Настоящее изобретение относится к области биологии, в частности, к биологии рака; а более конкретно, к способам выявления предзлокачественных или злокачественных тканей, заключающимся, в одном своем варианте, в обнаружении потерь синдикана в тканях, а в другом варианте, в обнаружении присутствия синдикана в биологических жидкостях человека; причем указанные способы могут быть осуществлены с использованием синдикан-специфических жидкостей, антител, и кДНК-зондов для детектирования.
Молекулы клеточной поверхности, которые участвуют в специфических взаимодействиях между клеточной поверхностью и внеклеточным матриксом (ECM), вызываются матриксными рецепторами. Распознавание матрикса этими рецепторами играет важную роль в регуляции клеточной формы, пролиферации и дифференциации клеток. а поэтому, оно имеет особое значение для нормального развития органов и для сохранения структуры тканей. Известными матриксными рецепторами являются, например, семейство трансмембранных гликопротеиновых рецепторов, которые имеют общие структурные и функциональные характеристики и называются интегринами (Hynes (1987), Cell 48: 549-554), 67-кДа-гликопротеином, который связывается с BI-цепью ламинина (Graf и др. (1987) Cell 48: 989-996), и протеогликанами (PG), которые в зависимости от строения глюкозаминогликана (например, гепарансульфат) могут взаимодействовать с рядом молекул матрикса (Jalkanen, M. (1987) Med. Biol. 65: 41-47).
Адгезия клеток, их конфигурация, пролиферация и дифференциация зависят от тесных контактов между клеточной поверхностью и окружающим внеклеточным матриксом (ECM). Избирательное использование взаимодействия "лиганд-рецептор" может обеспечивать разнообразие специфических клеточно-матриксных взаимодействий in vivo, необходимых для развития органов (Fkblom и др. (1986) Ann. Rev. Cell Biol. 2: 27-47). Известно, что трансформация влияет на ответную реакцию клеток на ECM (Liotta L. (1986) Caneer Res 46: 1-7), что позволяет предположить о возможном изменении экспрессии матриксных рецепторов в злокачественных клетках (Plansfaber и Hynes (1989) Cell 56: 281-290, Cheresh и др. (1989) Cell 57: 59-69). О таких изменениях, по большей части, ничего не известно, однако, они могут играть фундаментальную роль в злокачественном развитии некоторых типов клеток.
Протеогликан клеточной поверхности эпителиальных клеток мышиной молочной железы состоит из липофильного мембранного домена (Rapraeger и Bernfreld (1983) J. Biol. Chem. 258: 3632-3636, (1985) J. Biol. Chem. 260: 4103-4109) и из взаимодействующего с матриксом эктодомена, содержащего цепи гепарансульфата и хондроитинсульфата (Rapraeger и др. (1985) J. Biol. Chem. 260: 11046-11052). Недавно осуществленное кДНК-клонирование мышиного и человеческого синдиканов подтвердило присутствие указанных функциональных доменов на коровом белке, и эти PG были названы синдиканами (Sannders и др. (1989) J. Cell Biol. 108: 1547-1556, Mali и др. (1990) J. Biol. Chem. 265: 6884-6889). Эктодомен синдикана распознается моноклональными антителами (mab) 281-2 (Jalkanen и др. (1985) J. Cell Biol. 101: 976-984, Jalkanen и др. (1987) J. Cell Biol. 105: 3087-3096), и он с высокой степенью аффинности связывается с коллагеновыми фибриллами Типа I, III и V (Кода и др. (1985) J. Biol. Chem. 260: 81567-8162) и с низкой степенью аффинности связывается с C-концевым гепарин-связывающим доменом фибронектина (Sannders и Bernfreld (1988) J. Cell Biol. 106: 423-430), тромбоспондина (Sun и др. (1989) J. Biol. Chem. 264: 2885-2889) и тенаксина (Salmivicta и др. 1991) J. Biol. Chem. 266: в печати). Это связывание с лигандом стимулирует ассоциирование мембранного домена с актин-обогащенным цитоскелетом (Rapraeger и др. (1986) J. Cell Biol. 103: 2683-2696). Однако эпителиальные клетки могут также терять эктодомен со своей поверхности вследствие протеолитического расщепления корового белка, который отделяет эктодомен от мембранного домена (Jalkanen и др. (1987) J. Cell Biol. 105: 3087-3096, Weitzhandler и др. (1988) J. Biol. Chem. 263: 6949-6952). Следовательно, синдикан может связывать эпителиальный цитоскелет с матриксом, но при этом, он может также ослаблять связывание эпителиальной клетки с матриком. При таком связывании синдикан может опосредовать сборку матрикса в клеточную структуру и воздействовать на развитие клеток. Поскольку синдикан взаимодействует, главным образом, с компонентами стромы, такими как коллагеновые фибриллы и фибронектин, то он может служить в качестве посредника между эпителиальными и мезенхимальными тканями. Такой тип коммуникации играет важную роль для дифференциации нормальных эпителиальных тканей и формирования органов.
Экспрессия синдекана в процессе развития и формирования органов протекает в морфогенетических, а не гистологических границах (Tnesleff и др. (1988) Dev. Biol. 189: 5465-572), и этот отличительный признак также подтверждает активную роль синдикана как матриксного рецептора в процессе развития тканей. В основном, синдикан локализуется на различных эпителиальных клетках (Hayashi и др. (1987) J. Histochem. Cytochem. 35: 1079-1088), но он может быть также обнаружен и на поверхностях плазматических клеток (Sanderson и Bernfield (1988) Proc. Natl. Acad. Sci. USA 85: 9562-9566). Синдикан локализуется также до уплотнения мезенхимы, сразу после образования эпителия в процессе формирования органов (Thesleff и др. (1988) Dev. Biol. 189: 565-572). Было показано, что экспрессия синдикана в мезенхиме регулируется посредством эпителиального контакта в тканях зубов (Vainio и др. (1988)) и в тканях метанефроса (Vainio и др. (1989)), и в обеих этих тканях, его экспрессия может быть коррелирована как в пространстве, так и во времени до морфогенеза указанных органов.
Краткое описание изобретения
Целью настоящего изобретения является способ выявления предзлокачественных или злокачественных тканей путем обнаружения потерь синдикана тканями.
Другой целью настоящего изобретения является способ выявления предзлокачественных или злокачественных тканей путем обнаружения присутствия синдикана в физиологических жидкостях организма.
Следующей целью настоящего изобретения является способ, который может быть легко адаптирован для выявления злокачественных или предзлокачественных тканей у человека.
Очередной целью настоящего изобретения является биохимический, иммуногистологический или молекулярно-биологический способ обнаружения экспрессии или содержания синдикана в тканях или биологических жидкостях в целях выявления злокачественной или предзлокачественной трансформации клеток.
Другой целью настоящего изобретения является способ выявления гиперпластических изменений клеток.
Следующей целью настоящего изобретения является способ обнаружения предполагаемых гиперпластических изменений клеток.
Очередной целью настоящего изобретения является способ обнаружения предполагаемых морфологических изменений в клетках.
Еще одной целью настоящего изобретения является способ количественного определения уровня синдикана в тканях или физиологических жидкостях в целях получения оценочного значения.
Короче говоря, в более широких вариантах своего осуществления, настоящее изобретение относится к способу обнаружения потенциально неблагоприятной трансформации клеток организма, который заключается в том, что определяют уровень содержания синдикана в образце биологического материала, взятого из организма, и полученное значение содержания синдикана сравнивают со стандартным показателем содержания синдикана, определенным из эталонного биологического материала, предпочтительно, того же типа.
Рассматриваемый способ может быть осуществлен различными путями, например с использованием биохимических, иммуногистологических или молекулярно-биологических методов. Хотя указанный способ может быть применен для обнаружения трансформации, такой как злокачественное или предзлокачественное состояние, в широком ряде организмом, однако, особенно успешно он может быть использован для обнаружения неблагоприятных трансформаций в клетках, в частности, в клетках человека. Примерами таких трансформаций могут служить предзлокачественные или злокачественные перерождения клеток, включая их гиперпластические и морфологические изменения.
Выявление присутствия или отсутствия синдикана может быть осуществлено на различных биологических материалах организма, таких как клетки, ткани, и биологические жидкости, включая, биологические материалы человека, в частности, сыворотка, плазма, моча, цереброспинальная жидкость, или тканевые экстракты. Определение содержания синдикана в биологических материалах может быть осуществлено различными путями, например, с использованием синдикан-специфических лигандов или биохимических детерминант, синдикан-специфических антител, и синдикан-антисмысловые мРНК или кДНК или олигонуклеотиды.
Другие отличительные признаки, цели и преимущества настоящего изобретения будут более понятны из нижеследующего подробного описания настоящего изобретения, иллюстрируемого прилагаемыми чертежами.
Фиг. 1A представляет собой график зависимости содержания синдикана на клеточной поверхности от времени, иллюстрирующий потери синдикана эпителиальными клетками мышиной молочной железы, трансформированными посредством воздействия на них тестостероном.
Фиг. 1B представляет собой график зависимости уровня мРНК-экспрессии синдикана от времени, иллюстрирующий потери синдикана эпителиальными клетками мышиной молочной железы, трансформированными посредством воздействия на них тестостероном.
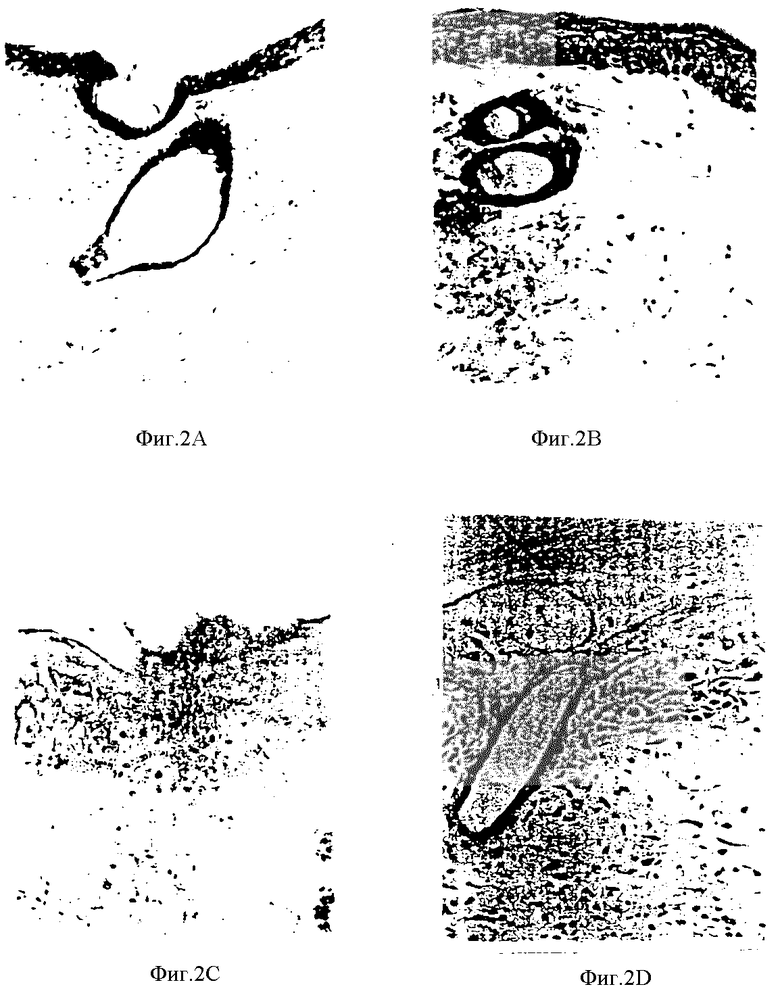
Фиг. 2A-2D представляют собой фотографии гистологических срезов мышиного биологического материала кожной ткани, иллюстрирующие течение экспрессии синдикана после УФ-облучения указанного материала.
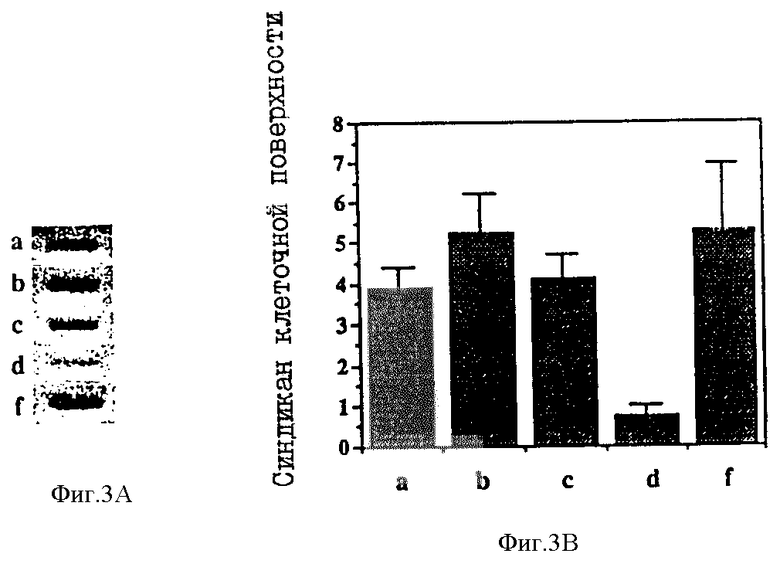
Фиг. 3A представляет собой фотографию, иллюстрирующую суспензию эпителиальных клеток мышиной молочной железы, культивированных в монослоях, один из которых был трансформирован ras-онкогеном, и
Фиг. 3B представляет собой график, иллюстрирующий содержание синдикана на поверхности клеток для монослоев, показанных на фиг. 3A.
Подробное описание предпочтительных вариантов изобретения
В основу настоящего изобретения был отчасти положен только что установленный факт, что обнаружение синдикана является ценным диагностическим критерием для оценки степени злокачественности биологических материалов, таких как эпителиальные клетки. В качестве примеров, были представлены три различных вида экспериментов по стимулированию трансформаций тканей (гормональное индуцирование, УФ-облучение, и индуцирование онкогеном), причем в каждом из этих экспериментов были обнаружены потери синдикана. Обнаружение потерь синдикана является важным показателем также и в других механизмах, приводящих к трансформации тканей, например, образование опухолей, а поэтому, настоящее изобретение не ограничивается лишь вышеприведенными примерами. Например, выявление синдикана может дать ценную информацию также относительно различных стадий пролиферации тканей (кожи, почки, головного мозга, легких, желчного пузыря), которые могут затем перейти в более злокачественную стадию. На этих так называемых предраковых стадиях, которые часто рассматриваются как дисплазии, экспрессируются различные уровни синдикана, являющиеся ценной информацией, необходимой для оценки серьезности заболевания и назначения соответствующего терапевтического лечения. Особенно важной областью обнаружения синдикана являются циркулирующие клетки плазмы. Поэтому их анализ с помощью рассматриваемого способа, в котором используются синдикан-специфические антитела, например FAS-анализ (флуоресцентный метод разделения клеток), позволяет получить важную информацию относительно патофизиологического состояния пациента с лимфомой, миеломой, лейкозом или заболеванием, характеризующимся другой пролиферативной стадией гематопоэтических клеток.
Известно также, что синдикан может "выпадать" из клеточных поверхностей (Jalkanen и др. (1987) J. Cell. Biol. 105: 3087-3096), что обычно имеет место при некоторых заболеваниях, и эта потеря синдикана клеточными поверхностями может приводить к его появлению в биожидкостях организма. Поэтому, в соответствии с настоящим изобретением, измерение количества синдикана в сыворотке, плазме, моче, цереброспинальной жидкости, или других тканевых экстрактах имеет важное диагностическое значение, и позволяет определить степень деструкции ткани и ее морфологические изменения по сравнению с нормальной тканью.
Как было упомянуто ранее, синдикан представляет собой протеогликан клеточной поверхности, состоящий из ковалентно связанных цепей гликозаминогликана (GAG) и корового белка, заключенного внутри мембраны (Saunders и др. (1988) J. Cell Biol. 108: 1547-1556). GAG-цепи синдикана, как известно, принадлежат к группам гепарансульфата и хондроитинсульфата (Rapraeger и др. (1985) J. Biol. Chem. 260: 11026-11052), и эти группы не ограничиваются лишь синдиканом. Коровый белок синдикана является уникальным и определяет эту молекулу из ряда подобных ей протеогликанов клеточной поверхности (Mali и др. (1990) J. Biol. Chem. 265: 6884-6889). Впервые был выделен мышиный синдикан, который чаще всего выявляется с помощью моноклонального антитела (MnAb 281-2) против корового белка синдикана (Jalkanen и др. (1985) J. Cell Biol. 101: 976-984).
Для более наглядного объяснения настоящего изобретения была проанализирована экспрессия синдикана в связи с индуцированием злокачественного фенотипа в трех различных независимых биологических моделях, каждая из которых приводит к злокачественному перерождению эпителиальных клеток. Во всех этих моделях, падение уровня экспрессии синдикана наблюдалось одновременно со злокачественной трансформацией. В целях иллюстрации настоящего изобретения, эти результаты вкратце представлены для следующих моделей.
В первой модели была использована линия клеток опухоли молочной железы мышей Shionеgi 115 (S115). Эти клетки представляют собой прекрасную модель для исследования трансформированного фенотипа, поскольку они являются восприимчивыми к андрогенам при повышенной скорости роста (Kind и др. (1976) J. Steroid Biochem 7: 869-873) и при морфологическом изменении ткани от эпителия до фибробластов (Yates и King (1981) Cancer Res. 41: 258-262). В присутствии андрогена, клетки легко прилипают к субстрату и приобретают способность к росту в суспензии, не связываясь с матриксом, тогда, как в отсутствие андрогена, они тесно связываются с матриксом, и полностью теряют способность к росту в суспензии (Yates и King (1981) Cancer Res. 41: 258-262). Причины такого морфологического изменения и безьякорного роста клеток не совсем ясны, однако для объяснения указанных фактов было предложено несколько механизмов, например, изменения в строении цитоскелета (Conchmon и др. (1981) Cancer Res. 41: 263-269) и в паракринных механизмах Darbre и King (1988) ln Breast cancer: Cellular Molecular Biologey, изд. M.E. Lippman и R.B. Dickson, Academic Publishers, стр. 307-341).
Обработанные тестостероном клетки S115 обнаруживают в высокой степени RGDS-зависимое связывание с фибронектином (FN), но не обнаруживают связывание с гепарин-связывающим доменом FN, что свидетельствует об экспрессии этими клетками интегрин-подобных молекул. И напротив, клетки S115, культицированные без тестостерона, обнаруживают эпителиальную морфологию и связывание с гепарин-связывающим доменом FN, на что указывает изменение экспрессии синдикана в обработанных гормоном клетках S115. В обработанных гормоном клетках S115 обнаруживалось снижение как количества матрикс-связывающего эктодоменаа синдикана (при количественной оценке с помощью радиоиммуноанализа и Вестернблот-анализза), так и количества мРНК синдикана (2,6 кв). Добавление антиандрогена ципротеронацетата в культуральную среду вызывало действие на мРНК синдикана, противоположное действию тестостерона. Таким образом, инактивация гена синдикана и последующее подавление экспрессии синдикана связаны с изменением адгезивных свойств, исчезновением эпителиального фенотипа, и, с другой стороны, с появлением трансформированного фенотипа в обработанных гормоном клетках S115.
Примеры полученных выше результатов представлены на фиг. 1A и 1B. На фиг. 1A изображен график зависимости содержания синдикана клеточных поверхностей от времени, иллюстрирующий потери синдикана эпителиальными клетками мышиной молочной железы S115, трансформированными посредством их обработки тестостероном, а на фиг. 1B изображен график зависимости уровня экспрессии мРНК синдикана от времени, иллюстрирующий потери синдикана эпителиальными клетками мышиной молочной железы, трансформированными путем их обработки тестостероном. Из фиг 1A, иллюстрирующего содержание белка, и фиг, 1B, иллюстрирующего уровень мРНУ, видно, что после обработки клеток наблюдается злокачественное развитие и потеря уровня экспрессии синдикана.
Как было упомянуто выше, добавление андрогена к S115-культурам приводит к подавлению экспрессии синдикана вследствие инактивации гена синдикана. Такое подавление тесно связано с приобретением клетками способности к безьякорному росту, а также с потерей эпителиального фенотипа, и является, поэтому, одной из причин, приводящих к развитию клеток S115 по типу трансформированных клеток в присутствии андрогена.
Во второй модели, проводили исследование на иммунореактивность к синдикану с использованием лысых (hr/hr) мышей, обработанных УФ-A- и УФ-B-облучением. Положительное окрашивание наблюдалось на поверхности нормальных эпидермальных клеток, а также в дермальных абортивных кистах волосяных фолликулов, характерных для этого типа мышей. Реакция немедленного типа к УФ-облучению, указывающая на гиперпластический эпидермис с легкой клеточной атипией, была также положительной, хотя, при этом, и наблюдалось более слабое окрашивание поверхностей эпидермальных клеток. Образцы с ярко выраженной дисплазией обнаруживали слабое окрашивание в слое гранулоцитов, тогда, как слой базальных клеток дал отрицательную реакцию. В папилломах и в единичной кератоакантоме, иммунореактивность на синдикан наблюдалась в доброкачественных гиперпластических эпидермальных клетках, а также при пролиферации эпидермальных клеток роговой кисты. Злокачественное перерождение, наблюдаемое в виде образования плоскоклеточного рака или саркомы, однозначно ассоциировалось со сниженнием синдикан-окрашивания. Эти результаты согласуются с высказанными ранее предположениями относительно того, что снижение уровня экспрессии синдикана ассоциируется со злокачественной трансформацией культивированных эпителиальных клеток, а также важной роли синдикана в сохранении структуры нормальных тканей и характера дифференциации кожной ткани.
Примеры описанных экспериментов по окрашиванию показаны на фиг. 2A-2D. На этих чертежах представлены фотографии гистологических срезов мышиного биоматериала кожной ткани, иллюстрирующие ход экспрессии синдикана после облучения материала УФ-светом. На фиг. 2A показан материал нормальной кожи, и на фиг. 2B и 2C показаны разные стадии гиперплазии, причем, на фиг. 2C показана гистология отчасти злокачественной ткани. Эти чертежи иллюстрируют предзлокачественную гиперплазию, индуцированную УФ-облучением, с постепенным снижением уровня экспрессии синдикана.
Из результатов, показанных выше, очевидно, что эпителиальные клетки in vivo, имеющие нормальную окружающую среду, обнаруживают снижение уровня экспрессии синдикана в процессе злокачественного перерождения, что опять же свидетельствует о том, что экспрессия синдикана необходима для сохранения нормальной морфологии клетки и целостности ткани, а также о том, что снижение экспрессии синдикана может быть связано со значительной дисплазией и озлокачествованием эпителиальных клеток.
В третьей модели использовали линию эпителиальных клеток мышиной молочной железы, трансфецированных c-Ha-ras-геном с точечной мутацией. Ген c-Ha-ras находится под контролем MMTV-LTR-промотора в указанных NOG-8ras-клетках, а поэтому его экспрессия может регулироваться дексаметазоном (Ciardiallo и др. 1989). В том случае, если эти клетки экспрессировали ген c-Ha-ras, то они образовывали бляшки на чашках с клеточной культурой и колонии в суспензии. Клетки NOG-8ras, культивированные на чашках, экспрессировали синдикан в таких же количествах, что и нормальные клетки в состоянии субсплошности. Однако экспрессия синдикана значительно снижалась в том случае, если клетки обнаруживали трансформацию фенотипа, т.е., если наблюдались бляшки на чашках или рост клеток в виде колонии в суспензии. При этом, уровень мРНК синдикана оставался неизменным в этих различных способах культивирования. Таким образом, в этих клетках наблюдалось посттранскрипциональное регулирование синдикана клеточной поверхности. Полученные результаты показывают, что во всех экспериментах наблюдается снижение уровня экспрессии синдикана в процессе трансформации клеток, однако способ регулирования уровня синдикана в различных клетках может варьироваться в зависимости от типа трансформированной клетки.
Примеры онкоген-индуцированного снижения уровня экспрессии синдикана показаны на фиг. 3A и 3B.
На фиг. 3A представлена фотография, иллюстрирующая суспензии эпителиальных клеток мышиной молочной железы, культивированных в монослоях, где один слой был трансформирован ras-онкогеном, а на фиг. 3B показан график, иллюстрирующий содержание синдикана клеточной поверхности для монослоев, показанных на фиг. 3A. Для каждых из эпителиальных клеток, культивированных в монослоях a, b, c и f, а также после ras-индуцирования в суспензии d, проводили количественную оценку экспрессии синдикана. Эта оценка показала потерю синдикана из ras-онкоген-трансформированных мышиных эпителиальных клеток.
Как показано выше, онкоген-индуцированная злокачественная трансформация также приводит к потере синдикана с поверхности эпителиальных клеток, что свидетельствует о том, что указанная потеря синдикана является общим отличительным признаком злокачественной трансформации, и что обнаружение такой потери синдикана является ценным диагностическим средством для определения типа ракового перерождения и степени его тяжести.
В соответствии со способом настоящего изобретения, предпочтительно использовать три метода определения содержания синдикана, а именно, биохимический, иммунохимический, и молекулярно-биологический. Первый метод основан на биохимическом распознавании синдикана, который может быть частью корового белка или ковалентно связанных GAG-цепей. Было показано, что синдикан может иметь специфические GAG-цепи, которые могут обеспечивать взаимодействие для синдикана (Elenins и др. (1990) J. Biol. Chem. 265: 17837-17943, Salmivirta и др. (1991), J. Biol. Chem. 266: в печати). Поэтому, синдикан может быть обнаружен путем использования соответствующего синдиканн-специфического лиганда. Однако, в целях настоящего изобретения, могут быть использованы и другие способы, описанные ниже, и предусматривающие применение синдикан-специфических антител и кДНК.
Путем клонирования человеческого синдикана (Mali и др. 1990 J. Biol. Chem, 265: 6884-6889) или его выделения из человеческого материала, такого как линия клеток молочной железы человека HBL-100 (Elenins и др. (1990) J. Biol. Chem, 265: 17837-17843), могут быть продуцированы антитела против синдикана человека. Это может быть осуществлено путем иммунизации животных для выработки у них поликлональных или моноклональных антител, либо путем обработки клеток in vitro для продуцирования моноклонального антитела с выделением интактного синдикана человека, или его фрагментов, синтетических пептидов или гибридных белков, предсказанных с помощью кДНК-клона человека. При осуществлении настоящего изобретения, для определения содержания синдикана могут быть использованы стандартные методы молекулярной биологии, микробиологии, иммунологии и методы рекомбинантных ДНК. Все эти методы подробно описаны в литературе. См., например, Current Protocols in Immunology (1990) by Greene Publishing Associates and John Wiley & Sons, Inc.
Экспрессия синдикана в тканях может быть исследована с помощью классических иммуногистологических методов, и с использованием синдикан-специфических антител, например, как показано в модели, проиллюстрированной на фиг. 2A-2D. В этих методах, специфическое распознавание осуществляется первичным антителом (поликлональным или моноклональным), но при этом может быть использована вторичная система обнаружения, в виде флуоресцентной метки, фермента, или другого конъюгированного второго антитела. В результате применения указанной техники получают иммуногистологическое окрашивание срезов тканей для патологического исследования. Эти ткани могут быть также экстрагированы, например, мочой или нейтральным детергентом, в целях высвобождения синдикана для проведения Вестерн-блоттинга или дот-слот-анализа (Jalkanen и др. (1985) J. Cell Biol. 105: 976-985, Jalkanen и др. (1987) J. Cell Biol. 105: 3087-3096). В этом способе, который основан на использовании катионных твердых фаз, количественная оценка синдикана может быть осуществлена с использованием выделенного синдикана в качестве стандарта. Этот способ может быть также применен для исследования биологических жидкостей организма. В случае использования образцов этих жидкостей, для установления молярной концентрации синдикана могут быть использованы стандартные величины содержания синдикана в различных физиологических жидкостях, таких как сыворотка, плазма, моча, цереброспинальная жидкость, и т.п. Нормальные количества синдикана могут быть установлены путем получения величин его содержания у здоровых волонтеров и сравнения этих величин с величинами, полученными от пациентов с различными заболеваниями и нарушениями.
Изменения в экспрессии синдикана могут быть также обнаружены с использованием кДНК синдикана, например, кДНК синдикана человека. Хотя синдиканы разных видов мало отличаются друг от друга, однако их структурные различия на нуклеотидном уровне являются достаточно большими, и не позволяют использовать мышиный синдикан для обнаружения мРНК человеческого синдикана. Вообще говоря, мРНК-уровни данного генного продукта могут быть определены исходя из специфического взаимодействия смысловой и антисмысловой форм тех же самых нуклеотидных цепей, а в данном случае, цепей синдикана. Указанная процедура может быть осуществлена с помощью in situ-техники для образцов ткани, мРНК-протективного анализа на выделенную РНК, или праймер-направленной полимеризации синдикан-специфических нуклеотидных цепей. Все указанные методы подробно описаны в литературе, например, в вышеупомянутых Current Protocols in Molecular Biology.
Конкретные методы, используемые в настоящем изобретении, представлены в приведенных ниже примерах. При этом, следует отметить, что эти примеры приводятся лишь в иллюстративных целях, и не должны рассматриваться как некое ограничение объема изобретения.
Пример I
Локализация синдикана в трансформированных эпителиальных клетках.
Клетки (штамм Shionogi S115) опухоли мышиной молочной железы культивировали традиционным способом в DMEM (модифицированная по способу Дульбекко среда Игла), дополненной 5% инактивированной нагреванием околоплодной сывороткой теленка (i-FCS), пируватом (1 мМ), глутамином (1 мМ), пенициллином (100 МЕ/мл), стрептомицином (100 мкг/мл) и тестостероном (10 нМ). Для исследования влияния обработки гормоном, использовали ростовую среду, дополненную обработанной 4% декстран-нагруженной угольной пылью плодной сывороткой теленка (DC-FCS), и содержащую, либо не содержащую 10 нМ тестостерона и/или 1 мкМ ципротерона ацетата. Для этих экспериментов, клетки пассировали с плотностью 10000 кл/см2 на чашки для тканевых культур Nunc. Затем клетки лизировали в 10 мМ Hepes, 1,5 mM MgCl2, содержащего Zodoglobin (Conlter Elpctronics, Ltd), а высвобожденные ядра суспендировали в изотоне (Conlter), и подсчитывали на цитомере Conlter.
Клетки S115 окрашивали на 4 день культивирования целых клеток, как описано Jalkanen и др. ((1985) J. Cell Biol. 101: 876-984). Протеогликан клеточной поверхности, т. е., синдикан, подвергали иммуноанализу для определения его локализации с использованием мышиного моноклонального антитела 281-2 против корового белка протеогликана (Jalkanen и др. (1985) J. Cell Biol. 10: 976-984), а указанные окрашивания контролировали с использованием другого моноклонального антитела Mel-14 (IgG 2a), специфичного к "хоминг"-рецептору лимфоцитов (Gallatin и др. (1981) Nature 304: 30-34). Обнаружение иммобилизованных мышиных антител осуществляли с помощью кроличьего антикроличьего ФИТЦ-коньюгата (Janssen Biochimica).
Пример II
Обнаружение изменения экспрессии синдикана при гормон-индуцированной трансформации эпителиальных клеток
Для количественной оценки синдикана на поверхностях клеток S115, клеточные монослои промывали несколько раз холодным фосфатно-буферным раствором (PBS), и с использованием бычьего панкреатического трипсина (Sigma, Type 111, 20 мкг/мл) выделяли эктодомен молекулы путем 10-минутного инкубирования на льду, как описано Rapraeger и Bernfielg (1983) J. Biol. Chem. 258: 3632-3636, (1985) J. Biol. Chem. 260: 4103-4109). После инактивации трипсиновым ингибитором (100 мкг/мл), клетки центрифугировали, супернатант отделяли для количественной оценки эктодомена, а клетки суспендировали в изотоне для подсчета. Моноклональное антитело метили радиоактивным иодом с использованием хлораминового метода окисления (Stahle и др. (1983) Meth. Enzymol 92: 242-253) до получения специфической активности 14,1•106 отсчетов в мин/мкг. Для проведения анализа, супернатанты от 2•205 клеток и очищенный контрольный синдикан из клеток NM и MG загружали в катионную найлоновую мембрану (Zeto-Probe, Riolag) в Minifolg-слот-аппарате (Schleicher & Scnuell), как описано Jalkanen и др. ((1987) J. Cell Biol. 105: 3087-3096). Затем мембрану вынимали из слот-аппарата и инкубировали при комнатной температуре один час в PBS, дополненном 10% околоплодной сывороткой теленка (FCS) для блокирования мембраны. После этого, мембрану инкубировали в течение ночи при +4oC в PBS, содержащем 125I-меченные антитела 281-2 (10000 отсчетов в мин/мл). После четырехкратного промывания фильтра фосфатно-буферным раствором, фильтр экспонировали с рентгеновской пленкой (Kodak X-Omat) для визуализации связанных 281-2. Для количественной оценки, каждую ячейку анализировали с помощью усовершенствованного лазерного денситометра (LKB Ultrosran XL), и результаты сравнивали с известным количеством синдикана из клеток NM и MG.
Трипсин-секретированные эктодомены из клеток S115 осаждали этанолом и фракционировали на ДСН-ПАА1-градиентном (4-10%) геле (O'-Farrell (1975) J. Biol. Chem. 250: 4007-4021). После электрофореза образцы трансформировали на мембране (Zeto-Probe) с использованием аппарата для электроблоттинга 2005 Transphor (LKB), как описано в литературе (Jalkanen и др. (1985) J. Cell Biol. 101: 976-894, Rapraeger и др. (1985) J. Biol. Chem. 260: 11046-11052) и мембрану экспонировали меченными 281-2, как описано выше. Для сравнения может быть снова использован эктодомен синдикана из клеток NM и MG.
Пример III
Обнаружение изменения экспрессии гена синдикана в трансформированных эпителиальных клетках
Полную PHK выделяли с использованием 4 М гуанидинизотиоцианата и C CI-осаждением, как описано Chirgwin и др. (1979) (Biochem. 18: 5294-5299). PHK-аликвоты (15 мкг) фракционировали на формальдегид-агароном геле (1%) и переносили на мембрану Gene Screen Plus. Гибридизацию с мульти-праймер (Amersham)-меченной вставкой PM-3-кДНК-зонда для синдикана (Saunders и др. (1989) J. Cell Biol. 108: 1547-1556) осуществляли в условиях, оговоренных изготовителем мембраны (New Englang Nuclear). Иммобилизованный зонд визуализировали путем экспонирования мембраны с рентгеновской пленкой (Kadak X-Omat) при 70oC, а количественную оценку 2,6 кв-мPHK проводили с помощью денситометрического анализа, описанного выше. Затем устанавливали корреляцию мPHK синдикана с рибосомной PHK, как описано Denis и др. ((1988) Nicl. Acid. Hes. 16: 2354-2359). Специфичность суппрессии синдикана, кроме того, исследовали путем зондирования тех же самых образцов определенной дозой глицеральдегид-3-фосфат-дегидрогеназы (GAPDH) (Fort и др. (1985) Nucl. Acid. Res. 13: 1431-1442) и мышинным-α--актином (Minty и др. (1981) J. Biol. Chem. 256: 1008-1014).
кPHK-гибридизацию in situ для парафиновых срезов осуществляли в соответствии с методом Wilkinson и др. ((1989) Develop. 105: 131-136). Sac I-Kpn I-фрагмент (535 п. с. ) из парциального кДНК-клона для мышиного синдикана (PM-4) (Saunders и др. (1989) J. Cell Biol. 108: 1547-1556) субклонировали (Vainio и др. (1991) Development, находится в процессе разработки) в рибозонд-вектор pGEM-4Z (Promega, Madison, W1). Клонированную плазмиду, содержащую вставку, линеаризовали с использованием ферментов EcoRI или Hind 111, и антисмысловые или смысловые транскрипты продуцировали из комплементарных нитей с использованием полимераз Т7 или SP6 в присутствии 35S-UTP (Amersham, Willshire, UK), соответственно. Максимальную длину транскриптов уменьшали до < 200 п. о. путем щелочного гидролиза, и фракции с наиболее высокой специфической активностью собирали, осаждали и солюбилизировали в гидридизационном буфере. Предварительно обработанные предметные стекла гибридизировали в течение ночи при 50oC, а процедуры для устранения неспецифического связывания зонда (включая промывки с высокой жесткостью и обработку рибонуклеазой A) и авторадиографию осуществляли как описано в литературе (Wiklinson и др. (1989) Development 105: 131-136).
Пример IV
Обнаружение изменения экспрессии синдикана в срезах тканей, происходящих от опухолей
Опухоли продуцировали у слегка пигментированных бесшерстных мышей-самцов штамма hr/hr C3H/Tif (Bomholdgaard, Дания) путем УФ-А и УФ-В-облучения животных, как описано Talve и др. ((1990) Photodermatol. Photoimmunol. Photomed. 7: 109-115). В течение периода 12 месяцев наблюдалось образование всего: 83 папиллом, 2 кератоакантомы, 4 плоскоклеточных карциномы, одной карциносаркомы, и 11 сарком, причем повышенное опухолеобразование ассоциировалось с высоким УФ-А (582 Дж/см2) в сочетании с высоким УФ-В (эритемально эффективным = EE) (1,0 Дж/см2) и с высоким УФ-В (EE) (0,8 Дж/см2) (78 по сравнению с 28 опухолями). Образцы брали от опухолей по грубо визуальным оценкам, а также от нормальной на вид ткани кожи, подверженной УФ-облучению. Часть образцов фиксировали в 10% забуфериренном формалине, заливали парафином, делали срезы, и традиционно окрашивали гематоксилином и эозином. Оставшиеся образцы замораживали в азоте, и приготавливали срезы. Степень поражения классифицировали в соответствии со стандартными гистопатологическими критериями. В некоторых случаях также использовали окрашивание с помощью Шифф-реакции (PAS) в целях визуализации нижних мембран, а также с помощью трихромных красителей Herovic, Welgert, Mosson и красителя Gamori для коллагеновых типов, либо с помощью других красителей, а затем анализировали с использованием электронного микроскопа, если это было необходимо.
Для иммуногистохимической локализации синдикана использовали мышиное моноклональное антитело 281-2. Это антитело является специфическим для корового белка мышиного эктодомена (18) синдикана. Для обнаружения иммобилизованных 281-2 использовали иммунопероксидазную технику с авидин-биотиновым усилением, описанную Hsu и др. ((1981) J. Histochem. Cytochem. 29: 577-580). После депарафинизации и повторной гидратации тканевых срезов, активность эндогенной пероксидазы блокировали путем инкубирования предметных стекол в 100% метаноле, содержащем 3% перекись водорода, в течение 30 минут. Затем срезы инкубировали с 2% нормальной козьей сывороткой Vector Laboratories Inc. Burlingame CA) в Трис-забуфериренном физиологичсеском растворе (pH 7,4) (TBS) в течение 30 минут при комнатной температуре для минимизации неспецифического окрашивания. После этого срезы покрывали первым антителом 281-2 при концентрации белка 2 мкг/мл в 1% (масс./об.) BSA-TBS, и инкубировали в течение ночи при 4oC. Затем предметные стекла инкубировали с биотинилированными козьими антикроличьими иммуноглобулинами (lgG) (Jaskson Immunoreslarch Laboratories, Inc. West Baltimore Пенсильвания) в течение 30 минут при комнатной температуре при разведении 1:1000 в 21% BATBS, а после этого инкубировали с авидинбиотин-пероксидазным комплексом (Vectastain Kit, Vector Laboratores, Burlingame, CA) в течение 30 минут, при комнатной температуре. После промывки, пероксидазная активность явно обнаруживалась при инкубировании предметных стекол с 0,5 мг/мл 3,3-диаминобензидина тетрагидрохлорида (DAB, Polysciences, Inc. Нортхемптон, Англия) в TBS, содержащем 0,68 мг/мл имидазола и 0,01% перекиси водорода в течение 5 минут в темном помещении. Затем предметные стекла подвергали контрастному окрашиванию с использованием гематоксилина Мейера, и заключали в заливочную среду Depex (BDH Limited Pool, Англия). В промежутках между всеми стадиями, предметные стекла три раза промывали Трис-забуфуриренным раствором (TBS). Для контрольных срезов, использовали нормальные мышиные lgG (Sigma, St. Louis MO) при концентрации 2 мкг/мл. Несколько замороженных срезов для каждого типа опухоли также окрашивали, что в результате давало идентичную иммунореактивность, как и в случае залитых парафином тканевых срезов. Изменения концентрации первого антитела и времени инкубации не оказывали значительного влияния на характер окрашивания.
Пример V
Обнаружение изменения экспрессии синдикана в процессе ras-онкоген-индуцированной трансформации
Клетки NOG-8 представляют собой субклон линии нормальных эпителиальных клеток мышиной молочной железы NM и MG, а NOG - ras - клетки представляют собой линию клеток NOG-8, которые были трафецированы плазмидов, содержащей глюкокортиконд-индуцируемый промотор MMTV-LTR, связанный с точечно-мутированным c-Ha-ras-геном. Клетки культивировали в RPMI-1640-среде для выращивания клеток (Gibco), дополнительной обработанной 5% декстран-нагруженной угольной пылью околоплодной сывороткой теленка (DCC-FCS), глутамином, пенициллином (100 ME/мл), и стрептомицином (100 мкг/мл). В этих клетках, экспрессия c-Ha-ras может быть максимально индуцирована путем добавления в культуральную среду 1 мкМ дексаметазона (Sigma). DCC-обаботку использовали для удаления лишних стероидов из сыворотки, которые могут повлиять на экспрессию трансфецированного гена. Для суспензионных культур, бактериологические чашки покрывали 1% агаром (Sigma) в RFMI-1640 для предупреждения возможной адгезии клеток с субстратом. Клетки засевали с плотностью 1 • 105/мл. Число клеток определяли после обработки трипсином на счетчике для подсчитывания клеток (Conlter Electronoc, Hialeah, FL). PM-4 представляет собой кДНК-зонд для синдикана, впервые клонированного Saunders и др. (1989) J. Cell Biol. 108: 1547-1556. От нормальных клеток мышиной молочной железы обнаруживали две полосы 2,6 кв и 3,4 кв.
Для получения РНК, клеточную культуру два раза промывали холодным фосфатно-буферным раствором (PBS), а затем солюбилизировали в 5 М GIT-буфере (4М гуанидинизотиосианата в 5 мМ цитрата натрия (pH 7,0), 0,1 М β-меркаптоэтанола и 0,5% N-лаурилсакрозина). Экстрацию полной РНК осуществляли путем центрифугирования в градиенте плотности хлористого цезия, как описано Chirgwin и др. (1979) Biochem 18: 5294-5299. Для Нозерн-блок-анализа, РНК-образцы выделяли с помощью электрофореза в 1% формальдегид-агарозном геле. Эти гели блотировали на гибридизационной мембране. Фильтры предварительно гибридизировали в 1 М NaCl, 1% додецилсульфате натрия, в 10% сульфате декстрана, растворе 5 • Денхардта, 100 мкг/мл ДНК спермы лосося, 50% формамида, при 42oC. Для гибридизации добавляли мультипраймер-меченные зонды PM-4 или BS-9. При этом использовали 32 P-изотоп (Amersham). Фильтры промывали при 65oC (для PM-4), и при 6oC (для BS-9 в 2 • SSC, 1% додецилсульфате натрия, и подвергали авторадиографии на рентгеновской пленке Kodax X-Omat или Fnji.
Для количественной оценки экспрессии синдикана клеточных поверхностей, клетки на чашках Петри 3 раза промывали охлажденным льдом фосфатно-буферным раствором. Затем чашки инкубировали в 0,5 мМ К-ЭДТК в PBS в течение 10 минут при +4oC. Затем добавляли трипсин (Sigma, T-8128) до конечной концентрации 20 мкг/мл, и инкубировали 10 минут при +4oC. Трипсин способствует высвобождению эктодомена синдикана в среду (Rapraeger & Bernfield (1985) J. Biol. Chem. 260: 4103-4109). Затем клетки соскребали резиновым скребком, либо колонии суспендировали, и добавляли трипсиновый ингибитор (Sigma, T-9128) до конечной концентрации 100 мкг/мл. Клетки центрифугировали (5 мин 100 • г), супернатант собирали, и число клеток подсчитывали с помощью счетчика Conlter. Клетки в суспензии собирали в пробирку, и колонии оставляли на 5 минут для седиментации, после чего супернатант удаляли, как и в случае культур на чашках Петри. Колонии 3 раза промывали охлажденным льдом PBS и обрабатывали трипсином.
Соответствующее количество супернатанта, которое соответствует 2 • 10 клеткам, блокировали на катионной найлоновой мембране (Zeta Probe, Bia-Rad). Мембрану предварительно инкубировали в буфере для инкубации (10% околоплодная сыворотка теленка, 1 мМ азида натрия, PBS) в течение 1 часа при комнатной температуре. Радиоактивно меченные (хлораминовый метод) MnAb 281-2 (Jalkanen и др. (1987) J. Cell Biol. 105: 3087-3096), распознающие эктодомен синдикана, добавляли к инкубированному буферу до конечной концентрации 10 000 СИР/мл и инкубировали в течение ночи при +4oC. Затем фильтр 10 минут промывали фосфатносолевым буфером при комнатной температуре. Промывание повторяли 10 раз, после чего проводили авторадиографию на рентгеновской пленке. Авторадиограмму анализировали с помощью денситометра (Gelscan XL Ultroscan, LKB) и программного обеспечения (Gelscom SL 2400) (LKB), и стандартизировали выделенным эктодоменом (Jalkanen и др. (1987) J. Cell Biol, 105: 3087-1096)
На основании вышеуказанного можно сделать вывод, что настоящее изобретение позволяет обнаруживать потенциально неблагоприятную трансформацию клеток путем оценки экспрессии синдикана и его количества, например, в тканях и в различных физиологических жидкостях. Указанный способ может быть осуществлен биохимическими методами, иммунногистохимическими методами или методами молекулярной биологии с использованием, например, синдиканспецифических лигандов, антител (поликлональных или моноклональных) и кДНК. Целью описанного способа является выявление злокачественных изменений эпителиальных клеток, в котором в процессе злокачественной трансформации клетки наблюдается значительное снижение уровня экспрессии синдикана. Рассматриваемый способ также включает в себя анализ экспрессии синдикана при гиперплазии, поскольку величина, характеризующая эту экспрессию, позволяет оценить степень опасности указанной предзлокачественной стадии. Анализы экспрессии синдикана представляют особую ценность для классификации различных предзлокачественных пролиферативных стадий дисплазии.
Изобретение относится к биологии, в частности к биологии рака. Определяют содержание синдикана в клетках, присутствующих в первом образце жидкости или ткани человека. Причем указанный первый образец берут из жидкости или ткани человека, в которых, как подозревается, имеются злокачественные клетки, предзлокачественные клетки, гиперпластические клетки или клетки, имеющие морфологические изменения по сравнению с нормальными клетками. Сравнивают определенное содержание синдикана в первом образце с уровнями синдикана, полученными от второго образца биологического материала. Причем указанный второй образец биологического материала берут из жидкости или ткани того же типа, что и первый образец биологического материала, за тем исключением, что второй образец содержит нормальные или менее трансформированные клетки, чем клетки первого образца. И обнаружение наличия указанных злокачественных или предзлокачественных изменений определяют на основании того, что в указанном первом образце биологического материала присутствует меньшее количество синдикана, чем во втором образце биологического материала. При этом указанным изменением в клетках может являться злокачественная трансформация, или предзлокачественная трансформация, или гиперплазия, или морфологическое изменение. В качестве биологического материала используют физиологическую жидкость: плазму, сыворотку, мочу или цереброспинальную жидкость. Способ информативен. 17 з.п.ф-лы, 3 ил.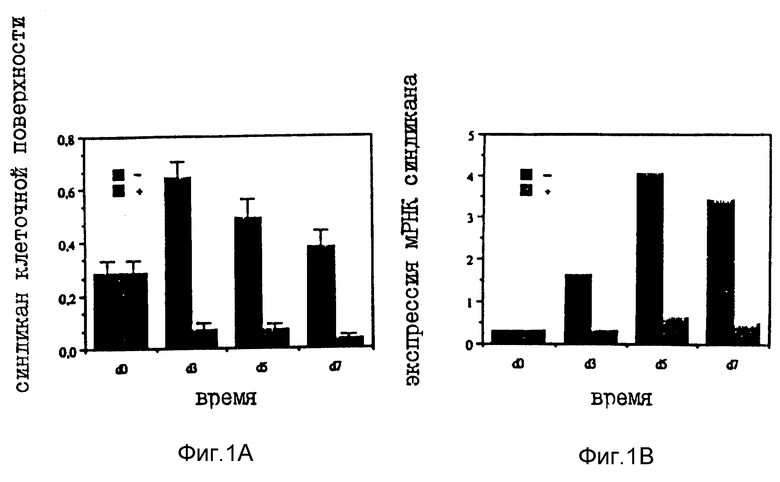
Приоритет по пунктам:
15.01.91 - по пп.1 - 18;
01.07.91 - по пп.1 - 18.
| Измерительный преобразователь температуры с частотным выходом | 1985 |
|
SU1278623A1 |
| Источник вторичного электропитания | 1985 |
|
SU1293802A1 |
| JP 01338686 A 28.12.89 | |||
| US 4698298 A 06.10.87 | |||
| US 5310653 A 10.05.94 | |||
| Автоматический огнетушитель | 0 |
|
SU92A1 |
| СПОСОБ ИММУНОЛОГИЧЕСКОЙ ДИАГНОСТИКИ ЗЛОКАЧЕСТВЕННОГО НОВООБРАЗОВАНИЯ | 1993 |
|
RU2093832C1 |
| СПОСОБ ПРОГНОЗИРОВАНИЯ ТЕЧЕНИЯ ОНКОЛОГИЧЕСКИХ ЗАБОЛЕВАНИЙ | 1990 |
|
RU2018835C1 |

