Предпосылки изобретения
Данное изобретение относится к способу получения L-глутаминовой кислоты с помощью ферментации. L-глутаминовая кислота широко используется в качестве сырья для приправ и так далее.
L-глутаминовую кислоту главным образом получают ферментацией с использованием так называемых коринеформных бактерий, продуцирующих L-глутаминовую кислоту, относящихся к роду Brevibacterium, Corynebacterium или Microbacterium, или их мутантных штаммов (Amino Acid Fermentation, Gakkai Shuppan Center, pp.195-215, 1986). В качестве способов получения L-глутаминовой кислоты ферментацией с использованием других бактериальных штаммов известен способ с использованием микроорганизма, относящегося к роду Bacillus, Streptomyces, Peniclllium или ему подобному (патент США No.3220929), способ с использованием микроорганизма, относящегося к роду Pseudomonas, Arthrobacter, Serratia, Candida или ему подобному (патент США No.3563857), способ с использованием микроорганизма, относящегося к роду Bacillus, Pseudomonas, Serratia, Aerobacter aerogenes (в настоящее время называемому Enterobacter aeroqenes) или ему подобному (Японская патентная публикация (Kokoku) No.32-9393), способ с использованием мутантного штамма Escherichia coli (выложенная заявка на патент Японии (Kokai) No.5-244970) и так далее. Кроме того, авторы данного изобретения предложили способ получения L-глутаминовой кислоты с использованием микроорганизма, относящегося к роду Klebsiella, Erwinia или Pantoea (выложенная заявка на патент Японии No.2000-106869).
Кроме того, обнаружены различные приемы повышения способности продуцировать L-глутаминовую кислоту путем усиления активностей ферментов биосинтеза L-глутаминовой кислоты с помощью использования технологии рекомбинантной ДНК. Например, сообщалось, что введение гена, кодирующего цитратсинтазу, происходящего из Escherichia coli или Corynebacterium glutarnicum, является эффективным для усиления способности продуцировать L-глутаминовую кислоту бактериями Corynebacterium или Brevibacterium (Японская патентная публикация (Kokoku.) No.7-121228). Кроме того, в выложенной заявке на патент Японии No.61-268185 сообщается о клетке, несущей рекомбинантную ДНК, содержащую ген глутаматдегидрогеназы, происходящий из бактерий Corynebacterium. Кроме того, в выложенной заявке на патент Японии No.63-214189 раскрывается способ повышения способности продуцировать L-глутаминовую кислоту амплификацией гена глутаматдегидрогеназы, гена изоцитратдегидрогеназы, гена аконитатгидратазы и гена цитратсинтазы.
Хотя продуктивность по L-глутаминовой кислоте значительно увеличивается с помощью указанной выше селекции микроорганизмов или усовершенствования способов получения, требуется разработка способов более эффективного получения L-глутаминовой кислоты при более низкой стоимости, чтобы удовлетворить дальнейший рост потребности в будущем.
Известен способ, при котором по мере того, как проводят ферментацию, кристаллизуют L-аминокислоту, накопленную в культуре (выложенная заявка на патент Японии No.62-288). В данном способе концентрацию L-аминокислоты в культуре поддерживают ниже определенного уровня путем осаждения накопленной в культуре L-аминокислоты. В частности, L-триптофан, L-тирозин или L-лейцин осаждают во время ферментации путем регулирования температуры и рН культуры, или добавлением к среде поверхностно-активного вещества.
Хотя, как описано выше, известен способ осуществления ферментации, сопровождаемой осаждением L-аминокислоты, подходящими для данного способа аминокислотами являются аминокислоты, проявляющие относительно низкую растворимость в воде, и не известно никаких примеров применения способа для высокорастворимых в воде аминокислот, таких как L-глутаминовая кислота. Кроме того, для осаждения L-глутаминовой кислоты среда должна иметь низкое значение рН. Однако бактерии, продуцирующие L-глутаминовую кислоту, такие как, упомянутые выше, не могут расти в кислых условиях, и поэтому ферментацию L-глутаминовой кислоты выполняют в нейтральных условиях (патенты США No.3220929 и 3032474; К.С. Chao and J.W. Foster, J. Bacteriol., 77, стр.715-725 (1959)). Таким образом, получение L-глутаминовой кислоты ферментацией, сопровождаемой осаждением, не известно. Более того, известно, что рост большинства ацидофильных бактерий ингибируется органическими кислотами, такими как уксусная кислота, молочная кислота и янтарная кислота (Yasuro Oshima Ed., "Extreme Environment Microorganism Handbook", стр.231, Science Forum; R.M. Borichewski, J. Bacteriol., 93, стр.597-599 (1967) и др.). Поэтому считается, что многие микроорганизмы в кислых условиях чувствительны к L-глутаминовой кислоте, которая также является органической кислотой, и не было сообщений о попытках поиска микроорганизмов, проявляющих способность продуцировать L-глутаминовую кислоту в кислых условиях.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Целью данного изобретения является способ получения L-глутаминовой кислоты ферментацией, который дает возможность эффективного получения L-глутаминовой кислоты даже в том случае, когда в качестве источника сахара используют материал, содержащий органическую кислоту, которая ингибирует рост микроорганизма при низком значении рН.
Авторы данного изобретения обнаружили, что при нейтральном значении рН бактерия, продуцирующая L-глутаминовую кислоту, потребляет органическую кислоту, которая ингибирует рост бактерии, продуцирующей L-глутаминовую кислоту, при низком значении рН, и что на основе данного свойства можно эффективно получать L-глутаминовую кислоту, используя в качестве источника сахара материал, содержащий органическую кислоту, которая ингибирует рост микроорганизма при низком рН. Таким образом, авторы осуществили данное изобретение.
Настоящее изобретение предоставляет следующее:
(1) Способ получения L-глутаминовой кислоты ферментацией, который включает культивирование микроорганизма, обладающего способностью продуцировать L-глутаминовую кислоту в среде, имеющей рН 5,0 или менее, в которой общее содержание органической кислоты, которая ингибирует рост микроорганизма при данном рН, является таким, при котором рост микроорганизма не ингибируется.
(2) Способ по п.(1), в котором L-глутаминовая кислота продуцируется и накапливается в среде с сопутствующим осаждением L-глутаминовой кислоты в течение культивирования в среде.
(3) Способ по п.(1) или (2), в котором общее содержание органической кислоты составляет 0,4 г/л или менее.
(4) Способ по любому из п.п. с (1) по (3), в котором органической кислотой является органическая кислота, имеющая число атомов углерода от 1 до 3.
(5) Способ по любому из п.п. с (1) по (4), в котором микроорганизм относится к роду Enterobacter.
(6) Способ по п.(5), в котором микроорганизмом является Enterobacter agglomerans.
(7) Способ по любому из п.п. с (1) по (6), в котором микроорганизм может метаболизировать источник углерода в жидкой среде, содержащей L-глутаминовую кислоту при концентрации насыщения и источник углерода, при определенном значении рН, и обладает способностью накапливать L-глутаминовую кислоту в количестве, превышающем концентрацию насыщения L-глутаминовой кислоты в жидкой среде при данном рН.
(8) Способ по п.(7), в котором значение рН составляет 5,0 или менее.
(9) Способ получения L-глутаминовой кислоты ферментацией, который включает культивирование микроорганизма, обладающего способностью продуцировать L-глутаминовую кислоту при первом значении рН, при котором рост микроорганизма не ингибируется органической кислотой в среде, и затем культивирование микроорганизма при втором значении рН, которое является подходящим для продуцирования микроорганизмом L-глутаминовой кислоты и является более низким, чем первое значение рН.
(10) Способ по п.(9), в котором органической кислотой является органическая кислота, имеющая количество атомов углерода от 1 до 3.
(11) Способ по п.(9) или (10), в котором второе значение рН составляет от 3,0 до 5,0.
(12) Способ по любому из п.п. с (9) по (11), в котором культивирование при первом значении рН выполняют, поддерживая рН среды на уровне первого значения рН добавлением к среде подщелачивающего вещества.
(13) Способ по п.(12), который включает снижение рН среды путем регулирования добавляемого количества подщелачивающего вещества после культивирования при первом рН.
(14) Способ по любому из п.п. с (9) по (13), в котором культивирование при первом рН продолжают до тех пор, пока не истощится органическая кислота в среде.
(15) Способ по любому из п.п. с (9) по (14), в котором микроорганизм относится к роду Enterobacter.
(16) Способ по п.(15), в котором микроорганизмом является Enterobacter agglormerans.
(17) Способ по любому из п.п. с (9) по (16), в котором микроорганизм может метаболизировать источник углерода в жидкой среде, содержащей L-глутаминовую кислоту при концентрации насыщения и источник углерода при определенном значении рН, и обладает способностью накапливать L-глутаминовую кислоту в количестве, превышающем концентрацию насыщения L-глутаминовой кислоты в жидкой среде при данном рН.
(18) Способ по п.(17), в котором значение рН составляет 5,0 или менее.
(19) Способ по п.(17) или (18), в котором значением рН, подходящим для продуцирования L-глутаминовой кислоты, является значение, при котором L-глутаминовая кислота, продуцируемая микроорганизмом, выпадает в осадок в среде, и L-глутаминовая кислота продуцируется и накапливается при сопутствующем осаждении L-глутаминовой кислоты во время культивирования в среде при данном значении рН.
Согласно способам настоящего изобретения, L-глутаминовую кислоту можно эффективно получать даже, когда в качестве источника сахара используют материал, содержащий органическую кислоту, которая ингибирует рост микроорганизма, такой как меласса.
КРАТКОЕ ПОЯСНЕНИЕ К ЧЕРТЕЖАМ
Фиг.1 представляет собой рестрикционную карту фрагмента ДНК, полученного из Enterobacter agglomerans, в pTWVEK101.
На фиг.2 показано сравнение аминокислотной последовательности, выведенной на основе нуклеотидной последовательности гена sucA, полученного из Enterobacter agglomerans, и гена sucA, полученного из Escherichia coli (наверху строка:
Enterobacter agglomerans, в графе Escherichia coli, то же самое применимо к последующим фигурам).
На фиг.3 показано сравнение аминокислотной последовательности, выведенной на основе нуклеотидной последовательности гена sucB, полученного из Enterobacter agglomerans, и гена sucB, полученного из Escherichia coli.
На фиг.4 показано сравнение аминокислотной последовательности, выведенной на основе нуклеотидной последовательности гена sucC, полученного из Enterobacter agglomerans, и гена sucC, полученного из Escherichia coli.
На фиг.5 показано сравнение аминокислотной последовательности, выведенной на основе нуклеотидной последовательности гена sdhB, полученного из Enterobacter agglomerans, и гена sdhB, полученного из Escherichia coli.
На фиг.6 показана конструкция плазмиды pMWCPG, содержащей ген gltA, ген ррс и ген gdhA.
На фиг.7 показана конструкция плазмиды RSF-Tet, содержащей начало репликации плазмиды RSF1010 широкого круга хозяев и ген устойчивости к тетрациклину.
На фиг.8 показана конструкция плазмиды RSFCPG, содержащей начало репликации плазмиды RSF1010 широкого круга хозяев, ген устойчивости к тетрациклину, ген gltA, ген ррс и ген gdhA.
На фиг.9 показана конструкция плазмиды pSTVCB, содержащей ген glta.
На фиг.10 показано ингибирующее рост действие различных органических кислот при кислом рН (рН 4,5).
На фиг.11 показаны результаты исследования рН, при котором вызывается ингибирование роста, с использованием муравьиной кислоты.
На фиг.12 показаны результаты исследования концентрации муравьиной кислоты, которая вызывает ингибирование роста в условиях рН 4,5.
На фиг.13 показан временной ход роста клеток во время культивирования.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Ниже будет дано подробное объяснение изобретения.
Способ получения согласно первому варианту данного изобретения (далее называемый также "первым способом получения согласно данному изобретению") представляет способ получения L-глутаминовой кислоты ферментацией, который включает культивирование микроорганизма, обладающего способностью продуцировать L-глутаминовую кислоту (далее называемого также "бактерией, продуцирующей L-глутаминовую кислоту") в среде, имеющей значение рН 5,0 или менее, в которой общее содержание органической кислоты, которая ингибирует рост микроорганизма при данном рН, равно содержанию, при котором рост микроорганизма не ингибируется.
В первом способе получения согласно данному изобретению предпочтительно, чтобы L-глутаминовая кислота продуцировалась и накапливалась при культивировании в среде с сопутствующим осаждением L-глутаминовой кислоты. Этого можно достичь доведением рН среды до значения рН, при котором продуцируемая L-глутаминовая кислота выпадает в осадок. Такое значение рН обычно составляет от 3,0 до 5,0. Считается, что при получении L-глутаминовой кислоты ферментацией снижение продуктивности L-глутаминовой кислотой, накапливаемой в среде в высокой концентрации, создает препятствие для повышения продуктивности. Например, клетка микроорганизма обладает системой выделения и системой поглощения L-глутаминовой кислоты, и если L-глутаминовая кислота, однажды выделенная в среду, снова поглощается клеткой, не только снижается эффективность продуцирования, но это также приводит к ингибированию реакций биосинтеза L-глутаминовой кислоты. Путем проведения культивирования при рН, при котором продуцируемая L-глутаминовая кислота выпадает в осадок, можно устранить такое снижение продуктивности вследствие накопления L-глутаминовой кислоты в высокой концентрации.
Величина рН среды предпочтительно составляет 4,5 или менее, более предпочтительно 4,0 или меньше.
В данном варианте воплощения под органической кислотой, которая ингибирует рост микроорганизма при некотором рН среды, подразумевают органическую кислоту, которая оказывает ингибирующее действие на ингибирование роста микроорганизма, когда она присутствует при определенной концентрации (обычно 0,5 г/л или более) в среде при данном рН, и обычно ею является органическая кислота, имеющая число атомов углерода от 1 до 3, т.е., муравьиная кислота, уксусная кислота или пропионовая кислота.
Общее содержание органической кислоты предпочтительно составляет 0,4 г/л или менее, более предпочтительно 0,3 г/л или менее, более предпочтительно 0,2 г/л или менее.
Способ получения согласно второму варианту данного изобретения (далее называемый также "вторым способом получения согласно данному изобретению") представляет собой способ получения L-глутаминовой кислоты ферментацией, который включает культивирование микроорганизма, обладающего способностью продуцировать L-глутаминовую кислоту при первом значении рН, при котором рост микроорганизма не ингибируется органической кислотой в среде, и затем культивирование микроорганизма при втором значении рН, которое является подходящим для продуцирования L-глутаминовой кислоты микроорганизмом и более низким, чем первое значение рН.
Обнаружено, что бактерия, продуцирующая L-глутаминовую кислоту, обычно претерпевает ингибирование роста органической кислотой в кислых условиях, тогда как она могла бы потреблять органическую кислоту в нейтральных условиях. На основе данного свойства, осуществляя выращивание клеток при нейтральном рН и затем, изменяя рН на кислое значение рН, чтобы продуцировать L-глутаминовую кислоту, становится возможным получение более высокой продуктивности, а также становится возможным применение в качестве источника сахара различных материалов.
В данном варианте под органической кислотой подразумевают органическую кислоту, которая обладает ингибирующим действием на рост микроорганизма, когда она присутствует в определенной концентрации (обычно 0,5 г/л или более) в среде при втором значении рН, и обычно ею является органическая кислота, имеющая количество атомов углерода от 1 до 3, т.е., муравьиная кислота, уксусная кислота или пропионовая кислота.
Первое значение рН и второе значение рН выбирают так, чтобы они соответствовали свойствам используемой бактерии, продуцирующей L-глутаминовую кислоту. Специалисты в данной области легко могут измерить указанные значения рН. Например, значение рН, при котором органическая кислота в среде не вызывает ингибирования роста микроорганизма, можно определить культивированием бактерии, продуцирующей L-глутаминовую кислоту, в средах, содержащих органическую кислоту, рН которых доведено до разных значений, измерением количества клеток на основе поглощения или тому подобным образом и сравнением количества клеток с количеством клеток бактерии, продуцирующей L-глутаминовую кислоту, культивируемой в таких же условиях, за исключением того, что при этом среды не содержат органической кислоты. К рН, подходящему для продуцирования L-глутаминовой кислоты, относится значение рН, при котором L-глутаминовая кислота накапливается в среде, определяемое при культивировании бактерии, продуцирующей L-глутаминовую кислоту, в средах с различными значениями рН. В частности, значение можно определить путем измерения количества L-глутаминовой кислоты, накапливаемой в средах при разных значениях рН и их сравнения.
Первое значение рН особо не ограничивается, при условии, что органическая кислота не ингибирует рост микроорганизма в среде, но обычно составляет от 5,0 до 8,0.
Вторым значением рН предпочтительно является рН, при котором продуцируемая L-глутаминовая кислота выпадает в осадок, и такое значение рН обычно составляет от 3,0 до 5,0. Как объяснялось для первого способа получения согласно данному изобретению, снижения продуктивности при накоплении L-глутаминовой кислоты в высокой концентрации можно избежать, проводя культивирование при рН, при котором продуцируемая L-глутаминовая кислота выпадает в осадок.
Первое значение рН и второе значение рН могут не быть строго постоянными в ходе культивирования, при условии, что могут быть достигнуты преимущества данного изобретения, и значения могут колебаться.
Бактерия, продуцирующая L-глутаминовую кислоту, продуцирует L-глутаминовую кислоту даже при первом рН, и поэтому рН понижается продуцируемой L-глутаминовой кислотой. Поэтому культивирование при первом рН предпочтительно осуществляют при поддержании рН среды на уровне первого значения рН добавлением в среду подщелачивающего вещества.
Хотя подщелачивающее вещество особым образом не ограничивается, при условии, что оно не оказывает неблагоприятного воздействия на рост бактерии, продуцирующей L-глутаминовую кислоту или на продуцирование L-глутаминовой кислоты, предпочтителен газообразный аммиак.
рН среды можно понизить от первого рН до второго рН добавлением кислого вещества. Однако рН в ходе культивирования понижается L-глутаминовой кислотой, продуцируемой бактерией, продуцирующей L-глутаминовую кислоту, как описано выше. Поэтому предпочтительно снижать рН среды от первого значения рН до второго значения рН посредством регулирования добавляемого количества подщелачивающего вещества, так как добавление кислого вещества можно исключить.
Культивирование при первом рН можно продолжать до тех пор, пока не истощится органическая кислота в среде. Истощение означает, что количество органической кислоты снижается до уровня, при котором рост бактерии, продуцирующей L-глутаминовую кислоту, не ингибируется в ходе культивирования при втором рН. Специалисты в данной области легко могут измерить такой уровень органической кислоты. Например, уровень можно определить посредством культивирования бактерии, продуцирующей L-глутаминовую кислоту, в средах, содержащих органическую кислоту в разных концентрациях, при втором значении рН, измерением количеств клеток бактерии, продуцирующей L-глутаминовую кислоту, и сравнения количеств клеток с количествами клеток бактерии, продуцирующей L-глутаминовую кислоту, культивируемой при таких условиях, за исключением того, что при этом среды не содержат органической кислоты. Как правило, по мере того как второе значение рН становится ниже, уровень органической кислоты также понижается.
Бактерия, продуцирующая L-глутаминовую кислоту, используемая в первом способе получения согласно данному изобретению и во втором способе получения согласно данному изобретению, представляет собой микроорганизм, который накапливает значительное количество L-глутаминовой кислоты в среде при его культивировании в среде. Примеры такой бактерии включают микроорганизмы, относящиеся к роду Enterobacter. Предпочтительной является Enterobacter agglomerans.
Кроме того, бактерией, продуцирующей L-глутаминовую кислоту, используемой в первом и втором способах получения согласно изобретению, предпочтительно является микроорганизм, который может метаболизировать источник углерода в жидкой среде, содержащей L-глутаминовую кислоту в концентрации насыщения и источник углерода, при определенном рН, и обладает способностью накапливать L-глутаминовую кислоту в количестве, превышающем концентрацию насыщения L-глутаминовой кислоты в жидкой среде при вышеупомянутом значении рН (далее называемый также "микроорганизмом, накапливающим L-глутаминовую кислоту"). Вышеуказанное определенное значение рН предпочтительно равно рН, при котором L-глутаминовая кислота выпадает в среде в осадок, и такое значение рН обычно составляет 5,0 или менее.
Под "концентрацией насыщения" подразумевается концентрация L-глутаминовой кислоты, растворенной в жидкой среде, когда жидкая среда насыщена L-глутаминовой кислотой.
В случае использования микроорганизма, накапливающего L-глутаминовую кислоту, значение рН, подходящее для продуцирования L-глутаминовой кислоты, предпочтительно равно рН, при котором L-глутаминовая кислота выпадает в среде в осадок. При осуществлении культивирования при данном рН L-глутаминовая кислота продуцируется и накапливается в среде при сопутствующем выпадении L-глутаминовой кислоты в осадок.
Микроорганизм, накапливающий L-глутаминовую кислоту, можно получить следующим образом. Образец, содержащий микроорганизмы, инокулируют в жидкую среду, содержащую L-глутаминовую кислоту в концентрации насыщения и источник углерода, при определенном значении рН, и отбирают штамм, который метаболизирует источник углерода. Хотя определенное значение рН особым образом не ограничивается, обычно оно составляет примерно 5,0 или меньше, предпочтительно примерно 4,5 или меньше, более предпочтительно примерно 4,3 или меньше. Микроорганизм, накапливающий L-глутаминовую кислоту, используют для получения L-глутаминовой кислоты ферментацией с сопутствующим выпадением L-глутаминовой кислоты в осадок. Если значение pH является слишком высоким, становится трудно обеспечить возможность для того, чтобы микроорганизм продуцировал L-глутаминовую кислоту в количестве, достаточном для осаждения. Поэтому рН предпочтительно находится в указанных выше пределах.
Если рН водного раствора, содержащего L-глутаминовую кислоту, понижают, растворимость L-глутаминовой кислоты в значительной степени снижается около рКа γ-карбоксильной группы (4,25, 25°С). Растворимость становится наименьшей в изоэлектрической точке (рН 3,2), и L-глутаминовая кислота, превышающая количество, соответствующее концентрации насыщения, выпадает в осадок. Хотя это зависит от состава среды, L-глутаминовую кислоту растворяют в количестве 10-20 г/л при рН 3,2, 30-40 г/л при рН 4,0 и 50-60 г/л при рН 4,7, примерно при 30°С. Обычно не требуется делать рН 3,0 или ниже, поскольку преципитирующее действие на L-глутаминовую кислоту достигает своего верхнего предела, когда рН становится ниже определенного значения. Однако рН может быть 3,0 или меньше.
Кроме того, выражение, что микроорганизм "может метаболизировать источник углерода" означает, что он может пролиферировать или может потреблять источник углерода, даже хотя он не может пролиферировать, то есть, выражение указывает, что микроорганизм катаболизирует источник углерода, такой как сахара или органические кислоты. В частности, например, если микроорганизм пролиферирует при его культивировании в жидкой среде, содержащей L-глутаминовую кислоту в концентрации насыщения при рН от 5,0 до 4,0, предпочтительно при рН от 4,5 до 4,0, более предпочтительно при рН от 4,3 до 4,0, особенно предпочтительно при рН 4,0 при соответствующей температуре, например, 28°С, 37°С или 50°С в течение от 2 до 4 дней, указанный микроорганизм может метаболизировать источник углерода в среде. Кроме того, например, если микроорганизм потребляет источник углерода даже несмотря на то, что микроорганизм не пролиферирует при культивировании в синтетической жидкой среде, содержащей L-глутаминовую кислоту в концентрации насыщения при рН от 5,0 до 4,0, предпочтительно при рН от 4,5 до 4,0, более предпочтительно при рН от 4,3 до 4,0, особенно предпочтительно при рН 4,0 при соответствующей температуре, например, 28°С, 37°С или 50°С в течение от 2 до 4 дней, указанным микроорганизмом является микроорганизм, который может метаболизировать источник углерода в среде.
Микроорганизмом, который может метаболизировать источник углерода, является микроорганизм, который может расти в вышеуказанной жидкой среде.
Кроме того, выражение, что микроорганизм "может расти", означает, что он может пролиферировать или может продуцировать L-глутаминовую кислоту даже несмотря на то, что он не может пролиферировать. В частности, например, если микроорганизм пролиферирует при культивировании в жидкой среде, содержащей L-глутаминовую кислоту в концентрации насыщения при рН от 5,0 до 4,0, предпочтительно при рН от 4,5 до 4,0, более предпочтительно при рН от 4,3 до 4,0, особенно предпочтительно при рН 4,0 при соответствующей температуре, например, 28°С, 37°С или 150°С в течение от 2 до 4 дней, указанный микроорганизм может расти в среде. Кроме того, например, если микроорганизм увеличивает количество L-глутаминовой кислоты в синтетической жидкой среде даже, несмотря на то, что микроорганизм не пролиферирует при культивировании микроорганизма в синтетической жидкой среде, содержащей L-глутаминовую кислоту в концентрации насыщения при рН от 5,0 до 4,0, предпочтительно при рН от 4,5 до 4,0, более предпочтительно при рН от 4,3 до 4,0, особенно предпочтительно при рН 4,0 при соответствующей температуре, например, 28°С, 37°С или 50°С в течение от 2 до 4 дней, указанным микроорганизмом является микроорганизм, который может расти в среде.
Отбор, описанный выше, можно повторить два или несколько раз при таких же условиях или с изменением рН или концентрации L-глутаминовой кислоты. Отбор на ранней стадии можно осуществлять в среде, содержащей L-глутаминовую кислоту в концентрации более низкой, чем концентрация насыщения, и после этого последующий отбор можно проводить в среде, содержащей L-глутаминовую кислоту в концентрации насыщения. Далее можно отобрать штаммы с предпочтительными свойствами, такими как самая высокая скорость пролиферации.
Микроорганизм, накапливающий L-глутаминовую кислоту, представляет собой микроорганизм, который в дополнение к описанным выше свойствам обладает способностью накапливать L-глутаминовую кислоту в количестве, превышающем количество, соответствующее концентрации насыщения L-глутаминовой кислоты в жидкой среде. Значение рН вышеупомянутой жидкой среды предпочтительно такое же или близко к значению рН среды, используемой для скрининга микроорганизма, обладающего вышеуказанными свойствами. Обычно микроорганизм становится чувствительным к L-глутаминовой кислоте при высокой концентрации по мере того, как рН снижается. Поэтому предпочтительно, чтобы рН не было низким с точки зрения устойчивости к L-глутаминовой кислоте, но низкое значение рН предпочтительно с точки зрения продукции L-глутаминовой кислоты с сопутствующим выпадением этой кислоты в осадок. Чтобы соответствовать указанным условиям, рН может быть в пределах от 3 до 5, предпочтительно от 4 до 5, более предпочтительно от 4 до 4,7, еще более предпочтительно от 4 до 4,5, особенно предпочтительно от 4,0 до 4,3.
В качестве микроорганизма, накапливающего L-глутаминовую кислоту, из материалов, полученных при селекции, или материалов селекции данного микроорганизма можно указать, например, микроорганизмы, относящиеся к роду Enterobacter, Klebsiella, Serratia, Pantoea, Erwinia, Escherichia, Corynebacterium, Alicyclobacillus, Bacillus, Saccharomyces и тому подобные. Из них предпочтительными являются микроорганизмы, относящиеся к роду Enterobacter. Далее объяснения по поводу микроорганизма согласно данному изобретению будут даны главным образом для микроорганизмов, относящихся к роду Enterobacter. Однако микроорганизм не ограничен организмами, относящимися к роду Enterobacter, и сходным образом можно использовать микроорганизмы, относящиеся к другим родам.
В качестве микроорганизма, относящегося к Enterobacter, в частности можно указать Enterobacter agglomerans, предпочтительно штамм AJ13355 Enterobacter agglomerans. Указанный штамм выделяли из почвы в Iwata-shi, Shizuoka, Japan в качестве штамма, который может пролиферировать в среде, содержащей L-глутаминовую кислоту и источник углерода, при низком рН.
Физиологические свойства AJ13355 показаны ниже:
(1) Окраска по Граму: отрицательная.
(2) Поведение в отношении кислорода: факультативный анаэроб.
(3) Каталаза: положительный.
(4) Оксидаза: отрицательный.
(5) Способность восстанавливать нитраты: отрицательный.
(6) Тест Voges-Proskauer: положительный.
(7) Тест с метиловым красным: отрицательный.
(8) Уреаза: отрицательный.
(9) Продукция индола: положительный.
(10) Подвижность: подвижный.
(11) Продукция H2S в среде TSI: слабо активен.
(12) В-Галактозидаза: положительный.
(13) Способность к ассимиляции сахаридов:
Арабиноза: положительный.
Сахароза: положительный.
Лактоза: положительный.
Ксилоза: положительный.
Сорбит: положительный.
Инозит: положительный.
Трегалоза: положительный.
Мальтоза: положительный.
Глюкоза: положительный
Адонит: отрицательный.
Раффиноза: положительный.
Салицин: отрицательный.
Мелибиоза: положительный
(14) Способность ассимилировать глицерозу: положительный.
(15) Способность ассимилировать органические кислоты:
Лимонная кислота: положительный.
Винная кислота: отрицательный.
Глюконовая кислота: положительный.
Уксусная кислота: положительный.
Малоновая кислота: отрицательный.
(16) Аргининдегидратаза: отрицательный.
(17) Орнитиндекарбоксилаза: отрицательный.
(18) Лизиндекарбоксилаза: отрицательный.
(19) Фенилаланиндезаминаза: отрицательный.
(20) Образование пигмента: желтый.
(21) Способность разжижать желатин: положительный.
(22) рН роста: рост возможен при рН 4, хорошо растет при рН от 4,5 до 7.
(23) Температура роста: хороший рост при 25°С, хороший рост при 30°С, хороший рост при 37°С, рост возможен при 42°С, рост невозможен при 45°С.
На основании указанных бактериологических свойств AJ13355 определили как Enterobacter agglomerans.
Штамм AJ13355 Enterobacter agglomerans был депонирован в National Institute of Bioscience and Human-Technology, Agency of Industrial Science and Technology, Ministry of International Trade and Industry (в настоящее время International Patent Organism Depositary, National Institute of Advanced Industrial Science and Technology) 19 февраля 1998 г. и получил инвентарный номер FERM Р-16644. Затем он был перенесен для международного депонирования по условиям Будапештского договора 11 января 1999 г. и получил инвентарный номер FERM BP-6614.
Микроорганизмом, накапливающим L-глутаминовую кислоту, может быть микроорганизм, исходно обладающий способностью продуцировать L-глутаминовую кислоту, или микроорганизм, обладающий способностью продуцировать L-глутаминовую кислоту, которая была ему придана или усилена при селекции благодаря использованию мутагенной обработки, технологии рекомбинантной ДНК или тому подобного.
Способность продуцировать L-глутаминовую кислоту можно придать или усилить, например, посредством увеличения активности фермента, который катализирует реакцию биосинтеза L-глутаминовой кислоты. Способность продуцировать L-глутаминовую кислоту также можно усилить посредством снижения или исключения активности фермента, который катализирует реакцию, которая является ответвлением пути биосинтеза L-глутаминовой кислоты и создает соединение, отличное от L-глутаминовой кислоты.
В качестве примеров фермента, который катализирует реакцию биосинтеза L-глутаминовой кислоты, можно указать глутаматдегидрогеназу (далее называемую "GDH"), глутаминсинтетазу, глутаматсинтазу, изоцитратдегидрогеназу, аконитатгидратазу, цитратсинтазу (далее также называемую "CS"), фосфоенолпируваткарбоксилазу (далее также называемую "РЕРС"), пируватдегидрогеназу, пируваткиназу, енолазу, фосфоглицеромутазу, фосфоглицераткиназу, глицеральдегид-3-фосфатдегидрогеназу, триозофосфатизомеразу, фруктозобифосфатальдолазу, фосфофруктокиназу, глюкозофосфатизомеразу и так далее. Среди этих ферментов предпочтительны один, два или три фермента CS, РЕРС и GDH. Кроме того, предпочтительно, чтобы активности всех трех ферментов CS, РЕРС и GDH были усилены у микроорганизма, накапливающего L-глутаминовую кислоту. В частности, предпочтительна CS Brevibacterium lactofermentum, поскольку она не подвержена ингибированию α-кетоглутаровой кислотой, L-глутаминовой кислотой и NADH.
Например, для того, чтобы усилить активность CS, РЕРС или GDH, ген, кодирующий CS, РЕРС или GDH можно клонировать в соответствующей плазмиде и полученной плазмидой можно трансформировать микроорганизм-хозяин. Количество копий гена, кодирующего CS, РЕРС или GDH (далее называемых сокращенно "ген gltA", "ген ррс" и "ген gdhA" соответственно) в трансформированных клетках штамма увеличивается, приводя в результате к увеличению активности CS, РЕРС или GDH.
Клонированные гены gltA, ррс и gdhA вводят в вышеуказанный исходный родительский штамм отдельно или в произвольной комбинации двух или трех их видов. В том случае, когда вводят два или три вида указанных генов, два или три вида генов можно клонировать в плазмиде одного вида и ввести в хозяина, или отдельно клонировать в двух или трех видах плазмид, которые могут существовать совместно, и ввести в хозяина.
Два или несколько видов генов, кодирующих фермент одного вида, но полученных из разных микроорганизмов, можно ввести в одного и того же хозяина.
Описанные выше плазмиды особым образом не ограничиваются, при условии, что они автономно реплицируются в клетке микроорганизма, относящегося, например, к роду Enterobacter или ему подобному. Однако можно указать, например, pUC19, pUC18, pBR322, pHSG299, pHSG298, pHSG399, pHSG398, RSF1010, pMW119, pMW118, pMW219, pMW218, pACYC177, pACYC184 и так далее. Кроме указанного можно также использовать векторы фаговой ДНК.
Трансформацию можно проводить, например, способом D.M. Morrison (Methods in Enzymology, 68, 326 (1979)), способом, при котором проницаемость для ДНК бактериальных клеток-реципиентов увеличивают обработкой клеток хлоридом кальция (Mandel M and Higa A., J. Mol. Biol., 53, 159 (1970)), электропорацией (Miller J. H., "A Short Course in Bacterial Genetics", Cold Spring Harbor Laboratory Press, U.S.A., 1992) и тому подобное.
Активность CS, РЕРС или GDH также можно увеличить, обеспечивая возможность для присутствия многочисленных копий гена gltA, гена ррс или гена gdhA в хромосомной ДНК вышеуказанного исходного родительского штамма, который будет хозяином. Для того чтобы ввести множественные копии гена gltA, гена ррс или гена gdhA в хромосомную ДНК микроорганизма, относящегося к роду Enterobacter или ему подобного, можно использовать последовательность, множественные копии которой присутствуют в хромосомной ДНК, такую как повторяющаяся ДНК и инвертированные повторы, присутствующие на концах транспозируемого элемента. В альтернативном случае множественные копии генов можно вводить в хромосомную ДНК, используя перенос транспозона, содержащего ген gltA, ген ррс или ген gdhA. В результате увеличивается количество копий гена gltA, гена ррс или гена gdhA в трансформированной линии клеток, и поэтому увеличивается активность CS, РЕРС или GDH.
В качестве организмов, используемых в качестве источника гена gltA, гена ррс или гена gdhA, количество копий которых требуется увеличить, можно использовать любой организм, при условии, что он обладает активностью CS, РЕРС или GDH. В числе других предпочтительны бактерии, которые являются прокариотами, например, бактерии, относящиеся к роду Enterobacter, Klebsiella, Erwinia, Pantoea, Serratia, Escherichia, Corynebacterium, Brevibacterium или Bacillus. В качестве конкретных примеров можно указать Escherichia coli, Brevibacterium lactofermentum и так далее. Ген gltA, ген ррс или ген gdhA можно получить из хромосомной ДНК описанных выше микроорганизмов.
Ген gltA, ген ррс и ген gdhA можно получить с использованием мутантного штамма, который дефицитен по активности CS, РЕРС или GDH, чтобы выделить фрагмент ДНК, который комплементирует их ауксотрофию, из хромосомной ДНК вышеуказанного микроорганизма. Кроме того, так как нуклеотидные последовательности указанных генов бактерий Escherichia и Corynebacterium уже выяснены (Biochemistry, 22, pp.5243-5249, (1983); J. Biochem., 95 pp.909-916, (1984); Gene, 27, pp.193-199, (1984); Microbiology, 140, pp.1817-1828, (1994); Mol. Gen. Genet., 218, pp.330-339, (1989); Molecular Microbiology, 6, pp.317-326, (1992)), эти гены также можно получить посредством ПЦР с использованием праймеров, синтезированных на основе каждой нуклеотидной последовательности, и хромосомной ДНК в качестве матрицы.
Кроме вышеуказанной амплификации генов, активность CS, РЕРС или GDH также можно увеличить, усиливая экспрессию гена gltA, гена ррс или гена gdhA. Например, экспрессию можно усилить заменой промотора гена gltA, гена ррс или гена gdhA другим более сильным промотором. Например, в качестве сильных промоторов известны промотор lac, промотор trp, промотор trc, промотор tac, промотор PR и промотор PL фага лямбда и так далее. Ген gltA, ген ррс или ген gdhA, промоторы которых заменены, клонируют в плазмиде и вводят в микроорганизм-хозяин, или вводят в хромосомную ДНК микроорганизма-хозяина, используя повторяющуюся ДНК, инвертированный повтор, транспозон или тому подобное.
Активность CS, РЕРС или GDH также можно увеличить, заменяя промотор гена gltA, гена ррс или гена gdhA на хромосоме другим более сильным промотором (смотри W087/03006 и выложенную заявку на патент Японии No.61-268183) или, встраивая сильный промотор выше кодирующей последовательности каждого гена (смотри Gene, 29, pp.231-241 (1984)). В частности, можно выполнить гомологичную рекомбинацию между геном gltA, геном ррс или геном gdhA, промотор которого заменяют более сильным промотором или ДНК, содержащей его часть, и соответствующим геном в хромосоме.
Примеры фермента, который катализирует реакцию, которая является ответвлением пути биосинтеза L-глутаминовой кислоты и создает соединение, отличное от L-глутаминовой кислоты, включают α-кетоглутаратдегидрогеназу (далее называемую "αKGDH"), изоцитратлиазу, фосфатацетилтрансферазу, ацетаткиназу, синтазу ацетогидроксикислоты, ацетолактатсинтазу, формиатацетилтрансферазу, лактатдегидрогеназу, глутаматдекарбоксилазу, 1-пирролиндегидрогеназу и так далее. Среди указанных ферментов предпочтительна α-KGDH.
Для того чтобы снизить или исключить активности вышеуказанных ферментов у микроорганизма, относящегося к роду Enterobacter или ему подобному, можно ввести мутации для снижения или элиминации внутриклеточной активности ферментов в гены вышеуказанных ферментов обычным способом мутагенной обработки или способом генетической инженерии.
Примеры способа мутагенной обработки включают, например, способы с использованием облучения рентгеновским излучением или ультрафиолетовым излучением, способы с использованием мутагенных агентов, таких как N-метил-N'-нитро-N-нитрозогуанидин и так далее. Сайт в гене, в который вводят мутацию, может быть в кодирующем районе, который кодирует ферментный белок, или в районе для регуляции экспрессии, таком как промотор.
Примеры способов генетической инженерии включают, например, способы с использованием рекомбинации генов, трансдукции, слияния клеток и так далее. Например, ген устойчивости к лекарственному препарату встраивают в клонированный ген-мишень для того, чтобы получить ген, который потерял свою функцию (дефектный ген). Затем указанный дефектный ген вводят в клетку микроорганизма-хозяина и заменяют ген-мишень в хромосоме вышеуказанным дефектным геном с использованием гомологичной рекомбинации (нарушение гена).
Снижение или отсутствие внутриклеточной активности фермента-мишени и степень снижения активности можно подтвердить измерением активности фермента в экстракте клеток или в его очищенной фракции, полученных из штамма-кандидата, и сравнением с активностью в штамме дикого типа. Например, активность αKGDH можно измерить способом Reed et al. (Reed L.J. and Mukherjee B.B., Methods in Enzymology, 13, pp.55-61 (1969)).
В зависимости от фермента-мишени мутантный штамм-мишень можно отобрать на основе фенотипа мутантного штамма. Например, мутантный штамм, в котором активность αKGDH элиминирована или снижена, не может пролиферировать или проявляет заметно сниженную скорость пролиферации в минимальной среде, содержащей глюкозу, или в минимальной среде, содержащей уксусную кислоту или L-глутаминовую кислоту в качестве единственного источника углерода в аэробных условиях культивирования. Однако нормальная пролиферация возможна даже в таких условиях при добавлении янтарной кислоты или лизина, метионина и диаминопимелиновой кислоты к минимальной среде, содержащей глюкозу. Используя указанные явления в качестве индикаторов, можно отобрать мутантный штамм с пониженной активностью αKGDH или дефицитный по активности αKGDH.
Способ получения штамма Brevibacterium lactofermentum, дефицитного по гену αKGDH, с использованием гомологичной рекомбинации подробно описан в W095/34672. Подобные способы можно применять для других микроорганизмов.
Кроме того, способы, такие как клонирование генов и расщепление и лигирование ДНК, трансформация и так далее, подробно описаны в Molecular Cloning, 2nd Edition, Cold Spring Harbor Press (1989) и так далее.
В качестве конкретного примера мутантного штамма, дефицитного по активности αKGDH или с пониженной активностью αKGDH, полученного как описано выше, можно указать AJ13356 Enterobacter agglomerans. Штамм AJ13356 Enterobacter agglomerans был депонирован в National Institute of Bioscience and Human-Technology, Agency of Industrial Science and Technology, Ministry of International Trade and Industry (в настоящее время International Patent Organism Depositary, National Institute of Advanced Industrial Science and Technology) 19 февраля 1998 г. и получил инвентарный номер FERM P-16645. Затем он был перенесен для международного депонирования по условиям Будапештского договора 11 января 1999 г. и получил инвентарный номер FERM ВР-6615. Штамм AJ13356 Enterobacter agglomerans дефицитен по активности αKGDH в результате нарушения гена субъединицы αKGDH-E1 (sucA).
В том случае, когда Enterobacter agglomerans, которая является примером микроорганизма, используемого в данном изобретении, культивируют в среде, содержащей сахарид, во внеклеточное пространство секретируется слизь, что в результате иногда приводит к низкой производительности. Поэтому когда используют Enterobacter agglomerans, обладающую таким свойством секретировать слизь, предпочтительно использовать мутантный штамм, который секретирует меньше слизи по сравнению со штаммом дикого типа. Примеры мутагенной обработки включают, например, способы с использованием облучения рентгеновским излучением или ультрафиолетовым излучением, способы с использованием обработки мутагенными агентами, такими как N-метил-N'-нитро-N-нитрозогуанидин и так далее. Мутантный штамм с пониженной секрецией слизи можно отобрать посредством инокуляции мутагенизированных бактериальных клеток в среду, содержащую сахарид, например, в чашку со средой LB, содержащей 5 г/л глюкозы, их культивирования в чашках под наклоном примерно в 45 градусов и отбора колонии, с которой не наблюдается стекание слизи.
В данном изобретении придание или усиление способности продуцировать L-глутаминовую кислоту и придание других подходящих свойств, таких как мутация для меньшей секреции слизи, описанная выше, можно осуществлять в произвольно выбранном порядке.
При культивировании микроорганизма, накапливающего L-глутаминовую кислоту, в жидкой среде, значение рН которой доводят до условия рН, которое делает возможным осаждение L-глутаминовой кислоты, L-глутаминовая кислота может продуцироваться и накапливаться при сопутствующем осаждении этой кислоты в среде.
Термин "условие, которое делает возможным осаждение L-глутаминовой кислоты, продуцируемой микроорганизмом" по отношению к данному изобретению означает условие, которое делает возможным осаждение L-глутаминовой кислоты в том случае, когда микроорганизм, накапливающий L-глутаминовую кислоту, продуцирует и накапливает L-глутаминовую кислоту. Хотя значение рН указанного условия может варьировать в зависимости от способности микроорганизма продуцировать L-глутаминовую кислоту, обычно оно составляет от 3 до 5 в том случае, когда микроорганизмом является бактерия Enterobacter.
В качестве сред, используемых для культивирования в первом способе получения согласно данному изобретению, культивирования при первом рН и культивирования при втором рН во втором способе получения согласно данному изобретению, можно использовать обычную питательную среду, содержащую источник углерода, источник азота, минеральные соли и следовые количества таких органических питательных веществ, как аминокислоты и витамины, которые необходимы, при условии, что значение рН доводят так, чтобы оно соответствовало предварительно определенному условию. Можно использовать или синтетическую среду, или природную среду. Источником углерода и источником азота, используемыми в среде, могут быть любые источники, при условии, что штамм, который необходимо культивировать, может их использовать.
В качестве источника углерода используют сахариды, такие как глюкоза, глицерин, фруктоза, сахароза, мальтоза, манноза, галактоза, гидролизат крахмала и меласса. Кроме того, можно использовать органические кислоты, такие как уксусная кислота и лимонная кислота, каждую в отдельности или в комбинации с другим источником углерода.
В качестве источника азота используют аммиак, соли аммония, такие как сульфат аммония, карбонат аммония, хлорид аммония, фосфат аммония и ацетат аммония, нитраты и так далее.
В качестве органических питательных веществ, используемых в следовых количествах, используют аминокислоты, витамины, жирные кислоты, нуклеиновые кислоты, питательные компоненты, содержащие указанные вещества, такие как пептон, казаминовая кислота, дрожжевой экстракт и продукты разложения соевого белка. В том случае, когда используют ауксотрофный мутантный штамм, который требует аминокислоты и так далее для метаболизма или роста, необходимо добавлять требуемое питательное вещество.
В качестве минеральных солей используют фосфаты, соли магния, соли кальция, соли железа, соли марганца и так далее.
Что касается способа культивирования, обычно выполняют культивирование в условиях аэрации при температуре от 20 до 42°С, при условии, что контролируют рН, чтобы оно было на уровне предварительно определенного значения.
После завершения культивирования осажденную в культуре L-глутаминовую кислоту можно собрать центрифугированием, фильтрацией и тому подобным способом. L-глутаминовую кислоту, растворенную в среде, также можно собрать известными способами. Например, L-глутаминовую кислоту можно выделить концентрированием культурального бульона для того, чтобы вызвать кристаллизацию L-глутаминовой кислоты, или выделить ионообменной хроматографией или тому подобными способами. Также можно кристаллизовать L-глутаминовую кислоту, растворенную в среде и затем собирать L-глутаминовую кислоту, выпавшую в осадок в культуральном бульоне, вместе с кристаллизованной L-глутаминовой кислотой.
В варианте, где L-глутаминовая кислота, превышающая концентрацию насыщения, выпадает в осадок, концентрация L-глутаминовой кислоты, растворенной в среде, поддерживается на постоянном уровне. Поэтому влияние L-глутаминовой кислоты в высокой концентрации на микроорганизмы может быть снижено. Соответственно также открывается возможность для селекции микроорганизма, обладающего дополнительной повышенной способностью продуцировать L-глутаминовую кислоту. Кроме того, так как L-глутаминовая кислота осаждается в виде кристаллов, подавляется закисление культурального бульона за счет накопления L-глутаминовой кислоты, и поэтому количество щелочи, используемой для поддержания рН культуры, может быть значительно уменьшено.
ПРИМЕРЫ
Ниже будет дано более конкретное объяснение данного изобретения со ссылками на следующие примеры. В примерах аминокислоты представляют собой L-аминокислоты, если не оговорено особо.
Справочный пример 1
<1> Скрининг микроорганизма, обладающего устойчивостью к L-глутаминовой кислоте в кислых условиях среды.
Скрининг микроорганизма, обладающего устойчивостью к L-глутаминовой кислоте в кислых условиях среды, выполняли следующим образом. Один (1) грамм каждого из примерно 500 образцов, полученных в природе, включая почву, фрукты, организм растений, речную воду и так далее, суспендировали в 5 мл стерилизованной воды, и 200 мкл образца наносили слоем на 20 мл твердой среды, рН которой доводили до рН 4,0 с помощью HCl. Состав среды был следующим: 3 г/л глюкозы, 1 г/л сульфата аммония, 0,2 г/л гептагидрата сульфата магния, 0,5 г/л однозамещенного фосфата калия, 0,2 г/л хлорида натрия, 0,1 г/л дигидрата хлорида кальция, 0,01 г/л гептагидрата сульфата железа, 0,01 г/л тетрагидрата сульфата марганца, 0,72 мг/л дигидрата сульфата цинка, 0,64 мг/л пентагидрата сульфата меди, 0,72 мг/л гексагидрата хлорида кобальта, 0,4 мг/л борной кислоты, 1,2 мг/л дигидрата молибдата натрия, 50 мкг/л биотина, 50 мкг/л пантотената кальция, 50 мкг/мл фолиевой кислоты, 50 мкг/мл инозита, 50 мкг/л ниацина, 50 мкг/л парааминобензойной кислоты, 50 мкг/л гидрохлорида пиридоксина, 50 мкг/л рибофлавина, 50 мкг/л гидрохлорида тиамина, 50 мг/л циклогексимида и 20 г/л агара.
Среды, покрытые слоем указанных выше образцов, инкубировали при 28°С, 37°С или 50°С в течение от 2 до 4 дней и получили 378 штаммов, образующих колонии.
Затем каждый из штаммов, полученных как описано выше, инокулировали в пробирку длиной 16, 5 см и диаметром 14 мм, содержащую 3 мл жидкой среды (рН доводили до 4,0 с помощью HCl), содержащей L-глутаминовую кислоту в концентрации насыщения, и культивировали при 28°С, 37°С или 50°С в течение от 24 часов до 3 дней при встряхивании. Затем отбирали выросшие штаммы. Состав вышеуказанной среды был следующим: 40 г/л глюкозы, 20 г/л сульфата аммония, 0,5 г/л гептагидрата сульфата магния, 2 г/л однозамещенного фосфата калия, 0,5 г/л хлорида натрия, 0,25 г/л дигидрата хлорида кальция, 0,02 г/л гептагидрата сульфата железа, 0,02 г/л тетрагидрата сульфата марганца, 0,72 мг/л дигидрата сульфата цинка, 0,64 мг/л пентагидрата сульфата меди, 0,72 мг/л гексагидрата хлорида кобальта, 0,4 мг/л борной кислоты, 1,2 мг/л дигидрата молибдата натрия и 2 г/л дрожжевого экстракта.
Таким образом, успешно получили 78 штаммов микроорганизмов, проявляющих устойчивость к L-глутаминовой кислоте в кислых условиях среды.
<2> Отбор штаммов, проявляющих самую высокую скорость роста, из микроорганизмов, обладающих устойчивостью к L-глутаминовой кислоте в кислых условиях среды.
Различные микроорганизмы, обладающие устойчивостью к L-глутаминовой кислоте в кислых условиях среды, полученные как описано выше, инокулировали каждый в пробирку длиной 16,5 см и диаметром 14 мм, содержащую 3 мл среды (рН доводили до 4,0 с помощью HCl), полученной добавлением 20 г/л L-глутаминовой кислоты и 2 г/л глюкозы в среду М9 (Sambrook, J. Fritsh, E.F. and Maniatis, Т. "Molecular Cloning", Cold Spring Harbor Laboratory Press, USA, 1989), и измеряли мутность среды в течение времени, чтобы отобрать штаммы, проявляющие подходящую скорость роста. В результате в качестве штамма, проявляющего подходящий рост, получили штамм AJ13355 из почвы в Iwata-shi, Shizuoka, Japan. Указанный штамм определили как Enterobacter agglomerans на основании его бактериологических свойств, описанных выше.
<3> Получение штамма с меньшей секрецией слизи из штамма AJ13355 Enterobacter agglomerans.
Так как штамм AJ13355 Enterobacter agglomerans секретирует слизь во внеклеточное пространство при культивировании в среде, содержащей сахарид, его производительность не достаточно подходит. Поэтому получили штамм, который секретирует меньше слизи, способом облучения ультрафиолетом (Miller, J.H. et al., "A Short Course in Bacterial Genetics; Laboratory Manual", P. 150, 1992, Cold Spring Harbor Laboratory Press, USA).
Штамм AJ13355 Enterobacter agglomerans облучали ультрафиолетовым излучением в течение 2 минут в положении на расстоянии 60 см от ультрафиолетовой лампы мощностью 60 Вт и культивировали в среде LB в течение ночи, чтобы закрепить мутацию. Мутагенизированный штамм разбавляли и инокулировали в среду LB, содержащую 5 г/л глюкозы и 20 г/л агара, так чтобы появлялось около 100 колоний на чашку, и культивировали при 30°С в течение ночи с наклоном чашки примерно в 45 градусов и затем отобрали 20 колоний, с которых не наблюдалось стекание слизи.
В качестве штамма, удовлетворяющего условию, чтобы не появлялось ревертанта даже после 5-кратного субкультивирования в среде LB, содержащей 5 г/л глюкозы и 20 г/л агара, и чтобы в нем наблюдался рост, эквивалентный росту родительского штамма в среде LB, среде LB, содержащей 5 г/л глюкозы, и среде М9 (Sambrook, J. Et al., Molecular Cloning, 2nd Edition, Cold Spring Harbor Press, USA, 1989) с добавлением 20 г/л L-глутаминовой кислоты и 2 г/л глюкозы и доведенной до значения рН 4,5 с помощью HCl, из штаммов отобранных, как указано выше, отобрали штамм SC17.
<4> Конструирование бактерии, продуцирующей глутаминовую кислоту на основе штамма SC17 Enterobacter agglomerans.
(1) Получение штамма, дефицитного по αKGDH, из штамма SC17 Enterobacter agglomerans.
Штамм, который был дефицитен по αKGDH и обладал усиленной системой биосинтеза L-глутаминовой кислоты, получали из штамма SC17 Enterobacter agglomerans.
(i) Клонирование гена αKGDH (далее называемого "sucAB") штамма AJ13355 Enterobacter agglomerans.
Ген sucAb Enterobacter agglomerans штамма AJ13355 клонировали посредством отбора фрагмента ДНК, комплементирующего неспособность дефицитного по гену субъединицы αKGDH-E1 (далее называемому "sucA") штамма Escherichia coli к ассимиляции уксусной кислоты, из хромосомной ДНК Enterobacter agglomerans штамма AJ13355.
Хромосомную ДНК Enterobacter agglomerans штамма AJ13355 выделяли способом, который обычно используют для экстракции хромосомной ДНК из Escherichia coii (Text for Bioengineering Experiments, Edited by the Society for Bioscience and Bioengineering, Japan, pp.97-98, Baifukan, 1992). Плазмида pTWV228 (резистентная к ампициллину), используемая в качестве вектора, представляла собой коммерческий продукт Takara Shuzo Co., Ltd.
Хромосомную ДНК штамма AJ13355, расщепленную ЕсоТ221, и pTWV228, расщепленную PstI, лигировали, используя лигазу Т4, и использовали для того, чтобы трансформировать Escherichia coli штамма JRG465, дефицитного по sucA (Herbert, J. et al., Mol. Gen. Genetics, 105, 182 (1969)). Из трансформированных штаммов, полученных как описано выше, отобрали штамм, растущий в минимальной среде с ацетатом, и из него экстрагировали плазмиду и обозначили ее pTWVEK101. В штамме JRG465 Escherichia coli, несущем pTWVEK101, наблюдалась компенсация ауксотрофии по янтарной кислоте или L-лизину и L-метионину, помимо характеристической особенности неспособности ассимилировать уксусную кислоту. Это свидетельствует о том, что pTWVEK101 содержала ген sucA Enterobacter agglomerans.
На фиг.1 показана рестрикционная карта фрагмента ДНК, полученного из Enterobacter agglomerans, в pTWVEK101. В нуклеотидной последовательности заштрихованной части на фиг.1 обнаружены нуклеотидные последовательности, рассматриваемые как две полноразмерные ОРС, и две нуклеотидные последовательности, рассматриваемые как неполные последовательности ОРС. В результате поиска их гомологов, обнаружено, что части, нуклеотидные последовательности которых были определены, содержали 3'-конец неполной последовательности гена железо-серосодержащего белка сукцинатдегидрогеназы (sdhB), полноразмерные sucA и ген субъединицы αKGDH-E2 (ген sucB) и 5'-конец неполной последовательности гена β-субъединицы сукцинил-КоА-синтетазы (ген sucC). Результаты сравнения аминокислотных последовательностей, выведенных на основе указанных нуклеотидных последовательностей, с последовательностями Escherichia coli (Eur. J. Biochem., 141, pp.351-359 (1984); Eur. J. Biochem., 141, pp.361-374 (1984); Biochemistry, 24, pp.6245-6252 (1985)) показаны на фиг.со 2 по 5. Таким образом, аминокислотные последовательности проявляют очень высокую гомологию друг с другом. Кроме того, было обнаружено, что кластер sdhB-sucA-sucB-sucC в хромосоме Enterobacter agglomerans образован так же, как и у Escherichia coli (Eur. J. Biochem., 141, pp.351-359 (1984); Eur. J. Biochem., 141, pp.361-374 (1984); Biochemistry, 24, pp.6245-6252 (1985)).
(ii) Отбор штамма, дефицитного по αKGDH, полученного из штамма SC17 Enterobaccer agglomerans.
Гомологичную рекомбинацию выполняли, используя ген sucAB Enterobacter agglomerans, полученный как описано выше, чтобы получить дефицитный по αKGDH штамм Enterobacter agglomerans.
После расщепления pTWVEK101 рестриктазой SphI, для того, чтобы вырезать фрагмент, содержащий sucA, фрагмент затупляли по концам с помощью фрагмента Кленова (Takara Shuzo Co., Ltd.) и лигировали с pBR322, расщепленной EcoRI и затупленной по концам фрагментом Кленова, используя ДНК-лигазу Т4 (Takara Shuzo Co., Ltd.). Полученную плазмиду расщепляли в сайте, узнаваемом рестриктазой BglII, расположенном примерно в центре sucA, используя фермент BglII, концы затупляли фрагментом Кленова и затем лигировали снова, используя ДНК-лигазу Т4. Предполагалось, что ген sucA стал нефункциональным, поскольку была введена мутация со сдвигом рамки в sucA плазмиды, заново сконструированной посредством указанного выше способа.
Плазмиду, сконструированную как описано выше, расщепляли рестриктазой ApaLI и подвергали электрофорезу в агарозном геле, чтобы извлечь фрагмент ДНК, содержащий sucA, в который была введена мутация сдвига рамки считывания, и ген устойчивости к тетрациклину, полученный из pBR322. Извлеченный фрагмент ДНК снова лигировали, используя ДНК-лигазу Т4, чтобы сконструировать плазмиду для нарушения гена αKGDH.
Плазмиду для нарушения гена αKGDH, полученную как описано выше, использовали для трансформации штамма SC17 Enterobacter agglomerans электропорацией (Miller, J.H., "A Short Course in Bacterial Genetics; Handbook", p.279, Cold Spring Harbor Laboratory Press, USA, 1992), и получили штамм, в котором sucA в хромосоме был заменен геном мутатного типа плазмиды посредством гомологичной рекомбинации, используя в качестве маркера устойчивость к тетрациклину. Полученный штамм назвали штаммом SC17sucA.
Для того чтобы подтвердить, что штамм SC17sucA был дефицитен по активности αKGDH, активность фермента измеряли способом Reed et al. (Reed, L.J. and Mukherjee, B.B., Methods in Enzymology, 13, pp.55-61, (1969)), используя клетки штамма, культивированные в среде LB в логарифмической фазе роста. В результате определили активность αKGDH для штамма SC17, равную 0,073 (ΔABS/мин/мг белка), в то время как активность αKGDH в штамме SC17sucA не выявлена, и таким образом, подтвердилось, что sucA был удален, как и планировалось.
(2) Усиление системы биосинтеза L-глутаминовой кислоты Enterobacter agglomerans штамма SC17sucA.
Затем в штамм SC17sucA вводили ген цитратсинтазы, ген фосфоенолпируваткарбоксилазы и ген глутаматдегидрогеназы, полученные из Escherichia coli.
(i) Получение плазмиды, имеющей в своем составе ген qltA, ген ррс и ген gdhA, полученные из Escherichia coli.
Объяснение процедур получения плазмиды, имеющей в своем составе ген qltA, ген ррс и ген gdhA, будет дано со ссылкой на фиг.6 и 7.
Плазмиду, несущую ген gdhA, полученную из Escherichia coli, pBRGDH (выложенная заявка на патент Японии No.7-203980), расщепляли HindIII и SphI, оба конца затупляли обработкой ДНК-полимеразой Т4, и затем фрагмент ДНК, имеющий в своем составе ген gdhA, очищали и выделяли. Отдельно плазмиду, несущую ген gltA и ген ррс, полученную из Escherichia coli, pMWCP (W097/08294), расщепляли XbaI, и затем оба конца затупляли, используя ДНК-полимеразу Т4. Полученную ДНК смешивали с указанным выше очищенным фрагментом ДНК, несущим ген gdhA, и лигировали, используя лигазу Т4, чтобы получить плазмиду pMWCPG, которая соответствовала pMWCP, дополнительно содержащей ген gdhA (фиг.6).
Одновременно плазмиду pVIC40 (выложенная заявка на патент Японии No.8-047397), имеющую в своем составе начало репликации плазмиды RSF1010 широкого круга хозяев, расщепляли NotI, обрабатывали ДНК-полимеразой Т4 DNA и расщепляли PstI. pBR322 расщепляли ЕсоТ141, обрабатывали ДНК-полимеразой Т4 и расщепляли PstI. Оба продукта смешивали и лигировали, используя лигазу Т4, чтобы получить плазмиду RSF-Tet, имеющую в своем составе начало репликации RSF1010 и ген устойчивости к тетрациклину (фиг.7).
Затем pMWCPG расщепляли EcoRI и PstI, и фрагмент ДНК, имеющий в своем составе ген qltA, ген ррс и ген gdhA, очищали и выделяли. RSF-Tet сходным образом расщепляли EcoRI и PstI, и фрагмент ДНК, имеющий в своем составе начало репликации RSF1010, очищали и выделяли. Оба продукта смешивали и лидировали, используя лигазу Т4, чтобы получить плазмиду RSFCPG, которая соответствовала RSF-Tet, содержащей ген gltA, ген ррс и ген gdhA (фиг.8). На основании комплементации ауксотрофии дефицитного по гену gltA, гену ррс или гену gdhA штамма, полученного из Escherichia coli, и измерения активности каждого фермента подтвердили, что полученная плазмида RSFCPG экспрессировала ген gltA, ген ррс и ген gdhA.
(ii) Получение плазмиды, имеющей в своем составе ген gltA, полученный из Brevibacterium iactofermentum.
Плазмиду, имеющую в своем составе ген gltA, полученный из Brevibacterium Iactofermentum, конструировали следующим образом. ПЦР выполняли, используя ДНК праймеров, которые получали на основании нуклеотидной последовательности гена gltA Corynebacterium glutamicum (Microbiology, 140, pp.1817-1828 (1994)), и хромосомную ДНК Brevibacterium lactofermentum ATCC13869 в качестве матрицы, чтобы получить фрагмент гена gltA, длиной около 3 т.п.н. Указанный фрагмент встраивали в плазмиду pHSG399 (приобретенную у Takara Shuzo Co., Ltd.), расщепленную Smal, чтобы получить плазмиду pHSGCB (фиг.9). Затем pHSGCB расщепляли HindIII, и вырезанный фрагмент гена gltA длиной примерно 3 т.п.н. встраивали в плазмиду pSTV29 (приобретенную у Takara Shuzo Co., Ltd.), расщепленную HindIII, чтобы получить плазмиду pSTVCB (фиг.9). Измеряя активность фермента в штамме AJ13355 Enterobacter agglomerans, подтвердили, что полученная плазмида pSTVCB экспрессировала ген gltA.
(iii) Введение RSFCPG и pSTVCB в штамм SC17sucA.
Штамм SC17sucA Enterobacter aggiomerans трансформировали RSFCPG посредством электропорации, чтобы получить трансформированный штамм SC17sucA/RSFCPG, проявляющий устойчивость к тетрациклину. Затем штамм SC17sucA/RSFCPG трансформировали pSTVCB путем электропорации, чтобы получить трансформированный штамм SC17sucA/RSFCPG + pSTVCB, проявляющий устойчивость к хлорамфениколу.
<5> Отбор штамма с повышенной устойчивостью к L-глутаминовой кислоте в кислых условиях среды.
Штамм с повышенной устойчивостью к L-глутаминовой кислоте при высокой концентрации в среде с низким значением рН (далее также называемый "штаммом, устойчивым к высокой концентрации Glu при низком рН") выделяли из штамма SC17sucA/RSFCPG+pSTVCB Enterobacter agglomerans.
Штамм SC17sucA/RSFCPG+pSTVCB культивировали в течение ночи при 30°С в среде LBG (10 г/л триптона, 5 г/л дрожжевого экстракта, 10 г/л NaCl, 5 г/л глюкозы), и клетки, промытые солевым раствором, соответствующим образом разбавляли и высевали в чашки со средой М9-Е (4 г/л глюкозы, 17 г/л Na2HPO4·12Н2О, 3 г/л КН2РО4, 0,5 г/л NaCl, 1 г/л NH4Cl, 10 мМ MgSOl, 10 мкм CaCl2, 50 мг/л L-лизина, 50 мг/л L-метионина, 50 мг/л DL-диаминопимелиновой кислоты, 25 мг/л тетрациклина, 25 мг/л хлорамфеникола, 30 г/л L-глутаминовой кислоты, рН которой доводили до 4,5 с помощью водного раствора аммиака). Получили колонию, появившуюся после культивирования при 32°С в течение 2 дней, в качестве штамма, устойчивого к высокой концентрации Glu при низком рН.
Измеряли уровень роста полученного штамма в жидкой среде М9-Е и тестировали способность продуцировать L-глутаминовую кислоту в большой пробирке объемом 50 мл, содержащей 5 мл среды для тестирования продукции L-глутаминовой кислоты (40 г/л глюкозы, 20 г/л сульфата аммония, 0,5 г/л гептагидрата сульфата магния, 2 г/л однозамещенного фосфата калия, 0,5 г/л хлорида натрия, 0,25 г/л дигидрата хлорида кальция, 0,02 г/л гептагидрата сульфата железа, 0,02 г/л тетрагидрата сульфата марганца, 0,72 мг/л дигидрата сульфата цинка, 0,64 мг/л пентагидрата сульфата меди, 0,72 мг/л гексагидрата хлорида кобальта, 0,4 мг/л борной кислоты, 1,2 мг/л дигидрата молибдата натрия, 2 г/л дрожжевого экстракта, 200 мг/л гидрохлорида L-лизина, 200 мг/л L-метионина, 200 мг/л DL-α, ε-диаминопимелиновой кислоты, 25 мг/л гидрохлорида тетрациклина и 25 мг/л хлорамфеникола). Штамм, который проявлял наилучший уровень роста и такую же способность продуцировать L-глутаминовую кислоту, как и его родительский штамм, штамм SC17/RSFCPG+pSTVCB, назвали AJ13601 Enterobacter agglomerans. Штамм AJ13601 был депонирован в National Institute of Bioscience and Human-Technology, Agency of Industrial Science and Technology, Ministry of International Trade and Industry (в настоящее время International Patent Organism Depositary, National Institute of Advanced Industrial Science and Technology; Central 6, Higashi 1-1-1, Tsukuba-shi, Ibaraki 305-8566, Japan) 18 августа 1999 г. и получил инвентарный номер FERM P-17516. Затем он был перенесен для международного депонирования по условиям Будапештского договора 6 июля 2000 г. и получил инвентраный номер FERM ВР-7207.
Пример 1: Ингибирование роста органической кислотой при кислом значении рН.
Хотя штамм AJ13601 Enterobacter agglomerans мог расти в широком диапазоне рН от нейтрального до кислого рН, ингибирование роста органической кислотой особенно заметно при кислом рН. Поэтому провели эксперимент, касающийся ингибирования роста органической кислотой при кислом рН.
Штамм AJ13601 Enterobacter agglomerans культивировали на агаризованной среде LBG (10 г/л триптона, 5 г/л дрожжевого экстракта, 10 г/л NaCl и 15 г/л агара), содержащей 25 мг/л гидрохлорида тетрациклина и 25 мг/л хлорамфеникола, при 30°С в течение 14 часов, и брали одну платиновую петлю клеток и инокулировали в 300 мл среды для посевной культуры, имеющей приведенный ниже состав, в ферментере с сосудом объемом 1 л, чтобы получить посевную культуру, при 34°С при рН 6,0.
[Состав среды для посевной культуры]
50 г/л сахарозы, 0,4 г/л гептагидрата сульфата магния, 4,0 г/л сульфата аммония, 2,0 г/л однозамещенного фосфата калия, 4,0 г/л дрожжевого экстракта, 0,01 г/л гептагидрата сульфата железа, 0,01 г/л пентагидрата сульфата марганца, 0,4 г/л гидрохлорида L-лизина, 0,4 г/л DL-метионина, 0,4 г/л DL-α,ε-диаминопимелиновой кислоты, 25 мг/л гидрохлорида тетрациклина и 25 мг/л хлорамфеникола.
рН в ходе культивирования контролировали добавлением газообразного аммиака. Выращивание посевной культуры останавливали при обнаружении истощения сахарида в среде для посевной культуры в качестве маркера, и бульон с посевной культурой инокулировали в 300 мл среды для основной культуры, находящейся в сосудах ферментеров объемом 1 л, в количестве, равном 20% объема среды для основной культуры, чтобы получить основную культуру. Состав среды для основной культуры показан ниже. При проведении эксперимента по ингибированию роста органической кислотой эксперимент выполняли, добавляя органическую кислоту в заранее определенной концентрации.
[Состав среды для основной культуры]
20 г/л глюкозы, 0,4 г/л гептагидрата сульфата магния, 5,0 г/л сульфата аммония, 6,0 г/л однозамещенного фосфата калия, 1,5 г/л хлорида натрия, 0,01 г/л гептагидрата сульфата железа, 0,01 г/л пентагидрата сульфата марганца, 0,8 г/л гидрохлорида L-лизина, 0,6 г/л DL-метионина, 0,6 г/л DL-α,ε-диаминопимелиновой кислоты, 25 мг/л гидрохлорида тетрациклина, 25 мг/л хлорамфеникола, 6/0 г/л дрожжевого экстракта и 0,75 г/л дигидрата хлорида кальция.
Температуру культуры доводили до 34°С, и рН регулировали, доводя до заранее определенного значения рН добавлением газообразного аммиака.
Эксперименты по ингибирующему действию на рост при кислом рН (рН 4.5) сначала выполняли с разными органическими кислотами. Эксперименты проводили, добавляя в среду для основной культуры муравьиную кислоту, уксусную кислоту, молочную кислоту, пропионовую кислоту, валериановую кислоту, янтарную кислоту, фумаровую кислоту, яблочную кислоту, щавелево-уксусную кислоту и лимонную кислоту в концентрации 0,5 г/л каждой. Часть результатов этих экспериментов показана на фиг.10.
Как показывают эти результаты, муравьиная кислота, уксусная кислота и пропионовая кислота показали явное ингибирование роста. С другой стороны, молочная кислота позволяет расти со скоростью, сравнимой со скоростью, полученной в контрольной среде без добавления органической кислоты (кроме того, янтарная кислота, фумаровая кислота и яблочная кислота также не показали ингибирования роста). Также имеется сообщение о том, что ингибирование роста микроорганизмов органической кислотой при кислом рН вызывается вследствие того, что органическая кислота, у которой отсутствует заряд в кислых условиях рН, проходит через цитоплазматическую мембрану и снова диссоциирует в нейтральных условиях рН в клетке, и это представление объясняется с использованием константы диссоциации рКа органической кислоты.
Однако результаты указанных выше экспериментов не могут быть объяснены на основе рКа, и предполагается, что ингибирование роста штамма AJ13601 Enterobacter agglomerans муравьиной кислотой, уксусной кислотой и пропионовой кислотой вызвано другим механизмом ингибирования, который нельзя объяснить с помощью концепции предыдущего сообщения.
Затем исследовали значения рН, ингибирующие рост, при использовании муравьиной кислоты (концентрация: 0,5 г/л). Результаты показаны на фиг.11. Стало ясно, что ингибирование роста муравьиной кислотой не наблюдалось в районе рН 5,5 или более, но наблюдалось при рН ниже данного значения. В частности, при рН 4,5 рост клеток едва наблюдается.
Затем также выясняли концентрацию муравьиной кислоты, вызывающую ингибирование роста в условиях рН 4,5. Результаты показаны на фиг.12. На основании результатов стало ясно, что ингибирование роста не наблюдалось в том случае, когда концентрация муравьиной кислоты в среде составляла 0,20 г/л или меньше, но ингибирование роста наблюдали при концентрации выше определенного уровня в пределах от 0,2 до 0,5 г/л.
На основании указанных выше экспериментальных результатов сделан вывод, что для того, чтобы обеспечить возможность культивирования штамма AJ13601 Enterobacter agglomerans при кислом рН с использованием источника углерода (сахарида), содержащего органическую кислоту, такого как меласса свеклы и меласса тростника, и источника азота, такого как кукурузный экстракт (CSL), раствор гидролизованного белка сои, раствор гидролизованного животного белка (мясной экстракт) и гидролизат казеина, необходимо снизить концентрацию в среде органической кислоты, проявляющей ингибирование, такой как муравьиная кислота и уксусная кислота, или для того, чтобы продуцировать L-глутаминовую кислоту при кислом рН необходим способ культивирования, включающий в себя культивирование клеток в пределах рН, в которых не наблюдается ингибирование роста, для того, чтобы поглощалась органическая кислота, проявляющая ингибирование, и затем культивирование клеток при значении рН, сдвинутом в сторону кислого рН, как указано в примере 2, приведенном ниже.
Пример 2: Снижение ингибирования роста органической кислотой посредством культивирования со сдвигом рН.
Рост клеток штамма AJ13601 Enterobacter agglomerans ингибируется низкомолекулярной органической кислотой, такой как муравьиная кислота, при кислом рН, как показано в примере 1. Поэтому, поскольку во многих случаях такое природное сырье, включая мелассу свеклы и мелассу тростника, содержит заметные количества таких органических кислот, штамм AJ13601 Enterobacter agglomerans нельзя культивировать при кислом рН, используя природное сырье. Однако, поскольку ингибирование роста органическими кислотами наблюдалось только при кислом рН, но не наблюдалось при нейтральном рН, были предприняты попытки продуцировать глутаминовую кислоту, проводя культивирование при нейтральном рН на ранней стадии культивирования, чтобы поглощать органические кислоты, вызывающие ингибирование роста, и затем сдвигая рН к кислому значению рН, чтобы продуцировать глутаминовую кислоту.
Штамм AJ13601 Enterobacter agglomerans культивировали на агаризованной среде LBG (10 г/л триптона, 5 г/л дрожжевого экстракта, 10 г/л NaCl и 15 г/л агара), содержащей 25 мг/л гидрохлорида тетрациклина и 25 мг/л хлорамфеникола при 30°С в течение 14 часов, и брали одну платиновую петлю клеток и инокулировали в 300 мл среды для посевной культуры, имеющей приведенный ниже состав, в сосуд ферментера объемом 1 л, чтобы вырастить посевную культуру при 34°С при рН 6,0. [Состав среды для посевной культуры]
50 г/л сахарозы, 0,4 г/л гептагидрата сульфата магния, 4,0 г/л сульфата аммония, 2,0 г/л однозамещенного фосфата калия, 4,0 г/л дрожжевого экстракта, 0,01 г/л гептагидрата сульфата железа, 0,01 г/л пентагидрата сульфата марганца, 0,4 г/л гидрохлорида L-лизина, 0,4 г/л DL-метионина, 0,4 г/л DL-α,ε-диаминопимелиновой кислоты, 25 мг/л гидрохлорида тетрациклина и 25 мг/л хлорамфеникола.
рН в ходе культивирования контролировали добавлением газообразного аммиака. Выращивание посевной культуры останавливали при обнаружении истощения сахарида в среде для посевной культуры в качестве маркера, и бульон с посевной культурой инокулировали в 300 мл среды для основной культуры, находящейся в сосуде ферментера объемом 1 л, в количестве, равном 20% объема среды для основной культуры, чтобы вырастить основную культуру. Состав среды для основной культуры показан ниже. [Состав среды для основной культуры]
20 г/л мелассы свеклы (или мелассы тростника), 0,4 г/л гептагидрата сульфата магния, 5,0 г/л сульфата аммония, 6,0 г/л однозамещенного фосфата калия, 1,5 г/л хлорида натрия, 0,01 г/л гептагидрата сульфата железа, 0,01 г/л пентагидрата сульфата марганца, 0,8 г/л гидрохлорида L-лизина, 0,6 г/л DL-метионина, 0,6 г/л DL-α,ε-диаминопимелиновой кислоты, 25 мг/л гидрохлорида тетрациклина, 25 мг/л хлорамфеникола, 6,0 г/л дрожжевого экстракта и 0,75 г/л дигидрата хлорида кальция.
Температуру культуры доводили до 34°С, и рН регулировали, доводя до заранее определенного значения рН добавлением газообразного аммиака. Культивирование начинали при рН 6,0 и проводили до тех пор, пока не поглощались сахарид и органическая кислота в среде для основной культуры. После истощения содержания сахарида, непрерывно (5 мл/час) добавляли 700 г/л водного раствора глюкозы. Наряду с непрерывным добавлением глюкозы, контролировали добавляемое количество газообразного аммиака, и снижение рН, связанное с продукцией глутаминовой кислоты, использовали для того, чтобы понизить рН до уровня 4,5 в течение примерно 2 часов, и затем культивирование продолжали при рН 4,5. Когда концентрация глутаминовой кислоты в культуральном бульоне достигала 45 г/л, в среду с основной культурой добавляли 1,0 г кристаллов глутаминовой кислоты в качестве зародышевых кристаллов, чтобы стимулировать осаждение кристаллов в культуральном бульоне.
В результате культивирования в течение 50 часов в сосуде ферментера осаждалось заметное количество кристаллов глутаминовой кислоты. Затем добавляли газообразный аммиак для того, чтобы увеличить рН до 6,0, чтобы растворить все кристаллы глутаминовой кислоты в сосуде ферментера, и затем измеряли количество полученной глутаминовой кислоты. Затем культивирование выполняли сходным образом, как описано выше, за исключением того, что поддерживали рН равным 4,5, начиная с ранней стадии культивирования, и измеряли количество полученной глутаминовой кислоты. В данном случае, поскольку количество клеток было снижено, культивирование прекращали через 28 часов. Результаты показаны в таблице 2.
Кроме того, измеряли временной ход роста клеток на основе оптической плотности (OD) при длине волны 620 нм. Результаты показаны на фиг.13.
В результате обнаружено, что в эксперименте, в котором культивирование осуществляли при кислом рН, начиная с ранней стадии культивирования, рост клеток был ингибирован и поэтому продукция L-глутаминовой кислоты была едва заметна, в то время как в эксперименте, в котором культивирование проводили с использованием сдвига рН, в культуральном бульоне выпадали кристаллы глутаминовой кислоты, и поэтому было показано, что в данном способе можно использовать природное сырье, содержащее органическую кислоту.
На основе указанных выше результатов получено подтверждение, что культивирование с использованием сдвига рН дает возможность для культивирования с выпадением кристаллов глутаминовой кислоты при кислом рН даже при использовании природного сырья, содержащего органические кислоты.








| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ L-ГЛУТАМИНОВОЙ КИСЛОТЫ | 2002 |
|
RU2282662C2 |
| ОРГАНИЧЕСКАЯ АЗОТСОДЕРЖАЩАЯ КОМПОЗИЦИЯ И УДОБРЕНИЕ, ВКЛЮЧАЮЩЕЕ ЕЕ | 2002 |
|
RU2291139C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-ГЛУТАМИНОВОЙ КИСЛОТЫ | 1999 |
|
RU2188236C2 |
| ШТАММ ENTEROBACTER AGGLOMERANS, СПОСОБ ПОЛУЧЕНИЯ L-ГЛУТАМИНОВОЙ КИСЛОТЫ (ВАРИАНТЫ) И СПОСОБ ПОЛУЧЕНИЯ МИКРООРГАНИЗМА ДЛЯ ВЫРАБОТКИ L-ГЛУТАМИНОВОЙ КИСЛОТЫ | 2000 |
|
RU2261272C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-ГЛУТАМИНОВОЙ КИСЛОТЫ | 1999 |
|
RU2194076C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ | 2003 |
|
RU2307165C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ СЕМЕЙСТВА ГЛУТАМАТА ИЛИ L-ВАЛИНА С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ Escherichia | 2009 |
|
RU2418064C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ Escherichia, В КОТОРОЙ ИНАКТИВИРОВАН ГЕН yafA | 2005 |
|
RU2315808C2 |
| СПОСОБ ПРОИЗВОДСТВА L-АМИНОКИСЛОТ | 2007 |
|
RU2422530C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-ТРЕОНИНА С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ Escherichia, В КОТОРОЙ ИНАКТИВИРОВАН ГЕН b2383 | 2006 |
|
RU2333952C2 |



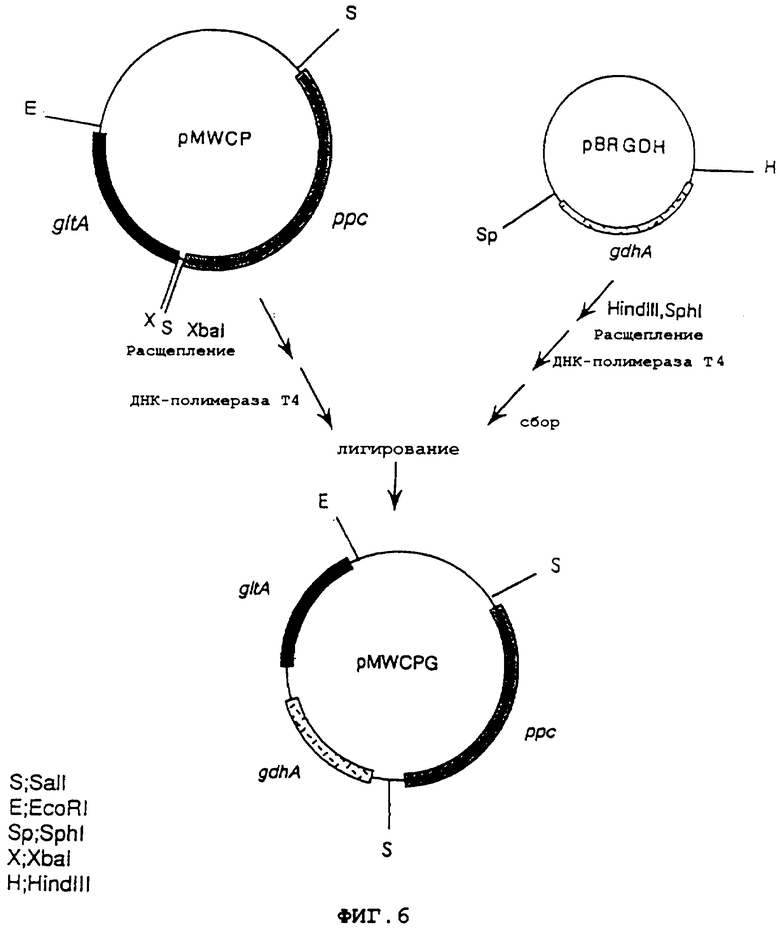
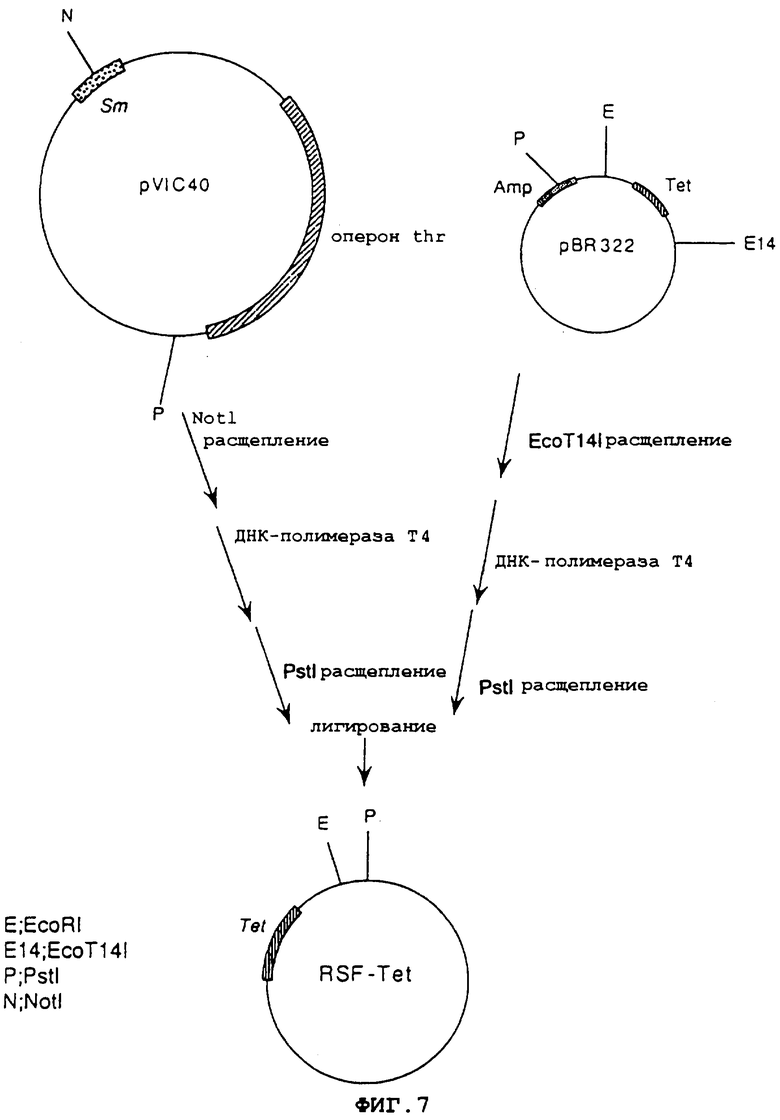
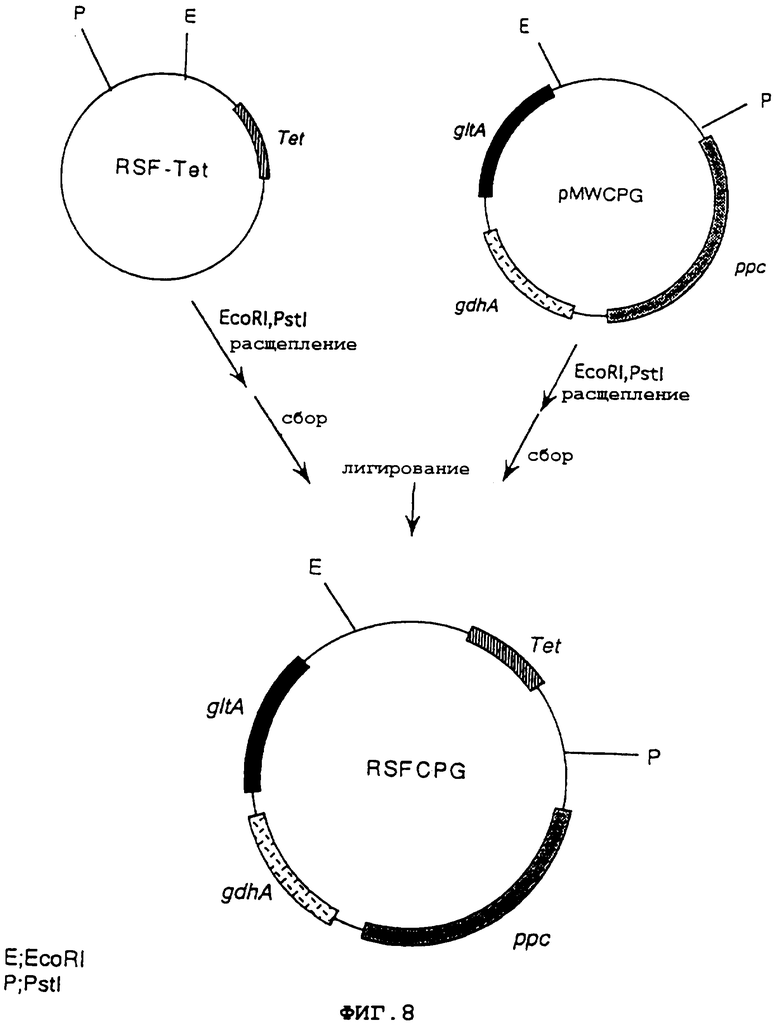

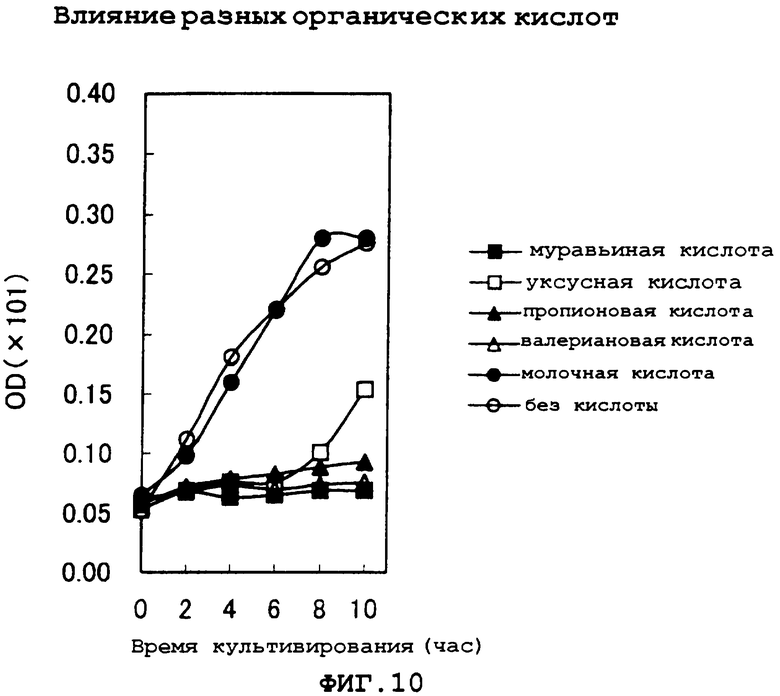
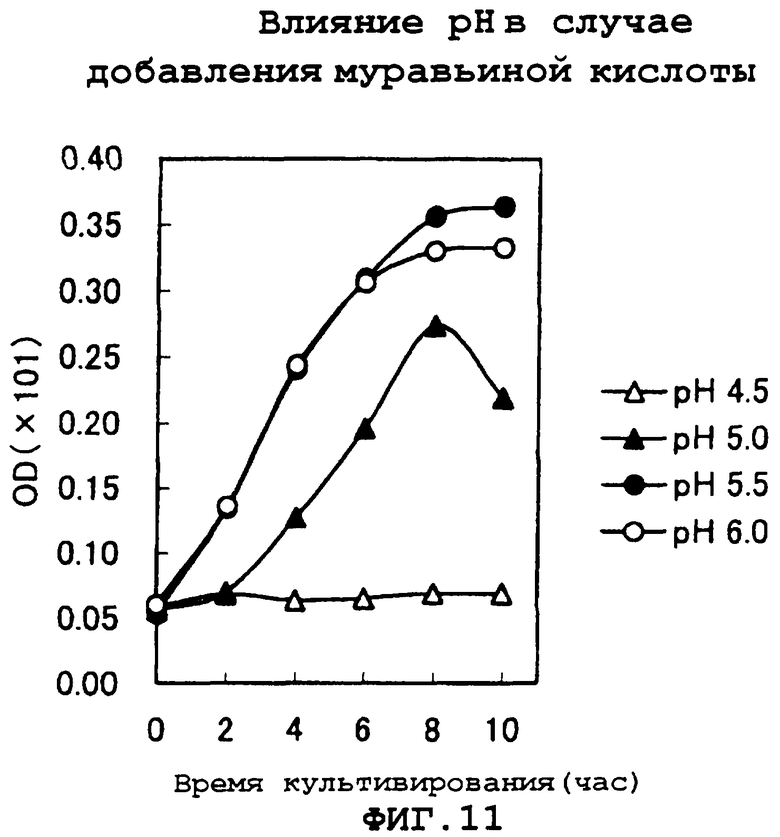
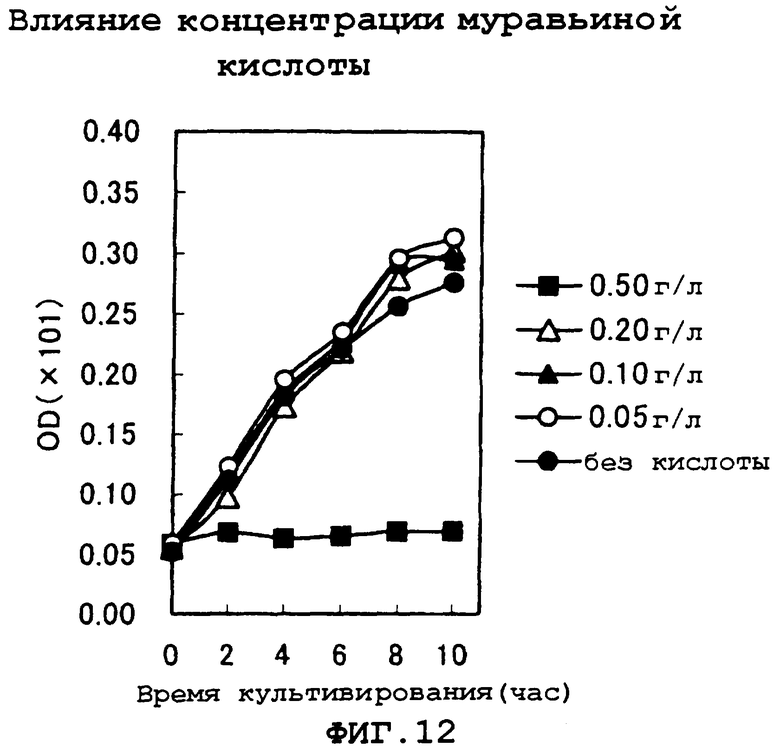
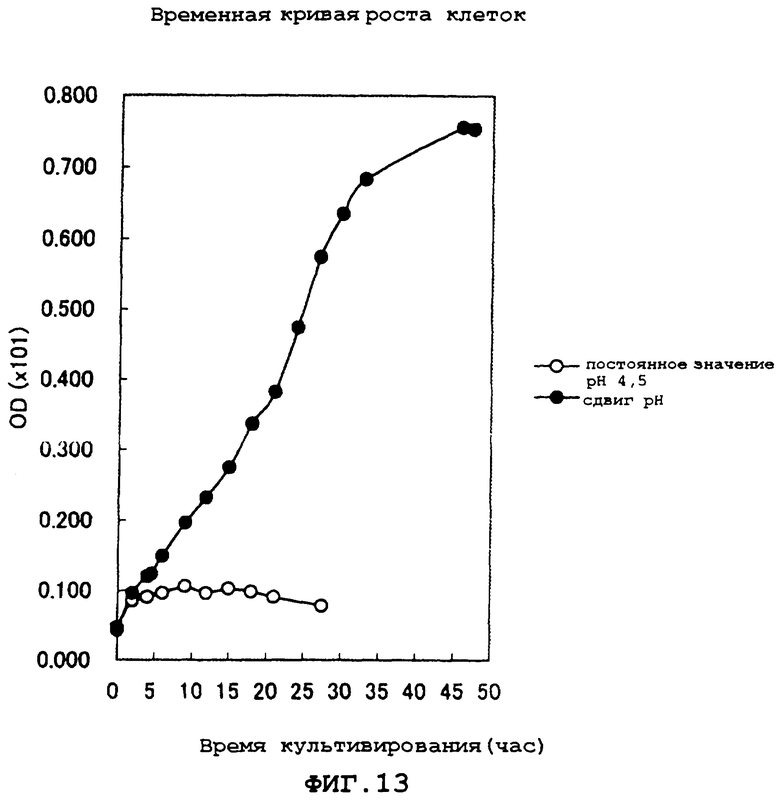
Изобретение относится к биотехнологии. L-глутаминовую кислоту получают ферментацией, включающей в себя культивирование микроорганизма, обладающего способностью продуцировать L-глутаминовую кислоту, в среде, имеющей значение рН 5,0 или меньше, при этом общее содержание органической кислоты, которая ингибирует рост микроорганизма при данном рН, равно такому количеству, при котором рост микроорганизма не ингибируется. L-глутаминовую кислоту получают также ферментацией, которая включает в себя культивирование микроорганизма, обладающего способностью продуцировать L-глутаминовую кислоту, при первом значении рН, при котором органическая кислота в среде не ингибирует рост микроорганизма, и затем культивирование микроорганизма при втором рН, которое подходит для продукции L-глутаминовой кислоты микроорганизмом и значение которого ниже, чем первое значение рН. При этом микроорганизм может метаболизировать источник углерода в жидкой среде, содержащей L-глутаминовую кислоту при концентрации насыщения и источник углерода, при рН 5 или меньше, и способен накапливать L-глутаминовую кислоту в количестве, превышающем концентрацию насыщения L-глутаминовой кислоты. Заявленное изобретение позволяет получать L-глутаминовую кислоту с высокой степенью эффективности. 2 н. и 8 з.п. ф-лы, 13 ил., 2 табл.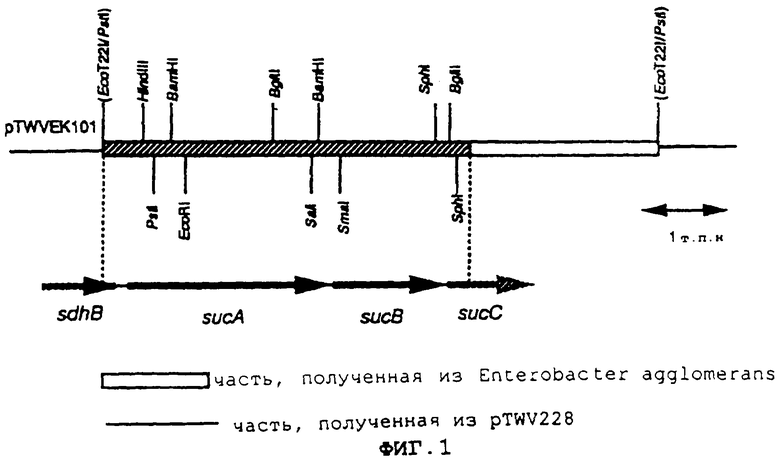
| US 3220929, 30.11.1965 | |||
| ШТАММ БАКТЕРИЙ CORYNEBACTERIUM GLUTAMICUM В-7198-ПРОДУЦЕНТ L-ГЛЮТАМИНОВОЙ КИСЛОТЫ | 1996 |
|
RU2107723C1 |
| Способ крашения тканей | 1922 |
|
SU62A1 |

