Изобретение относится к, по существу, чистому димеру apo-липопротеина AI-Milano/Apo AI-M/Apo-M/ и фармацевтическим составам, содержащим этот димер. Оно также относится к способу его получения путем рекомбинантной техники, а также путем выделения из плазмы. Этот продукт может быть использован для лечения атеросклероза и сердечно-сосудистых заболеваний; назначение Apo AI-M может быть способом замедления действия другого лекарства.
Предшествующий уровень техники.
Явная связь между повышенными уровнями холестерина в сыворотке крови и развитием болезней венечной артерии сердца /БВАС/ неоднократно подтверждалось данными, основанными на эпидемиологических исследованиях и исследованиях продольного разреза сердца. Однако определение сложных механизмов транспорта холестерина в плазме позволило признать избирательную функцию липопротеинов в кровеносной системе и установлении риска БВАС.
В действительности существует 4 основных липопротеина кровеносной системы: шарики жиров, которые появляются в крови во время переваривания жиросодержащей пищи /ШЖ/, липопротеины с очень низкой плотностью /ЛОНП/, липопротеины с низкой плотностью /ЛНП/ и липопротеины с высокой плотностью /ЛВП/. В то время как ШЖ представляет собой коротко живущий продут кишечного всасывания жиров, ЛОНП и в особенности ЛНП ответственны за транспорт холестерина в ткани, включающие, например, стенки артерий. В отличие от них ЛВП прямо вовлечены в удаление холестерина из периферических тканей путем перенесения их обратно либо в печень, либо к другим липопротеинам при помощи механизма, известного как "обратный транспорт холестерина" /ОТХ/.
"Защитная" роль ЛВП подтверждена в ряде исследований /например, Meller et al, Lancet, 1977: 965 - 968 и Whayne et al, Atherosclerosis, 1981; 39:411 - 419/. В этих исследованиях повышенные уровни ЛНП и в меньшей степени ЛОНП представляются явно связанными с увеличенным сердечно-сосудистым риском, тогда как высокие уровни ЛВП, по-видимому, придают сердечно-сосудистую защиту. Защитная роль ЛВП в дальнейшем была сильно поддержана исследованиями in vivo, показывающими, что внутривенные вливания ЛВП кроликам могут препятствовать развитию артериальных повреждений, вызванных холестерином /Badimon et al, Lab. Invest., 60, 455 - 61, 1989/, и/или способствовать их возвращению в прежнее состояние /Badimon et al, J. Clin Invest., 85, 1334 - 41, 1998/.
В настоящее время интерес в исследованиях защитного механизма(ов) ЛВП сосредоточен на apo-липопротеине AI /Apo AI/, основном компоненте ЛВП. Высокие уровни Apo AI в плазме связаны с уменьшенным риском БВАС и наличием повреждений венечной артерии /Maciejko et al, N Engl J. Med., 1983; 309: 385 - 389, Sedlis et al, Circulation, 1986; 73: 978 - 984/.
Плазменный Apo AI является отдельной полипептидной цепью из 243 аминокислот, первичная последовательность которых известна /Brewer et al, Biochem Biophys Res Commun. 1978; 80:623 - 630/. Apo AI синтезируется в клетке в виде предшественника из 267 аминокислот. Этот пред-apo-липопротеин подвергается N-концевому расщеплению - сначала внутриклеточному, где теряются 18 аминокислот, а затем дальнейшему отщеплению 6 аминокислот в плазме или лимфе при участии активности специфических протеаз.
Полагают, что основным требованием к структуре молекулы Apo AI является наличие повторяющихся единиц из 11 или 22 аминокислот, которые предпочтительно нужны для создания амфипатической спиральной конформации /Segurest et al, FEBS Lett 1974, 38: 247-253/. Эта структура позволяет осуществляться главным биологическим активностям Apo AI, а именно связыванию липидов и активации лецитин-холестерин-ацилтрансферазы /ЛХАТ/.
Другим недавно описанным свойством Apo AI является его антивирусная активность. О ней сообщалось по данным исследований in vitro; она осуществляется и против штаммов герпесного вируса /Srinivas R.V. et al, Virology. 1756, 48-57, 1990/, и также против вируса иммунодефецита человека (ВИЧ) /Owe et al, J. Chin Invest, 86, 1142-50, 1990/. По-видимому, эта активность осуществляется путем взаимодействия между амфипатическими спиральными участками Apo AI и гликопротеинами оболочки вирусов.
Исследования in vitro показывают, что комплексы Apo AI и лецитина могут способствовать удалению свободного холестерина из культивируемых клеток артериальных гладких мышц /Stein et al, Ciochem. Biophys. Acta. 1975; 380: 106-118/. При помощи этого механизма ЛВП также может уменьшить быстрое размножение этих клеток /Yoshida et al, Exp. Mol. Pathol. 1984; 41:258-266/.
Также было показано, что внутривенное вливание Apo AI или ЛВП экспериментальным животным вызывает значительные биохимические изменения, а также уменьшение протяженности и силы атеросклеротических повреждений. После первого сообщения, сделанного Maciejko и Mao /Arteriosclerosis, 1982: 2:407a/, Badimon и др. /смотри две работы, процитированные выше/ обнаружили, что эти вещества могут значительно уменьшить протяженность атеросклеротических повреждений /-45%/, а внутривенное вливание ЛВП /d = 1,063-1,325 г/мл/ кроликам, которых кормили холестерином, значительно снижает у них содержание эфира холестерина /-58,5%/. Эти исследования также нашли, что внутривенное вливание ЛВП проводит к возвращению в исходное состояние /почти на 50%/ установившихся повреждений. Также была возможность показать /Esper et al, Arteriosclerosis 1987; 7:523a/, что внутривенные вливания ЛВП могут заметно изменить состав липопротеинов в плазме кроликов Watanabe с наследственным повышенным содержанием холестерина в крови, у которых развиваются ранние артериальные повреждения. У этих кроликов внутривенные вливания ЛВП могут более чем удвоить соотношение между защищающим ЛВП и ЛНП, который способствует атеросклерозу.
Возможность с помощью ЛВП предотвратить артериальные заболевания на моделях животных далее была поддержана наблюдением того, что Apo AI может осуществлять in vitro активность, расщепляющую фибрин /Saku et al, Thromb. Res. , 1985; 39:1-8/. Роннебергер /Xth Int. Cohgr. Pharmacol., Sidney, 1987, p. 990/ продемонстрировал, что экстрагированный Apo AI может увеличивать расщепление фибрина у собак-ищеек и у Cynomologous обезьян. Похожая активность была отмечена in vitro в плазме человека. Этот исследователь смог подтвердить уменьшение липидных отложений и образования артериальных бляшек у животных, обработанных Apo AI.
Apo-липопротеин AI-Milano /Apo AI-M/ является первым описанным молекулярным вариантом человеческого Apo AI /Franceshini et al., J.Clin. Invest., 1980; 66: 892-900/. Он отличается замещением A 173 на Cy /Weisgraber et al., J. Miol Chem. , 1983; 258: 2508-2513/. Мутантный apo-потеин передается как аутосомный доминантный признак, и было идентифицировано 8 поколений носителей /Gualandri et al., Am. J. Hum. Genet, 1984; 37:1083-1097/.
Состояние индивидуального носителя Apo AI-M отличается замечательным снижением уровня холестерина, связанным с ЛВП. Несмотря на это организмы, имеющие этот признак, с очевидностью не показали какого-либо повышенного риска артериальных заболеваний; разумеется, из проверки их генеалогического древа явствовало, что эти организмы могут считаться "защищенными" от атеросклероза.
Механизм возможного защитного эффекта Apo AI-M у носителей, по-видимому, связан с изменением структуры мутантного apo-липопротеина из-за потери одной альфа-спирали и увеличенного обнажения гидрофобных остатков /Francheschini et al, J. Biol. Chem. 1985; 260: 1632-1635/. Потеря сжатой структуры многочисленных альфа-спиралей приводит к увеличению гибкости молекулы, которая легче соединяется с липидами по сравнению с нормальным AI. Более того, комплексы apo-липопротеина с липидами более чувствительны в отношении денатурации, таким образом подтверждая, что снабжение липидами также улучшено в случае мутантов.
Другой очень специфической чертой Apo AI-M является его способность образовывать собственные димеры и комплексы с Apo AII, в обоих случаях из-за присутствия Cys-остатка. Из исследований фракций крови, содержащих смесь apo-липопротеинов, получены указания на то, что присутствие димеров и комплексов в кровеносной системе может быть ответственным за увеличение времени полураспада, удаляющего apo-липопротеины, у носителей, недавно описанных в клинических исследованиях /Gregg et al., NATO ARW on Human Apolipoprotein Mutants: From Gene Structure to Phenotypic Expression, Limone, 1988/.
Димеры Apo AI-M /Apo AI-М/Apo AI-M/ действуют как подавляющий фактор взаимных превращений частиц ЛВП in vitro /Franceschini et al., J.Biol Chem., 1990; 265: 12224-12231/.
Ранние исследования смесей, содержащих димер, были основаны на Apo-AI-М, выделенном из натуральной крови людей с признаком Apo AI-M, и, таким образом, белок мог быть получен только в небольших количествах.
Трудность получения Apo AI и особенно Apo AI-M из фракционированной плазмы весьма значительна /Franceschini etal., J. Biol. Chem., 1985; 260: 16321-16325/. Выделение и получение не может быть проведено в большом масштабе, поскольку только небольшое количество неочищенного материала имеется в распоряжении исследователя. Более того, существует определенный риск, связанный с продуктами фракционированной плазмы, такой как загрязнение инфекционными агентами. Все это настолько существенно, что лучше этого избегать.
Были сделаны попытки получить человеческий Apo AI при помощи техники рекомбинантной ДНК. В опубликованном европейском патенте N 0267703 описано получение Apo AI из E.Coli. Процесс представляет химерные полипептиды, в которых группы Apo AI слиты с N-концевым аминокислотным остатком бета-галактозидазы или с одной или более областей, связывающих ИгG, белка A или с предпоследовательностью человеческого Apo AI.
Экспрессия Apo AI и Apo AI-М в штаммах дрожжей и использование полученных соединений в лечении атеросклероза и сердечно-сосудистых заболеваний раскрыто в W090/12879. Гены, кодирующие Apo AI и Apo AI-М, расположены на последовательности ДНК, которая кодирует выделение, узнаваемое в клетках дрожжей, и сигналы процессинга, присоединенные выше генов, кодирующих зрелые белки. В данном исследовании была использована измененная лидирующая последовательность MF-альфа-1, в которой последними остатками являются: His Gly Sor Leu Asp Lys Arg.
Настоящее изобретение.
В настоящее время мы неожиданно обнаружили, что очищенный димер Apo AI-М/Apo AI-М имеет продленный период полураспада в плазме по сравнению с мономером Apo AI-М; далее, что он имеет заметно улучшенное свойство способствовать расщеплению фибрина в сравнении с нормальным Apo AI, например его способность к прямой активации плазминогена /чего нормальный Apo AI-M делать не может/ - наблюдение, которое может иметь биологическую важность - и также возможность действовать в качестве про-лекарства для Apo AI-M. Димер также образует вновь составленные частицы ЛВП /липопротеин с высокой плотностью/ одинакового размера, которые не обнаружены среди частиц рекомбинантного ЛВП, содержащих Apo AI-M или Apo AI.
Настоящее изобретение относится к, по существу, чистому димеру apo-липопротеина Apo-Milano, впоследствии называемому Apo AI-M/Apo AI-M, с чистотой по меньшей мере 90%, предпочтительно по меньшей мере 98%, который сначала был выделен из плазмы и охарактеризован, а затем был также получен рекомбинантными методами. Настоящее изобретение также относится к фармацевтическим составам, включающим Apo AI-M/Apo AI-M, вместе /что необязательно/ со стабилизирующим агентом, например стабилизирующее липидное соединение, такое как фосфолипид и/или носитель.
Фармацевтические составы также могут содержать агент, снижающий содержание липидов и/или другой лекарство, уже известное в лечении атеросклероза и сердечно-сосудистых заболеваний, такое как гепарин, фракции гепарина и фрагменты гепарина или агенты, снижающие содержание липидов.
Apo-липопротеин Apo AI-M можно получить:
а) образованием apo-липопротеина AI-Milano путем рекомбинантной технологии в виде внутриклеточного слитого белка E.coli, отщеплением apo-липопротеина AI-Milano с помощью муравьиной кислоты и после этого превращением любого присутствующего мономера в димер, или
б) образованием apo-липопротеина AI-Milano путем рекомбинантной технологии, по которой apo-липопротеин AI-Milano, мономер или димер выделяется в среду бактериальной культуры в экспрессионной системе E.Coli, и любой присутствующий мономер после этого превращается в димер, в обоих случаях последним этапом является очистка димера до в основном чистой формы.
В соответствии с а) Apo AI-M образуется в виде слитого белка внутри клеток бактерий. Партнером по слиянию является измененная область белка А, которая связывается с ИгG, а сайт расщепления муравьиной кислотой по замыслу находится между партнером по слиянию и Apo AI-M. После лизисов бактерий слитый белок очищается на иммобилизованном ИгG и расщепляется муравьиной кислотой. Присутствие Apo AI-M и его димера было показано методом Вестерн-блоттинга на СДС-гель-электрофорезе.
В примере 3 показано, что искусственный Apo AI-M может быть получен в рекомбинантной E. coli и что образуются димеры. Однако использование муравьиной кислоты дает продукт, сокращенный на две аминокислоты с его N-конца. Предполагается, что такое сокращение не изменяет активности молекулы Apo AI-M.
Система в соответствии с б) показана в примере 4, в котором образуется полная правильная молекула.
Заявляемый димер Apo AI-M также можно получить:
с) сбором плазмы у носителей apo-липопротеина AI-Milano, выделением apo-липопротеина ЛВП, отделением димера при помощи хроматографии, например хроматографией на Сефакриле в несколько этапов, или
д) сбором плазмы у носителей apo-липопротеина AI-Milano, очисткой мономера и после этого превращением его в димер, а затем очисткой димера до по существу чистой формы.
Важным является то, что разделение в соответствии с с) осуществляется в несколько этапов, предпочтительно на длинной колонке, такой как 300 см.
Если присутствует мономер, то его всегда следует превращать в форму димера, как показано в нижеприведенных примерах.
Настоящее изобретение включает способ лечения атеросклероза и сердечно-сосудистых заболеваний, а также использование димера в приготовлении лекарств. Димер также может действовать как про-лекарство мономера при лечении атеросклероза и сердечно-сосудистых заболеваний.
Эти лекарства могут быть использованы для лечения атеросклероза и сердечно-сосудистых заболеваний, а также для предотвращения и лечения основных заболеваний сердца и кровеносной системы, таких как инфаркт миокарда, неустойчивая ангина, внезапные закупорки периферических сосудов, и во время восстановительного периода после пластических операций на венечных сосудах.
Когда осуществляется лечение хронических артериальных заболеваний, то лечатся и венечные и также периферические артерии, которые характеризуются закупоривающими бляшками. Димеры сами по себе будут использоваться для внутривенных вливаний с целью вызвать удаление жиров из бляшек или в соединении /что необязательно/ с уже назначенным лечением атеросклероза и сердечно-сосудистых заболеваний, таким как использование гепарина, фракций гепарина и фрагментов гепарина и/или лекарств, уменьшающих уровни в кровеносной системе липопротеинов, вызывающих атеросклероз.
Лекарства, содержащие димер, могут быть использованы для предотвращения и лечения тромбозов в различных клинических ситуациях, а также для стимуляции расщепления фибрина.
В димере Apo AI-M с высоким содержанием присутствуют амфипатические структуры, и предполагается, что димер обладает противовирусной активностью.
Сейчас на первое время путем использования настоящего изобретения возможно получать димер в, по существу, чистой форме - более 90% и даже выше чем 98%, а также продемонстрировать, что этот продукт обладает удивительно улучшенным действием на биологические системы по сравнению с Apo AI, которое показано в примерах 7-10, приведенных ниже.
Фиг. 1 и 2 иллюстрируют зависимость выхода в % от концентрации белка /пример 2б/.
Фиг. 3 иллюстрирует кинетику образования Apo AI-M/Apo AI-M в течение 24 часов /примеры 1б, 2/.
Фиг. 4 показывает синтетический линкер /пример 3а/.
Фиг. 5 показывает плазмиду /пример 3а/.
Фиг. 6 иллюстрирует Вестерн-анализ после расщепления слизистого белка /пример 3д/.
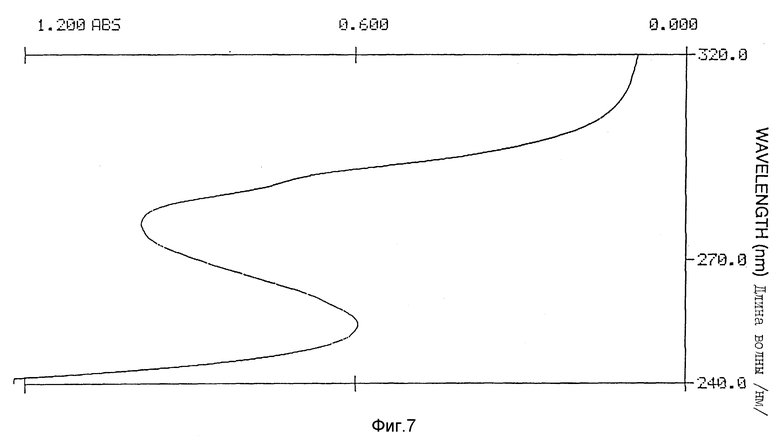
Фиг. 7 иллюстрирует УФ-спектр Apo AI-M/Apo AI-M /пример 5/.
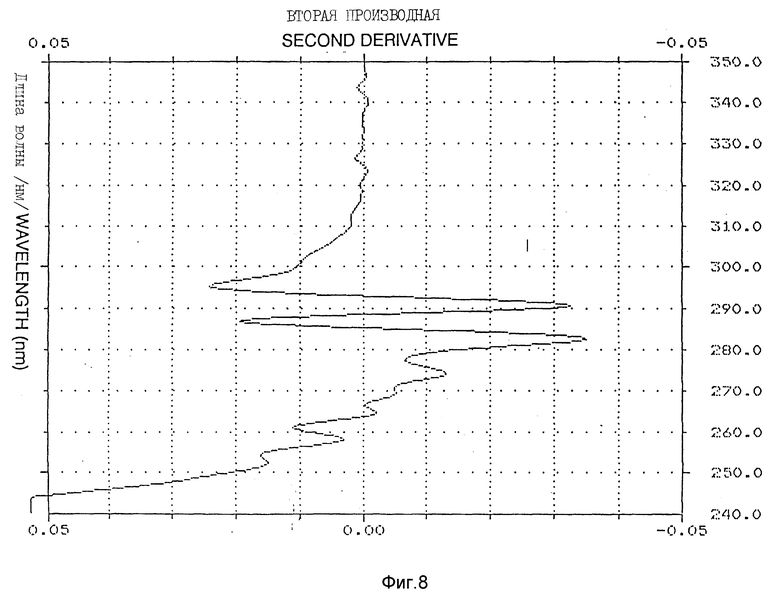
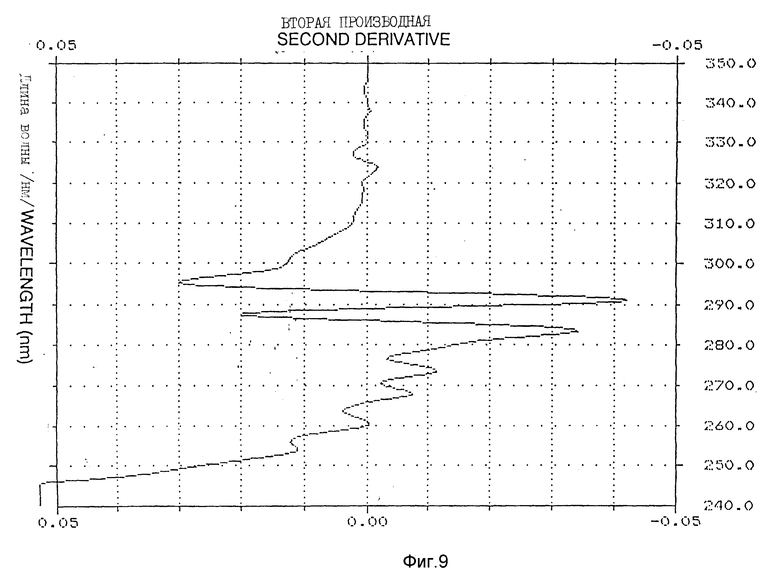
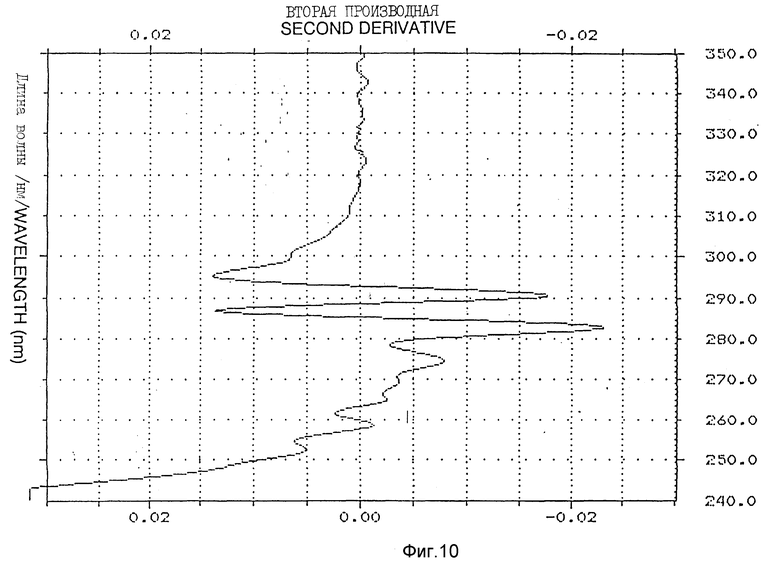
Фиг. 8-10 иллюстрируют вторую производную УФ-спектра /пример 5/.
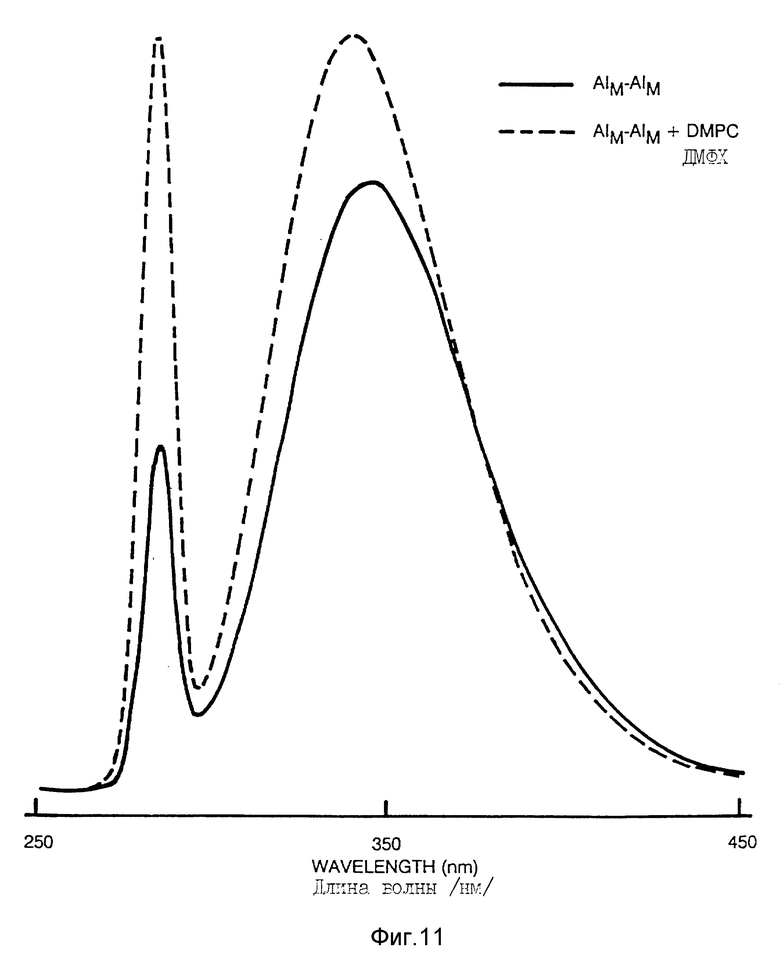
Фиг. 11 иллюстрирует флуоресцентную спектроскопию /пример 5/.
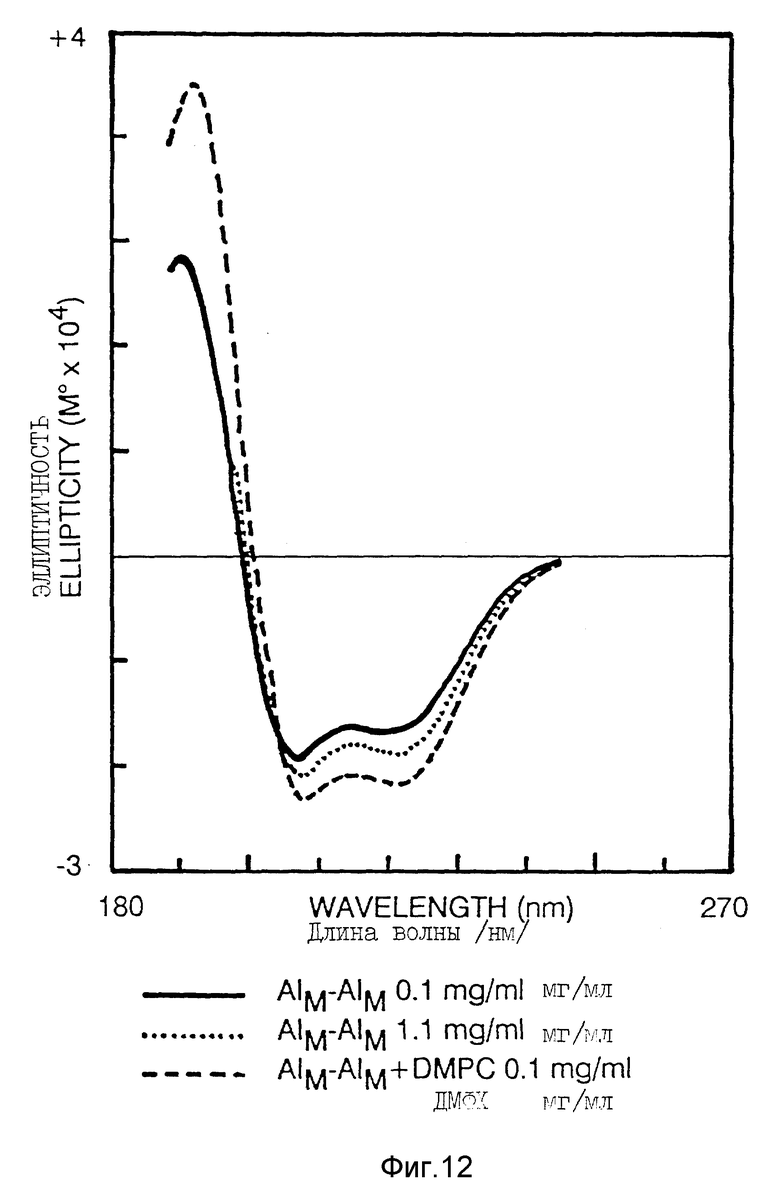
Фиг. 12 иллюстрирует КД-спектроскопию /пример 5/.
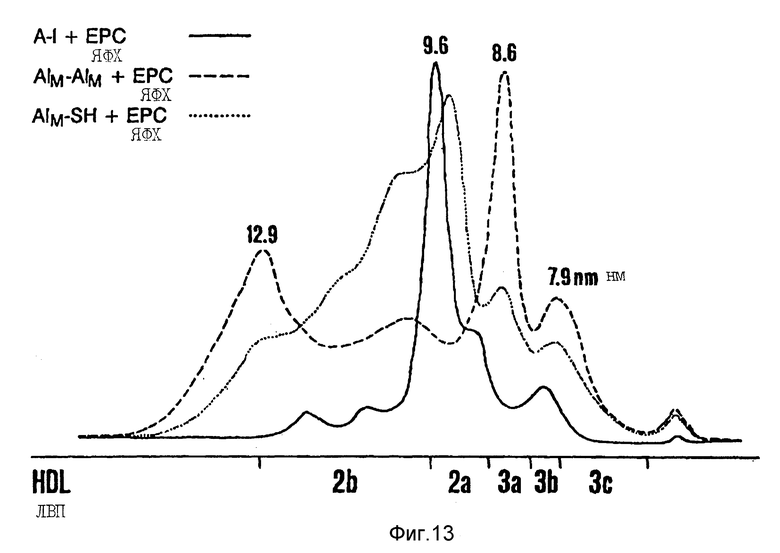
Фиг. 13 иллюстрирует рЛВП, содержащий Apo AI, Apo AI-M и Apo AI-М/Apo AI-M /пример 6/.
Экспериментальная часть.
Пример 1. Выделение димера из плазмы.
а). Приготовление apo-липопротеинов.
Образцы крови, взятые натощак у различных носителей apo-липопротеина Apo AI-М, собирали в раствор Na2-ЭДТА /1 мг/мл/, и готовили плазму путем центрифугирования с низкой скоростью при 4oC. Плазменные липопротеины с высокой плотностью /ЛВП, d= 1,063-1,21 г/мл/ выделяли последовательным ультрацентрифугированием /Хавел Р.Ж., Эдер Х.А., Брэгдон Ж.Х. Распределение и химический состав липопротеинов сыворотки человека, разделенных ультрацентрифугированием. J. Clin. Invest., 1955; 34: 1345-1354/ на ультрацентрифуге Beckman L5-50B, снабженной ротором 50,2 Ti. После ультрацентрифугирования в течение 48 часов при 40000 об/мин при 4oC верхнюю фракцию, содержащую ЛВП, разводили в соотношении 1:1 раствором 0,15 М NaCl, 0,01% Na2-ЭДТА, КВ /pH 7,4, d = 1,21 г/мл/ и повторно центрифугировали в течение 48 часов при 40000 об/мин и 4oC. ЛВП были диализованы с истощением против 5 мМ NH4HCO3, 0,01% Na2- ЭДТА, pH 7,4, лиофилизированы и обезжирены при помощи смеси диэтиловый эфир/этанол /3: 1, объем/объем/. Концентрация белка была измерена либо аминокислотным анализом, либо методом Лоури и др. /Лоури О.Н., Розенбрух Н.Ж., Фарр А.Л., Рандалл Р.Ж. Измерение белка с помощью фенольного реагента Фолина. J. Biol. Chem. 1951; 193: 265-275/.
б). Выделение Apo AI-M/Apo AI-M.
Для выделения Apo AI-M/Apo AI-M apo-липопротеины ЛВП из примера 1a растворяли в 0,1 М Трис-HCl, 0,04% Na2-ЭДТА, 0,01% NaN3, pH 7,4, содержащем 6 М Гуанидин-HCl /Гдн-HCl/. Apo-липопротеины наносили на колонку Сефакрил S-300 HR /2,6x300 см/, уравновешенную раствором 0,1 М Трис-HCl, 0,04% Na2-ЭДТА, 0,01% NaN3, pH 7,4, содержащим 4 M Гдн-HCl, apo-липопротеины элюировали тем же буфером при объемной скорости потока 1,5 мл/мин; фракции по 10 мл собирали. Объединенные фракции, содержащие Apo AI-M/Apo AI-M, концентрировали под вакуумом и повторно наносили на ту же колонку. Фракции, содержащие чистый Apo AI-M/Apo AI-M, собирали, диализовали против 5 мМ NH4HCO3, 0,01% Na2-ЭДТА, pH 7,4, лиофилизировали и хранили при -20oC в растворе 0,1 М Трис-HCl, 0,04% Na2-ЭДТА, 0,01% NaN3, pH 7,4, содержащем 3 М Гдн-HCl. Препараты Apo AI-M/Apo AI-M имели чистоту 90%, как было показано высокоэффективной эксклюзионной хроматографией /ВЭЭХ/ и СДС-полиакриламидным гель-электрофорезом /СДС-ПАГЭ/ при невосстанавливающих условиях. С использованием этого метода из плазмы носителей был выделен чистый димер Apo AI-V.
Пример 2. Очистка мономера из плазмы и после этого превращение его в димер.
а). Очистка Apo AI-M.
Для очистки мономера Apo AI-M apo-липопротеины ЛВП из примера 1а растворяли в 0,1 М Трис-HCl, 0,04% Na2-ЭДТА, 0.01% NaN3, pH 7,4, содержащем 6 М Гдн-HCl и 1% 2-меркаптоэтанола. После 4 часов инкубации при 37oC восстановленный apo-ЛВП наносили на колонку Сефакрил S-200 /2,6х150 см/, уравновешенную 0,1 М Трис-HCl, 0,04% Na2-ЭДТА, 0,01% NaN3, pH 7,4, содержащим 4 М Гдн-HCl и 0,1% 2-меркаптоэтанола. Apo-липопротеины элюировали тем же буфером при низкой объемной скорости потока 1,0 мл/мин; фракции по 5 мл собирали. Объединенные фракции, соответствующие apo AI+apo AI-M диализовали против 5 мМ NH4HCO3, 0,01% Na2-ЭДТА, pH 7,4 и лиофилизировали. Apo-липопротеины растворяли в 0,1 М Трис-HCl, 0,04% Na2-ЭДТА, 0,01% NaN3, pH 7,4, содержащем 6 М Гдн-HCl и 1% 2-меркаптоэтанола. После 4 часов инкубации при 25oC 2-меркаптоэтанол удаляли при помощи хроматографии на Сефадексе G25, а apo-липопротеины немедленно наносили на колонку тиопропил-сефароза /1х10 см/, уравновешенную 0,1 М Трис-HCl, 0,04% Na2-ЭДТА, 0,01 NaN3, pH 7,4, содержащим 4 М Гдн-HCl. После повторного пропускания через колонку в течение ночи с низкой объемной скоростью потока /0,15 мл/мин/, нормальный Apo AI элюировали 0,1 М Трис-HCl, 0,04% Na2-ЭДТА, 0,01% NaN3, pH 7,4, содержащим 4 M Гдн-HCl. Apo AI-M был затем элюирован тем же буфером, содержащим 4 М Гдн-HCl и 1% 2-меркаптоэтанола. Фракции, содержащие Apo AI-M, объединяли, диализировали против 5 мМ NH4HCO3, 0,01% Na2-ЭДТА, pH 7,4, лиофилизировали и хранили при -20oC в 0,1 М Трис-HCl, 0,04% Na2-ЭДТА, 0,01% NaN3, pH 7,4, содержащем 3 М Гдн-HCl и 0,1% 2-меркаптоэтанола. Препараты AI-M имели чистоту > 98%, как было показано ВЭЭХ, СДС-ПАГЭ и изоэлектрической фокусировкой.
б). Синтез Apo AI-M/Apo AI-M.
Раствор Apo AI-M диализировали против 25 мМ буфера Трис-HCl, pH 9,0. Восстановленный Apo AI-M разводили до желаемой конечной концентрации /3,6-53,6 мкМ/ 25 мМ буфером Трис-HCl, содержащим восстановленный глутатион /ГСН/ /1-5 мМ/, и предварительно инкубировали при 25oC в течение 5 минут. Окисление начинали путем добавления окисленного глутатиона /ГССГ/ /0,1-10,0 мМ/ и проводили в плотно закрытых пробирках при той же температуре в течение 24 часов. За окислением следили при помощи СДС-ПАГЭ /смотри выше/. После окрашивания и промывания от излишков краски гели сканировали при помощи лазерного денситометра LKB Ultroscan XL и процентное распределение полос индивидуальных белков рассчитывали с использованием программного обеспечения LKB 2400 Gelscan XL. За кинетикой окисления следили с помощью ВЭЭХ.
Для того чтобы оптимизировать синтез димера, проводили дополнительные эксперименты по окислению в присутствии ГСН/ГССГ + Гдн-HCl и ГСН/ГССГ + тиоредоксин /Pigiet VP et al, Proc. Natl. Acad. Sci. USA., 1986; 83: 7643-7647/.
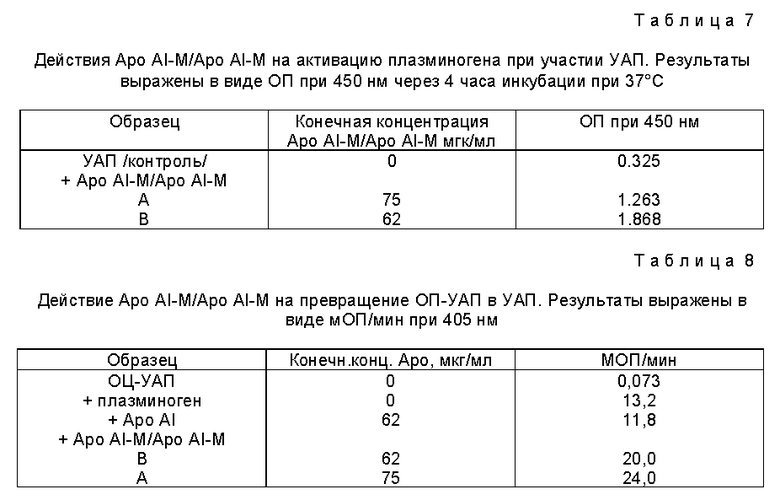
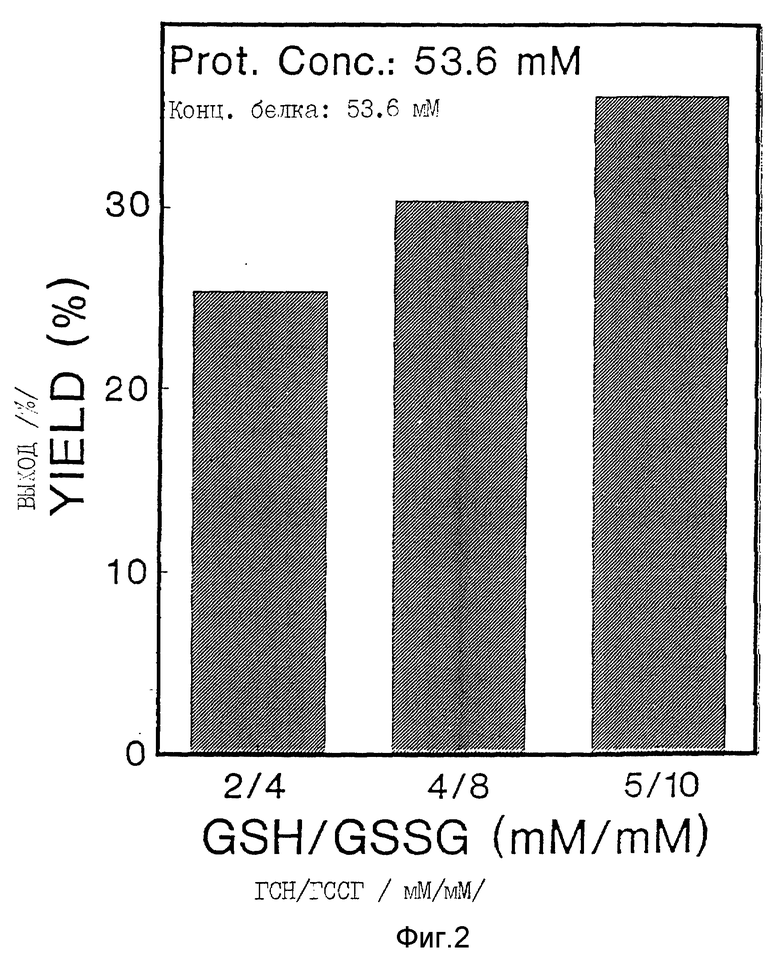
Окисление Apo AI-M проводили в закрытых пробирках в присутствии различных концентраций восстановленного/окисленного глутатиона /ГСН/ГССГ/. Выход димера в этой реакции зависел как от концентрации белка /фиг. 1, табл. 1/, так и от концентраций и соотношения ГСН/ГССГ /фиг. 2, табл. 1/. При увеличении концентрации белка с 8,9 мкМ до 53,6 мкМ процентный выход Apo AI-M/Apo AI-M увеличился с 26% до 51% /фиг. 1, табл. 1/. Уменьшение молярного соотношения ГСН/ГССГ с 1/2 до 1/16 привело к уменьшению на 43% выхода продукта; с другой стороны, повышение концентрации ГСН/ГССГ при постоянном молярном соотношении ГСН/ГССГ было связано с увеличением на 42% образования Apo AI-M/Apo AI-M /фиг. 2, табл. 1/. Ни температура реакции, ни присутствие денатурированного белка /Гдн-HCl/ не влияли на образования Apo AI-M/Apo AI-M. Значительное образование Apo Ai-M/Apo Ai-M также было достигнуто при инкубации Apo AI-M/ с ГСН/ГССГ в присутствии 0,2 мМ тиоредоксина /табл. 1/.
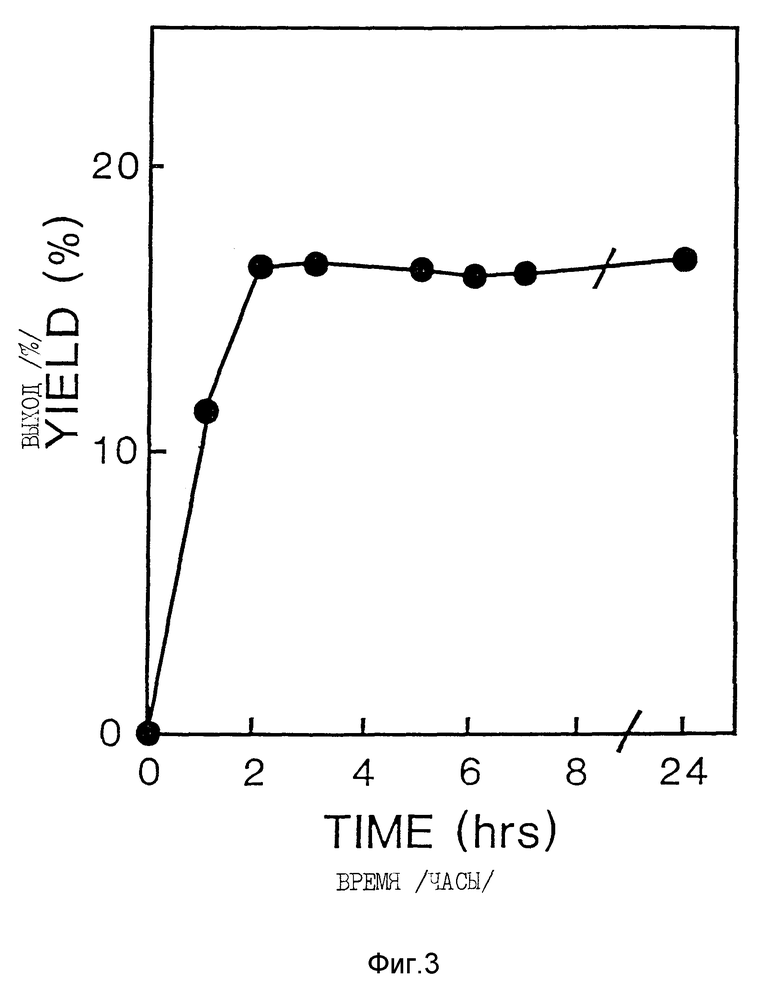
За кинетикой реакции окисления следили при помощи аналитической ВЭЭХ. Димерный и мономерный Apo AI-M давали отчетливые пики с временами задержки 10,8 и 12,7 минут, соответственно. Синтез AI-M/AI-M /ГСН/ГССГ 2 мМ/4 мМ, концентрация AI-M 8,9 мкМ/ почти завершился через 3 часа; продление инкубации до 24 часов не увеличило дальнейшего образования AI-M/AI-M /фиг. 3/.
Пример 3. Рекомбинантное образование Apo AI-M/Apo AI-M.
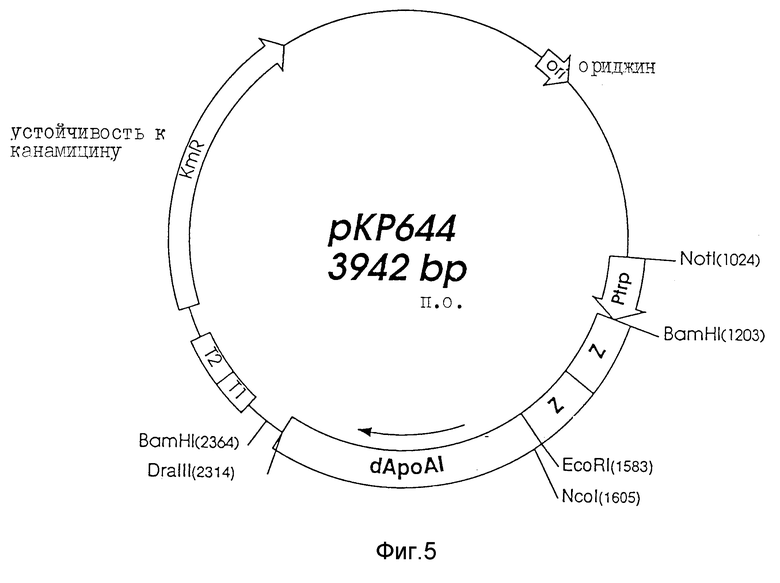
a). Конструкция экспрессионного вектора pKP644.
Репликативную форму бактериофага M13 mp18, содержащую кДНК, кодирующую apo-липопротеин AI-M /Sharpe C.R. et al., Nucleic Acids Research, Vol, 12, N 9, p.3917, 1984 и Cheung M.C. et al., Biochem. Biophys Acta, 960, pp.73-82, 1988/, расщепляли рестрикционным ферментом BamHI и очищали гель-электрофорезом и использованием агарозы с низкой температурой гелеобразования /НТГ/. Фрагмент из 822 п.о., соответствующий гену Apo AI-M, вырезали и лигировали в плазмидой pUC9, которую предварительно расщепляли BamHI и обрабатывали кишечной фосфодиэстеразой теленка.
Лигированную смесь использовали для трансформации компонентных клеток E. coli JM83; белые колонии снимали с агаровых чашек, в которых содержались ампициллин, X-Гал и ИПТГ. Выделенную плазмидную ДНК очищали на колонках Quia Gene /Quia Gene Inc. 9259, Etonave., Chatsworth, Cal. 91311 USA/ в соответствии с рекомендациями производителя. Производная плазмида, названная pUA/Apo AI-M, расщеплялась EcoRI и Ncol. Одну часть расщепленной смеси анализировали с помощью агарозного электрофореза для подтверждения правильной ориентации. Другую часть использовали для лигирования с синтетическим линкером "ΔApo AI-Eco" /фиг. 4/ по 5'-концу гена Apo AI-M для того, чтобы получить ген ΔApoAI, состоящий из 724 п.о. Последовательность ΔApo AI-M образует сайт Asp-Pro на N-конце закодированного белка, что дает возможность расщепления муравьиной кислотой. Компонентные клетки E.coli M83 трансформировали лигированной смесью, и производную плпзмиду /названную pUC/ ΔApo AI-M/ выделяли на колонках Quia Gene, как описано выше.
Плазмиду pUC/ΔApo AI-M и экспрессионный вектор pEZZ расщепляли EcoRI и BamHI, очищали электрофорезом с использованием агарозы НТГ, и фрагмент из 7999 п.о. pUC/ΔApo AI-M лигировали с фрагментом из 2763 п.о. оч. pEZZ. Лигированную смесь использовали для трансформации компетентных клеток E.coli RV 308; плазмидную ДНК получали, как описано выше.
Экспериментальные методики содержатся в Sambrook J., Fritsch. and Maniatis T. (1989). Molecular Cloning; A laboratory Manual 2nd edit. Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y.
Производная плазмида была обозначена pKP644 /фиг. 5/.
б). Экспрессия слитого белка ZZ - ΔApo AI-M.
40 мл ночной культуры E.coli RV 308/pKP644, выращенной при 30o на среде LB с 50 мг/мл канамицина, высевали на минимальную среду A /Curr. Meth. in Mol. Biol./ /2x500 мл/ с добавлением 0,2 г/л казаминовых кислот, 1 мМ MoO4, 0,25% глюкозы и 50 мг/мл канамицина. Культуру инкубировали при 37o в течение 24 часов при энергичном перемешивании.
с). Первичная очистка Apo AI-M.
1,0 л культуры E.coli /RV308/pKP644/, содержащей описанную плазмиду, центрифугировали, и осажденные клетки ресуспендировали в 30 мл 1xTS /25 мМ Трис-HCl, 0,2 M NaCl, 1 мМ ЭДТА/ и 6 М Гдн-HCl, а затем гомогенизировали путем однократного пропускания через французский пресс /SLM. Instruments Inc. /, с рабочим давлением 1000 ф/кв.д. Полученную суспензию инкубировали при комнатной температуре с мягким перемешиванием в течение 1 часа и центрифугировали. Супернатант разводили затем до конечной концентрации Гдн-HCl 1 М /а именно, в 6 раз/ и наносили на колонку с 15 мл Сефарозы-ИгG с быстрым прохождением, уравновешенную 1xTS. После нанесения колонку промывали 5-ю колоночными объемами 1xTS, а затем 20 мМ раствором ацетата аммония с pH 5,4 до тех пор, пока pH элюата не достиг 5,4. Связанный материал элюировали 25 мл 0,2 М HAc, прослеживая поглощение при 280 нм /A280/. Выход продукта с 1 л культуры на основании значений A280 составил 1,9 г.
д). Расщепление слитого белка.
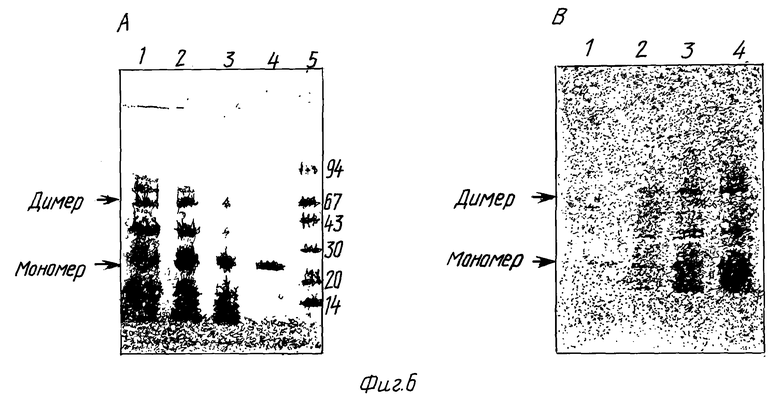
Элюат разделяли на равные части, лиофилизировали и ресуспендировали в 75%, 50% и 25% муравьиной кислоте, соответственно. Растворы инкубировали при 37oC в течение 28 часов и после этого лиофилизировали для удаления муравьиной кислоты. Продукт расщепления проанализировали с использованием СДС-ПАГЭ, сопровождавшимся Вестерн-анализом. Приблизительно 5 мг общего белка наносили на СДС-ПАГЭ с градиентом геля 8 - 25% в невосстанавливающих условиях. Образцы пропускали в двух повторностях. Один гель окрашивали Кумасси, а другой использовали для Вестерн-анализа. Результаты показаны на фиг. 6. Один из продуктов расщепления движется с одинаковой скоростью с очищенным натуральным Apo AI и дает начало сигналам при Вестерн-анализе. Вестерн-анализ проводили с использованием поликлональных антител, коньюгированных с пероксидазой хрена /The Binding site Ltd, Cambridge, Engl./, при появлении использовали стандартные методики.
Фиг. 6 показывает анализ полученных белков после расщепления слитого белка. Плоскость геля A: гель СДС-ПАГЭ /8 - 25%/, окрашенный Кумасси. Дорожка 1: 25% муравьиная кислота; дорожка 2: 50% муравьиная кислота; дорожка 3: 75% муравьиная кислота; дорожка 4: очищенный природный Apo AI /Sigma/ и дорожка 5: маркер LMW /Pharmacia/. Плоскость геля B: Вестерн-анализ второго экземпляра геля. Дорожка 1: очищенный природный Apo AI /Sigma/; дорожка 2: 75% муравьиная кислота; дорожка 3: 50% муравьиная кислота; дорожка 4: 25% муравьиная кислота. Присутствие полосы с удвоенным молекулярным весом Apo AI-M на Вестерн-анализе указывает на присутствие димера Apo AI-M.
Пример 4. Получение Apo AI-M в биореакторах.
Конструкция векторов для прямого выделения Apo AI-M в пориплазматическое пространство и среду для роста.
Штаммы и векторы. Штамм Escherichia coli K12, которые использовались в работе, имели следующие характеристики: HB101F-, hsd S20 (rB-, mB-) sup E44, HB101F-, hsd S20 (rB-, mB-), sup E44, ara 14, I-, gal K2, Iac Y1, pro A2, rsp L20, xyl-5, mtl-1, rec A13, rB-, mB-, mcr A(+), mcr B(-) (Boyer et al., 1969, J. Mol. Biol. 41: 459 - 472), DH5a F-, F80 DlacZDM15, D(lacZYA-argF)U 169, rec AI, end AI, gyr A, I-, thi-I, hsd R17, (rk -, mk +), sup E44, rel AI, (BRL USA), RV308 Dlac X74, gal OP:: I S2 (gal IOP308), strA, I- (Maurer et al., 1980. J. Mol. Biol. 139: 147 - 161) и BC 50 xyl-7, ara-14, T4-R, I-.
Штаммы HB101 и DH5a использовали для субклонирования фрагментов ДНК. Плазмида pUC9 /Vieira et al., 1982, Gene, 19: 259 - 68/ использовалась для субклонирования BamH1-фрагмента в 821 п.о. копии кДНК человеческого Apo AI, полученной от S. Sidoni (Milano). Нуклеотидная последовательность кДНК человеческого Apo AI может быть получена из Банка Генов под каталожным номером XO2162 /Seilhammer et al., 1984. DNA 3: 309 - 317/. Этот вектор был обозначен pKP575. На плазмиде pUC9 также был субклонирован EcoRI-Pst 1-фрагмент в 865 п. о. ДНК человеческого Apo AI-M /копия кДНК получена от S. Sidoni/. Это производное было обозначено pKP576. Плазмиды pKP683 и pKP764 являются производными плазмид pTR с 99 /Amann et al., 1988, Gene, 69: 301-15/ и производного pUC с маркером устойчивости к канамицину, произошедшему от транспозона /Tn903/ из pUC4-K /Vieira et al., 1982, Gene, 19: 259 - 268 и Oka et al. , 1981, J. Mol. Biol, . 147:217/ и терминаторами транскрипции /T1T2/ бактериофага fd из pUEX2 /Bressan et al. , 1987. Nucleic Acid Res. 15: 10056/.
Использованные методы. Бактериальные штаммы выращивали на среде Луриа Бертани /LB/ или на среде с дрожжевым экстрактом и триптоном -2хYT/ с добавлением ампициллина /Ap/ 50 мкг/мл или канамицина /Km/ 70 мкг/мл для получения плазмидной ДНК и для анализа экспрессии в небольшом масштабе /Sambrook et al. , 1989, Cold Spring Harbor Laboratory Press/. Щелочной триптозный кровяной агар /Difco, USA/, дополненный Ap 50 мкг/мл или Km 70 мкг/мл, использовали для выращивания клеток на агаровых чашках. Технику работы ce рекомбинантной ДНК выполняли в соответствии с Sambrook et al., 1989 /Cold Spring Harbor Laboratory Press/. Рестрикционные эндонуклеазы и ДНК-лигаза Т4 были получены от фирмы Boehringer Manheim (Germany), фирмы New England Biolab (Beverly, USA) или фирмы Pharmacia (Uppsala, Sweden). Изопропил-b-D-галактозид /ИПТГ/ был получен от фирмы Sigma (USA). Агароза с низкой температурой плавления и гелеобразования /Nu Sieve GTG, FMC Bioproducts, USA/ использовалась для выделения фрагментов ДНК, ПЦР-амплификации были выполнены с использованием прибора с циклическим изменением температурного режима для ДНК и ДНК-полимеразы Tag от фирмы Perkin-Elmer (Cetus Instuments/Norwalk, USA/. Олигонуклеотидные линкеры и праймеры были синтезированы на приборе Pharmacia-LKB Gene Assembler Plus от фирмы Pharmacia (Uppsala, Sweden/ с использованием фосфиттриэфирного метода на твердой фазе. Определение нуклеотидных последовательностей проводили на приборе Applied Biosystem 373A DNA sequencer с использованием набора Tag Dye DeoxyTM Terminator Cycle Sequencing от фирмы Applied Biosystem (USA).
Компьютерные программы, использованные при работе в ДНК. Для составления плазмидных карт использовали программу Macintosh Plasmid ARTIST /версия 1, 2/ /Clontech, USA/, а для определения последовательности ДНК использовали пакет программ по анализу ГСГ/последовательностей /Genetics Computer Group, Inc. , Madison. Wisconsin, USA). на цифровом компьютере VAX. Конструкция, экспрессия и выделение Apo AI-M из бактерий.
Целью конструирования вектора было получение системы образования и выделения Apo AI-M в E. coli с очень высоким уровнем Apo AI-M, выделяемого в среду для роста. Были синтезированы два олигонуклеотида для слияния кДНК Apo AI и Apo AI-M с фрагментами ДНК, кодирующими последовательности бактериальных сигналов. EcoRI-NcoI-фрагмент в 14 п.о. и NcoI-фрагмент в 40 п.о. плазмиды pKP575 был замещен синтетическим EcoRI-NcoI-фрагментом в 37 п.о. в плазмиде, обозначенной pKP580. Сайт расщепления BbsI на синтетическом фрагменте ДНК образует сайт расщепления, подобный сайту MluI, что дает возможность клонировать различные фрагменты, кодирующие последовательности бактериальных сигналов. Плазмида pKP631 была сконструирована путем замещения NcoI-DraIII-фрагмента в 702 п. о. плазмиды pKP575 /Apo AI/ NcoI-DraIII-фрагмента в 702 п.о. плазмиды pKP576 /Apo AI-M/. Из плазмиды pKP631 BbsI-HindIII-фрагмент в 820 п.о. был выделен и вставлен в сайты MluI и HindIII плазмидного вектора, который обозначили pKP682. Этот вектор содержит промотор tac /Пtac/, производное сигнальной последовательности ompA, два терминатора транскрипции и маркер устойчивости к канамицину. П I-П I-фрагмент в 1501 п. о. был выделен из плазмиды pKP682 и вставлен в подобный вектор, но Пtac был заменен промотором Пtrc. Экспрессионный вектор был обозначен pKP683. Плазмида pKP764 была сконструирована путем замещения DraIII-HinsIII-фрагмента в 88 п.о. плазмиды pKP683 на синтетический фрагмент ДНК в 14 п. о. , содержащий более сильные терминаторы трансляции и разрушенный сайт DraIII /разрушенный при помощи ввода A в конец выступающего 3'-конца DraIII/. Трансформацию штаммов E.coli выполняли как описано в Sambrook et al. , 1989, Cold Spring Harbor Laboratory Press. Плазмидные конструкции, использованные в экспериментах по экспрессии и для образования Apo AI-M, были проанализированы с использованием картирования рестрикционными ферментами, а структурные гены Apo AI-M проверяли определением нуклеотидной последовательности. Штаммы E. coli с подходящими плазмидами, которые использовали для выращивания в биореакторах, получали следующим образом. Клетки выращивали в течение ночи на среде LB или 2 х YT, дополненной Km, в качалочных колбах при 30oC. После центрифугирования клетки ресуспендировали в 1/2 объеме среды, хранящейся в условиях глубокого холода, в соответствии с Gergen et al., 1979. Nucleic Acid Res 7: 2115. Равные части разливали по специальным 1 мл-флаконам для хранения при низких температурах и до использования хранили при -75oC.
Питательные среды для клеток, выращиваемых в биореакторах.
Среда A: 16 г/л триптона /Difco, USA/, 8 г/л дрожжевого экстракта /Difco, USA/, 5 г/л NaCl и 0,05 г/л канамицина. Среда B: 2,5 г/л /NH4/2SO4, 3 г/л K2HPO4, 0,5 г/л Na3-цитрата, 5 г/л дрожжевого экстракта /Difco, USA/. После стерилизации среда дополнялась 10 г/л исходной глюкозы, 0,05 г/л канамицина, 1 г/л MgSO4 • 7H2O и 0,07 г/л тиамингидрохлорида. Добавляли также раствор микроэлементов /1 мл/л/ и раствор витаминов /0,65 мл/л/. Раствор микроэлементов содержал: 27 г/л FeCl3 • 6H2O, 4 г/л ZnSO4 • 7H2O, 7 г/л CoCl2 • 6H2O, 7 г/л Na2MoO4 • 2H2O, 8 г/л CuSO4 • 5H2O, 2 г/л H3BO3, 5 г/л MnSO4 • 4H2O, 11 г/л CaCl2 • 2H2O и 50 мл/л HCl. Раствор витаминов содержал: 0,5 г/л пантотената кальция, 0,5 г/л холин-хлорида, 0,5 г/л фолиевой кислоты, 1 г/л инозита, 0,5 г/л никотинамида, 0,5 г/л пиридоксин-гидрохлорида, 0,05 г/л рибофлавина и 0,5 г/л тиамингидрохлорида. В качестве пеногасителя использовался адеканол /0,2 мл/л/. В случае необходимости в течение культивирования делали дальнейшие добавки пеногасителя.
Культивирование RV308/pKP683 в биореакторе на 3,5 литра. В качестве посевного материала использовали культуру, хранящуюся в условиях глубокого холода; этой культурой засевали 500 мл среды A и сначала культивировали в колбах Эрленмейера объемом 2 л с отбойниками при 30oC в течение 8 - 10 часов. Объем посевного материала, соответствующий 10% от рабочего объема биореактора, переносили в биореактор. Культивирование проводили в биореакторе на 3,5 литра /Belach AB, Sweden/ с рабочим объемом 2,5 л. В течение фазы роста до индукции синтеза белка температура была 30oC, а затем повышалась до 37oC, pH поддерживался на уровне 7,0 с помощью 25% раствора аммиака. Интенсивность аэрации поддерживалась на уровне 1 оом, а давление растворенного кислорода /ДРК/ поддерживалось на уровне 30% при помощи регулирования скорости мешалки. После того как была израсходована начальная глюкоза, стали подавать подпитывающий раствор глюкозы, поддерживая в системе ограничение по глюкозе путем подпитывания 60%-ным раствором глюкозы. Начальная скорость подпитки, 0,04 г/мин, сохранялась в течение 3 часов, а затем постепенно повышалась до 0,4 г/мин в течение 3-х часов. За ростом клеток наблюдали по изменению оптической плотности /ОП/. Концентрацию Apo AI-M в надосадочной жидкости определяли при помощи радиоиммуноанализа /набор Apolipoprotein AI RTA 100, art. no. 109152-01, Kabi Pharmacia, Sweden/. После 16 часов культивирования, когда ОП составляла 58, индуцировали синтез белка путем добавления 0,5 мМ ИПТГ, а температуру увеличивали до 37oC. Через 4 часа после индукции концентрация Apo AI-M составляла 2,3 г/л, а через два дополнительных часа была 2,5 г/л.
Культивирование BC50/pKP764 в биореакторе на 3,5 л.
Культивирование выполняли, как описано выше, за исключением того, что к среде в биореакторе не добавляли канамицин. Через 15 часов, когда ОП составляла 60, добавляли ИПТГ и повышали температуру. Через 10 часов концентрация Apo AI-M в надосадочной жидкости составляла 3,7 г/л, а через 22 часа после индукции концентрация составляла 4,4 г/л.
Культивирование BC50/pKP764 в биореакторе на 300 литров.
Использовали биореактор на 300 литров /Chemoferm AB, Sweden/ с рабочим объемом в 180 литров. Посевной материал готовили, как описано выше для выращивания RV308/pKP683 в биореакторе на 3,5 литра, за исключением того, что время предварительного культивирования в качалочных колбах было 14 часов. Посевной материал переносили в биореактор для выращивания посевного материала на 50 литров с рабочим объемом 18 литров. Средой, которая использовалась в качалочных колбах, так же как и в биореакторе, была среда A. Среда в посевном биореакторе была дополнена 5 г/л глюкозы, а температура была 30oC, pH и аэрация были такими же, как описано выше для выращивания RV308/pKP683 в биореакторе на 3,5 литра, а ОП никогда не превышала 30%. Когда ОП культуры достигла 4, содержимое посевного биореактора было перенесено в биореактор на 300 литров. В этом биореакторе температура, pH и аэрация среды были такими же, как описаны выше для выращивания RV308/pKP683 в биореакторе на 3,5 литра. Перед индукцией ДРК поддерживали на уровне 30% или несколько выше при помощи увеличения скорости мешалки до ее максимального значения, а затем увеличивали давление воздуха. После индукции давление воздуха было увеличено до 2 бар, что привело к изменению ДРК на 15 - 20%. Через 16 часов культивирования в биореакторе, когда ОП культуры составляла 51, добавляли ИПТГ и повышали температуру до 37oC. Концентрация Apo AI-M, как мономера и как димера, составляла 1,3 г/л; через 5 часов после индукции и в течение следующего часа, в то время как охлаждался биореактор, концентрация Apo AI-M увеличилась до 1,5 г/л.
Все количество присутствующего мономера было превращено в димер и очищено в соответствии с традиционными методами.
Пример 5. Характеристика Apo AI-M/Apo AI-M из плазмы.
Очищенный Apo AI-M/Apo AI-M из примера 1 после прохождения через СДС-ПАГЭ в невосстанавливающих условиях дает одну полосу. В аналитическом СДС-ПАГЭ очевидный молекулярный вес белка составил, как и ожидалось, 56 кД, Apo AI-M/Apo AI-M показал сложный изоформный образец, характеризующийся присутствием по крайней мере 6 различных белковых полос в диапазоне 5,3 - 5,6 ИЭТ в невосстанавливающих, денатурированных гелях для ИЭФ.
Спектр УФ Apo AI-M/Apo AI-M /1,1 мг/мл показал типичный максимум поглощения при 280 нм с плечом на 290,2 нм /фиг. 7/. E расчетного белка /1 см, 1%/ при 280 нм составлял 16,9. Для оценки обнажения тирозиновых остатков в молекуле Apo AI-M/Apo AI-M выполняли анализ второй производной УФ-спектра, как описано у Рагона P. и др. "Определение обнажения тирозина в белках спектроскопией второй производной" /Biochemistry, 1984; 23: 1871 - 1875/. 2 УФ-спектр второй производной в типичном случае показывает два минимума, сосредоточенных вокруг 283 и 290,5 нм, и два максимума около 287 и 295 нм /фиг. 8 - 10/. Относительные степени обнажения тирозина /альфа/, рассчитанные для натурального Apo AI-M/Apo AI-M и Apo AI-M/Apo AI-M + ДМФХ были 0,75 и 0,49 соответственно /табл. 2/.
Были зарегистрированы спектры флуоресценции Apo AI-M/Apo AI-M /0,1 мг/мл/ и в возбужденном состоянии и во время эмиссии. Максимальная длина волны при возбуждении остатков триптофана в Apo AI-M/Apo AI-M составляла 280 нм, и не изменялась вследствие соединения с ДМФХ. Спектр эмиссии /возбуждение при 280 нм/ показал максимальную длину волны 344 нм /фиг. 11/; соединение с ДМФХ вызвало сдвиг этого максимума в "голубую" сторону /338 нм/ в связи с увеличением на 24% интенсивности флуоресценции в этом максимуме /фиг. 11/.
Спектр КД в дальней УФ-области Apo AI-M/Apo AI-M характеризуется типичными минимумами при 208 нм и 22 нм и максимумами около 195 нм /фиг. 12/. Содержание альфа-спиралей значительно увеличивалось с увеличением концентрации белка от 0,1 мг/мл до 1,1 мг/мл /фиг. 12, табл. 2/. Соединение Apo AI-M/Apo AI-M /0,1 мг/мл/ с ДМФХ вызвало дальнейшее увеличение альфа-спиральной структуры белка /фиг. 2, табл. 2/.
Методы характеристики продукта.
Инкубация с фосфолипидами.
Взвешенные количества димиристоилфосфатилхолина /ДМФХ/ растворяли в этаноле и выпаривали растворитель под N2; какие-либо остатки растворителя были удалены под вакуумом в течение 2 часов. Суспензию ДМФХ в 20 мМ фосфатном буфере, pH 7,4, смешивали с Apo AI-M/Apo AI-M /конечная концентрация 0,1 мг/мл/ в молярном соотношении ДМФХ /Apo AI-M/Apo AI-M 100:1.
Спектроскопия.
Растворы Apo AI-M/Apo AI-M диализовали против 20 мМ фосфатного буфера, pH 7,4, и разводили тем же буфером до желаемой концентрации белка.
УФ-спектры /нормальные и вторых производных/ растворов Apo AI-M/Apo AI-M и Apo AI-M/Apo AI-M 1 ДМФХ регистрировали на спектрометрах Jasco Uvides-610 и Perkin Elmer Lambda-2 при 25oC с использованием кварцевых кювет 1 см.
Топографическое расположение остатков тирозина было исследовано в соответствии с уравнением Рагона и др. /Рагон Р., Колонна Ж., Балестриери С., Сервилло Л., Ираке Ж. Определение обнажения тирозина в белках спектроскопией второй производной. Biochemistry, 1984; 23: 1871 - 1875/:
Альфа = (rn - ra)/(ru - ra),
где альфа является степенью обнажения тирозина в растворителе, rn и ru являются отношениями пиков производных /а/в/ для натурального и развернутого /в 6 М Гдн-HCl/ Apo AI-M/Apo AI-M соответственно, а ra является отношением пика второй производной раствора, содержащего свободные тирозин и триптофан, смешанные в таком же молярном соотношении, каким оно является в молекуле Apo AI-M/Apo AI-M.
Флуоресцентные спектры, присущие растворам Apo AI-M/Apo AI-M и Apo AI-M/Apo AI-M + ДМФХ, регистрировали на спектрофлуориметре Jasco FP-550 при 25oC.
Спектры кругового дихроизма /КД/ растворов Apo AI-M/Apo AI-M и Apo AI-M/Apo AI-M + ДМФХ регистрировали на спектрополяриметре Jasco J 500A при 25oC.
Значения величин эллиптичности остатка [THEYA] выражали в градусах • см2 • дмоль-1 и рассчитывали по уравнению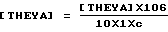
где [THEYA] является наблюдаемой эллиптичностью в градусах: 106 обозначает молекулярный вес остатка белка, 1 является длиной пробега в см, а с является концентрацией белка в г/мл.
Процент альфа-спиралей рассчитывался с использованием уравнения
/Гринфилд Н., Фасман Ж.Д. Расчет спектров кругового дихроизма для оценки конформации белка. Biochemistry, 1969; 8: 4108 - 4116/.
Электрофорез.
Аналитическую изоэлектрическую фокусировку с СДС-ПАГЭ выполняли как описано предварительно /Францесчини Ж., Сиртори М., Жанфранцесчи Ж., Сиртори Ц. Р. Отношения между apo-протеинами ЛВП и изопротеинами AI у организмов с признаком AI-M. Metabolism, 1981; 30: 502 - 509/.
Изоэлектрическую фокусировку выполняли в 10% акриламидных гелях, содержащих 6 М мочевину и 4% амфолинов /pH 4 - 6/. После фокусировки в течение ночи гели фиксировали и окрашивали красителем Кумасси бриллиантовой голубой Р-250, растворенным в смеси уксусная кислота/изопропиловый спирт. Изоэлектрические точки /ИЭТ/ полос любых неизвестных белков рассчитывали при построении графика ИЭТ известных белков /стандарты от фирмы Bio-Pad и apo-ЛВП/ против соответствующих расстояний перемещения.
Для СДС-ПАГЭ использовали 14% акриламидные гели, содержащие 0,1% СДС. Гели обрабатывали, как описано: молекулярный вес неизвестных белков вычисляли по графику log MB стандартных белков /KAB1-Pharmacia/ против расстояний перемещения.
Высокоэффективная эксклюзионная хроматография.
Аналитические разделения ВЭЭХ выполняли с использованием жидкостного хроматографа Jasco, снабженного колонкой 10 мкм TSK-G3000SW /7,5 x 300 мм/. Колонка ВЭЭХ была уравновешена и элюировалась 0,1 М фосфатным буфером, 0,1 М NaCl, pH 7,2, содержащим 8 М мочевину. Белки элюировали при объемной скорости потока 0,5 мл/мин, считывание показаний прибора производилось при 220 нм. Площадь пиков определяли с использованием интегратора НР-3390.
Пример 6. Производство частиц рДВП, содержащих Apo AI, Apo AI-M или Apo AI-M/Apo AI-M.
Вновь составленные частицы липопротеинов с высокой плотностью /рЛВП/ были получены с использованием методики, представленной Nichols A.V. et al. Biochim. Biophys. Acta., 750: 353 - 364 /1983/ и Matz C.E. et al., J. Biol. Chem. 257: 4535 - 4541 /1982/.
Димер рекомбинантного Apo AI-M /из примера 4/ и нормальный Apo AI, очищенный из человеческой плазмы, растворяли в 10 мМ Трис-HCl, 0,15 М NaCl, 0,01% ЭДТА, 0,006 NaN3, pH 8,0 /буфер A/, содержащем 4 М Гдн-HCl в концентрации 6 мг/мл. Для сравнения, дисульфидные связи в некоторой части Apo AI-M/Apo AI-M восстанавливали добавлением 20 мМ ДТГ к буферу A + Гдн-HCl. Белки подвергали глубокому диализу против буфера A и разводили этим же буфером до концентрации 5,2 мг/мл.
Фосфолипиды - либо яичный фосфатидилхолин /ЯФХ/, либо пальмитоилолеилфосфатидилхолин /ПОФХ/ - растворяли в CHCl3, высушивали по N2 и хранили под вакуумом в течение ночи. Добавляли хелат натрия в соотношении хелат/ФХ 0,55 /в/в/; смесь энергично перемешивали в течение 3 минут при комнатной температуре и инкубировали при 4oC в течение 2 часов. Затем добавляли белок в весовом соотношении ФХ/белок 2,17 /для ПОФХ/ или 2,47 /для ЯФХ/, смесь перемешивали в течение 3 минут при комнатной температуре и инкубировали при 4oC в течение ночи. После диализа против буфера A в течение 5 дней смесь центрифугировали при 11000 об/мин в течение 5 минут на микроцентрифуге Beckman и собирали надосадочную жидкость.
рЛВП отделяли при помощи неденатурирующего полиакриламидного градиентного гель-электрофореза /ГГЭ/, размер частиц определяли, как предварительно описано Nichols A.V. et al., Meth. Enzymol. 128: 417 - 431 /1986/.
Все проверенные apo-липопротеины после описанной процедуры почти полностью соединялись с липидами, что явствует из весьма незначительных пиков безлипидных apo-липопротеинов на гелях ГГЭ. Включение белка в рЛВП колебалось между 68% и 100% в 10 различных препаратах.
Профили ГГЭ вновь составленных частиц ЛВП показаны на фиг. 13. ЛВП, составленные из Apo AI и ЯФХ, дают основной пик на ГГЭ с диаметром 9,6 нм; также были обнаружены второстепенные компоненты как большего, так и меньшего размера. рЛВП, содержащие ЯФХ и Apo AI-M/Apo AI-M, состоят из двух основных /диаметры: 8,6 и 12,9 нм/ и двух второстепенных /диаметры: 7,9 и 10,8 нм/ компонентов /фиг. 13/; тот же самый размер частиц был получен, когда Apo AI-M/Apo AI-M включался в частицы вместе с ПОФХ.
Все три apo-липопротеина были почти полностью включены в стабильные липидно-белковые комплексы с различным размером частиц рЛВП, распределением и составом. В частности, рЛВП, сделанные из рекомбинантного Apo AI-M/Apo AI-M, состоят из двух основных компонентов, больший из которых является уникальным среди семейства Apo AI-M, содержащих рЛВП.
Биологическая оценка Apo AI-M/Apo AI-M.
Пример 7. Кинетическое поведение димера Apo AI-M в сравнении с мономером Apo AI-M у нормальных реципиентов.
Димеры продемонстрировали продленное присутствие в кровеносной системе, что было показано в исследованиях кинетически у человека, представленных ниже.
Здоровые добровольцы получали внутривенно 125I-меченый Apo AI-M или Apo AI-M/Apo AI-M. В табл. 3 показано время полураспада в плазме βt 1/2 /ч/, вычисленное в соответствии с двумя различными моделями, а также скорость дробленого разложения /СДР/. Оба показателя подтверждают заметно сниженный катаболизм димера по сравнению с мономером. Очень медленный катаболизм Apo AI-M/Apo AI-M указывает на то, что эти молекулы могут снижать превращение липопротеинов и, возможно, действовать в качестве эффективного предшественника Apo AI-M. Таким образом, после инъекции Apo AI-M/Apo AI-M можно предвидеть, что димер сможет оставаться в плазме в течение продолжительного периода времени и, следовательно, прямо воздействовать на метаболизм липопротеинов и на систему разрушения фибрина.
Влияние Apo AI-M/Apo AI-M на систему разрушения фибрина.
Вступление.
Система разрушения фибрина является главной защитой от фибриновых отложений на стенках сосудов и в этом качестве играет важную роль среди механизмов, которые предотвращают тромбоз.
Ферментом, ответственным за разрушение фибрина, является плазмин. Плазмин образуется из неактивного предшественника - плазминогена - под действием специфических активаторов /тканевый активатор плазминогена, ТАП, и урокиназа, УАП/. И процесс активации, и действие плазмина регулируются специфическими ингибиторами - ингибитор активатора плазминогена 1 /ИАП/ и альфа-2-антиплазмин соответственно. Схема фибриноразрушительной системы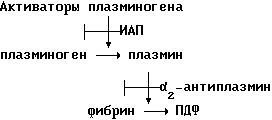
ПДФ = продукт деградации фибрина.
Целью настоящего исследования в примерах 7 - 9 было установить, как димер Apo AI-M действует на человеческую фибрино-разрушительную систему. И самоактивация плазминогена, и его активация ТАП и УАП были изучены в присутствии и в отсутствие Apo AI-M/Apo AI-M. Человеческий Apo AI, выделенный из плазмы, использовался в качестве контроля /продукт фирмы Sigma, N = A 9284/.
Активация фибриноразрушающей системы измерялась при помощи хромогенных субстратов. Эти субстраты содержат хромофорную группу паранитроанилина, который может отщепляться от молекулы субстрата под действием плазмина. Свободный п-НА имеет яркую желтую окраску, за которой можно легко следить при длине волны 405 нм. Количество п-НА, которое освобождается, прямо пропорционально количеству образовавшейся ферментной активности. Все измерения были сделаны с использованием устройства для считывания результатов с микрочашек THEPMO max, управляемого программой SOFTmaxTM, версия 2.01, из приборного обеспечения работ на молекулярном уровне /Menlo. Park. CA., USA/. Серии образцов, использованные в этих исследованиях, были получены рекомбинантным способом в соответствии с примером 4. Очистка включала все традиционные биохимические методы: ионообменную хроматографию, хроматографию с использованием гидрофобного взаимодействия и гель-фильтрационную хроматографию, сопровождающуюся ультрафильтрацией и высушиванием из замороженного состояния. Все исследованные серии образцов содержали 90% димерной формы, что было показано обратнофазовой ВЭЖХ. Концентрацию определяли с использованием Kabi Pharmacia apolipoprotein AIRIA-100.
Были исследованы три препарата - A, B и C.
Пример 8. Самоактивация плазминогена в присутствии Apo AI-M/Apo AI-M и Apo AI.
Глу-плазминоген /конечная концентрация 94 мкг/мл/ инкубировали в течение трех часов при 37oC в 0,1 моль/л Трис-буфере, pH 7,6. За образованием плазмина следили с помощью хромогенного субстрата S-2251 /H-DVal-L-Leu-L-Lys-пНА/, полученного от фирмы Chromogenix AB, Molndal, Sweden. Он был в конечной концентрации 0,6 ммоль/л. Плазминогены, использованные в этом исследовании, были получены от фирмы Chromogenix AB или от фирмы IMCO Inc. Stockholm, Sweden.
В этом исследовании были проверены серии образцов Apo AI-M/Apo AI-M /конечная концентрация 3,9 - 75 мкг/мл/; количество плазмина, образующееся в их присутствии, сравнивали с количеством плазмина, образующегося в присутствии Apo AI /конечная концентрация 125 мкг/мл/, и с количеством плазмина, образующегося в отсутствие каких-либо добавок /контроль/ /табл. 4/.
Неожиданно было обнаружено, что Apo AI-M/Apo AI-M может повышать активацию плазминогена в отсутствие каких-либо активаторов плазминогена. Apo AI, произошедший из плазмы, никаким образом не влиял на молекулу плазминогена.
Наблюдаемая активность может быть приписана Apo AI-M/Apo AI-M. Однако на основании этих данных нельзя исключить, что Apo AI-M/Apo AI-M был загрязнен каким-то протеолитическим ферментом(ами), который может активировать плазминоген. Были выполнены эксперименты для того, чтобы исключить такую возможность.
Все препараты Apo AI-M/Apo AI-M, использованные в фибриноразрушающих анализах, были проверены с хромогенными субстратами S-2251 /H-D-Val-L-Leu-L-Lys-пНА/, чувствительному к плазминоподобной активности, и S-2288 /H-D-Ile-Pro-Arg-пНА/, чувствительному к Arg-специфичным протеазам. Субстраты были получены от фирмы Chromogenix AB. Конечная концентрация Apo AI-M/Apo AI-M в этих анализах была одинаковой с концентрацией, использованной в фибриноразрушающих экспериментах и, таким образом, могла колебаться между различными сериями образцов.
Анализ: 25 мкл Apo AI-M/Apo AI-M /конечная концентрация 60 - 70 мкг/мл/, 150 мкл 0,1 моль/л Трис-буфера, pH 7,8, 50 мкл S-2251 /0,6 ммоль/л/ или S-2288 /1,0 ммоль/л/.
Образец, содержащий только буфер и субстрат, использовали в качестве контроля неспецифического гидролиза субстрата. Все образцы были проанализированы в двух повторностях.
Чашки для микротитрования инкубировали при 37oC и показатели поглощения регистрировали с часовыми интервалами.
В другой серии экспериментов Apo AI-M/Apo AI-M обрабатывали необратимым ингибитором сериновых протеаз - диизопропил -фторфосфатом, ДФФ /продукт фирмы Sigma, N = D0789/. Конечная концентрация Apo AI-M/Apo AI-M в буфере 0,2 моль/л KHCO3, pH 7,6, составляла, приблизительно, 75 мкг/мл. ДФФ, конечная концентрация 123 ммоль/л, добавляли к этому раствору. Через 4 часа инкубации образцы диализовали в течение ночи против двух смен карбонатного буфера.
Определение активности с использованием тех же условий, которые описаны раньше, выполняли с Apo AI-M/Apo AI-M, обработанным ДФФ, и с необработанным Apo AI-M/Apo AI-M. Через 3 часа инкубации с плазминогеном и S-2251, ОП при 405 нм составляла 0,209 для образцов, содержащих Apo AI-M/Apo AI-M, обработанный ДФФ, 0,234 для образцов, содержащих необработанный Apo AI-M/Apo AI-M, и 0,030 для образцов, содержащих только плазминоген.
Таким образом, можно заключить, что наблюдаемый эффект активации плазминогена прямо связан с присутствием Apo AI-M/Apo AI-M, а не вызван какими-либо возможными протеолитическими загрязнителями.
Пример 9. Действие Apo AI-M/Apo AI-M на активацию плазминогена в присутствии активаторов плазминогена ТАП и УАП.
И урокиназа /УАП/, и тканевый активатор плазминогена /ТАП/ превращают плазминоген в плазмин путем протеолитического расщепления одной пептидной связи - Arg 560-Val 561 в молекуле плазминогена. В то время как двухцепочечная урокиназа может прямо активировать плазминоген, ТАП требует присутствия фибрина для своей оптимальной активации плазминогена. Присутствие каталитических количеств фибрина, который вместе с ТАП и плазминогеном образует тройной комплекс, будет повышать ферментную эффективность ТАП приблизительно в 600 раз.
Активацию плазминогена с участием ТАП изучали при помощи выпускаемого промышленностью набора SpectrolyseR /фибрин/ ТАП/ИАП от фирмы Biopool AB, Umea, Sweden.
В этом анализе плазминоген инкубировали с ТАП в присутствии хромогенного субстрата D-But-CHT-Lys-пНА и дезАА-фибриногена /мономер фибрина/, который действует как стимулятор активации. Облучающийся плазмин расщепляет субстрат с образованием свободного пНА.
Apo-AI-M/Apo-AI-M добавляли к этой системе и сравнивали с препаратом Apo AI от фирмы Sigma. Действие обоих apo-липопротеинов проверяли в этой системе как в присутствии, так и в отсутствие фибрина.
Пример анализирующей системы:
25 мкл буфера Spectrolyse,
25 мкл препарата Apo или буфера 20 мкл ТАРП /конечная концентрация = 1,7 МЕ/мл/,
150 мкл реагента Spectrolyse PAR /=смесь плазминогена и субстрата/,
10 мкл Desa fib /=мономер фибрина/ или 10 мкл буфера.
Образцы инкубировали в чашках для микротитрования при 37oC в течение трех часов.
Значительная стимуляция активации плазминогена наблюдалась при действии Apo AI-M/Apo AI-M как в присутствии, так и в отсутствие фибрина. Apo AI стимулировал активацию в меньшей степени в присутствии фибрина. В отсутствие фибрина стимуляция при действии Apo AI-M/Apo AI-M была очень резко выраженной в сравнении с очень низкой стимуляцией при действии Apo AI.
Apo AI-M/Apo AI-M также показал значительный усиливающий эффект, когда плазминоген активировали при помощи УАП. Урокиназа, использованная в этом анализе, представляла собой препарат с высоким молекулярным весом, полученный от фирмы Calbiochem.
Урокиназу /конечная концентрация 2,5 МЕ/мл/ смешивали с плазминогеном /конечная концентрация 94 мкг/мл/ и хромогенным субстратом S-2251 /конечная концентрация 0,6 ммоль/л/. К этой смеси добавляли образец Apo AI-M/Apo AI-M в конечной концентрации 75 или 62 мкг/мл. Реакцию проводили в 0,1 моль/л Трис-буфере, pH 7,6.
Подобно ТАП, наблюдалась сильная стимуляция образования плазмина при участии УАП, когда к анализирующей смеси добавляли APO AI-M/Apo AI-M /табл. 7/.
Усиливающее действие на разрушение фибрина также сохраняется, когда Apo AI-M/Apo AI-M приготовлен в виде фармацевтического состава вместе с носителем. В качестве возможного носителя применялись липосомы, состоящие из фосфатидилхолина, ФХ /12 мг/мл/. Концентрация Apo AI-M/Apo AI-M /серия образцов C/ в липосомах была 3,6 мг/мл.
Активацию плазминогена /конечная концентрация 94 мкг/мл/ с участием УАП /конечная концентрация 2,5 МЕ/мл/ проверяли в присутствии липосом, заполненных Apo AI-M/Apo AI-M, и сравнивали с активацией, осуществляющейся в присутствии липосом, не содержащих белка. Образцы инкубировали в течение четырех часов при 37oC с S-2251; за образованием плазмина непрерывно следили.
Активация плазминогена с участием УАП. Действие APO AI-M/Apo AI-M /серия образцов C/ в липосомах. Результаты выражены в виде ОП при 405 нм.
Образец - ОП при 405 нм
УАП + плазминоген + липосомы, не содержащие белка - 0,221
250 мкг/мл ФХ + Apo AI-M/Apo AI-M , 75 мкг/мл - 0,499
в липосомах; 250 мкг/мл ФХ - 1,084
Присутствие пустых липосом стимулирует активацию плазминогена приблизительно в 2 раза. Добавление Apo AI-M/Apo AI-M в липосомы увеличивает этот эффект в пять раз - в сравнении с образцом, содержащим только урокиназный активатор.
Пример 10. Действие Apo AI-M/Apo AI-M на превращение одноцепочечной урокиназы в двухцепочечную урокиназу.
Одноцепочечная урокиназа /ОЦ-УАП/ является предшественником двухцепочечной урокиназы /УАП/. В отличие от УАП ОЦ-УАП обладает только очень низкой активностью, разрушающей амидную связь, в отношении небольших синтетических субстратов. Активность, разрушающая амидную связь, составляет самое большее 0,4% от активности УАП. Однако несмотря на то что ОЦ-УАП является про-ферментом, она обладает способностью активировать плазминоген в плазмин. Предполагается, что в смеси плазминогена и ОЦ-УАП в активацию плазминогена в плазмин вовлечена последовательность из трех реакций:
1) ОЦ-УАП + плазминоген ---> ОЦ-УАП + плазмин
2) плазмин + ОЦ-УАП ---> плазмин + УАП
3) УАП + плазминоген ---> УАП + плазмин
Мы изучили последовательность реакций, приводящих к превращению ОЦ-УАП в УАП в присутствии плазминогена. Урокиназную активность обнаруживали с помощью урокиназно-специфичного хромогенного субстрата S-2444 /пиро-Glu-Gly-Arg-пНА/ от фирмы Chromogenix AB. Серийные образцы Apo AI-M/Apo AI-M и Apo AI добавляли к системе, и величину образовавшейся активности УАП сравнивали с активностью, полученной в образцах без добавления apo-липопротеина. Одноцепочечная урокиназа, использованная в этих анализах, была рекомбинантным продуктом, полученным от фирмы Grunenthal GmbH, Aachen, Germany /N партии 0088808/.
25 мкг Apo или буфера;
75 мкл 0,05 моль/л Трис, pH 7,6, содержащего 0,1 моль/л NaCl и 0,02% Tween 80;
25 мкл ОЦ-УАП, конечная концентрация 5454 нмоль/л;
50 мкл S-2444, конечная концентрация 1 ммоль/л;
25 мкл/л плазминогена, конечная концентрация 52,1 нмоль/л.
Образцы инкубировали при 37oC в течение 90 минут. За увеличением оптической плотности, отражающей количество образующегося УАП, следили непрерывно в течение последних 20 минут инкубации /см. табл. 8/.
Превращение ОЦ-УАП в УАП в присутствии плазминогена стимулировалось Apo AI-M/Apo AI-M, в то время как Apo AI, выделенный из плазмы, не имел значительного влияния на эту систему.
Наблюдаемое действие Apo AI-M/Apo AI-M на систему, разрушающую фибрин, которая применялась в этом исследовании, было выше по сравнению с действием, показанным Apo AI, выделенным из плазмы. Apo AI-M/Apo AI-M обладает сильной способностью стимулировать активность, разрушающую фибрин, которая превышает подобную активность Apo AI. Вероятно, что этот превышающий эффект Apo AI-M/Apo AI-M по отношению к Apo AI также будет обнаружен in vivo.
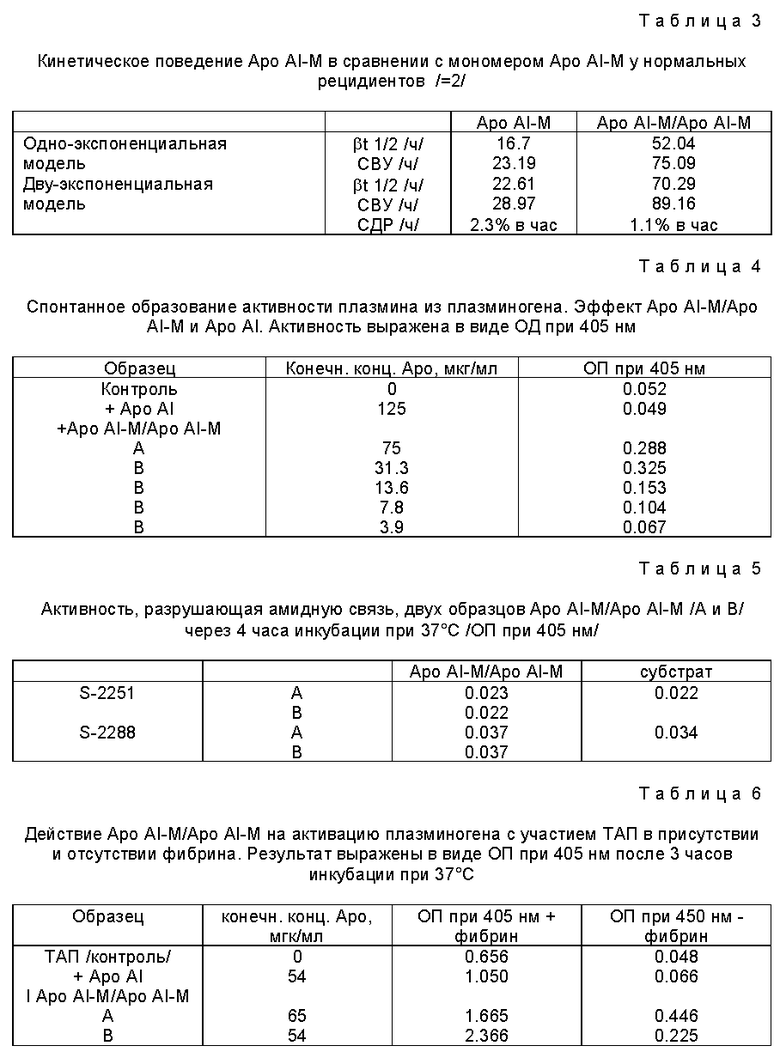

Изобретение относится к медицине и может быть использовано при получении димера молекулярного варианта апо-липопротеина AI-Milano и применении его для изготовления лекарств при профилактике и лечении атеросклероза и сердечно-сосудистых заболеваний. Заявленный димер характеризуется чистотой по меньшей мере 90%, предпочтительно по меньшей мере 98%. Димер апо-липопротеина AI-Milano получают из крови носителей или при помощи рекомбинантной технологии в экспрессионной системе в E. coli или в виде внутриклеточного слитого белка в E. coli. Фармацевтическая композиция, содержащая в качестве активного начала димер апо-липопротеина, дополнительно может содержать носитель и/или стабилизирующее соединение, предпочтительно липидное, и/или агент, снижающий содержание липидов в организме. Фармацевтическая композиция, содержащая димер, может быть использована для лечения атеросклероза и сердечно-сосудистых заболеваний, как пролекарство для мономера апо-липопротеина, для предотвращения тромбозов. Способ позволяет получить димер апо-липопротеина, по существу, в чистой форме, обладающий продленным периодом полураспада в плазме и улучшенной возможностью способствовать расщеплению фибрина в организме по сравнению с мономером. 6 с. и 21 з.п.ф-лы, 13 ил., 8 табл.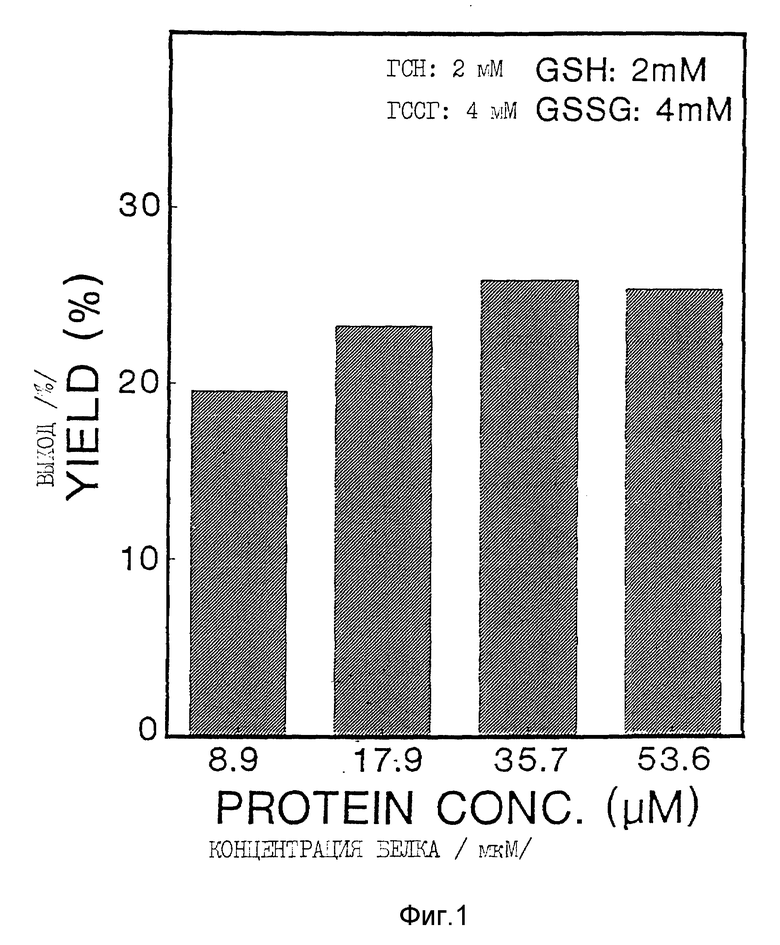
| Пожарный двухцилиндровый насос | 0 |
|
SU90A1 |
| ЫЙ ПЬЁЗОЭЛЕМЁНТ АТ-СРЁЗА | 0 |
|
SU267703A1 |
| Препарат фирмы Берингер Ингельхайм "Актелизе" | |||
| Топка с несколькими решетками для твердого топлива | 1918 |
|
SU8A1 |

