Изобретение относится к новому усовершенствованному тканевому активному плазминогена (усовершенствованному АПТ), имеющему пролонгированный полупериод существования в организме и повышенную стабильность к воздействию тепла и кислот, который может быть использован для подавления воспламенения вокруг области тромбообразования.
Изобретение также касается способа получения названного тканевого активатора плазминогена с помощью технологии рекомбинантных ДНК и средств, используемых для его осуществления.
Известно, что человеческий тканевый активатор плазминогена (АПТ) обладает полезной фибринолитической активностью и чрезвычайно эффективен в отношении фибринсвязанного плазминогена, тогда как плазминоген в фазе свободной циркуляции в организме он активирует не столь эффективно как обычные тромболитические средства, стрептокиназа (СК) и урокиназа (УК). Известны аминокислотная последовательность человеческого АПТ и нуклеотидная последовательность кДНК, кодирующей человеческий АПТ (Pennica. D., et al., Nature, 301, 214-221, 1983). Также известно, что человеческий АПТ растворяет сгустки венозной и артериальной крови. В крупномасштабных клинических исследованиях отмечается, что человеческий АПТ, вводимый внутривенно, эффективен при повторной перфузии закупоривающейся венечной артерии у пациента с острым инфарктом миокарда.
Однако недостатком применения этого препарата при лечении заболевания, связанного с тромбообразованием, является крайне короткий полупериод существования его ферментативной активности в крови (Rijken, D.C., et al., Thromb. Heamost. 54 (1), 61, 1985, Hubert, E.F., et al., Blood, 65, 539, 1985). При использовании для лечения человеческий АПТ приходится применять в виде непрерывной внутривенной инъекции с высокой дозой.
Известно, что встречающийся в природе человеческий АПТ имеет доменную структуру, начиная от N-конца молекулы следуют фингердомен, домен ФРЭ (фактор роста эпидермиса), два домена "крингл 1" и "крингл 2" и домер серин-протеазы. В работе Rijken et al., отмечается (Rijken D.C., et al., Thromb. Heamost., 54 (1), 61, 1985), что непродолжительность биологического полупериода существования человеческого АПТ может иметь отношение ко всем доменам человеческого АПТ, кроме домена серин-протеазы. В работе Zonneveld et al. (Zonneveld, A.J.V., et al, Proc. Natl. Acad. USA., 83, 4670, 1986) также отмечается, что фингердомен, домен ФРЭ и домен "крингл 2" могут иметь важное значение для фибринсвязывающей активности встречающегося в природе человеческого АПТ, а также для поддержания фибринзависимой активации АПТ. Однако до сих пор не разработаны какие-либо конкретные меры для поддержания фибринсвязующей активности, которой обладает встречающийся в природе человеческий АПТ, и его фибринзависимой активности, а также для пролонгирования биологического полупериода существования.
В опубликованной выложенной заявке на патент Японии N 48378/1987 описывается АПТ, полученный делецией 87-175 аминокислот встречающегося в природе человеческого АПТ, в котором "крингл 1" делетирован. Этот АПТ отличается дополнительной индуцированной точечной мутацией в области фактора роста эпидермиса. В заявке на патент Японии раскрывается, что модифицированный АПТ имеет способность связываться с фибрином, но взаимодействие с ингибитором тканевого активатора плазминогена является ослабленным.
В Европатенте N 241208 описывается АПТ, полученный делецией 92-179 аминокислот встречающегося в природе человеческого АПТ, в котором также делетирован "крингл 1". В данной работе упоминается, что этот АПТ имеет фибринолитическую активность.
Кроме того, Европатент N 231624 раскрывает модифицированный АПТ, обладающий пролонгированным полупериодом существования. Модифицированный АПТ, имеющий F-EGFK2-A - последовательность, лишен домена "крингл 1", однако какой-бы то ни было конкретный способ его получения не показан. В свете цитированного выше понятно, что модифицированный АПТ в соответствии с изобретением должен отличаться от встречающегося в природе АПТ аминокислотной последовательностью в области внутренних доменов.
В результате обширных исследований заявитель получил усовершенствованный АПТ, который содержит фингер-домен, ФРЭ-домен, крингл 2-домен и домен серин-протеазы, но при этом первый "крингл 1" - домен делетирован в специфическом сайте, а в сайт, связывающий домены "крингла 2" и серин-протеазы введена мутация, в результате чего получен усовершенствованный АПТ, проявляющий превосходную устойчивость против тепла и кислот, имеющий заметно пролонгированный биологический полупериод существования и выраженную противовоспалительную активность и при этом сохраняющий желательные свойства встречающегося в природе человеческого АПТ.
Изобретение относится к усовершенствованному АПТ. АПТ в соответствии с изобретением заметно отличается по своей химической структуре от встречающегося в природе человеческого АПТ и проявляет лучшие свойства.
Усовершенствованный АПТ в соответствии с изобретением представляет собой полипептид, имеющий аминокислотную последовательность, представленную общей формулой, представленной на фиг.28-29, где R является прямой связью, Y обозначает A-Ile-B (A обозначает Arg или Glu и B обозначает lys или Ile), предпочтительно Glu-Jle-Lys. H2N обозначает аминоконец и -COOH обозначает карбоксиконец).
В изобретении термин "усовершенствованный АПТ" используют для обозначения аналога АПТ, в котором A и B обозначают описанные ниже аминокислоты соответственно:
усовершенствованный АПТ (II): Arg, Lys;
усовершенствованный АПТ (V): Arg, Ile;
усовершенствованный АПТ (VI): Glu, Lys;
усовершенствованный АПТ (VIII): Glu, Ile.
Изобретение также направлено на экспрессию предлагаемого аналога АПТ с использованием методик рекомбинантной ДНК. С этим связаны новые ДНК, кодирующие усовершенствованный АПТ, и векторы экспрессии рекомбинантной ДНК.
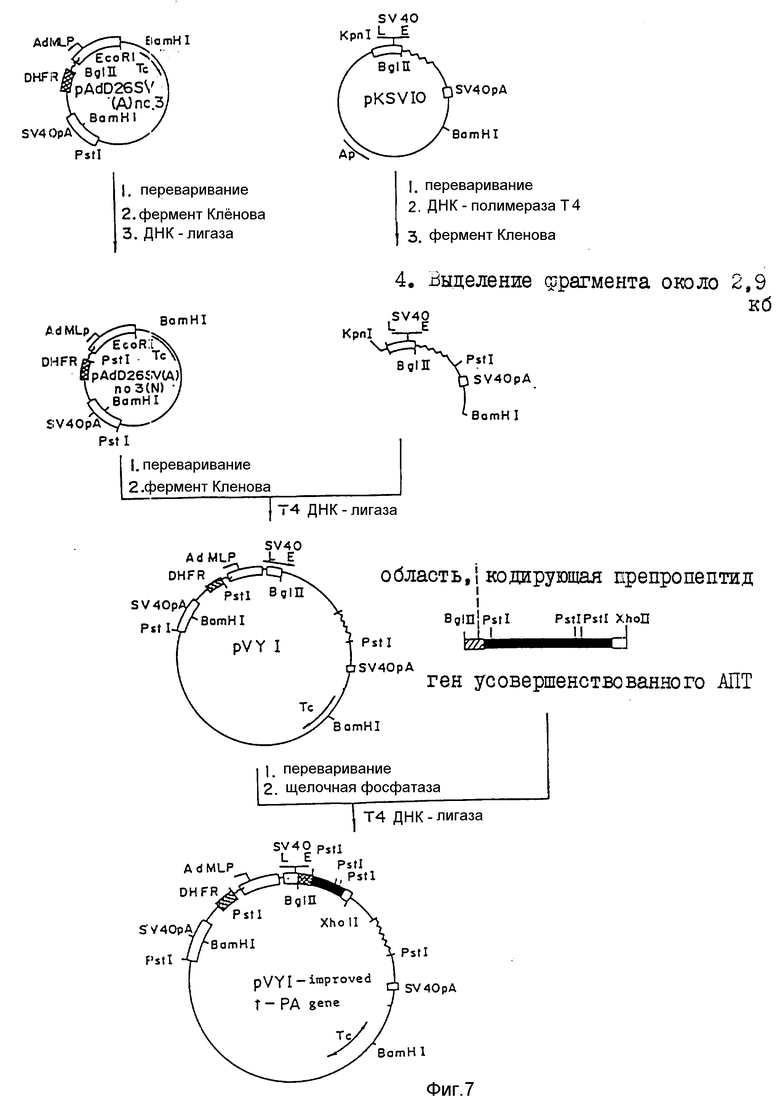
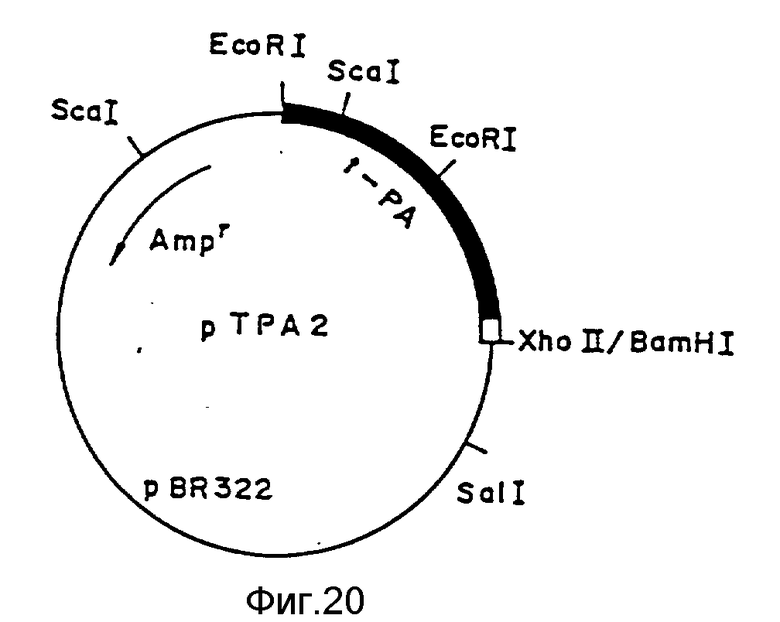
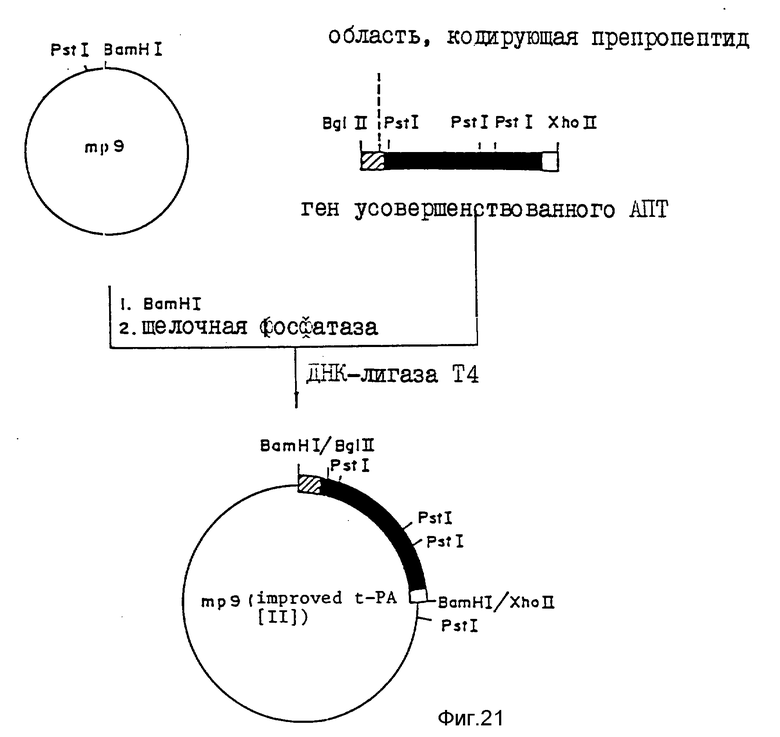
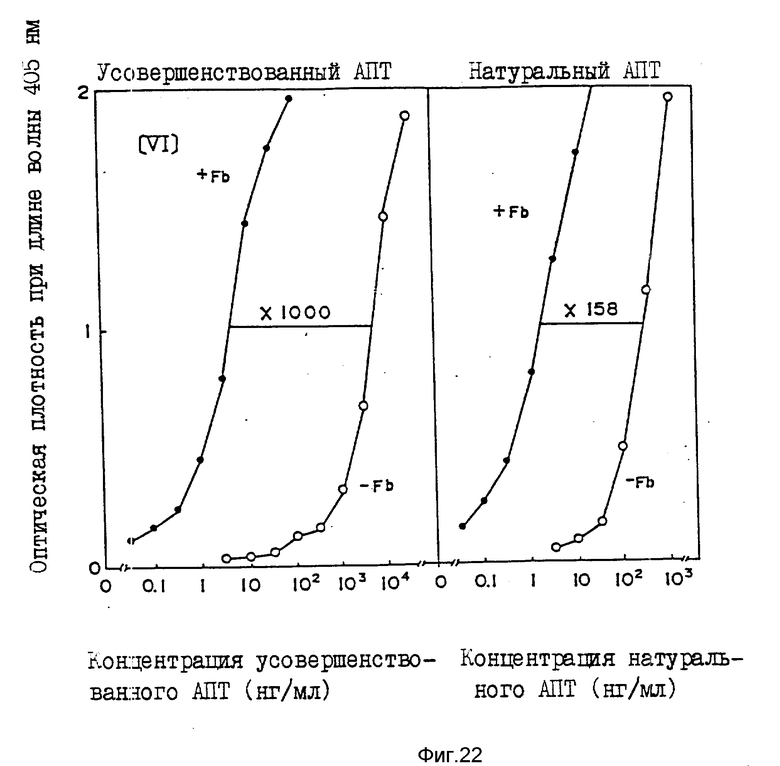
На фиг.1, 2 показана последовательность 16 олигодезоксинуклеотидов, используемая для конструирования фрагмента синтетического гена, кодирующего усовершенствованный АПТ (II); на фиг.3 - 4 - фрагмент синтетического гена для конструирования усовершенствованного АПТ (II) изобретения, содержащего концы рестрикции ферментами Bge 11 и Eco R1, который конструируют с использованием 16 олигодезоксинуклеотидов, показанных на фиг.1 - 2; на фиг.5 - методика конструирования усовершенствованного АПТ (II) (на рисунке черный участок, заштрихованный участок и незакрашенный участок обозначают область, кодирующую соответственно зрелый белок АПТ, область, кодирующую пропропептид и нетранслируемую область; на фиг.6 - метод проверки фрагмента синтетического гена блока IV путем определения последовательности оснований ДНК дидезоксиметодом и методом 7-DEAZA; на фиг.7 - методика построения вектора экспрессии pVY1 в животных клетках и интеграцию ДНК усовершенствованного АПТ в pVY1; на фиг.8 - 13 последовательности ДНК, кодирующие усовершенствованный АПТ (II) и усовершенствованный АПТ (V); на фиг.14 - 19 - аминокислотные последовательности, происходящие из последовательностей ДНК, кодирующих усовершенствованный АПТ (II) и усовершенствованный АПТ (V); на фиг.20 - рестрикционные ферменты и функциональная карта плазмиды pTPA 2, имеющей фрагмент Eco R1-Xho (около 1000 пар оснований) природного гена АПТ, интегрированный в вектор pBR322 по сайтам расщепления Eco R1 и Bam H1; на фиг.21 - mp9 (усовершенствованного АПТ (II), имеющую фрагмент BgL11-Xho 11 (около 1500 пар оснований) гена, усовершенствованного АПТ (II) интегрированный в двуцепочечную ДНК M13 mp9 в сайте расщепления BamH1; на фиг.22 - зависимость "доза-эффект" для активности АПТ усовершенствованного АПТ (VI) и встречающегося в природе АПТ методом S-2251 в присутствии (+Fb) и отсутствие (-Fb) заместителя фибрина; на фиг.23 - изменение активности усовершенствованного АПТ (VI) и нативного АПТ в крови кролика с течением времени; на фиг.24 - изменение остаточной активности усовершенствованного АПТ (VI) после термообработки; на фиг. 25 - ингибирование усовершенствованным АПТ (VI) фактора, активирующего лимфоциты (LAF); на фиг.26 - активирование с помощью денатурированного белка, усовершенствованного АПТ (VI); на фиг.27 - деградация денатурированного белка под действием усовершенствованного АПТ (VI).
Ниже подробно излагается способ получения рекомбинатных ДНК и трансформированных клеток.
Способ получения усовершенствованного АПТ.
Ген, кодирующий природный АПТ, на основе которого получают АПТ настоящего изобретения, выделяют из банка кДНК, изготовленного из клеток человеческой меланомы Bowes. Поли A+РНК выделяют из клеток человеческой меланомы Bowes и фракционируют центрифугированием в градиенте плотности сахарозы. Затем отбирают небольшое количество фракционированной поли (A)+РНК и фракцию мРНК, кодирующую ген АПТ идентифицируют методом дотгибридизации с использованием олигонуклеотидного зонда, способного распознавать специфическую последовательность мРНК АПТ. С использованием в качестве исходного вещества этой фракции, богатой мРНК АПТ, получают банк кДНК и подвергают скринингу при помощи зонда для идентификации мРНК АПТ, описанного выше. Поскольку не выделено ни одного клона, имеющего полную последовательность гена АПТ, недостающую последовательность оснований синтезируют ДНК-синтезатором с получением нужного гена. Затем желаемый ген конструируют методом индукции сайт-специфической мутации.
Фрагмент Eco R1-Xho 11 встречается в природе гена АПТ (около 1000 пар оснований), часть которого делетирована у N = окончания, введен в вектор pBR332 в сайтах расщепления Eco R1 и BamH1, при этом получена pTPA2. Штамм (E.coli HB 101/pTPA2), полученный трансформацией E.coli этой плазмидой, депонирован в институте ферментационных исследований Агенства по промышленной науке и технике Японии под регистрационным номером P-9649 (FERM BP-2107). Рестрикционная и функциональная карта плазмиды pTPA2 приведены на фиг.20.
Ген усовершенствованного АПТ встраивают в плазмиду pVY1.
Плазмиду pVY1 получают лигированием фрагмента BamH1-Kpn1 (около 2900 пар оснований) плазмиды pRSV10 (изготовленной фармация Файн Кемикалэ) с фрагментом от расщепления Eco R1 плазмиды pAdD26SV (A) N 3 (N) (полученной от доктора Хироши Ханда из Токийского университета (после получения у обоих тупых концов. Соответственно, данный вектор содержит кДНК гена дигидрофолат-редуктазы мыши под транскрипционным контролем основного позднего промотора аденовируса (Ad2), ранний промотор SV 40 вверх от сайта инсерции гена усовершенствованного АПТ и интрона и последовательность полиаденилирования, расположенные ниже гена. Ген настоящего изобретения может быть встроен и в другой подходящий вектор экспрессии. Вектор экспрессии интродуцируют далее в пригодную клетку хозяина с получением трансформантов. В качестве клеток хозяина могут быть использованы прокариотические клетки, также как E.coli, Bacillus subtilis и т.д., эукариотические микроорганизмы, такие как дрожжи, и т.д., а также клетки высших животных. В качестве представителя E.coli обычно используются штамм JM109, штамм W3110, Q и т.д., принадлежащие к штамму K12, в качестве представителя Bacillus subtilis используют штамм BD170, штамм BR151 и т.д. Из дрожжей можно использовать штамма RH218, штамм SHY1 и т.д. дрожжей Saccharomyces cerevisiae.
Для экспрессии обычно используют плазмидный вектор или фаговый вектор, содержащий репликон, происходящий от видов, совместимых с клетками хозяина, и регуляторную последовательность. Примерами вектора для E.coli являются, например, плазмиды pBR322, pUC18, pUC19 и т.д., λ - фаг, например λqt , Charon 4A и т.д., фаг M13 и др. В качестве вектора для Bacillus subtilis можно использовать pUB110, pSA2100 и т.д., а в качестве вектора для дрожжей можно использовать YRp7, YEp61 и т.д.
Вектор должен нести промотор, способный экспрессировать искомый белок. В качестве промотора для гена E.coli или фагового гена можно использовать, например, Lae, trp, tac, trc, pL и т.д.
В качестве хозяина можно использовать культивируемые клетки животных, такие как клетки почки макак резус, клетки личинок комара, клетки почки африканской зеленой мартышки, мышиный фетальный фибробласт, клетки яичника китайского хомячка, человеческие фетальные клетки почки, клетки ткани яйца бабочки, человеческие цервикальные эпителий-подобные клетки, человечески клетки миеломы, мышиные фибробласты и так далее. Как вектор можно использовать ранний промотор SV40, поздний промотор SV40, SV40, несущий промотор от эукариотного гена (например, эстроген-индуцируемый ген птичьего овальбумина, ген интерферона, глюкокортикоид-индуцируемый ген тирозин-аминотрансферазы, ген тимидин-киназы, ранний и поздний гены аденовируса, ген фосфоглицерат-киназы, ген α -фактора и т.д.), вирус коровьей папиломы или производные от них векторы.
Кроме того, известно, что АПТ, секретируемые и продуцируемые клетками, имеют различные N-окончания в зависимости от различия в сайтах расщепления.
В случае секретирования и продуцирования АПТ с использованием культуральных клеток в качестве хозяина способ расщепления сигнальной пептидазой или протеазой варьируется в зависимости от вида клеток, так что можно получить и виды АПТ, имеющие различные N-окончания.
Это явление подходит не только для случая секреции и производства при помощи культуральных клеток, так как считают, что аналогичное явление также может возникнуть при получении АПТ посредством E.coli, Bacillus sublitis, дрожжей и других клеток, подвергнутых специальной модификации.
Для трансформации хозяина с использованием вектора экспрессии с интегрированным в него геном усовершенствованного АПТ в случае использования E.coli можно применять метод Hanahan, Hanahan, D.J.Mol. Biol., 166, 557, 1983), в случае манипулирования с животными клетками можно использовать кальцийфосфатный метод (Vander Eb, A.J. and Graham, F.L., Method in Enrymoloqy, 65, 826, 1980, Academic Press) и так далее.
Как описано выше, усовершенствованный АПТ является пригодным для лечения различных приобретенных заболеваний, включая васкулярную коагуляцию (даже глубокой вены), эмболию легочной артерии, периферический артериальный тромбоз, эмболию в результате поражения сердца или периферической артерии, острый инфаркт миокарда и тромботический приступ.
Как и встречающийся в природе человеческий АПТ, усовершенствованный АПТ особенно пригоден для лечения острого инфаркта миокарда. Как недавно доказано, встречающийся в природе человеческий АПТ эффективен для растворения закупоривающего венечную артерию тромба, регенерации миокардиальной перфузии и восстановления большинства частей в ишемическом миокардиальном слое при введении внутривенно с дозировкой от 30 до 70 мг в течение 1-3 часов. Усовершенствованный АПТ отличается пролонгированным биологическим полупериодом существования в крови и поэтому эффективен в тех же случаях, что и встречающийся в природе человеческий АПТ. Ожидается, что усовершенствованный АПТ может дать клинический эффект, подобный природному человеческому АПТ, при дозе около 10% от дозы, которая рекомендуется при использовании встречающегося в природе человеческого АПТ, даже при однократном введении.
Кроме того, усовершенствованный АПТ настоящего изобретения проявляют следующие ценные свойства, которые до сих пор не были известны в отношении нативного человеческого АПТ и модифицированных АПТ.
а) Противовоспалительная активность.
На участке тромба выявляется не только образование самого тромба, но также и образование продуктов деградации фибрина или следовых количеств кинина. Известно, что эти вещества имеют индуцирующую воспаление активность и вызывают, таким образом, воспаление в области тромба. По этой причине желательно, чтобы средство, используемое для лечения тромбоза, обладало не только тромболитической активностью, но также и противовоспалительной активностью.
В результате проведенных исследований заявителю удалось придать усовершенствованному АПТ противовоспалительную активность на основе двух функций.
Одна из них состоит в том, что усовершенствованный АПТ ингибирует биологическую активность интерлейкина 1 (ИЛ-1), который является одним из медиаторов воспалительной реакции. ИЛ-1, продуцируемый макрофагом, как считают, принимает участие в воспалительной реакции посредством гипертермии, ускорения роста фибробласта, производства коллагеназы в синовиальной клеточной мембране и так далее, или за счет ускорения синтеза простациклина в васкулярных эндотелиальных клетках. Также известно, что ИЛ-1 воздействует на клетки печени, ускоряя производство белков (сывороточного амилоидного белка, фибриногена и т.д.) в острой фазе, которая возрастает при воспалении. Заявитель установил, что усовершенствованный АПТ ингибирует активность (LAF-активность) по повышению митогенной реакционной способности мышиного тимоцита, которая является одной из биологических активностей ИЛ-1.
Другая функция заключается в том, что усовершенствованный АПТ имеет сродство к денатурированному белку (денатурированному иммуноглобулину G, денатурированному альбумину и так далее), возникающему в результате воспаления в участке тромба, и дополнительно обладает свойством активироваться под действием этого денатурированного белка.
Благодаря этой активности усовершенствованный АПТ разлагает только денатурированный белок в области воспаления, и воспаление может временно ослабиться. Заявитель подтвердил посредством гель-электрофореза в додецил-сульфате натрия, что усовершенствованный АПТ разлагает только денатурированный белок.
Как показано на фиг. 26, активация и селективность усовершенствованного АПТ под действием денатурированного белка являются очевидными. С иммуноглобулином G, обработанным HCl, причем при в несколько раз меньшей концентрации, показана та же активность, что и с фибриногеном, обработанным BrCN. С другой стороны, нормальный иммуноглобулин C не проявляет активирующего действия по отношению к усовершенствованному АПТ даже при концентрации, равной 500 мкг/мл.
Предотвращение повторной окклюзии после восстановления перфузии закупоренного кровеносного сосуда.
Известно, что при лечении тромбоза натуральным АПТ отмечается повторная окклюзия с высокой частотой после восстановления кровотока закупоренного кровеносного сосуда. По этой причине осуществляют комбинированную терапию с ингибитором коагуляции тромбоцитов или антикоагулянтом. Однако комбинированная терапия заключает в себе проблемы взаимодействия лекарственных препаратов, контроля дозировок, подобных эффектов и так далее. Предпочтительно, чтобы сам АПТ дополнительно обладал активностью предотвращения повторной окклюзии.
Усовершенствованный АПТ настоящего изобретения обладает способностью предотвращать случаи повторной окклюзии за счет двух типов активности.
Первый тип представляет собой предотвращение быстрого снижения концентрации АПТ после введения усовершенствованного АПТ благодаря пролонгированной длительности действия, что приводит к устранению симптома Стюарта-Холмса и тем самым препятствует случаям возникновения повторной окклюзии.
Второй тип заключается в том, что благодаря предотвращению повреждения васкулярных эндотелиальных клеток, вызванного ИЛ-1, опосредованно ингибируется коагуляция тромбоцитов, что препятствует случаям возникновения повторной окклюзии.
с) Повышенная устойчивость.
Белковые препараты, как правило, неустойчивы, поэтому желательно хранить препараты в замороженном сухом состоянии или при низких температурах в виде раствора. При введении активатора плазминогена пациенту с острым инфарктом миокарда существует необходимость осуществлять процедуру в течение нескольких часов после начала приступа с тем, чтобы снизить коэффициент смертности. В таком случае желательны устойчивые препараты, которые можно хранить при комнатной температуре.
Кроме того, повышенная устойчивость позволяет осуществлять термообработку, обработку кислотами и т.п. во время приготовления препаратов. В частности, в отношении усовершенствованного АПТ настоящего изобретения, который продуцируют культуры клеток, становится возможным удалять ретровирус клеточного происхождения, который, как известно, является нестойким к воздействию тепла.
Ниже изобретение описано более конкретно со ссылкой на примеры, однако оно ими не ограничивается. Если не указано что-то иное, рекомбинантную ДНК продуцируют в соответствии с лабораторным руководством.
Маниатис Т и др., Молекулярное клонирование: лабораторное руководство, Коулд Спринг Харбор Лаборатори, Коулд Спринг Харбор, Нью-Йорк (1982).
Пример 1. Клонирование к ДНК АПТ.
Клетки человеческой меланомы Bowes (приобретены у доктора Роблина, Р. в Национальном институте по вопросам исследования рака, США) культивируют в соответствии со способом Opdenakker et al. (Opdenakker, G., et al., Eur. J. Biochem, 131, 481-487 (1983)). С целью индукции мРНК АПТ в культуральную смесь прибавляют ТФА (12-О-тетрадеканоилфорбол-13-ацетат) при конечной концентрации 100 нг/мл с последующим культивированным в течение 16 часов. Затем полную клеточную РНК экстрагируют из культивируемых клеток в соответствии с модифицированным методом Freeman et al. ((Okayama)Berqa DNA Manual, стр. 3, 1985, Фармация Файн Кемикалз). С использованием колонки с олиго-dT целлюлозой (изготовленной Фармация Файн Кемикалз) поли (A)+ РНК отделяют от всей клеточной РНК. В результате из числа приблизительно 10o клеток получают около 400 мкг поли(A)+ РНК.
Эту поли(A)+ РНК фракционируют центрифугированием в градиенте плотности сахарозы традиционным способом. Отбирают часть фракционированной поли(A)+ РНК, и проводят дот-блотгибридизацию (Perbal, B., Apractical Gube to Molecular Cloninq, 410, 1984, John Wiley and Sons, Inc) с использованием олигонуклеотидного зонда, специфического к мРНК АПТ. Зонд (зонд Y), используемый в данном случае, имеет последовательность оснований 5'-GCNNGGCAAAGATGGCA-3', которая комплементарна области мРНК, кодирующий аминокислотные остатки от +291 до +297 в последовательности АПТ, описанной Pennicaetal, и синтезируют β -цианофосфамидатным методом, используя ДНК-синтезатор, модель 380А, (изготовленный фирмой Applied Biosystems). Синтез ДНК-олигомера, отщепление защитной группы, отщепление от смолы и очистку осуществляют в соответствии с руководством по эксплуатации ДНК-синтезатора, Модель 380А. Мечение радиоактивным изотопом зонда Y на 5'-конце осуществляют в соответствии с лабораторным руководством, используя Т4-полинуклеотид-киназу (изготовленную Така-Ра Шузо Ко, Лтд) т и γ-(32P) АТФ.
Зонд Y сильно гибридизуется, главным образом, с 20-30S поли(A)+ РНК (эту фракцию называют фракцией M).
Используя матрицу, получают 10 мкг поли(A)+ РНК из фракции M; 3 мкг двуцепочечной кДНК синтезируют с использованием обратной транскриптазы (изготовленной Биокемикал Индастри Ко., Лтд) в соответствии с методом Gubler-Hoffman (Gubler, U. and Hoffman, B.J., Gene 25, 263, 1983), и прибавляют к двуцепочечной кДНК у 3'-окончания дезокси C-цепь в соответствии с методом Denq-Wu (Denq, G. R. and Wu, R., Nucleic Acids Res., 9, 4173, 1981). Затем двуцепочечную кДНК, удлиненную дезокси C-цепью подвергают гель-фильтрации на сефарозе CL 4B (изготовленной фармации Файн Кемикалз) с целью удаления низкомолекулярных нуклеиновых кислот, имеющих менее 500 пар оснований. После этого кДНК подвергают отжигу с помощью pBR322 (изготовленной Бетезда Рисерч), содержащей дезокси G-цепь в сайте Pst1, используя традиционную методику. Смесью, полученной после отжига, трансформируют компетентные клетки HB101 E.coli (изготовленные Такара Шузо Ко., Лтд). В результате получают банк кДНК, состоящий из приблизительно 4000 самостоятельных трансформантов.
Эту кДНК подвергают гибридизации колоний с использованием зонда Y, описанного выше, в соответствии с методом Woods (Woods, D., Focus, 6 (3), 1, 1984), изготовлен Бетесда Рисерч Лаб.), получая клоны, взаимодействующие с зондом Y. Среди клонов выявляют pTPA1 клон, содержащий наиболее длинную кДНК АПТ. Затем осуществляют дидезоксиметод (Carlson, J., et al., J. Biotechnoloqy, 1, 253, 1984), используя фаговый вектор M13 и метод 7-DEAZA (Mizusawa S. , et al., Nucleis Acids Res., 14, 1319, 1986). В результате установлено, что плазмида pTPA1 содержит последовательность оснований от Ty+441 до Ay+2544 для гена АПТ, описанного Pennicaetal.
Пример 2. Конструирование усовершенствованного АПТ (II).
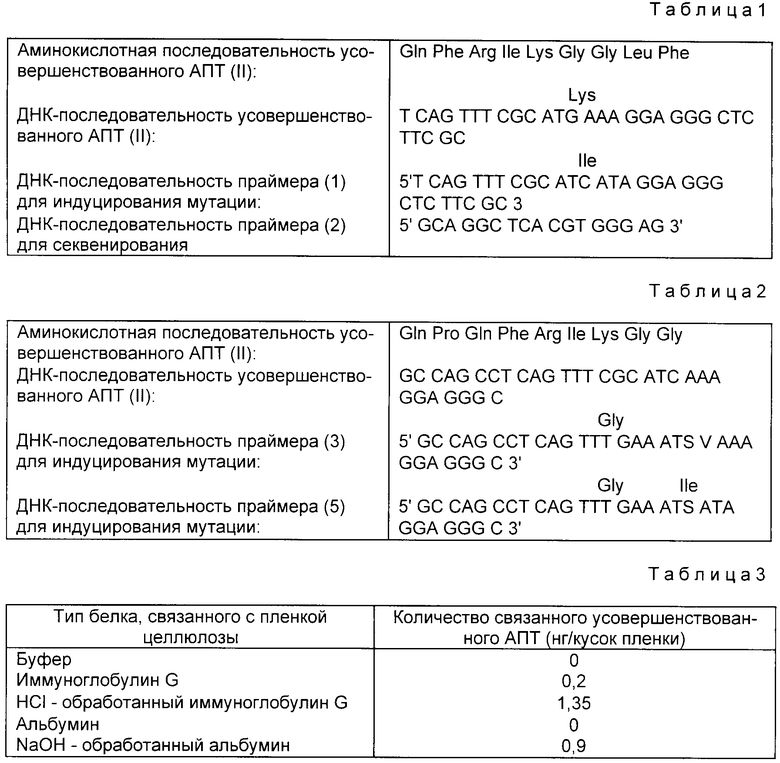
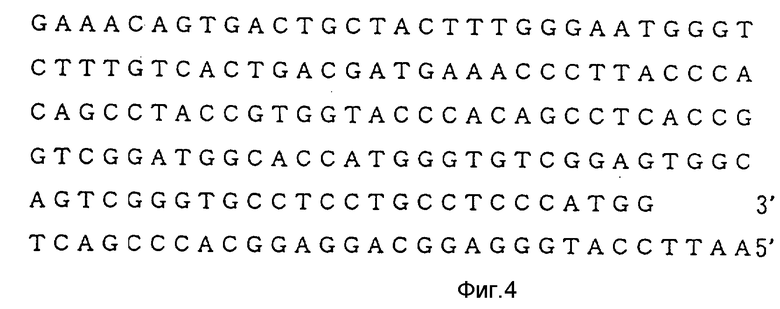
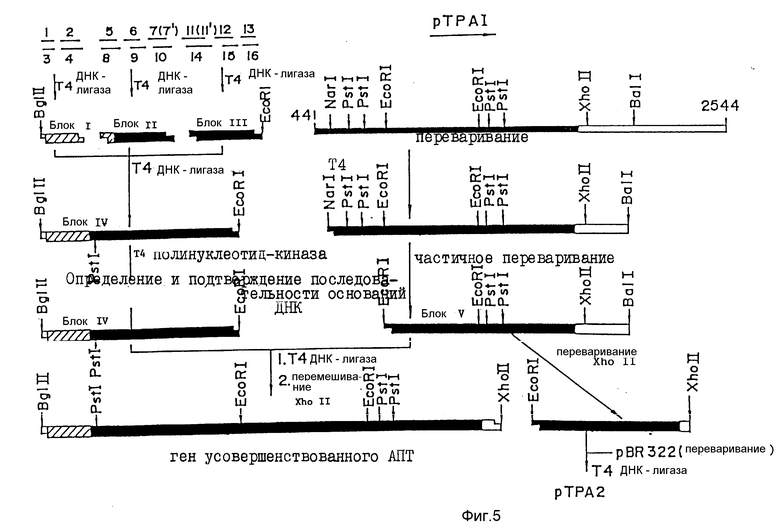
В плазмиде pTPA1, показанной в примере 1, N-концевая область является недостаточной для построения усовершенствованного АПТ (II), который лишен крингл 1-домена. Поэтому недостаточный ДНК-сегмент синтезируют, как описано выше, с использованием ДНК-синтезатора 380А (изготовленного Applied Biosystems). Последовательность оснований синтезированного олигомера и полная синтезированная последовательность показаны на фиг. 1-4. Специфические методики конструирования усовершенствованного АПТ (II) с использованием этих олигомеров приведены на фиг. 5-6.
2-1). Конструирование блока IV (фрагмент Bql II-Eco R1, около 480 пар оснований).
Фрагмент блока IV на фиг. 5 получают следующим образом.
Во-первых, в соответствии с лабораторным руководством, 40 пмоль каждого из синтетических олигонуклеотидов 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14 и 15, показанных на фиг. 1-2, фосфорилируют 10 единицами Т4-полинуклеотид-киназы (изготовленной Такара Шузо Ко., Лтд) при температуре 37oC в течение одного часа в 50 мкл реакционного раствора для каждого из них. Реакционный раствор обрабатывают фенолом. После преципитации этанолом осадки сушат при пониженном давлении и растворяют в стерильной дистиллированной воде. После отстаивания 40 пмоль каждого олигомера в 150 мкл раствора, содержащего 6 мМ Трис-HCl (pH 7,5), 20 мМ NaCl, 7 мМ MgCl2 и 0,1 мМ ЭДТА, при температуре 80oC в течение 5 минут, при температуре 60oC в течение 5 минут и при комнатной температуре в течение одного часа, в соответствующих блоках блока I (олигомеры 1, 2, 3 и 4), блока II (олигомеры 5, 6, 7, 8, 9 и 10) и блока III (олигомеры 11, 12, 13, 14, 15 и 16) осуществляют преципитацию этанолом и сушку при пониженном давлении. Остаток растворяют в 40 мкл стерильной дистиллированной воды. Реакцию осуществляют в 400 мкл реакционного раствора при температуре 4oC в течение 15 часов, используя набор для ДНК-лигирования (изготовленный такара Шузо Ко., Лтд). После преципитации этанолом и сушки при пониженном давлении осадок растворяют с терильной дистиллированной воде: в случае блока I (1) осуществляют гельэлектрофорез в 5%-ном полиакриламиде (лабораторное руководство), отделяют и очищают традиционным способом (лабораторное руководство), фрагмент около 100 пар оснований, а в случае блока II (2) и блока III (3) гель-электрофорез осуществляют в 3%-ном агарозном геле (агароза LMP, изготовлена BRL) (лабораторное руководство) и выделяют и очищают электроэлюцией (лабораторное руководство) фрагменты около 190 пар оснований.
Затем 0,1 мкг, 0,2 мкг и 0,2 мкг фрагментов блока I, блока II и блока III соответственно лигируют с использованием вышеуказанного набора для ДНК-лигирования. Осуществляют гель-электрофорез при концентрации агарозы 1,5 % с тем, чтобы выделить фрагмент Bgl II-Eco R1 (блок IV) размером около 480 пар оснований. Затем ДНК выделяют из агарозного геля с помощью электроэлюции. Эту ДНК затем фосфориллируют в 100 мкл реакционного раствора при температуре 37oC в течение одного часа с использованием 10 единиц вышеуказанной Т4-полинуклеотид-киназы, после чего обрабатывают фенолом, осаждают этанолом и сушат при пониженном давлении.
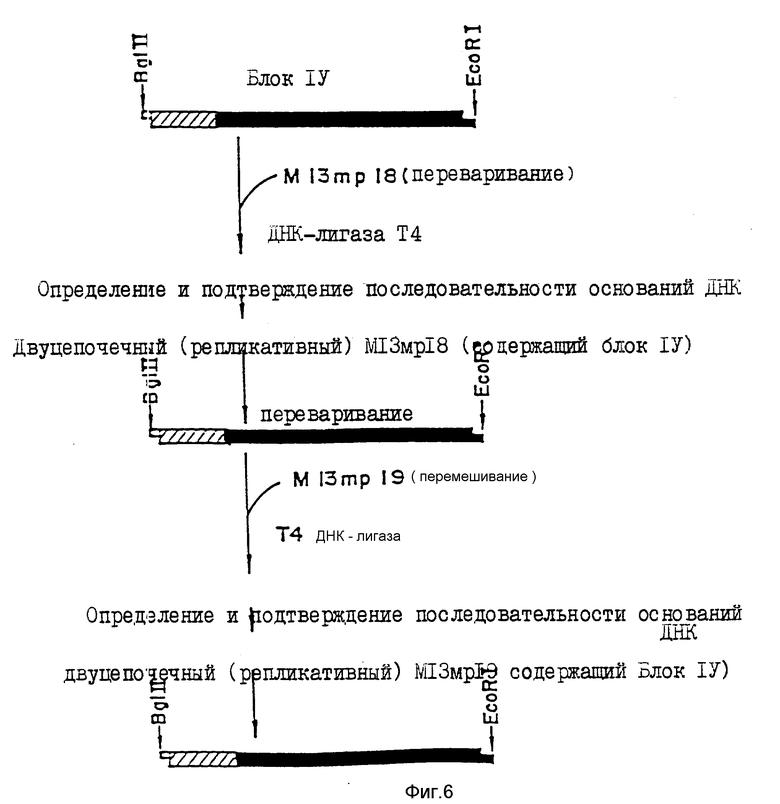
Этот фрагмент синтетического гена и последовательность оснований блока IV подтверждают, определяя последовательность оснований в соответствии с дидезоксиметодом, используя вектор фага М13. Специфические методики показаны на фиг. 6. После лигирования вышеописанного фрагмента Bgl II-Eco R1 блока IV с М13 mp18 ДНК (изготовлена Boehringer Mannheim-Yamanouchi Co., Ltd.), расщепленной рестрикционными ферментами BamH1 (изготовлен Boehringer Mannheim-Yamanouchi Co., Ltd.) и Eco R1 (изготовлен Boehringer Mannheim-Yamanouchi Co. , Ltd. ) определяют последовательность его оснований с использованием набора секвенирования M13 (изготовлен Тарака Шузо К., Лтд) и набора секвенирования 7 - DEAZA (изготовлен Такара Шузо Ко., Лтд).
Сайт расщепления рестрикционным ферментом Bgl11 и сайт расщепления рестрикционным ферментом BamH1 лигируют изошизомерным расположением через (BamH1 - расщепленный конец Bgl11-сайт расщепления), и лигированный фрагмент можно расщепить рестрикционным ферментом Xho 11, в результате чего появляются естественные концы расщепления Bgl 11 и Bamh1 соответственно.
Для более точного определения последовательности оснований фагом M 13mp18 (включающим фрагмент блока IV) заражают штамм E.cjli JM109 в соответствии с методом Messing/Messing J., Methods in Enzymology, 101, 20-78 (1983)), после чего получают двуцепочечную ДНК (репликативного типа). После расщепления данной ДНК (50 мкг) рестрикционными ферментами Xho 11 (изготовлен Boehringer Mannheim-Yamanouchi Co) и Eco R1 осуществляют гель-электрофорез в 1,5%-ном агарозном геле с выделением фрагмента (блок IV) около 480 пар оснований. Эту ДНК экстрагируют электроэлюцией. После лигирования экстрагированной ДНК с M13mp19 ДНК (изготовлена Boehringer Mannheim-Yamanouchi Co., Ltd) расщепленной рестрикционными ферментами Eco R1 и BamH1 аналогичным вышеописанному образом, используя набор для ДНК-лигирования, определяют последовательность оснований. Как описано выше данную последовательность можно проверить более точно, определяя последовательность обеих ДНК с использованием M13mP18 и M13mp19. Кроме того, двуцепочечную репликативную ДНК M13mp19 (с блоком IV) получают описанным методом. После расщепления этой ДНК (50 мкг) рестрикционными ферментами Eco R1 и Xho 11 осуществляют гель-электрофорез в 1,5%-ной агарозе, выделяя при этом фрагмент (блок IV) размером около 480 пар оснований.
2-2). Выделение блока V (фрагмент Eco R1-Bal1, около 1250 пар оснований).
Из клона pTRA1, полученного в примере 1, выделяют плазмидную ДНК в больших количествах в соответствии с методом, описанным в лабораторном руководстве, как показано на фиг. 5. После расщепления 70 мкг этой ДНК рестрикционными ферментами Bal1 (изготовлен Такара Шузо Ко. , Лтд) и Nar1 (изготовлен Нирро Ген Ко., Лтд) осуществляют электрофорез в 0,8%-ном агарозном геле, выделяя при этом фрагмент Nar1-Bal1 (около 1540 пар оснований). ДНК выделяют электроэлюированием. После дальнейшего частичного переваривания данной ДНК рестрикционным ферментом Eco R1 осуществляют электрофорез а 0,7%-ном агарозном геле, выделяя фрагмент Eco R1-Bal1 (около 1250 пар оснований). ДНК выделяют электроэлюированием.
2-3). Конструирование гена усовершенствованного АПТ (II) из блока IV и блока V.
Как показано на фиг. 5, ген усовершенствованного АПТ получает следующим образом.
После легирования блока IV (фрагмент Bgl11-Eco R1, около 480 пар оснований), полученного в примере 2-1, с блоком V (фрагмент Eco R1-Bal1, около 1250 пар оснований), полученным в примере 2-2, с использованием набора для ДНК-легирования, описанного выше, легированный продукт подвергают преципитации этанолом. После сушки при пониженном давлении преципитат расщепляют рестрикционным ферментом Xho 11 традиционным способом. Затем осуществляют электрофорез в 0,8% агарозном геле с тем, чтобы выделить фрагмент Bgl 11-Xho 11 (около 1500 пар оснований, содержит ген усовершенствованного АПТ). Затем электроэлюированием выделяют ДНК. Полная последовательность оснований полученного таким образом гена усовершенствованного АПТ (II), приведена на фиг. 8-13. Выведенная аминокислотная последовательность также приведена на фиг. 14-19.
Пример 3. Конструирование гена усовершенствованного АПТ V, VI и VIII.
Конструирование гена усовершенствованного АПТ V, VI или VIII, осуществляют на основе гена усовершенствованного АПТ (II) со ссылкой на следующие публикации.
Генетическую конверсию осуществляют методом индукции сайт-специфической мутации.
Публикации: Золлер М. Дж. и Смит.М., Метод в ферментологии, 100, стр. 468-500 (1983), Золлер М. Дж. и Смит. М. ДНК, 3, стр.479-488 (1984), Моринага Й. и др. Биотехнология, стр. 636-630 (июль 1984), Адельман Дж. П. и др. , ДНК, 2, стр. 183-193 (1983), 6. Руководство по секвенированию M13 (puC) издано Джин Сейенз Рум Ко., Лтд).
3-1). Конструирование гена усовершенствованного АПТ (V).
A) Создание M13mp19 (АПТ/П/) для мутирования.
Фрагмент гена усовершенствованного АПТ (II), описанный подробно в примере 2, 2-3), лигируют с двуцепочечной ДНК M13mp9, обработанной рестрикционным ферментом BamH1 и щелочной фосфатазой (изготовленной Такара Шузо Ко., Лтд.). Продуктом лигирования трансфецируют компетентные клетки E. cjli JM109 (изготовленные Такара Шузо Ко., Лтд). Каждый клон, дающий бесцветное стерильное пятно, используют для заражения E.Coli JM109. Одноцепочечную ДНК выделяют итз культурального супернатанта, а двуцепочечную (репликативную) ДНК выделяют из клеток E.cjli в соответствии с методом Мессинга (Мессинг Дж. , методы в ферментологии, 101, стр. 20-78, 1983). При анализе характера этих двухцепочечных ДНК после расщепления рестрикционным ферментом Pst1 с помощью электрофореза в агарозном геле получают клон mp9 (усовершенствованный АПТ (II), в котором ген АПТ (II) инсерцирован в ДНК mp9 в желательном направлении, как показано на фиг. 21.
После расщепления части этих ДНК рестриктазой Pst осуществляют электрофорез в 0,8%-ном агарозном геле, где клон mp9 (усовершенствованный АПТ (II) показывает простую полоску в положении 7300 пар оснований, 840 пар оснований, 430 пар оснований и 80 пар оснований, приблизительно. Одноцепочечную ДНК данного клона используют в последующем эксперименте на индуцирование сайт-специфической мутации.
B) Синтез праймера, способного индуцировать сайт-специфическую мутацию.
Синтетический олигонуклеотид, используемый для индуцирования сайт-специфической мутации в гене усовершенствованного АПТ (II), синтезирует β -цианоэтилфосфоамидатным методом с использованием ДНК-синтезатора, модель 380 A (изготовленного Applied Biosystems). Синтез ДНК-олигомера, удаление защитной группы, отщепление от смолы и очистку осуществляют в соответствии с инструкцией по эксплуатации ДНК-синтезатора 380 A. Для индуцирования мутации в специфическом сайте получают праймер (1), способный индуцировать сайт-специфическую мутацию и праймер (2) для секвенирования дидезоксиметодом с использованием вектора фага M13 (Карлсон Дж. и др., журнал биотехнологии, 1, стр. 253, 1984).
Приведены аминокислотная и нуклеотидная последовательности для усовершенствованного АПТ (II). Праймер (1), способный индуцировать мутацию, отличается подчеркнутым основанием от генной последовательности усовершенствованного АПТ (II) (см. табл.1).
C) Индуцирование сайт-специфической мутации.
Ниже приводится способ создания клона, содержащего последовательность оснований праймера (1), способного давать мутацию, а именно гена усовершенствованного АПТ (IV). После отжига (ренатурации) одноцепочечной ДНК, описанной в примере 3,3-1), A) клона mp9 (усовершенствованного АПТ (II)и праймера (1) продукт ренатурации превращают в двуцепочечную ДНК, которой затем трансформируют E. coli JM109. Затем, используя праймер для секвенирования, осуществляют скрининг ДНК-последовательностей, выделяя фаговый клон, несущий мутированный ген усовершенствованного АПТ (II), а именно ген усовершенствованного АПТ (V). Из этого клона извлекают двуцепочечную (репликативную) фаговую ДНК и выделяют ген усовершенствованного АПТ (V).
5'-Концевое фосфорилирование синтетического олигомера.
ДНК праймера (1) для индуцирования сайт-специфической мутации фосфорилируют методом, описанным в примере 2,2-1).
Получение гетеродуплексной ДНЕ.
0,5 мкг одноцепочечной ДНК M13mp9 (усовершенствованного АПТ (II)) и 1,5 мкг двуцепочечной ДНК M13mp9, расщепленной рестрикционным ферментом BamH1, нагревают в 30 мкг раствора, содержащего 2 пмоль фосфорилированного праймера (1) 10 мМ Трис-HCl (pH 7,5), 0,1 мМ ЭДТА и 50 мМ NaCl, при температуре 90oC (2 мин), 50oC (5 мин), 37oC (5 мин) и при комнатной температуре (10 мин). К раствору прибавляют 36 мкл раствора 50 мМ Трис-HCl (pH 8,0), содержащего 4 единицы фермента Кленова, 7 единиц ДНК-лигазы фага T4, 0,1 мМ ЭДТА, 12 мМ MgCl2, 10 мМ дитиотрейтола, 0,7 мМ АТФ, 0,07 дАТФ и 0,2 мМ каждого из дГТФ, дТТФ и дЦТФ, с тем, чтобы стимулировать элонгацию праймера. Смесь подвергают взаимодействию при температуре 20oC в течение 2 часов и при температуре 4oC в течение 15 часов.
Трансформацию осуществляют с использованием раствора, описанного выше, и компетентных клеток E. coli JM109 (изготовленных Такара Шузо Ко., Лтд) до образования пятен лизиса. После отделения бесцветного пятна фагом заражают E. coli JN109 для пролиферации. Затем матричную одноцепочечную ДНК получают из культурального супернатанта относительно каждого клона. Эти одноцепочечные ДНК подвергают только реакции "T" (реакция "A" и "T" в примере 3-2) дидезоксиметода, используя праймер (2) для секвенирования, с последующим электрофорезом в полиакриламидном геле. После сушки гель анализируют радиоавтографией. На основе результатов идентифицируют клон, имеющий искомую мутантную последовательность. Культуральный супернатант клона используют для заражения клеток E.coli JM109 и вновь инокулируют на чашку с тем, чтобы осуществить выделение единственного пятна. Из полученного единственного пятна одноцепочечную ДНК выделяют в соответствии с вышеприведенным способом. Используя эти ДНК, во-первых, определяют последовательность оснований ДНК дидезоксиметодом, используя праймер (2) для секвенирования, получая клон, мутированный в желательную последовательность оснований. После заражения этого фагового клона клетками JM-109 E. coli с использованием метода Мессинга, описанного в примере 2, получают двуцепочечную ДНК. Эту двуцепочечную ДНК расщепляют рестрикционным ферментом Xho 11, осуществляют электрофорез в 0,8%-ном агарозном геле с выделением фрагмента (гена усовершенствованного АПТ (V) размером около 1500 пар оснований, содержащего ген усовершенствованного АПТ. Затем электроэлюированием экстрагируют ДНК.
Кроме того, дидезоксиметодом определяют полную последовательность оснований в отношении полученной таким образом ДНК, в результате чего находят, что ДНК представляет собой ген усовершенствованного АПТ (V). Полная последовательность оснований полученного таким образом гена усовершенствованного АПТ (V) (однако содержащая сигнальный пептид от -35 до -1) показана на фиг. 11 - 13. Выведенная из нее аминокислотная последовательность показана также на фиг. 17 - 19.
3-2) Конструирование усовершенствованных АПТ (VI) и (VIII).
Методики аналогичны тем, которые описаны в примере 3, 3-1). Во-первых, конструируют M13mp3 (усовершенствованный АПТ (II)), после чего синтезируют праймеры для индуцирования сайт-специфической мутации. Последовательность оснований этих праймеров описана выше, однако для конструирования гена усовершенствованного АПТ (VI) и гена усовершенствованного АПТ (VIII) используют соответственно фосфорилированный с 5'-конца праймер (3) и фосфорилированный с 5'-конца праймер (5) (см. табл. 2). Вслед за индуцированием сайт-специфической мутации определяют дидезоксиметодом полную последовательность оснований. Подтверждено, что они имеют желательные последовательности оснований. Таким образом, получают гены усовершенствованного АПТ (VI) и усовершенствованного АПТ (VIII).
Затем эти гены интегрируют в вектор pVY1 в соответствии с методикой, описанной в примерах 4 и 5.
Пример 4. Интеграция гена усовершенствованного АПТ (II) в вектор pVY1.
4-1) Конструирование вектора pVY1.
Вектор pVY1 получают как показано на фиг. 7.
A) Конструирование pAdD26SV (A) N3 (N) и придание тупых концов сайту расщепления Eco R1.
Во-первых, ДНК pAdD26SV(A) N3 (приобретена у доктора Хироши Ханда в Токийском университете, известна по тезисам в Mo1, Ce 11. Biol, 2 (11, (1982)) расцепляют рестрикционным ферментом Bgl11 (изготовлен Boehringer Mannheim-Yamanouchi Co. , Ltd.) традиционным способом. Затем ДНК делают тупоконечной традиционным способом с использованием фермента Кленова (изготовлен Boehringer Mannheim-Yamanouchi Co., Ltd). После обработки фенолом, преципитации этанолом и сушки при пониженном давлении преципитеты растворяют в стерильной дистиллированной воде. После дальнейшего лигирования трансформируют реакционной смесью компетентные клетки HB101 E.coli (изготовлены Такра Шузо, Ко. Лтд). Плазмидные ДНК получают из трансформантов, проявляющих устойчивость к тетрациклину, обычным образом.
После расщепления части этих ДНК рестрикционным ферментом BgL 1 осуществляют электрофорез в 0,7%-ной агарозе. В результате получают клон, несущий ДНК, которая не расщеплена рестрикционным ферментом BgL 11. После переваривания (pAdD26SV(A) N3 (N)) ДНК этого клона рестрикционным ферментом Eco R1 традиционным способом, ДНК делают тупоконечной с использованием фермента Кленова, как описано выше. После обработки фенолом, преципитации этанолом и сушки при пониженном давлении преципитаты растворяют в дистиллированной стерильной воде.
B) Выделение фрагмента Kpn 1-BamH1 (около 2900 пар оснований) из pKSV10 и формирование тупых концов.
После расщепления ДНК pKSV10 (изготовлена фармация Файн Кемикалз) рестрикционными ферментами Kpn1 и BamH1 традиционным способом ДНК делают тупоконечной с использованием ДНК-полимеразы T4 (лабораторное руководство, стр. 114 - 121). Затем осуществляют электрофорез в геле 0,7%-ной агарозы с выделением фрагмента размером около 2900 пар оснований. Затем фрагмент подвергают электроилюированию для экстракции ДНК
C) Конструирование pVY1.
После лигирования ДНК-фрагмента, полученного в A), и ДНК-фрагмента, полученного в B), осуществляют трансформацию компетентных клеток HB101 E. coli (описаны выше).
Из трансформантов, проявляющих устойчивость к тетрациклину, традиционным способом получают плазмидные ДНК. После расщепления части этих плазмидных ДНК рестрикционным ферментом Pst1 (изготовлен Boehringer Mannheim-Yamanouchi Co. , Ltd) осуществляют электрофорез в 1,0%-ном геле агарозы. В результате получают клон (плазмиду pVY1), характеризующийся полосами около 3400 пар оснований, около 3200 пар оснований и около 1400 пар оснований. Этот клон E/coli HB101 (pVY1 депонирован в научном Институте исследований ферментации Агентства по промышленной науке и технике Японии под регистрационным номером P-9625 (FEPM BP 2106).
4-2) Интеграция гена усовершенствованного АПТ (II) в вектор pVY1.
После расщепления ДНК плазмиды pVY1, полученной в примере 4-1), рестрикционным ферментом BgL 11 традиционным способом, осуществляют дефосфорилирование с использованием щелочной фосфатазы (изготовлена Такара Шузо. Ко. Лтд). Затем осуществляют обработку фенолом три раза. А после преципитации этанолом и сушки при пониженном давлении осадки растворяют в стерильной дистиллированной воде.
После лигирования этой ДНК с фрагментом BgL 11-Xho 11 (около 1500 пар оснований), полученным в примерах 3, 3-1), и компетентные клетки HB101 E.coli трансформируют продуктом лигирования в соответствии с методом, описанным выше. Из транспормантов, имеющих устойчивость к тетрациклину, получают традиционным образом плазмидные ДНК. После расщепления этих ДНК рестрикционными ферментами (BqL 11, Pst 1) выбирают клон, имеющий ген усовершенствованного АПТ (II) в векторе pVY1, интегрированный в требуемом направлении, причем отбор осуществляют на основе анализа картины электрофореза в геле агарозы. Во-первых, часть этих ДНК расщепляют рестрикционным ферментом BqL 11, после чего осуществляют электрофорез в 0,8%-ном геле агарозы, получая клон, имеющий полосу фрагмента размером около 1500 пар оснований, когда фрагмент BqL 11-Xho 11 лигируют с фрагментом BqL 11 плазмиды pVY1, лигированную часть Xho 11 и BqL 11 можно отрезать рестрикционным ферментом BqL 11.
Часть плазмидных ДНК этих клонов дополнительно расщепляют рестрикционным ферментом Pst1, и ДНК подвергают электрофорезу в 0,8%-ном геле агарозы с получением клона, имеющего одну полосу размером около 3400 пар оснований, две полосы размером около 2300 пар оснований, одну полосу размером около 1400 пар оснований и одну полосу размером около 80 пар оснований. С использованием этого клона (плазмиды pVY1-АПТ (II) в соответствии с лабораторным руководством получают плазмидные ДНК.
Пример 5. Интеграция генов усовершенствованных АПТ (V), (VI) и (VIII) в вектор pVY1.
После расщепления ДНК плазмиды pVY1, полученной в примере 4-1), рестрикционным ферментом BqL 11 традиционным образом осуществляют дефосфорилирование с использованием щелочной фосфатазы (изготовленной Такара Шузо, Ко., Лтд) с последующими обработкой (3 раза) фенолом, преципитацией этанолам и сушкой при пониженном давлении. Затем осадок растворяют в стерильной дистиллированной воде.
После лигирования данной ДНК с фрагментом BqLII-Xho 11 размером около 1500 пар оснований, полученным в примерах 2, 2-3), продуктом лигирования трансформируют вышеуказанные компетентные клетки HB101 E.coli. Плазмидные ДНК получают из трансформантов, проявляющих устойчивость к тетрациклину, в соответствии с традиционным способом. После расщепления этих ДНК рестрикционными ферментами BqL11 и Pstl осуществляют электрофорез в геле агарозы. Посредством анализа характера разделения в геле агарозы отбирают клоны, в которых ген усовершенствованного АПТ (V) встроен в вектор pVYI в требуемом направлении. Во-первых, после расщепления части этих ДНК рестрикционным ферментом BqL11 осуществляют электрофорез в 0,8%-ном геле агарозы с получением клонов и получают полосу размером около 1500 пар оснований. Когда фрагмент BqL11-Xholl связан с фрагментом BqL11 вектора pVYI, часть Xholl и BqL11 можно отщепить рестрикционным ферментом BqL11 благодаря вышеупомянутой конфигурации изошизомера.
После дальнейшего расщепления части плазменных ДНК этих клонов рестрикционным ферментом Pstl осуществляют электрофорез при концентрации геля агарозы, равной 0,8%, с получением клона, дающего полосу размером около 3400 пар оснований, полосу размером около 2300 пар оснований, две полосы размером около 1400 пар оснований, одну полосу размером около 800 пар оснований и одну полосу размером около 80 пар оснований. С использованием клона (плазмиды pVYI-АПТ (V)) плазмидную ДНК получают в больших количествах, основываясь на лабораторном руководстве.
Аналогичным образом в вектор pVYI интегрируют гены усовершенствованных АПТ (VI) и (VIII).
Пример 6. Экспрессия усовершенствованного АПТ в клетках CHO.
Плазмидой pVYI - усовершенствованный АПТ (VI), АПТ (II), АПТ (V) или АПТ (VIII) трансфецируют ДГФР-дефицитные клетки CHO (Urlaub, et al., Proc.Natl., Acad. Sci. USA, 77(7), 4216-4224, 1980) кальцийфосфатным методом (Graham, et al., Viroloqy, 52, 456, 1973). Обнаружено, что трансформатный клон, полученный на селективной среде (MEM A LPHA (-), GIBCO) в присутствии метотрексата (MTX), проявляет активность АПТ на уровне от 50 до 100 единиц/мл (значение, определенное фибрин/агарозным чашечным методом, описанным ниже). Этот клон используют для последующих исследований. В качестве среды для продуцирования используют среду GIT (изготовленную Уако Пьюэ Кемикал Индастри Ко., Лтд), обогащенную 20 международными единицами/мл (SIGMA) апротинина.
Пример 7. Очистка усовершенствованного АПТ от культурального супернатанта клеток CHO.
Культуральный супернатант, полученный в примере 6, частично очищают на аффинной колонке с анти-АПТ моноклональным антителом. Гибридному, продуцирующую моноклональные антитела, получают для АПТ, имеющего происхождение из клеток меланомы человека, традиционным образом. Антителопродуцирующую гибридному инокулируют мышам, и моноклональное антитело (подкласс: IgGM1), развившееся в асците, экстрагируют и очищают с использованием Целлюлофина Белка A (изготовлен Биокемикал Индастри Ко., Лтд) и буферной системы MAPS для очистки моноклонального антитела, изготовленной Биорад Лабораториз. Антитело связывают с CN3r-активированной сефарозой (производства компании Фармация Файн Кемикалз) в отношении 4 мг на 1 мл геля традиционным способом.
Гель с антителом (24 мл) смешивают с четырьмя литрами культурального супернатанта. После осторожного встряхивания в течение ночи при температуре 4oC гель загружают в колонку (диаметр 1,5 см х 20 см). Затем гель последовательно промывают 125 мл каждого из следующих растворов (1) Трис-HCl буфера pH 7,4 (буфер A), содержащего 25 международных единиц/мл апротинина (производства SIGMA) и 0,01% (масса/объем) Твин 80, (2) буфера A, содержащего 0,5 М NaCl, (3) буфера A, содержащего 4 М мочевины, и (4) буфера A. Усовершенствованный АПТ, связанный с гелем, элюируют 0,2 М глицин-HCl буфером pH 2,5, содержащим 25 международных единиц/мл апротинина и 0,01% (масса/объем) Твин 80. Активные фракции восстанавливают и объединяют. После диализа против 10 мМ Трис-HCl буфера, pH 7,4, содержащего 25 международных единиц/мл апротинина и 0,01% (масса/объема) Твин 80, в течение ночи диализат концентрируют в 20-30 раз вакуумным центробежным концентратом (Speed VAC, производство фирмы SAVANT Инк). Концентрат вновь диализируют против 10 мМ Трис-HCl буфера, pH 7,4, содержащего 0,15 М NaCl, 25 международных единиц/мл апротинина и 0,01% (масса/объем) Твин 80, в течение ночи, и используют для последующей оценки in vitro и in vivo. Окончательно удельная активность повышается в 3700-5000 раз, а выход составляет от 36 до 42% активности АПТ (определено фибрин/агарозным чашечным методом).
Эту активную фракцию анализируют электрофорезом с додецилсульфатом натрия и окрашиванием серебром. В восстановительных условиях на уровне 54 килодальтон отмечается очень сильная полоса вместе с несколькими другими полосами. Далее гель после электрофореза обрабатывают 2,5% (масса/объем) Тритон X-100 и помещают на фибрин/агарозную чашку с целью осуществления автографии фибрина при температуре 37oC, благодаря чему растворенную полосу обнаруживают на уровне около 50 килодальтон. На той же чашке природный АПТ появляется на уровне около 60 килодальтон. Результаты указывают на то, что АПТ, абсорбированный на колонке со сродством к антителу и элюированный указанным методом, соответствует усовершенствованному АПТ, имеющему молекулярную массу, которая примерно на 10000 меньше, чем молекулярная масса встречающегося в природе типа.
Пример 8. Измерение удельной активности усовершенствованного АПТ.
Количество белка в частично очищенном усовершенствованном АПТ определяют путем измерения всего белка в соответствии с методом BradFord (Bradford, Anal.Bochem., 72, 248 (1976)), используя бычий сывороточный альбумин в качестве эталонного белка. Количество антигена АПТ измеряют иммуноферментным анализом (ELISA).
Фибринолитическую активность определяют фибрин/агарозным чашечным методом и методом растворения пленки 1251-меченого фибрина. Фибрин/агарозную чашку получают путем добавления агара к 95% коагулированному фибриногену. Метод растворения пленки 1251-меченого фибрина осуществляют в соответствии с описанием Hoyraeerts et al. (J.Biol. Chem. 257, 2912, 1982), используя в качестве эталона АПТ из клеток меланомы человека, изготовленный Биоскотт Инк. и стандартизованный в соответствии с Международным Стандартом АПТ (Gaffuey and Curtis, Thromb. Haemostas, 53, 34, 1985).
Значение удельной активности, рассчитанное из значения активности, определенной методом растворения пленки 1251-фибрина, и количества антигена, определенного иммуноферментным анализом (ELISA), составляла от 300000 до 420000 единиц/мг антигена.
Пример 9. Сродство усовершенствованного АПТ к фибрину и активация фибрином
В соответствии с работой Verheijen, et al./EMBOJ, 5, 3525, 1986) исследуют сродство усовершенствованного АПТ к фибрину. К фибриногену при различных концентрациях прибавляют усовершенствованный или встречающийся в природе АПТ (1000 едини/мл), после чего прибавляют одну единицу тромбона с последующей реакцией при комнатной температуре в течение 3 минут. Образованный сгусток фибрина преципицируют центрифугированием со скоростью вращения 16000 об/мин в течение 8 минут и определяют количество АПТ, который не связан с фибрином, путем измерения активности фибрин/агарозным чашечным методом. В результате обнаружено, что усовершенствованный АПТ (VI) проявляет такое же сродство к фибрину, что и природная форма. Для того, чтобы исследовать степень активации плазминогена усовершенствованным АПТ в присутствии или отсутствии фибрина, осуществляют следующий эксперимент. С использованием планшета для титрования встречающийся в природе или усовершенствованный АПТ прибавляют к 0,1 М трис-HCl буферу, pH 7,5, содержащему 0,3 мМ синтетический субстрат п-нироанилид-трипептид S-2251 (H-D-Val-leulys-pNA. HCl, производство Каби Инк), 0,13 мкМ плазминогена без плазмина, 120 мкг/мл DESAFIBTM (производство Американ Диагност ика Инк.) и 0,1% Твин 80, с получением полного объема, равного 200 мкл. Систему поддерживают при температуре 37oC. По истечении определенного периода времени измеряют поглощение (оптическую плотность) при длине волны 405 нм, используя Titertech Multiscan 310 Model.
Кривая "доза-эффект" для амидолитической активности усовершенствованного АПТ (VI) и встречающегося в природе АПТ приведена на фиг. 22. Сдвиг кривой в зависимости "доза-эффект" вследствие прибавления DESAFIBTM для встречающегося в природе АПТ соответствует значению в 158 раз, тогда как для усовершенствованного АПТ достигает 100 раз. Это связано с тем, что активность усовершенствованного АПТ (VI) в отсутствие препарата DESAFIBTM ниже, приблизительно на 1/20, нежели активность натурального АПТ.
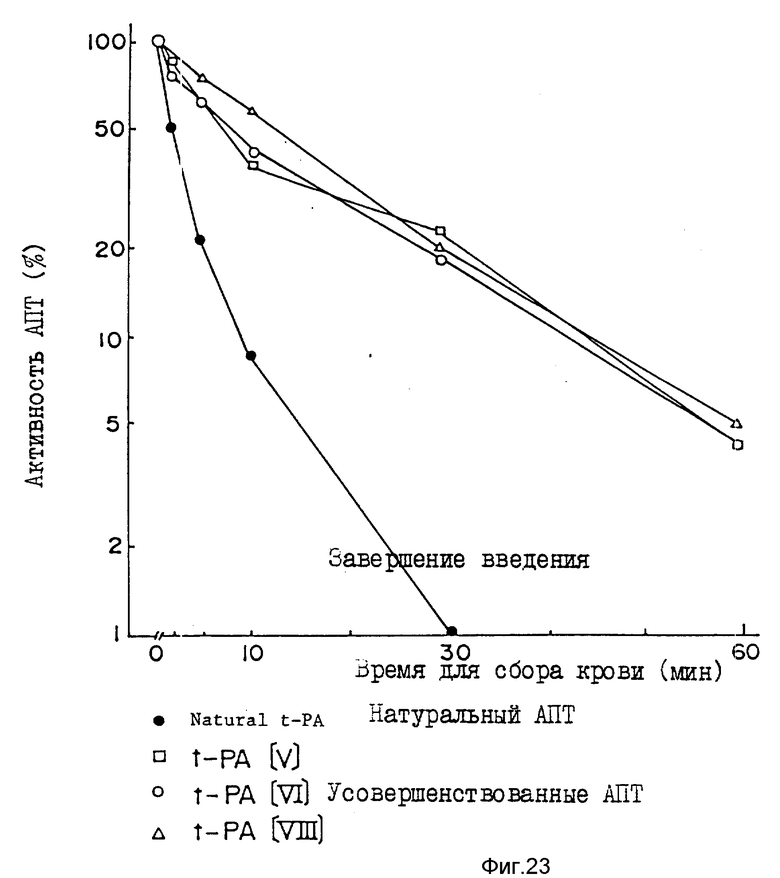
Пример 10. Анализ усовершенствованного АПТ на фибринолитическую активность в кровотоке кролика.
Фармакинетику путем сравнения активности встречающегося в природе АПТ (н-АПТ) и усовершенствованного АПТ настоящего изобретения на кролике. Как явствует из фиг. 23, усовершенствованный АПТ показывает заметное пролонгирование биологического полупериода существования в активном состоянии (натуральный АПТ показывает полупериод существования в течение 1-2 мин, тогда как усовершенствованный АПТ биологически активен в течение 8-15 минут). Кроме того, очевидно, что значение активности, равное 5% (значение через 30 с после введения составляет 100%), все еще остается в усовершенствованном АПТ даже через 60 минут после его введения (природный АПТ через 60 минут проявляет активность, равную 0,1% от начальной).
Данный эксперимент осуществляют следующим образом
Для испытания отбирают японского белого кролика весом 2,4 кг. Под анестезией пентобарбиталом АПТ вводят через периферическую вену уха. Доза составляет 15400 единиц (0,8 мл) усовершенствованного АПТ на кролика и 5400 единиц (0,8 мл) н-АПТ на кролика (значения определены фибрин-чашечным методом). Затем собирают 2,5 мл крови из бедренной артерии с использованием катетера в различные временные интервалы (от 0,5 до 60 минут) и добавляют в 1/9 объема лимонно-кислого натрия (3,8%). В течение 30 минут после сбора крови осуществляют центрифугирование при малой скорости, отделяя плазму. С использованием отделенной плазмы измеряют активность АПТ в крови.
(1) Измерение активности АПТ.
После разбавления 0,2 мл плазмы 3мМ ледяной уксусной кислотой в 16 раз осуществляют центрифугирование разбавленного продукта при малой скорости вращения с получением преципитатов. Преципитаты растворяют 20 мМ Трис-HCl, pH 7,4, с 140мМ NaCl в объеме, эквивалентном объему плазмы, с получением фракции эуглобулина. Активность АПТ определяют путем прибавления этой фракции эуглобулина в чашку фибрин/агарозы. После инкубирования чашки при температуре 37oC в течение 16 часов активность АПТ наблюдают в виде бляшки. Стандартную кривую для фибрин/агарозного чашечного метода получают путем разбавления АПТ, используемого для введения животному, до 0,1-10000 единиц/мл. Определенную таким образом активность АПТ крови выражают в процентах, используя активность АПТ, полученную сбором крови через 30 с после введения, взятую за 100%.
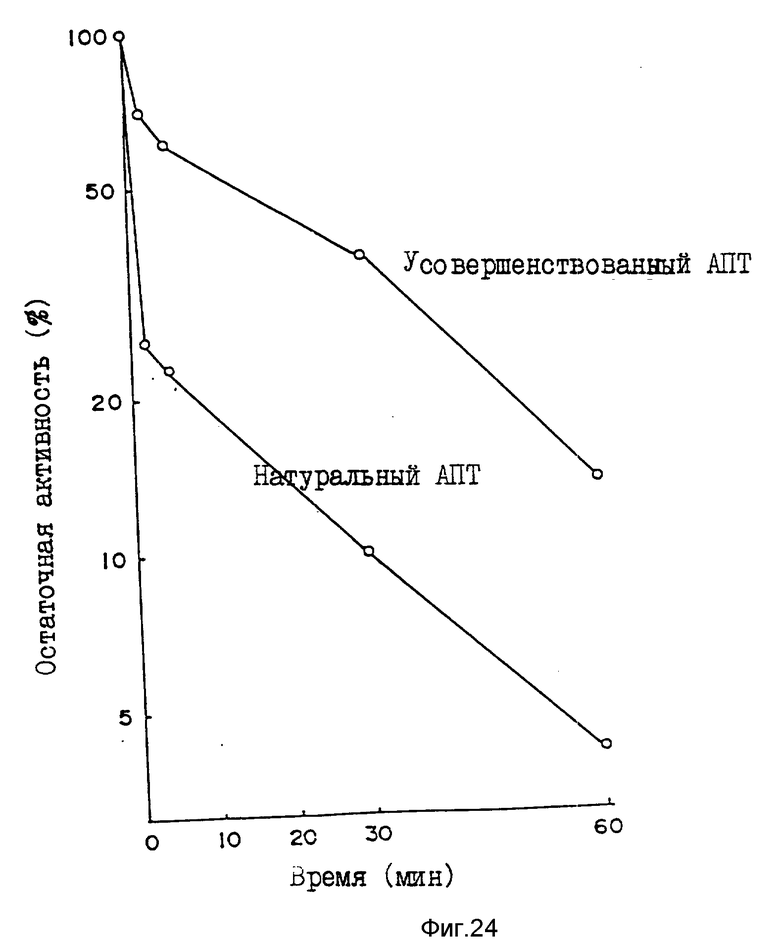
Пример 11. Устойчивость усовершенствованного АПТ (VI) к воздействию тепла и кислот.
Для определения устойчивости к воздействию тепла усовершенствованный АПТ (VI) и натуральный АПТ разбавляют 50 мМ Трис-буфером, содержащим 100 мМ NaCl и 0,01% Твин 80, pH 7,4, до концентрации 100 мкг/мл соответственно. Каждый раствор выдерживают в кипящей воде (температура 98oC) в течение 2-60 минут. После охлаждения определяют остаточную активность методом фибрин-чашки. Как показано на фиг. 24, снижение активности усовершенствованного АПТ (VI) незначительно по сравнению со снижением активности натурального АПТ. Например, после термообработки в течение 2 минут активность натурального АПТ снижается до 25%, тогда как усовершенствованный АПТ (VI) все еще сохраняет активность на уровне 71%.
Для исследования кислотоустойчивости усовершенствованный АПТ (VI) и натуральный АПТ растворяют в 0,5н. растворе HCl при концентрации 100 мкг/мл с последующим отстаиванием при комнатной температуре в течение 30 минут. После нейтрализации определяют активность фибрин-чашечным методом. Усовершенствованный АПТ не обнаруживает никакого изменения активности, тогда как активность натурального АПТ снижается на 50%.
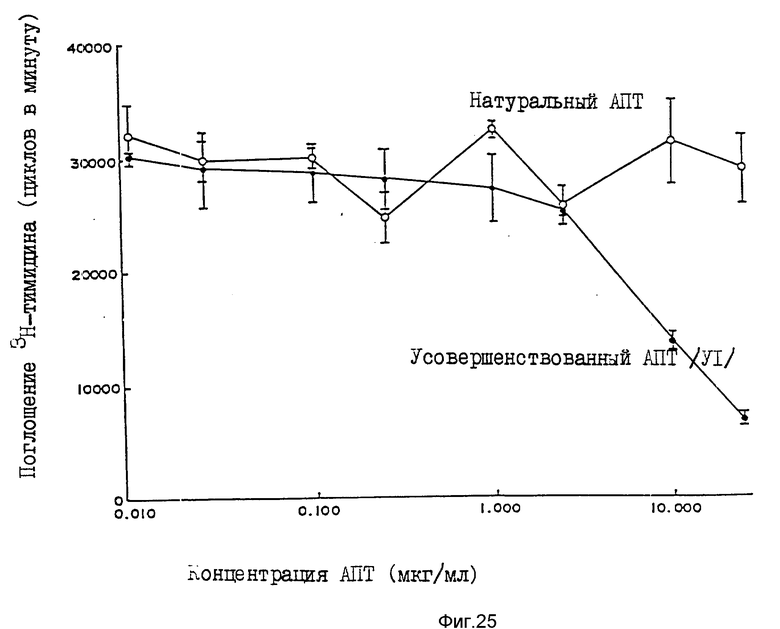
Пример 12. Ингибирование активного фактора, стимулирующего лимфоциты, усовершенствованным АПТ (VI)
Усовершенствованный АПТ (VI) и натуральный АПТ соответствующим образом разбавляют средой тканевой культуры PPM1 1640, содержащей 7%-ную околоплодную сыворотку теленка и 58 мкМ 2-меркаптоэтанол. 100 мкл разведения загружают в 96-луночный планшет для культуры тканей, после чего прибавляют по 50 мкл клеточной суспензии, содержащей тимоциты (2•107 клеток/мл) от мышей C3H/HeJмужского пола в возрасте от 4 до 6 недель, конканавалин A (1,2 мкг/мл), а также по 50 мкл ИЛ-1 (4 единицы/мл, Aenzyme Inc), с последующим культивированием в течение 48 часов в инкубаторе при температуре 37oC, содержащем 5% двуокись углерода. Затем прибавляют H3-тимидин при концентрации 0,5 мк. куб. дюйм /20 мкл/лунку. После культивирования в течение 18 часов клетки собирают на стекловолоконный фильтр и количество 3H-тимидина, введенное в клетки, измеряют жидкостным сцинтилляционным счетчиком, определяя активность фактора, стимулирующего лимфоциты.
Как показано на фиг.25, натуральный АПТ не ингибирует активность фактора, стимулирующего лимфоциты, а усовершенствованный АПТ существенно подавляет ее. При испытании только с растворителем не отмечено никакого воздействия.
Пример 13. Противовоспалительная активность, основанная на действии по отношению к денатурированному белку.
1) Получение денатурированного белка.
После инкубирования белкового раствора (5 мг/мл) в 0,1 н. растворе HCl или 0,1 н. растворе NaOH при температуре 37oC в течение 2-3 часов белковый раствор нейтрализуют тем же количеством NaOH или HCl.
2) Сродство усовершенствованного АПТ (VI) к денатурированному белку.
Метод: в соответствии с методикой, приведенной ниже, денатурированный белок "сцепляют" с нитроцеллюлозной пленкой. Затем измеряют количество усовершенствованного АПТ, связанного с обработкой белком и нитроцеллюлозной пленкой, оценивая таким образом сродство усовершенствованного АПТ к денатурированному белку.
Кусок нитроцеллюлозной пленки погружают в 20 мМ Трис-HCl буферный раствор, pH 7,5, содержащий 140 мМ NaCl.
Сушка. Денатурированный белок (50 мкг/10мкл) выпускают по капле на кусок нитроцеллюлозной пленки.
Сушка. Блокирование 3%-ным раствором желатины.
Промывка. Кусок нитроцеллюлозной пленки погружают в раствор усовершенствованного АПТ /1мкг/мл/.
Промывка. Прибавляют плазминоген и синтетический субстрат S-2251, после чего осуществляют инкубирование при температуре 37oC (количественный анализ на абсорбированный усовершенствованный АПТ).
Измерение поглощения при 405 нм.
Результаты: как показано в таблице 3, усовершенствованный АПТ показывает сродство к иммуноглобулину G, обработанному HCl, к альбумину, обработанному HCl, к альбумину, обработанному NaOH. С другой стороны, усовершенствованный АПТ не проявляет сродства к интактному иммуноглобулину G и альбумину.
3) Активация усовершенствованного АПТ (VI) денатурированным белком.
Метод: плазминоген (0,0078 единицы в 10 мкл), 100 мкл 3 мМ синтетического субстрата S-2251 и различные количества буфера TBS прибавляют в реакционный раствор активатора усовершенствованного АПТ (денатурированного белка, BrCN - обработанного фибриногена и т.п.) при различных концентрациях, получая 0,275 мл реакционного раствора. Усовершенствованный АПТ (2,5 н/г в 25 мкл) прибавляют в реакционный раствор с целью инициации реакции. После взаимодействия в течение определенного периода времени в реакционный раствор прибавляют 2% додецилсульфат натрия (эквимолярное количество) с тем, чтобы остановить реакцию. Путем измерения оптической плотности (OD 405) определяют активность усовершенствованного АПТ.
Результаты: как показано на фиг.26, NaOH - обработанный альбумин и HCl - обработанный иммуноглобулин G демонстрируют выраженное активирующее действие усовершенствованного АПТ. В частности, в HCl-обработанном иммуноглобулине G активация является сильной, а активность HCl-обработанного иммуноглобулина G примерно равна активности BrCN-обработанного фибриногена, причем при концентрации, которая в несколько раз меньше. Интактные альбумин и иммуноглобулин G не проявляют активацию.
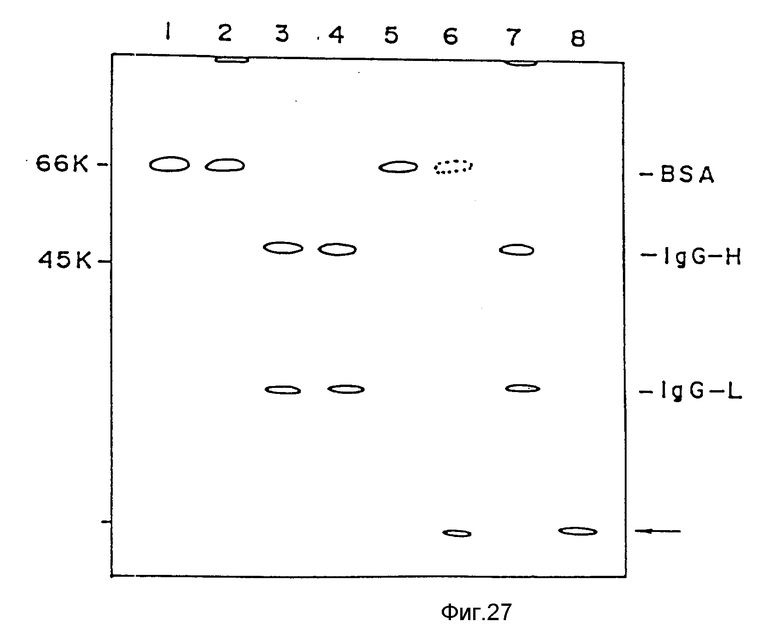
4) Деградация денатурированного белка под действием усовершенствованного АПТ (VI).
Метод: после взаимодействия денатурированного белка с усовершенствованным АПТ при таких же условиях, как и в методе, описанном в предыдущем подпункте, за исключением того, что в реакционную систему не прибавляют синтетический субстрат S - 2251, а количество денатурированного белка составляет 133 мкг/мл, осуществляют электрофорез в полиакриламидном геле с додецилсульфатом натрия в присутствии β -меркаптоэтанола.
Результаты: как показано на фиг.27, денатурированный обработкой NaOH или обработкой HCL белок приводит к исчезновению на картине эф-белковых полос и образованию продуктов деградации, что указывает на его разложение. С другой стороны, при использовании интактного альбумина не обнаружено никакого изменения в эф-картине после взаимодействия с усовершенствованным АПТ, а следовательно, не обнаружено никакой деградации денатурированного белка.


















Изобретение относится к получению тканевого активатора плазминогена (усовершенствованный АПТ), имеющего пролонгированный биологический полупериод существования, повышенную устойчивость к воздействию тепла и кислот и эффективный в качестве ингибитора воспаления вокруг сайта, где образован тромб. Для его получения проводят культивирование хозяйских клеток, трансформированных рекомбинантной ДНК, содержащей конкретную последовательность ДНК, кодирующую аналог tpА. 2 с.п.ф-лы, 3 табл. 29 ил.
где Y - Glu-Ile-Lys;
H2N - аминоконец;
-СООН - карбоксиконец;
R - прямая связь или аналогичная ей последовательность, содержащая замены, и/или делеции, и/или вставки, не связанные с изменениями активности,
и обладающий следующими свойствами: фибринолитической активностью, определенной методом растворения пленки 1 2 5I-фибрина, способностью активироваться фибрином и активностью улучшенного tpA в отсутствии фибрина, являющейся более низкой, чем активность природного tpA, увеличенным, по сравнению с природной формой, временем полужизни, повышенной, по сравнению с природным tpA, устойчивостью к кислотам и нагреванию, способностью ингибировать фактор активации лимфоцитов, способностью активироваться денатурированным белком.

