Изобретение относится к новым ДНК-последовательностям, рекомбинантным ДНК-молекулам, способам получения новых трансмембранных белков, экспрессированных в процессе развития Т-клеток, и к новым трансмембранным белкам, в основном, в чистом виде. В частности, настоящее изобретение относится к новым ДНК-последовательностям, экспрессированным в соответствующих хозяевах, и к новым белкам, два из которых являются интегральными мембранными белками, продуцированными в этих хозяевах. ДНК-последовательности и рекомбинантные ДНК-молекулы настоящего изобретения отличаются тем, что каждая из них кодирует новый белок, обладающий, по крайней мере, двумя из нижеперечисленных свойств, а именно: этот белок является /1/ экспрессированным Т-клетками лимфомы; /2/ экспрессированным в нормальной ткани тимуса, активированных клетках селезенки; или в кишечно-ассоциированной лимфоидной ткани; /3/ экспрессированным в ткани яичника, нормальных клетках печени или иммортализированных или раковых клеточных линиях; /4/ экспрессированным при эмбрионном росте; и/или /5/ этот белок имеет множество мембранных промежуточных доластей. Как можно видеть из последующего раскрытия изобретения, ДНК-последовательности, рекомбинантные ДНК-молекулы и способы продуцирования новых белков, экспрессируемых при развитии Т-клеток, и новых, в основном, чистых Т-клеточных белков могут быть использованы при манипулировании регуляцией Т-клеточного роста, для модификации онкогенного фенотипа, а также для локализации метастатических очагов опухолей. Настоящее изобретение также относится к новым антителам, которые связываются с эпитопами белков настоящего изобретения, и к использованию этих антител для идентификации и доставки лекарственных средств или других агентов к типоспецифическим клеткам, экспрессирующим новые белки.
В процессе развития иммунной системы Т-лимфоциты образуются из стволовых клеток предшественников, которые поступают в тимус, где происходит их дифференцировка и созревание. По мере происхождения Т-клетки через различные стадии развития в тимусе, многие гены либо активируются, либо подавляются. Например, в течение этого периода клетки приобретают IL-2-рецептор, CD4 и/или CD8 на своей поверхности. Эти маркеры дифференцировки являются важными для развития и/или функции Т-клеток. При развитии Т-лимфоцитов уровень экспрессии многих генных продуктов возрастает. Такими генными продуктами являются Т-клеточный рецептор для антигена, а также маркеры CD4 и CD8. В качестве Т-клеточных маркеров служили много других антигенов, прежде чем была установлена их точная функция. Лишь недавно было обнаружено, что Т-клеточный антиген Pgp1 "помогает" тимоцитам в их "хоминге" в тимус, а Т200 /CD45/ служит в качестве компонента для межклеточной передачи сигнала. Что касается другого Т-клеточного маркера Thy1, то его функция еще не выяснена.
Клетки SL 12. 4 обнаруживают CD4 CD8-двойной негативный фенотип, а поэтому являются схожими с тимоцитами на относительно ранней стадии развития. К тому же, они не экспрессируют α -субъединицу Т-клеточного рецептора. Однако, клетки SL 12.4 могут быть индуцированы для стабильной экспрессии CD4 и CD8 на их поверхности после совместного культивирования на тимусных эпителиальных монослоях. После таких обработок также индуцируется TCR-альфа мРНК. Таким образом, очевидно, что клетки SL12,4 обладают способностью к дифференцировке и созреванию. Эта уникальная in vitro биологическая система, до некоторой степени, имитирует микроокружение тимуса.
Был идентифицирован ряд генов, которые первыми экспрессируются в процессе развития тимоцитов. Многие из этих генов кодируют белки, которые должны быть экспрессированы для Т-клеток-предшественников для того, чтобы стать функциональными в иммунной системе, например, 1/ TCR для антигена, который необходим для распознавания антигена; 2/ CD25 /IL2-рецептор/, который должен быть экспрессирован в клетке, чтобы сообщить ей способность к ответу на цитокин IL2; 3/ генные продукты, играющие важную роль для трансдукции сигнала при распознавании антигена, такие, как CD3, CD4, CD45; 4/ некоторые из генных продуктов, участвующих в "хоминге" тимоцитов и обеспечивающих доставку к органам-мишеням; и 5/ генные продукты, участвующие в активации Т-клеток /Fowlkes и Pardoll, Advances in Immunology 44:207-264 /1989/; Hood и др., /1985/ Cell 40, 225-229; Rothenberg и Lugo, Develop. Biol 112, 1-17 /1985/; Adkins и др. , Ann. Rev. Immuol. 5:325-365 /1987/; Crabtree, Science 243: 343-355 /1989/; Kwon и Weissman, Proc. Natl. Acad. Sci. USA 86:1963-1967 /1989/.
Была отмечена гетерогенность в субпопуляциях тимоцитов, которые экспрессируют различные комбинации экспрессированных генов. Экспрессию генов детально анализировали во многих /но не во всех/ различных классах тимоцитов, и возможно, что остались неидентифицированные гены, которые кодируют продукты, способствующие Т развитию и "хомингу" Т-клеток, в частности, те продукты, которые экспрессируются в численно редких, переходных претимоцитах.
Благодаря экстенсивной гетерогенности тимоцитов практически невозможно получить фракционированные претимоциты в достаточных количествах или достаточной чистоты для полной характеризации каскада генной экспрессии, происходящей в процессе развития. По этим причинам клеточные линии лимфомы и лейкоза широко использовали для исследования экспрессии генов при развитии лимфоидных клеток /Greaves, Science 234: 697-704 /1986/; Hanley-Hyde и Lynch Ann. Rev. Immunol. 4: 621-649 /1986/. В значительной части литературных источников указывается, что редкие по своему числу, переходные клетки-предшественники являются мишенью для трансформации в злокачественные опухоли; и кроме того, в литературе указывается, что некоторые свойства трансформированных клеток-мишеней сохраняются в опухолевых клетках. Неожиданная экспрессия генов в опухолевых клетках часто не рассматривается как отклонение в трансформации. Однако, тщательный анализ "аберантной" генной экспрессии в гематопоэтических опухолевых клетках обнаружил редкие субпопуляции нормальных клеток-предшественников, которые экспрессируют указанные гены /Greaves, Science 234: 697-704 /1986/; Hanley-Hyde и Lynch Ann. Rev. Immunol. 4: 621-649 /1986/; Pierce и Speers Cancer Res., 48: 1996-2004 /1988/.
Гетерогенность клеточных линий лимфомы мыши и человека, происходящих от одного индивидуума, может быть следствием различной степени созревания отдельных клеток. Гетерогенность известных клеточных линий лимфомы была использована для получения близких по средству клеточных клонов, которые отличаются ограниченным количеством признаков. Hedrick и др., /Hedrick и др., Nature 308: 149-153 /1984/, используя субтрактивную технику клонирования, установили, что T- и B-клетки различаются в экспрессии около 100 генов. Возможно, что близкие по сродству T-клетки лимфомы могут различаться в экспрессии даже меньшего количества генов. Такие клеточные клоны являются удобными для работы с чистыми популяциями клеток с определенными и стабильными фенотипами, которые отличаются ограниченным числом признаков. В настоящей заявке, для получения такой близкой по сродству клеточной популяции разработали и использовали систему T-клеточной модели лимфомы, SL12. /Hays и др., Int.J.Cancer 30:597-601 /1986/; Macleod и др., Cancer Pesearch 44: 1784-1790 /1984/; Macleod и др., J.Nat. Cancer Inst. 74: 875-882 /1985/; Macleod и др. , Proc. Natl. Acad. Sci. VS A 83: 6989-6993 /1986/; Siegal и др., J. Exp. Med. 166: 1702-1715 /1987/.
Настоящее изобретение относится к новым ДНК-последовательностям, рекомбинантным ДНК /рРНК/-молекулам, способам получения новых T-клеточных белков, экспрессированных в процессе развития T-клеток; к новым, в основном, чистым T-клеточным белкам; и к антителам, которые связываются с указанными белками. В частности, настоящее изобретение относится к новым ДНК-последовательностям, экспрессированным в соответствующих хозяевах, а также к новым T-клеточным белкам, продуцированным в этих хозяевах. Настоящее изобретение также относится к новым трансмембранным белкам, являющимся, в основном, чистыми; к рДНК-молекулам, кодирующим указанные трансмембранные белки; и к способам продуцирования новых трансмембранных белков. ДНК-последовательности и рекомбинантные ДНК-молекулы настоящего изобретения отличаются тем, что они экспрессируются T-клетками лимфомы и имеют, по крайней мере, один из перечисленных ниже признаков, а именно: /1/ они экспрессируются в нормальном тимусе, в активированных клетках селезенки, или в кишечно-ассоциированной лимфоидной ткани; /2/ они экспрессируются в ткани яичника, нормальной ткани печени и/или эмбриональной ткани на определенной стадии развития; и /3/ они кодируют новые трансмембранные белки, имеющие множество мембранных промежуточных доменов.
В другом своем варианте настоящее изобретение относится к новому гену, 19,5, обозначаемому также Lov, который может быть индуцирован в клетках SL12.4 после совместного культивирования на эпителиальных монослоях тимуса. Настоящее изобретение также относится к поликлональным антителам против олигопептидной конструкции на основе Lov-последовательности кДНК. Индукция Lov, очевидно, является стабильной, поскольку клеточные клоны, выделенные из популяции клеток SL12.4 после сокультивирования, обнаруживают более высокий уровень экспрессии Lov в мРНК, а также более высокий уровень поверхностного белка. Ген Lov был картирован для мышиной хромосомы 16. Lov-генный продукт является эволюционно регулируемым и играет определенную роль в развитии T-клеток.
Была сконструирована кДНК-библиотека SL12.4, из которой были выделены шесть новых кДНК посредством субтрактивной гибридизации против сестринской клеточной линии лимфомы, SL12.3. Клетки SL12. 3 имеют признаки тимоцитов в более незрелой стадии развития, чем SL12. 4. Один из кДНК-клонов, 19. 5, экспрессировался в SL12.4, но не экспрессировался в SL12. 3-клетках. Эта последовательность была названа Lov /лимфоидная и овариально-клеточная экспрессия/. Предсказанный белок оказался в высокой степени гидрофобным, и как было установлено на основе компьютерного анализа, он содержит четыре трансмембранные промежуточные области. Причем, каких-либо значительных гомологий между Lov-кДНК, либо гомологий послеловательности предсказанного белка с другими известными последовательностями обнаружено не было. Lov является консервативным для таких видов млекопитающих, как, например, человек, грызуны, кролики, морские львы, а также для птиц; и в высокой степени экспрессируемым в яичниках и в кишечно-ассоциированной лимфоидной ткани /GALT/, а также в тимусе. В настоящей заявке раскрываются Lov-экспрессия и индуцируемость Lov в био-системе, физические свойства Lov-белка, а также локализация этого гена в мышиной хромосоме.
После получения ДНК-последовательностей и рекомбинантных ДНК-молекул настоящее изобретение также относится к зондам и способам идентификации клеток, содержащих или не содержащих эти последовательности, а также к средствам введения этих последовательностей в клетки, не содержащие данные последовательности. Кроме того, настоящее изобретение относится к способу ингибирования экспрессии новых последовательностей путем получения антисмысловой РНК-последовательности, которая при введении в клетку, или при введении ДНК, кодирующий указанную антисмысловую РНК в клетку, содержащую указанную ДНК-последовательность, будет продуцировать антисмысловую РНК, которая может связываться, а следовательно и блокировать синтез РНК, кодирующей новые белки настоящего изобретения. Из раскрытия настоящего изобретения каждому специалисту будет понятно, что для блокирования связывания лигандов с белками и для доставки лекарственных средств или других агентов /таких, как метки/ в клетки, экспрессирующих эти белки, могут быть использованы антитела против любых белков настоящего изобретения.
Настоящее изобретение также относится к кДНК-клону, 20.5, обозначаемому в данной заявке также Tea /SEQ IDN0:5/, который позволяет идентифицировать транскрипты, обнаруживаемые лишь в ограниченном числе тканей. Tea-транскрипты индуцируются в спленоцитах, активированных T-клеточным митогеном КонА. В отличие от других известных генов, экспрессируемых в активированных T-клетках, Tea-ген очевидно кодирует белок, который пересекает мембрану несколько раз, тогда как большинство известных интегральных мембранных белков, индуцируемые при активации T-клеток, являются простыми мембранными белками с одной мембранной областью /Crabtree, 1989, Science, 243:355-361/.
Настоящее изобретение также относится к способам продуцирования новых трансмембранных белков, являющихся, в основном, чистыми, которые могут быть использованы для регуляции развития T-клеток, для регуляции онкогенного фенотипа, а также для ингибирования активации T-клеток при аутоиммунном заболевании.
Новые кДНК-клоны, которые являются дифференциально экспрессированными между двумя близкими по сродству T-клеточными клонами лимфомы, были выделены с использованием дифференциального скрининга по методу исключения. Клетки SL12. 4, из которых выделяли кДНК, имеют признаки тимоцитов в промежуточной стадии развития и вызывают явно выраженные внеузловые овариальные опухоли у сингенных животных. Сестринский клеточный клон, SL12. 3, происходящий от этой же опухоли, имеет другой фенотип и вызывает более агрессивные, диффузные лимфомы. Четыре из пяти новых генов экспрессируются в нормальном тимусе, в активированных клетках селезенки, или в кишечно-ассоциированной лимфоидной ткани. ДНК-последовательности и предсказанные белковые последовательности для новых кДНК-клонов представлены на фиг. 3, 4, 15 - 18 и 33 - 37. Новые кДНК-клоны 19.5 обнаруживают мРНК в нормальном тимусе, в кишечно-ассоциированной лимфоидной ткани и овариальной ткани. Предсказанный белок имеет четыре предполагаемые трансмембранные промежуточные области. Экспрессия транскрипта подавляется в соматических клеточных гибридах, образованных из клеток SL12. 4, слитых с тремя различными T-клеточными линиями лимфомы, в которых отсутствует обнаруживаемая мРНК, комплементарная новому, кДНК-клону. Эта транс-негативная регуляция свидетельствует о том, что экспрессия гена регулируется по ингибирующему механизму.
На фиг. 1 иллюстрируют экспрессию 5 различных SL12. 4-специфичных кДНК-клонов с помощью Назерн-блоттинга; на фиг. 2 - гибридизацию методом Саузерн-блоттинга четырех различных SL12. 4-специфических кДНК-клонов; на фиг. 3 - 4 - ДНК и предсказанную белковую последовательность кДНК-клона 19.5 /SEQ IDN0: 4/. A, ДНК-последовательность и предсказанную аминокислотную последовательность получали с помощью двухцепочечного секвенирования. Для сборки ДНК-последовательности и получения предсказанной аминокислотной последовательности использовали Microgenie /Beckman/. B, показаны сайты рестрикции в кДНК для субклонирования, и стратегия секвенирования. Открытая рамка считывания показана пунктирной горизонтальной линией. Обе нити были секвенированы от интактной вставки и 4 субклонов с использованием праймеров для T3 и T7-областей плазмиды pT7T3 и одного синтетического олигонуклеотидного праймера. Для оценки физических свойств предсказанного белка и получения схематической структуры, показанной на фиг. 4c, использовали программы системы программного обеспечения PC Gene SOAP, HE LXMEM, NONOTNY и RAOARGOS.
На фиг. 5-6 иллюстрируют картины экспрессии кДНК 19. 5 и 20. 2 в гибридах соматических клеток, и 19. 5 в нормальной мышиной ткани; на фиг. 7 - с помощью метода "нозерн"-блоттинга, и с использованием последовательностей, комплементарных 19. 5, тот факт, что кДНК содержится в клеточных линиях карциномы яичника человека; на фиг. 8 - посредством Саузерн-блоттинга тот факт, что последовательности, комплементарные 19. 5, присутствуют во многих видах млекопитающих и являются, таким образом, консервативными в процессе развития; на фиг. 9 иллюстрируют посредством "нозерн"-блоттинга тот факт, что оба транскрипта Lov индуцируются в ответ на совместное культивирование; на фиг. 10 - анализ клеток SL12. 4 на поверхностную экспрессию Lov с использованием флуоресцентного антитела; на фиг. 11 - специфичность антитела 19. 5 для иммунизации антигена; на фиг. 12 - способность к индуцированию Lov-белка тимусными эпителиальными клетками; на фиг. 13 - стабильность Lov-экспрессии при "нозерн" блот- и FACS-анализах; на фиг. 14 иллюстрируют локализацию Lov в хромосоме 16; на фиг. 15 - 18 иллюстрируют ДНК и последовательность предсказанного белка клона кДНК 20. 5 /SEQ IDN0:5/; на фиг. 19 - экспрессию Tea-гена; на фиг. 20 - кинетику индукции Tea-гена в активированных спленоцитах; на фиг. 21 - 25 - сравнительный анализ кДНК-последовательностей 20. 5 и EPP; на фиг. 26 - 28 иллюстрируют сравнительный анализ Tea-последовательности предсказанного белка и рецепторной последовательности мышиного экотропного ретровируса.
На фиг. 29 - 30 иллюстрируют физические свойства предсказанных белковых продуктов Tea- и Rec-1-генов; на фиг. 31 - 32 иллюстрируют Саузерн-анализ ДНК от различных видов и анализ рекомбинантной инбредной ДНК на положение гена Tea на хромосоме 8; на фиг. 33 - ДНК и предсказанную белковую последовательность кДНК-клона 19. 1 /SEQ ID N0:1/; на фиг. 34 - ДНК и предсказанную белковую последовательность кДНК-клона 19. 2 /SEQ ID N0:2/; на фиг. 35 - 36 - ДНК- и предсказанную белковую последовательность кДНК-клона 19. 4 /SEQ ID N0: 3/; на фиг. 37 изображают схематическое представление pNEO/TfR-NC; на фиг. 38 изображают схематическое представление pMAMneoBlue.
В данном описании используется следующие термины:
термин "хозяин" включает в себя не только прокариоты, но также и эукариоты, такие как дрожжи и нитевидные организмы, а также растительные и животные клетки;
термин "прокариот" включает в себя все бактерии, которые могут быть трансформированы с помощью ДНК в целях экспрессии трансмембранных или рекомбинантных трансмембранных T-клеточных белков /ртТСР/ настоящего изобретения;
термин "эукариот" включает в себя все дрожжи, грибки, животные и растительные клетки, которые могут быть трансформированы с помощью ДНК в целях экспрессии трансмембранных или рекомбинантных трансмембранных T-клеточных белков настоящего изобретения;
ДНК для T-клеточных белков настоящего изобретения могут происходить от любых видов млекопитающих. При этом необходимо только, чтобы генетическая последовательность для T-клеточных белков /TCP/ экспрессировалась в прокариотическом или эукариотическом организме. Предпочтительной является T-клеточная ДНК, которая экспрессирует TCP-белки, происходящие от мыши. Особенно предпочтительной является последовательность T-клеточной ДНК, которая является иммунологически перекрестно реактивной между несколькими видами животных /например, таких как человек, мышь, кролик или морской лев/;
рекомбинантная ДНК-молекула, кодирующая любой T-клеточный белок настоящего изобретения, может быть использована для трансформации хозяина с помощью стандартной техники, хорошо известной специалистам. Особенно предпочтительно использовать вектор, содержащий последовательность, кодирующую T-клеточные белки настоящего изобретения, в целях трансформации прокариота;
рекомбинантный T-клеточный белок /pTCT/ настоящего изобретения может иметь больше или меньше аминокислот на фланкирующих концах по сравнению с аминокислотной последовательностью нативных T-клеточных белков;
термин "в основном, чистый", используемый в отношении трансмембранного T-клеточного белка настоящего изобретения, означает, что полипептид, в основном, не содержит других белков, обычно ассоциированных с T-клеточным белком в его натуральном виде, и обнаруживает постоянный и репродуцируемый электрофоретический или хроматографический ответы, профили элюции, и антигенную активность. Термин "в основном, чистый" не включает в себя искусственные или синтетические смеси T-клеточного протеина с другими соединениями.
Способы получения слитых, правильно присоединенных генов, и их экспрессии в бактериях хорошо известны специалистам и описаны, например, в патенте США 4 366 246, который вводится в настоящее описание посредством ссылки. Генетические конструкции и методы, описанные в этом патенте, могут быть использованы для экспрессии трансмембранного T-клеточного белка в прокариотических или эукариотических хозяевах.
Прокариотическими хозяевами могут быть Грам-отрицательные а также Грам-положительные бактерии, такие как E. coli, S. tymphimurium, Serratia marcescens и Bacillus subtilis.
Эукариотическими хозяевами могут быть дрожжи, такие как Pichia pastoris или клетки млекопитающих.
В основном, векторы экспрессии, содержащие промоторные последовательности для облегчения эффективной транскрипции вставленного ДНК-фрагмента, используются в соответствии с используемым хозяином. Вектор экспрессии обычно содержит начало репликации, промотор /промоторы/, терминатор /терминаторы/, а также специфические гены, обладающие способностью к фенотипической селекции в трансформированной клетке. Трансформированные хозяева могут быть подвергнуты ферментации или культивированию известными способами, обычно используемыми для достижения оптимального клеточного роста.
Примерами промоторов, которые могут быть использованы в настоящем изобретении /при этом указанные примеры не ограничивают настоящее изобретение/, являются: rec A, trp, lac, tac, бактериофаг лямбда pR, или pL, MMTV, SV40. Примеры некоторых плазмид или бактериофагов, которые могут быть использованы в настоящем изобретении, приводятся в Molecular Cloning Maniatis и др., Cold Spring Harbor laboratories, 1982, и в других хорошо известных литературных источниках, и могут быть легко подобраны любым специалистом.
Настоящее изобретение относится к любому хозяину, модифицированному в соответствии с описанными способами, или модифицированному любым другим способом, обычно применяемым специалистами в таких случаях, например, путем переноса генетического материала с использованием лизогенного фага, и способствующим экспрессии гена, кодирующего трансмембранный T-клеточный белок, в прокариотах или эукариотах.
Ген представляет собой ДНК-последовательность, которая кодирует, посредством ее матрицы или матричной РНК, последовательность аминокислот специфического пептида. Термин кДНК включает в себя гены, из которых удалены интроны. Термин рДНК включает в себя молекулу, подвергнутую рекомбинации путем сплайсинга кДНК- или геномной ДНК-последовательностей in vitro.
Клонирующей системой является плазмидная или фаговая ДНК или другая ДНК-последовательность, которая способна к репликации в хозяйской клетке; которая содержит один или небольшое число сайтов, узнаваемых эндонуклеазой, в которых указанная ДНК-последовательность может быть разрезана без потери ее основной биологической функции; и которая содержит маркер, подходящий для использования в идентификации трансформированных клеток. Такими маркерами могут быть, например, устойчивость к тетрациклину, неомицину или ампициллину. Слово "вектор" иногда используется для обозначения системы клонирования.
Система экспрессии является аналогичной системе клонирования, но при этом она обладает способностью к экспрессии данного структурного гена в хозяине, обычно под контролем определенных регуляторных последовательностей.
Хозяева, трансформированные трансмембранными T-клеточным геномом для трансмембранных T-клеточных белков, являются особенно подходящими для продуцирования трансмембранных T-клеточных полипептидов и белков.
Рекомбинантный T-клеточный белок может содержать полную аминокислотную последовательность T-клеточного белка, или он может содержать лишь специфическую детерминанту. Животное, иммунизированное T-клеточным рекомбинантным белком, будет продуцировать антитела, которые будут связываться с детерминантами, присутствующими на рекомбинантном или натуральном полипептидах. Таким образом, может быть осуществлено промышленное производство T-клеточных рекомбинантных белков.
Термин "индивидуум", используемый в настоящем описании, подразумевает любое животное, предпочтительно, млекопитающее, а наиболее предпочтительно, такие как грызуны, кошки, собаки, коровы или человек.
Обнаруживаемыми метками могут быть любые обнаруживаемые молекулы. Обычно используются радиоактивные метки, примерами которых могут служить /однако, эти примеры не ограничивают настоящего изобретения/ 32P, 34C, 125I, 3H, и 35S. Биотин-меченные нуклеотиды могут быть введены в ДНК или РНК путем "ник"-трансляции, или с помощью ферментных или химических средств. Биотилированные зонды могут быть обнаружены после гибридизации с использованием авидин/стрептавидинового, флуоресцентного, ферментного конъюгатов или конъюгатов с коллоидальным золотом. Нуклеиновые кислоты могут быть также мечены другими флуоресцентными соединениями, иммунодетектируемыми флуоресцентными производными, или аналогами биотина. Нуклеиновые кислоты могут быть также меченными путем связывания белка. Могут быть также использованы нуклеиновые кислоты, сшитые с радиоактивным или флуоресцентным гистоном H1, ферментами /щелочной фосфатазой и пероксидазой/, или с белком путем однонитевого связывания /SSB/.
Таким образом, в настоящем изобретении антитело против трансмембранного T-клеточного белка используется в качестве зонда для трансмембранных белков настоящего изобретения, и в качестве ингибиторов связывания натуральных лигандов трансмембранных T-клеточных белков настоящего изобретения, а также в качестве системы, обеспечивающей доставку лекарственного препарата или метки к органу-мишени.
Два клеточных клона, происходящие от T-клеточной линии лимфомы SL12, выбирали для выделения новых дифференциально экспрессированных генов, исходя из известных различий в экспрессии генов и их различной способности вызывать образование опухолей у сингенных животных-хозяев /Hays и др., Int.J. Cancer 38: 597-601 /1986/; Macleod и др., Cancer Research 44: 1784-/1790/ 1784-1790 /1984/; Macleiod и др., J. Nat. Cancer Inst 74: 875-882 /1985/; Macleiod и др., Proc. Natl. Acad. Sci. USA 83: 6989-6993; /1986/; Siegel и др., J. Exp. Med. 166: 1702-1715 /1987/; Weinzoth и др., Cancer Pesearch 45: 4804-4809 /1985/; Wilkinson и др., EMBO J. 7: 101-109 /1988/ и табл. 1, где систематизированы фенотипы/. Клеточная линия SL12. 3 экспрессирует очень небольшое количество генов, необходимых для функции T-клеток; при этом, эта линия клеток вызывает у сингенных животных образование в высокой степени злокачественных, диффузных, агрессивных опухолей. В противоположность этому, клетки SL12.4 экспрессируют мРНК для всех компонентов TCR/CD3-комплекса, за исключением TCP-альфа, и в некоторых отношениях, эти клетки аналогичны тимоцитам в промежуточной стадии развития указанных тимоцитов. Клетки SL12. 4 являются гораздо менее онкогенными и индуцируют истиные экстранодальные опухоли. У самок животных-хозяев, местом образования опухолей в первую очередь является яичник. Новые трансмембранные белки могут принимать участие в доставке опухолевых клеток к яичнику.
Новые кДНК-клоны выделяли из двух клеточных линий, отличающихся своей онкогенной способностью, способностью к "хомингу" и состоянием созревания. Эти кДНК-клоны представляют гены, которые кодируют продукты, связанные с различной способностью двух клеточных линий образовывать опухоли и/или гены, которые участвуют в развитии T-клеток. В настоящем изобретении раскрывается выделение и характеризация пяти новых кДНК-клонов, представляющих гены, которые предпочтительно экспрессируются в T-клеточном клоне SL12. 4, и не экспрессируются или очень незначительно экспрессируются в сестринском клеточном клоне SL12. 3. кДНК-клоны были получены путем комбинирования субтрактивной гибридизации обогащенных зондов и классического дифференциального скрининга. В настоящей заявке раскрываются новые кДНК-клоны, представляющие гены, дифференциально экспрессированные в двух клеточных клонах. Все из указанных генов экспрессируются в ограниченном количестве субпопулляций тканей. Некоторые транскрипты не находятся лишь исключительно в лимфоидных клетках, но указывают на порядок экспрессии. Последовательности двух кДНК /19. 5 и 20. 5/ /SEQ ID N: 4 и 5 соответственно/, свидетельствуют о том, что генными продуктами являются новые мембранные белки с несколькими промежуточными областями, экспрессирующиеся в нормальной ткани мышиного яичника, в тимусе, кишечной лимфоидной ткани, в селезенке и печени. Одна последовательность была также гибридизирована с клетками карциномы яичника человека и с нормальной тканью.
Для более ясного понимания настоящего изобретения ниже представлены примеры его осуществления. Эти примеры имеют лишь иллюстративный характер и ни в коем случае не должны рассматриваться как некое ограничение объема настоящего изобретения, если, разумеется, это не оговорено особо.
Пример 1. Выделение, характеризация и культивирование клеток.
A. Клеточные линии лимфомы. Требования к выделению, характеризации и культивированию T-клеточных линий лимфомы SL12.1, SL12.3, SL12.4 и гибридов соматических клеток, образованных между ними, подробно описаны в следующих работах Hays и др., Int. J Cancer 38: 597-601 (1986); Macleod и др., Cancer Research 44: 1784-1790 (1984); Macleod и др. J. Nat. Cancer Inst. 74: 875-882 (1985); Macleod и др., Proc. Natl Acad, Sci. USA 83: 6989-6993 (1986) и Weinroth и др. , 1985, Cancer Rese arch 45: 4804-4809, которые вводятся в настоящее описание посредством ссылки.
Суммарные данные по фенотипам клеточных клонов SL12.3 и SL12.4 представлены в табл. 1. Экспрессия транскрипта, экспрессия поверхностного белка, онкогенность и тип опухоли определяли с помощью "нозерн"-анализа, проточной цитометрии, и in vivo-инъекции клонированных клеток в сингенных животных, соответственно. 1,0 кв и 1,3 кв TCR - β -транскрипты кодируют (D)-T-C- и V-D-T-C-последовательности, соответственно. Глюкокортикоидный ответ определяли посредством культивирования клеток в 1 мМ дексаметазоне.
Клетки SAK8 (Gasson и Bourgeois, T. Cell. Biol. 96, 409-415 (1983) были получены у Dr. Gasson. Клетки лимфомы культивировали в модифицированной по способу Дульбекко среде Игла, дополненной 10% околоплодной сывороткой теленка, глутамином, пенициллином и стрептомицином. Две клеточные линии карциномы яичника человека 2008 (Disaea и др., Am. T. Obst et. Gynecol. 114: 979-989 (1972)) и COLO 316 (Woods и др., Cancer Res. 39: 4449-4459 (1979) культивировали в RPMI-среде 1640, дополненной 5% телячьей сывороткой, глутаминов, и 1% грибковой бактерии (Irvine Scientific, Santa Ana, CA). Когда клетки использовали для получения РНК, то эти клетки собирали в течение экспоненциального роста при плотности, близкой 5-8 • 105 клеток/мл (Wilkinson, и Macleod, EMBO T. 7: 101-109 (1988). Спленоциты, происходящие от мышей BALB/с, засевали с плотностью 3 • 106 клеток/мл и активировали 10 мкг/мл КонА в течение двух дней перед сбором РНК.
B. Совместное культивирование клеток SL12.4 и тимусных эпителиальных монослоев
Условия совместного культивирования SL12.4-клеток и тимусных эпителиальных слоев, в общих чертах, являются следующими.
Клетки SL12.4 засевали при такой плотности, чтобы их конечная концентрация после 3-х дневного периода совместного культивирования составляла 1 • 106 клеток/мл. На третий день культивирования, TEL или TEPI были в состоянии сплошности. Клетки культивировали в модифицированной по способу Дульбенко среде Игла, содержащей 10% плодную сыворотку теленка и дополненной глутамином и пенициллином/стрептомицином, при 37oC.
C. Клеточные линии для исследования экспрессии 20.5
В исследованиях экспрессии 20.5 были использованы клеточные линии из следующих источников: эмбриональные карциномы F9 и PCC4 (Bernstine и др., Proc. Natl. Acad. Sci. USA. 70: 3899-3903 (1973), опухоль гипофиза ATt20 (Buonassisi и др. , Proc, Natl. Acad Sci. USA 48: 1184-1192 (1962)), TEPI эпителия (Bearodsley и др. , Proc. Natl. Acad. Sci. USA. 80: 6005-6009 (1983)), эпителий млекопитающих (Evans, (1988) Science 240: 889-894) 12,9), 3TC (ATCC N 92), а MEF получали согласно описанию Freshney (Freshney, (1983), Culture of Animal Cells. Alan R. Liss, Inc. стр. 99 - 110). Клетки культивировали в модифицированной по способу Дульбенко среде Игла, дополненной 10% околоплодной сывороткой теленка, глутамином, пенициллином и стрептомицином. Клетки, использованные для получения РНК, собирали в течение экспоненциального роста при плотности культур 5-8 • 105 клеток/мл. Спленоциты, полученные от BALB/с мышей, засевали при 3 • 106 клеток/мл в среду RPMI 1640, дополненную как указано выше, и активировали с помощью 10 мкг/мл КонА в течение 6, 24, 48 или 72 ч. перед сбором РНК.
Пример 2. Стратегия клонирования и скрининга.
Poly (A)+ мРНК от клеток SL12.4 использовали в качестве матрицы для получения двухцепочечной (дц) кДНК (Gubler и Hoffman, Gene 25: 263-269 (1983). К предварительно метилицированной дцДНК добавляли EcoRI-линкеры. Дефосфорилированные λ gt 10-плечи (stratagene) лигировали с кДНК и упаковывали в мябла-фаг с использованием экстракта упаковки Stratagene в соответствии с инструкциями производителей (Huynh и др., IN. D. Glover (изд.), DNA Cloning Techni gues : A Practical Approach. IRL. Press, Oxford. U. K. (1984)).
Субтрактивную гибридизацию осуществляли, в основном, в соответствии с описанием Hedrick и др., Nature 308: 149-153 (1984); Timberlake, Dev. Biol. 78: 497-503 (1980). Одноцепочечную кДНК получали из 10 мг поли (A)+РНК SL12.4 с использованием 250 мК32P dCTP (Anmersham) в присутствии 100 мкг/мл актиномицина D и гибридизировали с Rot 1260 (молекула нуклеотида/литр • сек) с 25 мг о (A)+РНК от клеток SL12.3 в объеме 8 мл при 68oC в течение 18 ч. После гибридизации, оцкДНК собирали при помощи хроматографии на колонке с гидроксиапатитом. Из 1 мкг исходной кДНК SL12.4 выделяли приблизительно 120 нг (12% от исходной кДНК, содержащей 3 • 107 им/мин) и использовали в качестве зонда для гибридизации с двумя 150 мм нитроцеллюлозными фильтрами, содержащими 20000 бляшек λ gt10 на фильтр. Первый из двух отпечатков дубликатных фильтров из библиотеки λ gt 10 SL12.4 зондировали с использованием всей кДНК от мРНК SL12.3, а отпечаток второго фильтра зондировали с использованием субтрактивно обогащенный кДНК, полученной, как описано выше. Используемая техника аналогична технике, описанной Filmus и др. Очищенные от бляшек клоны λ-фага идентифицировали на SL12.4-специфичность с помощью двух скринингов (используя отдельно полученные субтрактивные зонды). Затем проводили "Нозерн"-анализ для подтверждения того, что клоны гибридизировались только с мРНК из клеток SL12.4, а не из клеток SL12.3. Вставки кДНК удаляли из лямбда-ДНК путем переваривания рестриктирующими ферментами Hind III и Bg III, изолирования в легкоплавкой агарозе (Sea Kem) и субклонирования в плазмидный вектор pT7/T3 (Bethesda Research Laboratory), переваренный Hind III и BamHI. Вставки не могли быть вырезаны из фага посредством EcoRI, поскольку EcoRI-сайты были разрушены во всех изолятах Kuziel и др., Nucl.Acid Res 15: 3181 (1987).
Пример 3. "Нозерн" - блот-анализ.
A. Общие процедуры. Полную клеточную РНК выделяли из клеточных линий и тканей методом с использованием гуанидинизотиоцианата (Maniatis и др., Molecular Cloning: A Laboratory Manual. Cold Spring Harborlaboratory Press, Cold Spring Harbor, Нью-Йорк (1983), модифицированным в соответствии с описанием (Wilkinson и др., EMBO T. 7 : 101-109 (1988)). Для "Нозерн"-анализа, 10 мкг РНК подвергали электрофорезу в 1% агарозном геле, содержащем формальдегид, и переносили на нитроцеллюлозные мембраны (Meinkoth, Wahl, Anal, Biochem. 13: 267-284 (1984)).
Равную загрузку и перенос РНК на полосу анализировали путем окрашивания акридиновым оранжевым (Maniatis и др., Molecular Cloning: A Laboratory Manual. Cold Spring Harbor Laboratory Press, Cold Spring Harbor N.J. (1983)) и путем гибридизации с актином, CHO-A и/или циклофиллином. "Нозерн" - блоты были гибридизированы со случайно праймированными (Amersham)32P-меченными кДНК-вставками в присутствии 10% декстрансульфата и 50% формамида в течение 12 - 18 ч. при 42oC, последовательно промывали с конечной 30-минутной промывкой в 0,1 • SSPE, 0,1% ДНС при 42oC или 50oC. Для удаления меченного зонда РНК-блоты промывали 0,1 • SSPE и 0,1% ДНС при 90oC, охлаждали до комнатной температуры, осушали воздухом и хранили под вакуумом до следующей гибридизации.
B. "Нозерн"-анализ РНК (Lov (19,5). После совместного культивирования клетки SL12.4 собирали и лизировали в 4 М изотиоцианата гуанидиния, 1 М 2-меркаптоэтанола и 25 мМ ацетата натрия (pH 5,2). Затем лизат помещали на слой 5,7 М хлорида цезия/2мМ ЭДТК и центрифугировали при 38000 об/мин в течение 18 ч. при 4oC. РНК-дебрис затем ресуспендировали в TE-буферов и дважды экстрагировали хлороформом/бутанолом. 10 мкг РНК подвергали электрофорезу в каждой полосе 1% формальдегид-агарозного геля и переносили на нитроцеллюлозные мембраны. "Нозерн"-блоты гибридизировали с ДНК-вставками Lov-гена, который был мечен 32P с использованием набора для мечения методом рандомизированного праймирования Amersham. Гибридизация протекала в течение 12 - 18 ч. при 42oC в присутствии 10% декстрансульфата и 50% формальдегида. Затем полученные авторадиограммы анализировали на изменения в РНКLov путем сканирующей лазерной денситометрии. Кажущиеся индукции были нормированы к РНК-загрузке, используя последовательную гибридизацию с актином и CHO-A, которые не индуцируются при совместном культивировании (Macleod и др., Proc, Natl. Sci, USA, 83: 6989-6993 (1986)). Поэтому указанные величины представляют собой скорректированный кратный прирост экспрессии Lov при сокультивировании по отношению к необработанным клеткам SL12.4.
C. "Нозерн"-блок-анализ РНК 20,5 (Tea). Полную клеточную РНК выделяли из SL12.4, SL12.3 и свежих тканевых препаратов от Balb/c-мышей с помощью метода, использующего изотиоцианат гуанидина, описанного выше. Цитоплазматическую или ядерную РНК получали согласно приведенному выше описанию. Для "Нозерн"-анализа, 10 мкг РНК подвергали электрофорезу в 1% формальдегидном агарозном геле и переносили на нитроцеллюлозные мембраны (Maniatis и др., (1983), Molecular Cloning; A Laboratory Manual. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, Нью-Йорк); Meinkoth, и Wahl (1984) Anal, Biochem. 13: 267-284).
Пример 4. Саузерн-блот-анализ.
A. Общая процедура. Полную клеточную ДНК выделяли из клеток T-лимфомы и соматических клеточных гибридов мыши и хомячка и из тканей других видов и переваривали рестриктирующими ферментами, указанными в надписях к фигурам согласно условиям поставщиков. 10 мкг переваренной ДГК наносили на каждую полосу 0,7% агарозного геля, подвергали электрофорезу и блотировали на носителе Нитран (Nytran) согласно способу, описанному (Meinkoth и Wahl, Anal, Biochem. 13: 267-284 (1984), а затем гибридизировали и промывали в соответствии с приведенным выше описанием для "Нозерн"-блот-анализа.
B. Саузерн-блот-анализ 20,5. Саузерн-блот-анализ 20,5 осуществляли согласно приведенному выше описанию, за теми лишь исключениями, которые указаны ниже.
Полную клеточную ДНК выделяли из клеток SL12.4, печени мыши и хомячка и из гибридов соматических клеток. ДНК от печени цыпленка и человека получали в готовом виде от Clonetec, Palo Alto, Калифорния. ДНК переваривали рестриктирующими ферментами, указанными в примерах согласно условиям поставщиков. 10 мкг переваренной ДНК наносили на каждую полосу 0,8% агарозного геля, подвергали электрофорезу в Трис-ацетатном буфере в течение, по крайней мере, 48 ч. , блотировали на носителях Нитран, гибридизировали и промывали в соответствии с описанным выше Нозерн-блот-анализом. Блоты, содержащие ДНК от других видов, промывали в условиях меньшей жесткости. Конечную промывку проводили при комнатной температуре в 2 • SSPE.
Пример 5. Выделение новых кДНК-клонов с использованием дифференциального скрининга по методу исключения.
Новые кДНК-клоны, которые были экспрессированы исключительно или преимущественно клетками SL12.4, чем клетками SL12.3, идентифицировали и выделяли как описано в Примерах 1- 4. Начальный скрининг 40000 рекомбинантного фага ggt10 дал лишь 11 кандидатов, восемь из которых были выделены после повторного скрининга с использованием субстрактивных зондов, в результате чего получали шесть различных генных продуктов.
Проводили характеризацию пяти кДНК-клонов. Нозерн-блоты РНК от клеток SL12.3 и SL12.4 показали, что изолированные кДНК-клоны были дифференциально экспрессированы в клетках SL12.3 и SL12.4, как показано на фиг. 1. Очищенные вставки от соответствующих SL12.4 - T-клеточных специфических кДНК клонов метили и использовали для зондирования Нозерн-блотов. Как указывалось, каждая дорожка содержала 10 мкг полной клеточной РНК от клеточных линий SL12.4 и SL12.3. Затем блоты зондировали вышеуказанной радиоактивно меченной кДНК-вставкой.
Стрелки указывают на относительную мобильность 18S - и 28S-транскриптов. Количество SL12.3- и SL12.4-РНК, загруженной на одну дорожку, является эквивалентным количеству, определенному путем гибридизации с CHO-A-кДНК-зондом, как описано в примере 3.
Некоторые кДНК-клоны были экспрессированы исключительно в клетках SL12. 4 /19 .5 и 20. 5/ /Seq IDN: 4 и 5, соответственно/, а другие /19. 1, 19. 2 и 19. 4/ /SEQ ID N 1-3, соответственно были в более высокой степени экспрессированы в клетках SL12. 4. Тот факт, что некоторые из кДНК, которые согласно идентификации, имеют более чем один класс размера транскрипта, позволяет предположить, что их инициация или терминация имеет место у различных сайтов, либо имеет место различный сплайсинг, либо они могут происходить от генов с близким родством.
кДНК-клоны, соответствующие низкокопийным или однокопийным генам, имеются в обеих клеточных линиях SL12. Поскольку в клетках SL12. 3 отсутствует обнаружимая экспрессия транскриптов 19. 5 и 20. 5, то были проведены исследования для выяснения, можно ли делецией или реаранжировкой гена объяснить отсутствие транскрипта. Геномную ДНК от клеток SL 12. 3, SL12. 4 и нормальной печени переваривали EcoRI, Hind III или Pst I, и анализировали с использованием очищенных вставок в качестве зондов для кДНК-клонов. На фиг. 2 проиллюстрирована Саузерн-блот-гибридизация SL12. 4 4-специфических кДНК-клонов. Геномную ДНК /10 мкг на дорожку/ из клеток AKP-мышиной печени, из SL12. 3-, SL12. 40 и SAK-клонированных T-клеточных линий лимфомы, и из гибридных клеток SL12. 3 • SL12. 4, SAK • SL12. 4, SAK • SL12. 3 переваривали EcoRI и анализировали с помощью Саузерн-блотов, гибридизированных с указанными кДНК-клонами, как указано в Примере 4. Размеры фрагментов /в кв = тысячи пар оснований/, определенные путем совместной миграции лямбда-Hind III-переваренной ДНК, указаны на полях. Один или несколько рестрикционных фрагментов "узнавались" зондами, что указывало на то, что соответствующие гены присутствуют в обеих клеточных линиях при низком числе копий в мышином геноме. Интенсивность гибридизации была аналогичной для всех дорожек, что свидетельствует о том, что гены присутствуют в SL12. 3-клеточной ДНК приблизительно в таких же количествах, и не имеют обнаруживаемых реаранжировок, как показал анализ с использованием 4-х различных ферментов /фиг. 2/. Поэтому, отсюда можно сделать вывод, что различие в экспрессии генов в SL12-клеточных клонах, очевидно, обусловлено клеточно-специфической регуляцией, и не потерей или обнаружимой реаранжировкой в соответствующих генах. Однако, небольшие реаранжировки или точечные мутации в клетках SL12. 3 не могут быть полностью исключены. SL12. 4-специфические кДНК-клоны представляют новые гены. кДНК-клоны 19. 1, 19. 2, 19. 5 и 20. 5 /SEQ ID NN 1, 2, 4 и 5, соответственно/ секвенировали полностью. Это секвенирование не показало значительных гомологий с известными последовательностями. ДНК-последовательности показаны на фиг. 3-4, 15-18, 33-36.
Хотя 19, 4-последовательность |SEQ II N:3/ была определена лишь частично /приблизительно 2,8 кв из 3,6 кв/, однако, значительной гомологии этой последовательности с имеющимися в банке данных последовательностями ДНК обнаружено не было; не было также установлено идентичности в отношении размеров транскрипта, уровней экспрессии и характера экспрессии в ткани. Кроме того, было установлено, что ни один из исследуемых кДНК-клонов не соответствует известным T-клеточным генам, онкогенам или мышиным ретровирусным генам, которые, как известно экспрессируются в AKR-кдеточных линиях, в отношении размера транскрипта или картины происхождения экспрессии Hagiwara и др., J. Immiol. 138: 2514-2517 /1987/; Heckford и др., J. Immiol 137: 3652-3663 /1986/; LeClair и др., EMBO. J. 5: 3227-3234 /1986/; Mushinski, и др., Science 220: 795-798 /1983/; Yagie и др., Cell 42: 81-87 /1985/; Quint и др., J. Virol. 39: 1-10 /1981/; Selton и др., EMBO T. 4: 1793-1798 /1985/.
Размер кДНК и транскриптов, распознаваемых ими, а также длина открытой рамки считывания /OPF4/ показаны в табл. 2.
На основании сравнения размеров транскрипта и кДНК-вставки, можно сделать вывод, что ни одна из кДНК не имеет полную длину. 1,6 кв-ДНК-последовательность кДНК 19.1 не содержит какой-либо из длинных OPF, имеет полосу polyA, и очевидно, представляет собой 3-нетранслированную последовательность. 19.2-кДНК составляет 3,5 кв в длину, но имеет относительно короткую OPF /471 п.о. 5, табл. 2/. Частичная последовательность 19.4-кДНК, как обнаружилось, имеет OPF длиной 2,4 кв. Подобно 19.4, кДНК-клон 19.5 содержит полную кодирующую последовательность приблизительно полной длины.
Пример 6. Анализ кДНК 19.5.
кДНК-клон 19,5, очевидно, кодирует множественный мембранный белок с промежуточными областями. 19.5-кДНК и предсказанная аминокислотная последовательность показаны на фиг. 3. Стратегия секвенирования показана на фиг. 4b. кДНК-последовательность имеет несколько участков, свидетельствующих о том, что полная кодирующая область присутствует в 19.5-клоне:
1/ имеются стоп-сигналы во всех трех рамках считывания перед предсказанным старт-положением и внутри предсказанной 3-нетранслированной области;
2/ предсказанный стартовый метиониновый кодон окружен последовательностью, которая соответствует GXC AUG G /где X может быть A, C, G или U., обобщающей типичной последовательности Kozak, для оптимального участка инициации трансляции /30/;
3/ потенциальный сигнал полиаденилирования, ATTAAA, /31/ присутствует у 3 терминирующего кодона.
Выведенная аминокислотная последовательность дает размер предсказанного /немодифицированного/ белка 32981 Да и обнаруживает два потенциальных сайта гликосилирования, которые отмечены звездочками на фиг. 3. Предсказанная аминокислотная последовательность дает точную оценку молекулярной массы, которая может быть изменена либо путем гликосилирования, либо путем фосфорилирования, либо тем и другим. С использованием программ системы программного обеспечения Intelli Genetics, были идентифицированы четыре в высокой степени гидрофобных, потенциальных трансмембранных промежуточных областей /подчеркнуты на фиг. 3/. Сигнальной последовательности обнаружено не было. На фиг. 4c показана схема возможной структуры мембранного белка на основе физических свойств предсказанного белка. Хотя предсказанная структура напоминает никотиновый ацетил-холиновый рецептор а, однако, сходства с белковой или ДНК-последовательностью обнаружено не было. На фиг. 3 показаны последовательности ДНК и предсказанного белка кДНК-клона 19.5. На фиг. 3 представлены кДНК-последовательность и предсказанная аминокислотная последовательность, полученная путем двухцепочечного секвенирования. Для сборки ДНК-последовательности и получения предсказанной аминокислотной последовательности использовали программу Microgenie /Beckman/. На фиг. 4b показаны сайты рестрикции в кДНК, использованные для субклонирования и секвенирования. Открытая рамка считывания показана пунктирной горизонтальной линией. Секвенирование обеих цепей проводили от интактной вставки и 4 субклонов с использованием праймеров для T3- и T7-областей плазмиды pT7T3 и одного синтетического олигонуклеотидного праймера. Для оценки физических свойств предсказанного белка и получения схематической структуры, показанной на фиг. 4c, использовали программы системы программного обеспечения SOAP, HELIXMEM, NONOTNY, RAOARGOS.
Пример 7. Негативная регуляция экспрессии гена 19.5 в соматических клеточных гибридах.
С использованием соматических клеточных гибридов, образованных путем слияния SL12.3 и SL12.4-клеток /SL12.3 • SL12.4/, исследовали различную экспрессию 19.5-родственных транскриптов в клетках SL12.3 и SL12.4. Эти гибриды представляют особый интерес, поскольку они имеют необычно стабильное и почти тетраплоидное число хромосом. Гибридные клетки SL12.3 • SL12.4 содержат 79 хромосом и не теряют ни одной хромосомы в течение нескольких лет. На фиг. 5 видно, что экспрессия 19.5-транскриптов сильно подавляется в гибридных клетках SL12.3 • SL12.4. Кроме того, слияния, имеющие место между клетками SL12.4 и двумя другими T-клеточными линиями лимфомы, SAK8 или SL12.1, в которых отсутствует экспрессия мРНК 19.5 /фиг. 5/, также приводит к образованию гибридных клеток, которые не содержат обнаружимых транскриптов, как показано на фиг. 5.
Низкая экспрессия гена 19.5 во всех трех соматических клеточных гибридах не является результатом потери генов или их явной перегруппировки, поскольку сравнение Саузерн-блотов ДНК от SL12.3 и SL12.4-клеточных линий не показало какого-либо значительного различия в размерах полос или интенсивности, показанных на фиг. 2. Кроме того, представляется маловероятным, что обе хромосомы, содержащие ген, сообщенный SL12.4-родителем, теряются в трех независимых гибридах, имеющих почти тетраплоидное число клеточных хромосом и стабильность карио-типа. Гены, представляемые 19.5-кДНК, являются негативно регулируемыми в соматических клеточных гибридах, образованных между клетками SL12.4 и тремя независимыми T-клеточными линиями, в которых отсутствует экспрессия гена 19.5. Полученные результаты позволяют предположить, что факторы супрессии, сообщаемые клетками SL12.3, могут быть, по крайней мере, частично ответственными за отличающуюся экспрессию гена, обнаруживаемого с помощью 19.5-кДНК-зонда. Аналогично, было показано, что TCR-бета- и CD3-дельта-транскрипты регулируются негативными факторами, присутствующими в клетках SL12.3. Поскольку в качестве гибридных партнеров для клеток SL12.4 использовали три различные T-клеточные линии лимфомы, и во всех трех гибридах отсутствует обнаружимая экспрессия мРНК 19.5, то очевидно, что в регуляции экспрессии гена 19.5 участвуют негативные факторы. Представляется маловероятным, что мутации или необнаружимые делеции обеих 19.5-аллелей от SL12.4-родительской клетки могут иметь место во всех трех независимых гибридах, происходящих от различных клеточных источников.
При изучении соматических клеточных гибридов между клетками различного происхождения, наблюдается феномен фенотипической экстинкции, в результате чего аккумуляции транскриптов с сильно дифференцированной специфичностью не происходит /Davidson, Ann. Rev. Genet. 8: 195-218 /1974/; Hyman и Stallings, Immunogenetics 6: 447-458 /1978//. Фенотипическая экстинкция /т.е., ослабление прим. пер./ является редким явлением для гибридов, образованных между близкородственными клетками, происходящими от одного источника /Hyman и др., Immunogenetics 10, 261-271 /1980//. Если супрессия происходит в гибридах соматических клеток одного и того же происхождения то, как правило, она обусловлена действием специфических трансактивирующих депрессантов, экспрессированных одной из родительских клеточных линий. В случае гена гормона роста, супрессия в соматических гибридных клетках оказывает косвенное воздействие на специфический позитивный активатор, GHFI McCormick и др., Cell 55: 379-389 /1988//.
Пример 8. Тканевое распределение SL12.4-клон-специфических последовательностей
РНК, происходящие от ряда мышиных тканей, оценивали для определения, везде ли происходит экспрессия гена, соответствующего 19.5, либо эта экспрессия является ткане-специфической.
Фиг. 5-6 иллюстрируют картины экспрессии кДНК 19.5 и 20.2 в гибридах соматических клеток и 19.5 в нормальной мышиной ткани. Нозерн-блоты получали согласно описанию в примере 3 и на фиг. 1. На фиг. 5 показаны РНК от SL12.3, SL12.4, SAK8 и SL12.1-родственных клеточных линий и гибридных клеток, проанализированных на экспрессию генов 19.5 и 20.2. На фиг. 6 показаны РНК от нормальных тканей и органов, происходящих от мышей Balb/c. Размеры транскриптов оценивали на основании их относительной миграции с актином, циклофиллином, 18- и 28S-рибосомной РНК. 19.5-кДНК гибридизировался с двумя транскриптами в 1,7 и 1,5 кв. Блоты зондировали с использованием произвольно праймированной 19.5-кДНК-вставки, а затем зондировали с использованием CHO-A, актина и/или циклофиллина /CP/ для проверки, все ли дорожки содержат примерно эквивалентные количества РНК. CP-контроль, очевидно, позволяет с большей надежностью измерить РНК-загрузку для нормальных тканей, чем актин, при исследовании РНК от нормальных клеток. Все, кажущиеся одинаково загруженными дорожки окрашивали акридиновым оранжевым перед их блотированием.
Небольшие количества 19.5-мРНК присутствуют в GALT /т.е., в кишечно-ассоциированной лимфоидной ткани /gnt-associated Iymphoid tissue/, включая клетки пейеровых бляшек/, мозге, сердце и легких, и большое количество этого транскрипта имеется в яичнике /фиг. 6/. Однако, 19.5-транскрипты не были обнаружены ни в печени, ни в тонких кишках /фиг. 6/, ни в поджелудочной железе, ни в яичках, ни в костном мозге, ни в неподвижных спленоцитах, ни в спленоцитах, активированных T-клеточным митогеном Кон А /табл. 3/. Хотя 19,5-родственные транскрипты не были обнаружены в нефракционированных тимоцитах /фиг. 6/, а также в очищенных CD4 CD8-би-негативных тимоцитах, однако, мРНК присутствует в целом тимусе /фиг. 6/ и в одной тимусной эпителиальной клеточной линии /TEL/. Поэтому, по всей вероятности, транскрипт экспрессируется стромальными клетками тимуса.
Некоторые из новых кДНК-клонов распознают мРНК, экспрессия которых не ограничена клетками лимфоидного происхождения. Аналогичное явление наблюдается и в отношении других генов, обладающих известной T-клеточной функцией, например, таких, как Thy-1 и CD4, которые также экспрессируются в нормальных клетках мозга /Lonberg и др. , Mol. Cell Biol. 8: 2224-2228 /1988//; и антигена 6C3, маркера для пре-B-клеточных опухолей, который также был обнаружен на кортикальных эпителиальных клетках тимуса /Adkins и др., Ann. Rev. Immunol. 5: 325-365 /1987//. Экспрессия 19.5-транскриптов в нормальном яичнике и в клеточной линии карциномы яичника является довольно неожиданным фактом, если учесть, что клеточная линия SL12.4 индуцирует истинные экстранодальные опухоли в яичнике у 95% реципиентов-самок животных /14/. В противоположность этому, T-SL12.3-клетки лимфомы образуют диффузные опухоли, которые не возникают в яичнике /Macleod и др., J. Nat. Cancer Inst 74: 875-882 /1985// и эти клетки не экспрессируют 19.5-транскрипты. Известно, что B-лимфома человека /Burkitts'/ часто метастазирует в яичках /Harrisons Principle of Internal Medicine. Ilth Edition, /Ed. Braunwald и др., A, McGraw Hill, N.Y. /1987//. Поскольку 19.5-кДНК гибридизируется с двумя различными клеточными линиями яичника человека, то она может быть функцией распознавания или "хоминга" для 19.5-генного продукта, которая позволяет ей самой обосноваться и развиваться в яичнике. Даже если 19.5-мРНК не находится во всех тимоцитах или во фракционированных би-негативных тимоцитах, все же, возможно, что этот ген экспрессируется в редких суб-популяциях тимоцитов или он экспрессируется лишь временно в эмбриональной или зрелой стадиях лимфопоэза. Исследование гибридизации in situ позволит определить типы клеток в тимусе, яичнике и сердце, которые экспрессируют 19.5-родственные транскрипты.
В табл. 3 систематизированы исследования экспрессии, которые проводились на всех пяти кДНК-клонах /19,5 включен для сравнения/.
Относительную экспрессию транскриптов, комплементарных новым кДНК-клонам, оценивали с помощью Нозерн-анализа. Знак "+" означает, что данный зонд обнаруживал транскрипт от указанной ткани. Во всех случаях, транскрипты от мышиной ткани мигрируют вместе с РНК от клеточной линии SL12.4. Знак "-" означает экспрессию, по крайней мере, в 10 - 20 раз меньшую, чем в клетках SL12.4.
Четыре из пяти кДНК-клонов обнаруживают транскрипты в лимфоидной ткани: 19.1, 19.2, 19.4 и 19.5 заметно экспрессируются в тимусе; экспрессия 19.1, 19.2 и 19.4 индуцируется в Кон А-активированных спленоцитах; 19.1, 19.2 и 19.5 экспрессируются в GALT. Некоторые нелимфоидные ткани содержат транскрипты, которые гибридизируются с 19.2- и 19.5-кДНК-зондами /фиг. 6 и табл. 3/.
Клеточная линия карциномы яичника человека экспрессирует последовательности, комплементарные кДНК 19.5. Поскольку в мышиной ткани яичника наблюдался высокий уровень экспрессии 19.5-гена, то клеточные линии карциномы яичника были исследованы на присутствие последовательности 19.5. Нозерн-блоты 10 мкг полной РНК от клеток SL12.4 и двух клеточных линий карциномы яичника были зондированы, как описано на фиг. 2. Конечную промывку блота слева осуществляли 2 • SSPE при комнатной температуре. Такой же блот, показанный справа, затем промывали 0,2 • SSPE при комнатной температуре, что приводило к потере сигнала из дорожек, содержащих РНК человека, но не из дорожки, содержащей РНКSL12.4, происходящей от мыши. РНК от двух установленных клеточных линий человека 2008 и COLO 316 исследовали с помощью Нозерн-анализа, в результате чего было установлено, что они содержат 1,7 кв-транскрипт, который гибридизируется с 19.5-кДНК-зондом /фиг. 7/. Обнаружение того факта, что клетки мезодермального происхождения экспрессируют транскрипт, является до некоторой степени неожиданным, если учесть, что эти клетки составляют лишь небольшой процент от массы нормального яичника. Они находятся в одном толстом слое клеток на внешней поверхности яичника. Стромальные клетки яичника также могут экспрессировать этот ген. Присутствие транскриптов, которые гибридизируются с 19.5-кДНК, позволяет предположить, что ген, соответствующий гену, кодирующему 19.5-мРНК, находится в геноме человека.
Клетки некоторых видов млекопитающих экспрессируют последовательности, комплементарные 19.5-кДНК. Для того, чтобы определить, содержат ли ДНК других млекопитающих последовательности, общие с 19,5, приводили зондирование Саузерн-блотов ДНК от льва, хомячка, мыши и человека. Саузерн-блоты 10 мкг ДНК от указанных источников, переваренные Pst1, зондировали произвольно праймированной 32P-меченной 19.5-кДНК-вставкой. Эти блоты промывали с конечной жесткостью 1 • SSPE при комнатной температуре. Дорожка, содержащая ДНК человека, имела относительно высокий фон. Точки с правой стороны дорожки указывают на репродуцируемо-обнаружимые фрагменты ДНК, которые перекрестно гибридизируются с мышиным зондом. Все ДНК имеют простые картины гибридизации, что позволяет предположить, что этот ген является консервативным для млекопитающих /фиг. 8/. Как видно из фиг. 8, комплементарные последовательности были также обнаружены в ДНК кролика, крысы, лемура и орангутана. Таким образом, очевидно, что ген, представленный 19.5-кДНК-клоном, является консервативным внутри видов млекопитающих. Консервативность этого гена позволяет предположить, что он имеет важную, но еще неизвестную функцию.
Количество транскриптов, распознаваемых новыми кДНК-клонами, не было определено непосредственно. Однако, при сравнении на Нозерн-блотах, 19.2 и 19.5 оказались относительно не обогащенными мРНК, которые экспрессируются в меньшем числе копий, чем TCP-бета-мРНК. Тот факт, что 4 из генов, клонированных из клеток SL12.4, обнаруживают заметную экспрессию в тимусе, 3 гена - в активированных спленоцитах, и 3 гена в клетках GALT, свидетельствует о том, что модельная система лимфомы может быть использована для выделения генов, регулируемых в нормальных лимфоидных тканях, и в T-клеточном онтогенезе.
Ни один из новых кДНК-клонов, описанных в настоящей работе, не соответствует известным генным последовательностям. Для оценки предсказанных белков, представляемых 19.2, 19.4, 19.5 и 20.5-кДНК, на нуклеотидные или ДНК-сайты связывания, цинковые "пальцы" /"фингеры"-"finger"/, лейциновые "застежки молнии" /"zipper"/, центры ферментной активности, клеточные центры доставки, или на признаки структурных белков, на рецепторы, цитокины и факторы роста использовали программу PC Gene, Prosite, /Intell Genetic/. Эти анализы не обнаружили перечисленных выше сайтов ни в одном из предсказанных белков. В то же время не имеется каких-либо свидетельств на их потенциальную функцию.
По всей вероятности, эти гены кодируют продукты, участвующие в процессе дифференцировки, поскольку:
1/ эти гены дифференциально экспрессируются в клеточных клонах SL12.3 и SL12.4, которые, очевидно, представляют собой различные стадии созревания тимоцитов;
2/ эти гены обнаруживают тканевую специфичность при их экспрессии;
3/ экспрессия генов 19.1, 19.2 и 19.4 индицируется в T-клетках селезенки, активированных T-клеточным митогеном Кон А;
4/ 20.5- и 19.5-родственные гены экспрессируются специфическим образом в стадии раннего эмбрионального развития.
Серии новых кДНК-клонов кодируются генами, которые имеют представляющие интерес картины экспрессии регуляции. Они могут быть использованы для оценки молекулярных механизмов, регулирующих клеточную дифференцировку, "хоминга" T-лимфомы, и онкогенных свойств трансформированных T-клеток-предшественников. Указанные кДНК-клоны могут быть также использованы в качестве зондов для локализации клеток, несущих эти новые T-клеточные гены, и в качестве источников для синтеза в основном, чистых новых T-клеточных белков.
Пример 9. Получение поликлональных антител против Lov
14-аминокислотный олигопептид синтезировали с использованием VCSDCore. Сконструированная последовательность пептида представляла собой K-G*-H-A-D-R-E-Q-E-S-E-E-E-V. Эта последовательность представляет собой последние 13 аминокислот в предсказанном C-конце Lov с добавлением 14C-глицина для использования его в качестве индикатора связывания с носителем. 2 мг этого пептида связывали с 4 мг гемоцианина лимфы улитки, используемого в качестве белка-носителя /K H/, путем медленного /в течение 30 мин/ добавления глутарового альдегида. После этого в течение ночи диализовали против PB. Перед иммунизацией получали 10 мл сыворотки от 2 самок новозеландских белых кроликов /Holbarts, возраст 6 месяцев/. Затем животным инъецировали конъюгированный пептид в присутствии полного адъюванта Фрейнда. Животным инъецировали 0,2 мг полного пептида подкожно в 5 местах. Повторную бустер-иммунизацию проводили через 4 недели путем введения 0,2 мг пептида в неполном адъюванте Фрейнда. Еженедельно проводили анализ сыворотки крови на продуцирование антител против Lov с помощью ELISA.
Пример 10. Анализ антисыворотки к Lov.
Присутствие антител, распознающих Lov, контролировали с помощью ELISA-анализа. Олигопептид и KLH осушали непосредственно на ELISA-планшетах в лунках. Затем добавляли различные разведения антисыворотки и оставляли для реакции на 2 ч. После промывки в Blotto /реагент для проведения анализа методом "вестерн-блоттинг" - прим. пер./ образцы инкубировали в 1:1000 разведении "второго" антитела /козий антикроличий иммуноглобулин/ IgG /Fc/, связанный с пероксидазой хрена/ в течение 2 ч. Пероксидаза хрена катализует колометрическую реакцию с использованием H2O2 и OPD в качестве субстрата. Результаты анализировали на автоматическом спектрофотометре для прочтения ELISA-планшетов /планшет-ридере/. Когда титр антител достигал плато /титрования/, то кроликам выпускали кровь /приблизительно через 9 недель после начальной инъекции/. В ELISA-анализах использовали клеточные компоненты, клетки лизировали ультразвуком. Мембранные фракции выделяли из цитоплазмы путем центрифугирования лизата в течение 30 мин при 45000 г. Затем мембрану и цитозольные фракции осушали на ELISA-лунках в течение ночи.
Пример 11. Иммунофлуоресцентный анализ.
106 совместно культивированных или необработанных клеток SL12.4 суспендировали в 25 мкл буфера A/PBS, содержащий 5% околоплодной телячьей сыворотки и 0,02% азида натрия/. Затем добавляли 50 мкл 1:100-разведенной антисыворотки против Lov, и инкубировали на льду в течение 30 мин. После трех промывок в буфере A, клетки инкубировали на льду в течение 30 мин вместе с козьим антикроличьим IgG /разведенным 1:1000/. В экспериментах по испытанию на специфичность взаимодействия, к антисыворотке, до ее добавления к клеткам, добавляли в течение 30 мин равный объем 1 мг/мл раствора олигопептида. Клетки анализировали на флуоресцентно-активированном клеточном носителе цитофлуорографа 50Н с использованием лазерного излучения при 488 нм. Погибшие клетки удаляли благодаря окрашиванию пропидийиодидом. 5000 клеток анализировали для построения гистограмм.
Пример 12. Иммунопреципитация Lov-белка.
Иммунопреципитацию осуществляли методом, описанным Harlow и Lane, In Antibodies: A Laboratory Manual Cold Spring Harbor Press, N. Y. /1988/. Клетки подвергали импульсному мечению 35метионином в безметиониновой среде в течение 4 ч. Затем клетки лизировали либо в РИПА-буфере, либо в лизирующем буфере, содержащем NP40. Лизаты подвергали предварительному клиренсу путем добавления 50 мкл преиммунной сыворотки, инкубированной на льду в течение 1 ч и перекрестно-реактивные белки подвергали иммунопреципитации, используя Staphylococcus aureus /Calbiochem/. Затем добавляли еще 5 мкл иммунной сыворотки, инкубировали в течение 1 ч., и осаждали. После этого материал промывали и проводили электрофорез в ПААГ с ДСН. После иммуностимуляции гель экспонировали с рентгеновской пленкой и проводили анализ с помощью радиоавтографии, хорошо известный специалистам.
Пример 13. Генетическое картирование Lov.
Продуцирование и характеризация гибридов соматических клеток китайского хомячка и мыши были описаны ранее /Kozak /1983/ J. Virol. 48: 300-303/. Из этих гибридов выделяли ДНК и переваривали Pst1, ферментом, который давал уникальную карту для каждого вида, после того, как перевары ДНК мыши или хомячка зондировали с использованием 10 кв 19.5-кДНК-вставки. Переваренную ДНК подвергали электрофорезу на 0,8% агарозном геле в течение 72 ч. при 25 В, а затем переносили на нитрановые мембраны. Полученные блоты зондировали 32P-меченной Lov-кДНК и экспонировали с рентгеновской пленкой Kodak XAR. Для нескольких клеточных линий получали данные о содержании мышиных хромосом, гибридизирующихся с Lov, и полученные результаты анализировали на компьютере для определения наилучшего кандидата для локализации мыши.
Пример 14. Индуцирование экспрессии Lov в клетках SL12.4, после совместного культивирования на тимусных эпителиальных монослоях.
TEL /Glimcher и др., Scand.J. Immunol. 17: 1-11 /1983/ и TEPI /Beardsley и Hays Proc. Natl. Acad. Sci. USA 80: 6005-6009 /1983// представляют собой две линии иммортализированных тимусных эпителиальных клеток, которые могут индуцировать стабильную экспрессию гена CD4 и CD8 в клетках SL12.4. После совместного культивирования клеток SL12.4 либо на TEL-либо на TEPI-монослоях в течение трех дней, оценивали мРНК-уровни Lov. Нозерн-блоты на фиг. 9 показывают, что оба транскрипта /1,7 и 1,5 кв/ Lov индуцируются в ответ на совместное культивирование. Обе линии тимусных эпителиальных клеток позволяют выявить эту индукцию, хотя после совместного культивирования с TEL уровень экспрессии Lov выше по сравнению с TEPI /табл. 4/. Полученные результаты представляют данные денситометрического анализа /± станд. ошибка/ по пяти отдельным экспериментам. Более того, повышенные уровни Lov-мРНК в SL 12.4-клетках сохраняются даже после удаления стимулирующих тимусных монослоев. Нетимусные прилипающие клетки /MME: линия эпителия мышиной молочной железы, и AKRI-2B: линия фибробластов/ не оказывали индуцирующего действия на экспрессию Lov в SL12.4-клеткх. 100 п.о.-фрагменты, соответствующие 5'- и 3' - областям кДНК Lov были использованы для Нозерн-зондов. Оба зонда распознавали оба транскрипта Lov.
Авторадиограммы, полученные в результате последовательной гибридизации Нозерн-блотов с CD4, CD8, TCR-α, 19.5 /Lov/, и наконец с актин/CHO-A, сканировали с помощью лазерной денситометрии. Денситометрические сигналы нормализовали для загруженной РНК с использованием в качестве контроля актин/CHO-A. Полученные величины представляют собой отношение интенсивности гибридизации индуцированной мРНК к неиндуцированной мРНК. Таким образом, они представляют собой величину /± станд. ош./, показывающую, во сколько раз уровень накопления мРНК в указанных выше условиях превышает мРНК-уровень в необработанных SL12.4-клетках.
Совместное культивирование этих клеток на тимусном эпителиальном монослое, очевидно, вызывает созревание SL12.4-клеток. В настоящей заявке показано, что новый ген Lov также индуцируется в SL12.4-клетках после указанной обработки. Последовательность Lov свидетельствует о том, что предсказанный кодированный белок должен быть поверхностной молекулой, содержащей несколько мембранных промежуточных областей. Увеличение уровней обеих мРНК /3-4-х кратная индукция/ и поверхностной экспрессии Lov наблюдалось после совместного культивирования на тимусном эпителии. Такое индуцирование экспрессии Lov оказалось также стабильным в клонах, выделенных после совместного культивирования, и этот уровень высокой экспрессии Lov сохранялся даже через месяц после культивирования. Вследствие относительно малого уровня индукции, дополнительные ядерные эксперименты, чтобы определить, объясняется ли индукция транскрипционным или пост-транскрипционным механизмами, не могут быть осуществлены.
мРНК Lov состоит из транскриптов двух размеров, 1,7 кв и 1,5 кв. Оба эти транскрипта равным образом индуцируются после совместного культивирования клеток SL12.4 на тимусных эпителиальных монослоях. Кроме того, оба транскрипта распознаются 5'- и 3'-зондами, происходящими от кДНК Lov. При этом, неизвестно, происходят ли эти транскрипты в результате альтернативных сайтов инициации, расщепления, сигналов полиаденилирования, или в результате каких-либо других механизмов. Однако, в других зрелых тканях, которые, как показали исследования, экспрессируют Lov /т.е. яичник, сердце/, наблюдался лишь больший транскрипт. Более того, в процессе развития мышиного эмбриона происходит изменение, заключающееся в том, что вместо экспрессии обоих транскриптов начинает экспрессироваться только больший транскрипт /примерно на 15-й день, данные не приводятся/. Возможно, что только 1,7 кв-транскрипт кодирует функциональный белок у взрослого животного.
Пример 15. Обнаружение Lov-белка на мембранах клеток SL124.
Антитела против Lov распознают эпитопы, обнаруженные на плазматических мембранах клеток SL12.4. Поликлональные антитела против 13 последних аминокислот Lov были продуцированы у кроликов с использованием в качестве антигена синтетического пептида. Эти антитела распознают антиген, ассоциированный с плазматической мембраной клеток SL12.4. Фиг. 10 иллюстрирует иммунофлуоресцентный анализ клеток SL12.4 на поверхностную экспрессию Lov. Гистограммы, изображенные на фиг. 10, представляют флуоресценцию клеток SL12.4, окрашенных иммунной сывороткой в зависимости от флуоресценции клеток SL12.4, окрашенных преимунной сывороткой. При исследовании клеток с искусственно увеличенной проницаемостью мембраны под микроскопом, связывание антитела наблюдалось лишь на плазматической мембране и не окрашивало цитоплазму. Данные, полученные в результате флуоресценции сесринского клеточного клона лимфомы SL12.3, показали, что клетки SL12.3 не экспрессируют мРНК для Lov и не обнаруживают какой-либо экспрессии Lov на их клеточной поверхности. Связывание иммунных сывороточных антител с антигеном было также специфически блокировано путем добавления конкурирующих количеств иммунизирующего олигопептида /фиг. 11/, но не какого-либо другого неспецифического олигопептида. Мембранные препараты, но не цитозольные фракции, клеток SL12.4 также обнаруживали Lov-белок при исследовании с помощью ELISA-анализа.
Антитело, продуцированное против синтетического олигопептида, соответствующего карбокси-концу Lov, специфически "узнает" эпитоп, находящийся на поверхности клеток SL12.4. Родственная клеточная линия, SL12.3, не экспрессирующая Lov-мРНК, также не экспрессирует белок, исследуемый путем флуоресценции. Были проведены иммунопреципитационные исследования Lov-белка, которые однако, не показали необычного уровня неспецифических взаимодействий с преиммунной сывороткой. Тот факт, что Lov-белок, вероятно, не имеется в изобилии по отношению к базисной экспрессии мРНК в этих клетках, еще более осложняет эти эксперименты. Тем не менее, после 35S-метионинового мечения в SL12.4 и F6 /клеточный клон с высокой степенью экспрессии Lov/ появились три специфические полосы, а в клетках SL12.3 они отсутствовали. Эти три полосы также исчезали после конкурентного добавления специфического олигонуклеотида. 32 кД-полоса соответствует предсказанной молекулярной массе стова Lov. 22 кД-полоса может быть продуктом расщепления, а 50 кД-полоса может представлять гликозилированную форму Lov.
Пример 16. Индукция поверхностной экспрессии Lov.
Уровни поверхностной экспрессии Lov были также индуцированы после совместного культивирования на тимусном эпителиальном монослое в течение 3 дней. На клетках SL12.4, после совместного культивирования на тимусном эпителиальном монослое в течение 3-х дней, проводили FACS-анализ /на клеточном сортере с возбуждением флуоресценции. - Прим. пер./. На фиг. 12 показано, что количество обнаруживаемого Lov-белка на поверхности клеток SL12.4 было индуцибельным после обработки тимусным эпителием. Представленные гистограммы иллюстрируют реакцию с антисывороткой и неиммунной сывороткой в стимулированных и нестимулированных SL12.4-клетках. Графики также иллюстрируют результаты совместного культивирования клеток SL12.4 на TEL по сравнению с результатом, полученным от необработанных клеток SL12.4 /левый график/. Эти графики построены с использованием логарифмической шкалы. TEPI-клетки показали аналогичный ответ в отношении индукции Lov.
Клеточные клоны, предварительно сокультивированные на тимусных эпителиальных монослоях, обнаруживали более высокий базальный уровень экспрессии 19.5. В некоторых клетках, совместное культивирование давало более высокий уровень экспрессии Lov. Из SL12.4, после совместного культивирования либо на TEL, либо на TEPI, был выделен ряд клеточных клонов. Клонирование осуществляли посредством лимитирующего разведения. Например, F6 являются клетками, происходящими и клонированными из клеточной популяции SL12.4, после ее совместного культивирования на TEL монослое в течение 3-х дней. Эти клеточные клоны были культивированы в нормальной среде, не содержащей стимулирующего тимусного эпителия, в течение нескольких месяцев. Нозерн-анализ РНК и FACS-анализ поверхностного белка на F6 показали, что этот клеточный клон обеспечивает более высокий стабильный уровень экспрессии Lov, чем родительские SL12.4-клетки /фиг. 13/. Другие выделенные и проанализированные клеточные клоны показали различные уровни Lov-экспрессии. Дополнительное сокультивирование этих клеточных клонов на тимусных эпителиальных монослоях не дало какого-либо значительного увеличения Lov-экспрессии.
Пример 17. Хромосомная локализация.
Геномную ДНК получали из "хомячок-мышь"-гибридных соматических клеток, содержащих комплемент специфического набора мышиных хромосом. Переваривание рестриктирующим ферментом Pst 1 дало различные рестрикционные картины для хомячка и мыши /фиг. 14, дорожки 1, 2/. Саузерн-блоты ДНК из этих гибридных клеток, зондированных Lov, показали, что лишь немногие из гибридов содержали мышиную картину гибридизации /фиг. 14/. На основании Саузерн-анализов некоторых из указанных гибридных клеточных линий, наилучшую корреляцию получали с мышиной хромосомой 16. Образец номер 77 представлял собой гибридную клетку, где хромосома 16 является лишь мышиной составляющей. В табл. 5 приводятся результаты компьютерного анализа, иллюстрирующего тот факт, что хромосома 16 имеет наименьшее расхождение для Lov в тестированных гибридных клетках. Кроме того, Lov-ген был картирован для специфической области хромосомы 16.
При картировании 19.5 Lov-гена использовали гибриды соматических клеток мыши и хомячка. Символ (+/) означает присутствие, а символ (-/) означает отсутствие мышиного рестрикционного Lov-фрагмента по отношению к присутствию (/+) или отсутствию (/-) конкретной мышиной хромосомы, указанной цифрой в левой колонке, обнаруженной путем гибридизации с 20.5-кДНК-зондом. Число наблюдаемых расхождений представляет собой сумму (+/-) и (-/+) - наблюдений. Знак /*/ означает, что ни один из этих гибридов не был кариотипирован. Эти гибриды были типированы по другим маркерам, так, что возможно, что указанные исключения представляют собой нескорректированные распределения хромосом.
Пример 18. Иммунопреципитация Lov-белка.
Для определения размера Lov-белка проводили исследования методом иммунопреципитации. В этом эксперименте использовали обе метки: 35S для вновь синтезированного полного белка и 125I для поверхностного белка. Преиммунная сыворотка от обоих кроликов имела аналогичные неспецифические полосы. Хотя в лизатах присутствовало много неспецифических полос, однако, несколько специфических полос было идентифицировано. Две полосы, соответствующие молекулярной массе 22 и 32 кД, оказались специфичными в клетках SL12.4, меченных 35S-метионином. Ни одна из полос не присутствовала ни в преиммунной сыворотке, ни в лизатах, полученных от клеток SL12.3. Однако, 2 полосы не наблюдались в 125I-меченных клетках, а вместо этого присутствовала полоса 50 кД в SL12.4, но не в SL 12.3-клетках. Были предприняты попытки уменьшить количество фоновых полос /т. е. , проводили осаждение сульфатом аммония IgG-фракций, аффинную очистку антитела на колонке с сефарозой, связанной с синтетическим олигопептидом, используя F6-клон в качестве индуктора высокого уровня экспрессии Lov/, однако, попытки снизить наблюдаемые неспецифические взаимодействия оказались безуспешными.
Таким образом, ген Lov кодирует белок, который экспрессируется на клеточной поверхности. Кроме того, уровни Lov очевидно, коррелируют со стадией созревания указанных T-клеточных линий лимфомы. После сокультивации с тимусным эпителием, в клетках SL12,4 не только происходят изменения в направлении двойного позитивного фенотипа CD4+CD8+, но в этих клетках также устанавливается повышенный уровень экспрессии Lov. Lov представляет новую кДНК и белок, не имеющие значительной гомологии с какими-либо известными последовательностями. Перед тем как была определена их функция, было описано много других T-клеточных маркеров, например, таких как Thy-1 и T-200. Хотя Lov был выделен из T-клеточной линии лимфомы, этот ген могут экспрессировать и другие клеточные типы. Картина экспрессии Lov может дать некоторые ключи к разгадке его возможной функции. К тканям, экспрессирующим наиболее высокие уровни Lov-мРНК, относятся тимус, кишечно-ассоциированные лимфоидные ткани /GALT/, и яичники. Из них, первые два вида ткани являются обычными местами "обитания" лимфоцитов. Любопытно отметить, что при инъекции клеток SL12.4 сингенным мышам, первыми очагами образования опухоли являются ткани яичника. Кроме того, яичники включают в себя экстенсивную лимфатическую систему. GALT-клетки, называемые также, пейеровыми бляшками, ограничены высокоэндотелиальными сосудами, и как было показано, антиген Mel-14 должен проходить через них. Клетки SL12.4 экспрессирует Mel-14 на своей клеточной поверхности, а поэтому, по всей вероятности должны присутствовать в вышеуказанных специализированных областях тонкой кишки. Поскольку Lov экспрессируется на поверхности клеток, то он может играть определенную роль в формировании эффекта "дома" /или "хоминге"/ лимфоцитов. Lov не был обнаружен ни в одной из популяций лимфоцитов и тимоцитов. Вероятно, он экспрессируется лишь в некоторых очень небольших субпопуляциях T-клеток и не является доминантным. Клетки SL12.4 могут представлять собой обычный редко встречающийся тимоцит.
Пример 19. Конструирование кДНК-библиотеки и скрининг 20.5-гена.
Так же как и другие клоны, конструирование кДНК-библиотеки и скрининг 20.5-гена осуществляли в соответствии с описанием в Примерах 1-4. кДНК-вставку удаляли из лямбда-ДНК путем переваривания рестриктирующими ферментами HindIII и BgIII и субклонировали в плазмидный вектор pT7/T3 /Bethesda Research/ Laboratories.
Пример 20. Анализ ДНК-последовательности 20.5 или Tea.
Проводили рестрикционно-эндонуклеазное картирование, и фрагменты субклонировали в pT7T3, плазмиду очищали на хлориде цезия, и осуществляли прямое секвенирование при помощи двухнитевого дидезокси-метода с использованием реагентов секвеназы /Seguenase/ /U.S. Biochemical Corp., Cleveland, Ohio./ Часть последовательности определяли с использованием праймеров для хозяйской плазмиды, а другие специфические олигонуклеотидные праймеры /17-меры/ получали для кДНК в лаборатории UCSD Cancer Center Core Molecular Biology Facility. Обе нити ДНК были секвенированы, и все последовательности были определены, по крайней мере, в двух реакциях в дубликате. Для сбора информации о перекрывающихся последовательности и проведения предварительного анализа ДНК-последовательности использовали компьютерные программы Microgenie.
Пример 21. Генетическое картирование 20.5
Получение и характеризация гибридов соматических клеток китайского хомячка и мыши были ранее описаны Hoggan и др., J. Virol. 62: 1055-1056 (1988). Штамм мыши NF/N был получен от Division of Natural Resources, NIH, Bethesda, Мериленд. Мышей Mus musculus mis, получали из лабораторной колонии, происходящей от мыши, пойманной в Скиве (Дания), и сохраненной Dr. M. Potter (NCI, NIH, Contract N01-CD2-5584) в лабораториях Hazelton, Роквилл, Мериленд. Для получения экспериментальных животных, самок гибридной линии NF /N • X.m.musculus подвергали обратному скрещиванию с самцами M.m. musculus. ДНК экстрагировали из мышиной печени, переваривали Sacl и BamHI, подвергали электрофорезу в 0,4% агарозном геле в течение 48 ч при 24В и переносили на найлоновые мембраны (Hybond N+, Amersham). Мембраны гибридизировали с 32P-меченной кДНК 20.5 и зондом 438 п.о., представляющим собой ген ДНК-полимеразы B, который присутствует на хромосоме 8 (MacBride и др., в печати). Мембраны промывали и зондировали в соответствии с приведенными ранее описаниями. Образцы почки, взятые от тех же мышей, типировали на наследование маркеров, Gr-1 и Es-1 путем гистохимического окрашивания после проведения электрофореза на крахмальных гелях (Harris и Hopkinson "Handbook of Enzyme Electrophoresis Human Genetic" North Holland Publishing Co., Амстердам (1976)).
Пример 22. Выделение и ДНК-последовательность кДНК-клона 20.5.
40000-член кДНК-библиотеки SL12.4 в ggt10 (16.24) скринировали с использованием кДНК-зонда SL12.4, субстрактивного против SL12.3-мРНК, и одновременно на дубликатных фильтрах с полной кДНК SL12.3, в соответствии с приведенными выше описаниями. Этот метод скринирования использовали для характеризации 20.5-гена. Исходя из экспрессирующих свойств, описанных ниже, кДНК-клон обозначали Tea (ранний ген активации T-клеток). Вставку из этого клона секвенировали на обеих нитях; полученная последовательность представлена на фиг. 15-18. кДНК-вставка составляет 2397 пар оснований в длину и содержит одну длинную открытую рамку считывания, начиная от пары оснований 409 и простираясь до 1769. кДНК не включает в себя сигнальную последовательность полиаденирования или poly A-участок. кДНК-последовательность 20.5 позволяет предсказать множественный мембранный белок с промежуточными областями. На фиг. 15 - 18 показана предсказанная 453-аминокислотная последовательность предсказанного белка с молекулярной массой 49,57 кДа (немодифицированный). Очевидно, кДНК-последовательность содержит полную кодирующую область, поскольку предсказанный T-концевой метиониновый кодон окружен обобщающей последовательностью Kozak (GXC AUG, где X может быть A, U, G или C), которая является областью инициации оптимальной транляции, а предсказанные 5'- и 3'-нетранслированные области содержат несколько стоп-кодонов во всех трех рамках считывания. Физические свойства предсказанного белка анализировали с помощью программ Microggenie и PC-ген. На фиг. 15 - 18 звездочками отмечены три потенциальные сайта N-гликосилирования. Предсказанный белок имеет девять в высокой степени гидрофобных областей (на фиг. 15 - 18 подчеркнуты), семь из которых обладают свойствами трансмембранных промежуточных областей, как было показано анализом с использованием программ системы программного обеспечения Intelli - Genetics : SOAP, HELIXMEM, NOVOTNY и PAOARGOS.
Tea-ген дифференциально экспрессировался в T-клетках лимфомы и активировал T-лимфоидные клетки от нормальной селезенки. Анализировали экспрессию транскриптов, распознаваемых кДНК-клоном 20.5. Фиг 19 иллюстрирует экспрессию Tea-гена. Очищенную вставку, содержащую полную кодирующую область, 32P-метили путем произвольного праймирования и использовали для зондирования Нозерн-блотов, в которых каждая дорожка содержит 10 мкг полной клеточной РНК или (см. фиг. 19b, цитоплазматической и ядерной РНК), происходящей от клеток SL12.4, клеток SL12.3, указанных клеточных линиях или тканей от нормальных Balb /c-мышей. Авторадиограммы Нозерн-блотов показаны на рис. a-d. Фиг. 19a иллюстрируется сравнение экспрессии Tea-мРНК в клеточных линиях SL 12.3 и SL12.4. На фиг. 19c проиллюстрирована экспрессия Tea-мРНК в ряде мышиных клеточных линий, указанных в надписях, а на фиг. 19d показана РНК от указанных нормальных тканей мыши Balb /c (GALT - кишечно-ассоциированная лимфоидная ткань). На каждой фигуре указан размер транскриптов (в кв = тысяча пар оснований), который был оценен по их относительной миграции в отношении BRL-маркерам и эндогенным рибосомным РНК 18 и 28S. Эквивалентную нагрузку и перенос РНК во всех дорожках оценивали путем окрашивания акридиновым оранжевым и путем гибридизации с 32P-циклифиллин (Cyc)- и/или 32P-Cho-A-меченной кДНК.
20.5-кДНК-зонд "узнавал" два транскрипта приблизительно 4 кв и 7 кв, которые присутствуют в клетках SL12.4, но не в клетках SL12.3 T-лимфомы (фиг. 14a). Оценивали субклеточную локализацию двух указанных транскриптов для того, чтобы определить, имеются ли оба зрелых транскрипта в цитоплазме, или большая РНК является ядерным предшественником. Оказалось, что оба транскрипта являются полностью процессированными РНК, поскольку они были обнаружены в цитоплазме (фиг. 19b). Происхождение этих двух транскриптов не совсем понятно; они, по всей вероятности, могут образовываться в результате альтернативной инициации транскрипции, альтернативного сплайсинга транскрипта, или использования альтернативных сигналов полиаденилинирования. Оба зрелых цитоплазматических транскрипта (7 и 4 кв) по своему размеру больше кДНК-клона (2,4 кв). Таким образом, кДНК не является полностью процессированной, хотя она содержит полную кодирующую область. Оба транскрипта обнаруживаются с помощью зондов, полученных для 5' - (нуклеотиды 1/380) и 3' -(нуклеотиды 2005 - 2394) области кДНК-клона, что указывает на то, что кДНК не является клонирующим артефактом, который соединяет два различных транскрипта. Помимо указанных двух зрелых РНК, в ядерных и полноклеточных РНК-препаратах имеются более крупные, гораздо более редко встречающиеся транскрипты, которые могут быть ядерными предшественниками, не подвергавшимися или частично подвергающимися сплайсингу (фиг. 19b).
Несколько мышиных клеточных линий эмбрионного (F9 и PC4), эпителиального (эпителия молочной железы (MME) и нейронного (ATt-20) происхождения были исследованы на экспрессию Tea-РНК. Ни одна из этих клеточных линий не экспрессировала обнаруживаемую Tea-РНК. В противоположность этому, клеточные линии тримусного эпителия (TEPI) и фибробласта (3T3), MEF) содержали Tea-мРНК, хотя она гораздо чаще встречается в T-клеточной лимфоме SL 12.4 (фиг. 19c).
Для того, чтобы определить, экспрессируют ли нормальные ткани и клетки лимфоидного происхождения Tea-ген, исследовали Нозерн-блоты, полученные от мРНК мышиных тканей. В клетках тимуса, неактивированной селезенки, кишечно-ассоциированной лимфоидной ткани GALT и костного мозга отсутствовала обнаруживаемая экспрессия Tea-мРНК (фиг. 12). Однако, Tea-транскрипты индуцировались в нормальных клетках селезенки, активированных T-клеточным митогеном, Конканавалином А (Кон А, фиг. 19d). Кон А использовали для моделирования активации T-клеток селезенки, обычно происходящей специфическим образом в клеточном клоне после соответствующей презентации чужеродного антигена. Из испытываемых тканей, которые экспрессировали умеренные количества Tea-мРНК, нелимфоидной тканью была только печень. Tea-транскрипты не были обнаружены в кишках, желудке, яичнике (фиг. 19d), головном мозге, сердце, легких, почках, поджелудочной железе или яичках. Таким образом, экспрессия Tea-гена ограничивается несколькими типами клеток, такими, как активированные клетки селезенки, тимусные эпителиальные клетки, клетки-T-лимфомы и печени.
Пример 23. Индукция tea-мРНК.
Для дополнительного исследования индукции Tea-мРНК, РНК получали через 6, 24, 48 и 72 ч. после добавления Кон А к спленоцитам. На фиг. 20 показана кинетика индукции Tea-гена в активированных спленоцитах. Проводили Нозерн-анализы РНК, полученной от неактивных и активированных клеток селезенки, собранных в указанные интервалы времени, после активации T-клеточным митогеном Кон А. На фиг. 20a указан период времени 72 ч. На фиг. 20b показан другой блок РНК вместе с контрольным зондом циклофиллином (Cyc) для иллюстрации относительного количества мРНК, загруженной в каждой дорожке. В отличие от Cyc-мРНК, загрузка рРНК в каждой дорожке была эквивалентной, как было показано посредством окрашивания акридиновым оранжевым. Tea-транскрипты не обнаруживались в неактивных лимфоцитах селезенки, и стали обнаруживаться лишь через 6 ч. после активации, достигая пика через 48 ч. Хотя полное количество РНК является эквивалентным для каждой дорожки (10 мкг), как было показано путем окрашивания акридиновым оранжевым, однако, относительное количество рибосомной РНК по отношению к мРНК изменяется в процессе активации T-клеток, поскольку количество актинового, CHO-A- и циклофиллинового транскриптов/10 мкг всех РНК возрастает (фиг. 20). Однако, факт индуцирования экспрессии Tea-гена по отношению к указанным контрольным РНК в процессе T-клеточной активации совершенно очевиден.
Пример 24. Гомология 20.5-ДНК и аминокислотной последовательности с мышиным экотропным ретровирусным рецептором.
Исследования гомологии с использованием базы данных Bionet не обнаружили значительного сходства между 20-5-ДНК- и другими ДНК-последовательностями, описанными ранее. Однако, сравнение вышеуказанной последовательности с полученным недавно (Albritton и др. Cell 57, 659-666 (1989)) кДНК-клоном мышиного экотропного ретровирусного рецептора (ERR) обнаружило значительную гомологию этих последовательностей. Ген, кодирующий экотропный ретровирусный рецептор, был обозначен Rec-1. На фиг. 21-24 иллюстрируется сравнение 20.5-кДНК-последовательности с ERR-кДНК-последовательностью.
На фиг. 21-24 продемонстрирован сравнительный анализ 20.5- и ERR-кДНК-последовательностей. На фиг. 21, области ERR- и 20.5-кДНК-последовательностей, совпадающие в указанном сравнительном анализе, указаны вертикальными черточками. Для каждой кДНК указаны открытые рамки считывания. На фиг. 23 полная кДНК-последовательность 20.5 показана в верхней строке из каждой пары строк, а кДНК-последовательность ERR (400 - 2425 п.о.), в которой отсутствуют первые 400 пар оснований, показана в нижней строке каждой пары. Горизонтальными линиями отмечены положения идентичности последовательностей. Сравнение проводили с помощью программы Microgenie, специально образуя, при этом, бреши для сравнительного анализа наиболее высокогомологичных последовательностей. Процент идентичности определяли только лишь для явно перекрывающихся областей кДНК. Следует отметить, что 20.5-кДНК гораздо длиннее у своего 3'-конца, а ERR-кДНК гораздо длиннее у своего 5'-конца.
На схематическом изображении двух кДНК (фиг. 21-24) показано, что хотя ERR-кДНК длиннее у 5'-конца, а 20.5-кДНК длиннее у 3'-конца, однако, в целом, обе кДНК имеют аналогичную длину. Кодирующая область каждой последовательности показана в виде затрированного участка каждого кДНК-клона. Сравнительный анализ показал, что идентичность ДНК-последовательностей между перекрывающимися областями 20.5-кДНК (1-2047 п.о.)- и ERR-кДНК (400-2425 п.о.) составляет 59%; а область 20.5- кДНК (1011-1088 п.о.) имеет идентичность на 80%. 5'-некодирующая область 20.5-кДНК-последовательности (1-410 п.о.) на 68% идентична перекрывающейся 5'-кодирующей области ERR-кДНК-последовательности, что позволяет предположить, что эти два гена произошли от общей последовательности посредством дупликации гена.
Предсказанный Tea-белок имеет структурное сходство с другими белками в отношении их множественных мембранных промежуточных областей, таких как трансдуцирующие белки, ионные каналы, ионные насосы и переносчики сахара. Однако, при этом не было обнаружено сколь-нибудь значительного сходства этой последовательности ни с семействами этого гена, ни с другими известными белковыми последовательностями, за тем лишь существенным исключением, что предсказанный ERR-белок происходит от ERR-кДНК-последовательности. Сравнительный анализ предсказанного Tea-белка и ERR-белка выявил две области аминокислотной последовательности с высокой степенью сходства (фиг. 26-28).
На фиг. 26 проиллюстрирован сравнительный анализ предсказанной последовательности Tea-белка и последовательности мышиного экотропного ретровирусного рецептора. На фиг. 26 схематически показана область двух сравниваемых белковых продуктов; Tea-белок простирается от аминокислот 1 - 404, а ERR-белок от аминокислот 204 - 603. На фиг. 27 проиллюстрирован сравнительный анализ двух предсказанных белков, где аминокислотная последовательность 20.5-кДНК показана в верхней строке, а последовательность ERR-кДНК показана в нижней строке. Жирными скобками выделены две области аминокислотных последовательностей с высокой степенью идентичности: Область 1 имеет 81,3% идентичности на протяжении 192 аминокислот, Область 2 имеет 51,9% идентичности на протяжении 79 аминокислот. Область 1, указанная в скобках, имеет 81% идентичности и 91% схожести на протяжении 193 аминокислот. Область 2 имеет 62% идентичности и 75% схожести на протяжении 60 аминокислот. Различия консервативных аминокислот (Doolittle R. Of Urfs and Orts : A primer on how to analyze derived amino acid Seguences. (University Science Books, Mill Valley, Калифорния 1986) указаны двоеточием. Аминокислотные соответствия показаны вертикальными черточками.
В противоположность предсказанному Tea-генному продукту, EPP-белок является крупнее и имеет 13-14 предсказанных трансмембранных промежуточных областей. Кроме того, он имеет более короткий гидрофильный карбокси-конец, а его амино-конец имеет такую же длину. Карбокси- и амино-концы указанных белков обнаруживают наибольшее расхождение в своих последовательностях. Аналогично ERR и другим белкам, имеющим множество мембранных промежуточных областей, продукт Tea-гена не содержит сигнальной последовательности. Однако, несмотря на наличие областей с высокой степенью идентичности, эти два белка представляют собой явно различные генные продукты.
На фиг. 29 схематически представлена структура белка, полученная с использованием программ PC-ген для оценки физических свойств, обозначенных надписями на фигуре.
На фиг. 29 проиллюстрированы физические свойства предсказанных белковых продуктов Tea- и Rec-1-генов. На фиг. 29a представлена модель возможной структуры Tea-белка. Жирными линиями показаны две области, имеющие высокую степень идентичности с ERR-белком. Для оценки физических свойств предсказанного белка при построении указанной модели использовали PC-ген-программы из Intelli Genetics : SOAP, HELIXMEM, NOVOTNY, PAOAPGOS.
На фиг. 29b показана область двух предсказанных белков, которые анализировали для сравнения их гидрофобности (аминокислоты 1 - 403 для Tea-белка, и аминокислоты 204-603 для ERR-белка). На фиг. 30 представлены диаграммы гидрофобности предсказанного Tea-белка (верхний график) и предсказанного ERR-белка (нижний график). Представленная схематическая модель явно иллюстрирует области высокой степени идентичности с ERR-белком. Поскольку ERR кодирует клеточный поверхностный белок, то были проведены сравнения профилей гидрофобности двух предсказанных белков в областях перекрывания (аминокислоты 1-401 Tea-белка и аминокислоты 204-600 ERR-белка).
На фиг. 29 в верхней части показана область, в которой сравнивались два белка для построения диаграмм гидрофобности. Это сравнение покрывает область из 401 сопряженных аминокислот и обнаруживает значительную идентичность. Таким образом, продукт tea-гена, по всей вероятности, также является клеточно-поверхностным белком.
У высших позвоночных, Tea-ген, очевидно, является консервативным. Для того, чтобы определить, являются ли Tea-генные последовательности консервативными в процессе эволюции, ДНК от человека, хомячка, мыши и курицы испытывали на перекрестную гибридизацию с 20.5-кДНК-зондом. При использовании непроцессированного кДНК-клона для зондирования Саузерн-блотов, во всех испытываемых геномных ДНК наблюдалась заметная гибридизация (фиг. 31-32).
На фиг. 31-32 проиллюстрирован Саузерн-анализ ДНК от различных видов и анализ рекомбинантной инбредной ДНК для определения местоположения Tea-гена на хромосоме 8. 32P-меченный кДНК-зонд, используемый для обоих анализов, представлял собой почти полный Ball-фрагмент 20.5-кДНК-клона. На фиг. 31 A показана авторадиограмма, иллюстрирующая картину гибридизации Pst1-переваренной ДНК человека, хомячка, мыши и курицы, которую зондировали с использованием Ball-фрагмента, описанного в Материалах и Методах.
На фиг. 32 представлена авторадиограмма ДНК печени, происходящей от рекомбинантных инбредных животных Фиг. 31-32 иллюстрирует пример картины гибридизации, полученной от нескольких вышеуказанных рекомбинантных животных и от NFS/N - родительской ДНК, переваренной Sacl.
Для определения любые ли обнаруженные ДНК-фрагменты происходят от Rec-1 -гена, проводили испытания, которые показали, что зонд к 3'-области 20.5-кДНК-клона (который обнаруживает значительное расхождение с ERR-кДНК-последовательностью) гибридизируется с ДНК-фрагментами одного и того же размера. Эти результаты позволяют предположить, что последовательности, обнаруженные у дивергентного 3'-конца, также являются консервативными в процессе эволюции, и что установленная перекрестная гибридизация ДНК других видов имела место с наиболее дивергентной последовательностью, находящейся между двумя кДНК-клонами. Эти данные, взятые вместе с данными, полученными в предыдущих разделах, свидетельствуют о том, что Tea-ген кодирует белок, отличающийся от ERR, который обнаруживается в других видах. Расхождение в аминокислотных последовательностях в амино- и карбокси-концах позволяет предположить, что эти два белка могут иметь различные функции.
Пример 25. Картирование Tea-гена для хромосомы 8.
Поскольку продукт ERR-гена действует как вирусный рецептор, Tea-ген картировали для определения, находится ли этот ген на хромосоме, которая кодирует один из других известных ретровирусных рецепторов (Kozak (1983) J. Virol. 48: 300-303). Несколько различных гибридов, образованных от соматических клеток китайского хомячка и мыши, (каждый) содержат ограниченное количество различных мышиных хромосом. Эти гибриды использовали для картирования Tea-гена (Hoggan и др., (1988) T. Virol. 62: 1055-1056). Саузерн-анализы гибридных клеточных ДНК, переваренных Psr 1, показали, что 7 из 21 гибрида содержали мышь-специфические ДНК-фрагменты. Сравнение состава известных мышиных хромосом посредством положительной гибридизации с мышь-специфическими ДНК-фрагментами показало наилучшую корреляцию с мышиной хромосомой 8 (табл. 6).
При картировании tea-гена использовали гибриды соматических клеток мыши и хомячка. Символы в заголовках граф указывают на присутствие (+/) или отсутствие (-/) мышиного рестрикционного Tea-фрагмента по отношению к присутствию (/+) или (отсутствию (/-) конкретной мышиной хромосомы, указанной цифрой в левой колонке и обнаруженной путем гибридизации с 20.5-кДНК-зондом. Число несоответствий представляет сумму (+/-) и (-/+)-наблюдений.
Знак (+) означает, что ни один из этих гибридов не был кариотипирован. Они были типированы для других маркеров. Таким образом, возможно, что (+/-) гибридные клетки содержат фрагменты хромосомы 8 или небольшой процент клеток содержит эту хромосому. В качестве исключения (-/+) может содержать часть хромосомы 8, но при этом не содержит области, включающей в себя Tea-ген.
Для подтверждения этого наблюдения, определяли положение Tea-гена на хромосоме 8 с помощью анализа путем межвидового обратного скрещивания. Было показано, что Sst1-переваренная ДНК NFS/N-мыши продуцирует перекрестно-реактивные полосы 10,0, 7,4 и 5,5 кв, ДНК M.m. musculus продуцирует вставка гибридизации 20.5-кДНК-зонда при обратном скрещивании мышей NFS\N • M.m musculus F1 с мышами M. m. musculus. Картина сегрегации полиформизма этих рестрикционных фрагментов по другим маркерам на хромосоме 8 свидетельствует о том, что этот ген связан с Gr-1 и Es-1 в следующем порядке:
центромер Polb-Gr-I-Tea-ES-I (табл. 7 и 8).
Вставка: "...10,0, 6,4 и 5,5 кв - фрагменты. На фиг. 32 показана картина ..."
Расположение Tea-гена на хромосоме 8 свидетельствует о том, что этот ген отличается от гена Rec-1, который кодирует ERR, и расположен на хромосоме 5. Это также исключает возможность того, что Tea является MCF-ретровирусным рецептором, который расположен на хромосоме 1, хотя пока еще не имеется достаточных данных для определения, является ли он родственным рецептору для вируса A/10, который еще не был картирован на хромосомах.
Известно, что 70 генов или генных продуктов обнаруживают возрастание в экспрессии, если T-клетки являются активированными в ответ на воздействие антигена вместе с молекулами аутогистосовместимости на поверхности антиген-представляющих клеток. Поликлональные активаторы, такие, как лектины, кальций, ионофоры, или антитела к T-клеточному рецептору для антигена, могут имитировать ответ, индуцированный антигеном, Некоторые из этих генов "активации" участвуют в переходе между G0 и G1 клеточного цикла. Некоторые кодируют цитокины и их рецепторы, ядерные регуляторные белки, а другие участвуют в транспорте ионов и питательных веществ в клетки для стимуляции их роста. По крайней мере, 26 генных продуктов "активации" T-клеток были локализированы для клеточной мембраны. 20.5 является первым примером клонированного гена или кДНК, который (которая) обладает способностью кодировать трансмембранный белок с множеством промежуточных доменов, индуцируемый в процессе активации T-клеток.
Процесс активации T-клеток селезенки, инициированной КонА или антигеном, начинается через несколько минут после начала взаимодействия с пектином или после презентации антигена и продолжается в течение около 7 - 14 дней. Изменения в экспрессии гена происходят на протяжении всего периода активации. Crabtree классифицировал эти изменения в генной экспрессии по аналогии с активацией вирусного гена (немедленная, ранняя, поздняя и очень поздняя). По критерию Crabtree, Tea является ранним геном, поскольку Tea-мРНК, фактически, не обнаруживается в нормальных покоящихся T-клетках, и ее экспрессия возрастает до обнаружимого уровня через 6 ч. после активации, достигая пикового значения, примерно через 24 ч. Функция Tea-гена остается пока неясной; по всей вероятности, она заключается в трансдукции сигналов или в транспорте небольших молекул, которые являются сигнальными трансдукторами, либо этот ген играет роль рецептора для неидентифицировнаного лиганда. Довольно длинный карбокси-конец предлагаемого Tea-белка может служить в качестве сигнального трансдуктора, хотя подтверждения этого предположения пока не найдено. Поскольку T-клетки лимфомы SL12.3 и многочисленные другие T- и B-клеточные опухолевые линии не экспрессируют этот ген, то можно сделать вывод, что для клеточного роста не требуется экспрессия Tea-гена. Однако, исследования показали, что не исключена возможность того, что экспрессия Tea-гена необходима для пролиферации нормальных клеток, которая сопровождает активацию T-клеток. Подавление экспрессии этого гена может блокировать активацию T-клеток при аутоиммунных заболеваниях, таких, как артриты, диабет и другие иммунные заболевания, опосредованные T-клетками.
Высокая степень гомологии между Tea- и ERR-белками позволяет предположить, что Tea-генный продукт может служить в качестве мышиного ретровирусного рецептора, так как, по крайней мере, 4 класса мышиных ретровирусных рецепторов были определены генетически, и лишь один был молекулярно клонирован. Локализация Tea-гена на хромосоме 8 исключает возможность того, что Tea кодирует рецептор для рекомбинантного экотропного вируса MCF, но допускает возможность того, что этот ген кодирует рецептор для амфотропного вируса, расположенный в хромосоме 8. В отличие от Rec-1-гена (кодирующего ERR), который экспрессируется во всех мышиных тканях, Tea-ген имеет гораздо более ограниченное распределение в различных тканях. Если Tea-ген кодирует белок, действующий как ретровирусный рецептор, то такое ограниченное тканевое распределение должно обеспечивать его специфичность для ретровирусов лишь лимфоидного происхождения. В качестве прецедента для рецептор-опосредованного ограничения можно указать на ретровирус человека ВИЧ, который использует белок CD4 в качестве своего раннего рецептора.
Однако, маловероятно, чтобы клеточно-специфические рецепторы были полностью ответственны за тканевую специфичность мышиных ретровирусов. Тканевый тропизм, обнаруживаемый ретровирусами, является, очевидно, результатом воздействия целого ряда факторов, таких, как тканевая специфичность LTR, изменчивость белков env вируса gp 70, клеточные факторы, и/или экспрессия соответствующих поверхностно-клеточных рецепторов. Если будет идентифицирован сайт ERR-связывания для вируса, то можно будет определить, находится ли это сайт в области высокой степени гомологичности с Tea-белком. Исследования вирусного связывания и инфицирования необходимы для того, чтобы определить, играет ли Tea-белок роль вирусного рецептора.
Несмотря на высокую степень сходства с ERR-генным продуктом, физические свойства Tea- и ERR-белков значительно отличаются друг от друга. Они имеют различные хромосомные карты и различные картины тканевой экспрессии. Поскольку естественные клеточные функции обоих этих генов неизвестны, то необходимо провести дополнительные исследования для того, чтобы выяснить, имеется ли какое-либо функциональное сходство между этими двумя белками. Анализ консервативных и неконсервативных аминокислот может внести ясность в определение сходных или различных функций ERR-белка и Tea-генного продукта. В частности, идентификация этого нового семейства генов и участков ДНК-последовательности, которые являются в высокой степени консервативными для этих двух молекул, может способствовать поиску новых членов этого семейства генов.
Векторы, обозначенные pT7T3-19.1 (входящий N 68298), pT7T3-19.2 (входящий N 69299), pT7T3-19.4 (входящие NN 68300, 68301, 68302), pT7T3-19.5 (входящий N 68303) и pT7T3-20.5 (входящий N 68305) были депонированы в Американской коллекции типовых культур (АТСС, Роквилл, Мэриленд, США) 13 апреля 1990 г. Эти депозиты являются доступными согласно правилам выдачи и законам США и тех иностранных государств, где были поданы дубликаты данной заявки. Доступность депозита, однако, не дает права на применение изобретения данной заявки при аннулировании любого патента, выданного на эту заявку, или любого разделения или продолжения этой заявки.


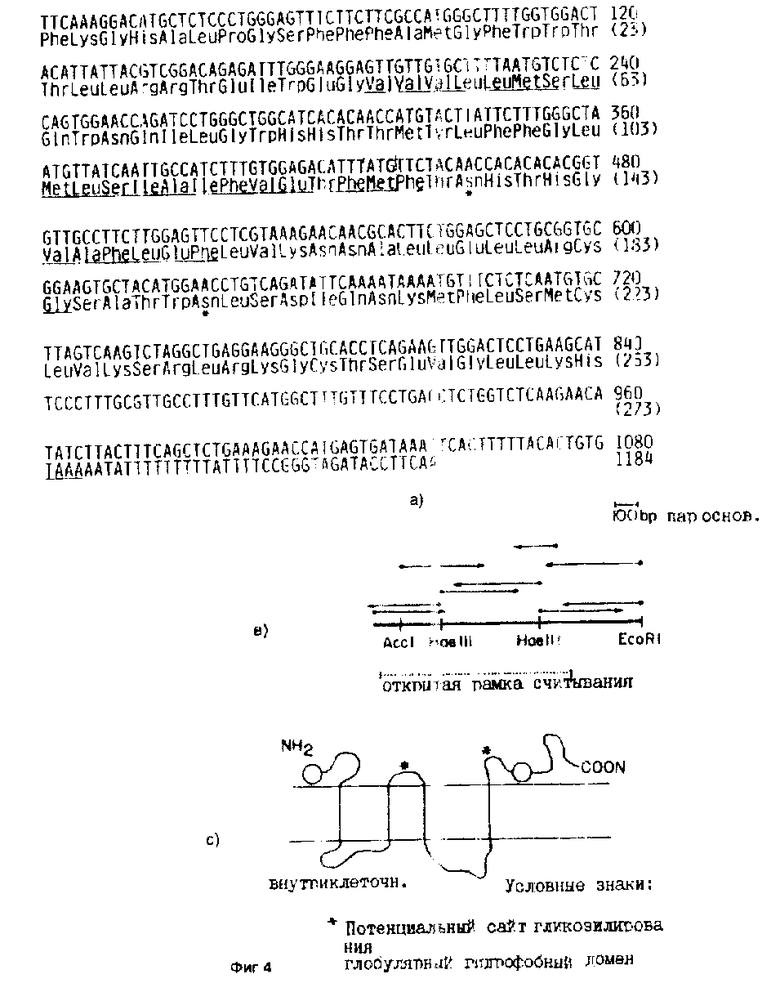

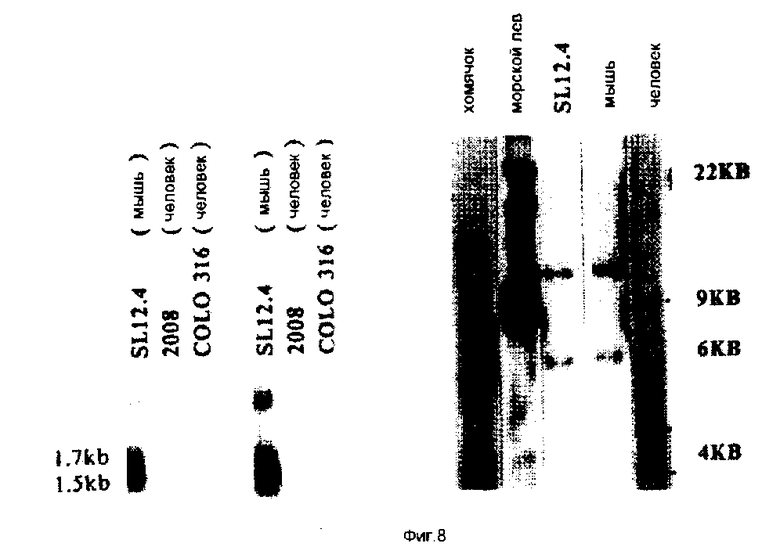
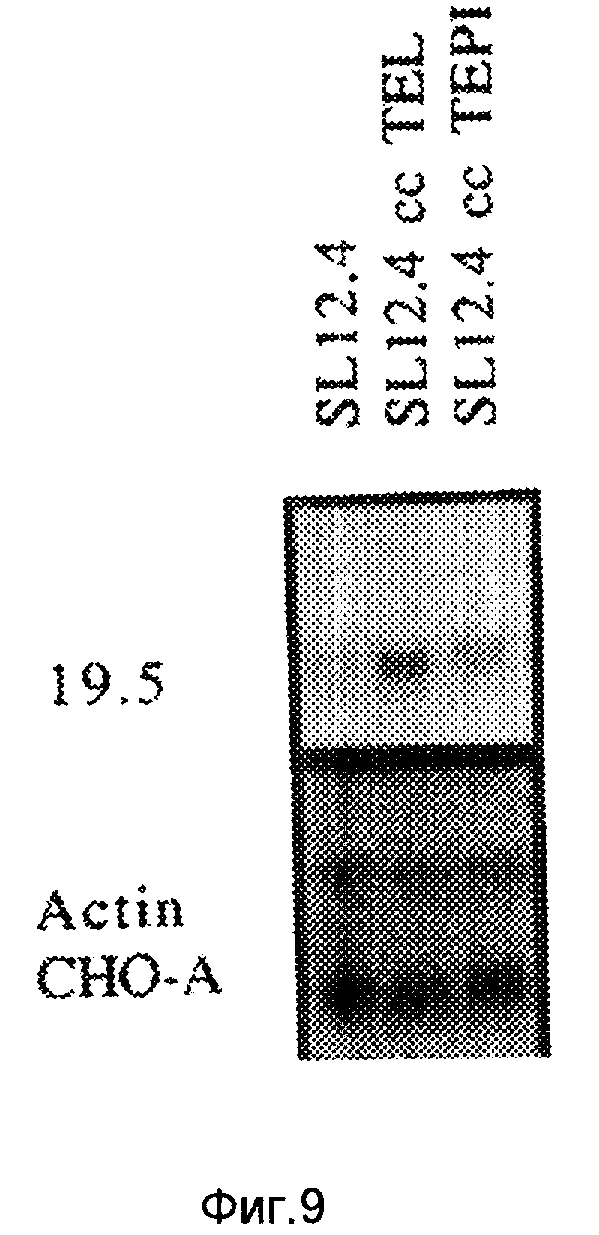

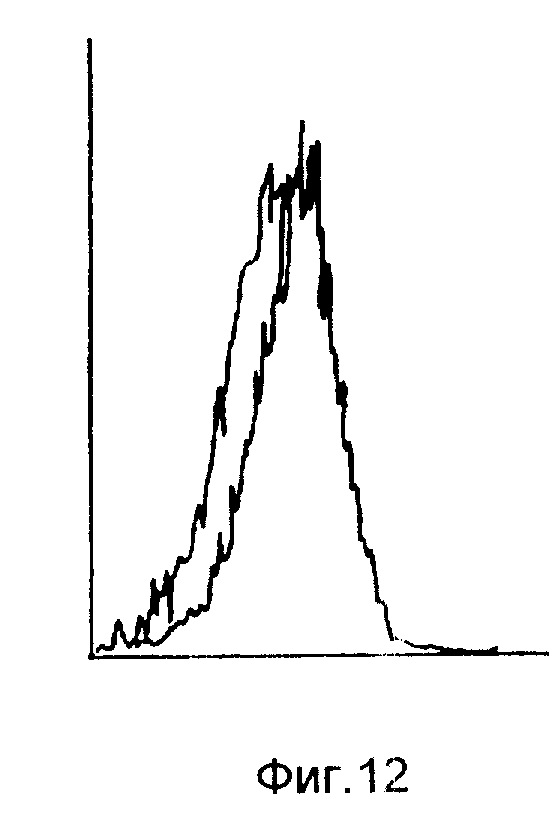






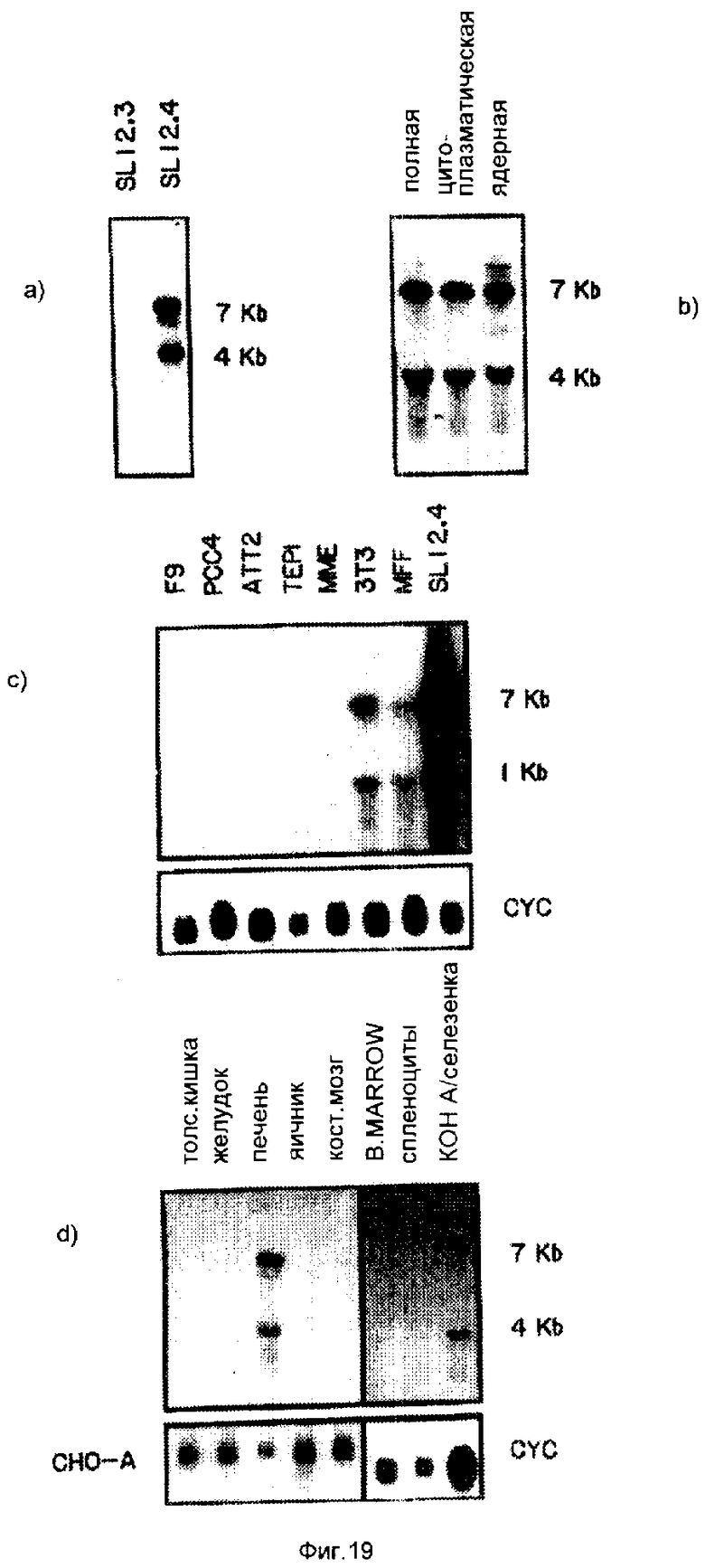








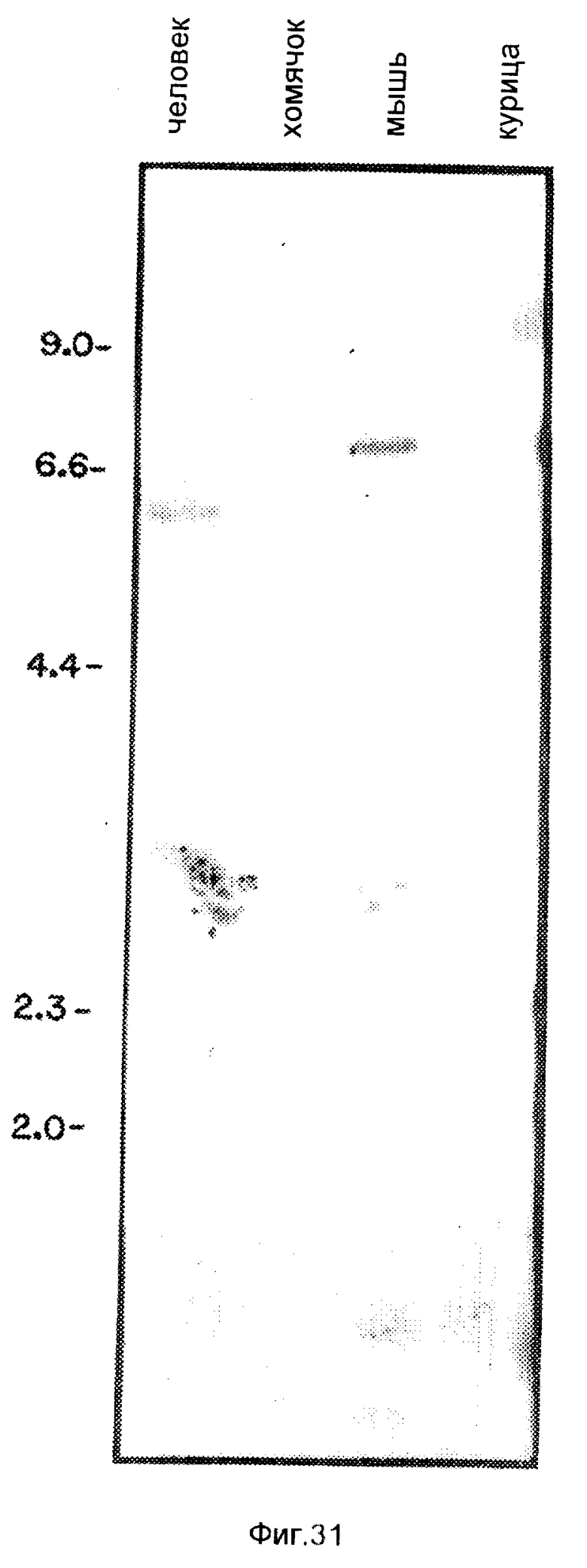





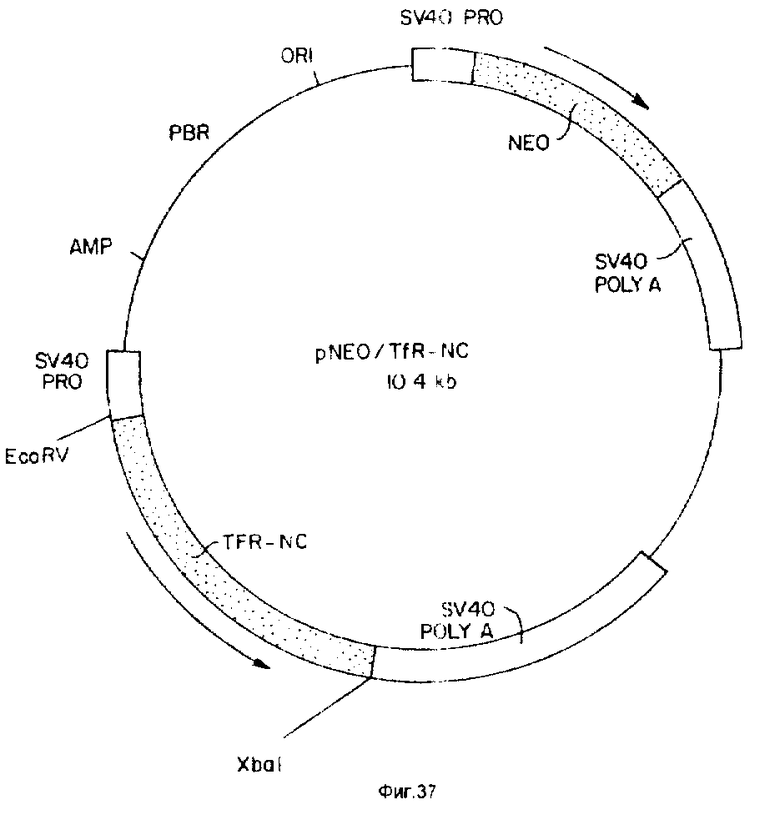

Изобретение относится к биотехнологии, в частности к рекомбинантным векторам клонирования, и может быть использовано при манипулировании регуляцией Т-клеточного роста, для модификации онкогенного фенотипа, а также для локализации метастатических очагов опухолей. Рекомбинантный вектор клонирования состоит из Hind III - Bam H I фрагмента плазмиды pT7T3 и фрагмента ДНК, кодирующего белок T-лимфы, и имеет определенную нуклеотидную последовательность. 4 с.п. ф-лы, 38 ил., 8 табл.
| WO, международная заявка, 90/02180, C 12 N 15/12, 1990. |

