ICAM-2, используют для получения клонов кДНК полной длины.
В любом из описанных ранее способов аутентичность клонов можно подтвердить, экспрессируя клоны полной длины, например в COS клетках, и тестируя по поводу реакционной способности ICAM-3 MAG.
VI. Агенты настоящего изобретения: ICAM-3 и их функциональные производные, агонисты и антагонисты.
Далее настоящее изобретение направлено на ICAM-3, его "функциональные производные", его "агонисты" и "антагонисты".
А. Функциональные производные ICAM-3.
"Функциональным производным" ICAM-3 является соединение, которое обладает биологической активностью (либо функциональной, либо структурной), то есть практически аналогично биологической активности ICAM-3. Термин "функциональное производное" включает "фрагменты", "варианты" и "химические производные" родственных ICAM-3 молекул.
Термин "фрагмент" ICAM-3 означает любой полипептидный субсет молекулы. Фрагмент ICAM-3, который обладает ICAM-3 активностью и который растворим (то есть не связан с мембраной), наиболее предпочтителен. Растворимый фрагмент предпочтительно создают в результате делеции участка мембраны родственной молекулы или за счет делеции или замены гидрофильных аминокислотных остатков на гидрофобные остатки. Идентификация таких остатков хорошо известна специалистам.
Фрагменты, содержащие любое число полных Ig - подобных доменов, предпочтительны, например домен 1(Д1), Д1 и Д2, Д1-3, Д1-4 и Д1-5.
Под "вариантом" ICAM-3 подразумевают практически аналогичную по структуре и функциям либо целой молекулы, либо ее фрагменту.
Говорят, что молекула "практически аналогична" другой молекуле, если обе молекулы имеют практически аналогичную структуру или обе молекулы обладают аналогичной биологической активностью. Таким образом, при условии, что две молекулы обладают аналогичной активностью, их называют вариантами в том смысле, как используют этот термин здесь, даже если одна из этих молекул обладает структурой, который нет в другой молекуле или если последовательности аминокислотных остатков не идентичны.
Как здесь использовано, говорят, что молекула является "химическим производным" другой молекулы, если она содержит дополнительные химические фрагменты, которые не являются обычно частью нативных молекул. Такие фрагменты могут усовершенствовать растворимость молекул, абсорбцию, биологический срок полужизни и т.д. Фрагменты могут альтернативно снижать токсичность молекул или исключать или ослаблять любые нежелательные побочные эффекты молекул. Фрагменты, способные осуществлять такие эффекты, раскрыты в Remington's Pharmaceutical Sciences (1980).
"Токсин-производные" молекул составляют специальный класс "химических производных". "Токсин-производные" молекулы (такие как ICAM-3 или анти-ICAM-3 антитела), которые ковалентно связаны с фрагментом токсина. Процедура присоединения таких фрагментов к молекуле хорошо известна специалистам.
Связывание таких молекул с клеткой приводит фрагмент токсина в тесную близость к клетке и в результате приводит к гибели клеток. Можно использовать любой подходящий фрагмент токсина, однако предпочтительно использовать токсин сам по себе, например токсин рицина, токсин холеры, токсин дифтерии, радиоизотопные токсины или токсины, образующие каналы в мембранах.
Функциональные производные ICAM-3, содержащие вплоть до около 100 остатков, можно легко получить в результате in vitro синтеза. При желании такие фрагменты можно модифицировать, используя известные специалистам способы, подвергая взаимодействию мишеневые аминокислотные остатки очищенных или неочищенных протеинов с органическими агентами дериватизации, которые способны реагировать с выбранными боковыми цепями или концевыми остатками. Полученные ковалентные производные можно использовать для идентификации остатков, важных для биологической активности.
Многие способы можно использовать для создания и выделения фрагментов ICAM-3. В результате дериватизации бифункциональными агентами можно получить ICAM-3, сшитые с нерастворимой в воде матрицей носителя. В другом варианте такие реакционноспособные матрицы, как цианогенбромидом активированные углеводороды и реакционноспособные субстраты, описанные в патентах США 3 969 287, 3 691 016, 4 195 128, 4 247 642, 4 229 537 и 4 330 440, используют для иммобилизации протеина.
Функциональные производные ICAM-3, имеющие чередующиеся аминокислотные последовательности, также можно получить в результате мутаций ДНК, кодирующих ICAM-3. Такие функциональные производные включают, например, делеции, вставки или замены аминокислотных остатков внутри аминокислотной последовательности ICAM-3. Любые сочетания делеций, вставок и замен можно использовать для создания окончательной конструкции при условии, что конечная структура обладает нужной активностью. Очевидно, что мутации, которые могут быть осуществлены в ДНК, кодирующей функциональное производное, не должны выводить последовательность из считывающей рамки и предпочтительно не должны создавать комплементарные участки, которые могут продуцировать вторичные мРНК структуры (см. EP патентную заявку 75 444).
На генетическом уровне функциональные производные можно получить направленным мутагенезом ДНК кодирующей ICAM-3, получая таким образом ДНК, кодирующую функциональное производное, после чего экспрессируя ДНК в рекомбинантной клеточной культуре.
Хотя место введения вариации в аминокислотную последовательность заранее известно, мутации perse не обязательно известны заранее. Так, например, для оптимизации характера мутации в указанном месте статический мутагенез, например линкерной сканирующий мутагенез, можно проводить по целевому кодону или целевому участку для создания большого числа производных, которые можно затем экспрессировать и скринировать для оптимальной комбинации целевой активности.
Способы создания мутаций в заранее определенном участке ДНК известной последовательности хорошо известны, например таким способом является направленный мутагенез.
Препараты функциональных производных ICAM-3 в соответствии с изложенным предпочтительно получают в результате направленного мутагенеза ДНК, которая кодирует ICAM-3, или ранее полученного функционального производного ICAM-3. Способы такого сайт-специфического мутагенеза хорошо известны специалистам и изложены, например, в таких публикациях, как Maniatis, T. et al., 14: Molecular Cloning, а Laborafory-Manual Caldspring Harbor, NY (1982), раскрытие которой включено по ссылке. Направленный мутагенез позволяет получать ICAM-3 функциональные производные за счет использования специфических олигонуклеитидных последователей, которые кодируют ДНК последовательность целевой мутации.
Можно осуществить аминокислотные делеции, доставки или замены, используя сайт-направленный мутагенез. Делеции аминокислотных последовательностей обычно охватывают от 1 до 30 остатков, более предпочтительно от 1 до 10 остатков, которые обычно бывают соседними. Наиболее предпочтительными делециями являются такие, которые осуществляют для создания растворимых форм ICAM-3. Растворимые формы ICAM-3 наиболее предпочтительно создаются за счет делеции либо мембранного участка ICAM-3, либо гидрофобных остатков внутри ICAM-3.
Аминокислотные вставки включают вставки единичных или множественных остатков внутри ICAM-3 кодирующих последовательностей, а также терминальных слияний с полипептидами практически неограниченной длины. Вставки внутри последовательности (то есть вставки внутри полной последовательности молекулы ICAM-3) могут составлять обычно от около 1 до 10 остатков, более предпочтительно от 1 до 5. Примеры терминальных вставок включают слияние сигнальной последовательности (независимо гетерологической или гомологической) с клеткой хозяина, с N - концов молекулы для облегчения секреции производного из рекомбинантного хозяина.
Третью группу функциональных производных представляют те, в которых по одному или более из аминокислотных остатков в молекуле ICAM-3 удалено и различные остатки вставлены на их место. Такие замены предпочтительно осуществляют в соответствии с таблицей 1, если необходимо слегка изменить характеристики молекулы ICAM-3.
Замещения, изменяющие функциональные или иммунологические идентичности осуществляют, выбирая заместители, которые менее консервативны, нежели те, которые приведены в таблице 1, то есть выбирая остатки, которые отличаются более заметно по их влиянию на сохранение: (a) структуры полипептидной основной цепи в области замещения, например, как плоской или спиральной конформации; (b) заряда или гидрофобности молекулы в мишеневом участке; или (c) объема боковой цепи. Замещения, которые вообще менее консервативны, представляют собой те, в которых: (a) глицин и/или пролин замещен другой аминокислотой или исключен, или вставлен; (b) гидрофильный остаток замещен на гидрофобный остаток; (c) цистеиновый остаток замещен на другой остаток; (d) остаток, содержащий электроположительную боковую цепь, замещен на остаток, имеющий электроотрицательный заряд; или (e) остаток, имеющий объемную боковую цепь, замещен таким, у которого нет такой боковой цепи.
Более предпочтительными являются замещения, которые влияют на растворимость ICAM-3. Такие наиболее предпочтительно создаются за счет замены гидрофобных аминокислот на гидрофильные аминокислоты.
Мутации, предназначенные для повышения средства ICAM-3, можно осуществить за счет введения аминокислотных остатков, которые представляют гомологичные положения в ICAM-1 или ICAM-2. Аналогично, такие мутанты ICAM-3 молекул можно получить без n-связанного СНО в гомологическом положении в ICAM-1 или ICAM-2.
Трудно предсказать точно, какое действие, какое-либо конкретное замещение, делеция или вставка окажут влияние на биологическую активность ICAM-3 прежде, чем это будет проделано. Однако специалистам должно быть ясно, что этот эффект можно оценить в рутинном анализе скринирования. Так, например, обычно производное получают линкерным сканированием сайт-направленного мутагенеза ДНК, кодирующей нативную ICAM-3 молекулу. Затем производное экспрессируют в рекомбинантного хозяина и необязательно выделяют из клеточной культуры, например с помощью иммуноаффинной хроматографии.
Затем активности клеточного лизата или очищенного производного скринируют в подходящем анализе скринирования по целевым характеристикам. Так, например, изменение иммунологического характера функционального производного, например аффинности данного антитела, определяют с помощью конкурирующего типа иммуноанализа. Изменения в иммуномодуляторной активности определяют соответствующим анализом. Модификация таких протеиновых характеристик, как окисление-восстановление или термостабильность, биологический срок полужизни, гидрофобность, подверженность протеолитическому разложению или тенденция к аггрегации с носителями или в мультимеры, анализируют способами, известными специалистам.
Б. Агонисты и антагонисты ICAM-3.
"Агонистом" ICAM-3 является соединение, которое повышает или понижает способность ICAM-3 выполнять какую-либо из его биологических функций. Примером такого агониста является агент, который повышает способность ICAM-3 связываться с клеточным рецептором.
"Антагонистом" ICAM-3 является соединение, которое уменьшает или предотвращает способность ICAM-3 осуществлять какую-либо из его биологических функцией. Примеры таких антагонистов включают ICAM-1 и ICAM-2, функциональные производные ICAM-1 и ICAM-2, анти-ICAM-3 антитела, анти-LFA-1 и антитела и т.д.
Клеточный аггрегационный анализ, описанный в патентных заявках США N 07/045963, 07/115798, 07/155943, 07/189915 или 07/250446 (причем все заявки включены по ссылке), способен определять LFA-1 зависимую аггрегацию и, таким образом, может быть использован для идентификации агентов, которые влияют на степень ICAM-3/LFA-1 аггрегации. Таким образом, такой анализ может быть использован для идентификации агонистов и антагонистов ICAM-3. Антагонисты могут действовать, придавая способность LFA-1 или ICAM-3 реализовать аггрегацию. Кроме того, могут быть исследованы и неиммуноглобулиновые (то есть химические) агенты с применением вышеуказанного анализа для определения того факта, являются ли они агонистами или антагонистами ICAM-3/LFA-1, осуществляющими аггрегацию. Такой аггрегационный анализ обычно осуществляют, используя периферические T-клетки, стимулированные PMA. Так связывание периферических кровяных клеток с LFA-1 используют для анализа ICAM-3, осуществляющих аггрегацию.
С. Анти-ICAM-3 антитела.
Предпочтительный иммуноглобулиновый антагонист настоящего изобретения представляет антитела к ICAM-3, например CBR-IC3/1, раскрытые здесь. Здесь подходящие анти-ICAM-3 антитела можно получить любым из известных способов.
Антитела к ICAM-3 (или функциональные производные ICAM-3) могут быть либо поликлональными, либо моноклональными. Кроме того, антитела рассматриваемой заявки могут быть гуманизированы с помощью процедур, известных специалистам, или процедур, описанных в PCT заявках PCT/US 91/02942 и PCT/US 91/02946, поданных в приемный оффис США 22 апреля 1991 г.
Анти-ICAM-3 антитела можно получить вводя ICAM-3 или его пептидные фрагменты соответствующим животным. Иммунизованные животные продуцируют поликлональные антитела в ответ на иммунизацию. Использование пептидных фрагментов ICAM-3 позволяет получать участки специфических антител, которые реактивны только с эпитопом (эпитопами), содержащимися в пептидных фрагментах, использованных при иммунизации животных.
В других вариантах анти-ICAM-3 антитела можно получить, используя ICAM-3, которая нативно экспрессирована на поверхности лимфоцитов. Интродукция таких клеток в соответствующие животные, например внутрибрюшинной инъекцией, приводит к продуцированию антител, способных связываться с ICAM-3 или членами CD-18 семейства молекул. При желании сыворотку таких животных можно отобрать и использовать в качестве источника поликлональных антител, способных связывать эти молекулы.
В другом варианте анти-ICAM-3 антитела можно продуцировать, адаптируя способ Selden R.F. (Европейская патентная заявка N 289 034) или Selden R.F. et al. (Science 236: 714-718 (1987)). Более конкретно, клетки подходящего животного (например, мыши) трансфектируют вектором, способным экспрессировать либо интактную ICAM-3 молекулу, либо ее фрагмент. Продуцирование ICAM-3 в трансфектированных клетках животного вызывает иммунную реакцию у животного и приводит к выработке анти-ICAM-3 антител этим животным.
Предпочтительно, однако, удалять спленоциты из животных, иммунизованных любым из описанных выше способов, для создания моноклональных антител, способных связывать ICAM-3.
Гибридные клетки, полученные описанным выше способом, можно скринировать различными способами для идентификации гибридомных клеток, которые секретируют антитела, способные связываться с ICAM-3. В предпочтительном анализе скринирования такие молекулы идентифицируют по их способности ингибировать аггрегацию ICAM-3 -экспрессирующих, ICAM-1 и ICAM-2 неэкспрессирующих клеток. Антитела, способные ингибировать такую аггрегацию, затем скринируют для определения того, могут ли они ингибировать такую аггрегацию за счет связывания с ICAM-3 или с членами CD-18 семейства молекул. Для такого скринирования можно использовать любые средства, способные различать ICAM-3 от семейства молекул CD-18. Так, например, антиген, связанный с антителом, можно проанализировать как иммуноосаждением, так и полиакриламидным гель-электрофорезом. Имеется возможность провести различие между теми антителами, которые связываются с членами CD-18 семейства молекул, и теми, которые связывают ICAM-3 за счет скринирования способности антител связываться с клетками, которые экспрессируют LFA-1, но не ICAM-3 (или наоборот). Способность антител связываться с клетками, экспрессирующими LFA-1, но не ICAM-3, можно определить за счет обычно используемых специалистами способов. Такие средства включают иммуноанализ (особенно те, в которых используют иммунофлуоресценцию), клеточную агглютинацию, исследования связывания фильтра, осаждение антитела и т.д.
В предпочтительном варианте антитела выбирают по их способности связываться с клетками, экспресирующими ICAM-3, но не с клетками, которые не экспресcируют ICAM-3.
В дополнение к вышеописанным агонистам и антагонистам ICAM-3 можно использовать и другие агенты в соответствии со способом настоящего изобретения для лечения воспалений.
HIV инфекция, астма и т.д. включают анти-идиопатические антитела к анти-ICAM-3 антителам и рецепторные молекулы или фрагменты таких молекул, которые способны связываться с ICAM-3.
Анти-идиопатические антитела, представляющие интерес для настоящего изобретения, способны конкурентно связываться с ICAM-3 (или исключительно с ними). Такие антитела можно получить, например, вырабатывая антитела к анти-ICAM-3 антителам, а затем скринируя антитела по их способности связываться с ICAM-3 природными связывающими лигандами.
Так как молекулы семейства CD-18 способны связываться с ICAM-3, введение таких молекул (например, гетеродимеров, содержащих как альфа, так и бета субъединицы, или молекул, состоящих только из альфа или бета субъединиц, или молекул, содержащих фрагменты либо одного типа, либо обоих типов субъединиц) будет конкурировать с клетками для связывания с ICAM-3, присутствующими на клетке.
Анти-аггрегационные антитела настоящего изобретения можно идентифицировать и титровать любым из множества способов. Так, например, можно измерять способность антител к дифференциальному связыванию с клетками, которые экспрессируют ICAM-3 (например, T-клетки), и по их неспособности связываться с клетками, которые не могут экспрессировать ICAM-3. Подходящие анализы клеточной аггрегации описаны в патентных заявках США NN 07/045963, 07/115798, 07/155943, 07/189815 или 07/250446 (все они включены сюда по ссылке). В другом варианте можно измерять способность антител связываться с ICAM-3 или пептидным фрагментом ICAM-3. Как хорошо понятно специалистам, вышеуказанные анализы можно модифицировать или осуществлять в другом порядке для получения различных потенциальных анализов скринирования, каждый из которых способен идентифицировать и проводить различия между антителами, способными связываться с ICAM-3 и членами CD-18 семейства молекул.
Д. Получение агентов настоящего изобретения.
Агенты настоящего изобретения можно получить: в природных процессах (например, включающих животных, растения, грибки, бактерии и т.д. для получения неиммуноглобулинового антагониста ICAM-3, или индуцируя животных для продуцирования поликлональных антител, способных связываться с ICAM-3); синтетическими способами (например, синтезируя ICAM-3, функциональные производные ICAM-3 или протеиновые антагонисты ICAM-3 (либо иммуноглобулин, либо неиммуноглобулин)); с помощью гибридомной технологии (например, для получения моноклональных антител, способных связываться с ICAM-3) или с помощью рекомбинантной технологии (например, для получения агентов настоящего изобретения в различных хозяйствах), например: в дрожжах, бактериях, грибках, культивируемых клетках млекопитающих и т.д., используя рекомбинантные плазмиды или вирусные векторы. Выбор используемого способа зависит от таких факторов, как удобство, желательный выход и т.д. Однако нет необходимости использовать только один из вышеописанных способов, процессов или технологий для продуцирования конкретного противовоспалительного агента; вышеуказанные процессы, способы и технологии можно сочетать для получения конкретного агента.
VII. Применение ICAM-3 и их функциональных производных агонистов и антагонистов.
Агенты настоящего изобретения можно использовать для осуществления различных биологических функций, которые осуществляются с помощью ICAM-3.
А. Супрессия воспалительных процессов.
Один из аспектов настоящего изобретения проистекает из способности ICAM-3 и их функциональных производных взаимодействовать с рецепторами CD-18 семейства молекул, особенно LFA-1. За счет способности ICAM-3 взаимодействовать с членами семейства CD-18 гликопротеинов их можно использовать для супрессии (то есть для предотвращения или ослабления) воспалений.
Термин "воспаление" в том смысле, как здесь использован, означает индуцирование как реакций специфической защитной системы, так и реакций неспецифической защитной системы.
В том смысле, как здесь использован термин "специфическая защитная система", он относится к той компоненте иммунной системы, которая реагирует на присутствие антигенов. Говорят, что воспаление возникает как реакция специфической защитной системы, если воспаление вызвано, опосредствовано или связано с реакцией специфической защитной системы. Примеры воспалений, являющихся результатом реакций специфический защитной системы, включают реакции на такие антигены, как вирус краснухи, автоиммунные заболевания и замедленного типа реакция гиперчувствительности, связанная с T-клетками (что видно, например, у индивидуумов с "положительным" тестом Манту). Хронические воспалительные заболевания и отторжение твердых трансплантированных тканей и органов, например почек и трансплантатов костного мозга, - это дальнейшие примеры воспалительных реакций специфической защитной системы.
Как использовано здесь, реакция "неспецифической защитной системы" должна означать реакцию, осуществляемую за счет лейкоцитов, которые не способны к иммунологической памяти. Такие клетки включают гранулоциты и макрофаги. Как здесь использовано, говорят, что воспаление является результатом реакции неспецифической защитной системы, если воспаление вызвано или опосредовано, или связано с реакцией неспецифической защитной системы. Примеры воспалений, которые происходит в результате по крайней мере частично за счет реакции неспецифической защитной системы, включают воспаления, связанные с такими состояниями, как синдром расстройств респираторной системы у взрослых (ARDS) или синдрома множественного поражения органов, вторичные после сепсиса, травм или кровотечений; реперфузионные поражения тканей миокарда или других тканей; острый гломерулонефрит; реактивные артриты; дерматиты с острыми воспалительными компонентами; острые пурулентные менингиты или другие воспалительные расстройства центральной нервной системы, например удар, ожог, гемодиализ, лейкаферез, язвенные колиты; болезнь Крона, некротизация энтероколитов; синдром, связанный с трансфузией гранулоцитов; токсикоз, вызванный цитокином.
Как обсуждалось ранее, связывание ICAM-3 молекул с мембранами CD-18 семейства молекул играет главную роль в клеточной адгезии.
За счет процесса адгезии лимфоциты способны непрерывно контролировать животное на предмет присутствия чужеродных антигенов. Хотя эти процессы обычно желательны, они также вызывают отторжение трансплантатов твердых органов (например, почек), отторжение трансплантатов нетвердых органов (например, костного мозга), отторжение трансплантатов тканей и многие автоиммунные заболевания. Следовательно, любые средства, которые способны ослабить или ингибировать клеточную адгезию, были бы весьма желательны для пациентов с трансплантатами твердых органов (особенно трансплантатами почек), трансплантатами нетвердых органов (особенно с трансплантатами костного мозга), трансплантатами тканей или для пациентов с автоиммунными заболеваниями.
Моноклональные антитела к членам CD-18 семейства ингибируют многие функции, зависящие от адгезии лейкоцитов, включая связывание с эндотелием (Haskarol, D. et al. , J. Immunol. 137: 2901-2906 (1988)), гомотипические адгезии (Rothlein, R., et al., J. Exp. Mad 163: 1132-1149 (1986)), индуцируемую антигеном и митогеном пролиферацию лимфоцитов (Davignon, D. et al., Proc. Natl. Acad. Sci, USA 78: 3435-4539 (1981)), образование антител (Fischer. A. et al., J. Immunol. 136: 3198-3203 (1986)) и эффекторные функции всех лейкоцитов, такие как литическая активность питотоксических T клеток (Krensky, A.M.,: et al., J. Immunol. 132: 2180-2181 (1984)), макрофагов (Strassman, I. et al. , J. Immunol. 136: 4328-4333 (1986)) и всех клеток, вовлеченных в зависящие от антител клеточные цитотоксические реакции (Kohl. S. et al., J. Immunol 133: 2972-2978 (1984)). Во всех вышеуказанных функциях антитела ингибируют способность лейкоцитов к адгезии к соответствующему клеточному субстрату, что, в свою очередь, ингибирует биологические функции, связанные со связыванием. Такие функции в той степени, в которой они включают ICAM-3/LFA-1 взаимодействия, могут быть подавлены анти-ICAM-3 антителами. Таким образом, моноклональные антитела, способные связываться с ICAM-3, можно использовать в качестве противовоспалительных агентов для млекопитающих. Существенно, что такие агенты отличаются от обычных противовоспалительных агентов тем, что они способны селективно ингибировать адгезию и не вызывают другие побочные эффекты, такие как нефротоксичность, которые сопровождают обычные агенты.
Так как ICAM, особенно в растворимой форме, способны действовать таким же образом, как и антитела к членам семейства CD-18, их можно использовать для супрессии воспалений. Более того, функциональные производные и антагонисты ICAM-3 можно также использовать для подавления воспалительных процессов.
1. Супрессия реакций гиперчувствительности замедленного типа.
ICAM-3 осуществляет частично адгезионные акты, необходимые для создания воспалительных реакций, таких как реакции гиперчувствительности замедленного типа. Так антитела (особенно моноклональные антитела), способные связываться с ICAM-3, обладают терапевтическим потенциалом по ослаблению или исключению таких реакций.
В другом варианте, так как ICAM-3 является антагонистом взаимодействия ICAM-3/LFA-1, ICAM-3 (особенно в растворимой форме) или его функциональные производные можно использовать для супрессии реакций гиперчувствительности замедленного типа.
Такие потенциальные терапевтические применения можно использовать двояким образом. Во-первых, композиции, содержащие анти-ICAM-3 моноклональные антитела или растворимые производные ICAM-3, можно вводить пациентам с реакциями гиперчувствительности замедленного типа. Так, например, такие композиции можно представить пациентам, которые были в контакте с такими антигенами, как сумах укореняющийся и т.д. В другом варианте моноклональные антитела, способные связываться с ICAM-3, вводят пациенту вместе с антигеном для предотвращения последующем воспалительной реакции. Так дополнительное введение антигена с анти-ICAM-3 антителами может способствовать временной толерантности пациента до последующего представления антигена.
2. Лечение хронических воспалительных заболеваний.
Так как у LAD пациентов из-за отсутствия LFA-1 не возникает воспалительной реакции, как считают, антагонизм LFA-1 природных лигандов будет также ингибировать воспалительную реакцию. Способность антител против ICAM-3 ингибировать воспаление обеспечивает основу для терапевтического применения при лечении хронических воспалительных заболеваний и автоиммунных заболеваний, таких как крапивница, автоиммунные тироидиты, экспериментальные аллергические энцефаломиелиты (EAE), множественные склеродозы, некоторые формы диабетов, синдром Рейнауда, ревматоидные артриты и т.д. Такие антитела можно также использовать в качестве терапевтических агентов при лечении псориазов.
Вообще анти-ICAM-3 антитела настоящего изобретения можно вводить отдельно или в сочетании с анти-ICAM-1 и/или анти-ICAM-2 антителами при лечении таких заболеваний, которые в настоящее время лечат стероидными препаратами.
В соответствии с настоящим изобретением такие воспалительные и иммунные реакции отторжения можно подавить (например, либо предотврать, либо ослабить) за счет введения нуждающемуся в таком лечении субъекту такого количества противовоспалительного агента, которого достаточно для подавления указанного воспаления. Подходящие противовоспалительные агенты включают: антитела, способные связываться с ICAM-3; фрагменты указанных антител, причем указанные фрагменты способны связываться с ICAM-3; практически чистый ICAM-3; функциональные производные ICAM-3; неиммуноглобулиновые антагонисты ICAM-3 или неиммуноглобулиновые антагонисты ICAM-3, отличные от LFA-1. Наиболее предпочтительны противовоспалительные агенты, состоящие из растворимых функциональных производных ICAM-3. Такое противовоспалительное лечение может также включать дополнительное введение агентов, выбранных из группы, состоящей из антител, способных связываться с LFA-1; неиммуноглобулиновых антагонистов LFA-1; практически чистого ICAM-1 и/или ICAM-2, или производных, или анти-ICAM-1 и/или анти-ICAM-2 антител или их фрагментов.
Далее настоящее изобретение включает вышеуказанные способы супрессии воспалительных реакций специфической защитной системы, где иммуносупрессор дополнительно вводят субъекту. Такие агенты предпочтительно вводят в дозах, которые ниже (то есть в субоптимальных дозах), нежели те, которые необходимы обычно. Использование субоптимальных доз возможно благодаря синергическому действию агентов настоящего изобретения. Примеры подходящих иммуносупрессоров включают (но не ограничиваются ими) дексаметазон, азатиоприн, ICAM-1, ICAM-2 или пиклоспорин A.
3. Терапия неспецифических воспалений.
Многие воспалительные реакции связаны с реакциями "неспецифической защитной системы" и осуществляются за счет лейкоцитов, которые не способны к иммунологический памяти. Такие клетки включают гранулоциты и макрофаги. Как здесь использовано, говорят, что воспаления являются результатом реакции неспецифической защитной системы, если воспаление вызывается, осуществляется за счет или связано с реакцией неспецифической защитной системы. Примеры воспалений, которые приводят, по крайней мере частично, к реакции неспецифической защитной системы, включают воспаления, связанные с такими состояниями, как синдром расстройства распираторной системы у взрослых (APD) или синдромы множественного поражения органов, вторичного после сепсисов, травм и кровотечений; реперфузионных поражений тканей миокарда и других; острые гломерулонефриты; реактивные артриты; дерматозы с острыми воспалительными компонентами; острые пурулентные менингиты или другие воспалительные расстройства центральной нервной системы, такие как удар, ожог, гемодиализ, лейкаферез; язвенные колиты; болезнь Крона; некротизирующие энтероколиты; синдром, связанный с трансфузией гранулоцитов; и токсичность, вызванная цитокином.
Противовоспалительные агенты настоящего изобретения являются соединениями, способными специфически антагонизировать взаимодействие CD-18 комплекса на гранулоцитах с эндотеллиальными клетками. Такие антагонисты включают: ICAM-3; функциональные производные ICAM-3; неиммуноглобулиновые антагонисты ICAM-3, отличные от ICAM-1, ICAM-2 или членов CD-18 семейства молекул.
В. Супрессоры отторжения органов и тканей.
Так как ICAM-3, особенно в растворимой форме, способны действовать таким же образом, что и антитела к членам семейства CD-18, их можно использовать для супрессии отторжения органов или тканей, вызываемого любой из функций, зависящей от клеточной адгезии. Более того, анти-ICAM-3 антитела, функциональные производные ICAM-3 и антагонисты ICAM-3 можно использовать для супрессии таких отторжений.
ICAM-3 и анти-ICAM-3 антитела можно использовать для предотвращения отторжения твердых органов или тканей, например почек, отторжения нетвердых органов, например костного мозга, или модифицированных автоиммунных реакций у млекопитающих. Важно, что использование моноклональных антител, способных распознавать ICAM-3, может позволить осуществить трансплантацию органов даже от индивидуумов с HLA несовместимостью
С. Добавление к интродукции антигенных материалов, вводимых для терапевтических или диагностических целей.
Иммунные реакции на терапевтические или диагностические агенты, такие как, например, бычий инсулин, интерферон, тканевого типа плазминогенный активатор или мышиные моноклональные антитела, существенно придают терапевтическую или диагностическую ценность таким агентам и в некоторых случаях вызывают такие заболевания, как сывороточная болезнь. Такие ситуации можно исправить, используя антитела настоящего изобретения. В этом варианте настоящего изобретения анти-ICAM-3 антитела следует вводить в сочетании с терапевтическим и диагностическим агентом. Такое дополнение к антителам предотвращает распознавание агента реципиентом, и поэтому предотвращает инициирование иммунной реакции реципиента против них. Отсутствие такой иммунной реакции приводит к способности пациента получать дополнительные введения терапевтического или диагностического агента.
ICAM-3 (особенно в растворимой форме) или его функциональные производные можно использовать отдельно или в сочетании с ICAM-1 или ICAM-2, или с антителами, способными связываться LFA-1 при лечении заболеваний. Так в солюбилизированной форме такие молекулы можно использовать для ингибирования отторжения органа или трансплантата. ICAM-3 или его функциональные производные можно использовать таким же образом, что и анти-ICAM-3 антитела для снижения иммуногеничности терапевтических или диагностических агентов.
Д. Супрессоры опухолевых метастазов.
Агенты настоящего изобретения можно также использовать для супрессии метастазов кроветворных опухолевых клеток, которые требуют функционального члена CD-18 семейства для миграции. В соответствии с этим вариантом настоящего изобретения пациенту, нуждающемуся в таком лечении, предоставляется такое количество агента (например, антител, способных связываться с ICAM-3, токсин-производных указанного антитела; фрагментов антител, причем указанные фрагменты способны связываться с ICAM-3; токсин-производные указанного фрагмента; практически чистого ICAM-3; функциональных производных ICAM-3; или неиммуноглобулиновых антагонистов ICAM-3, отличных от ICAM-1 или ICAM-2), которого достаточно для супрессии указанных метастазов.
В следующем варианте настоящего изобретения предложен способ супрессии роста ICAM-3 экспрессирующих опухолевых клеток. Конкретно, указанный способ включает предоставление пациенту, нуждающемуся в лечении, такого количества агента, которого достаточно для супрессии указанного роста. Подходящие агенты включают антитела, способные связываться с ICAM-3; токсин-производными указанного антитела; фрагменты антител, причем указанные фрагменты способны связываться с ICAM-3; токсин-производные указанного фрагмента; токсин-производные члена CD-18 семейства молекул; или токсин-производные функционального производного члена CD-18 семейства молекул.
В следующем варианте настоящего изобретения предложен способ супрессии роста LFA-1 экспрессирующих опухолевых клеток. Конкретно, указанный способ включает обеспечение пациента, нуждающегося в таком лечении, таким количеством токсина, которого достаточно для супрессии указанного роста. Подходящие токсины включают токсин-производные ICAM-3 или токсин-производные функционального производного ICAM-3.
E. Супрессия HIV инфекции и предотвращение распространения HIV инфицированных клеток.
В следующем варианте настоящего изобретения предложен способ супрессии инфекции HIV. Конкретно, указанный способ включает введение HIV-инфицированному субъекту эффективного количества агента супрессии HIV-инфекции. Хотя настоящее изобретение конкретно касается способа супрессии HIV-1 инфекции, следует учитывать, что этот способ можно применять к любому HIV варианту (например, HIV-2), который может инфицировать клетки таким способом, который может быть супрессирован агентами настоящего изобретения. Такие варианты являются эквивалентами HIV-1 для целей настоящего изобретения.
Один из аспектов настоящего изобретения проистекает из понимания того факта, что экспрессия LFA-1 и в некоторых случаях LFA-1 связывающих лигандов, стимулированных HIV инфекцией, промотирует реакции адгезии клетка-клетка, которые могут повысить время контакта инфицированных и неинфицированных клеток, облегчая перенос вируса от инфицированных к неинфицированным клеткам. Так агенты настоящего изобретения, которые способны модулировать LFA-1 (лиганд взаимодействия), способны подавлять инфекцию HIV и, в частности, HIV-1.
Следует понять, что за счет каких молекул, ингибирующих взаимодействия LFA-1 /лиганд, можно подавить HIV инфекцию за счет придания способности LFA-1 лиганду, экспрессируемому HIV-инфицированными клетками, связываться с CD11/CD18 рецепторами здоровых T клеток. В другом варианте молекулы, которые ингибируют LFA-1 (лиганд взаимодействия), могут придавать способность CD11/CD18 рецепторам, экспрессированным HIV-инфицированными клетками, связываться с LFA-1 здоровых T клеток. Для придания способности клеткам связываться с CD11a/CD18 рецептором или LFA-1 связывающего лиганда имеется возможность использовать ICAM-3, фрагмент ICAM-3, функциональное производное ICAM-3 или анти-ICAM-3 антитела.
Агенты изобретения предназначены для того, чтобы обеспечить реципиентов агентами в количествах, достаточных для достижения супрессии HIV- инфекции. Говорят, что количество достаточно для "супрессии" HIV-инфекции, если дозы, способ введение и т.д. агента достаточны для ослабления или предотвращения такой HIV инфекции. Агенты должны быть предложены пациентам, которые экспонированы или подвержены HIV инфекции.
Агенты изобретения могут быть либо "профилактическими", либо "терапевтическими" при лечении HIV инфекций. Если предполагается профилактика, агент предоставляют до того, как появляются симптомы вирусного заболевания (например, до, во время или сразу после времени такой инфекции, но до появления симптомов такой инфекции). Профилактическое введение агента служит для предотвращения или ослабления любой последующей HIV инфекции. Если агент используют в терапевтических целях, его предоставляют при определении (или сразу после) инфицированных вирусом клеток. Терапевтическое введение агента служит для ослабления любой дополнительной HIV инфекции.
Агенты изобретения могут, таким образом, быть введены либо до установления вирусной инфекции (для того, чтобы подавить нежелательную HIV инфекцию), или после реального определения таких инфицированных вирусом клеток (для подавления дальнейшей инфекции).
В следующем варианте изобретение предлагает улучшенную терапию для AIDS и улучшает средства для подавления HIV инфекции и, особенно, HIV-1 инфекции, которая включает совместное введение:
(I) ICAM-3, растворимого производного ICAM-3, CD11 (либо CD11a, CD11b или CD11c), растворимого производного CD11, CD18, растворимого производного CD18 или CD11/CD18 гетеродимера, или растворимого производного CD11/CD18 гетеродимера и/или
(II) антитела, способного связываться с ICAM-3 с
(III) клетками или частицами, связанными с CD-4 или растворимым производным CD-4 и/или
(IV) молекулами (предпочтительно, антителами или фрагментами антител), способными связываться с CD-4.
В следующем варианте настоящего изобретения предложен способ супрессии миграции HIV инфицированных клеток. Более конкретно, указанный способ включает введение эффективного количества анти-миграционного агента HIV - инфицированному субъекту.
Анти-миграционные агенты настоящего изобретения включают ICAM-3, фрагмент ICAM-3, функциональное производное ICAM-3 или анти-ICAM-3 антитела, которые способны придавать способность инфицированным HIV T-клеткам связываться с LFA-1 лигандом. Антитела, которые связываются с ICAM-3, будут подавлять миграцию, придавая способность ICAM-3 экспрессированным HIV - инфицированным T клеткам, способность связываться с клетками, экспрессирующими CD11/CD18 рецептор. Для придания способности клеткам связываться с CD11/CD18 рецепторам имеется возможность использовать антитела, способные связываться с ICAM-3.
Агенты настоящего изобретения предназначены для придания реципиентам агентов в количестве, достаточном для супрессии миграции HIV (или других вирусов) - инфицированных T клеток. Говорят, что количество достаточно для "супрессии" миграции T клеток, если дозы, способы введения и т.д. агента достаточны для ослабления или предотвращения такой миграции.
Введение такого соединения (соединений) может быть либо с "профилактическими", либо с "терапевтическими" целями. Если предусматривается профилактика, агенты настоящего изобретения вводят до появления каких-либо симптомов вирусной инфекции (например, до, во время или сразу после момента такой инфекции, но до проявления каких-либо симптомов такой инфекции). Профилактическое введение агентов служит для предотвращения или ослабления любой последующей миграции инфицированных вирусом T клеток. Терапевтическое введение агента служит для ослабления любой дополнительной миграции таких T клеток.
Агенты настоящего изобретения могут, таким образом, быть представлены либо до установления вирусной инфекции (для подавления нежелательной миграции инфицированных T клеток), или после реального определения таких инфицированных вирусом клеток.
Г. Лечение астмы.
В следующем варианте настоящего изобретения агент, способный модулировать LFA-1/ICAM-3 взаимодействия, используют при лечении астмы. Конкретно, указанный способ включает введение эффективного количества анти-астматического агента индивидууму, нуждающемуся в таком лечении.
Анти-астматические агенты настоящего изобретения включают ICAM-3, фрагмент ICAM-3, функциональное производное ICAM-3 или ICAM-3 антитела, которые способны придавать способность клеткам связываться с LFA-1. Антитела, которые связываются с ICAM-3 будут подавлять миграцию эозинофилов путем придания способности ICAM-3 экспрессированным на этих клетках, связываться с клетками, экспрессирующими CD11/CD18 рецептор.
Анти-астмастические агенты настоящего изобретения предназначены для предоставления реципиенту агента в количестве, достаточном для снижения или ослабления серьезности, степени или длительности симптомов астмы. Для терапевтических целей агент предоставляют во время (или сразу после) проявления симптомов астмы. Терапевтическое введение агента служит для ослабления любых острых приступов астмы. Таким образом, агенты настоящего изобретения могут вводиться либо до нежелательного приступа астмы (для ослабления серьезности приступа, длительности, степени приступа) или после начала приступа.
С. Диагностические и прогностические применения.
Моноклональные антитела, способные связываться с ICAM-3, можно использовать как средства изображения или визуализации сайтов ICAM-3 экспрессии и воспалений у пациента. При таком использовании анти- ICAM-3 моноклональные антитела метят детектируемым образом, например, используя радиоизотопы, метки сродства (например, биотин, авидин и т.д.), флуоресцентные метки или парамагнитные атомы. Процедуры введения таких меток хорошо известны специалистам. Меченые антитела можно затем использовать в получении диагностических изображений. Клиническое применение антител для получения диагностических изобретений описано в обзоре Grossman, H.B. Urol. Clin. North Amer. 13: 465-474 (1986), Unger, E.C. et al., Invest. Radiol. 20: 693-700 (1985) и Khaw, B.A. et al., Science, 209: 195-197 (1980)).
Наличие ICAM-3 - экспрессии можно также детектировать за счет использования зондов гибридизации, таких как мРНК, кДНК, геномная ДНК, или синтетического олигонуклеотидного зонда, который связывается с ICAM-3 геномной последовательностью, или с ICAM-3 мРНК последовательностью, присутствующими в клетках, которые экспрессируют ICAM-3. Методики осуществления такого гибридизационного анализа раскрыты Maniatis, T et al., Molecular Cloning, a Laboratory Manual, Coldspring Harber, Ny (1982) и Haymes, B.D. et al., Nucleic Acid Hybridisation, а Practical Approach, IRL Press, Washington, DC (1985) (которые включены сюда по ссылке)).
Определение очага ICAM-3 экспрессии с помощью использования меченных антител или зондов нуклеиновых кислот можно использовать для диагностики развития опухолей. В одном из диагностических вариантов образцы тканей или крови отбирают у субъекта и инкубируют в присутствии антител, которые детектируемо помечены или которые могут быть детектируемо помечены. В предпочтительном варианте такую методику осуществляют неинвазивным способом, используя магнитные изображения, флюорографию и т.д. Такие диагностические тесты можно использовать для контроля за реципиентами с трансплантированными органами, например почкой, для определения ранних сигналов возможного отторжения тканей. Такой анализ можно проводить в попытках определить предрасположенность индивидуумов к ревматоидным артритам или другим хроническим воспалительным заболеваниям.
Так, например, пометив радиоактивно анти-ICAM-3 антитела или фрагменты антител, можно детектировать антигены, используя радиоиммунологический анализ. Хорошее описание радиоиммунологического анализа (RIA) можно найти в Laboratory Technigues and Biochemistry in Molecular Biology by Work T.S. et al., North Holland Publishing Company N.V. (1978) и преимущественно в главе, озаглавленной "Введение в радиоиммунологический анализ и связанные с ним методики" Chard. T. (включено сюда по ссылке). В другом варианте антитела можно метить флуоресцирующим соединением, энзимом или другими подходящими метками, известными специалистам.
В дополнении к локализации сайтов воспаления, антитела, которые могут связываться с ICAM-3, можно использовать для анализа биологических жидкостей на предмет присутствия циркулирующих ICAM-3, используя любую из обычно используемых сред для анализа жидкостей. Наличие циркулирующих ICAM-3 в образце жидкости является показателем воспалительной реакции или других биологических функций, осуществляемых за счет ICAM-3. Кроме того, наличие ICAM-3 в аминотических жидкостях связано с осложнениями, возникающими при беременности. Для анализа могут быть использованы любые биологические жидкости. Однако предпочтительными биологическими жидкостями являются: кровь, сыворотка, плазма, синовиальная жидкость, спинномозговая жидкость или моча.
VIII. Введение композиций настоящего изобретения.
Терапевтический эффект ICAM-3 может быть достигнут при введении пациенту полной ICAM-3 молекулы или ее терапевтически активного пептидного фрагмента. Особый интерес представляют терапевтически активные пептидные фрагменты ICAM-3, которые растворимы.
ICAM-3 и его функциональные производные можно получить синтетически за счет использования рекомбинантной ДНК технологии, протеолизом или сочетанием этих способов. Терапевтические преимущества ICAM-3 можно усилить за счет использования функциональных производных ICAM-3, обладающих дополнительными аминокислотными остатками, которые добавляют, чтобы повысить способность присоединяться к носителю или повысить активность ICAM-3. В объем настоящего изобретения входят также функциональные производные ICAM-3, в которых отсутствуют определенные аминокислотные остатки или которые содержат измененные аминокислотные остатки, при условии, что такие производные обладают или осуществляют биологические или фармакологические функции ICAM-3.
Говорят, что как антитела настоящего изобретения, так и раскрытые здесь молекулы ICAM-3 "практически не содержат природных примесей", если препараты, которые их содержат, практически свободны от материалов, с которыми эти продукты обычно и природно находятся.
Изобретение относится также к антителам и их биологически активным фрагментам (как поликлональным, так и моноклональным), которые способны связываться с ICAM-3. Такие антитела можно получить либо из животных, либо на культурах тканей, либо с помощью рекомбинантных ДНК.
Введение ICAM-3 или молекул, полученных из ICAM-3, можно осуществить отдельно или в сочетании с ICAM-1 и/или ICAM-2. Введение анти-ICAM-3 антител или других молекул, способных связываться с ICAM-3 или с молекулами, полученными на ICAM-3, можно осуществить отдельно или в сочетании с анти-ICAM-1 антителами и/или анти- ICAM-2 антителами. При введении пациенту антител или фрагментов антител, способных связываться с ICAM-3, или вводя ICAM-3 (или фрагмент, вариант или их производные) пациенту, дозы вводимого агента должны меняться в зависимости от таких факторов как возраст пациента, вес, рост, секс, общее состояние здоровья, медицинская предыстория и т.д. Обычно желательно вводить реципиенту дозы антител, которые находятся в интервале от около 1 мг/кг до 10 мг/кг (веса пациента), хотя можно вводить и более низкие или более высокие дозы. При введении пациенту ICAM-3 молекул или их функциональных производных предпочтительно вводить такие молекулы в дозах, которые также находятся в интервале от около 1 мг/кг до 10 мг/кг (веса пациента), хотя возможно вводить и более низкие и более высокие дозы. Как будет обсуждаться далее, терапевтически эффективная доза может быть снижена, если анти-ICAM-3 антитела вводят вместе с анти- ICAM-1 антителами, анти-ICAM-1 антителами и/или анти-ICAM-2 антителами. В том смысле, как здесь использовано, говорят, что соединение вводится дополнительно с другим соединением, если введение этих двух соединений настолько близко по времени, что их можно одновременно детектировать в сыворотке пациента.
Оба антитела, способные связываться с ICAM-3, и сами ICAM-3 можно вводить пациенту внутривенно, внутримышечно, подкожно, энтерально, поверхностно или парентерально. При введении антител или ICAM-3 с помощью инъекций такое введение может быть непрерывной инъекцией или единичной или множественными дозами.
Агенты изобретения предназначены для введения реципиенту в количестве, достаточном для того, чтобы быть "физиологически эффективными". Говорят, что количество "физиологически эффективно", если доза, способ введения агента и т. п. достаточны для ослабления или предотвращения физиологических эффектов, связанных с ICAM-3. Так, например, один из агентов настоящего изобретения вводят пациенту с целью подавления воспаления и говорят, что он физиологически эффективен, если его вводят в дозе, достаточной для подавления воспаления.
Кроме того, анти-ICAM-3 антитела или фрагменты можно вводить либо отдельно, либо в сочетании с одним или более из дополнительных иммуносуппрессорных агентов (особенно, пациентам с трансплантатами органов или тканей). Введение таких соединений (или соединения) может быть как с целью "профилактики", так и с "терапевтическими" целями. В случае профилактики иммуносупрессорные соединения вводят до проявления каких-либо воспалительных реакций или симптомов (например, до, во время или сразу после момента трансплантации органа или ткани, но до появления каких-либо симптомов отторжения тканей или органа). Профилактическое введение соединения (соединений) служит для предотвращения или ослабления любых последующих воспалительных реакций (например, отторжения трансплантированных органов или тканей и т.д.). При терапевтическом введении соединение (соединения) вводят во время (или сразу после установления симптома острого воспаления, например отторжения органа или ткани). Терапевтическое введение соединения (соединений) служит для ослабления любых острых воспалений (например, отторжения трансплантированных твердых органов или тканей, например почки, или нетвердых органов, например костного мозга). Таким образом, противовоспалительные агенты настоящего изобретения можно вводить либо до установления воспаления (например, для супрессии нежелательного воспаления), либо после начала воспаления.
Говорят, что композиция "фармакологически приемлема", если пациент переносит ее введение. Когда говорят, что агент вводят в "терапевтически эффективном количестве", подразумевают, что это количество физиологически значимо. Агент является физиологически значимым, если его присутствие приводит к детектируемым изменениям физиологии реципиента-пациента.
Антитела и ICAM-3 молекулы настоящего изобретения можно сформулировать в соответствии с известными способами приготовления фармацевтически полезных композиций, где такие материалы или их функциональные производные объединяют с фармацевтически приемлемым носителем. Подходящие носители и их композиции, включая другие протеины человека, например альбумин сыворотки человека, описаны, например, в Remington's Pharmaceutical. Sciens (16th ed. Osol, A., Ed Mack. Easton PA. (1980). Для получения фармацевтически приемлемой композиции для эффективного введения такие композиции будут содержать эффективное количество агента настоящего изобретения вместе с подходящим количеством носителя. Кроме того, антитела настоящего изобретения можно гуманизировать за счет химеризации или CDR трансплантатов, чтобы они стали более "фармакологически приемлемы" пациентами. Такие способы химеризации антител, особенно анти-ICAM-1 антител, описаны в патентной заявке Великобритании N 90 09 548.0, 90 09 549.8 и PCT заявках N PCT/US 91/02942 и PCT/US 91/02946, поданных в приемный оффис США 27 апреля 1991 г. (включено сюда по ссылке).
Дополнительные фармацевтические способы можно использовать для контроля длительности действия. Препараты с контролируемым высвобождением можно получить за счет использования полимеров, которые образуют комплексы или абсорбируют агенты настоящего изобретения. Скорость и длительность контролируемого высвобождения можно регулировать в некоторой степени за счет выбора соответствующих макромолекулярных матриц, изменяя концентрации включенных макромолекул, а также за счет способов включения. Другим возможным способом контроля длительности действия за счет контролируемого высвобождения препаратов является включение агентов настоящего изобретения в частицы полимерного материала, например в полиэфиры, полиаминокислоты, гидрогели, полимеры молочной кислоты или сополимеры этилена и винилацетата. В другом варианте вместо включения этих агентов в полимерные частицы можно заключить эти материалы в микрокапсулы, полученные, например, в результате коацервации или в результате межфазной полимеризации, микрокапсулированием в желатине или поли(метилметакрилате), или в системе постановки коллоидных лекарств, например липосом, микросфер альбумина, микроэмульсий, наночастиц и нанокапсул или в макроэмульсиях.
Далее изобретение включает фармацевтическую композицию, содержащую: (a) противовоспалительный агент (например, антитела, способные связываться с ICAM-3; фрагмент указанных антител, способных связываться с ICAM-3; ICAM-3; функциональное производное ICAM-3; или неиммуноглобулиновый антагонист ICAM-3, отличный от ICAM-1 и ICAM-2, и (b) по крайней мере один иммуносупрессорный агент. Примеры подходящих иммуносупрессорных агентов включают дексаметазон, азатиоприн и циклоспорин A.
После того, как были в общем описаны изобретение, агенты и способы их получения, изобретение будет легче понять со ссылкой на следующие примеры, которые приведены лишь с целью иллюстрации и не должны лимитировать настоящее изобретение, если только нет специальных указаний.
Пример I. Характеристика ICAM-3, нового адгезионного лиганда для ICAM-1.
Адгезию клеточных линий и лимфоцитов Dustin et al., Cold spring Harbor Symp. Quant. Biol., 64: 753-765 (1989) к пластинам, покрытым LFA-1, осуществляют в соответствии с описанием. LFA-1, очищенные от JY лизатов, абсорбируют на микротитровальные пластины с 96 ячейками (Linbro-titertek) в количестве 1100 сайтов/мкм2. Неспецифические связывающие сайты блокируют 1% BSA и промывают PBS (5% FBS/2 мм MgCl/ 0,5% HSA/ среда анализа). Специфического ингибирования LFA-1 достигают за счет инкубирования в течение 30 минут при комнатной температуре микротитровальных пластин с 1/200 разбавлениями TS/22 (анти-LFA-1) асцитами. Покоящиеся T клетки выделяют из целой крови за счет прилипания к пластику и фильтрации на найлоновой вате, и составили 91% CD2+, тогда как PHA-бласты генерируют за счет культивирования клеток в присутствии 10 мкг/мл фитогемагглютинина (PHA). Клетки метят флуорохромом BCECF (Molecular Probes, Inc. ), промывают и снова суспендируют в среде анализа. MAb предварительная обработка клеток состоит в инкубировании с 1/200 разбавлениями для асцитов в течение 45 минут при 4oC, после чего 105 клеток переносят в каждую ячейку. Клеточные линии прилипают к твердой фазе LFA-1 в течение 1 часа при 37oC, а не прилипшие клетки удаляют 6 раз отсасыванием (23 gauge иглой), тогда как лимфоциты осаждают при 30 x g в течение 5 минут и инкубируют при 37oC в течение 30 минут. Постоянные лимфоциты, которые удаляют, стряхивая среду с пластин 8 раз, добавляя по 100 ψ между каждыми промывками. Стряхивание более эффективно для тщательного удаления несвязанных T лимфоцитов, которые более трудно удалять из-за их малого размера. Флуоресценцию непосредственно количественно определяют из 96 ячеечных пластин, используя флуоресцентный концентрационный анализатор (Baxte). Все MAb используют при насыщенных концентрациях (1/200 разбавлений асцитов) и включают ТSI/22 (анти-CD11a, Ig G1) (Sanches-Madrid. et al., Proc. Natl.. Acad. Sci USA 79: 7489 - 7493 (1982)), RRI/1 (анти-ICAM-1, IgG 1) (Rothlein et al. , J. Immunol. 137: 1270 - 1274 (1986)), CBP-102/2 (анти-ICAM-2, IgG1a) (de Fouderolles et al., J. Exp. Med. (1991) в печати) и CBP-IC3/1 (анти-ICAM-3, IgG1). CBP-IC3/1 получают при слиянии мышиной миеломы P3X63A 8,653 с клетками селезенки из SKW3-инъектированных BaIb/c мышей (Geffer et al., Som. Cell Gen. 3.231 - 236 (1977)) и 600 гибридом скринируют по их способности ингибировать SKW3 связывание с очищенными LFA-1. CRB-IC3/1, как было показано, не реагируют ни с очищенным LFA-1, ни с COS клетками, трансфектированными либо ICAM-1, ICAM-2 или LFA-1 кДНК (данные не представлены). Один из четырех представительных экспериментов представлен, а прямоугольники ошибок указывают стандартные отклонения.
Пример II. Цитометрический анализ в потоке клеточной экспрессии ICAM-1, ICAM-2 и ICAM-3.
Все использованные клеточные линии были ранее описаны Hibbs et al., J. Clin Invest 85: 674 - 681 (1990). Периферические одноядерные клетки крови (РВМС) были получены при седиментации декстрана и Ficoll-Hypaque (1,077) центрифугированием, как описано (Dusfin et al., J. Cell. Biol. 107: 321 - 331 (1988)).
Нейтрофилы удаляют из клеточного осадка, а примесные эритроциты удаляют гипотоническим лизисом. Лимфоциты и моноциты разделяют цитометрическим анализом, используя взаимноперпендикулярное светорассеяние, и подтверждают моноцитанами и T-клеткам и специфичным MAb. Прилипание к пластику используют для обогащения лимфоцитами из РВМС и клетки культивируют в присутствии 10 мкг/мл фитогемагглютинина (PHA). Образцы анализируют, используя EPICS цитометр в потоке, и количество определяют, используя EPICS Immune-Brite флуоресцентные шарики (Coulter) для калибровки цитометра. Экспрессия ICAM-3 на покоящихся лимфоцитах в 2 - 3 раза больше, нежели CD3 или LFA-1, тогда как моноциты экспрессируют в 3 - 4 раза больше ICAM-3, нежели LFA-1. Уровни экспрессии Mac-1 на нейтрофилах эквивалентны экспрессии Mac-1 (CD11/CD18). Обработка клеток фосфолипазой C выявила отсутствие PI-связанной формы ICAM-3.
В таблице 2 представлена относительная антигенная поверхностная экспрессия ICAM-3 по данным иммунофлуоресцентной цитометрии в потоке.
Примечания к таблице 2:
* - Мембранная экспрессия, определенная иммунофлуоресцентным анализом в потоке в соответствии с указанными материалами и способами. Величины являются результатом по крайней мере двух экспериментов. Для калибровки цитометра используют флуоресцентные шарики таким образом, что каждая единица соответствует приблизительно 103 эквивалентам флуоресцина (Caulte Diagnostics, Hialeah, FL). "Незначительные" клеточные линии включают: эндотелиальные клетки умбиликальной вены человека, huvec; карциномы груди человека, Hep G2; клеточную линию эпителоидной карциномы человека HeLa; рабдомиссаркомы человека, PD 3/5; фибросаркомы человека и FS 1, 2, 3; глиобластомы человека.
Пример III. Иммуноосаждение ICAM-3.
Введение меток 125I на поверхности клеток осуществляют используя йодоген, как указано (Kishimoto et al. J. Biol., Chem. 264: 3588 - 3595 (1989)). Клетки лизируют тритоном X-100 (1%). Лизаты предварительно очищают бычьим IgG на сефарозе, а затем инкубируют с соответствующей MAb-связанной сефарозой в течение 2 часов. Шарики промывают и кипятят в образце буфера, содержащем 50 мМ Tris, 1% SDS и 1% 2-меркаптоэтанола. Невосстановленные образцы кипятят в буфере без 2-меркаптоэтанола и обрабатывают 20 мМ йодоацетамида. Образцы анализируют на 7% полиакриламидном геле в виде вертикальных столбиков, как было указано ранее (Laemmlin, H.K., Nature 227: 680 - 685 (1970)) и протеины визуализируют авторадиографией. Обработка образцов N-гликанозой (Lenzyme), как было описано ранее (Tarentino et al., Biochemestry 24: 4665 - 4671 (1985)), с использованием концентрации 10 ед/мл при 37oC в течение 18 часов, как было показано, приводит к оптимальному отщеплению всех -связанных олигосахаридов от пептидной цепи. МНС класс I содержит N-связанный углеводород, тогда как ICAM-3/Мr 60000, 6 N-связанных сайтов, основная цепь Мr 28383) сильно гликоэилирована.
Пример IV. Распределение ICAM-3.
Анти-ICAM-3 антитела настоящего изобретения используют для дальнейшей оценки тканевого распределения ICAM-3 протеина с использованием цитометрии в потоке. В таблице 3 представлены суммарно типы клеток, которые демонстрируют поверхностную экспрессию ICAM-3. Экспрессия ICAM-3, по-видимому, ограничена кроветворными клетками. Данные цитометрии в потоке подтверждаются в процессе иммуногистохимического окрашивания ткани с использованием меченых анти-ICAM-3 антител (данные не представлены). Что касается данных цитометрии в потоке, иммуногистохимические исследования показывают, что ICAM-3 экспрессия, по-видимому, ограничена кроветворными клетками.
В таблице 3 представлена относительная поверхностная антигенная экспрессия ICAM-3 по данным иммунофлуоресцентной цитометрии в потоке.
Примечания к таблице 3:
* - Мембранную экспрессию определяют иммунофлуоресцентной цитометрией в потоке, как указано в материалах и способах. Указанные значения представляют собой среднее из по крайней мере двух экспериментов. Для калибровки цитометра используют флуоресцентные шарики таким образом, что одна единица составляет приблизительно 103 эквивалентов флуоресцина (Culte Diagnostics, Hialeah, FL).
± - Незначительные клеточные линии включают: Эндотелиальные клетки умбиликальной вены человека, HUVEC; карциномы груди человека, Hep C2; клеточную линию эпителоидной карциномы человека HeLa; рабдомиосаркомы человека, RID 3/5; фибросаркомы человека и FS 1, 2, 3; глисбластомы человека.
Пример V. Очистка ICAM-3.
ICAM-3 очищают, используя иммуноафинную хроматографию, где анти-ICAM-3 антитела CBP-ICAM-3 иммобилизуют на матрицах, используя известные специалистам способы. В качестве исходных материалов для выделения ICAM-3 были использованы многочисленные источники. Они включают, хотя и не ограничиваются ими, SKW3, клеточную линию тимомы человека и миндалины человека. Способы, которые используют для очистки ICAM-3 соответствуют тем, которые описаны de Fougerolles et al., в J. Exp. Med. 175: 185-190 (1992). Специалистам известны режимы промывки и элюирования (реагенты варьируются слегка для каждого антигена, подлежащего очистке, и для каждого антитела, которое используют в иммуноафинной хроматографии).
Как было указано ранее, ICAM-3 очищенные от SKW3 клеток или клеток миндалин человека, по-видимому, имеют молекулярный вес от около 120 до 124 кД, тогда как ICAM-3, выделенный из нейтрофильных лизатов, приводит к широкой полосе, соответствующей молекулярным весам от около 120 до 150 кД.
Было показано, что ICAM-3, очищенный таким образом, сохраняет свою способность связывать анти-ICAM-3 антитела и LFA-I на различных клетках (фиг. 6 и 7).
Пример VI. Идентификация различных молекулярных весов ICAM-3.
Иммуноосаждение и анализ с помощью электрофореза на полиакриламидном геле используют для определения молекулярных весов ICAM-3, на клетках различных типов. Молекулярный вес ICAM-3, иммуноосажденных с нейтрофилов, как было обнаружено, отличается от молекулярного веса ICAM-3, иммуноосажденного с лимфоцитов. ICAM-3 выделенные из лимфоцитных и лимфоидных клеточных линий, проявляются как полоса с молекулярным весом от около 120 до 124 кД. Молекулярный вес ICAM-3, выделенных с нейтрофилов, несколько выше того, который имеют ICAM-3, экспрессированные лимфоидными клетками, и проявляются как диффузная полоса от около 120 до 150 кД (фиг. 4).
Такие вариации в размерах не являются необычными для членов семейства ICAM гликопротеинов. ICAM-1 демонстрирует аналогичные вариации молекулярного веса среди различного типа клеток. Такие варианты молекулярного веса, видные для ICAM-1, как было показано, вызываются различиями в степени гликозилирования. Поэтому вариации в молекулярных весах ICAM-3 наиболее вероятно связаны с различиями в гликозилировании. Специалисты могут легко подтвердить это, выделяя ICAM-3 из нейтрофилов различными гликозидазами и другими энзиматическими обработками и наблюдая за их влиянием на молекулярные веса.
Как было показано, различные степени гликозилирования ICAM-1 влияют на MAC-1/CD11 /CD18/ связывание. Поэтому, меняя степень гликозилирования ICAM-3, можно создавать производные ICAM-3, которые обладают модифицированными афинностями для связывания с различными лигандами ICAM-3, например, членам и CD11/CD18 семейства гликопротеинов.
Пример VII. Антитела ICAM-3.
Мышам вводят инъекцией комбинацию адъюванта и ICAM-3 протеина, предварительно очищенного от SKW3 или клеток миндалин, как описано ранее. Моноклональные антитела иммунизованных животных получают, используя известные специалистам способы. В таблице 4 представлены различные полученные анти-ICAM-3 антитела. Различные антитела были идентифицированы на основании их способности реагировать с очищенными иммобилизованными ICAM-3 с помощью ELISA. Затем положительные MAb скринируют на различных клеточных линиях, которые, как известно, являются ICAM-3 положительными, а затем иммуноосаждают из радиомеченых ICAM-3 положительных клеточных лизатов. Затем MAb тестируют по их способности блокировать PMA-индуцированную SKW3 клеточную аггрегацию в присутствии анти-ICAM-1 и анти-ICAM-2 антител. В другом варианте анти-ICAM-3 антитела можно идентифицировать по их способности ингибировать связывание SKW3 клеток с иммобилизованными очищенными ICAM-3.
Одним из доказательств того, что существует третий лиганд LFA-1 является наличие ICAM-1/ICAM-2 независимого пути для аггрегации SKW3 клеток, стимулированной PMA. Авторы смогли блокировать эту аггрегацию либо мышиной анти-ICAM-3 поликлональной антисывороткой, либо комбинацией CBR-IC3/1 и CBR-1C3/2.
Примечания к таблице 4:
* - Относится к шкале аггрегации в J. Exp. Med (1991) 0 = нет аггрегации
** - Другой ICAM-3 MAb из лаборатории в Испании (F. Sanches-Madrid). После представления наших результатов они напомнили им о мA, который они получили, но никогда не характеризовали. Оказалось, что это ICAM-3. CBR-IC3/2 3/3, 3/5 иммуноосаждают ту же полосу 120 кД, что и CBR-IC3/1. Проверка MAb по их способности к вестерн блотту.
Пример VIII. Иммунологический анализ с анти-ICAM-3-антителами.
Анти-ICAM-1 антитела (RR1/1) и анти-ICAM-2 антитела (CBR-IC2/2) и анти-ICAM-3 антитела (комбинация CBR-IC3/1 и CBR-IC3/3) тестируют по их способности блокировать адгезию клеток SKW3, стимулированную: (1) PMA, к очищенным ICAM; (2) клеточное деление, стимулированное фитогемагглютином (PHA), и (3) смешанную реакцию лимфоцитов (MLR).
Ранее было показано, что PMA может активировать LFA-1, повышая тем самым их способность связываться с ICAM-1 (Dustin et al. Nature 341: 619 (1989)). На фиг. 5 показано, что аналогичный механизм активации присутствует в LFA-1/ICAM-3 связывании.
Способность различных антител блокировать связывание SKW3 клеток, стимулированных PMA, с очищенными ICAM-1 и ICAM-3 была исследована (фиг. 6). Антитела CBR-IC3/1 и CBR-IC3/2, если они присутствуют индивидуально, не блокируют эффективно связывание стимулированных PMA SKW3 клеток с очищенными ICAM-3. Однако связывание стимулированных PMA SKW3 клеток с ICAM-3 можно блокировать, используя комбинацию этих двух антител. Как было отмечено de Faugerolles в J. Exp. Med, CBR-IC3/1, моноклональные антитела сами способны блокировать связывание ICAM-3 с очищенными LFA-1.
Далее проводили анализ связывания PMA стимулированных SKW3 клеток с ICAM-3 для определения того, существует ли температурная зависимость и зависимость от катиона. Было показано, что существует как температурная (фиг. 8) зависимость, так и зависимость от катиона (данные не представлены) для ICAM-1, ICAM-3/LFA-1 связывания.
На фиг. 9 представлен суммарный эффект различных антител на PHA стимуляцию клеточного деления. Как было указано ранее (Krensky et al., J. Immunol 131: 611/616 (1983)), PHA стимулирует клеточную пролиферацию таким способом, что ее можно ингибировать анти-LFA-1 и анти-ICAM-1 антителами, анти-ICAM-2 антителами или комбинацией анти-ICAM-1 и анти-ICAM-2 антител и они оказывают незначительное влияние на деление клеток, стимулированное PHA. Однако комбинация 1) анти-ICAM-1, ICAM-2 и анти-ICAM-3 антител, 2) двух анти-ICAM-3 антител (IC3/1 и IC3/2) и 3) анти-ICAM-1 и анти-ICAM-3 антител оказывается эффективной при блокировании PHA стимулированного деления клеток.
Пример IX. Смешанная лимфоцитная реакция.
Для определения иммунологической реакции используют анализ. Этот анализ в основном включает добавление химически фиксированных чужеродных клеток (стимуляторных клеток) к раствору, содержащему PВL от различных индивидуумов (клетки респондеров), и определение уровня реакции, используя пролиферацию клеток респондеров как индекс, как было указано ранее (Krensky et al., J. Immunol. 131: 611-616 (1983)). Этот анализ представляет собой первый шаг по идентификации композиций, пригодных для предотвращения отторжения трансплантатов.
Влияние различных антител и комбинаций антител на MLR реакции представлено на фиг. 10. В сумме, отдельно анти-ICAM-3 демонстрирует малое влияние. Однако было обнаружено, что более высокий уровень эффективности блокирования MLR наблюдается для комбинаций анти-ICAM-1 и анти-ICAM-3 антител. Наиболее сильную реакцию наблюдают для комбинации анти-ICAM-1, анти-ICAM-2 и анти-ICAM-3 антител.
Пример X. Пептидные последовательности ICAM-3.
ICAM-3 очищают от клеток миндалин человека иммуноафинной хроматографией, как указано в примере 5. Очищенные ICAM-3 энзиматически переваривают с помощью Lys-C энзима, используя известные способы, и пептидные фрагменты разделяют с помощью ВЖХ. Фиг. 11 представляет газовую хроматограмму различных пиков протеинов, которые наблюдаются при типичном переваривании. Пики 10 и 17 были идентифицированы как содержащие пептидные фрагменты достаточного размера и структуры, которые следует секвенировать (здесь и далее NK-10 и NK-17 пептиды соответственно).
Аминокислотные последовательности NK-10 и NK-17 пептидов определяют стандартными способами. Было найдено, что последовательность NK-10 протеина представляет собой KIDRAYCRQHLK (последовательность ID N 1). Было предположено, что первый остаток лизина (K), изображенный в последовательности, существует, так как Lys-C отщепляет протеины после лизинового остатка.
Последовательность NK-17 пептида была представлена как KIAZETSLK (последовательность ID N 2). Как и в случае NK-10 протеина, первый лизиновый остаток был предположен, а позднее его присутствие было подтверждено с помощью анализа последовательности ДНК.
Сравнение последовательностей NK-10 и NK-17 протеинов с известными ICAM-1 и ICAM-2 последовательностями выявило высокую степень гомологичности. Пептид NK-17 демонстрирует значительную гомологичность с последовательностями, содержащимися в первом Ig домене ICAM-2. Пептид NK-10 демонстрирует слабую гомологичность с последовательностью внутри домена 4 ICAM-1.
Пример XI. кДНК клонирование ICAM-3.
Дегенеративные олигонуклеотидные зонды были созданы на базе полученных ранее последовательностей пептидов. Далее зонды были сконструированы для включения ранее идентифицированного кодона, предпочтительно наблюдавшегося в ICAM-1 и ICAM-2 нуклеотидных последовательностях. Нуклеотидные последовательности зонда на основании аминокислотной последовательности NK-17 и NK-10 протеинов выглядят следующим образом: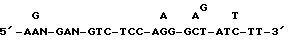 (последовательность ID N 3) NK-17 зонд. Олигомер 23 с 24-кратной дегенеративностью, содержащий 2N (инозин)
(последовательность ID N 3) NK-17 зонд. Олигомер 23 с 24-кратной дегенеративностью, содержащий 2N (инозин)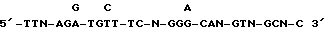 (последовательность ID N 4) NK-10 зонд. Олигомер 25 с 8-кратной дегенеративностью, содержащей 5N (инозин)
(последовательность ID N 4) NK-10 зонд. Олигомер 25 с 8-кратной дегенеративностью, содержащей 5N (инозин)
Была создана кДНК экспрессионная библиотека, созданная из РНК миндалин в соответствии с описанным ранее (Wong et al., Proc. Natl. Acad. Sci, USA 82: 7711-7716 (1985)). Эту библиотеку скринируют, используя NK-17 зонд, и бляшки, демонстрирующие положительную гибридизацию, повторно скринируют, используя NK-10 зонд. Один клон, клон 11, 2, как было обнаружено, имеет вставку от около 1,6 до 1,8 кб длиной. На фиг. 12 представлено схематическое изображение клона 11.2 и положение NK-10 и NK-17 пептидов.
Концы вставки были секвенированы в соответствии с известными специалистам способами и, используя праймеры, полученные из прилегающего вектора, тогда как NK-10 зонд используют в качестве внутреннего секвенирующего праймера. Таким образом, получают последовательности, которые считывались только по одной нити. Специалисты легко поймут, что такая последовательность может быть получена обычными способами.
Резюме полученных последовательностей представлено на фиг. 18. По этому поводу следует заметить следующее. Во-первых, была идентифицирована последовательность с высокой степенью гомологичности с трансмембранным участком ICAM-1. Поэтому, если нужно получить растворимые фрагменты ICAM-3, нужно исключить все последовательности после начала трансмембранного участка, отмеченные на фиг. 12. Во-вторых, последовательность, полученная с 5 конца клона 11.2, по-видимому, близка к 5 концу природного гена, что следует из гомологичности последовательностей, которые существуют между ICAM-1, ICAM-2 и ICAM-3.
Пример XII. Гомологичность последовательностей между различными молекулами ICAM.
Нуклеотидные и аминокислотные последовательности NK-10 и NK-17 пептидов, так же как и полученные ранее нуклеотидные последовательности, сравнивали с известными последовательностями ICAM-1 и ICAM-2 (фиг. 13-17).
Полученные результаты показывают, что ICAM-3 демонстрируют высокий уровень гомологичности как ICAM-1, так и ICAM-2. Гомологичности между ICAM-1 и ICAM-3 наблюдаются внутри первого (фиг. 16) и четвертого/пятого доменов ICAM-1, а также трансмембранного домена и части цитоплазмического домена ICAM-1 (фиг. 13 и 14). Дополнительные рассмотрения банка данных не выявили другой последовательности, которая была бы идентична или аналогична последовательности, полученной для ICAM-3. Значительная гомологичность наблюдалась также для ICAM-3 и первых доменов ICAM-2 (фиг. 17).
Клон 11.2 использовали для дальнейшего скринирования библиотек. Были обнаружены дополнительные клоны для получения кДНК вставок от около 2 до 2,4 кД. По всей видимости, они представляют клоны кДНК полной длины, так как ничего не было обнаружено большего размера.




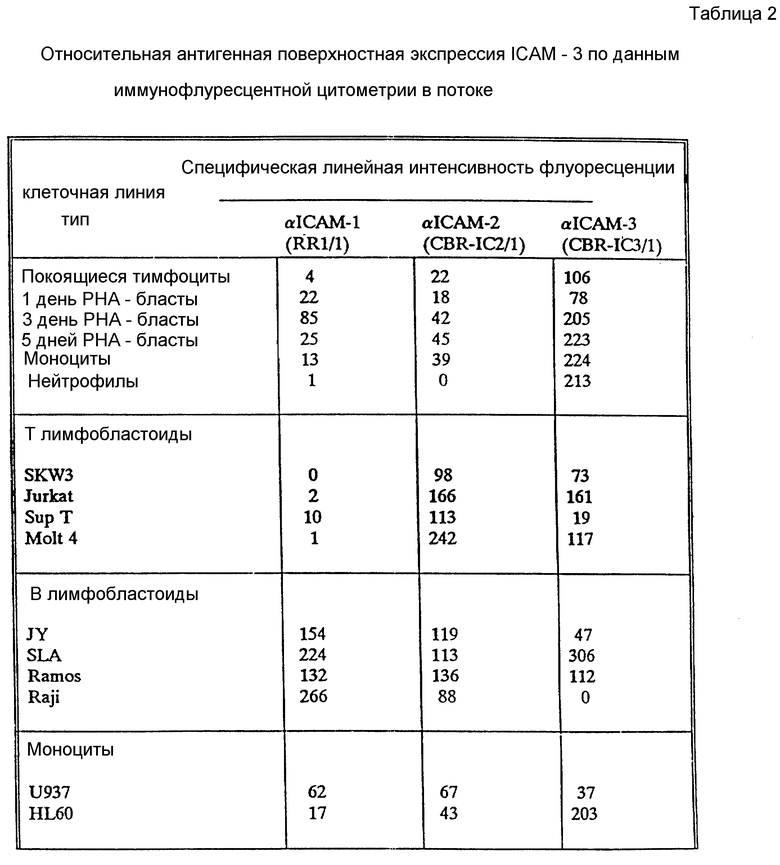
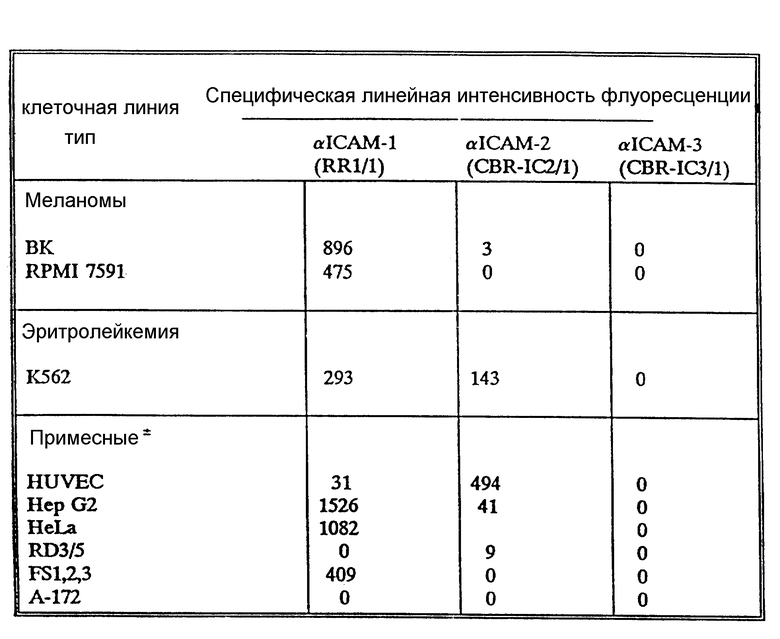
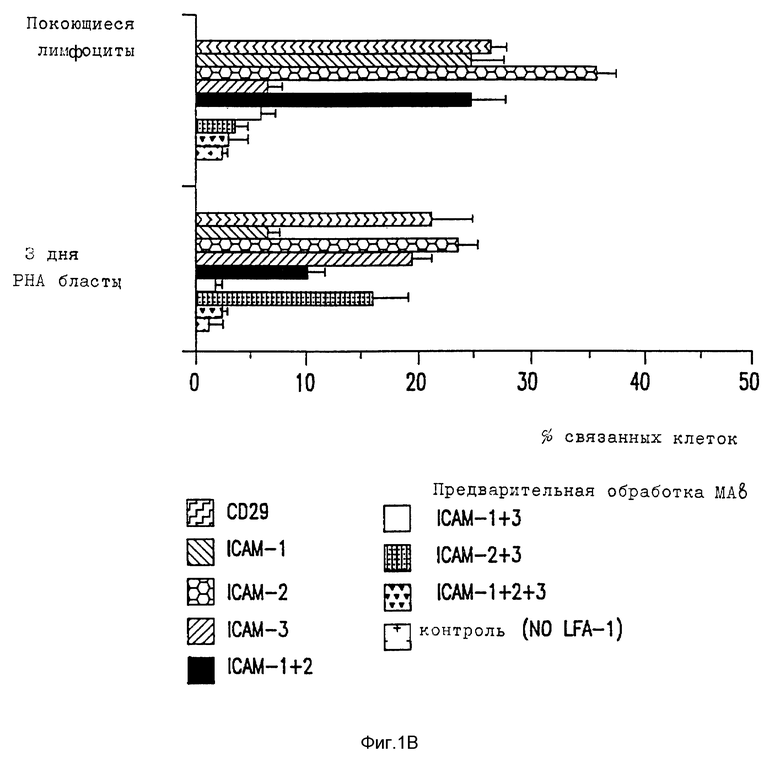
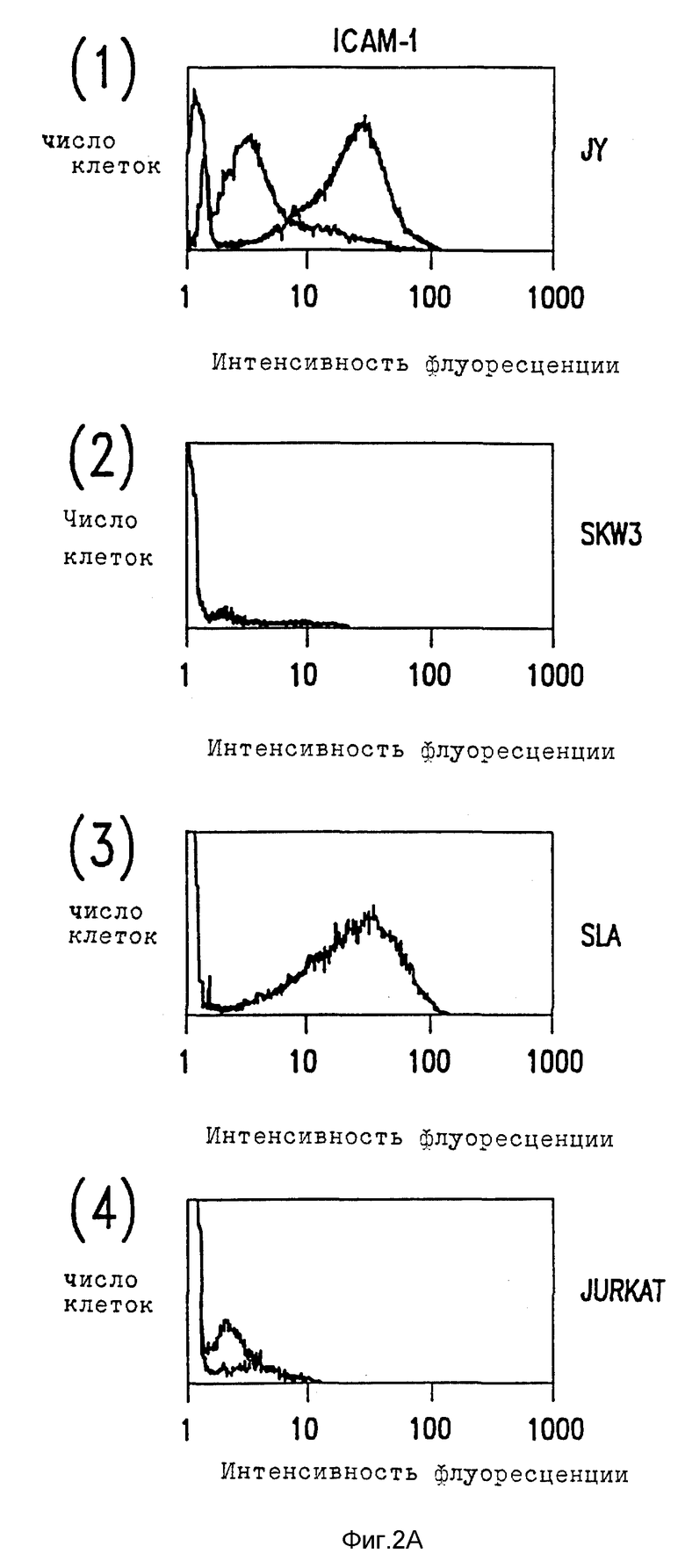
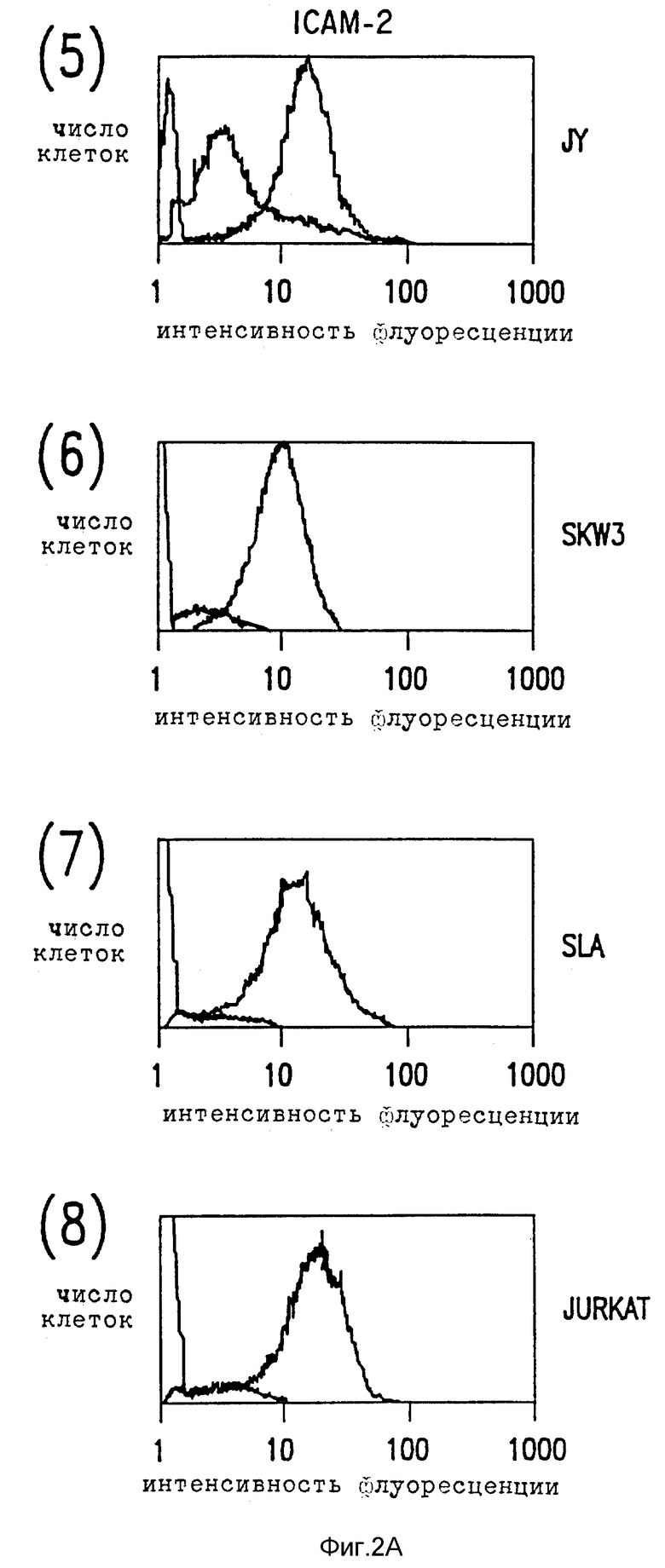
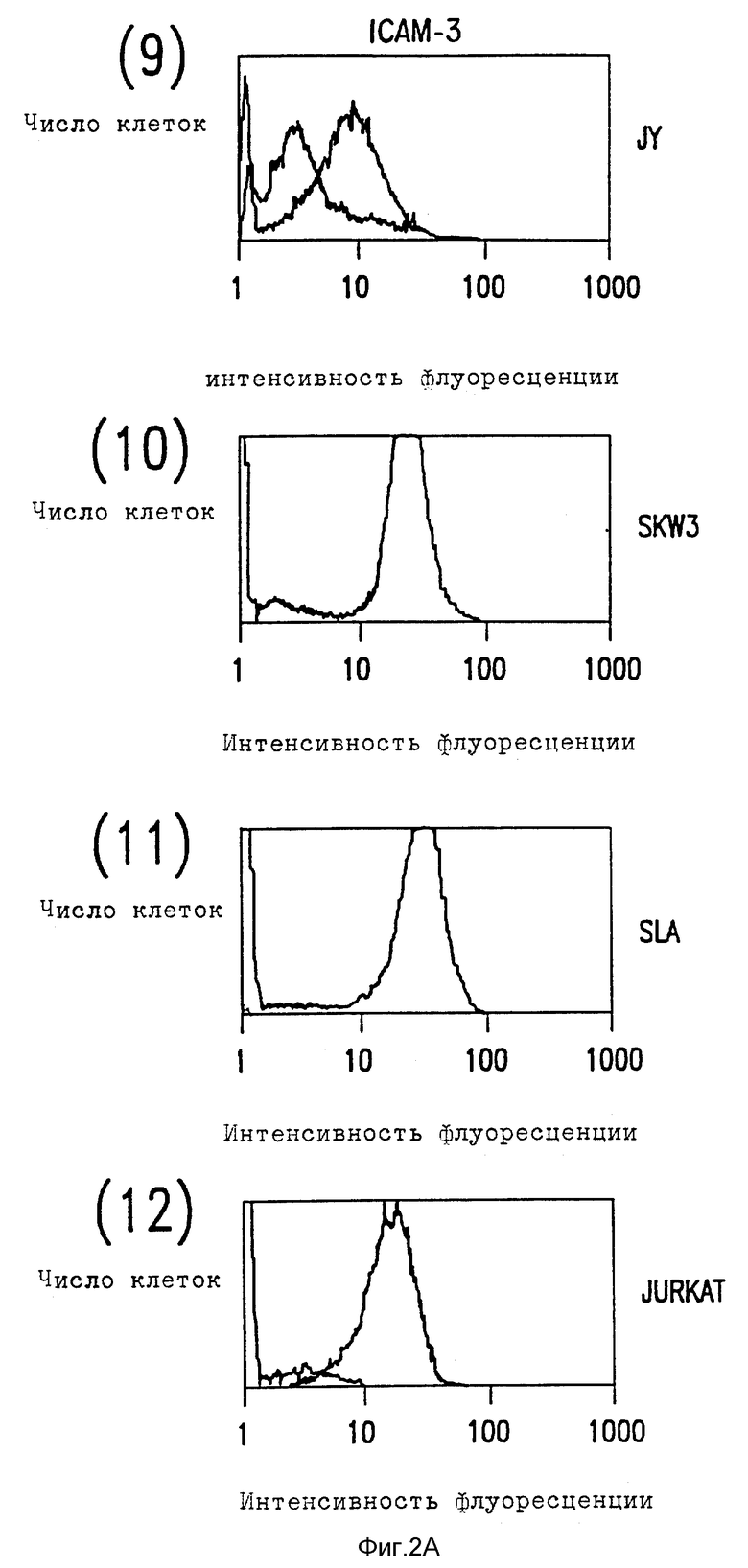

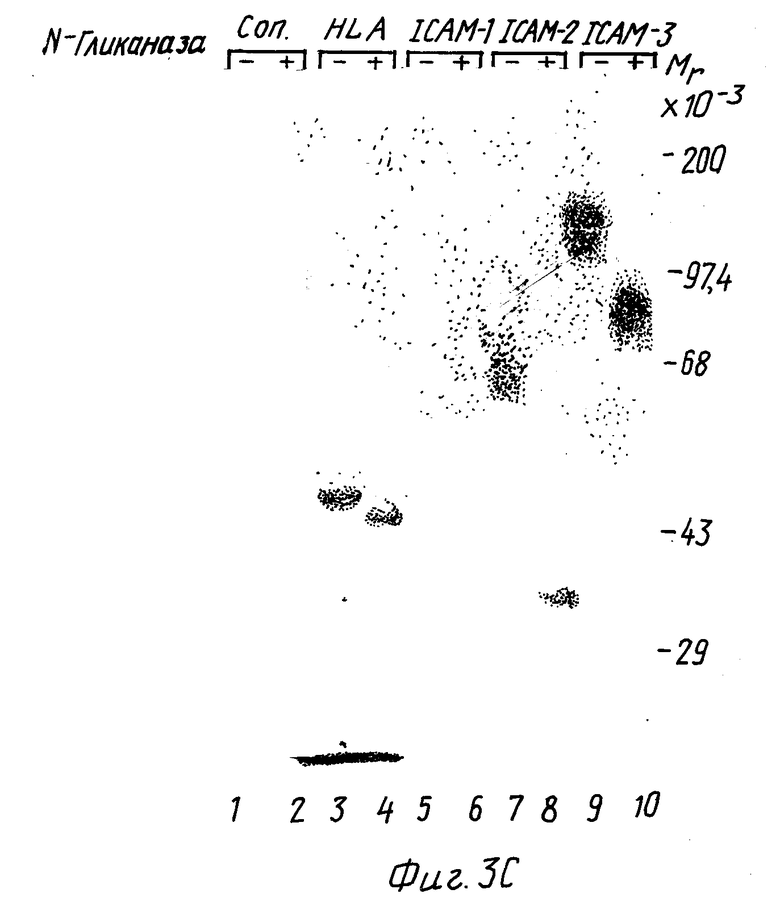
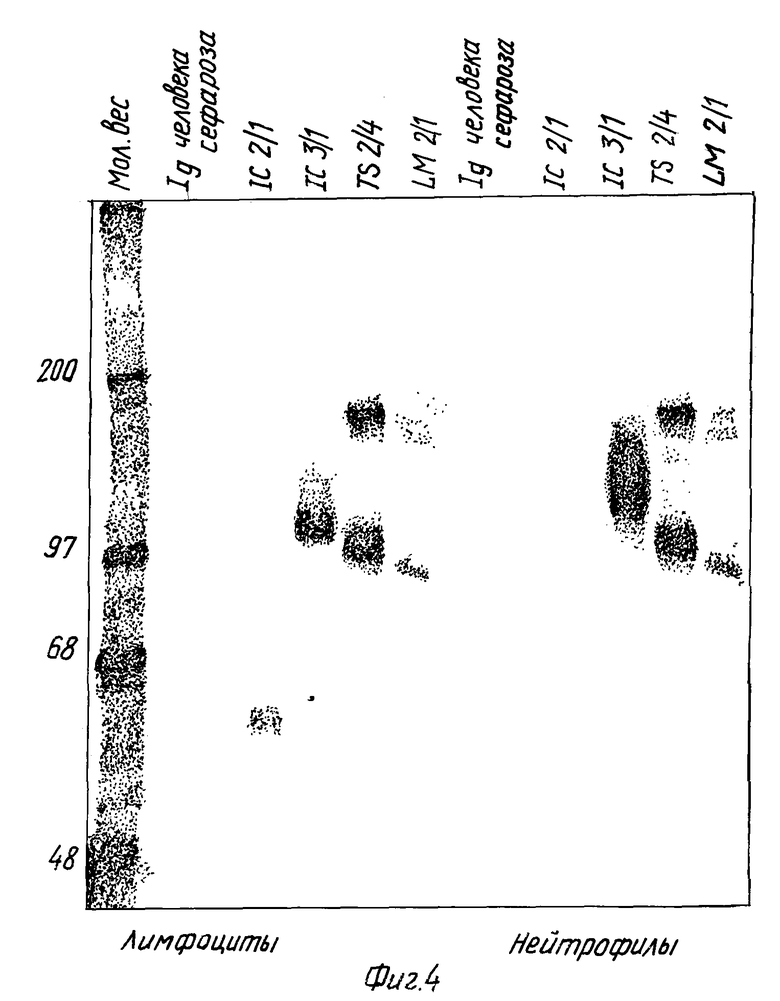
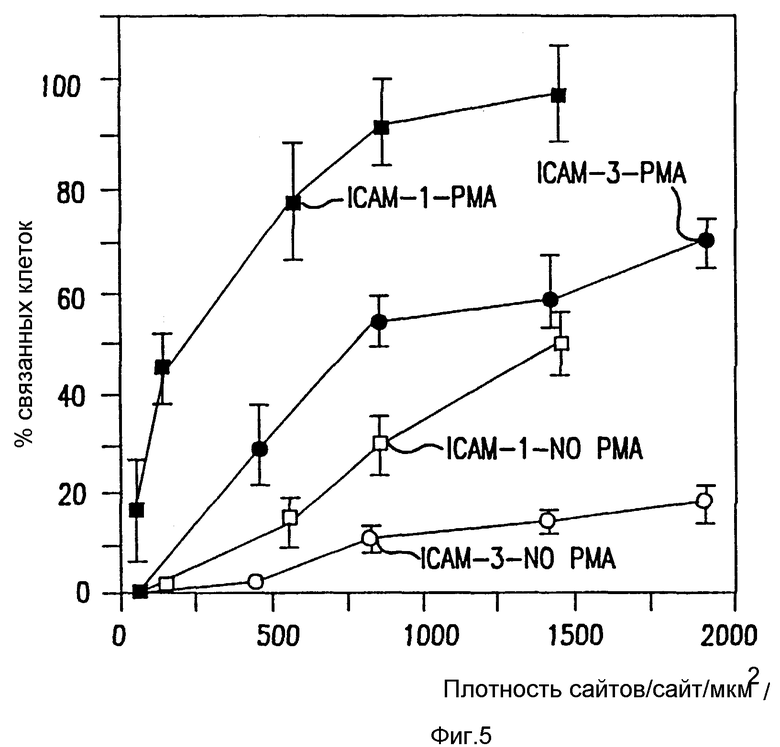
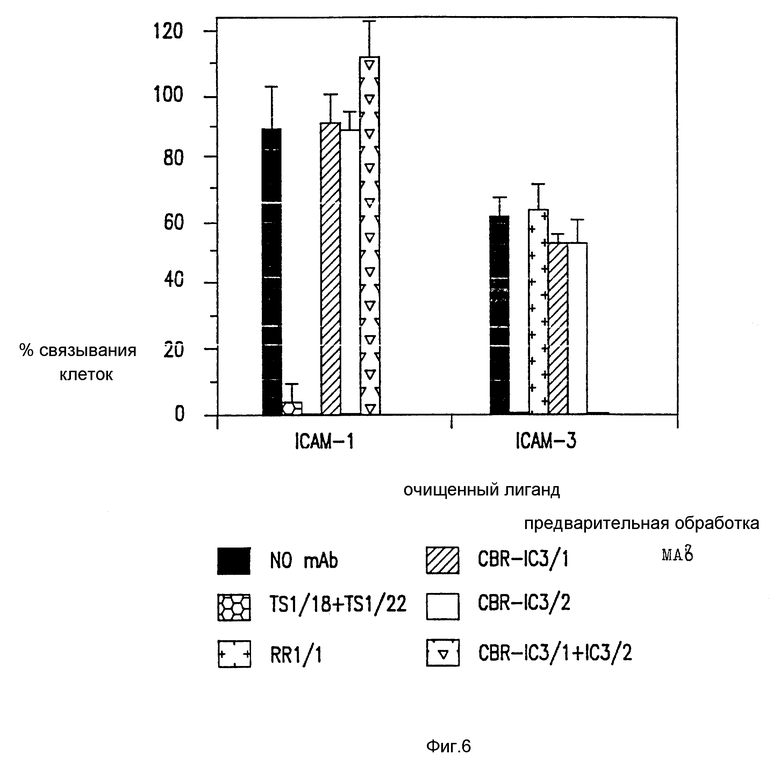
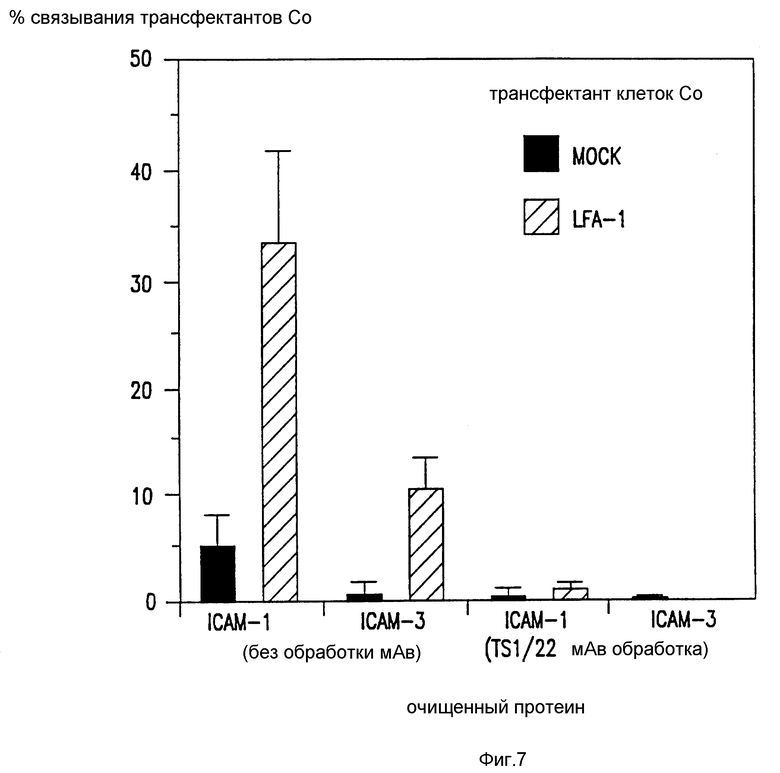
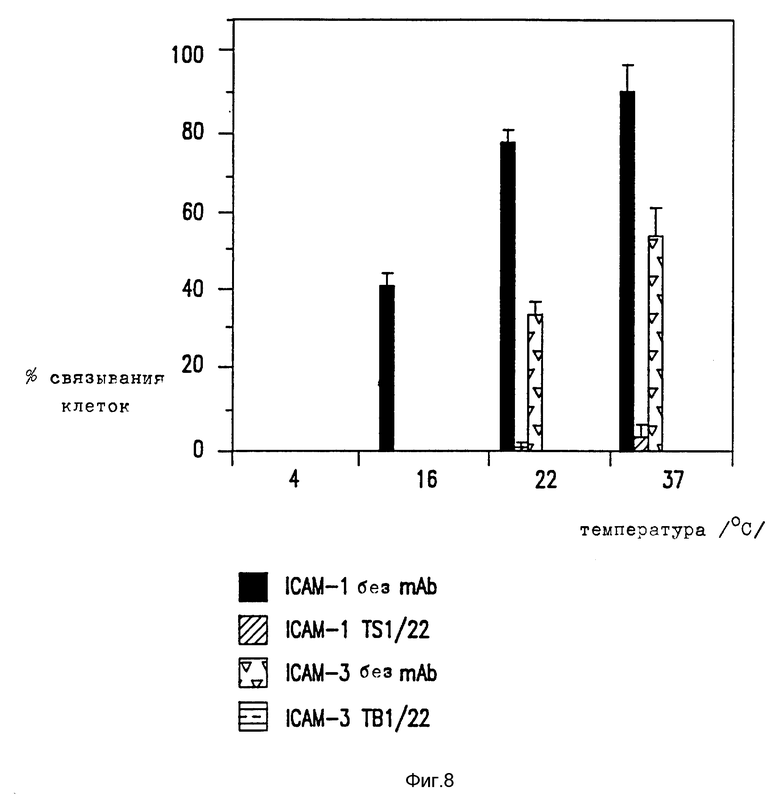
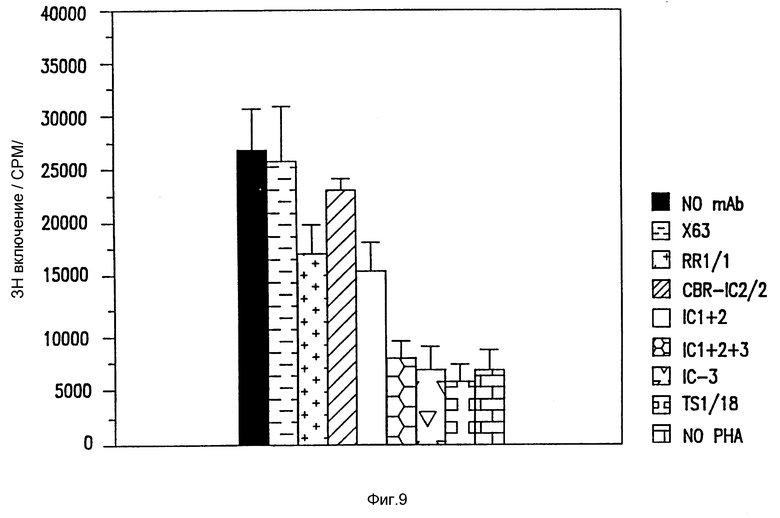
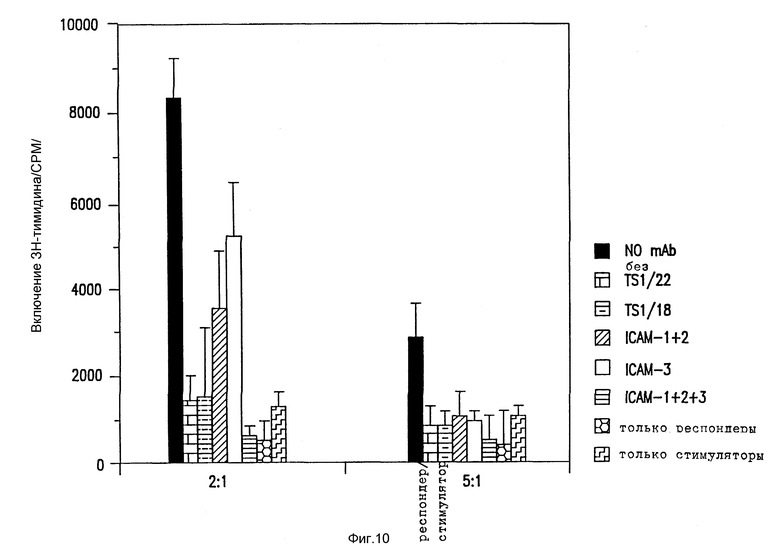
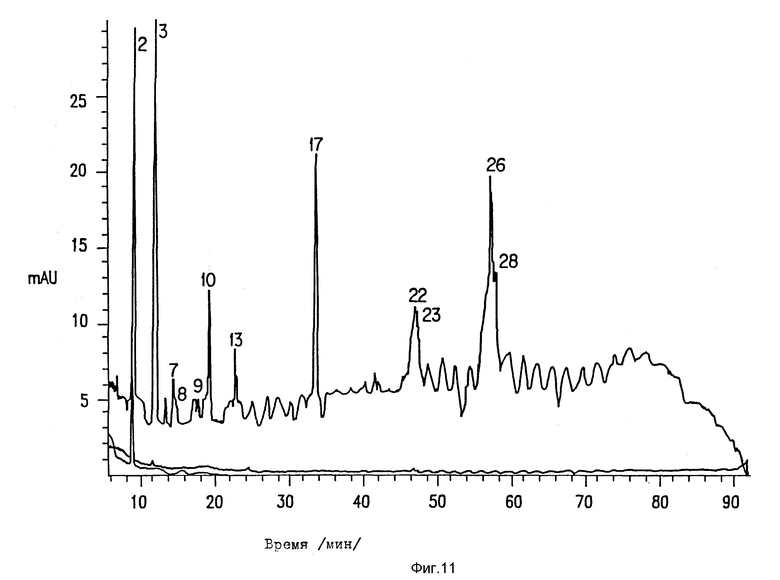
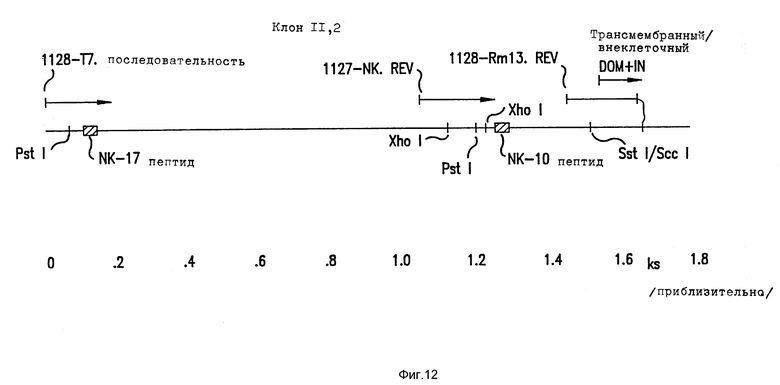








Изобретение относится к области медицины, точнее - медицинской генетики. Сущность изобретения заключается в создании рекомбинантной молекулы ДНК, кодирующей молекулу адгезии ICAM-3, а также в экспрессии с помощью ДНК, молекулы адгезии ICAM-3. Изобретение включает также создание антитела или его фрагмента, способного связываться рецептором в организме больного. Создана также фармацевтическая композиция на основе антител. Технический результат изобретения заключается в расширении арсенала иммунокомпетентных средств. 4 с. и 11 з.п. ф-лы, 18 ил., 4 табл.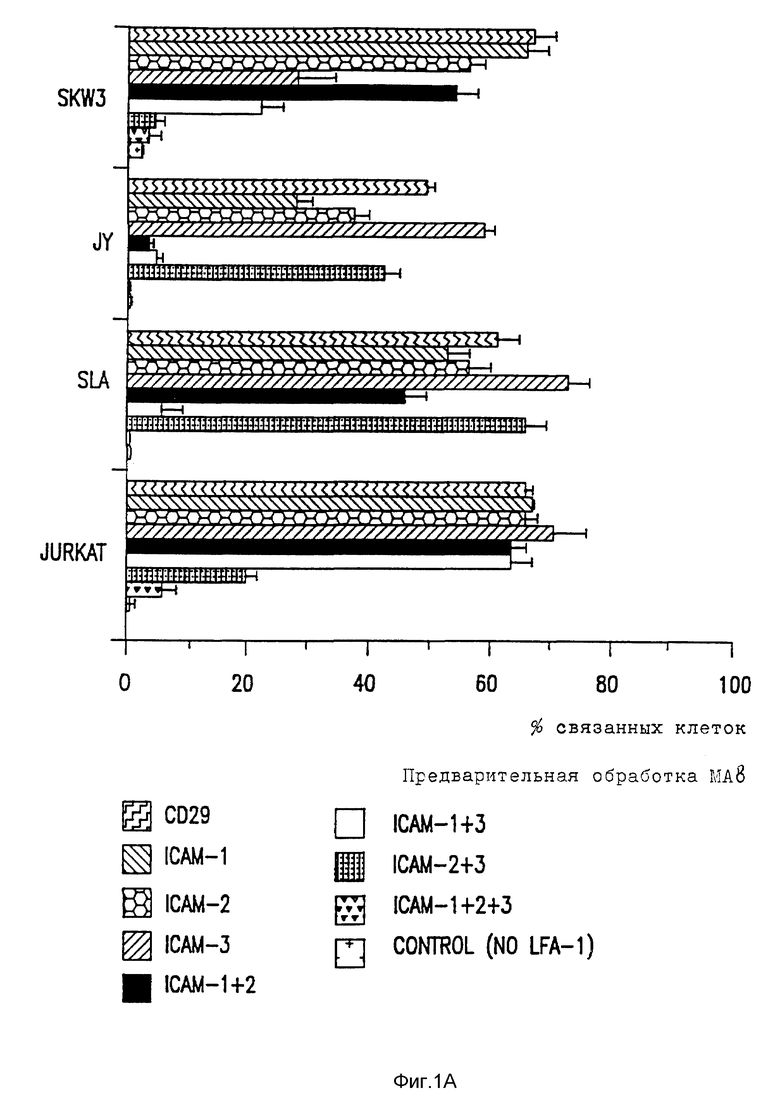
| Пожарный двухцилиндровый насос | 0 |
|
SU90A1 |
| Способ размножения копий рисунков, текста и т.п. | 1921 |
|
SU89A1 |
| Endotelial Leucocyte Adhesirum Molecule..., Science 243, p.1160, 1989. | |||

