Настоящее изобретение относится к области биологии и терапии рака. Главным образом, настоящее изобретение относится к способу замедления или нормализации скорости роста клетки, особенно злокачественной клетки, в результате воздействия на такую клетку эффективных количеств эктодоменовой части синдекана-1. Способ настоящего изобретения облегчает и обеспечивает нормализацию скорости роста и состояние дифференциации злокачественных клеток.
Описание уровня техники
Клеточная дифференцировка основана на селективном использовании генетической информации, запрограмированной внеклеточными раздражителями, которые, например, могут включать клеточные взаимодействия и связывание молекул внеклеточного эффектора поверхностными рецепторами клетки. Становится все более очевидным, что протеогликаны клеточной поверхности играют важную роль в регуляции поведения клеток. Синдеканы представляют собой поверхностные протеогликаны клеток, которые, как было показано, участвуют как в распознавании матрицы, так и в связывании фактора роста, на основании чего предполагается, что они участвуют в регуляции клеток. Известны последовательности человеческого, мышиного, крысиного и хомячьего синдеканов. Недавно появились обзоры по синдеканам (Jalkanen с сотр. в кн. "Рецепторы для внеклеточной матрицы", изд. Дж. Макдональд и Р. Мэшэм, Академик Пресс, Сан-Диего, стр. 1-37 (1991) и Bernfield О., с сотр., Annu. Rev. Cell Biol. 8:365-393 (1992)).
Синдекан-1 представляет собой наиболее хорошо охарактеризованный клеточный поверхностный протеогликан (Saunders с сотр., J. Cell Biol. 108:1547-1556 (1989); Mali с сотр., J. Biol. Chem. 265:6884-6889 (1990)).
В международной заявке на патент WO 90/12033 раскрывается аминокислотная последовательность и последовательность соответствующей кДНК молекулы мышиного синдекана-1. Диагностический метод детекции трансформированных клеток по детекции изменений представляет собой экспрессию синдекана в трансформированных клетках, и такой метод описан в международных заявках на патент WO 92/13274 и WO 93/05167.
Энхансер синдеканового гена, а также способ уменьшения роста злокачественных клеток путем индукции экспрессии синдекана в таких злокачественных клетках описаны в международной заявке на патент (PCT/FI93/00514).
Краткое описание чертежей
Фиг. 1 изображает последовательность человеческого синдекана-1.
Кружки: возможные сайты GAG присоединения; подчеркнуто длинной линией: трансмембранный домен; подчеркнуто короткой линией: сигнал aataa полиадениляции.
На фиг. 2 изображена последовательность мышиного синдекана-1.
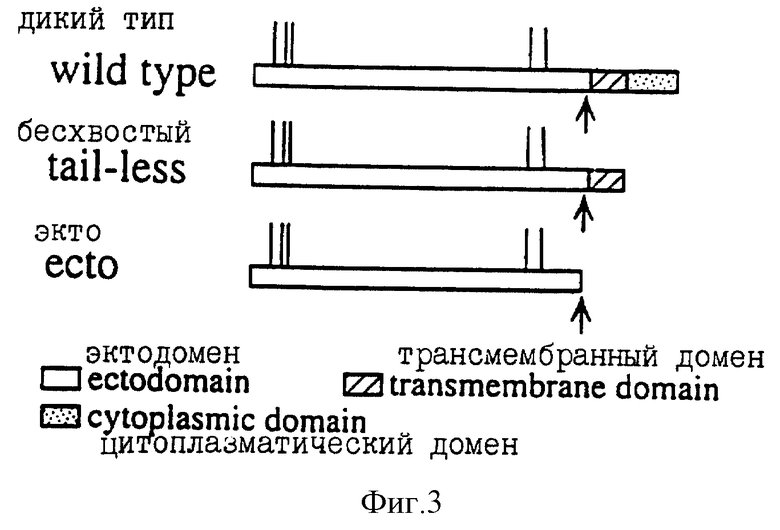
Фиг. 3 изображает схематизированную структуру капсидных белков дикого типа, бесхвостных и экто трансфекционных конструкций. Конструкция дикого типа содержит эктодомен мышиного синдекана полной длины (Mali, М. с сотр., J. Biol. Chem. 268:24215 (1993)). Бесхвостная конструкция образуется с использованием сайтнаправленного мутагенеза с использованием олигонуклеотида с образованием делеционного мутанта с одним аргининовым остатком в цитоплазматическом домене в соответствии с описанным в примерах (Miettinen, Н.М. с сотр., J. Cell Sci., в печати (1994)). Эктоконструкция также может быть получена с использованием описанного в примерах сайтнаправленного мутагенеза с применением олигонуклеотида, и она имеет стоп-кодон в протеаза-чувствительном сайте, соседствующем с поверхностью клетки, вертикальные линии обозначают предполагаемые сайты GAG присоединения, а стрелки - сайт, чувствительный к двухосновной протеазе.
На фиг. 4 изображена организация актинового филамента и иммунофлуоресцентная локализация синдекана-1 на поверхности клетки.
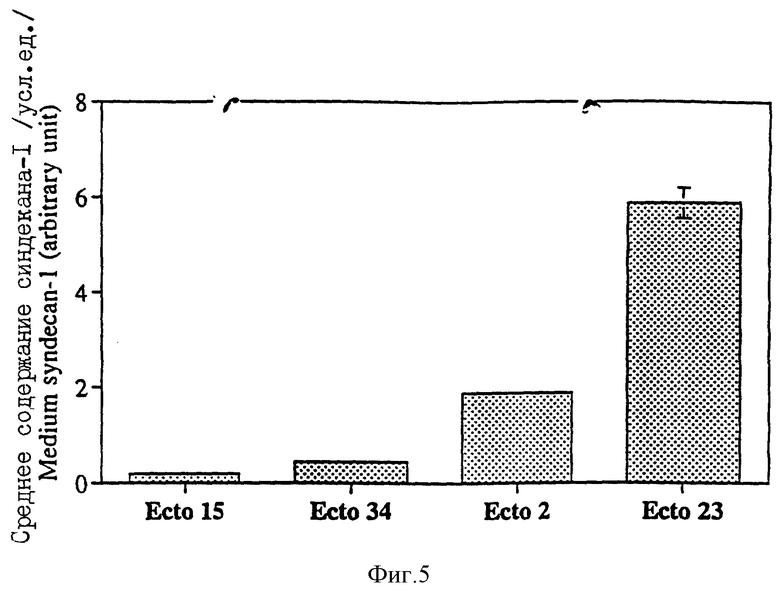
Фиг. 5 - количество секретированного эктодомена синдекана-1 из кондиционированной среды клонов Экто клеток (Экто 15, 34, 2 и 23). Клетки культивировали в течение двух дней в присутствии 10 нМ тестостерона и использовали эктодомен синдекана-1, накопленного в среде. Культурную среду использовали непосредственно. Образцы нормализовали к числу клеток и эквивалентные количества слот-блоттировали на мембране Hybond-H+. Эктодомен синдекана-1 определяли, как описано в примерах, методом усиленной хемилюминесценции с использованием 281-2 (Miettinen, Н. М. с сотр. , J. Cell Sci., в печати (1994)). Количественное определение проводили с использованием аналитической системы с компьютерным изображением ((Имэйджинг Рэсерч Инк.). Представлены средние значения и SEM для двух параллельных образцов.
На фиг. 6 изображена организация актинового филамента клонов Экто клеток. Экто клетки культивировали в присутствии 10 нМ тестостерона и актиновые филаменты визуализировали с помощью родаминконъюгированного фаллоидина.
На фиг. 7 изображено образование колоний эктоклеточных клонов на мягком агаре. Клетки культивировали в течение 12 дней в 0,33% мягком агаре, DMEM+5% FCS в присутствии 10 нМ тестостерона, в соответствии с описанным ранее (Leppa, S. с сотр., Proc. Natl. Acad. Sci. USA 89:932 (1992))).
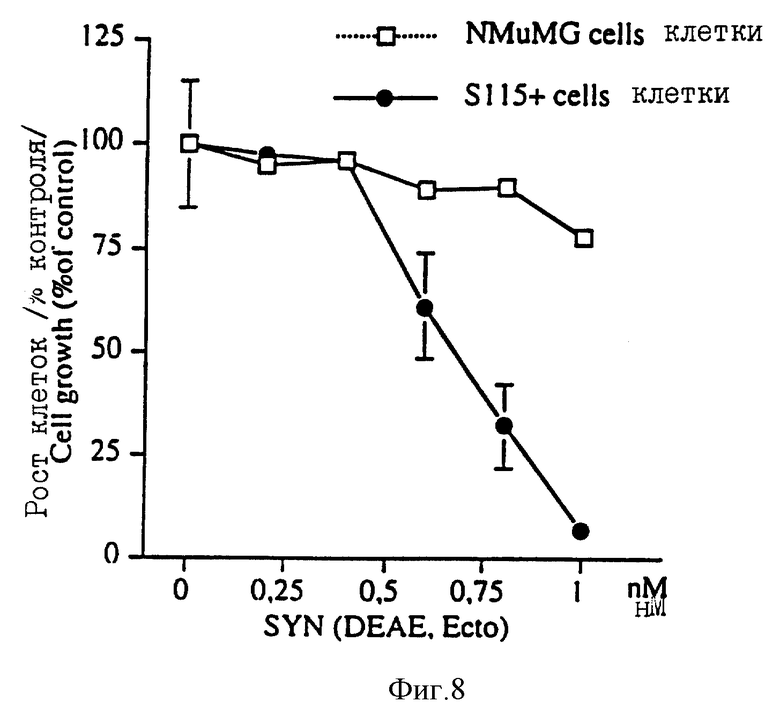
На фиг. 8 показано влияние DEAE-выделенного эктодомена синдекана-1 (примеры) из кондиционированной среды клеток Экто 2 на рост NMuMG и обработанных тестостероном (10нМ) клеток S115 (S115+). 1500 клеток переносили на 96-луночные микротитрационные планшеты и клетки культивировали в присутствии DEAE-выделенного эктодомена синдекана-1 до достижения 75-85% слияния контрольных (без эктодомена синдекана-1) клеток (четыре дня для клеток NMuMG и три дня для клеток S115+). Затем клетки фиксировали 2% параформальдегида, окрашивали с помощью 0,5% красителя - кристаллического фиолетового и промывали дистиллированной водой. Окрашенные клетки суспендировали в 10% уксусной кислоте и их количество определяли спектрофотометрически при длине волны 595 нм.
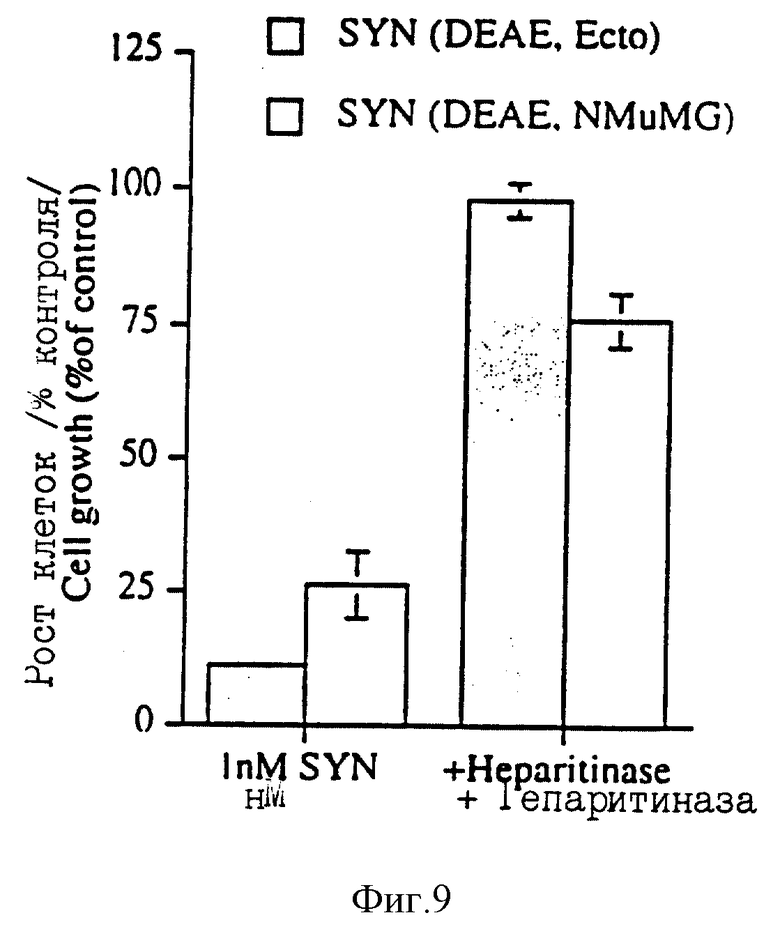
На фиг. 9 изображено влияние гепаритиназной обработки DEAE-выделенного эктодомена синдекана-1 на ингибирование роста клеток S115+. Клетки S115+ культивировали в присутствии 1 нМ DEAE-выделенного синдекана-1 из культуральной среды Экто 2 клеток и из среды NMuMG клеток или в присутствии таких же препаратов, предварительно обработанных гепаритиназой (Сейкагаку Когио Ко.) в течение 1 часа при 37oC.
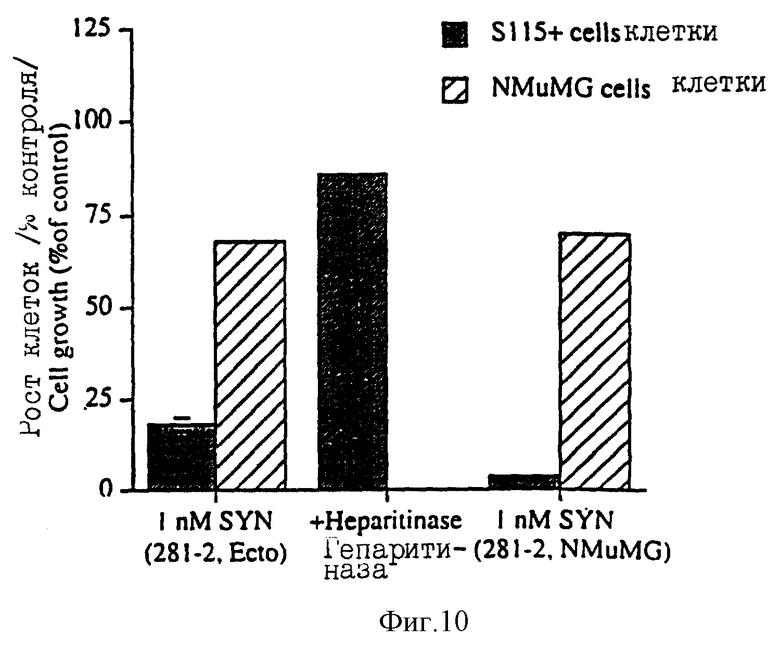
На фиг.10 показано влияние очищенного иммуно-сорбционным методом эктодомена синдекана-1 на рост клеток S115+ и NMuMG. DEAE-выделенный эктодомен синдекана-1 дополнительно очищали на 281-2 иммуноаффинной колонке (примеры). Клетки S115+ и NMuMG культивировали в присутствии 1 нМ иммуноаффинно-очищенного эктодомена синдекана-1.
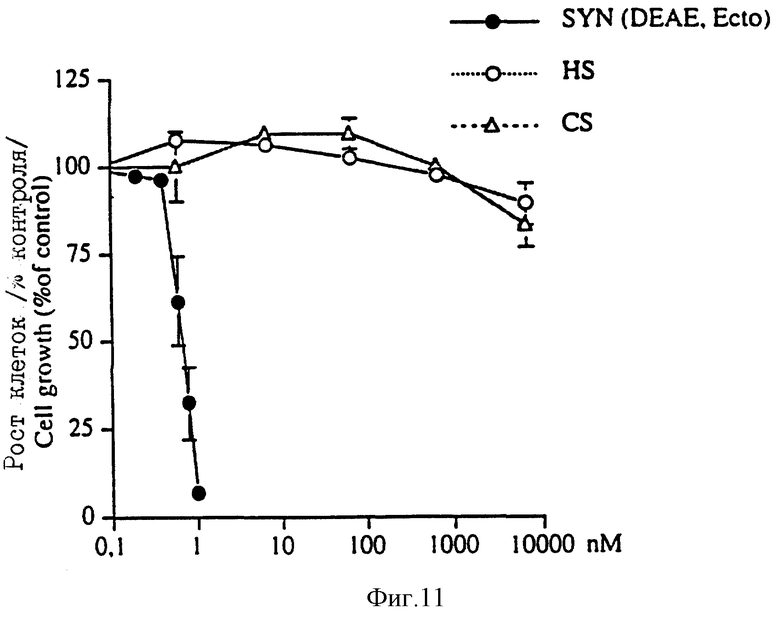
На фиг. 11 показано, что DEAE-выделенный эктодомен синдекана-1, а не HS или CS GAG ингибирует рост клеток S115+.
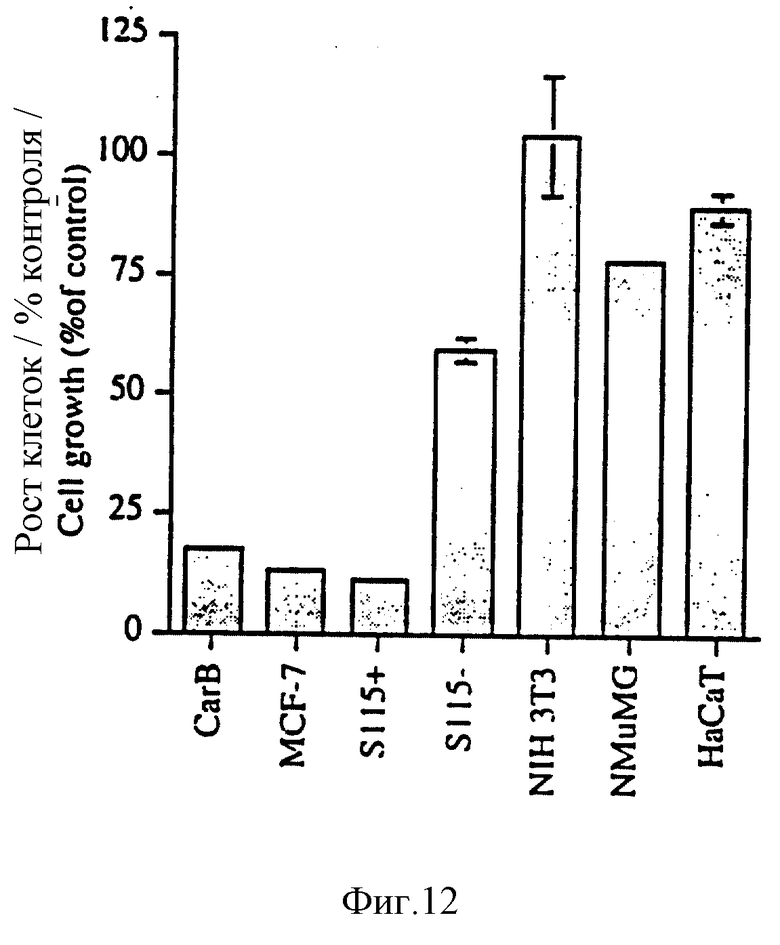
На фиг. 12 показано ингибирование роста клеток различных клеточных линий (CarB, MCF-7, S115+ в присутствии 10 нМ тестостерона, S115+ без тестостерона, NIH 3Т3, NMuMG и НаСаТ) с помощью 1 нМ DEAE-выделенного эктодомена синдекана-1 (примеры). Рост клеток анализировали на всех панелях аналогично показанному для панели (А) и проводили сравнение с поведением клеток без обработки (% контроля, ось "у"/. Представлены средние значения и значения SEM для двух параллельных образцов.
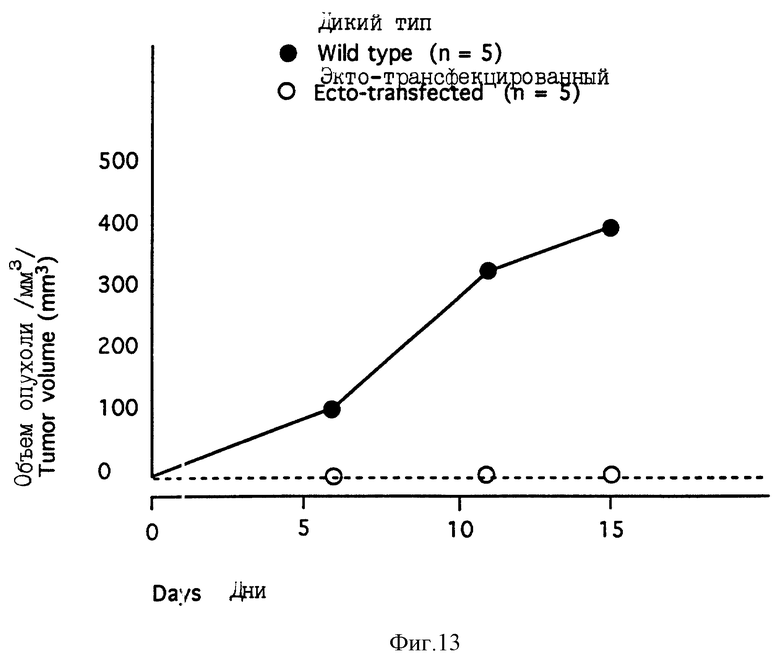
На фиг. 13 показано подавление опухолевого роста на лишенных волосяного покрова мышах с помощью эктодомена синдекана-1.
Краткое изложение сущности изобретения
Настоящее изобретение, прежде всего, относится к фармацевтически приемлемой композиции, содержащей синдекановый эктодомен.
Кроме этого, настоящее изобретение относится к способу уменьшений или нормализации роста опухолевых клеток путем доставки такого синдеканового эктодоменового протеина к опухолевой клетки во внеклеточной среде окружающей клетку.
Способы настоящего изобретения применимы как на злокачественных, так и на неэлокачественных опухолевых клетках и они особенно применимы на опухолях, характеризующихся потерей синдекана-1, таких, как глиомы, миеломы, карциномы, саркомы, лимфомы или аденомы.
Определения
Для более ясного и последовательного понимания описания и формулы изобретения, включающих указанные ниже термины, ниже приведена расшифровка таких терминов.
Рост клеток. Под термином "рост клеток" подразумевается клеточная репликация или скорость деления клеток как в контролируемых, так и в неконтролируемых условиях. Поэтому рост клеток подразумевает скорость деления и репликацию.
Злокачественный. Tepмин "злокачественный" относится к неконтролируемому росту клеток.
Более зрелый (дефинитивный) фенотип. При упоминании клеток, имеющих "более зрелый фенотип" подразумевается, что клетка обладает фенотипом, обычно характерным для некоторых типов клеток более зрелых, чем данная клетка. Фенотип может быть определен одной или более фенотипическими характеристиками. Так, например, форма эпителиальной клетки представляет собой более зрелый фенотип мезенхимально-подобной формы; поэтому в таком примере "более зрелый фенотип" представляет собой морфологию эпителиальной клетки, а не мезенхимально-подобную форму. Терминально дифференцированная мезенхимальная клетка представляет собой "более зрелый фенотип", чем конденсирующая мезенхимальная клетка. В тексте может также упоминаться актинсодержащий цитоскелет; дезорганизованные актиновые филаменты являются индикаторами менее зрелого фенотипа, чем организованные филаменты.
Эффективное количество. "Эффективное количество" какого-либо агента представляет собой количество такого агента, достаточное для получения желаемого результата, особенно при применении такого агента на животном или человеке. Эффективное количество эктодомена синдекана-1 в композициях и способах настоящего изобретения представляет собой количество, достаточное для уменьшения роста опухолевых клеток, предпочтительно до скоростей нормального роста конкретных клеточных типов.
Применение. Под термином "применение" подразумевается введение эктодомена синдекана в соответствии с изобретением в организм животного или человека с помощью соответствующих средств, известных в данной области медицины, причем такой термин охватывает инъекционное, оральное, энтеральное, подкожное и парентеральное (например, внутривенное) применение, но не ограничивается ими.
Воздействие эктодоменом синдекана. Под "воздействием" на клетку синдеканового эктодомена в композициях изобретения подразумевается, что внешняя среда клетки обеспечивается количествами синдеканового эктодомена, являющимися эффективными для промотирования желаемого эффекта, особенно для замедления скорости роста опухолевых клеток.
Фармацевтически приемлемая соль. Подразумевается, что термин "фармацевтически приемлемые соли" охватывает соли синдеканового эктодомена настоящего изобретения. Такие соли могут образовываться из фармацевтически приемлемых кислот или оснований, например из таких кислот, как серная, хлористо-водородная, азотная, фосфорная и т.д., или таких оснований, как гидроксиды щелочноземельных металлов, гидроксиды аммония, гидроксиды алкиламмония и т.п.
Фармацевтически приемлемая композиция. Подразумевается, что термин "фармацевтически приемлемая композиция" охватывает растворители, носители, разбавители и т. п. , которые используются в качестве добавок в препаратах синдеканового эктодомена настоящего изобретения для применения соединений на пациентах (человеке или животном), нуждающихся в лечении. Такие добавки могут выполнять некоторые функции, например, обеспечивать надлежащие ионные условия для применения, стабилизировать синдекановый эктодомен от дезактивации или деградации и/или увеличивать период полураспада синдеканового эктодомена. Фармацевтически приемлемая композиции в медицинском отношении совместима с организмом хозяина, на котором осуществляют применение лекарственного средства.
Лечение. Подразумевается, что термин "лечение" или "обработка" охватывает применение фармацевтически приемлемых композиций изобретения, включающих эффективные количества синдеканового эктодомена изобретения, на пациенте в целях, которые могут включать профилактику, улучшение состояния, предотвращение или лечение медицинского нарушения, включая подавление роста опухоли.
Практически не содержащий природных загрязнений. Материал считается "практически не содержащим природных загрязнений", если он был подвергнут существенной очистке от веществ, которые обычно присутствуют в нем в естественном состоянии, до такой очистки, и такие примеси, обычно обнаруживаемые при естественном состоянии вещества in vivo или in vitro, практически отсутствуют в конечном препарате такого материала. При применении на объекте, нуждающемся в лечении, синдекановый эктодомен изобретения практически не содержит природных загрязнений, которые связаны с синдекановым эктодоменом как in vivo (в организме хозяина, из которого выделяют эктодомен) так и in vitro (в результате химического синтеза). Под термином "практически отсутствует" подразумевается, что такие примеси либо полностью отсутствуют, или присутствуют в таких низких концентрациях, что их наличие (1) не препятствует желаемому терапевтическому эффекту активного агента (в данном случае способности синдеканового эктодомена ингибировать рост опухоли) в терапевтически приемлемой композиции при ее применении на пациенте, нуждающемся в лечении и (2) не вредит пациенту при применении такой композиции.
Описание изобретения
Настоящее изобретение базируется на открытии того факта, что эктодомены синдекана обладают некоторыми биологическими функциями и способны обеспечивать такими функциями клетки в присутствии на внешней поверхности клетки, отличной от тех, что синтезируют такой синдекановый эктодомен. Синдеканы представляют собой протеины, связанные с мембраной. Неожиданно было установлено, что внеклеточно образовавшийся синдекановый эктодомен сам по себе достаточен для реконструкции более дифференцированной морфологии опухолевых клеток и подавления роста злокачественных клеток. В этом случае примером изобретения служит синдекан-1.
Все синдеканы содержат цитоплазмический домен, трансмембранный домен и внеклеточный домен. Внеклеточный домен представляет собой эктодомен. Как обсуждается Jalkanen с сотр. в книге "Рецепторы для внеклеточной матрицы", изд. Дж. Макдональд и Р.Мэшем, Академик Пресс, Сан-Диего, стр. 1-37 (1991), синдеканы содержат высококонсервативные гомологические последовательности в трех отдельных областях их эктодоменов. Двухосновная последовательность непосредственно соседствует с N-терминальным концом гидрофобного трансмембраного домена, что предполагает ее местонахождение за внешней створкой мембраны плазмы и способность выполнять функции протеаза-восприимчивого сайта, который обеспечивает отщепление интактного эктодомена от поверхности клетки.
Коровый белок человеческого синдекана-1 содержит 310 аминокислотных остатков. Мышиный и человеческий синдеканы-1 в высокой степени структурно и функционально гомологичны друг другу. Человеческий синдекан-1 имеет размер, заряд, гидростатическую плотность и GAG состав, идентичные соответствующим характеристикам мышиного синдекана-1. Эктодомен человеческого синдекана-1, подобно мышиному аналогу, связывается с фибриллами коллагена типа 1 и с фибронектином, но не способен связываться с ламинином или витронектином.
Последовательность человеческого синдекана-1 является известной, и она была подвергнута клонированию (Mali с сотр., J. Biol. Chem. 265:6884-6889 (1990)).
В соответствии с нумерацией согласно фиг. 2 в статье Mali с сотр., J. Biol. Chem. 265:6884-6889 (1990) аминокислоты 1-251 представляют собой эктодомен человеческого синдекана-1 (с присоединенным сигналом секреции), гидрофобный, охватывающий мембрану домен содержит следующие 25 аминокислотных остатков (аминокислоты 252-276), а цитоплазматический домен содержит последние 34 аминокислотных остатка (аминокислоты 277-310). Сигнальная пептидная последовательность представляет собой первые 17 аминокислот эктодомена. Использующийся для промотирования секреции синдекана-1 из синтезирующей его клетки секреционный сигнал не является необходимым для подавления роста опухоли или дифференцировочных функций эктодомена изобретения.
В связи с этим последовательность эктодомена настоящего изобретения содержит такие фрагменты синдекановых аминокислотных остатков 1-251, которые сохраняют сайты присоединения GAG и желаемую функцию эктодомена, как это имеет место в случае, например, эктодоменов, содержащих аминокислоты 1- 251 (с секреционным сигналом и расщеплением на RK сайте), 18-251 (без сигнала секреции, но с расщеплением на RK сайте), 1-231 (с сигналом секреции, но с расщеплением на RR сайте) и 18-251 (без секреционного сигнала, но с расщеплением на RR сайте). Эктодомен с карбоксиокончанием на сайте, расположенном между аминокислотными остатками 231-251, или фрагмент сигнала секреции, меньший фрагмента из аминокислот 1-17, также могут использоваться, поскольку можно ожидать, что такие решения сохраняют биологические свойства эктодомена.
Хотя человеческий и мышиный эктодомены идентичны по аминокислотному уровню лишь на 70%, все предполагаемые сайты присоединения гликосаминогликана (GAG) в мышиной и человеческой последовательностях являются идентичными. Пять возможных сайтов прикрепления гликозаминогликана эктодомена человеческого синдекана представляют собой позиции 37, 45, 47, 206 и 216. Два из таких сайтов принадлежат к консенсусной последовательности SGXG, а три других - к (E/D)GSG(E/D). Кроме этого, мышиный и человеческий синдеканы содержат идентичные сигнальный сайт N-гликозиляцин м протеиназа-чувствительный двухосновный RK сайт, соседний с внеклеточной лицевой поверхностью трансмембранного домена. Человеческий синдекан также содержит вторую двухосновную RR последовательность из 18 остатков помимо RK последовательности. Протеолитическое расщепление в этом сайте также может приводить к образованию эктодомена настоящего изобретения, который содержит все незатронутые GAG сайты.
Трансмембранные домены человеческого и мышиного синдекана-1 идентичны на 96% (единственным отличием в человеческом синдекане является замена аланина на глицин), а цитоплазматические домены на 100% идентичны в мышином и человеческом синдеканах.
Синдекановый эктодомен, например эктодомен человеческого синдекана, может быть получен рекомбинантными методами в организме любого желательного хозяина. Однако предпочтительно, но не обязательно, использовать хозяина с клеточным типом, аналогичным клеточному типу опухоли с тем, чтобы обеспечить максимально аналогичный состав GAG относительно клетки в неопухолевом состоянии. Имеется в распоряжении большое число депонированных клеточных линий, которые специфичны в отношении человеческой ткани или обладают характеристиками различных клеточных типов.
Так например, клоны мышиного синдекана-1 настоящего изобретения конструировали с использованием липосомной трансфекции и генетицина для последующей селекции устойчиво трансфекцированных клеточных клонов. Клоны линии клеток S115 (см. фиг. 3) экспрессируют дикий тип мышиного синдекана-1 (дикий тип), делеционный мутант с единственным аргининовым остатком только в цитоплазматическом домене (бесхвостый) или плоский эктодомен синдекана-1 (экто). Дикий тип синдекана-1 и цитоплазматический делеционный мутант (бесхвостый) клонировали в EcoRI сайт pBGS эукариотического вектора экспрессии. Эктодоменовую конструкцию клонировали в рМАМnео вектор с тем, чтобы также обеспечить эффективные уровни экспрессии в присутствии гормона, поскольку ММТ LTR промотор индуцируется тем же стероидным гормоном, что и клетки. Необязательно использовать именно такой вектор, поскольку в этой области известно большое число таких векторов экспрессии. Экспрессию синдекана-1 на поверхностях клеток детектируют с использованием моноклонального антитела, примером которого может служить упомянутое выше mAB 218-2, которое распознает эктодомен корового протеина мышиного синдекана-1, а актиновые филаменты визуализируют с использованием родамин-конъюгированного фаллоидина как индикатора состояния дифференцировки и роста клетки.
В отсутствие тестостерона клетки S115 демонстрируют наличие организованных актиновых филаментов, типичных для эпителиодального состояния таких клеток. В присутствии тестостерона актин дезорганизуется и приобретает шарообразную форму, при этом также подавляется экспрессия синдекана-1 на поверхности клеток, клоны дикого типа и бесхвостые клоны, экспрессирующие синдекан-1 на клеточных поверхностях восстанавливают организацию актинового филамента, несмотря на обработку тестостероном. Из-за трансфекции бесхвостного мутанта также индуцируются изменения, аналогичные для синдекана-1 дикого типа, клетки S115 трансфекцируются простым эктодоменом и образуется более 50 независимых клонов, секретирующих различные уровни эктодомена в культурную среду. Поверхности таких клеток приобретают лишь слабую окраску под действием синдекана-1, и такие клетки все еще демонстрируют хорошо организованные актиновые филаменты и эпителоидную морфологию. Эти результаты показывают, что эктодомен синдекана-1 обладает вполне достаточными свойствами для восстановления эпителиоидной морфологии клеток S115, обработанных тестостероном до морфологического состояния, характерного для более зрелого фенотипа, и является ценным противораковым лекарственным средством.
В случае неопухолевых клеток синдекан экспрессируется в эпителиальных клетках, мезенхимальных клетках, пре-B клетках и плазмацитах, но не экспрессируется B-клетками. Синдекан также экспрессируется в тканях, содержащих клетки таких типов, например, в человеческой мозговой ткани. Поэтому способы настоящего изобретения особенно полезны в борьбе с опухолями эпителиальных, мезенхимальных, пре-B и плазматических клеток. Главным образом, способы настоящего изобретения используются для замедления роста стероидо-реактивных опухолей, особенно эстроген или андроген-реактивных опухолей (опухолей, которые лучше растут в присутствии стероидов, например, эстрогена или андрогена), включающих опухоли клеток молочной железы, опухоли клеток слизистой оболочки матки и опухоли клеток предстательной железы.
При лечении людей и животных эктодомен синдекана-1 применяют в фармацевтически приемлемом растворе при дозировках, достаточных для восстановления со стояния нормального роста опухоли или злокачественных клеток, свидетельством чего является замедление скорости роста. Синдекансодержащий фармацевтически приемлемый раствор может применяться в любой форме, оказывающей профилактическое, паллиативное, предотвращающее или регрессирующее действие на рост опухоли.
Количество композиций эктодомена синдекана-1 на стоящего изобретения, которое применяют на пациенте и длительность такого применения, могут определяться путем мониторинга опухолевого роста у пациента в ходе лечения и регулироваться в соответствии с реакцией пациента. Синдекановый эктодомен настоящего изобретения предпочтительно применяют на опухолевой клетке-мишени при внеклеточных концентрациях 0,7-1,0 нМ (см. фиг. 11), однако могут использоваться любые концентрации, достаточные для уменьшения роста опухоли. Эктодомен может применяться либо локально (как это имеет место при концентрированной доставке непосредственно к органу-мишени) или системно (например, путем до ставки через кровеносную систему). Доза синдекана, применяемая на пациенте (как на человеке, так и на животном), таким образом, должна выбираться с учетом объема (например, объема крови), в котором применяют эктодомен, и типа опухоли, являющейся мишенью для лекарства.
Так например, если необходимо постоянное воздействие синдеканового эктодомена, то требуется более частый прием дозировок лекарства, чем для случая, когда необходимо лишь временное воздействие синдеканового эктодомена на опухоль. Так, например, 1 нМ синдеканового эктодомена, содержащего аминокислоты 1-251, соответствует концентрации 0,2 мг/л (200 мкг/л) в крови или при локальном применении в зоне действия. Типичные системные дозировки синдеканового эктодомена, используемые в способах настоящего изобретения для лечения людей или животных, включают количества, обеспечивающие конечную концентрацию в крови, наиболее предпочтительно, 0,2 мг синдеканового эктодомена на литр крови. Объем крови у людей составляет 6% от веса тела, следовательно, на 70 кг веса тела приходится 4,2 литра крови. Однако, поскольку предполагается, что эффекты синдеканового эктодомена носят локальный (например, воздействие на специфическую клеточную мембрану), изолированный и кинетически ограничительный характер, теоретически минимальная дозировка может быть выше указанной для достижения полезных терапевтических эффектов.
Синдекановый эктодомен может применяться любым путем, обеспечивающим доставку эффективных количеств лекарства в желаемую активную область, например путем инъекции. В случае парентерального применения препараты, содержащие синдекановый эктодомен, могут использоваться на пациенте, нуждающемся в таком лечении в комбинации с фармацевтически приемлемыми стерильными водными или неводными растворителями, суспензиями и эмульсиями, примерами неводных растворителей могут служить пропиленгликоль, полиэтиленгликоль, растительное масло, рыбий жир и подходящие для инъекций сложные органические эфиры. Водные носители включают воду, водно-спиртовые растворы, эмульсии или суспензии, включая физиологический раствор и буфферные медицинские парентеральные носители, включающие раствор хлористого натрия, декстрозный раствор Рингера, раствор декстрозы и хлористого натрия, раствор Рингера, содержащий лактозу или фиксированные масла. Внутривенные носители включают жидкие и питательные дополнители, электролитические дополнители, например добавки на основе декстрозы Рингера и т.п.
Медикамент, содержащий синдекановый эктодомен (фармацевтически приемлемый раствор, содержащий терапевтически активный синдекановый эктодомен), может применяться с помощью катетеров или насосов, особенно в тех случаях, когда желательно обеспечить доставку эктодомена в локализованных высоких концентрациях. Медикамент, содержащий эктодомен синдекана-1, может применяться подкожно или непосредственно в мягкую ткань с помощью имплантированных устройств, инертных в отношении жидкостной среды тела. Такие устройства и имплантационные системы известны в данной области. Так например, керамическая система для доставки протеинов описана в WO 92/00109.
Медикамент, содержащий эктодомен синдекана-1, может применяться с использованием такой молекулы, как часть химерной молекулы (или комплекса), предназначенной для воздействия на конкретный орган-мишень, например как часть антитела, распознающего детерминанты целевой ткани, органа или клетки как в опухолевом, так и неопухолевом состоянии.
Фармацевтически приемлемый раствор, содержащий эктодомен синдекана-1, может применяться локально. Хотя эктодомен синдекана- 1 может применяться на пациенте в режиме, включающем использование других лекарств для борьбы с раком, оптимальное применение синдекансодержащих композиций изобретения особенно полезно в этом случае.
Местное применение предпочтительно осуществляют одним из двух методов. Согласно первому из них терапевтически активный синдекановый эктодомен может смешиваться с подходящими фармацевтически приемлемыми носителями и (необязательно) с усилителями пенетрации с целью облегчения доставки активного агента через кожу с образованием мазей, эмульсий, лосьонов, растворов, кремов, гелей и т.п. и затем приготовленный препарат применяют на определенном участке кожи. Согласно второму методу терапевтически активный синдекановый эктодомен может вводиться в пластырь или систему трансдермальной доставки в соответствии с известной технологией приготовления таких пластырей и систем доставки.
Применение лекарства в форме пролонгированного выделения более удобно для пациента в том случае, когда необходимы повторяющиеся инъекции в течение длительного времени или когда желательно длительное воздействие эктодомена на опухолевую клетку. При использовании в виде внутривенных дозировочных форм композиции настоящего изобретения демонстрируют достаточно быстрое начало действия, что необходимо для быстрого регулирования опухолевого роста.
Применение может быть локализовано непосредственно на клетке, если она связана с тканью или органом тела, или применение может быть системным, в среде, где присутствует такая клетка, например в крови или спинно-мозговой жидкости. Системное применение на теле пациента, например применение медикамента через кровеносную систему, облегчает лечение пациентов, у которых опухолевые клетки обнаружены в более чем одном участке тела.
Обеспечение синдеканового эктодомена в виде продукта конструкции на основе экспрессии синдеканового эктодомена, которая секретирует эктодомен в эффективных количествах, также рассматривается как "применение". Так, например, применение через гематоэнцефалитический барьер может быть достигнуто в использовании известных вирусных векторных систем для доставки ДНК синдеканового эктодомена, обеспечивающих экспрессию эктодомена и его секрецию во внеклеточную среду, как это имеет место, например, в ретровирусных системах, описанных в WO 93/03743, WO 090/09441, и в статьях Breakefield, X.А. с сотр. The New Biologist 3:203-218 (1991) и Huang, Q. с сотр., Exp. Neurol. 115:303-315 (1992).
Фармацевтически приемлемую композицию изобретения, содержащую эктодомен синдекана-1, получают известным способом, например с помощью традиционных процессов смешивания, растворения, лиофилизации или т.п. Композиции настоящего изобретения, обеспечивающие эктодомен синдекана-1, находят применение в связи со своей способностью замедлять или предотвращать рост опухоли или их рецидивы и способность изменять генотип клеток на генотип более дифференцированного состояния как на людях, так и на животных. В соответствии с настоящим изобретением композиции эктодомена синдекаца-1 используют собственные механизмы тела для промотирования дифференциации конкретных клеточных типов до их максимального потенциала.
Подразумевается, что композиции и способы изобретения не ограничиваются использованием только синдекана-1. Известно, что синдекан-1, синдекан-2, синдекан-3 и синдекан-4 содержат аналогичные доменные структуры. Известно, что дифференцировка некоторых клеточных типов связана с потерей синдекана-1, но с появлением другого члена синдеканового семейства (Bernfield, О., с сотр., Annu. Rev. Cell Biol., 8:365-393)).
Так, например, в том случае когда бронхиальный эпителий образует ростки, легочная мезенхима теряет синдекан-1, но приобретает синдекан-2. В опухолях из клеточных типов, которые теряют синдекан-1 в ходе дифференцировки, но экспрессируют другой синдекан, можно ожидать использования эктодомена из синдекана, которые экспрессируется в состоянии дифференциации.
Представленные ниже примеры носят лишь иллюстративный характер и не ограничивают сферу изобретения.
ПРИМЕРЫ
Следующие ниже примеры иллюстрируют, но не ограничивают изобретение.
Пример 1
Делеционные мутантные синдекановые конструкции
С использованием липосомной трансфекции и последующей селекции устойчиво трансфекцированных клеточных клонов в присутствии генетицина, как описано Leppa с сотр. , Proc. Natl. Acad. Sci. США. 89:932 (1992), получали клоны линии клеток S115 (см. фиг. 3), которые экспрессировали дикий тип мышиного синдекана-1 (дикий тип), делеционный мутант с единственным аргининовым остатком только в цитоплазматическом домене (бесхвостый) или только эктодомен синдекана-1 (Экто 2; см. фиг.3). Три такие формы и соответствующих хозяев конструировали следующим образом.
кДНК мышиного синдекана-1 полной длины, описанную в статье Mali с сотр., J. Biol. Chem. 268:24215-24222 (1993), клонировали в EcoRI сайт Bluescript SK+ (Промега).
1) EcoRI вставку конструкции Bluescript клонировали в EcoRI сайт вектора pBGS (Mali с сотр. , J. Biol. Chem. 268:24215-24222(1993)) и подтверждали ориентацию. Такую конструкцию обозначали как "дикий тип".
Мутагенный олигонуклеотид из 25 оснований, имеющий последовательность: 5' G CTG TAС CGC TAG CAG AAG GAC-3' [SEQ ID N:1], содержащий стоп-кодон и Nhel рестрикционный сайт (подчеркнут), использовали для превращения кодона по второй аминокислоте (метионин) цитоплазматического домена, следующего за трансмембранным доменом, в стоп-кодон. Мутацию подтверждали рестрикционным перевариванием и дидеокси секвенированием. EcoRI вставку конструкции Bluescript клонировали в EcoRI сайт амплифицируемого pBGS вектора (Mali с сотр. , J. Biol. Chem. 268:24215-24222 (1993). Такой мутантный синдекан-1, содержащий одну аминокислоту (аргинин) в его предполагаемом цитоплазматическом домене, обозначали как "бесхвостый".
Мутагенный олигонуклеотид из 33 оснований:
5'-GACACCTCCCAGTACTCACTTCCTGTCCAAAAG-3' [SEQ ID N:2], содержащий стоп-кодон, и Scal-сайт (подчеркнут) использовали для превращения первого кодона (Е) после сайта эктодомена, чувствительного к двухосновной протеазе, в стоп-кодон. Мутацию подтверждали рестрикционным перевариванием и дидеокси секвенированием. Было установлено, что анализируемый объект представляет собой Bluescript-эктоконструкцию. EcoRI вставку Bluescript-эктоконструкции клонировали в EcoRI сайт вектора pJC119R (Miettinen с сотр., J. Cell Sci. 107: в печати, (1994)). Xhol переваренную экто-вставку из pJC119R-эктоконструкции лигировали в Xhol сайт pMAMneo эвкариотного трансфекционного вектора, полученного из Контех, Пало Альто (Leppa с сотр., Proc. Natl. Acad. Sci. США. 89, 932(1992)) и ориентацию подтверждали рестрикционными перевариваниями.
Пример 2
Экспрессия мутантного синдекана-1, нормализующего злокачественный рост в клетках S115
Синдекан-1 дикого типа и цитоплазматический делеционный мутант (бесхвостый) клонировали в EcoRI сайт pBGS эукариотического экспрессионного вектора (Mali с сотр. , J. Biol. Chem. 268:24215 (1993), а эктодоменовую конструкцию клонировали в pMAMneo вектор с целью достижения эффективных уровней экспрессии также в присутствии гормона (персональное сообщение, S. AIa-Uoti, Турецкий центр биотехнологии). pBGS система не подавлялась тестостероном. Экспрессию синдекана-1 на поверхностях клеток детектировали с использованием mAb 281-2 (Jalkanen с сотр., J.Cell Biol. 101:976 (1985), который распознает эктодомен корового протеина мышиного синлекана-1, а актиновые филаменты визуализировали с использованием родамин-конъюгированного фаллоидина.
Клетки (S115+, дикого типа, бесхвостые и Экто 2) культивировали в течение четырех дней на предметных стеклах в среде DMEM-5%FCS-lмM пирувата Na в присутствии 10 нМ тестостерона, за исключением клеток S115, которые культивировали без тестостерона в среде ДМЕМ-4%DCC-FCS (обработка древесным углем, на который нанесен дейстран, обеспечивает удаление из сыворотки эндогенных стероидов) в присутствии 1мМ пирувата Na (пировинограднокислого натрия - прим. пер.) Клетки фиксировали с помощью 0,1% Тритона-Х-100, 2% параформальдегида и инкубировали в присутствии родамин-конъюгированного фаллоидина (Сигма). Экспрессию синдекана-1 на поверхности клетки визуализировали путем инкубации живых клеток в течение 1 часа на льду в присутствии крысиного mAb 281-2 (распознающего эктодомен мышиного синдекана-1); затем их фиксировали 2% параформальдегида и связанный mAb 281-2 визуализировали с использованием FITC-конъюгированного кроличьего антикрысиного IgC.
В отсутствие тестостерона клетки S115 демонстрируют организованные актиновые филаменты, типичные для этих клеток в эпителиоидальном состоянии. В присутствии
гормона актин дезорганизуется и приобретает шарикообразную форму и наблюдается подавление экспрессии синдекана-1 на поверхности клеток, как это было показано ранее Leppa с сотр. Cell Reg. 2, 1 (1991), фиг.4.
Клоны дикого типа и бесхвостые клоны, экспрессирующие синдекан-1 на поверхности клеток, восстанавливают организацию актинового филамента несмотря на обработку тестостероном, фиг. 4.
Пример 3
Влияние секретированного эктодомена синдекана-1 на культивированные S115 клетки
Поскольку трансфекция бесхвостого мутанта вызывает такие же изменения, что и в случае синдекана-1 дикого типа, клетки S115 трансфекцировали с помощью эктодомена. Получали более 50 независимых клонов, секретирующих различные количества эктодомена в культурную среду (см. фиг. 5, 6 и 7). Поверхности таких клеток лишь слабо окрашивались синдеканом-1, но они все еще демонстрировали организованные актиновые филаменты и эпителоидную морфологию (фиг. 4). Эти результаты подтверждают тот факт, что эктодомен синдекана-1 обладает достаточной совокупностью свойств для реконструкции эпителиоидной морфологии обработанных тестостероном клеток S115.
С целью подробного анализа Экто клонов методом усиленной хемилюминесценции измеряли количества секретированного эктодомена синдекана-1 в культурной среде с использованием mAb 281-2 к эктодомену корового протеина синдекана-1. Два отдельных устойчиво трансфекцированных клеточных клона, секретирующих высокие количества синдекана-1 в культурную среду (Экто 2 и Экто 23) и два клеточных клона с низкой экспрессией (Экто 15 и Экто 34) выбирали для дальнейшего анализа (фиг.5).
Явная корреляция между экспрессией эктодомена синдекана-1 и реорганизацией актиновых филламентов наблюдалась в присутствии 10 нМ тестостерона: Экто 15 и Экто 34 с низкой экспрессией синдекана- 1 содержали дезорганизованный, главным образом, глобулярный актин, тогда как Экто 2 и Экто 23 клоны, экспрессирующие эктодомен синдекана-1, демонстрировали эпителиоидную морфологию с пучками организованного актинового филамента (фиг.6). Как было показано ранее, повышенная экспрессия интактного синдекана-1 подавляет опухолевый рост обработанных тестостероном клеток S115 (Leppa с сотр., см. цитированную выше ссылку) и в данном случае клоны Экто 2 и Экто 23 с высокой экспрессией эктодомена синдекана-1 также демонстрируют ограниченный рост на мягком агаре. Однако клоны Экто 15 и Экто 34 с низкой экспрессией эктодомена синдекана-1 демонстрируют рост на мягком агаре, типичный для родственных клеток S115 (фиг.7). Эксперимент на мягком агаре показывает, что помимо морфологии экспрессия эктодомена синдекана-1 также достаточна для ограничения онкогенного роста клеток S115.
Пример 4
Выделение и очистка синдеканового эктодомена от Экто клеточных культур
Поскольку эктодомен синдекана-1, по-видимому, ответственен за подавление злокачественного роста андроген-обработанных клеток S115, осуществляли сбор кондиционированной среды из Экто клеточных культур для выделения эктодомена. Кондиционированную клеточную культурную среду денатурировали с помощью 2 М мочевины и кипятили перед загрузкой в DEAE-сефацеловую колонку, добавляли 50 мМ ацетата Na (pH=4,5) и среду замораживали до +4oC. Колонку промывали 0,2 М NaCl, 2М мочевины, 50 мМ ацетата Na (pH=4,5) и связанный материал элюировали с использованием 1 М NaCl, 2 М мочевины, 50 мМ ацетата Na (pH=4,5). Фракции, содержащие эктодомен синдекана-1 подвергали диализу противотоком к забуференному фосфатом физиологическому раствору (PBS) при 4oC. Количество эктодомена синдекана-1 во фракциях определяли методами слот-блоттинга и последующей усиленной хемилюминесценции, с использованием mAb 281-2 (Пример 2 и Miettinen Н.М. с сотр., J. Cell. Sci. в печати (1994)) и сравнивали это количество с известным стандартным синдеканом-1.
Эктодомен синдекана-1 из культурированной среды Экто клеток биохимически аналогичен эктодомену синдекана-1, выделенному из нормальных эпителиальных клеток грудной железы мышей (NMuMG). После выделения измеряли содержание в препарате синдекана-1 и препарат тестировали на гормон-обработанных клетках S115. Как показано на фиг. 8, такие низкие концентрации DЕАЕ-выделенного эктодомена синдекана-1 как 1 нМ подавляют рост обработанных тестостероном клеток S115 (фиг. 8). Такая же концентрация лишь незначительно ингибирует рост NMuMG клеток, которые использовались в качестве нормальных эпителиальных клеток (фиг. 8). Эктодомен синдекана-1 также выделяли из культурной среды NMuMG клеток и опять-таки 1 нМ концентрация такого препарата ингибировала рост обработанных гормоном клеток S115 (фиг. 9). Обработка DEAE-выделенного эктодомена гепаритиназой полностью подавляла ростингибирующую активность таких препаратов (фиг. 9), что предполагает невовлечение в такой процесс корового протеина синдекана-1.
DEAE-выделенный эктодомен синдекана-1 дополнительно очищали с использованием mAb 281-2 иммуноаффинной колонки: DEAE-выделенный эктодомен синдекана-1 в PBS загружали в mAb 281-2 Сефароза CL-4B иммуноаффинную колонку в соответствии с методикой, описанной Jalkanen с сотр., J. Cell. Biol. 105: 3087 (1987) и связанный материал элюировали 50 мМ триэтиламина (pH=11,5). Фракции, содержащие эктодомен синдекана-1, подвергали диализу противотоком к дистиллированной воде и затем лиофилизировали. После этого эктодомен синдекана-1 суспендировали в DMEM. (Гибко) и определяли его количество по описанной выше методике. Опять-таки при 1 нМ концентрации такого иммуноаффинно-очищенного эктодомена синдекана-1 наблюдалось ингибирование роста обработанных тестостероном клеток S115 и лишь незначительный эффект достигался на клетках NMuMG (фиг.10). С другой стороны, гепарин сульфатные (HS) или хондроитин сульфатные (CS) гликозаминогликановые цепи сами по себе не подавляют рост клеток S115, даже если их используют с концентрацией, в тысячи раз превышающей концентрацию эктодомена синдекана-1 (фиг.11).
Пример 5
Влияние выделенного эктодомена синдекана-1 на культивированные клеточные линии
Ингибиторный эффект выделенного эктодомена синдекана-1 также тестировали на нескольких других клеточных линиях. Такие объекты включают плохо дифференцированные клетки карциномы чешуйчатых клеток (Carb), клетки опухоли грудной железы человека (MCF-7; АТСС НТВ 22), клетки S115, содержащие (S115+) и не содержащие гормон (S115-), NIH 3Т3 фибробласты (АТСС CRL, 1658), нормальные эпителиальные клетки грудной железы (NMuMG; АТСС CRL 1636), и человеческие кератиноцитные клетки (НаСаТ; фиг. 12).
Клетки культивировали и анализировали, как показано на фиг.8, в следующих средах в течение указанных периодов времени: CarB клетки (М. Quintanilla, К. Brown, М. Ramsden, A. Balmain, Nature 322, 78 (1991)) культивировали в HAM-F12-10% FCS в течение четырех дней; клетки MCF-7 культивировали в DMEM-5% FCS, дополненной 10 нМ эстрадиола (E2) и 10 мкг/мл инсулина в течение 4 дней; клетки S115+ и S115- культивировали, как показано на фиг.3, в течение 3 дней; клетки NIH 3Т3 культивировали в DMEM-5% FCS в течение 4 дней, NMuMG и HaCaT клетки культивировали в 10% FCS-DMEM в течение 4 дней. Поскольку клетки S115- растут значительно медленнее, чем клетки S115+, 3000 клеток S115- (для других клеточных линий 1500 клеток) пропорционально добавляли в лунку с целью получения результатов, сравнимых с полученными для клеток S115+. В связи с этим в случае клеток S115- 3000 клеток трансфекцировали на пластину в отличие от 1500 клеток для других образцов.
Линии клеток, образующие опухоли (CarB, MCF-7, S115+), демонстрировали сильное подавление роста при воздействии эктодомена синдекана-1 в концентрации 1 нМ (фиг.12). В отличие от этого наблюдалось лишь незначительное ингибирование или его отсутствие в случае других испытанных клеточных линий (S115-, NIH 3Т3, NMuMG, HaCaT; фиг.12), которые во всех отношениях являются неонкогенными. Воздействие гормона удваивает скорость роста клеток S115 (Leppa с сотр. , см. ссылку, цитированную выше), однако, если в культуру вводят эктодомен синдекана-1, рост клеток S115 в отсутствие андрогена происходит в 5,4 раза быстрее, чем рост этих же клеток S115 в присутствии тестостерона (фиг.12). Этот результат связан с ингибированием "злокачественного" поведения клеток S115+ и с ненарушенным ростом эпителоидных S115- клеток.
Пример 6
Подавление роста опухоли in vivo с помощью эктодомена синдекана-1
Эктоконструкцию получали в соответствии с методикой описанных выше примеров с использованием кДНК мышиного синдекана-1 полной длины, клонированной в Bluescript SK+ вектор (10) и мутагенного 33-основного олигонуклеотида 5'- GACACCTCCCAGTACTCACTTCCTGTCCAAAAG-3' [SEQ ID N:2), содержащего стоп-кодон (Bold) и Sсal сайт (CAGTAC), с целью превращения первой аминокислоты (Е), следующей после эктодоменового сайта, чувствительного к двухосновной протеазе, в стоп-кодон. Мутацию селектировали ограничительным перевариванием и подтверждали дидеокси секвенированием. Синдекан-1 дикого типа и цитоплазматический делеционный мутант клонировали в EcoRI сайт pBGS эвкариотного экспрессионного вектора (Mali с сотр., J. Biol. Chem. (1993) 268, 24215-24222). Экто мутант лигировали в Xhol сайт pMAMneo эукариотного трансфекционного вектора /Leppa с сотр., PNAS (1992) 89, 932-936), поскольку было известно, что pMAMneo трансфекцированные S115 клетки хорошо работают в биореакторной системе (персональное сообщение. Sari Ala-Uotila, Турецкий центр биотехнологии). S115 клетки трансфекцировали с использованием липосомной трансфекции и последующей селекции с помощью Генетицина, как было описано выше (Leppa с сотр., PNAS (1992) 89, 932-936).
Клетки S115 и трансфекцированные клеточные клоны культивировали в ДМЕМ-5% FBS-1 мМ пирувата Na в присутствии 10 нМ тестостерона, за исключением клеток S115-, которые культивировали без тестостерона в DMEM-4% DCC-PBS (фетальную телячью сыворотку обрабатывали древесным углем с декстрановым покрытием для удаления эндогенных стероидов из сыворотки) в присутствии 1 мМ пирувата Na.
Для роста опухоли субконфлюентные культуры отделяли с помощью трипсина, промывали DMEM и подсчитывали их количество с помощью счетчика Коултера (Коултер Электроникс). Клетки ресуспендировали в DMEM при плотности 5·107/мл и хранили на льду до инъекции. Атимным (Athymic) самцам мышей, лишенных волосяного покрова (nu/nu-BALB/cABom) в возрасте 6-8 недель (Бомхольтгард, Рай, Дания) делали подкожную инъекцию 0,2 мл клеточной суспензии. Одновременно имплантировали силатическую тестостероновую капсулу. У мышей регулярно наблюдали развитие опухоли и размер опухолей измеряли через определенные промежутки времени в двух перпендикулярных направлениях. После умертвления животных легкие и печень исследовали на возможное появление метастаз. Размер опухолей измеряли через 6, 11 и 15 дней после инъекции и наносили на график, представленный на фиг.13, средние значения для пяти индивидуальных опухолей. Трансфекцированные эктодоменом клетки давали лишь острую воспалительную реакцию и не обнаруживали опухолевого роста в отличие от клеток дикого типа, которые образовывали быстро растущие опухоли. Этот эксперимент демонстрирует эффективность эктодомена синдекана-1 в качестве агента, подавляющего опухоль in vivo.
На все цитированные ссылки указывается в тексте описания. После приведенного выше подробного описания изобретения специалисты в данной области должны понимать, что сфера изобретения может быть представлена широким и эквивалентным интервалом условий, параметров и т.д., не затрагивая при этом сущность сферы изобретения или любого его воплощения.










Изобретение относится к медицине, в частности к онкологии, и касается уменьшения роста опухолевой клетки. Для этого создают во внеклеточной среде указанной клетки концентрацию синдеканового эктодомена, равную 0,7-1,0 нМ. Для лечения опухолей предлагают также фармацевтическую композицию, содержащую в качестве активного ингредиента синдикановый эктодомен. Способ позволяет повысить эффективность подавления роста злокачественных клеток. 3 с. и 19 з. п. ф-лы, 13 ил.
| WO 9412162 A1, 09.06.94. |

