Область изобретения
Областью техники данного изобретения является получение рекомбинантных белков с помощью генетической инженерии эукариотических клеток.
Предпосылки изобретения
Многие ферменты или гормоны продуцируются клеткой в виде неактивных проферментов или прогормонов и затем превращаются in vivo в месте «использования» или в «момент использования» в соответствующие активные вещества. Примерами таких ферментов являются некоторые металлопротеазы матрикса, такие как коллагеназа I, плазминоген, химотрипсиноген, трипсиноген. Указанный механизм «активации» является общим для гормонов, таких как хромогранин, кальцитонин и т.д.
Среди молекул, синтезируемых в клетке в виде предшественников, типичным примером можно считать урокиназу (активатор плазминогена уринарного типа, у-АП). Этот фермент представляет собой гликопротеид плазмы, относящийся к обширному семейству сериновых протеаз. Его основная функция, наряду со многими другими функциями, состоит в физиологической активации плазминогена в плазмин (Barlow, G.H., Methods in Enzymology, 45: 239-244, (1976)). Плазмин является ключевым протеолитическим ферментом в фибринолитическом процессе, который приводит к фибринолизу (Robbins, K.C. and Summaria, L., Methods in Enzymology, 45: 257-273, (1976)).
У человека у-АП экспрессируется в различных тканях в виде пре-профермента (препроурокиназы) и затем секретируется в кровь в виде проурокиназы в одноцепочечной форме. Поэтому указанную зимогенную форму часто сокращенно обозначают оц-уАП (одноцепочечный у-АП). оц-уАП в SDS-ПААГ имеет видимую молекулярную массу 50000-54000 дальтон и является каталитически неактивным.
В крови плазмин осуществляет протеолитическое расщепление, которое превращает профермент оц-уАП в двухцепочечный полипептид, называемый дц-уАП, который является каталитически и физиологически активным.
дц-уАП состоит из А- и В-цепи, связанных друг с другом дисульфидной связью. В-цепь несет в себе каталитический сайт, а также сайт N-гликозилирования, расположенный у Asn302 (Bergwerff A.A., van Oostrum J., Kamerling J.P. and Vliegenthart J.F., Eur. J. Biochemistry, 1995, 228: 1009-1019). Указанную форму у-АП называют двухцепочечным у-АП с высокой молекулярной массой (дц-уАП ВММ). Дальнейшее протеолитическое расщепление, осуществляемое плазмином в А-цепи дц-уАП ВММ, приводит к образованию более короткой двухцепочечной молекулы урокиназы, называемой дц-уАП НММ (двухцепочечный у-АП с низкой молекулярной массой).
Многочисленные попытки получения дц-уАП посредством технологий рекомбинантной ДНК с одной стороны подтвердили пригодность такой молекулы для клиники, и с другой стороны, потребность в таком подходе, необходимом по причинам безопасности и чистоты.
В патенте EP 154272 описано получение рекомбинантного гликозилированного оц-уАП, полученного путем введения последовательности кДНК в клетки животных.
В патенте EP 303028 описано получение рекомбинантного гликозилированного оц-уАП, полученного путем введения геномной последовательности в клетки млекопитающих.
Получение, описанное в указанных патентах, относится к рекомбинантной урокиназе (оц-уАП) в энзиматически неактивной форме.
Получение активного фермента (дц-уАП) с помощью рекомбинантной ДНК еще остается открытым вопросом, главным образом вследствие его сложного внеклеточного процессинга. На самом деле, в некоторых рекомбинантных эукариотических системах, описанных до настоящего времени, получают небольшие количества рекомбинантного активного дц-уАП. В этих эукариотических системах полученный в результате продукт является смесью оц-уАП и дц-уАП (Cheng D. Et al., Chinese Journal of Biotechnology, 1994, 9: 151-159). Это главным образом является следствием неэффективности стадий процессинга, вовлеченных в активацию оц-уАП. Данный факт выдвигает некоторые проблемы, касающиеся очистки двух отдельных форм.
Клиническое применение смеси уАП-ВММ и уАП-НММ привело бы к другим проблемам, связанным с точной воспроизводимостью соотношения соответствующих компонентов, которые, кроме того, проявляют разные фармакологические свойства.
С другой стороны, получение рекомбинантного оц-уАП привело к другой стратегии терапии, включающей в себя введение оц-уАП, как такового, превращаемого затем эндогенным плазмином в активный фермент в потоке крови. В этом случае возникающие проблемы являются следствием неопределенности дозы из-за непредсказуемости скорости превращения in vivo.
Однако в настоящее время молекулярные формы дц-уАП ВММ и НММ остаются единственными молекулами, представляющими обоснованный интерес с точки зрения терапии, на основании их успешного применения в течение многих лет для лечения при нарушениях процесса фибринолиза в клинике.
Обе молекулы, которые доступны в настоящее время в виде фармацевтических продуктов, не являются рекомбинантными (для обзора см.: Scripp's thrombolytic report, PJB Publications Ltd., (1993)). Форму ВММ в настоящее время получают экстракцией из мочи человека, как описано, например, в DE 3439980, тогда как форма НММ была получена из культур клеток эмбриональной почки человека в присутствии сыворотки, как описано в DT 2551017. В этом случае сыворотка снабжает клетки не только факторами роста, но также плазмином для превращения проурокиназы в форму дц-уАП НММ. Однако в последнее время по отношению к этому продукту возникло беспокойство, связанное с безопасностью, из-за его происхождения из эмбриональных клеток ("The pink sheet", February 1, 1999, page 6).
Сущность изобретения
Данное изобретение относится к способу получения зрелых рекомбинантных белков с использованием генетически трансфицированных линий эукариотических клеток. Процесс созревания рекомбинантного предшественника достигается выращиванием линии клеток в присутствии алкановых кислот или их производных, или их солей в течение, по меньшей мере, 24 часов. Согласно предпочтительному варианту данного изобретения, зрелый рекомбинантный белок представляет собой двухцепочечный уАП (дц-уАП), экспрессированный с клонированной кДНК предшественника.
Поэтому следующим предметом данного изобретения является способ получения, выделения и очистки рекомбинантного дц-уАП ВММ и НММ и полученные продукты, соответствующие двум формам зрелой урокиназы, получаемым согласно таким способам.
Следующие предметы данного изобретения представляют собой способ лечения тромбоэмболических нарушений, при котором используют рекомбинантный зрелый дц-уАП ВММ и НММ и фармацевтические композиции, содержащие в качестве активных агентов рекомбинантный дц-уАП ВММ и НММ, полученный согласно способу настоящего изобретения.
Подробное описание изобретения
Данное изобретение относится к способу получения зрелых рекомбинантных белков в культуральной среде генетически модифицированных эукариотических клеток.
Заявители неожиданно обнаружили, что в том случае, когда генетически модифицированные эукариотические клетки в течение периода времени, по меньшей мере, равного 24 часам, обрабатывают алкановыми кислотами или их производными, или их солями, которые добавляют в среды для культур клеток, превращение рекомбинантного белка-предшественника в соответствующую зрелую форму происходит очень эффективно, и зрелый активный белок накапливается в большом количестве в надосадке культуры клеток.
Превращение белка-предшественника в зрелую форму, в частности, эффективно в том случае, когда обработку алкановыми кислотами или их производными, или их солями, комбинируют с понижением температуры культуры клеток до значений, равных или ниже 37°С, предпочтительно составляющими от 30°С до 36°С, наиболее предпочтительно 34°С, что указывает на синергетическое действие.
Термин «клетки, подвергнутые генетическим манипуляциям», относится к клеткам, которые были трансфицированы или трансформированы экзогенной ДНК, предпочтительно кДНК, кодирующей требуемый белок-предшественник. В предпочтительном варианте последовательностью кДНК является нуклеиновая кислота, кодирующая предшественник урокиназы (препроурокиназу или проурокиназу). Термин «белок-предшественник» относится к полипептиду, который секретируется, или другим образом продуцируется клеткой, и который требует дальнейшего протеолитического расщепления или «процессинга» для его активации, или для его продукции в форме близкой, насколько это возможно, к биологически активной или природной форме. Другими примерами таких белков-предшественников, кодируемых соответствующими последовательностями ДНК или кДНК, являются такие зимогены, как: трипсиноген, химотрипсиноген, плазминоген, проамилаза, пролипаза, металлопротеазы матрикса, т.е. коллагеназа I, факторы, относящиеся к каскаду системы комплемента, и прогормоны, такие как: профактор роста гепатоцитов (Pro-HGF), пре-проинсулин, соматостатин, хромогранин-А и т.д. В предпочтительном варианте белок-предшественник представляет собой пре-профермент, и наиболее предпочтительно является пре-проурокиназой или проурокиназой, которая становится активной в форме дц-уАП (двухцепочечная урокиназа). Урокиназа (уАП) представляет собой сериновую протеиназу, главной функцией которой in vivo является активация плазминогена в плазмин. У человека уАП экспрессируется в разных тканях в виде пре-профермента, и затем секретируется в кровь в виде каталитически неактивной проурокиназы в одноцепочечной форме (оц-уАП), которая в свою очередь процессируется плазмином в двухцепочечный уАП (дц-уАП), каталитически активный. дц-уАП построен из А-цепи и В-цепи, связанных друг с другом дисульфидной связью, причем последняя несет каталитический сайт. Активный дц-уАП в природе выявляется в крови и моче в двух разных формах: дц-уАП ВММ и дц-уАП НММ, которые отличаются своими А-цепями, более короткая А-цепь в дц-уАП НММ.
Алкановые кислоты или их производные, или их соли в предшествующем уровне техники до настоящего времени использовали для того, чтобы повысить выход продукции/секреции рекомбинантных белков в системах культур клеток млекопитающих. Согласно данному изобретению алкановые кислоты или их производные, или их соли действуют как «усилители процессинга». Насколько известно авторам настоящего изобретения, алкановые кислоты или их производные или их соли впервые используются в качестве «усилителей процессинга» белков-предшественников, как описано выше.
Алкановые кислоты или их соли, или их производные предпочтительно представляют собой прямую цепь С2-С10, и более предпочтительно C3-C4. Алкановые кислоты или их соли и/или производные предпочтительно выбирают из бутирата или пропионата, предпочтительно в виде их натриевых солей, или трибутирина или фенилбутирата. Особенно предпочтительным является бутират натрия.
Алкановые кислоты или их производные добавляют в среду культивирования генетически модифицированных клеток в концентрации, составляющей от 0,01 до 500 мМ. В случае клеток млекопитающих предпочтительные пределы концентрации составляют от 0,1 до 20 мМ, наиболее предпочтительно 0,5-2,5 мМ. Однако будет понятно, что такие концентрации могут варьировать в соответствии с используемой линией клеток и в соответствии с другими факторами, такими как жизнеспособность культуры клеток во время или в конце обработки. Согласно предпочтительному варианту эукариотические клетки являются клетками млекопитающих, выбранными из клеток, обычно используемых для продукции рекомбинантных белков: клеток HEK-293, CV-1, COS, BSC-1, MDCK, A-431, BHK, CHO. В предпочтительном варианте клетки СНО представляют собой клетки СНО-Messi (ECACC № 93080520).
Следующим предметом данного изобретения является способ получения рекомбинантного дц-уАП человека в клетках СНО, включающий добавление алкановых кислот или их производных, или их солей в культуральную среду, не содержащую сыворотки, в которой линия клеток СНО, генетически модифицированная эукариотическим экспрессирующим вектором, несущим кДНК препроурокиназы, поддерживается в течение периода времени, по меньшей мере, равного 24 часам при температуре, равной или ниже 37°С, предпочтительно находящейся в интервале от 33 до 35°С, наиболее предпочтительно при 34°С. Следующим предметом данного изобретения является способ превращения проурокиназы (оц-уАП) в дц-урокиназу (дц-уАП) добавлением алкановых кислот или их производных, или их солей в среду культивирования клеток СНО и ферментацией культуры клеток при температурах, равных или ниже 37°С и в течение периода времени, по меньшей мере, равного 24 часам.
Как указано выше относительно природной формы, также для рекомбинантной урокиназы термин «дц-уАП» относится к каталитически активной урокиназе, которая может быть в форме дц-уАП ВММ и НММ. дц-уАП ВММ и НММ отличаются по дифференциально процессированной А-цепи и проявляют очень похожие функциональные активности. дц-уАП ВММ и НММ можно отличить друг от друга, например, в целях анализа по разной электрофоретической картине в не восстанавливающем SDS-ПААГ: дц-уАП ВММ мигрирует в области 50-54 кДа, в то время как дц-уАП НММ при тех же условиях мигрирует в области 30-33 кДа.
Следующим предметом данного изобретения является способ получения рекомбинантного дц-уАП человека, который получают способом, включающим в себя следующие стадии:
а) культивирование подвергнутых генетическим манипуляциям клеток СНО, трансфицированных кДНК или геном пре-проУК, в культуральной среде, содержащей алкановые кислоты или их производные, или их соли, предпочтительно бутират или его соли в концентрации, находящейся в пределах от 0,1 до 20 мМ, и при температуре, в интервале от 30°С до 37°С, еще более предпочтительно в интервале от 33°С до 35°С наиболее предпочтительно при 34°С;
b) продолжение указанного культивирования в течение периода времени, по меньшей мере, равного 24 часам, или предпочтительно находящегося в пределах от 48 до 200 часов, наиболее предпочтительно примерно 120 часов (около пяти дней) и
с) получение надосадка культуры клеток для выделения указанного рекомбинантного дц-уАП человека.
В соответствии с предпочтительным вариантом способа согласно настоящему изобретению линией клеток, подвергнутой генетическим манипуляциям, является стабильный трансформант СНО, наиболее предпочтительно трансформант СНО-Messi (ECACC № 93080520), отобранный на основе приобретения метаболического маркерного гена. Средой для культивирования предпочтительно является культуральная среда без сыворотки, еще более предпочтительно определенная среда, не содержащая сыворотки и белка, такая как коммерчески доступные среды. В предпочтительном варианте средой для культивирования клеток является CHOMaster®.
Под стабильным трансформантом или трансфектантом СНО подразумевают клон СНО, трансфицированный экспрессирующим вектором, который стабильно интегрирован в геном клетки. Экспрессирующий эукариотический вектор выбирают согласно критериям, хорошо известным в данной области: наличие сильного эукариотического или вирусного промотора, такого как CMV-IE, поздний или ранний промотор SV40, RSV, чтобы направлять транскрипцию экзогенной ДНК; сигнал полиаденилирования; энхансеры транскрипции и другие регуляторные районы, которые выбирают согласно способам, хорошо известным в данной области. Другими характерными признаками экспрессирующих векторов являются: начало репликации прокариот, ген для селекции в эукариотических, а также в прокариотических клетках, такой как ген в-лактамазы или kanR, или neoR, или tet-F, или гигромицин-R, которые хорошо известны специалистам в данной области.
Согласно предпочтительному варианту настоящего изобретения селекцию эукариотических клеток осуществляют благодаря экспрессии метаболического маркерного гена: Trp-синтазы (ген trpB) на Trp-ауксотрофных клетках СНО, или по гистидинолдегидрогеназе (ген hisD) на His-ауксотрофных клетках СНО.
Стабильные клоны предпочтительно выбирают на основе их ростовых свойств, уровней продуктивности и их стабильности в культуре.
Культивирование выбранного клона СНО обычно проводят в биореакторе согласно протоколам, хорошо известным в данной области. Согласно предпочтительному варианту культивирование выполняют партиями. Предпочтительная исходная концентрация клеток составляет около 3×105 живых клеток/мл, и жизнеспособность клеток в инокуляте обычно составляет более 95%, согласно измерению способом исключения с красителем трипановым синим. Обычно соотношение «объем инокулята/объем свежей среды» варьирует от 1:1 до 1:5, а также в соответствии с общей емкостью биореактора и ростом клеток.
При плотности клеток в пределах от 1×106 до 4×106 клеток/мл, обычно достигаемой через 3-5 дней роста, клетки отделяют от использованной среды, например, тангенциальным фильтрованием или центрифугированием, и ресуспендируют в таком же исходном объеме свежей среды, в которую были добавлены алкановые кислоты или их соли, или их производные в конечных концентрациях в пределах от 0,1 до 20 мМ. Алкановые кислоты или их соли или их производные добавляют в момент внесения инокулята клеток или после этого. Добавление алкановых кислот или их солей или их производных необязательно можно повторять во время роста или ферментации. Алкановые кислоты, соли и/или производные предпочтительно выбраны из: бутирата или пропионата, предпочтительно их натриевых солей, трибутирина и фенилбутирата. Особенно предпочтительным является бутират и его натриевая соль, которую добавляют в предпочтительной концентрации от 0,5 мМ до 2,5 мМ, наиболее предпочтительно от 1 мМ до 1,5 мМ.
Влияние алкановых кислот на продукцию рекомбинантного процессированного белка дополнительно усиливают понижением температуры культуры клеток ниже 37°С, предпочтительно в пределах температур, составляющих от 30°С до 36°С, еще более предпочтительно 33-35°С, наиболее предпочтительно 34±0,5°С. Во время ферментации также контролируют уровни глюкозы, и предпочтительно поддерживают выше 1 г/л. Ферментацию партиями во время фазы продукции предпочтительно проводят в соответствии со следующими параметрами:
Температура: 34±1°С, предпочтительно 34±0,5°С
рН: 7,15±0,1
рО2: 50±20%
В соответствии с предпочтительными условиями ферментации более высокую концентрацию дц-уАП получают через 5 дней культивирования после добавления алкановой кислоты, их производных, или их солей. За продукцией активного продукта можно проследить с помощью функциональных или иммунологических анализов. Например, за продукцией дц-уАП можно проследить хромогенным анализом, таким как анализ Pefachrome®. В альтернативном случае продукцию активного дц-уАП или исчезновение оц-уАП можно отслеживать в SDS-ПААГ в денатурирующих и восстанавливающих условиях, благодаря разной картине миграции при электрофорезе: оц-уАП в действительности мигрирует как одноцепочечный полипептид ˜50-54 кДа, в то время как дц-уАП ВММ разделяется на А-цепь и В-цепь, соответственно мигрирующие в области ˜20 и ˜33 кДа.
Согласно предпочтительному варианту истощенный надосадок культуры клеток (т.е. культуральная среда, в которой выросли клетки), содержащий дц-уАП, обычно собирают после 3-8 дней культивирования, обычно на пятый день, когда баланс между уровнями рекомбинантного белка и выживаемостью клеток (последняя предпочтительно сохраняется выше 70%) оптимален. Альтернативно надосадок после истощения питательных веществ собирают тогда, когда оц-уАП отсутствует согласно измерениям в SDS-восстанавливающем ПААГ, и максимально превращен в дц-уАП, где под дц-уАП имеют в виду смесь дц-уАП ВММ и НММ. Обычно оптимальное время для выделения зрелого дц-уАП находится в пределах от 48 до 200 часов, при этом предпочтительный период времени равен 120 часам культивирования в присутствии алкановых кислот или их производных, или их солей, и обычно соответствует уровню продукции дц-уАП около 4000 IU/мл. Согласно описанным вариантам настоящего изобретения превращение форм предшественников (пре-про-уАП, про-уАП, оц-уАП) в каталитически активный дц-уАП характеризуется эффективностью более 95% согласно определению аналитическим электрофорезом в восстанавливающем SDS-ПААГ. Из общего количества полученного дц-уАП около 80% находится в ВММ-форме, и оставшиеся 20% - в НММ-форме. Поэтому следующим предметом данного изобретения является надосадок культуры клеток, содержащий дц-уАП, получаемый согласно способу, описанному и охарактеризованному стадиями с а) по с).
В одном из следующих вариантов настоящее изобретение относится к способу получения рекомбинантного каталитически активного дц-уАП ВММ и НММ, который образуется в результате эффективного превращения каталитически неактивного оц-уАП или про-уАП или проурокиназы, непосредственно осуществляемого в истощенной культуральной среде и характеризуемого скоростью превращения предшественника в зрелый белок, которая составляет более 95%.
Следующий предмет данного изобретения представляет собой хроматографический способ выделения рекомбинантных форм молекул дц-уАП ВММ и дц-уАП НММ, характерной особенностью которого является использование надосадка культуры клеток, полученного на стадии с) процесса получения дц-уАП.
Согласно предпочтительному варианту изобретения дц-уАП НММ отделяют от дц-уАП ВММ способом, включающим в себя ионообменную хроматографию, и предпочтительно в соответствии со следующими дополнительными стадиями: d) подкисление надосадка культуры клеток слабой кислотой до рН в пределах от 5,0 до 5,8, необязательно с добавлением неионного детергента и фильтрацией, е) контактирование надосадка с колонкой для ионообменной хроматографии при значении рН в пределах от 5,5 до 6,5; f) высвобождение дц-уАП НММ добавлением буферного раствора со значением рН в пределах от 5,5 до 6,5, кроме того, содержащего моновалентный ион в концентрации от 200 до 300 мМ, как, например, 250 мМ NaCl в фосфатном буфере; g) высвобождение дц-уАП ВММ добавлением буферного раствора при значениях рН от 5,5 до 6,5, дополнительно содержащего моновалентные ионы в концентрации, равной, по меньшей мере, 400 мМ, как, например, 500 мМ NaCl в фосфатном буфере. В ходе хроматографии также проводят промежуточные промывки, чтобы избавиться от всех компонентов, не специфично связанных с дц-уАП ВММ и НММ, и промывают буферами и/или растворами, хорошо известными в данной области.
Согласно еще одному варианту настоящее изобретение относится к дополнительной очистке рекомбинантного дц-уАП ВММ и НММ вплоть до терапевтической степени очистки, при которой используют две формы, выделенные, соответственно, на стадиях g) и f) процесса разделения, и которая, кроме того, включает в себя аффинную хроматографию на колонке с бензамидином. Указанная колонка позволяет проводит процедуру очистки, чтобы избавиться от возможных следов оц-уАП, если они присутствуют.
Хроматографию на бензамидине обычно выполняют согласно способам, хорошо известным в данной области. Согласно предпочтительному варианту способа очистки дц-уАП ВММ очищают хроматографией на бензамидине, дополнительно подвергая элюат, полученный на стадии g) обработке на следующих дополнительных стадиях: g') контактирование элюата, содержащего дц-уАП ВММ с бензамидиновой колонкой при значениях рН от 6,2 до 6,8; g'') высвобождение дц-уАП ВММ буферным раствором при значениях рН в пределах от 3,8 до 4,2, кроме того, содержащим: ацетат натрия в концентрации от 50 до 150 мМ, NaCl в концентрации от 300 до 500 мМ; g''') необязательно дальнейшее контактирование выделенного дц-уАП ВММ с колонкой для гель-фильтрации и высвобождение дц-уАП ВММ фосфатным или ацетатным буфером с низким содержанием соли, как, например, 5 мМ натрий-фосфатным буфером, при значениях рН от 4 до 7.
Согласно еще одному варианту настоящего изобретения дц-уАП НММ очищают хроматографией на бензамидине посредством дальнейшей обработки элюата со стадии f) согласно следующим дополнительным стадиям: f') контактирование элюата, содержащего дц-уАП НММ с бензамидиновой колонкой при значениях рН от 6 до 8; f'') высвобождение дц-уАП НММ раствором при значениях рН от 3,8 до 4,2, кроме того, содержащего ацетат натрия в концентрации в пределах от 50 до 150 мМ, NaCl в концентрации от 300 до 500 мМ; f''') кроме того, необязательно контактирование выделенного дц-уАП НММ с колонкой для гель-фильтрации и высвобождение дц-уАП НММ буферным раствором при значениях рН от 4 до 7, как например, 5 мМ натрий-фосфатным или ацетатным буфером.
Продукт(ты), получаемые при комбинировании способов продукции (стадии от а до с), разделения (стадии от d до g) и очистки (стадии от g' до g''' и от f' до f'''), отличающемся тем, что использовали надосадок, полученный из истощенной среды культуры клеток, полученной на стадии с) процесса получения, представляет собой рекомбинантный дц-уАП. Рекомбинантный дц-уАП является молекулярной формой ВММ, как определено ранее, и его получают при уровне очистки, составляющем более 90%, и/или дц-уАП в НММ-форме, как определено ранее, при уровне очистки выше 90% согласно определению аналитическим электрофорезом в SDS-ПААГ.
Очищенный рекомбинантный дц-уАП (ВММ и/или НММ) является активной формой, что подтверждается функциональными и биохимическими анализами, и имеет терапевтическую степень очистки в соответствии с требованиями Европейской фармакопеи. Поэтому не требуется никакой дальнейшей обработки и/или очистки, в противоположность рекомбинантным про-уАП или оц-уАП, полученным посредством технологий рекомбинантной ДНК, относящимся к предшествующему уровню техники. Молекулярная форма продукта подтверждается структурными данными, полученными масс-спектроскопией и анализом N-концов способом деградации по Эдману.
Как определено посредством функциональных анализов, таких как анализ фибронолиза, определение константы Михаэлиса-Ментена, анализ связывания ингибитора активатора плазминогена (ИАП-1), рекомбинантные продукты, полученные согласно данному изобретению, функционально неотличимы от экстрагируемого дц-уАП, так как их активности в анализах полностью соизмеримы с активностями экстрагируемого натурального продукта.
Кроме того, рекомбинантные продукты преимущественно получают из клеток СНО, которые обладают хорошо установленной надежностью в смысле продукции рекомбинантных белков.
Очищенные рекомбинантные дц-уАП ВММ и НММ согласно данному изобретению используют в качестве мощных фибринолитических средств для лечения тромбоза и патологических явлений любого другого вида, при которых необходимо фармакологическим путем удалить сгусток плазмы. Их применение подтверждается хорошо проверенным клиническим применением соответствующих природных экстрагируемых форм.
Поэтому следующий вариант изобретения относится к рекомбинантному дц-уАП ВММ/НММ, получаемому согласно способу настоящего изобретения, для лечения тромбоэмболических явлений, требующих удаления сгустков фармакологическим способом, таких как окклюзия периферических артерий, клиренс катетера, эмболия сосудов легких, тромбоз глубоких вен или лечение инфаркта миокарда.
Наилучший способ реализации данного изобретения описан посредством следующих экспериментальных примеров.
Описание чертежей
Фиг.1: восстанавливающий SDS-ПААГ рекомбинантного дц-уАП.
Показан восстанавливающий SDS-ПААГ очищенного дц-уАП, который получен после добавления в среду с культурой клеток 1,2 мМ бутирата и ферментации в течение 5 дней.
При восстанавливающих условиях дц-уАП расщепляется на А-цепь (20 кДа) и В-цепь (33 кДа), а оц-уАП мигрирует примерно в районе 55 кДа. Дорожка 1: очищенная урокиназа из рекомбинантной культуры СНО, выращенной в отсутствии Na-бутирата; дорожка 2: очищенная урокиназа из рекомбинантной культуры СНО, выращенной в течение пяти дней в присутствии 1,2 мМ Na-бутирата. Условия роста описаны в тексте; дорожка 3: стандарт молекулярной массы.
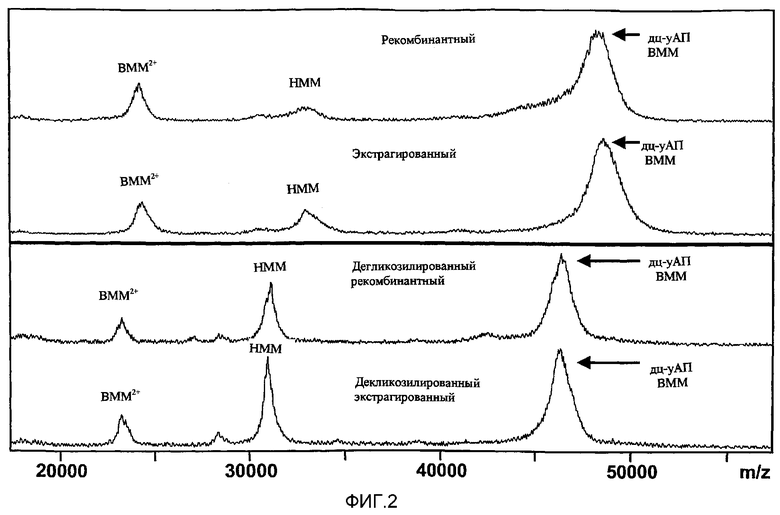
Фиг.2: Анализ масс рекомбинантного и экстрагируемого дц-уАП.
На фигуре показан спектр, полученный с помощью масс-спектрометрического анализа рекомбинантного и экстрагируемого дц-уАП (ВММ и НММ) как в нативной (гликозилированной; два верхних спектра), так и в дегликозилированной формах (два нижних спектра).
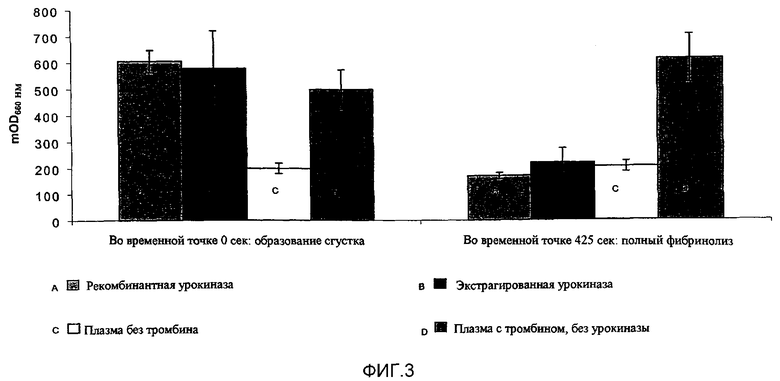
Фиг.3: Анализ фибринолиза.
Эксперимент выполняли добавлением 0,5 мл плазмы человека к 100 мкл урокиназы (1000 U/мл), и инкубировали при 42°С в течение 5 мин. К смеси добавляли 100 мкл тромбина (20 U/мл), и измеряли поглощение при 660 нм в течение 20 мин при 42°С.
Заштрихованные области слева направо во временной точке «0 сек» и временной точке «425 сек» представляют: А) рекомбинантную урокиназу; В) экстрагируемую урокиназу; С) плазму без добавления тромбина (контроль без образования сгустков); D) плазму с тромбином, но без добавления урокиназы (позитивный контроль с образованием сгустков).
Во временной точке 425 сек наблюдается полный лизис в том случае, если в сгустке присутствует рекомбинантная или экстрагируемая урокиназа.
Показано, что рекомбинантная и экстрагируемая урокиназы (дц-уАП ВММ) показывают одно и то же время фибринолиза.
ЭКСПЕРИМЕНТАЛЬНАЯ ЧАСТЬ
Пример 1: Клонирование и отбор стабильных клонов, экспрессирующих пре-про-УК
Последовательность кДНК, кодирующую пре-проурокиназу человека (соответствующую последовательности ID D00244 в Genbank), синтезировали с мРНК линии клеток почки человека (CAKI-1) согласно способам, хорошо известным в данной области, описанным, например, в Molecular Cloning: A laboratory Manual, Sambrook et al., Cold Spring Harbor Laboratory press, (1989).
Коротко, кДНК синтезировали в реакции с обратной транскриптазой AMV (Bцhringer-Mannheim) в присутствии смеси мРНК, олиго dT18 (Bцhringer-Mannheim) и смеси четырех дезоксинуклеотидов (dATP, dGTP, dCTP, dTTP), согласно инструкциям производителя.
Смесь молекул кДНК специфично амплифицировали в ПЦР со следующими 5'- и 3'-праймерами:
Олиго1 (5'): 5'TAGCGCCGGTACCTCGCCACCATGAGA3'
Олиго2 (3'): 5'TGGAGATGACTCTAGAGCAAAATGACAACCA3'
Получили последовательность кДНК длиной 1296 нуклеотидов, кодирующую последовательность пре-проУК человека, и клонировали в интегрирующий экспрессирующий вектор, полученный из pBR322, несущий следующие элементы:
- последовательность кДНК пре-проУК человека под контролем раннего промотора вируса SV40 (Benoist C. and Chambon P. 1981, Nature 290: 304-310).
- селектируемый маркер TrpB для метаболической селекции в клетках СНО-Messi (Hartman, SC Mulligan, RC (1988). Proc. Natl. Acad. Sci. USA, 85: 8047-51).
- устойчивость к ампициллину (ген bla) в качестве маркера устойчивости к антибиотикам в E. coli.
- начало репликации E. coli для амплификации в E. coli.
Конечный полученный рекомбинантный экспрессирующий вектор назвали pTZA9.
Линией клеток СНО, используемой для получения рекомбинантного дц-уАП, была линия клеток СНО-Messi (ECACC, Porton Down, Salisbury, United Kingdom, Рег. № 93080520).
Указанная линия клеток способна расти в суспензии в химически определенной среде CHOMaster® (Ferruccio Messi Cell Culture Technologies, Zürich, Switzerland) без добавления какой-либо сыворотки или белкового компонента. Время удвоения указанной линии клеток в таких средах (полной и селектирующей средах) составляет около 24 часов.
Трансфекцию клеток СНО-Messi вектором pTZA9 осуществляли согласно способу, описанному в Felgner et al., (1987). Proc. Natl. Acad. Sci. USA, 84, 7413-7417, и соблюдали рекомендации производителя реагентов для трансфекции. Коротко, 1 мкг ДНК экспрессирующего вектора pTZA9 (в растворе 100 мкг/мл) смешивали с 30 мкл Lipofectin® (GIBCO BRL, Life Technologies) примерно в 500 мкл экспоненциально растущих клеток СНО-Messi (примерно 1×106 клеток) Смесь для трансфекции выдерживали 30 мин при комнатной температуре перед добавлением среды CHOMaster® с добавлением 25,1 мг/л триптофана и инкубированием в течение ночи при 37°С.
Линия клеток СНО-Messi является ауксотрофом по триптофану, и поэтому обладает абсолютной потребностью в указанной аминокислоте для роста. После трансфекции клеток СНО-Messi вектором, экспрессирующим пре-проурокиназу, несущим также ген триптофансинтазы, клетки СНО-Messi получают преимущество благодаря приобретенной способности продуцировать триптофан, и теперь могут расти в химически определенной среде в отсутствии триптофана при добавлении серина и индола.
Стабильно трансфицированные клетки получили после лимитирующего разведения в селектирующей среде CHOMaster® (без триптофана) с добавлением серина (0,02 г/л) и индола (0,35 г/л).
Селекцию клонов, продуцирующих урокиназу, проводили разведением смеси для трансфекции селективной средой CHOMaster® в лунках для микротитрования. Другая система, используемая для получения отобранных отдельных клонов, продуцирующих урокиназу, представляла собой разведение 1:10 суспензии клеток, содержащей примерно 103 клеток/мл, раствором высокой вязкости, содержащим 0,2 г/л Methocel в селектирующей среде CHOMaster® с добавлением 4% диализованной фетальной сыворотки теленка. Вязкую смесь высевали в 24-ячеечные планшеты и через 2 недели стерильным наконечником пипетки собирали отдельные клоны и начинали новую культуру в суспензии.
Следующая стратегия культивирования была адаптирована для получения посевного материала в биореакторе. Культуру клеток, выращенную в лунках планшета для микротитрования, затем разделяли 1:3 селективной средой CHOMaster® в большие ячейки (24-ячеечные и 6-ячеечные планшеты) и затем во флаконы объемом 25 и 75 см2, после того как плотность клеток достигала примерно 4-5×105 клеток/мл.
Соблюдая постоянное отношение деления на части (1:3) и плотность клеток, при которой их делят на части в Т-флаконы, суспензионные культуры переносили во вращающиеся флаконы объемом 2 л (Integra Bioscience, Switzerland), и во время процесса роста использовали рабочий объем 1 л.
Пример 2: Определение оптимальных условий секреции/процессинга дц-уАП.
Сначала определили оптимальные условия относительно концентрации бутирата и температуры при получении в лабораторном масштабе. Предварительные эксперименты, нацеленные на оптимизацию концентрации Na-бутирата, выполняли в культуре рекомбинантной линии клеток СНО объемом 1 л (1×106 клеток/мл), чтобы добиться наилучшей продукции дц-уАП (ВММ и НММ) и наибольшей выживаемости клеток после 5 дней культивирования во вращающихся флаконах.
В таблице 1 представлены данные по жизнеспособности клеток и выходу уАП через 5 дней выращивания культуры во вращающихся флаконах объемом 1 л при 37°С и при разных концентрациях Na-бутирата. За активностью уАП следили с помощью хромогенного анализа, используя в качестве хромогенного субстрата Pefachrome® UK (54-46) (Pentapharm, LTD, Basel, Switzerland), согласно инструкциям производителя; за жизнеспособностью клеток следили каждый день и измеряли с помощью микроскопа после исключения погибших клеток методом окрашивания трипановым синим, как описано в Doyle et al. A, Griffiths, JB and Newell, DG (Eds.) (1994), в "Cell and Tissue Culture: Laboratory Procedures". John Wiley & Sons. New York.
Как показано в таблице 1, наиболее эффективная концентрация Na-бутирата в отношении выхода уАП через 5 дней культивирования во вращающихся флаконах объемом 1 л при 37°С составляет 1,2 мМ. При этой концентрации жизнеспособность клеток еще довольно высокая (76%), даже по сравнению с контролем (92%). Высокая жизнеспособность клеток предотвращает слишком большое загрязнение надосадка обломками клеток и белками, имеющими отношение к хозяину, и снижает нагрузку примесями, которые нужно будет удалять в ходе процесса очистки.
Вторую серию экспериментов выполняли при концентрации бутирата, равной 1,2 мМ, при 5-дневной ферментации, чтобы определить оптимальную температуру для культуры рекомбинантной линии клеток СНО объемом 2 л (1,5-2×106 клеток/мл) в биореакторе.
В таблице 2 показано, что 34°С является температурой, которая позволяет достичь высокой степени продуктивности уАП наряду с высокой жизнеспособностью клеток. Интересно, что когда такой же эксперимент выполнили при 32°С, активность, равная 3961 U/мл, была достигнута только через 12 дней при низкой жизнеспособности клеток (63%).
Уменьшение времени культивирования в высокой мере желательно по экономическим причинам, а также для сохранения целостности белка. В действительности желательно избегать длительной экспозиции рекомбинантного продукта с различными протеолитическими и гликолитическими ферментами, вышедшими из лизированных клеток.
Исходя из данных, показанных в таблице 2, становится ясно, что снижение температуры культивирования до 34°С наряду с добавлением Na-бутирата (1,2 мМ) в среду культивирования, обусловливает синергетическое действие, результатом которого является повышенный выход активного уАП.
На основании указанных данных можно сделать вывод, что обработка 1,2 мМ Na-бутиратом при температуре 34°С является оптимальным условием для получения активного дц-уАП и жизнеспособности клеток, в частности в течение 5-дневного периода культивирования.
Пример 3: Получение рекомбинантного дц-уАП в клетках СНО в биореакторе объемом 2 л
Инокуляцию клеток проводили в биореакторе объемом 2,4 л (Infors HT, Type Labforce, Bottmingen, Switzerland) разделением экспоненциально растущей культуры клеток. Соотношение «объем инокулята/объем свежей среды» выбирали от 1:1 до 1:5, в соответствии с общей емкостью биореактора и условиями культивирования. Выбирали рабочий объем 2 л и конечную плотность клеток инокулята 3×105 клеток/мл.
Культура, используемая для первичного инокулята, имела степень жизнеспособности не менее 95%. В ходе роста суспензионной культуры в биореакторе следили за концентрацией глюкозы, которая никогда не оставалась ниже 1 г/л в истощенной среде.
Для культуры устанавливали следующие параметры ферментации:
Температура: 37±0,5°С
рН: 7,15±0,1
рО2: 50±20%
Когда плотность клеток достигала значений около 2×106 живых клеток/мл, клетки отделяли от истощенной среды тангенциальным фильтрованием (или центрифугированием). Затем клетки ресуспендировали в биореакторе в таком же исходном объеме свежей среды CHOMaster® с добавлением бутирата натрия, чтобы достичь конечной концентрации 1,2 мМ.
Температуру культуры понижали до 34±0,5°С, а остальные параметры ферментации сохраняли такими, которые устанавливали раньше, как указано ниже:
Температура: 34±0,5°С
рН: 7,15±0,1
рО2: 50±20%
За продукцией активного у-АП (дц-уАП ВММ и НММ) следили с помощью хромогенного теста со специфичным субстратом, Pefachrome® UK. Наблюдали постепенное увеличение активности вплоть до максимальных значений до 7000 IU/мл, достигаемых через 4-5 дней ферментации. В этой точке клетки собирали и затем отбрасывали, а истощенную культуральную среду, содержащую дц-уАП, далее обрабатывали для очистки.
Средние уровни продукции, относящиеся к четырем независимым ферментациям, представлены в таблице 3:
Результаты ферментации
В биореакторе наблюдали постепенное снижение жизнеспособности клеток с первого по пятый день культивирования после добавления бутирата натрия. Минимальное значение жизнеспособности клеток при 1,2 мМ бутирата наблюдалось на пятый день культивирования, и не было значительно ниже 70%, и следовательно, было еще достаточно высоким. При указанных значениях предполагается ограниченное высвобождение деградирующих лизосомальных ферментов в среду роста.
Как показано в таблице 3, значительное увеличение экспрессии дц-уАП в процессе получения, как описано, главным образом происходит в течение последних 2/3 дней ферментации после добавления бутирата натрия. Стабильность большей части секретируемой урокиназы в истощенную среду оптимально при ограничении 2/3 днями при температуре 34°С. Указанная комбинация параметров снижает воздействие на белок деградирующей активности протеолитических и гликолитических ферментов, и обеспечивает высокое качество рекомбинантного белка для очистки. На фиг.1 показаны продукты, полученные после ферментации в отсутствии или в присутствии 1,2 мМ бутирата при температуре 34°С. При последних условиях можно выявить полное превращение оц-уАП в дц-уАП по сравнению с культурой, в которую не добавляли Na-бутират.
Пример 4: Очистка дц-уАП ВММ и НММ
Очистка дц-уАП ВММ.
Надосадок культуры клеток, выращенной в биореакторе, получали, как описано в примере 3, и подкисляли добавлением CH3COOH до рН 5,5, и очищали от остатков клеток фильтрацией через фильтр 0,45 мкм. Добавляли твин-80 до 0,01%, и надосадок наносили на колонку для ионообменной хроматографии (SP Sepharose Big Beads, Amersham-Pharmacia), предварительно уравновешенную 20 мМ буферным раствором фосфата натрия рН 6,0. Размер колонки составлял 10 см в высоту и 2,6 см в диаметре. Скорость потока в ходе нанесения и промывки составляла 10 мл/мин и во время элюирования 2 мл/мин.
После нанесения колонку сначала промывали 3 объемами буферного раствора, содержащего 20 мМ фосфата натрия, 150 мМ NaCl, рН 6,0, для того, чтобы удалить не относящиеся к урокиназе примеси, и затем еще 3 объемами буферного раствора, содержащего 20 мМ фосфат натрия, 250 мМ NaCl, рН 6,0, для того, чтобы элюировать в основном дц-уАП НММ. Последний хранили в замороженном виде для дальнейшей очистки.
Элюирование дц-уАП ВММ проводили пропусканием через ионообменную колонку буферного раствора, содержащего 20 мМ фосфат натрия, 500 мМ хлорида натрия, рН 6,0.
Полученный элюат, содержащий урокиназу (дц-уАП ВММ) доводили до рН 6,5 добавлением 1 н. NaOH. Затем его наносили на аффинную колонку с бензамидин-сефарозой 6В, предварительно уравновешенную, по меньшей мере, 2 объемами буферного раствора, содержащего 20 мМ фосфат натрия, 400 мМ NaCl, рН 6,5.
Размер колонки составлял 10 см в высоту, 2,6 см в диаметре. Скорость потока во время нанесения, промывки и элюирования составляла 2,5 мл/мин.
Затем колонку промывали 2 объемами буферного раствора, содержащего 20 мМ фосфат натрия, 400 мМ NaCl, рН 6,5, и, наконец, элюировали 2,5 объемами буферного раствора, содержащего 100 мМ ацетат натрия, 400 мМ NaCl, рН 4,0.
Колонка с бензамидином позволяла избавиться от примесей, не относящихся к урокиназе, а также от нерегистрируемых количеств, если они имеются, оц-уАП, который может присутствовать в загружаемом материале.
Фракции, содержащие дц-уАП ВММ, идентифицировали как фракции, относящиеся к относительно устойчивому и уникальному пику на хроматограмме, и объединяли.
Полученные объединенные фракции наносили на колонку для гель-фильтрации (эксклюзионная хроматография на основе разделения по размеру молекул). Колонка для гель-фильтрации имела высоту 30 см и диаметр 2,6 см, и предварительно была уравновешена буферным раствором, содержащим 5 мМ фосфат натрия, рН 4,9. Используемая скорость потока составляла примерно 3 мл/мин.
Таким образом, дц-уАП урокиназного типа ВММ элюировали в чистом виде, осуществляя элюирование буферным раствором, содержащем 5 мМ фосфат натрия, рН 4,9, как показано на фиг.1 (дорожка 2).
Наконец, урокиназа была готова для приготовления в буфере, подходящем для окончательной лиофилизации.
Очистка дц-уАП НММ.
Фракцию(ии), соответствующие дц-уАП НММ, собранные из промывки ионообменной колонки буферным раствором, содержащим 20 мМ фосфат натрия, 250 мМ NaCl, рН 6,0, объединяли и затем очищали, пропуская через аффинную хроматографическую колонку с бензамидином. Перед нанесением на указанную колонку, пул фракций доводили до рН 6,5 или 7,0 добавлением 1 н. NaOH, и колонку предварительно уравновешивали, по меньшей мере, 2 объемами буферного раствора, содержащего 20 мМ фосфат натрия, 400 мМ NaCl, рН 6,5. После загрузки колонку промывали 2 объемами буферного раствора, содержащего 20 мМ фосфат натрия, 400 мМ NaCl, рН 6,5, и наконец, элюировали 2,5 объемами буферного раствора, содержащего 100 мМ ацетат натрия, 400 мМ NaCl, рН 4,0. Фракции, содержащие дц-уАП НММ, идентифицировали как фракции, относящиеся к относительно устойчивому и уникальному пику на хроматограмме, и объединяли.
Полученные объединенные фракции наносили на колонку для гель-фильтрации (эксклюзионная хроматография на основе разделения по размеру молекул) высотой 30 см и диаметром 2,6 см, предварительно уравновешенную буферным раствором, содержащим 5 мМ фосфат натрия, рН 4,9. Используемая скорость потока составляла примерно 3 мл/мин.
Таким образом, дц-уАП урокиназного типа НММ элюировали в чистом виде, осуществляя элюирование буферным раствором, содержащем 5 мМ фосфат натрия, рН 4,9, и, наконец, урокиназа была готова для приготовления в буфере, подходящем для окончательной лиофилизации.
Пример 5: Характеристика рекомбинантного дц-уАП
Процессинг рекомбинантной молекулы.
Рекомбинантный дц-уАП ВММ характеризовали в сравнительных исследованиях с коммерческим препаратом экстрагированного дц-уАП ВММ (Ukidan®, Serono) посредством масс-спектроскопии и функциональных исследований.
Молекулярная масса.
Данные масс-спектроскопии, точность которых варьировала в пределах от +/- 50 Да до +/- 100 Да, подтвердили, что молекулярная масса соответствовала молекулярной массе, ожидаемой после правильного процессинга белка-предшественника (оц-уАП) и, в частности, как показано на фиг.2, подтвердили, что:
- рекомбинантный, полученный согласно способу изобретения, и коммерческий экстрагированный дц-уАП ВММ очень сходны по молекулярным массам, составляющим 48267 Да и 48565 Да, соответственно;
- в случае дегликозилирования две формы также имеют очень сходные молекулярные массы, 46382 (рекомбинантный) и 46313 Да (экстрагированный).
- сходным образом, анализ рекомбинантного, полученного согласно способу настоящего изобретения, и экстрагированного дц-уАП НММ показал очень близкие массы гликозилированных (33249 Да и 33189 Да, соответственно) и негликозилированных форм (31029 и 30969, соответственно).
Кроме того, масс-спектрометрический анализ MALDI-MS показал, что очищенные молекулы в основном интактны (>95%), другими словами, это подтверждает, что продукты деградации, связанные с уАП, присутствуют в несущественном количестве, и что, поэтому выбранные способы очистки не влияют на целостность рекомбинантной молекулы.
Правильный процессинг N-конца подтвердили способом деградации по Эдману очищенного дц-уАП ВММ. Как предполагалось, NH2-концевая последовательность В-цепи определена, как последовательность:
-IIGGEF-,
тогда как NH2-концевая последовательность А-цепи определена, как и ожидалось в виде последовательности:
-SNELHQ-.
Представленные данные свидетельствуют, что протеолитическое расщепление осуществляется точно и специфично в месте связи Lys158-Ile159, и Lys158 точно удаляется из оставшейся молекулы. Кроме того, анализ пептидного картирования подтвердил наличие правильных NH2- и С-концов как А-, так и В-цепи рекомбинантного дц-уАП ВММ.
Характер гликозилирования.
На очищенном рекомбинантном дц-уАП анализировали гликаны с помощью масс-спектроскопии и электрофореза углеводов с использованием флюоресценции (FACE). Оба способа выявили стабильное гликозилирование: N-гликаны рекомбинантной урокиназы состоят из фукозилированных по центральной части, двух-, трех- и четырех-усиковых сложных цепей со степенью сиалирования 80-90%. С помощью масс-спектроскопии определили сайт гликозилирования и подтвердили, что гликозилирование происходит у Asn302.
Функциональные исследования.
Также определяли биологическую активность рекомбинантного дц-уАП ВММ путем измерения следующих параметров:
- анализ Kd связывания с природным рецептором урокиназы (смотри таблицу 4);
- анализ стехиометрической ингибирующей активности ингибитора активатора плазминогена (ИАП-1), который является природным ингибитором урокиназы;
- анализ кинетики ингибирования ИАП-1 (смотри таблицу 4);
- изучение энзиматического параметра Km с хромогенным субстратом (смотри таблицу 4);
- изучение кинетики активации плазминогена;
- способность к фибринолизу (фиг.3);
- деградация фибрина.
Данные, показанные в таблице 4, свидетельствуют о значительной функциональной идентичности рекомбинантного дц-уАП ВММ, полученного согласно способу настоящего изобретения, и коммерческого экстрагированного дц-уАП ВММ. В частности, показано, что два продукта имеют:
(а) сходные константы скорости (k1) образования комплекса с ИАП-1, которые измеряли согласно Chmielweska et al. Biochem. J. 1988, 251: 327-332,
(b) сходные константы Михаэлиса-Ментена (Km) для субстрата урокиназы, которые измеряли согласно Briggs, GE and Haldane JBS, Biochem. J. 1925, 29: 338-339 и Lijnen, HR et al. Eur. J. Biochem. 1994, 224: 567-574, и
(с) сходные константы аффинности по отношению к рецептору урокиназы (Kd), которые измеряли согласно: Cubellis, M.V. et al., J. Biol. Chem., 1986, 261: 15819-15822.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ ФАРМАЦЕВТИЧЕСКИ АКТИВНЫХ РЕКОМБИНАНТНЫХ БЕЛКОВ | 2002 |
|
RU2227745C2 |
| СПОСОБ ВЫДЕЛЕНИЯ И ОЧИСТКИ РЕКОМБИНАНТНОЙ ЧЕЛОВЕЧЕСКОЙ ПРОУРОКИНАЗЫ М5 | 2013 |
|
RU2553533C2 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК рВК415, КОДИРУЮЩАЯ ПОЛИПЕПТИД РЕКОМБИНАНТНОГО ТКАНЕВОГО АКТИВАТОРА ПЛАЗМИНОГЕНА ЧЕЛОВЕКА, ЛИНИЯ КЛЕТОК Cricetulus griseus CHO 1F8 - ПРОДУЦЕНТ РЕКОМБИНАНТНОГО ТКАНЕВОГО АКТИВАТОРА ПЛАЗМИНОГЕНА ЧЕЛОВЕКА И СПОСОБ ПОЛУЧЕНИЯ И ВЫДЕЛЕНИЯ ПОЛИПЕПТИДА, ОБЛАДАЮЩЕГО АКТИВНОСТЬЮ ТКАНЕВОГО АКТИВАТОРА ПЛАЗМИНОГЕНА | 2012 |
|
RU2500817C1 |
| ДИМЕР МОЛЕКУЛЯРНОГО ВАРИАНТА АПО-ЛИПОПРОТЕИНА, СПОСОБ ЕГО ПОЛУЧЕНИЯ И ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ НА ЕГО ОСНОВЕ | 1992 |
|
RU2134696C1 |
| СПОСОБ ПОЛУЧЕНИЯ АНТИТЕЛА ИЛИ ЕГО ФРАГМЕНТА С ПОДПИТКОЙ (ВАРИАНТЫ) | 2007 |
|
RU2518289C2 |
| Способ получения рекомбинантного фолликулостимулирующего гормона человека, клеточная линия - продуцент и плазмидные экспрессионные векторы | 2019 |
|
RU2697273C1 |
| Способ получения полипептида со свойствами проурокиназы и штамм бактерий ЕSснеRIсна coLI - продуцент полипептида со свойствами проурокиназы (варианты) | 1986 |
|
SU1695827A3 |
| СПОСОБ МАССОВОГО ПРОИЗВОДСТВА МУЛЬТИМЕРНОГО МАННОЗОСВЯЗЫВАЮЩЕГО ЛЕКТИНА | 2004 |
|
RU2350654C2 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pOptiVEC-MBP-Fc, КОДИРУЮЩАЯ КОНСТАНТНЫЙ ФРАГМЕНТ ИММУНОГЛОБУЛИНА ЧЕЛОВЕКА, СЛИТНОГО С ФРАГМЕНТОМ ОСНОВНОГО БЕЛКА МИЕЛИНА, ЛИНИИ ЭУКАРИОТИЧЕСКИХ КЛЕТОК - ПРОДУЦЕНТОВ УКАЗАННОГО БЕЛКА И СПОСОБ ПОЛУЧЕНИЯ БЕЛКА MBP-Fc ДЛЯ ТЕРАПИИ РАССЕЯННОГО СКЛЕРОЗА | 2016 |
|
RU2679055C2 |
| СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО БЕЛКА, МОДИФИЦИРОВАННОГО ПРИСОЕДИНЕНИЕМ АЛЬБУМИНА ЧЕЛОВЕКА, ИЗ КУЛЬТУРАЛЬНОЙ ЖИДКОСТИ ДРОЖЖЕЙ | 2007 |
|
RU2373286C2 |
Изобретение относится к медицинской промышленности и касается способа получения рекомбинантного дц-уАП (двухцепочечного фермента урокиназы). Сущность изобретения включает способ культивирования подвергнутых генетическим манипуляциям клеток, CHO-Messi, стабильно трансфицированных кДНК препроурокиназы в среде для культивирования, содержащей алкановые кислоты или их производные при температуре в интервале от 30 до 37°С, разделения полученного дц-уАП на низкомолекулярную и высокомолекулярную формы. Изобретение также включает рекомбинантный дц-уАП, а также дц-уАП ВММ и дц-уАП НММ на разных стадиях получения. Преимущество изобретения заключается в разработке способа получения рекомбинантных форм зрелого двухцепочечного белка дц-уАП, а также его низкомолекулярной и высокомолекулярной форм. 10 н. и 9 з.п. ф-лы, 4 табл., 3 ил.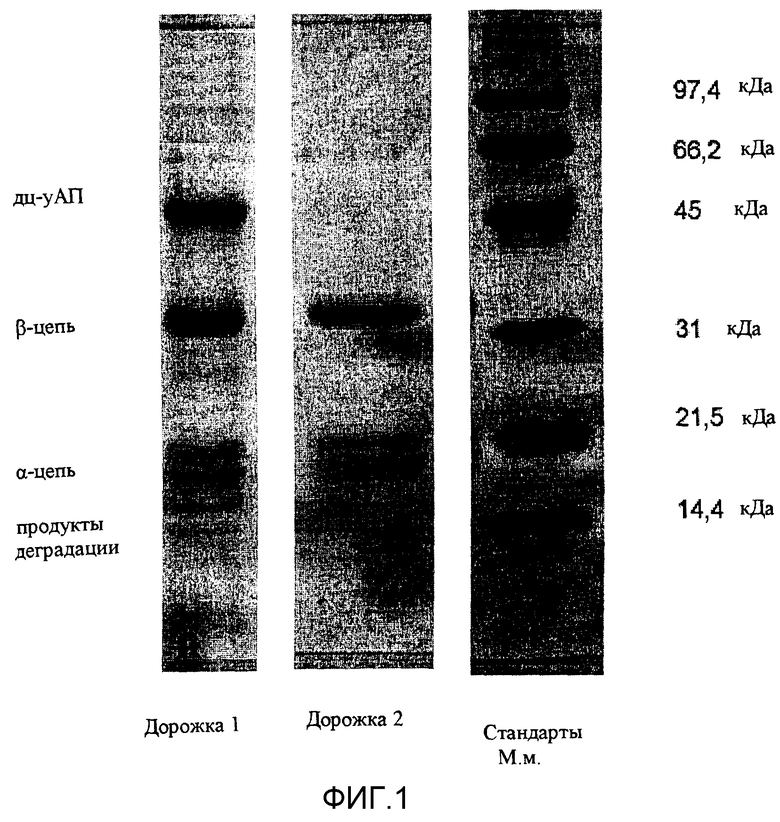
a) Культивирование подвергнутых генетическим манипуляциям клеток CHO-Messi, стабильно трансфицированных кДНК пре-проурокиназы, в среде для культивирования, содержащей алкановые кислоты или их производные, или их соли при температуре в интервале от 30 до 37°С;
b) продолжение указанного культивирования клеток в течение периода времени, по меньше мере, равного 24 ч;
c) получение надосадочной жидкости культуры клеток.
d) подкисление надосадочной жидкости культуры клеток, полученной по п.1, слабой кислотой до значений рН в пределах от 5,0 до 5,8, необязательно с добавлением неионного детергента;
e) пропускание подкисленной надосадочной жидкости через колонку с SP-сефарозой при значениях рН в пределах от 5,5 до 6,5;
f) элюирование дц-уАП НММ добавлением буферного раствора со значением рН в пределах от 5,5 до 6,5, содержащего моновалентный ион в концентрации от 200 до 300 мМ;
g) элюирование дц-уАП ВММ добавлением буферного раствора со значением рН 6-7,5, содержащего моновалентные ионы в концентрации, по меньшей мере, равной 400 мМ.
g') контактирование буферного раствора со стадии g), содержащего элюированный дц-уАП ВММ, с бензамидиновой колонкой при значениях рН в пределах от 6,2 до 6,8;
g'') элюирование дц-уАП ВММ буферным раствором при значении рН в пределах от 3,8 до 4,2, кроме того, содержащим моновалентные ионы в концентрации от 300 до 500 мМ;
g''') необязательно дальнейшее контактирование элюированного дц-уАП ВММ с колонкой для гель-фильтрации, и элюирование дц-уАП ВММ буферным раствором с низким содержанием солей при значениях рН в пределах от 4 до 7.
f) контактирование раствора, содержащего элюированный дц-уАП НММ, полученного на стадии f), с бензамидиновой колонкой при значениях рН в пределах от 6 до 8;
f) элюирование дц-уАП НММ буферным раствором со значениями рН от 3,8 до 4,2, кроме того, содержащего моновалентные ионы в концентрации в пределах от 300 до 500 мМ;
f'') кроме того, необязательно контактирование элюированного дц-уАП НММ с колонкой для гель-фильтрации и элюирование дц-уАП НММ буферным раствором с низким содержанием солей при значениях рН в пределах от 4 до 7.
| Способ контроля работы форсунок в распылительной сушилке | 1977 |
|
SU620279A1 |
| Устройство для цементирования обсадных колонн | 1980 |
|
SU866132A2 |
| Способ приготовления антикристаллизатора | 1950 |
|
SU92182A1 |
| Спеченный фрикционный материал | 1974 |
|
SU496327A1 |

