Настоящая заявка является частичным продолжением заявки на патент США NN 444881, поданной 1 декабря 1989 г, которая в свою очередь является частичным продолжением заявки на патент США N 118234, поданной 6 ноября 1987 г.
Настоящее изобретение осуществленно при поддержке правительства по программам AR37318 и AR36794, финансируемым Национальными институтами здоровья. Правительство имеет определенные права на данное изобретение.
Настоящее изобретение относится к способам обнаружения и регистрации разрушения коллагена in vivo. Более конкретно, изобретение относится к способам количественного определения сшитых телопептидов, образованных in vivo при разрушении коллагена типа II и III.
К настоящему времени описано три класса коллагенов. Известно, что коллагены класса I, подразделяемые на типы I, II, III, V и IX, образуют фибрилы. Коллагены всех этих типов синтезируются в виде молекул предколлагена (проколлагена), состоящих из N-концевых и C-концевых предпептидов (пропептидов), присоединенных к центральной молекуле коллагена. После удаления этих предпептидов, что в природе происходит in vivo в ходе синтеза коллагена, и оставшаяся центральная часть молекулы коллагена в основном состоит из трехспирального домена, обладающего концевыми телопептидными последовательностями, не являющимися трехспиральными. Такие телопептидные последовательности играют важную роль в качестве участков межмолекулярного сшивания вне клеток фибрилл коллагена.
Настоящее изобретение относится к способам обнаружения разрушения коллагена, основанным на анализе конкретных сшитых телопептидов, образованных in vivo в ходе разрушения коллагена. В прошлом были созданы методы анализа с регистрацией разрушения коллагена in vivo путем определения различных биохимических маркеров, некоторые из которых являются продуктами разрушения коллагена. Например, деформацию костей, связанную с болезнью Педжета, регистрируют путем определения небольших пептидов, содержащих гидроксипролин и выделяемых в мочу после разрушения костного коллагена. Russell и др. Metab.Bone Dis. and Rel.Res. 4 и 5, 255 - 262 (1981), Singer F.R. и др. Метаболические костные заболевания, т. II (ред. Aviol, L.V и Kane S.M.), 489 - 575 (1975), Академик Пресс, Нью-Йорк.
Другими исследованиями определялось в моче такое соединение с поперечными связями (сшитое), как пиридинолин в качестве показателя разрушения коллагена при заболеваниях суставов. См. например, в качестве прототипа Wu и Eyre, Biochemistry, 23 1850 (1984), Black и др. Annals of the Rheumatic Diseases 48 641 - 644 (1989), Robins и др. Annals of the Rhevmatic Diseases 45 969-973 (1986), Seibel и др. The Jovrnal of Rhevmatology, 16 964 (1989). В отличие от настоящего изобретения некоторыми прежними исследователями пептиды в биологических жидкостях подвергались гидролизу, после чего проводился анализ на присутствие отдельных гидроксипиридиниевых остатков. Ни одним из этих исследователей не сообщалось об определении телопептида, содержащего первичную связь и образовавшегося естественным путем in vivo при разрушении коллагена, что характерно для настоящего изобретения.
В заявке на патент Великобритании CB 2205643 сообщается, что разрушение в организме коллагена типа III определяют количественно путем определения в биологической жидкости концентрации II-концевого телопептида из коллагена типа III. в той же ссылке утверждается, что сшитые области телопептида нежелательны. В самом деле, в ссылке говорится, что для получения телопептида необходимо использовать несшитый источник коллагена. Пептиды настоящего изобретения имеют поперечные связи (сшиты) во всех случаях. Поперечные связи в коллагене более подробно рассматриваются ниже в разделе "Сшивание коллагена".
Имеется целый ряд сообщений о том, что разрушение коллагена может быть определено количественным анализом определенных предколлагеновых пептидов. В настоящем изобретении речь идет о телопептидах, а не о предпептидах, отличающихся друг от друга их расположением в молекуле коллагена, а также временем их отщепления in vivo См. патент США 4504587, патент США 4312853, Pierard и др. Analytical Biochemistry 141, 127 - 136 (1984), Niemela, Clin, Chem. 31(8), 1301 - 1304 (1985), Rohde, и др. European Jovrhal of Clinical Investigation, 9 451 - 459 (1979), WO 89/04491.
Патент США 4778768 относится к способу определения изменений, происходящих в хряще артикулятора, путем количественного анализа мономера протеогликана или его антигенных фрагментов в образце синовиальной жидкости. Данный патент не относится к обнаружению сшитых телопептидов, происходящих из разрушенного коллагена.
В работе Dodge, J.Clin., Unvest. 83, 647 - 661 (1981) раскрываются способы анализа разрушения коллагена типа II с применением поликлональной антисыворотки, специфично реагирующей с нераспленными альфа-цепями и пептидами, происходящими из коллагенов типа II человека и коровы под воздействием бромистого циана. Пептиды, о которых идет речь, не являются сшитыми телопептидами, как в настоящем изобретении.
Исследователями нескольких групп исследованы и определены аминокислотные последовательности коллагена человека типа III, преда l (II) коллагена человека и всей цепи полностью предшественника преда l (III) коллагена человека, а также соответствующих клоны кДНК. См. Loidl и др. Nucleic Acids Research 12, 9383 - 9394 (1984) Sangiorgi и др. Nucleic Acids Research, 13, 2207 - 2225 (1985), Baldwin и др. Biochem. J. 262 521 - 528 (1989), Ala-Kokko и др. Biochem. J. 260, 509 - 516 (1989) Ни в одной из приведенных ссылок не указано строение конкретных телопептидов в качестве продуктов разрушения, анализом которых можно определить in vivo количество разрушенного фибриллярного коллагена.
Несмотря на вышеприведенные прототипы, сохраняется необходимость в простых и эффективных методах анализа для определения разрушения коллагена in vivo. Подобные методы анализа могут быть использованы для обнаружения и регистрации протекания заболевания, такого как остеоартрит (разрушения коллагена типа II) и различные воспалительные нарушения, такие как васкулитный синдром (разрушение коллагена типа III).
Методы анализа разрушения коллагена типа I раскрыты в первичной заявке на патент США N 118234 и могут быть использованы для обнаружения и оценки костной резорбции. Обнаружение костной резорбции может представлять интерес для обнаружения и регистрации таких заболеваний, как остеопороз. Остеопороз - это наиболее обычное костное заболевание человека. Первичный остеопороз с повышенной склонностью к переломам возникает в результате прогрессирующей общей потери скелетной костной массы. Установлено, что в США им поражено 15 - 20 миллионов людей. Причиной заболевания является дисбаланс в реконструкции костей, т.е. в скоростях синтеза и разрушения костной ткани.
Ежегодно для пожилых людей отмечается около 1,2 миллиона случаев перелома, связанных с остеопорозом, в том числе: примерно 538000 случаев компрессионного перелома позвоночника, примерно 22700 случаев перелома бедра и значительное число преждевременных переломов периферических костей. В случае перелома бедра 12 - 20% заканчиваются фатально вследствие тяжких травм и кровотечения, а половина выживших больных требует ухода в домашних условиях. Общие потери от связанных с остеопорозом травм в настоящее время достигают по меньшей мере 7 миллионов долларов (Barnes O.M. Science, 236, 914 (1987).
Остеопороз наиболее обычен для женщин в состоянии менопаузы, которые в среднем теряют 15% костной массы в течение 10 лет после наступления менопаузы. Это заболевание характерно также и для мужчин по мере их старения и для молодых спортсменок с аменореей. Несмотря на большие и все более возрастающие социальные и экономические последствия остеопороза, метода определения скорости костной резорбции у больных и нормальных субъектов не существует. Основная трудность в регистрации заболевания заключается в отсутствии специфического анализа для определения скорости костной резорбции.
Способы оценки костной массы зачастую основаны на определении кальция во всем организме в анализе с активацией нейтронами или определении минеральной массы в данной кости по поглощению фотонов. Подобные определения способны создать лишь долговременные впечатления о возможном уменьшении костной массы. Определение кальциевого равновесия путем сравнения его поглощения с расходом утомительно, ненадежно и позволяет всего лишь косвенно оценить возможные потери костных минералов за длительный период. Другие применяемые в настоящее время методы оценки костной массы и затрудненного костного метаболизма включают количественную сканирующую радиометрию в избранных месторасположениях костной (запястье, пяточная кость и т.д.) и гистоморфометрию подвздошного гребня.
В первом случае дается грубая мера содержания минеральной части в специфичном участке единственной кости. Гистоморфометрия дает полуколичественную оценку равновесия между вновь осажденным костным швом и резорбированными поверхностями.
Анализ мочи на объем разрушения кости во всем организме за 24 часа мог бы оказаться гораздо более полезным. Изучением минеральной части (например, кальциевого равновесия) не позволяет сделать этого надежно или просто. Поскольку костная резорбция включает разрушение минеральной и органической матрицы, идеальным показателем мог бы оказаться специфичный биохимический маркер в биологических жидкостях для вновь образовавшихся продуктов разрушения кости. Испытаны некоторые потенциальные органические метки. Например, гидроксипролин-аминокислота, сильно ограниченная в коллагене, и основной структурный белок в костной и прочих соединительных тканях, экстретируются в мочу. Как известно, степень его экскреции возрастает при определенных условиях, особенно в случае болезни Педжета - нарушение костного метаболизма, при котором, как указано выше, резко возрастает деформация костей. По этой причине гидроксипролин интенсивно использовался в качестве аминокислотного маркера разрушения коллагена (Singer F.R. и др. 1978, см. выше).
В патенте США N 3600132 раскрыт способ определения гидроксипролина в биологических жидкостях, таких как моча, сыворотка, поясничная жидкость и другие межклеточные жидкости, с целью регистрации отклонений в метаболизме коллагена. В частности, создатель этого патента отмечает, что в патологических состояниях, таких как болезнь Педжета, синдром Марфана, остеопсатароз, рост непластов в коллагеновых тканях и различные формы карликовости, может быть определен путем анализа содержания гидроксипролина повышенный анаболизм или катаболизм коллагена. Изобретатель определяет гидроксипролин его окислением перекисью водорода и N-хлор-п-толуолсульфонамидов в производное пиррола с последующим колориметрическим определением в п-диметиламинобензальдегиде.
В случае болезни Педжета повышенное содержание в моче гидроксипролина вероятно в основном является результатом разрушения костей, однако, гидроксипролин не может быть, как правило, в качестве специфического показателя. Большая часть гидроксипролина в моче может возникать в результате синтеза нового коллагена (значительные количества вновь образованного белка разрушаются и экскретируются, даже не войдя в состав ткани) и в результате оборота определенных белков крови, а также и других белков, содержащих гидроксипролин. Более того, примерно 80% свободного гидроксипролина, происходящего в результате разрушения белка, подвергается метаболизму в печени и вовсе не попадает в мочу.
Kivirko K.I.Int. Rev. Connect. Tissue Res. 5 93 (1970) и Weis P.H. Klein L. Clin. Invest 48 I (1969)
Гидроксилизин и его гликозидные производные, принадлежащие к коллагеновым белкам, считаются более действенными в качестве маркеров разрушения коллагена, чем гидроксипролин. Однако по тем же причинам, приведенным выше для гидроксипролина, гидроксилизин и его гликозиды, вероятно в равной степени не могут служить специфическими маркерами резорбции кости. Krane S.M. Simon L. S.Develop. Biochem. 22, 185 (1981).
Помимо уникальных для коллагена аминокислот различные неколлагеновые белки костной ткани, такие как остеокальцин, или продукты разрушения таких белков создали основу иммуноализа, направленного на определения костного метаболизма. Pvice P.A. и др. J. Clin. Invest, 66 878 (1980) и Gundber C.M. т др. Meth. Enzymol, 107 516 (1984). Однако сейчас очевидно, что неколлагеновые белки костного происхождения, являясь потенциально полезными метками костной метаболической активности, вряд ли сами по себе способны служить количественной мерой костной резорбции. Например, концентрация остеокальцина в сыворотке крови колеблется в довольно широких пределах как при отсутствии, так и при наличии метаболического костного заболевания. Концентрация остеокальцина повышена в состоянии повышенного разрушения скелета, однако, неясно является ли это следствием возросшего синтеза или разрешения костей. Krane S.M. и др. Develop. Biochem, 22 (1981), Price P.A. и др. j. Clin. Invest, 66 878 (1980) и Gundber C.M. и др. Meth. Enzymol, 107, 516 (1984)
Полимеры типа, принадлежащего к коллагену позвоночных, для нормального функционирования требуют образования за счет альдегидных групп поперечных связей. Альдегиды коллагена происходят от нескольких специчных лизиновых или гидроксилизиновых боковых цепей под действием лизилоксидазы. Различные ди-, три- и тетрафункциональные сшивающие аминокислоты образуются в спонтанных внутри- и межмолекулярных реакциях таких альдегидов в пределах вновь образованных коллагеновых полимеров, при этом тип сшивающего остатка меняется специфично в зависимости от типа ткани (см. Eyre D.R. и др. Ann. Rev. Biochem, 53 717 - 748 (1984).
Можно отличить два основных пути сшивания для полосатого (повтор в 67 нм) фибриллярного коллагена, один из которых основан на лизиновых альдегидах, а другой - на гидроксилизиновых альдегидах. Путь с участием лизиновых альдегидов доминирует в коже взрослого субъекта, роговице, склере и сухожилие крысиного хвоста, а также часто протекает в других мягких соединительных тканях. Путь с участием гидроксилизиновых альдегидов доминирует в костях, хрящах, связках, большинстве сухожилий и большинстве внутренних соединительных тканей организма (Eyre D.R. и др. (1974) см. выше). Выбор пути зависит от того, гидроксилированы или нет лизиновые остатки в тех участках телопептида, в которых позднее под действием лизилоксидазы будут образованы альдегидные остатки (Barnes M.J. и др. Biochem. J. 139 461 (1974)
Химическое строение зрелых сшитых аминокислот, образованных на пути с участием лизиновых альдегидов, неизвестно, однако остатки гидроксипиридиния идентифицированы в качестве зрелых продуктов на пути с участием гидроксилизиновых альдегидов. На обоих путях и в большинстве тканей промежуточные, восстанавливаемые боргидридом остатки исчезают по мере вызревания вновь образованного коллагена, указывая на то, что они являются сравнительно короткоживущими промежутками образования (Bailey, A.J. и др. FEBS Lett, 16 86 (1971). Исключения составляют кости и дентин, в которых способные восстанавливаться остатки сохраняются в заметной концентрации в течение всей жизни, что видимо частично связано с быстрой минерализацией вновь образованных коллагеновых фибрилл, которые ингибируют дальнейшее спонтанное сшивающее взаимодействие (Eyre D.R. в книге "Химия и биология минерализованных соединительных" ред. Veis A. , стр. 51 - 55 (1981), Эльзевир, Нью-Йорк, а также Walter S.C. и др. Calc. Tiss Inll, 35 401 - 405 (1983).
Идентифицированы две химические формы 3-гидроксипиридиевой поперечной связи (формулы I и II). Оба соединения характеризуются естественной флуоресценцией с одинаковым характеристическим спектром возбуждения и эмиссии (Fujim oto D. и др. Biochem. Biophys. ReS. Commun, 76 1124 (1977) и Eyre D. P. Develop. Biochem., 22 50 (1981). Данные аминокислоты могут быть разделены и подвергнуты анализу непосредственно в тканевых гидролизатах с хорошей чувствительностью с использованием ВЭЖХ с обращением фаз и флуоресцентного детектирования (Eyre D. R. и др. Analyle. Biochem. 137, 380 - 388 (1984) Необходимо отметить, что настоящее изобретение включает количественное определение конкретных пептидов, а не аминокислот.

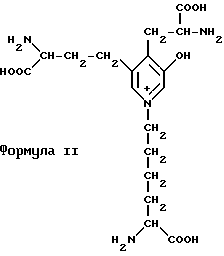
Известно, что у растущих животных указанные вызревшие поперечные связи могут концентрироваться в большей степени в неминерализованной части костного коллагена по сравнению с минерализованным коллагеном (Banes A.J. и др. Biochem. Biophys. Res. Commun. 113 (1983) Однако другие исследования, проведенные на костях телят или взрослого человека, не подтверждают данной концепции (Eyre D.R. в книге "Химия и биология минерализованных тканей" (ред. Bullev. W.T.), стр. 105, (1985), Эбско Медиа Инк, Бирмингем, Алабама).
О присутствии коллагеновых гидроксипиридиниевых сшивок в моче впервые сообщено Gunja-Smith и Boucek (Gunja-Smith. Z и Boucek R.J., Biochem. J., 187, 759 - 762 (1981), которые использовали длительную процедуру выделения пептидов и обычный минокислотный анализ. В то время им была известна только ГП-форма сшивки. В работе Robins SP, Biochem. J., 207 617 - 620 (1982) сообщается о ферментивно связанном иммуноаналиазе для определения ГП в моче, в котором создают поликлональные антитела к свободной аминокислоте, сопряженной с альбумином бычьей сыворотки. Данный анализ направлен на создание указателя для регистрации повышенного разрушения суставов, происходящего в случае артритных заболеваний, и основан, согласно Robins, на том открытии, что пиридинолин в гораздо большей степени преобладает в хрящах, а не в костном коллагене.
В более поздней работе, относящейся к ферментативно связанному иммуноанализу, Robins сообщает, что лизилпиридинолин нереакционноспособен по отношению к антисыворотке к пиридинолину, ковалентно связанному с альбумином бычьей сыворотки (Robins и др. , Ann. Pheum. Diseases, 45 969 - 973 (1986) Указатель Робинса в моче для разрушения хрящей основан на том открытии, что гидроксилизилпиридинолин, происходящий в первую очередь из хрящей, обнаруживается в моче в концентрациях, пропорциональных степени резорбции (т.е. разрушения) хряща суставов. В принципе этот указатель может быть использован и для определения потерь хряща во всем организме, однако, при этом невозможно получить никакой информации о костной резорбции.
Также представляет интерес международная публикация N WO 89/12824 Исследовательского Института Роуэтта. Данная последняя публикация раскрывает способ регистрации разрушения коллагена, включающий анализ образца биологической жидкости, содержащего фрагмент коллагена, включающий лизилпиридинолин или гидроксилизилпиридинолин, или их замещенную форму и метод определения ткани, из которой происходит разрушенный коллаген.
Таким образом, существует необходимость в методе, позволяющем определить степень костной резорбции во всем человеческом организме. Наиболее полезный метод будет в том случае, если его можно будет применить к биологическим жидкостям, особенно к моче. Метод должен быть чувствительным, т.е. допускающим количественные определения вплоть до 1 пикомоля, и быстр, т.е. позволяющий определять степень костной резорбции за 24 часа, с тем, чтобы можно было оценить эффективность различных видов лечения (например, эстрогеном)
Настоящее изобретение основано на открытии присутствия в биологических жидкостях больных и нормальных людей конкретных сшитых телопептидов. Эти телопептиды образуются in vivo в ходе разрушения и регенерации коллагена. Термин "телопептиды" используется здесь в широком смысле для обозначения сшитых пептидов, обладающих последовательностями, которые связаны с телопептидной областью, например коллагена типа II и типа III, и которые могут быть сшиты с остатком или пептидом, относящимся к трехспиральному домену коллагена. Как правило, описываемые здесь телопептиды имеют меньшее число аминокислотных остатков, чем полные телопептидные домены коллагенов типа II и типа III. Обычно телопептиды настоящего изобретения состоят из двух пептидов, соединенных пиридиниевой поперечной связью, и, кроме того, соединенных пиридиниевой поперечной связью с остатком или пептидом трехспирального домена коллагена. Раскрывая здесь строение этих телопептидов, необходимо указать (а для специалиста это очевидно), что они могут быть получены не только in vivo, например синтетически. Указанные пептиды, как правило, получают в очищенном виде, т. е. по существу без примесей, в частности без примесей других пептидов.
Настоящее изобретение, кроме того, относится к способам определения in vivo разрушения коллагенов типа II и типа III. Способы включают количественное определение в биологической жидкости концентрации конкретного телопептида, обладающего 3-гидроксипиридиниевой поперечной связью и происходящего от разрушения коллагена. Раскрываемые настоящим изобретением способы аналогичны способам, ранее раскрытым в заявке на патент США N 118234, поданной 6 ноября 1987 г, для определения абсолютной степени костной резорбции in vivo. Данные способы включают количественное определение в биологической жидкости концентрации телопептидов, обладающих 3-гидроксипиридиниевой поперечной связью и возникающих в результате костной коллагеновой резорбции.
В представительном анализе биологическую жидкость пациента контактируют с иммунологическим связывающим партнером, специфичным на телопептид, обладающий 3-гидроксипиридиниевой поперечной связью и происходящий из коллагена типа II и типа III. Биологическая жидкость может быть использована как таковая или же может быть очищена перед стадией контактирования. Такая стадия очистки может быть осуществлена с помощью целого ряда стандартных методик, в том числе: адсорбции и элюирования в патронах, хроматографии на молекулярных ситах, диализа, ионообменной хроматографии, хроматографии на окиси алюминия, хроматографии на гидроксиапатите и сочетании таких методик.
Другие представленные воплощения количественного определения концентрации фрагментов пептида, обладающего 3-гидроксипиридиниевой поперечной связью, в биологической жидкости включают электрохимическое титрование, спектроскопию естественной флуоресценции и поглощение в ультрафиолетовой области спектра. Электрохимическое титрование может быть проведено непосредственно на биологической жидкости без ее дополнительной очистки. Однако, если это невозможно из-за большого количества загрязняющих веществ биологическую жидкость перед стадией электрохимического титрования вначале очищают. Приемлемые способы очистки перед электрохимическим детектированием включают: диализ, ионообменную хроматографию, хроматографию на окиси алюминия, хроматографию на молекулярных ситах, хроматографию на гидроксиапатите и ионообменную абсорбцию и элюирование.
Флуорометрический анализ биологической жидкости, содержащей 3-гидроксипиридиниевую поперечную связь, является альтернативным путем количественного определения разрушения коллагена (а следовательно, и костной резорбции, если количественно определяют пептид типа I). Флуорометрический анализ может быть осуществлен непосредственно на биологической жидкости без дополнительной ее очистки. Однако для определенных биологических жидкостей, в частности мочи, рекомендуется перед флуорометрическим анализом проведение очистки. Такая стадия очистки состоит из диализа аликвотного количества биологической жидкости, например мочи, относительно водного раствора с получением в результате частично очищенных фрагментов пептида, удержанных в непродиффундировавшем веществе (Ретентате). Непродиффундировавшее вещество затем лиофилизуют, растворяют в растворе с ионной парой и адсорбируют на колонке для аффинной хроматографии. Такие очищенные фрагменты пептида затем могут быть гидролизованы, а продукты гидролиза могут быть разделены хроматографически. Хроматографическое разделение может быть осуществлено либо с помощью высоко эффективной жидкостной хроматографии, либо с помощью микроканальной высокоэффективной жидкостной хроматографии.
Изобретение включает пептиды, строение которых идентично строению пептидов, образующихся при разрушении коллагена и по существу несодержащих других пептидов человеческого организма, и которые могут быть получены из биологической жидкости. Такие пептиды имеют по меньшей мере одну 3-гидроксипиридиниевую поперечную связь, в частности лизилпиридинолиновую поперечную связь или гидроксилизилпиридинолиновую поперечную связь, и происходят из телопептидной области коллагена типа II или типа III, связанной с одним или несколькими остатками из трехспирального домена, обычно под воздействием эндогенных протеаз и/или пептидаз.
Строение телопептидов типа II и типа III обсуждается ниже. Кроме того, включена информация о телопептидах типа I, первоначальным источником которой является заявка на патент США N 118234.
Другой аспект настоящего изобретения включает анализ на раскрываемые здесь пептиды, в которых циклы пиридиния интактны и расщеплены. Поскольку подозревают, что некоторые из циклов пиридиния происходят in vivo, анализ, в котором обнаруживают как интактные, так и расщепленные циклы пиридиния, может привести к более тонкой оценке разрушения коллагена. В связи с данным аспектом настоящего изобретения в анализе могут быть использованы специфические связующие партнеры для индивидуальных пептидов, содержащих интактные или расщепленные циклы пиридиния. Могут быть использованы индивидуальные специфичные связующие партнеры, распознающие пептиды обоих типов (пептиды, содержащие как интактные, так и расщепленные циклы пиридиния). Или же могут быть применены специфичные связующие партнеры, различающие пептиды, содержащие интактный цикл пиридиния, и пептиды, в которых цикл пиридиния расщеплен.
Строение сшитых телопептидов, происходящих из коллагена типа I
Специфичный телопептид, обладающий 3-гидроксипиридиниевой поперечной связью, происходящей из N-концевого (аминоконцевого) телопептидного домена костного коллагена типа I, характеризуется следующей аминокислотной последовательностью:
Формула III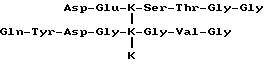
где
представляет гидроксилизилпиридинолин или лизилпиридинолин и Gln-глутамин или пирролидинкарбоновая кислота.
Изобретение также направлено на пептид, содержащий по меньшей мере одну 3-гидроксипиридиниевую поперечную связь, происходящую из C-концевого (карбоксиконцевого) телопептидного домена костного коллагена типа I. Такая C-концевая телопептидная последовательность сшита либо лизилпиридинолином, либо гидроксилизилпиридинолином. Пример подобной пептидной последовательности представлен формулой:
Формула IV
где
представляет гидроксилизил- или лизилпиридинолин.
Со времени подачи заявки на патент США N 118234 изобретателем получены доказательства существования в биологических жидкостях двух дополнительных телопептидов коллагена типа I, имеющих следующее строение:
Формула V
и
Формула VI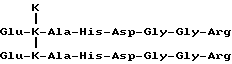
Данные телопептиды могут также быть количественно определены способом настоящего изобретения
Строение сшитого телопептида, происходящего из коллагена типа II
Специфичный телопептид, обладающий гидроксилизилпиридинолиновой поперечной связью, происходящей из C-концевого телопептидного домена коллагена типа II, характеризуется следующей аминокислотной последовательностью (далее называемой центральной пептидной структурой):
Формула VII
где сшивающий остаток, обозначенный как Hyl-Hyl-Hyl, представлен гидроксипиридинолином (ГП)-естественным 3-гироксипиридиниевым остатком, присутствующим в зрелых коллагеновых фибрилах различных тканей.
Аминоконцевые телопептиды из коллагена типа II не обнаружены в биологических жидкостях и подозревают, что потенциальные пептиды, происходящие из N-концевой телопептидной области колагена типа II по существу разрушаются in vivo, возможно с перекрыванием всех путей к свободной ГП сшивающей аминокислоте.
Строение сшитых телопептидов, происходящих из коллагена типа III
По аналогии с вышеизложенным в биологических жидкостях могут присутствовать сшитые пептиды, возникающие в результате протеолиза человеческого коллагена типа III. Такие пептиды обладают центральной структурой, отраженной следующими схожими формулами:
Формула VIII
Формула IX
где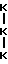
представляет гидроксилизил- или лизилпиридинолин и Gln - глутамин или пирролидинкарбоновая кислота
Скорее всего в биологических жидкостях сшитый пептид, происходящий из коллагена типа III, обладает центральной структурой формулы:
Формула X
происходящей из двух aI(III)N-телопептидных доменов, связанных с остатком гидроксилизилпиридинолина (Hyl-Hyl-Hyl).
Второй возможный фрагмент C-телопептидного сшивающего домена, если основываться на наблюдаемых в моче пептидах коллагена типа I и II, обладает центральной структурой формулы:
Формула XI
В биологических жидкостях, кроме того, могут присутствовать и подвергнуться определению меньшие и большие варианты (отличающиеся на одну-три аминокислоту в каждом компоненте цепи) указанных двух пептидов, соответствующих родственным последовательностям, приведенным выше (формулы VIII и IX). Аналогичные меньшие и большие варианты каждого пептида, охарактеризованного здесь, составляют часть настоящего изобретения.
Изобретение в целом включает всех специфичных связующих партнеров для охарактеризованных здесь пептидов. "Специфичные связующие партнеры" - это молекулы, способные связываться с пептидами настоящего изобретения. Этот термин включает иммунологических связующих партнеров, таких как антитела (моноклональные и поликлональные), антиген-связующие фрагменты антител (например, Fab и F(ab')2 фрагменты), одноцепные антиген-связующие молекулы и т.п., полученные гибридомной или рДНК технологией.
Изобретение включает слитые клеточные гибриды (гибридомы), продуцирующие моноклональные антитела, специфичные к вышеохарактеризованным коллагеновым пептидам, обладающим 3-гидроксипиридиниевыми поперечными связями (как с интактным циклом пиридиния, так и с циклом, который расщеплен).
Изобретение, кроме того, включает моноклональные антитела, продуцированные слитыми клеточными гибридами, и эти же антитела (а также их связующие фрагменты, например Fab) в сочетании с поддающимся обнаружению маркером. Примеры поддающихся обнаружению маркеров включают: ферменты, хромофоры, флуорофоры, коферменты, ингибиторы ферментов, хемилюминесцетные вещества, парамагнитные металлы, спиновые метки и радиозотопы. Или же подобные специфичные связующие партнеры могут быть связаны с одним из членов лиганд-связующего комплекса (например, авидин-биотин), и в этом случае поддающийся обнаружению маркер может поставляться связанным с дополняющим членом комплекса.
Изобретение также включает тест-набор, применимый для количественного определения пептидов, обладающих 3-гидроксипиридиниевыми поперечными связями, возникающими в биологической жидкости в результате разрушения коллагена. Набор может включать специфичный связующий партнер для пептида, происходящего из разрушенного коллагена. Специфичные связующие партнеры могут в тест-наборе находится в сочетании с поддающимся обнаружению маркером или членом лиганд-связующего комплекса (см. выше)
На фиг. 1 представлен коллаген типа I и предполагаемый источник телопептида. Не установлено, происходят ли два показанных телопептида из одной молекулы коллагена, как показано на фиг. 1, или же из двух молекул коллагена.
На фиг. 2 показана относительная флуоресценция (возбуждение при 297 нм, эмиссия при 390 нм) относительного номера фракции (4 мл), полученных в ходе очистки сшитых телопептидов хроматографией на молекулярных ситах. Сшитые коллагеновые телопептиды типа II содержатся во фракциях, обозначенных, как II.
На фиг. 3A показана относительная флуоресценция (возбуждение при 330 нм, эмиссия при 390 нм) относительно времени удерживания фракций ионообменной ВЭЖХ (ЕАЕ-5Р). Сшитые коллагеновые телопептиды типа II содержатся во фракции, обозначенной цифрой IV.
На фиг. 3B показано поглощение (297 нм) относительно времени элюирования в минутах для той же хроматографии.
На фиг. 4A показана относительная флуоресценции (возбуждение при 297 нм, эмиссия при 390 нм) по отношению к времени элюирования фракций, полученных в ходе ВЭЖХ с образованием фаз. Сшитые коллагеновые телопептиды типа II элюируются, как показано. Фракции, указанные черточкой (-), свидетельствуют на основании секвенсционного и композиционного анализа указанных пептидов о сохранении или потери в них gly(C) или pro(P) остатков.
На фиг. 4B показано поглощение (220 нм) как функция времени элюирования ВЭЖХ с обращением фаз.
На фиг. 5 приведено сравнение концентрации ГП и ЛП в кортикальном слое и губчатом веществе костей человека в зависимости от возраста.
На фиг. 6 отражено молярное отношение сшивок в ГЛ-ЛП, как функции возроста.
На фиг. 7A показана относительная флуоресценция (возбуждение при 297 нм, эмиссия при > 370 нм) как функция объема элюирования в ходе разделения сшитых N-телопептидов коллагена типа I с помощью ВЭЖХ с обращением фаз.
На фиг. 7B показана относительная флуоресценция (возбуждение при 297 нм, эмиссия при > 370 нм) относительно объема элюирования в ходе разделения сшитых C-телопептидов коллагена типа I с помощью ВЭЖХ с обращением фаз.
На фиг. 8A показана относительная флуоресценция (возбуждение при 297 нм, эмиссия при 380 нм) как функция времени элюирования сшитых коллагеновых телопептидов типа I.
На фиг. 8B показана относительная флуоресценция (возбуждение при 297 нм, эмиссия при 380 нм) как функция времени элюирования сшитых коллагеновых телопептидов типа I.
На фиг. 9 представлены опыты по связыванию представительных моноклональных антител HB 10611 с P1 пептидом (вышеприведенной формулы III, чистые квадратики), a2 (I) N-телопептидом (QYDGKGVGC, черные ромбики) и a1 (I) N-телопептидом (YDEKSTGGC, черные квадратики).
На фиг. 10 показано строение N-телопептидной области декальцинированного костного коллагена человека. P1 пептид (формула III) заключен в рамку, пептид содержит эпитоп, коорелированный с костной резорбцией.
Телопептиды коллагена типа II
Центральная пептидная структура пептидов коллагена типа II может быть обнаружена в биологических жидкостях в виде компонента более крупных пептидов, несущих дополнительные аминокислоты или аминокислотные последовательности на одном или нескольких концах трехпептидных последовательностей, соединенных остатком ГП. На фиг. 1 показано, как телопептиды коллагена типа II, соединенные с трехспиральной последовательностью, могут быть образованы in vivo из человеческого источника под воздействием протеолитических ферментов-пепсина и трипсина. В биологических жидкостях могут также присутствовать и фрагменты меньшего размера, потерявшие в центральной пептидной структуре аминокислоты, в частности фрагменты из спиральной последовательности.
Как правило, вставки или делеции аминокислот в центральной пептидной структуре составляют 1-3 аминокислоты. Дополнительные аминокислоты, как правило, будут определяться телопептидной последовательностью коллагена типа II, встречающейся в естественных условиях in vivo. В качестве примеров приведены пептиды следующего строения:
Формула XII
и
формула XIII
которые могут быть выделены хроматографически из мочи, и пептид иного строения:
формула XIV
также может быть выделен из мочи. Кроме того, могут присутствовать гликозилированные варианты центральной структуры и ее большие или меньшие варианты, в которых остаток галактозы или остаток глюкозилгалактозы присоединен к гидроксилу боковой цепи сшивающего ГП остатка. Каждый пик на графиках, приведенных на фиг. 4A и 4B, может соответствовать сшитому фрагменту, конкретного строения, который может быть количественно определен в целях настоящего изобретения.
Приведенные структуры отвечают сайту их происхождения в количестве типа II фибрил человека, т.е. сайту молекулярного сшивания, образованному между двумя a1(II)C - телопептидами и остаток 87 трехспирального домена, для которых известны следующие последовательности:
формула XV
Выделенные пептидные фрагменты представляют собой продукты протеолитического разрушения в организме коллагена типа II фибрил. Центральная структура, содержащая остаток ГП, сравнительно устойчива к дальнейшему протеолизу и может служить количественной мерой разрушения коллагена типа II.
Коллаген типа II присутствует в стекловидном хряще суставов скелета взрослого индивидуума. Количественное определение телопептидов коллагена типа II в биологической жидкости, например, с помощью моноклональных антител, распознающих эпитоп в пептидной структуре, могло бы послужить количественной мерой разрушения или регенерации хряща во всем организме. Настоящее изобретение в его рекомендуемом воплощении включает анализ разрушения хрящевой ткани человека, основанный на количественном определении степени экскреции в мочу по меньшей мере одного из членов такого семейства телопептидов. Такой анализ может быть использован, например, для:
(1) отбора взрослых людей с выявлением индивидуумов с ненормально высокой степенью разрушения хряща в качестве раннего индикатора остеоартрита,
(2) регистрации действия потенциальных противоартритных лекарств на хрящевой метаболизм у больных остеоартритом и ревматоидным артридом,
(3) регистрации развития вырожденных заболеваний суставов у больных остеоартритом и ревматоидным артритом и их реакцию на различное лечебное вмешательство.
Остеоартрит - это вырожденное заболевание суставных хрящей. На ранних этапах в большинстве случаев не имеет воспалительного характера (т.е. отличен от ревматоидного артрита). Это не какая-то единственная болезнь, а последние этапы выхода сустава из строя в результате различных факторов (например: генетической предрасположенности, механических перегрузок, врожденных пороков суставов, прежней травмы и т.д.). Разрушение суставного хряща - это центральный признак прогрессирующего остеоартрита. Случаи остеоартрита, основываясь на радиографических обследованиях, составляют 4% для возрастной группы в 18-24 года и до 85% для возрастной группы в 75-79 лет. В настоящее время болезнь может быть диагностирована по боли и радиографически или по другим отраженным симптомам эрозии хряща.
Вышеприведенный анализ может быть использован для обнаружения ранних свидетельств ускоренного разрушения хряща у больных со слабовыраженными симптомами для регистрации развития болезни у больных с прогрессирующим заболеванием и в качестве средства регистрации действия лекарств или другого лечебного вмешательства.
У нормальных молодых людей (со сформированных скелетом) разрушение хрящевого коллагена очень маловероятно. Тест, позволяющий определить фрагменты хрящевого коллагена в моче (а также в крови и синовиальной жидкости), был бы очень полезен для оценки "здоровья" хрящей во всем организме и отдельных суставах. Вышеприведенный специфичный для коллагена типа I анализ может быть использован для пептида коллагена типа II анализ позволяет решить эту задачу. В недалеком будущем подобный анализ мог быть стать рутинным диагностическим инструментом для выявления тех индивидуумов, суставы которых изношены. Их можно было бы раньше направлять на профилактическое лечение, например, следующим поклонением так называемых хондрозащитных лекарств, разрабатываемых в настоящее время основными фармацевтическими фирмами, которые активно ищут лучшие средства лечения остеоартрита.
Другие заболевания, при которых разрушается суставный хрящ, включают: ревматоидный артрит, анкилозирующий спондилоартрит, болезнь Стилла, псориазный артрит, синдром Рейтера, рецидивный полихондрит, синдром боли в нижней части спины и другие инфекционные формы артрита. Вышеприведенный специфичный для коллагена типа II анализ может быть использован для диагностирования и регистрации указанных заболеваний и выявления из реакции на лечение, что показано выше на примере остеоартрита.
Телопептиды коллагена типа III
Как указано выше, телопептиды человеческого коллагена типа III, которые могут присутствовать в биологических жидкостях, как можно ожидать, будут обладать центральной структурой, отраженной следующими родственными формулами:
Формула VIII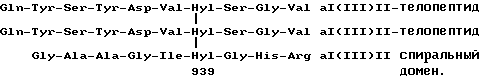
и
Формула IX
где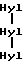
представляет гидроксилизилпиридинолин.
По аналогии с пептидами типа II пептиды коллагена типа III могут существовать в гликозилированной форме их центральной структуры. К примеру, остатки галактозы или остатки глюкозилгалактозы могут быть присоединены к центральной структуре, например, через гидроксил.
Сшивающий остаток в пептидах коллагена типа III может быть представлен, как остаток 3-гидроксипиридиния (гидроксилизилпиридинолин). Структура телопептида типа II, как найдено, в первую очередь включает сшивающие остатки гидроксилизилпиридинолина. Однако, если пептиды коллагена типа II происходят из N-концевой телопептидной области коллагена типа II, пептиды коллагена типа III могут происходить как из N-конца, так и из C-конца коллагена типа III, при обязательном наличии по меньшей мере одного сшивающего остатка.
Коллаген типа III присутствует во многих соединительных тканях в ассоциации с коллагеном типа I. Он особенно сконцентрирован в стенках сосудов, в коже, и, например, в синовиальных мембранах суставов, где его ускоренное разрушение можно наблюдать в случае воспалительных заболеваний суставов, таких как ревматоидный артрит.
Для обнаружения, диагностирования и регистрации различных воспалительных нарушений, возможно, в особенности в случае васкулитного симптомокомплекса может быть использован специфический анализ разрушения коллагена типа III путем количественного определения вышеописанным способом сшитых пептидов коллагена типа III. В сочетании с анализами для определения степени разрушения костного коллагена типа I и хрящевого коллагена типа II подобный анализ может быть использован в качестве дифференцирующего диагностического инструмента для больных с различными вырожденными и воспалительными нарушениями, которые приводят к разрушениям или патологическим процессам в соединительных тканях.
Выделение телопептидов коллагена типа II и типа III
Общая методика.
Мочу отбирают у нормальных подростков в фазе быстрого роста скелета. С помощью последовательности хроматографических стадий, включающей, но без ограничения только перечисленным, адсорбцию на селективных патронах гидрофобного взаимодействующего носителя и ионообменного носителя, и на молекулярных ситах, стадии ионообменной и колоночной ВЭЖХ с обращением фаз хроматографии, выделяют индивидуальные пептиды. Сшитые пептиды, содержащие остатки ГП (и ЛП), детектируют в ходе колоночной хроматографии по их естественной флуоресценции (Ex max 297 нм, pH 4, Ex max 330 нм, > pH 6, Em max 390 нм). В нижеследующем примере дается конкретная методика выделения.
Характерный пример
Свежую мочу (при 4oC), разбавленную 5-кратно водой и с добавленной до концентрации 2% (об/об) трифторуксусной кислотой, пропускают через C-18 гидрофобные связующие патроны (Уотерс C-18 Сеп-пак, предварительно увлажненный 80% (об/об) ацетонитрилом и затем промытый водой) Удержанные пептиды промывают водой, после чего элюируют 3 мл 20% (об/об) ацетонитрила и полученный элюент до 0,05 М NH4HCO3, 10% (об/об) ацетонитрила добавлением равного объема 0,1 М NH4HCO3. Полученный раствор пропускают через ОМА-Сеп-пак (Уотерс), промытый 10 мл 0,1 М NaCl, 20% (об/об) ацетонитрила, затем 10 мл воды, после чего пептиды элюируют 3 мл 1% (об/об) трифторуксусной кислоты и сушат в приборе Спид-Вак (Савант).
Пептиды фракционируют на трех хроматографических стадиях. На первой стадии используется хроматография на молекулярных ситах с колонкой Био-Геля Р 10 (Био Рад Лаб, 2,5 см • 90 см) с элюированием 10% (об/об) уксусной кислотой и регистрацией эффлюента на ГП флуоресценцию, как показано на фиг. 2. На фиг. 2 на оси Y представлена относительная флуоресцентная эмиссия при 390 нм (возбуждение при 297 нм), а на оси X - номер фракции. Объем фракции 4 мл. Фракция обозначенная цифрой II, обогащена сшитыми телопептидами коллагена типа II. Сшитые телопептиды коллагена типа I содержатся во фракциях, обозначенных цифрами III и IV. Узкие фракции под цифрой II (обогащены сшитыми пептидами коллагена типа II) объединяют, сушат вымораживанием и фракционируют ионообменной хроматографией на ЕАЕ-ВЭЖХ колонке (TSL-DEAE-5PW, 7,5 мм • 7,5 мм, Био-Рад Лабс), приведенной в равновесие 0,02 М Трис-HCl, 10% (об/об) ацетонитрила (pH 7,5) с элюированием градиентом 0 - 0,5 М NaCl в том же буфере, как показано на фиг. 3.
На фиг. 3 А представлен график относительной флуоресцентной эмиссии при 390 нм (возбуждение при 330 нм) как функции времени элюирования. Сшитые телопептиды коллагена типа II в первую очередь обнаруживаются в сегменте, обозначенным цифрой IV.
На фиг. 3B представлен график поглощения при 220 нм как функции времени элюирования в минутах. Объединенная фракция IV содержит сшитые пептиды коллагена типа II. Индивидуальные пептиды затем разделяют в общей фракции IV с помощью ВЭЖХ с обращением фаз на C-18 колонке (Аквапор RP-300, 25 см • 4,6 с мм, Броунли Лабс) с элюированием 0 - 30% (об/об) ацетонитрила в 0,1% (об/об) трифторуксусной кислоте. На фиг. 4A представлен график относительной интенсивности флуоресценции при 390 нм (возбуждение при 297 нм) как функции времени элюирования. Пики, связанные с конкретными пептидами, указаны на фиг. 4A. На фиг. 4B представлено относительное поглощение при 220 нм как функция времени.
Фрагменты сшитых пептидов коллагена типа III, содержащие сшивающие ГП остатки, могут быть выделены применением аналогичной комбинации стадий из мочи нормальных растущих субъектов или, например, из мочи больных с воспалительными нарушениями сосудистой системы.
Телопептиды коллагена типа I
Данный аспект изобретения основан на открытии того, что как лизилпиридинолиновые (ЛП), так и гидроксилизилпиридинолиновые (ГП) пептидные фрагменты (т.е. телопептиды в применяемом здесь значении), происходящие из резорбированного костного коллагена, экскретируются в мочу без их метаболизма. Изобретение также основано и на том открытии, что никакие другие соединительные ткани не содержат в значительных количествах ЛП и что отношение ГП к ЛП в зрелом костном коллагене остается примерно постоянным на всем протяжении жизни субъекта.
На фиг. 5 приведено сравнение концентрации ГП и ЛП как в кортикальном слое кости, так и в губчатом веществе кости в зависимости от возраста. Замечено, что концентрация ГП плюс ЛП сшивок в костном коллагене достигает максимума в возрасте 10 - 15 лет и остается почти постоянным в течение всей взрослой жизни. Более того, отношение ГП к ЛП, приведенное на фиг. 6, мало изменяется в течение жизни, оставаясь постоянно на уровне 3,5 - 1. Такие основополагающие данные показывают, что сшивки 3-гидроксипиридиния в костном коллагене остаются сравнительно постоянными, вследствие чего биологические жидкости, происходящие при разрушении костного коллагена, будут содержать сшитые 3-гидроксипиридинием пептидные фрагменты в концентрации, пропорционаьной абсолютной степени костной резорбции.
Поскольку ЛП - это уникальная для костного коллагена 3-гидроксипиридиниевая поперечная связь (сшивка), способ определения абсолютной степени костной резорбции в его простейшей форме основан на количественном определении в биологической жидкости концентрации пептидных фрагментов, содержащих сшивки 3-гидроксипиридиния, предпочтительно сшивки лизилпиридинолина (ЛП).
В применяемом в данном описании и в прилагаемой формуле изобретения значении термином "количественное определение" в применении к телопептидам типа I, II и III обозначается определение любыми приемлимыми средствами, в том числе (но без ограничения только ими): спектрометрические, гравиметрические, объемометрические, кулонометрические, иммунометрические, потенциометрические или амперометрические средства определения концентрации пептидных фрагментов, содержащих сшивки 3-гидроксипиридиния, в аликвотном количестве биологической жидкости. Приемлемые биологические жидкости включают мочу, сыворотку и синовиальную жидкость. Рекомендуемой биологической жидкостью является моча.
Поскольку концентрация пептидов в моче будет уменьшаться по мере увеличения объема мочи, кроме того, рекомендуется при выборе в качестве биологической жидкости мочи аликвоты для анализа отбирать из общего количества мочи, собранной в фиксированный период времени, например за 24 часа. Таким путем подсчитывают абсолютную степень костной резорбции или разрушения коллагена за период в 24 часа. Или же пептиды в моче могут быть определены в виде отношения относительно вещества-маркера, присутствующего в моче, такого как креатинин. При этом показатель разрушения коллагена или костной резорбции по моче будет оставаться независимым от объема мочи.
В одном из воплощений изобретения продуцируют моноклональные или поликлональные антитела, специфичные к пептидным фрагментам, содержащие сшивки лизилпиридинолина и присутствующие в биологической жидкости, например моче. Фрагменты телопептида типа I могут быть выделены из биологической жидкости любого пациента, однако, рекомендуется, чтобы эти пептиды были выделены из биологической жидкости пациентов с болезнью Педжета или быстро растущих подростков вследствие высокой концентрации у них фрагментов пептида типа I. Телопептиды типа II и типа III могут быть выделены из биологической жидкости любого пациента, но более легко могут быть получены от больных, страдающих заболеваниями, включающими разрушения коллагена типа II или типа III, или от быстро растущих подростков.
Выделение телопептидов коллагена типа I.
Мочу пациентов с болезнью Педжета в активной форме диализуют 48 ч при 4oC в диализной трубочке с пониженной пористостью (Спектропор с предельной мол. массой < 3500) с удалением большей части растворимых веществ. В этих условиях представляющие интерес пептиды в основном задерживаются. Высушенную замораживанием непродиффудировавшую часть затем элюируют (аликвоты по 200 мг) из колонки (90 см • 2,5 см) Био-Геля Р2 (200 - 400 меш) в 10% уксусной кислоте и при комнатной температуре. Область эффлюента, объединяющую сшитые пептиды, определяют измерением флуоресценции собранных фракций при возбуждении в 297 нм/эмиссии в 395 нм и объединенную фракцию сушат вымораживанием. Дальнейшего разделения полученного продукта достигают на колонке Био-Геля Р-4 (200 - 400 меш, 90 см • 2,5 см) с элюированием 10% уксусной кислотой.
Регистрацией флуоресценции вышеполученного элюента определяют две следующие друг за другом общие фракции. Более ранняя фракция обогащена пептидными фрагментами, имеющими две аминокислотные последовательности, происходящими из C-концевого телопептидного домена aI(I) цепи костного коллагена типа I, соединенной с третьей последовательностью, происходящей из трехспиральной части костного коллагена типа I. Эти три пептидные последовательности сшиты 3-гидроксипиридинием. Перекрывающая более поздняя фракция обогащена пептидными фрагментами с аминокислотной последовательностью, происходящей из N-концевого телопептидного домена костного коллагена типа I, соединенной с участием 3-гидроксипиридиниевой сшивки.
Индивидуальные пептиды затем разделяют в каждой из двух вышеполученных фракций ионообменной ВЭЖХ на TSК DEAE-5-PW колонке (Био Рад, 7,5 см • 7,5 мм) с элюированием градиентом NaCl (0 - 0,2 М) в 0,02 М Трис-HCl (pH 7,5) содержащим 10% (об/об) ацетонитрила. Сшитые пептиды, основанные на N-концевом телопептиде и C-концевом телопептиде, элюируются в виде 3 - 4 пиков флуоресценции в интервале между 0,08 М и 0,15 М NaCl.
Сшитые пептиды, основанные на C-концевом телопептиде, элюируются первыми в виде ряда пиков флуоресценции, а основной и присутствующий в меньшем количестве сшитые пептиды, основанные на N-концевом телопептиде, элюируются к концу градиента в виде характеристических пиков. Каждый из них собирают, сушат вымораживанием и хроматографируют на C-18 колонке для ВЭЖХ с обращением фаз (Видак 218ТР54, 25 см • 4,6 мм) с элюированием градиентом (0 - 10%) ацетонитрил: н-пропанол 3: 1 об/об) в 0,01 М трифторуксусной кислоте. По этой методике из одного 24-часового сбора мочи больного болезнью Педжета можно выделить примерно 100 - 500 г индивидуальных пептидных фрагментов, содержащих сшивки 3-гидроксипиридиния.
С помощью полной стехиометрии выделенных аминокислот подтверждены чистота и молекулярные размеры аминокислотных составов основных выделенных пептидов. N-концевой секвенсционный анализ деградацией по Эдману подтвердили базовые центральные структуры, соответствующие последовательностям известных сшивающих сайтов коллагена типа I и в дополняющих аминокислотных составах. II - концевая телопептидная последовательность а2(1) цепи блокирована для секвенсционного анализа предположительно вследствие известной циклизации II - концевого глутамина в пирролидонкарбоновую кислоту.
Типичный характер элюирования N-концевых телопептидов, полученных по вышеприведенной методике, показан на фиг. 7A. Основной полученный пептидный фрагмент имеет аминокислотный состав формулы: (Asx)2(Glx)2(Gly)5Val-Tyr-Ser-Thr, где Asx - аминокислота Asp или Asn и Glx - аминокислота Gln или Clu. Последовательность этого пептида представлена нижеследующей формулой III.
Сшитые пептиды, основанные на C-концевом телопептиде, разделенные ВЭЖХ с обращением фаз по вышеприведенной методике, представлены на фиг. 7B. Как видно из фиг. 7B, эти пептиды в свою очередь разделяются на ряд C-концевых телопептидов, каждый из которых содержит 3-гидроксипиридиниевые сшивки. Основной пептид, показан на фиг. 7B, анализируют вышеприведенным способом и аминокислотный состав формулы (Asp)5(Glu)4(Gly)10(His)2- (Arg)2(Hyp)2(Ala)5. Последовательность этого пептида представлена нижеследующей формулой IV. Полагают, что другие сшитые пептиды, основанные на C-концевом телопептиде, представленные на фиг. 7B меньшими пиками, представляют собой вставки и делеции аминокислот в структуру формулы IV. Любой из пептидов этих меньших пиков может быть использован в качестве иммуногена в нижеприведенной методике.
Формула III
формула IV
Формула V
Формула VI
где
представляет ГП или ЛП сшивки и Gln представляет глутамин или пирролидонкарбоновую кислоту.
Эквиваленты пептидам, представленным вышеприведенными формулами, с точки зрения их присутствия в биологической жидкости вследствие разрушения коллагена, включают те случаи, когда в структуре пептида случаются определенные отклонения. Например, в моче человека в сравнительно небольших количествах обнаружен пептид, соответствующий формуле III, но имеющий остаток тирозина, присоединенный к аминоконцу II - концевого остатка аспартата. Особенно очевидно присутствие в моче меньших пептидных фрагментов молекулы, представленной формулой IV, происходящих в результате костной резорбции. Эти пептиды представлены меньшими пиками C-концевой телопептидной фракции на фиг. 7B и их можно идентифицировать по аминокислотному составу и секвенсционным анализом.
Примеры методик для количественного определения пептидов
A. Иммунологическая методика для количественного определения пептидов.
Партнеры в иммунологическом связывании, способные специфично связываться с пептидными фрагментами, происходящими из костного коллагена, и полученные из физиологической жидкости, могут быть приготовлены хорошо известными способами. Рекомендуемый способ выделения таких пептидных фрагментов приведен выше. Под иммунологическими партнерами в применяемом здесь значении имеются в виду антитела и фрагменты антител, способные связываться с телопептидом.
Как моноклональные, так и поликлональные антитела, специфично связывающиеся с охарактеризованными здесь пептидами и их эквивалентами, получают хорошо известными методами. См. например. Campbell A.M. Лабораторные методики в биохимии и молекулярной биологии т. 13 (1984). Эльзевир, которая вводится здесь в качестве ссылки. Можно создавать антитела к указанным пептидам и их фрагментам по их выделению. Однако поскольку молекулярная масса указанных пептидных фрагментов, как правило, меньше 5000, рекомендуется, чтобы гаптен был связан с молекулой носителя. Приемлемые молекулы носителя включают, но без ограничения только ими: альбумин бычьей сыворотки, авальбумин, тироглобулин и гемоцианин лимфы улитки (ГЛУ) К рекомендуемым носителям относятся тироглобулин и ГЛУ.
Хорошо известно, что ориентация гаптена при его связывании с белком-носителем имеет решающее значение для специфичности антисыворотки. Более того, не все гаптен-белковые конъюгаты являются равно эффективными иммуногенами. Выбор методики для связывания конкретного гаптена с белком-носителем, таким образом, зависит от аминокислотной последовательности выбранных пептидных фрагментов мочи. Например, при выборе пептида формулы III рекомендуемая методика заключается в связывании этого гаптена с гемоцианином лимфы улитки (ГЛУ) или иным приемлимым носителем и глутаральдегидом. Альтернативная методика заключается в связывании пептида с ГЛУ при участии карбодиимида. Данные методики позволяют гарантировать то, что в праймированных продуцирующих антитела клетках позвоночных (например, B лимфоциты) будут присутствовать предпочтительные эпитопы (обсуждаемые ниже в разделе под заглавием "Характеристика предпочтительного эпитопа").
Другие пептиды в зависимости от их источника могут потребовать других методик. Соответственно в зависимости от обстоятельств может быть использован целый ряд связующих средств, в том числе (но без ограничения только ими): карбодиимиды, глутаральдегид, смешанные ангидриды, а также как гомобифункциональные, так и гетеробифункциональные реагенты (см. например, каталог Пирса за 1986-87 гг, Пирс Кемикал Ко, Рокфорд, шт. ИЛ). Рекомендуемые связующие средства включают карбодиимиды и гетеробифункциональные реагенты, такие как м-малеоимидобензил-N-гидроксисукцинимидовый эфир (МБС).
Методы связывания гаптена с носителем известны. См., например Charo T. Лабораторные методики в биохимии и молекулярной биологии т. 6 (1986) Парц Эльзевир, шт. Н-Й, которая вводится здесь в качестве ссылки.
Могут быть созданы как моноклональные, так и поликлональные антитела к иммуногенной молекуле гаптен-носитель. Однако рекомендуется получение моноклональных антител (МАо). По той же причине рекомендуется, чтобы иммунизация осуществлялась на мышах. Методика иммунизации мышей обычно включает участие адъюванта. Примеры приемлимых методик приведены у Charo T. (1987) см. выше. Клетки селезенки иммунизированной мыши собирают и гомогенизируют, после чего сливают с раковыми клетками в присутствии полиэтиленгликоля с получением слитого клеточного гибрида, продуцирующего моноклональные антитела, специфичные к пептидным фрагментам, происходящим из коллагена. Примеры подобных пептидов, представлены вышеприведенными формулами. Приемлимые раковые клетки включают клетки миеломы, гепатомы, карциномы и саркомы. Подробное описание такой методики, включая способ отбора, способ выращивания отобранных гибридных клеток и сбора моноклональных антител, продуцированных отобранными гибридными клетками, приведена в работе Galfre C. Milstein C. Meth. Enzymol. 73 1 (1981). Рекомендуемая методика первичного отбора включает применение пептидных фрагментов, возникающих при резорбции костного коллагена и содержащих сшивки 3-гидроксипиридиния, в твердофазном радиоиммуноанализе. Ниже приводится характерный пример, описывающий рекомендуемые антитела.
Для создания моноклональных антител могут быть также применены биотехнологические методы. Подобные методы могут быть осуществлены обычным специалистом путем создания библиотеки экспрессии и праймеров (см. William D. Huse и др, "Создание большой комбинационной библиотеки иммуноглобулинового репертуара в лямбда-фаге" Science 246 1275 - 1281, декабрь 1989 г, см. также L. Sastry и др. "Клонирование иммунологического репертуара в Escheri chia coli для образования моноклональных каталитических антител. Создание библиотеки сДНК, специфичной к тяжелоцепным переменным областям, Ppoc. Nall. Acad. Sci. США, 5728 - 5732 август 1989, г, см. также Michelle Altinq-Mees и др. "Библиотеки экспрессии моноклональных антител. Быстрая альтернатива гибридомам", Strafegiesin Molekular Biology, 3, 1 - 9, январь 1990 г.) или использованием продажных наборов, предназначенных для этой цели (например, набор фирмы Стратацит, Ла Джолла, Калифорния. Вкратце в объеме данного воплощения изобретения, из популяции B клеток выделяют мРНК и используют для создания библиотеки экспрессии к ДНК тяжело- и легкоцепного иммуноглобулина в векторах Иммуно Zap(H) и Иммуно Zap(L). Такие векторы могут быть отобраны индивидуально или подвергнуты совместной экспрессии с образованием фрагментов Fab или антител (Huse и др, см. выше, а также Sastry и др. см. выше). Положительные бляки могут быть затем превращены в нелитическую плазмиду, позволяющую экспрессировать в E. coli на высоком уровне фрагменты моноклональных антител.
Моноклональные антитела или другие иммунологические связующие партнеры, применяемые в связи с настоящим изобретением, предпочтительно специфичны к конкретному типу коллагенового телопептида. Например, анализ на телопептида разрушенного коллагена типа II или типа III предпочтительно должен позволять различать пептиды типа I, типа II и типа III. Однако в некоторых случаях в такой селективности нет необходимости, например, если известно, что больной не страдает от разрушения коллагена одного типа, но его страдания, как подозревают, вызваны разрушением коллагена анализируемого типа. Вследствие отличий в аминокислотных последовательностях семейств телопептидов типа I, типа II и типа III значительного пересечения реакционноспособностей не может произойти. В самом деле, в ходе отбора спленоцитных слитых клонов могут быть подобраны такие гибридомы, которые продуцируют моноклональные антитела, специфичные к представляющим интерес сшитым телопептидам (но без сродства к коллагенам двух других типов). Основываясь на различных в последовательностях выделенных пептидных структур, такая специфичность вполне осуществима. Пептидные фрагменты коллагенов родственных типов I, II и III, приемлемые для такого отбора гибридомы, могут быть получены из костей, хрящей и других человеческих тканей и использованы для подбора клонов от мышей, соответствующим образом иммунизированных антигеном к индивидуальному сшитому пептиду, выделенному из биологической жидкости.
Иммунологические связующие партнеры, в особенности моноклональные антитела, полученные по вышеприведенной методике или эквивалентными методами, применяют в различных иммунометрических анализах для количественного определения концентрации вышеохарактеризованных пептидов, имеющих сшивки 3-гидроксипиридиния. В таких иммунометрических анализах рекомендуется использовать моноклональные антитела или фрагмент антител, соединенные с подающимся обнаружению маркером. Примеры приемлимых поддающихся обнаружению маркеров включают, но без ограничения только ими: ферменты, коферменты, ингибиторы ферментов, хромофоры, флуорофоры, хемилюминесцентные вещества, парамагнитные металлы, спиновые метки и радионуклеиды. Примеры стандартных иммунометрических методов анализа, приемлимые для количественного определения телопептидов, включают, но без ограничения только ими: ферментный иммуносорбентный анализ (ФИСА) (Ingvall E. Meth. Enzymol. 70 (1981), радиоиммунный анализ (РИА) и "сэндвичевый иммунорадиометрический анализ (ИРМА) и "сэндвичевый" иммунорадиометрический анализ (ИРМА).
В упрощенном виде указанные иммунометрические методы анализа могут быть использованы для определения абсолютной степени костной резорбции или разрушения коллагена путем простого контактирования биологической жидкости с иммунологическим связующим партнером, специфичным к коллагеновому телопептиду, имеющему сшивку 3-гидроксипиридиния.
Рекомендуется, чтобы вышеуказанные иммунометрические анализы проводились непосредственно на необработанных биологических жидкостях (например, моче, крови, сыворотке или синовиальной жидкости). Однако в тех случаях, когда загрязняющие вещества могут повлиять на результаты анализа, возникает необходимость в частичной очистке биологической жидкости. Методы частичной очистки включают, но без ограничения только ими: адсорбцию и элюирование в патронах, хроматографию на молекулярных ситах, диализ, ионный обмен, хроматографию на окиси алюминия, хроматографию на гидроксиапатите и сочетание указанных методов.
Тест-наборы, пригодные для использования в соответствии с настоящим изобретением, содержат специфичные связующие партнеры, такие как моноклональные антитела, полученные по вышеприведенным методикам и которые специфично связываются с пептидными фрагментами, возникающими при разрушении коллагена и обнаруживаются в биологической жидкости. Рекомендуется, чтобы специфичные связующие партнеры в тест-наборе были связаны с поддающимся обнаружению маркером вышеуказанного типа. Также приемлимы наборы, содержащие панель из двух или более специфичных связующих партнеров, в частности иммунологических связующих партнеров. Каждый иммунологический связующий партнер в таком тест-наборе по своей реакционноспособности предпочтительно существенно не пересекается с телопептидами, происходящими из коллагена другого типа. Например, иммунологический связующий партнер, специфично связывающийся с телопептидом коллагена типа II, предпочтительно не должен вступать в перекрестную реакцию с телопептидами как коллагена типа I, так и коллагена типа II. Небольшая степень (напр. 5 - 10%) перекрестной реакционноспособности допустима. Другие тест-наборы могут включать первый специфичный связующий партнер к происходящему из коллагена телопептиду, имеющему сшивку с пиридиниевым циклом (который может быть OH-замещен), и второй специфичный связующий партнер к телопептиду, обладающему тем же строением, что и первый телопептид, за тем исключением, что пиридиниевый цикл расщеплен, например, фотолитически.
(I) Продуцирование моноклональных антител
Ниже приводится пример получения моноклональных антител к пептидному иммуногену, основанному на вышеприведенной формуле III.
Фракцию, обогащенную пептидом формулы III (свидетельство разрушенного костного коллагена), получают из мочи подростка с помощью хроматографии с обращением фаз и хроматографии на молекулярных ситах. Пептид соединяют по стандартной методике с гемоцианином лимфы улитки (ГЛУ) в присутствии глутаральдегида. Мышей (Balb/c) иммунизируют подкожно полученным конъюгатом (50 - 70 г) сначала в полном адъюванте Фройнда, затем активируют (25 г) с интервалом в 3 недели в неполном адъюванте Фройнда внутрибрюшинно. После того, как анализ крови показал высокий титр к пептиду формулы III (далее обозначаемый как P1), соединенный с альбумином бычьей сыворотки (АБС), на основании ФИСА, мышей ревакцинируют внутривенно низкой дозой (5 г) иммуногена в стерильном P1. Спустя три дня использованием стандартной гибридомной технологии клетки селезенки отдельной мыши сливают с клетками мышиной миеломы. Надосадочные жидкости гибридомных клонов, выращенных в отдельных ячейках 96-ячеистого планшета, отбирают на наличие реакционноспособных моноклональных антител, первоначально используя препарат сырого P1, присоединенного к АБС. После формального клонирования путем ограничения разбавления антитела, продуцирование индивидуальными гибридомами, характеризуют использованием ФИСА относительно панели отборочных антигенов. Этими антигенами являются пептиды P1 (формула III) и P2 (формула VII), связанные с АБС. Используют ингибирующее испытание, в котором P1, связанный с АБС, наносят на поверхность пластиковых ячеек, и антитела предварительно инкубируют с раствором потенциального антигена. Для цветового проявления применением соответствующего субстрата применяют вторичные антитела (козий антимышинный ИгГ, связанный с пероксидазой из хрена (ПОХ)). Идентифицируют целевые моноклональные антитела с высоким связующим сродством к пептиду P1. При использовании в виде препарата асцитной жидкости антитела работают в ингибирующем испытании с оптимальным выходом по цвету при разбавлении в 2 миллиона раз (что указывает на константу связывания в интервале 10-9 - 10-11 М-1, наиболее вероятно примерно 10-10 М-1). При использовании ФИСА антитела способны обнаруживать и определять P1, присутствующий в моче нормального человека, без каких-либо стадий концентрирования или очистки. Гибридома, продуцирующая рекомендуемые моноклональные антитела, депонированы в Американской коллекции типовых культур (АТСС), 12301 Парклоун Драйв, Росвиль, Мерилэнд 20852 под регистрационным номером НВ 10611. Далее эту гибридому обозначают как 1Р11, а продуцируемые ею моноклональные антитела далее обозначают как MAb-1Н11.
Показано также, что сэндвичевые анализы работают с применением специфичных к P1 моноклональных антител и поликлональной антисыворотки, созданной у кроликов к связанному P1. Для специфичного связывания с P1 из мочи с последующей возможностью обнаружения с помощью стандартного ФИСА или иного иммуноанализа могут быть использованы специфичные к P1 моноклональные антитела, поликлональная сыворотка, их связующие фрагменты и т.п.
(II) Характеристика предпочтительного эпитопа
В структуру P1 включен эпитоп, распознаваемый антителами MAb-1Н11. Эпитоп распознается в чистом P1 и некоторых больших пептидах, содержащих структуры P1 (напр. P1, присоединенный к остатку тирозина через II-концевой остаток аспарата P1). Эпитоп включает химические признаки каждой из двух телопептидных последовательностей, заключенных в структуре пептида P1. Пептиды, синтезированные для соответствия а1 (I) и а2 (I) N-концевым телопептидным последовательностям человека с добавлением C-концевого цистеина для присоединения к альбумину бычьей сыворотки (т.е. YDEKSTGGC и QYDGKGVGC) не распознаются MAb-1H11. Это показано с помощью ФИСА и использованием свободных пептидов, конкурентных с нанесенным на поверхность P1 (см. фиг. 9), или непосредственно в качестве связующих партнеров, связанных с АБС и нанесенных на поверхность. Что касается фиг. 9, то поглощение при g = 450 нм поддающегося обнаружению маркера отложено на графике относительно концентрации свободного пептида P1. По мере увеличения количества свободного P1 количество поддающегося обнаружению маркера, связанного с иммунобилизованным (нанесенным на поверхность) P1, уменьшается. Для сравнения а2(I) и а1(I) N-телопептиды демонстрируют незначительное, если вообще демонстрируют, конкурентное связывание с MAb-1Н11.
Кроме того, большие формы P1, несущие остаток тирозина у N-концевой аспаратиновой кислоты, выделены из мочи аффинным связыванием с MAb-1Н11, но с меньшим выходом, чем P1. В еще меньших количествах выделены также другие несколько более крупные пептиды, несущие эпитоп P1.
Антитела не отличаются селективностью по отношению к природе сшивки в P1, т.е. не различают гидроксилизилпиридинолин (ГП) и лизилпиридинолин (ЛП). Связываются как ГП-содержащие, так и ЛП-содержащие формы, вероятно с одинаковым сродством, о чем можно судить на основании анализа пептидов, выделенных из мочи на аффинной колонке, содержащей MAb-1Н11 на агарозе. Свободные сшивающие аминокислоты (ГП и ЛП), полученные либо кислотным гидролизом костного коллагена, либо присутствующие естественным путем в моче, не распознаются MAb-1Н11.
После фотолитического раскрытия в пептиде P1 цикла 3-пиридинола под действием УФ-облучения (длиноволновый УФ) специфичное связывание с антителами также не подтверждено влиянию, предположительно вследствие того, что индивидуальные пептиды остаются сшитыми друг с другом. Опыт проводят следующим образом
В условиях, в которых, как показано, полностью разрушаются ГП и ЛП (о чем свидетельствует исчезновение флуоресценции характеристических флуоресцентных пиков при ВЭЖХ ОФ), как в виде свободных аминокислот, так и нерастворимых пептидов и интактных белковых цепей, препарат P1 облучают УФ (длиноволновая установка, лампа Минралайт UVSL-25, Ультра-Вайолет Продактс Инк, Сан Габриэль, Калифорния).
Полученный раствор анализируют на связывание с MAb-1Н11 с применением контрольного раствора точного того же продукта, но необлученного. Результаты ФИСА с 1Н11 показали по существу отсутствие потерь связывания с P1, что предполагает отсутствие существенного влияния расщепления цикла на эпитоп. В условиях УФ облучения (pH 9) следует ожидать расщепление единственной связи с раскрытием цикла, а не двойное расщепление с удалением циклического азота. См. например, Eyre D. Met. Enzymol, 144 139 - 155 (1987)
Эпитопу, распознаваемому MAb-1Н11, таким образом, присуще по меньшей мере сочетание химических и конформационных признаков, заключенных в двух телопептидных последовательностях, обведенных на фиг. 10 рамкой, вместе со стерическими признаками, обусловленными трехвалентной сшивающей аминокислотой, которая их соединяет. Особенно важной частью эпитопа является а2(I) N-телопептидная последовательность (ОУ СК).
Тот факт, что эпитоп, распознаваемый MAb-1Н11, не зависит от интактного цикла пиридиния. является неожиданным открытием. Если раскрытие цикла происходит in vivo или даже in vitro в рутинных условиях обработки, что видимо вероятно, тогда количественный анализ целевого пептида -(ов) с интактными циклами пиридиния приведет к недооценке степени костной резорбции. Предварительные наблюдения указывают на то, что разрушение циклов пиридиния в целевых пептидах видимо происходит в моче, в частности, и/или образцах мочи, даже при хранении в холодильнике. Соответственно можно ожидать, что анализ, основанный на данном описании, будет более точным. Рассматриваются два варианта: с применением единственного специфичного связующего партнера, распознающего целевой пептид -(ы) как с замкнутым, так и раскрытым циклом, или с применением двух специфичных связующих партнеров, дифференцировано распознающих эпитопы с замкнутым или раскрытым циклом соответственно.
Специфически связующие партнеры, различающие целевые пептиды с замкнутым или раскрытым циклом, могут быть получены введением соответствующей стадии отбора в стандартные методики получения таких специфически связующих партнеров. Например, для получения моноклональных антител, специфически связывающихся с пептидом P1 с раскрытым циклом, может быть отобрана библиотека кандидатов в моноклональные антитела по их способности связываться с P1, имеющим открытый пиридинолиновый цикл (например, под действием ультрафиолетового облучения), и неспособности связываться с P1, имеющим интактный пиридинолиновый цикл. Пептиды с открытыми и интактными циклами, применимые на такой стадии отбора, могут быть получены известными способами очистки, такими как ВЭЖХ ОФ, описанная выше.
Дальнейшие опыты показали, что эпитоп наблюдается и в костном коллагене человека, однако проявляет себя и связывается с MAb-1Н11 только после интенсивного протеолиза. Так пептиды, полученные из декальцинированного костного коллагена человека действием бактериальной коллагеназы, связываются с MAb-1Н11 и, как показано, происходят из N-телопептидного до спирального сайта, показанного на фиг. 10. Одна форма содержит гексапептид СIКСНР (вместо нетелопептидного K-луча в P1), явно происходящей из а1(I) остатков 928 - 933. Другая форма включает эквивалентный, но отличный гексапептид, происходящий из а2(I) цепи. Фрагменты костного коллагена человека, солюбилизованные пепсином CNBr или трипсином, не распознаются MAb-1Н11, как с помощью ФИСА при их использовании в качестве конкурентных ингибиторов, так Вестерн блотированием, после электрофореза на НДС-полиакриламиде с указанием на то, что указанными солюбилизующими средствами не продуцируется эпитоп, распознаваемый MAb-1Н11.
B. Методика электрохимического анализа пептидов
Альтернативная методика анализа вышеохарактеризовнаных пептидов состоит в определении физического показателя пептидов, имеющих сшивки 3-гидроксипиридиния. Один из подобных физических показателей основывается на электрохимическом детектировании. Метод заключается в инъектировании аликвоты биологической жидкости, например мочи, в электрохимический детектор, основанный на окислительно-восстановительном потенциале, приемлемом для детектирования пептидов, содержащих цикл 3-гидроксипиридиния. Цикл 3-гидроксипиридиния - фенольный по своему характеру способен к обратимому окислению, поэтому электрохимический детектор (например, Модель 5100А Кулохем фирмы Esa 45 Виггинс Ав, Бедфорд МА) представляется в высшей степени желательным инструментом, пригодным для количественного определения концентрации пептидов настоящего изобретения. Базовые модели электрохимического детектора в настоящее время выпускаются промышленностью: амперометрический (например, фирмы Био-Аналитикал-Системс) и кулонометрический (фирмы ESa Инк, Ведфорд, МА 01730). Оба пригодны для применения в соответствии с настоящим изобретением, однако последняя система изначально более чувствительна, поэтому и рекомендуется, поскольку достигается полное окисление или восстановление анализируемой молекулы в колоночном эффлюенте. Кроме того, отсеивающие или охранные электроды могут быть помещены в восходящем направлении от аналитического электрода с селективным окислением или восстановлением мешающих веществ со значительным улучшением в результате селективности. По существу напряжение аналитического электрода на окислительновосстановительный потенциал анализируемой молекулы, а для разрушения мешающих веществ устанавливают одну или несколько ячеек предварительной обработки.
В рекомендуемом методе анализа строят стандартную кривую ток/напряжение для стандартных пептидов, содержащих лизилпиридинолин или гидроксилизилпиридинолин, с целью определения напряжения, необходимого для достижения оптимальной чувствительности. Специалистам известны и электрохимические детекторы, и оптимальные условия их применения. С помощью электрохимического детектора сложные смеси биологических жидкостей часто могут быть без помех подвергнуты непосредственному анализу. Соответственно для большинства пациентов нет никакой необходимости в предварительной обработке биологической жидкости. Однако в некоторых случаях мешающие соединения могут снизить надежность определения. В таких случаях может потребоваться предварительная обработка биологической жидкости (например, мочи).
Соответственно в альтернативном воплощении изобретения сначала очищают перед электрохимическим титрованием очищенных пептидных фрагментов. Стадия очистки может быть осуществлена самыми различными способами, в том числе, но без ограничения только ими: диализом, ионообменной хроматографией, хроматографией на окиси алюминия, хроматографией на гидроксиапатите, хроматографией на молекулярных ситах или сочетанием перечисленных способов. В рекомендуемой методике очистки отмеренную аликвоту (25 мл) 24-часового образца мочи диализуют в диализной трубке с пониженной пористостью с удалением основной массы растворимых люминесцентных примесей. Недиффундировавшую часть затем лиофилизуют, вновь растворяют в 1% гептафтормасляной кислоте (ГФМК) растворе ионной пары и пептиды адсорбируют на патроне Сеп-Пак C-18 фирмы Уотерс. Затем патроны промывают 5 мл 1% ГФМК, после чего элюируют 3 мл 50% метанола в 1% ГФМК
Другой рекомендуемый метод очистки заключается в адсорбировании отмеренной аликвоты мочи на ионообменном адсорбционном фильтре и элюировании адсорбционного фильтра буферным элюирующим раствором. Фракции элюата, содержащие пептидные фрагменты с 3-гидроксипиридиевыми сшивками затем собирают для анализа.
В еще одном рекомендуемом методе очистки применяют хроматографию на молекулярных ситах. Например, аликвоту мочи наносят на колонку с Био-Гелем P2 или Сефадексом C-20 и собирают фракции, элюирующиеся в интервале 1000 - 5000 Дальтон. Для специалиста очевидно, что для очистки или частичной очистки мочи или иной биологической жидкости с целью выделения пептидных фрагментов, содержащих 3-гидроксипиридиниевые сшивки, может быть использовано сочетание вышеприведенных методов. Очищенные или частично очищенные пептидные фрагменты могут быть подвергнуты дополнительной очистке, дополнительно обработаны или могут быть проанализированы непосредственно в частично очищенном виде. Дополнительная очистка, если желательна повышенная чувствительность, включает разделение частично очищенных пептидных фрагментов с применением высокоэффективной жидкостной хроматографии (ВЭЖХ) или микроканальной ВЭЖХ. Такие пептиды затем могут быть определены количественно электрохимическим титрованием.
Рекомендуемая методика электрохимического титрования состоит в настройке окислительно-восстановительного потенциала детектирующей ячейки электрохимического детектора (Кулохем, Модели 5100А) на максимальный сигнал для чистого ЛП. Затем детектор используют для регистрации эффлюента с C-18 колонки ВЭЖХ, применяемой для разделения частично очищенных пептидов.
C. Методика флуорометрического количественного определения пептидов
Альтернативный рекомендуемый метод количественного определения концентрации вышеохарактеризованных пептидов, имеющих сшивки 3-гидроксипиридиния, заключается в определении характеристической естественной флуоресценции этих пептидов. Для тех биологических жидкостей, которые содержат немного природных флуоресцентных веществ, отличных от 3-гидроксипиридиниевых сшивок, флуорометрический анализ может быть осуществлен непосредственно на биологической жидкости без ее дополнительной очистки. В этом случае пептиды разделяют с помощью ВЭЖХ и естественную флуоресценцию ГП и ЛП аминокислотных остатков определяют при 395 нм и возбуждении при 297 нм по существу так, как описано в работе Eyre D.P. и др. Analyte. Biochem. 137 (1984), которая вводится здесь в качестве ссылки.
Согласно настоящему изобретению рекомендуется, чтобы флуорметрический анализ проводился на моче. Однако моча обычно содержит значительные количества флуоресцентных примесей естественного происхождения, которые следует удалить перед проведением флуоресцентного анализа. Соответственно образцы мочи вначале частично очищают по методике, приведенной выше для электрохимического детектирования. Такой частично очищенный образец мочи затем может быть подвергнут флуорометрическому анализу. Или же ГП и ЛП сшитые пептиды в частично очищенных образцах мочи или иной биологической жидкости могут быть гидролизованы в 6 М HCl примерно при 108oC в течение 24 часов по методике Eyre и др. (1984), см. выше. В результате гидролиза аминокислоты, соединенные с лизиновыми предшественниками "трехпептидных" ГП и ЛП сшивок, образуют свободные ГП или ЛП аминокислоты формул I и II. Такие небольшие трипептиды затем разделяют вышеописанными способами, предпочтительно ВЭЖХ с последующим определением естественной флуоресценции (Ex 297 нм, E 390 нм).
Возможно биологическую жидкость (предпочтительно мочу) пропускают непосредственно через C-18 аффинный патрон с обращением фаз после добавления ацетонитрил-метанола (5 - 10% об/об) Неудержанную часть доводят до 0,05 - 0,10 М катионного ионопарного средства, такого как тетрабутиламмонийгидроксид, и пропускают через второй C-18 патрон с обращением фаз. Промытую удержанную часть, содержащую флуоресцентные пептиды, из второго патрона элюируют смесью ацетонитрил-вода (или метанол-вода), сушат и флуоресцентные пептиды анализируют с помощью ВЭЖХ с обращением фаз анализируют с помощью ВЭЖХ с обращением фаз или микроканальной ВЭЖХ с применением в элюенте анионного ионопарного средства, такого как 0,01 М трифторуксусная кислота.
На фиг. 8A показан характер элюирования разделенных с помощью ВЭЖХ пептидных фрагментов в гидролизате мочи нормального человека и их естественная флуоресценция. Определение интегрированной области в пределах конверта данного компонента применяют для определения концентрации этого компонента. Отношение ГП: ЛП, обнаруживаемое в моче нормального человека и моче больных болезнью Педжета, согласно фиг. 8B, в обоих случаях примерно 4,5:1. Это чуть выше, чем отношение 4: 1, обнаруживаемое в самой кости (Eyre и др. 1984). Более высокие отношения, обнаруживаемые в моче, указывает на то, что часть ГП фракции в моче может происходить отлично от костного источника, например пищи или других источников разрушения коллагена, например в результате катаболизма хряща. Именно по этой причине рекомендуется, чтобы ЛП, происходящий исключительно из костей, применялся в качестве абсолютного показателя костной резорбции. Однако при отсутствии избыточного разрушения, например при ревматоидном артрите или в случае быстрого абсорбирования кости, в качестве показателя костной резорбции можно использовать ГП или комбинацию ГП плюс ЛП.
В то время как изобретение описано со ссылкой на его рекомендуемые воплощения, рядовой специалист после прочтения вышеприведенного описания сможет внести различные изменения, замены и сокращения в объект изобретения. Т. е. изобретение на практике может быть осуществлено путями, отличными от специально приведенных здесь. Подразумевается, что защита изобретения, гарантируемая патентным законодательством, будет ограничена только прилагаемой формулой изобретения и ее эквивалентами.




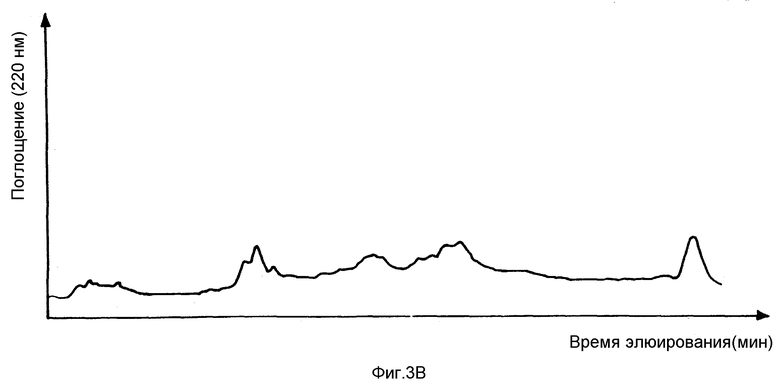



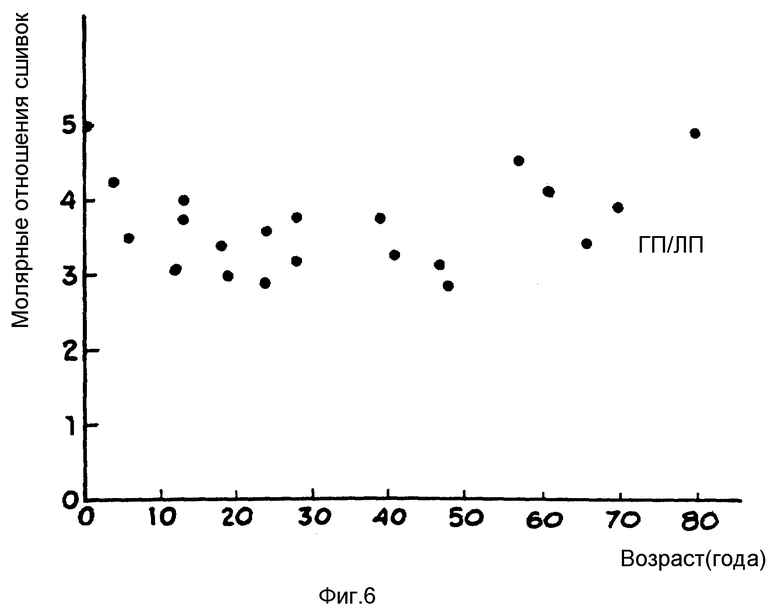
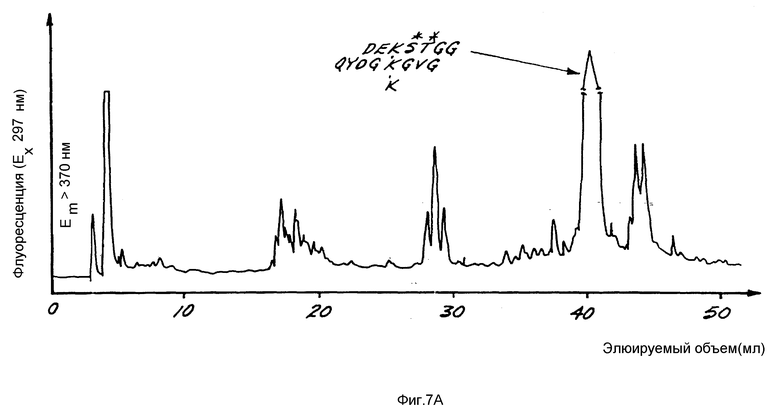


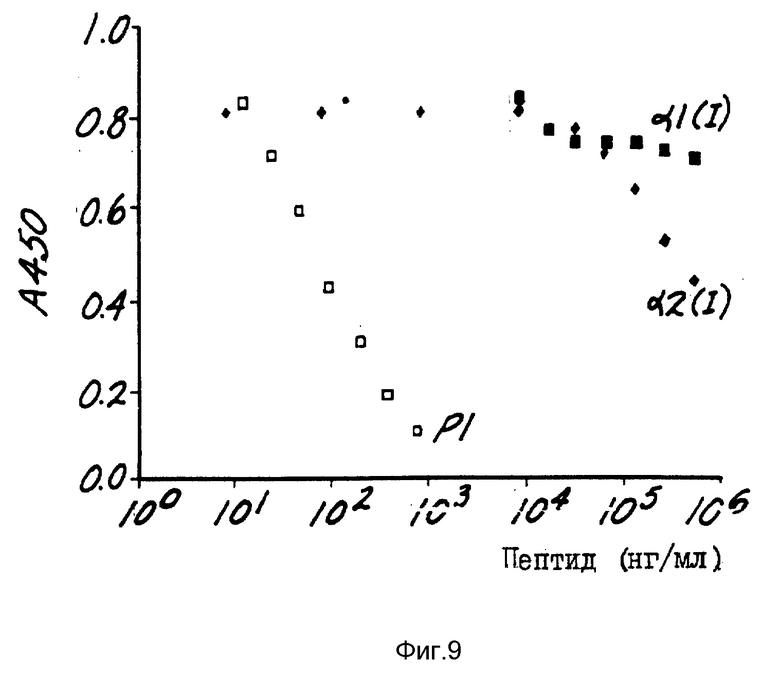

Изобретение относится к биотехнологии и медицине и может быть использовано для определения разрушения коллагена in vivo. Определение и регистрацию разрушения коллагена осуществляют путем контактирования иммунологически связывающих партнеров к сшитым телопептидам, происходящим из коллагена типа I, II, III и содержащим раскрытое или нераскрытое пиридиниевое кольцо 3-гидроксипиридиниевой сшивки с биологической жидкостью. Сшивка представляет собой гидроксилизилпиридинолин или лизилпиридинолин. Изобретение позволяет проводить диагностику заболеваний, связанных с разрушением коллагена. 15 с. и 14 з.п.ф-лы, 14 ил.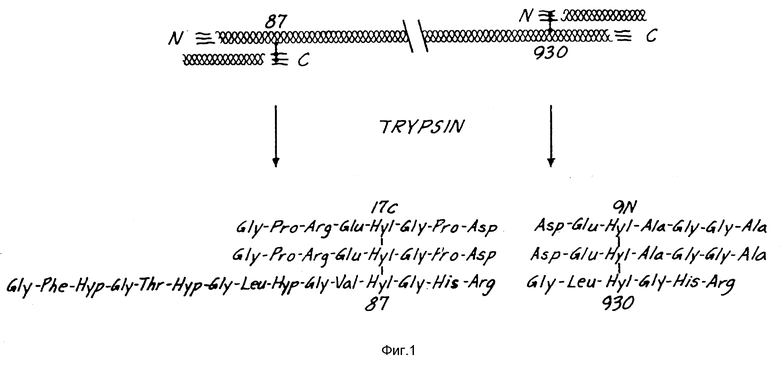

где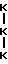
представляет собой 3-гидроксипиридиниевую сшивку, выбранную из гидроксилизилпиридинолина и лизилпиридинолина;
Gln является глутамином или пирролидинкарбоновой кислотой,
отличающийся тем, что биологическая жидкость либо контактирует с первым иммунологическим связывающим партнером, специфическим к первому пептиду и ко второму пептиду, имеющему приведенную выше формулу с тем исключением, что пиридиниевое кольцо 3-гидроксипиридиниевой сшивки было раскрыто, или биологическая жидкость контактирует как со вторым иммунологическим связывающим партнером, специфическим к первому пептиду, так и с третьим иммунологическим связывающим партнером, специфическим ко второму пептиду.
где
представляет 4-гидроксипиридиниевую сшивку, выбранную из гидроксилизилпиридинолина и лизилпиридинолина;
Gln представляет глутамин или пирролидинкарбоновую кислоту,
и для второго пептида, имеющего ту же структуру за исключением того, что пиридиниевое кольцо 3-гидроксипиридиниевой сшивки раскрыто.
где
представляет гидроксилизилпиридинолин, причем первый и второй пептиды содержат нераскрытое и раскрытое пиридиниевое кольцо соответственно.

или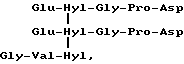
где
представляет собой гидроксилизилпиридинолин, причем указанные первый и второй пептиды содержат нераскрытое или раскрытое кольцо пиридиния соответственно.
или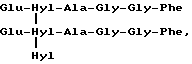
отличающийся тем, что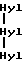
представляет гидроксилизилпиридинолин, причем первый и второй пептиды содержат нерасщепленное или расщепленное кольцо соответственно.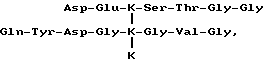
где
представляет 3-гидроксипиридиниевую сшивку, выбранную из гидроксилизилпиридинолина и лизилпиридинолина;
Gln представляет глутамин или пирролидинкарбоновую кислоту,
причем пиридиниевое кольцо 3-гидроксипиридиниевой сшивки в первом и втором пептидах не раскрыто и раскрыто соответственно.
где
представляет гидроксилизилпиридинолин.

или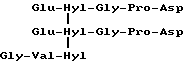
где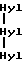
представляет собой гидроксилизилпиридинолин.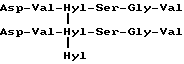
или
где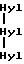
представляет гидроксилизилпиридинолин.
где
представляет гидроксилизилпиридинолин.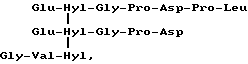
где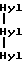
представляет гидроксилизилпиридинолин.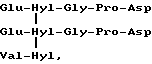
где
представляет гидроксилизилпиридинолин.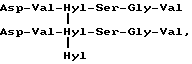
где
представляет гидроксилизилпиридинолин.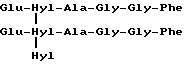
где
представляет гидроксилизилпиридинолин.
где
представляет лизилпиридинолин или гидроксилизилпиридинолин.
где
представляет 3-гидроксипиридиниевую сшивку, выбранную из лизилпиридинолина и гидроксилизилпиридинолина, где пиридиниевое кольцо 3-гидроксипиридиниевой сшивки раскрыто либо не раскрыто.
Glu-K-Ala-His-Asp-Gly-Gly-Arg,
где К представляет собой часть 3-гидроксипиридиниевой сшивки, выбранной из гидроксилизилпиридинолина и лизилпиридинолина, с нераскрытым или раскрытым пиридиниевым кольцом.
Приоритет по пунктам:
21.11.90 по пп.1-8, 10-12, 26, 28, 29;
01.12.89 по пп.9, 13-25, 27.
| Способ размножения копий рисунков, текста и т.п. | 1921 |
|
SU89A1 |
| Шланговое соединение | 0 |
|
SU88A1 |

