Изобретение относится к химерным регуляторным участкам, применяемым для контроля экспрессии генов в растениях. Эти химерные регуляторные участки выделены из генов опин-синтазы патогена растений Agrobacterium tumefaciens.
A. tumefaciens - грамотрицательная почвенная бактерия, которая инфицирует большинство двудольных и ряд однодольных растений. Инфекция A. tumefaciens часто приводит к образованию корончатых галлов на инфицированных растениях. В процессе инфекции A. tumefaciens определенный сегмент ДНК (Т-ДНК) большой плазмиды, индуцирующей опухоли (Ti), переносится в клетку чувствительного растения и интегрируются в ядерный геном растения. При этом происходит экспрессия генов Т-ДНК. Некоторые гены Т-ДНК кодируют ферменты, участвующие в синтезе гормонов, активных в растениях. Эти гормоны могут вызывать образование опухоли у инфицированных растений. Другие гены Т-ДНК направляют синтез и секрецию уникальных производных аминокислот и сахаров, называемых опинами. A. tumefaciens может использовать эти опины в качестве источников углерода и (иногда) азота. См. Gelvin, Plant Physiol. 92: 281-85 (1990); Gelvin, Transgenic Plants (Academic Press 1993); Ream, Ann. Rev. Phytopathol. 27: 583-618 (1989; Zambryski, Ann. Rev. Plant Physiol. Plant Mol. Biol. 43: 465-90 (1992).
Гены Т-ДНК содержат участки, работающие в растениях и обладающие сходством с регуляторными участками растений. Например, большинство промоторов растений содержат цис-действующие элементы, такие как расположенные против хода транскрипции активирующие последовательности (UAS) (часто называемые энхансерами), которые, связывая транс-действующие факторы, определяют или влияют на силу промотора и тканеспецифичную экспрессию. Atchison, Annu. Rev. Cell Biol. 4:127-53 (1988). Общая сила промотора, а также экспрессия соответствующего ему гена может определяться комбинацией и пространственной ориентацией цис-действующих элементов, а также наличием ядерных факторов, взаимодействующих с этими элементами. Dynan, Cell 58:1-4 (1989). Изначально находясь на прокариотной плазмиде, гены Т-ДНК сохраняют все элементы последовательности (промоторы и UAS), необходимые для транскрипции в растениях. Например, гены Т-ДНК содержат области ТАТА, которые находятся в сайте инициации транскрипции и содержат расположенные перед ними элементы на расстоянии более ста пар осн. от сайта инициации транскрипции, что модулирует уровни транскрипции. См. Gelvin, Transgenic Plants (Academic Press 1993).
Два гена, которые содержат расположенные против хода транскрипции активирующие последовательности, представлены генами октопин-синтазы (ocs) и маннопин-синтазы (mas). Ген ocs кодирует продукт, который конденсирует аргинин и пируват с образованием октопина. Hack and Kemp, Plant Physiol. 65:949-55 (1980). Палиндром из 16 пар оснований, расположенный перед геном ocs, активирует гетерологичный промотор кукурузы adhl в временной экспрессирующей системе. Ellis et al., EMBO J. 6:11-16 (1987); Ellis et al., EMBO J. 6:3203-08 (1987). Этот палиндром также важен для активности промотора ocs в стабильно трансформированном каллюсе табака. Leisner and Gelvin, Proc. Nat'l Acad. Sci. USA 85: 2553-57 (1988); Leisner and Gelvin, Plant Cell 1: 925-36 (1989).
Гены mas 1' и 2' делят двойной двунаправленный промотор и интергенный участок из 479 пар осн. Эти гены кодируют ферменты двустадийного пути синтеза маннопина Ellis et al., Mol. Gen. Genet. 195:466-73 (1984); Komro et al., Plant Mol. Biol. 4: 253-63 (1985). Транскрипция генов mas является дивергентной, а интергенный участок содержит все цис-действующие элементы, необходимые для транскрипции обоих генов. DiRita and Gelvin, Mol. Gen. Genet. 207: 233-41 (1987); Fox et al. Plant Mol. Biol. 20: 219-33 (1992); Leung et al., Mol. Gen. Genet. 230: 463-74 (1991); Guevara-Garcia et al., Plant J. 4: 495-505 (1993).
Промоторы генов ocs и mas использовали для направления экспрессии связанных генов в трансгенных растениях. Однако применение этих промоторов было ограничено низким уровнем экспрессии в определенных тканях трансгенных растений. DiRita and Gelvin, supra; Harpster et al., Mol. Gen. Genet. 212: 182-90 (1988); Sanger et al., Plant Mol. Biol. 14: 433-43 (1990). Например, промотор ocs направляет определенную, специфичную для клеток экспрессию в трансгенном табаке. Kononowicz et al., Plant Cell 4: 17-27(1992). Ген mas отличается слабой экспрессией в листьях и стеблях при более сильной экспрессии в корнях. Он проявляет повышенную экспрессию в повреждениях и индуцируется ауксином. Langridge et al. , Proc. Nat'l Acad. Sci 86: 7890-94 (1989); Teeri et al., EMBO J., 8: 343-50 (1989); Saito et al., Planta 184: 40-46 (1991) Guevara-Garcia et al., loc. cit.
Поскольку промоторы и другие регуляторные участки имеют различную силу и тканевую специфичность, были разработаны определенные рекомбинантные регуляторные участки. Например, энхансерные элементы, которые специфично связывают определенные транс-действующие факторы, могут модулировать активность транскрипции и экспрессию, специфичную для клеток. Bienz and Pelham, Cell 45: 753-60 (1986).
Известно также применение определенных конститутивных промоторов, например 35S конструкций вируса мозаики цветной капусты (CaMV). Промотор CaMV 35S имеет активаторы с множественными доменами, которые активируют промотор 35S в зависимости от стадии развития и тканевой специфичности. См. Benfey et al. , EMBO J. 8: 2195-2202 (1989); Benfey et al., EMBO J. 9: 1677-1684 (1990): Benfey et al., EMBO J. 9: 1685-96.
Публикация Koziel et al., Bio/Technology 11: 194-199 (1993) относится к промоторам, применяемым в конструкциях, получаемых для тканеспецифичной стимуляции гетерологичного гена. Koziel предлагает конструкцию системы экспрессии гена, которая содержит усеченный ген crylA(b) (фрагмент гена, кодирующий первые 648 аминокислот инсектицидного белка из Bacillus thuringiensis, состоящего из 1155 аминокислот), связанный с промотором CaMV 35S или комбинацией 2-х тканеспецифичных промоторов, выделенных из кукурузы (промотор фосфоенолпируват-карбоксилазы (PEPC) и промотор, специфичный для пыльцы). Koziel отмечает высокие уровни экспрессии при использовании любой промоторной конфигурации. Koziel et al. также использовали (1) промотор PEPC, который, как известно, вызывает специфичную для зеленых тканей экспрессию, и (2) промотор, специфичный для пыльцы кукурузы. Полученная экспрессия инсектицидного белка на уровне 1500 - 4000 нг/мг является достаточно высоким уровнем экспрессии. Кроме того, применение промоторов PEPC/специфичных к пыльце приводило к тканеспецифичной экспрессии.
Известны также попытки получить рекомбинантную экспрессию иными способами. Изобретение Bevan et al., PCT/GB92/00566 (1992) в основном относится к применению единственного промотора, отличного от промотора, описанного Koziel et al., для получения тканеспецифичной экспрессии гетерологичного гена. Это применение связано с использованием промотора фенилаланин-аммоний-лиазы бобов (PAL) для получения тканеспецифичной экспрессии. Был сконструирован гибридный ген, в котором слиты участки регуляции транскрипции геномного клона PAL (клона, содержащего в дополнение к кодирующим последовательностям, промежуточные последовательности, являющиеся частью последовательностей гена PAL) и открытая рамка считывания (ORF), содержащая 68 аминокислот N-конца белка PAL с полной ORF бета-глюкуронидазы (GUS) после участка полиаденилирования и терминации транскрипции нопалин-синтазы (слияние двух последних элементов типично для большинства экспрессирующих векторов эукариотных растений и обеспечивает эффективную экспрессию). Поскольку исходная конструкция обладала сильной активностью в ряде растений, исследователи создали делеции в участках промотора PAL гибридного гена. Минимальный промотор PAL, необходимый для полной экспрессии, как было показано, состоит из 253 пар осн. стартового сайта (инициации) транскрипции PAL. Варьированием участков делеции, главным образом в этом минимальном участке, исследователи установили возможность модификации степени тканеспецифичной экспрессии.
Патент США N 5034322 в основном относится к применению промоторов нопалин-синазы с малой субъединицей гена рибулоза-1,5-бис-фосфат-карбоксилазы.
Было также показано, что химерный промотор, названный "Mac", который содержит участок от +65 до -301 mas и участок от -90 до -941 энхансера 35S, обладает активностью GUS на уровне, в несколько раз превышающем уровень двойного промотора CaMV 35S в трансгенных растениях табака Comai et al., Plant Mol. Biol. 15: 373-81 (1990).
Описанные выше конструкции имеют ряд ограничений в плане эффективности и контролируемости экспрессии. Например, известные подходы не обеспечивают сильную конститутивную экспрессию в условиях, когда она желательна.
Сущность изобретения
Целью настоящего изобретения является получение химерных регуляторных участков для улучшенной экспрессии генов в растениях.
Изобретение также позволяет получать химерные регуляторные участки, которые содержат промоторы и находящиеся против хода транскрипции активирующие последовательности из A. tumefaciens.
Согласно изобретению также возможно получение генных кластеров, содержащих гены, экспрессирующиеся под контролем химерных регуляторных участков, которые содержат промоторы и находящиеся против хода транскрипции активирующие последовательности из A. tumefaciens.
Еще одним объектом изобретения является способ получения индуцибельной экспрессии чужеродных генов в растениях.
И, наконец, получение плазмид и трансгенных растений, содержащих генные кластеры, также предусмотрено настоящим изобретением.
В соответствии этим, согласно изобретению представлены химерные регуляторные участки для экспрессии генов в растениях, которые содержат указанную активирующую последовательность, выделенную из первого гена опин-синтазы A. tumefaciens, оперативно связанную с промотором, выделенным из второго гена опин-синтазы A. tumefaciens, отличного от первого гена опин-синтазы A. tumefaciens. Первый и второй гены опин-синтазы A. tumefaciens предпочтительно представлены генами маннопин-синтазы или октопин-синтазы.
Также представлены химерные регуляторные участки для экспрессии генов в растениях, содержащие по меньшей мере две активирующие последовательности опин-синтазы A. tumefaciens, оперативно связанные с промотором опин-синтазы A. tumefaciens. Указанные активирующие последовательности могут быть выделены из одного и того же или различных генов опин-синтазы A. tumefaciens, например маннопин-синтазы и октопин-синтазы. Дополнительно одна или обе активирующие последовательности и промотор могут быть выделены из одного и того же гена опин-синтазы A. tumefaciens.
В соответствии с изобретением предложены также генные кластеры, содержащие экспрессируемый ген, оперативно связанный с химерным регуляторным участком, как описано выше. Генный кластер может содержать также терминаторы транскрипции и сигналы полиаденилирования, например сигнал полиаденилирования нопалин-синтазы.
Кроме того, предложен кластер для индуцибельной экспрессии чужеродного гена, который содержит чужеродный ген, оперативно связанный с регуляторным участком, который содержит промотор, выделенный из гена маннопин-синтазы A. tumefaciens и расположенную против хода транскрипции активирующую последовательность, выделенную из гена маннопин-синтазы A. tumefaciens. Регуляторный участок может также содержать активирующие последовательности, выделенные из гена октопин-синтазы A. tumefaciens.
Объектами изобретения являются также способы экспрессии генов в растениях, которые предусматривают стадии связывания гена с химерным регуляторным участком, соответствующим изобретению, инсерцию гена и химерного регуляторного участка в растение и обеспечение экспрессии гена в растении.
Кроме того, представлены плазмиды и трансгенные растения, несущие описанные в изобретении химерные регуляторные участки и генные кластеры.
Иные объекты, отличия и преимущества настоящего изобретения будут рассмотрены при обсуждении экспериментальных данных и чертежей.
Краткое описание чертежей.
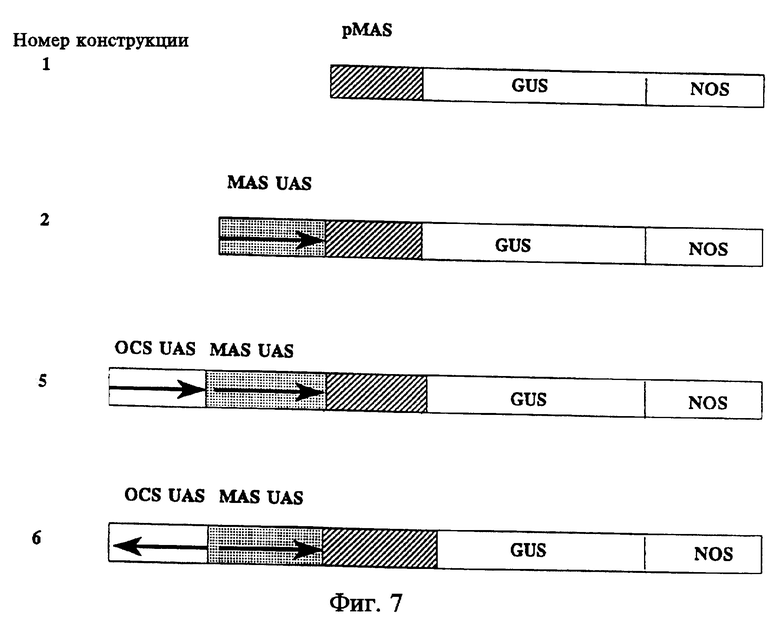
На фиг. 1 схематически представлена структура химерных регуляторных участков на основе mas и ocs. Стрелки указывают на ориентацию расположенных против хода транскрипции активирующих последовательностей относительно промоторов mas и ocs. Цифры означают положение нуклеотидов относительно сайтов инициации транскрипции. Тример активирующей последовательности ocs был использован в конструкциях 3 и 4. Мономер или тример активирующей последовательности ocs был использован в конструкциях 5 и 6.
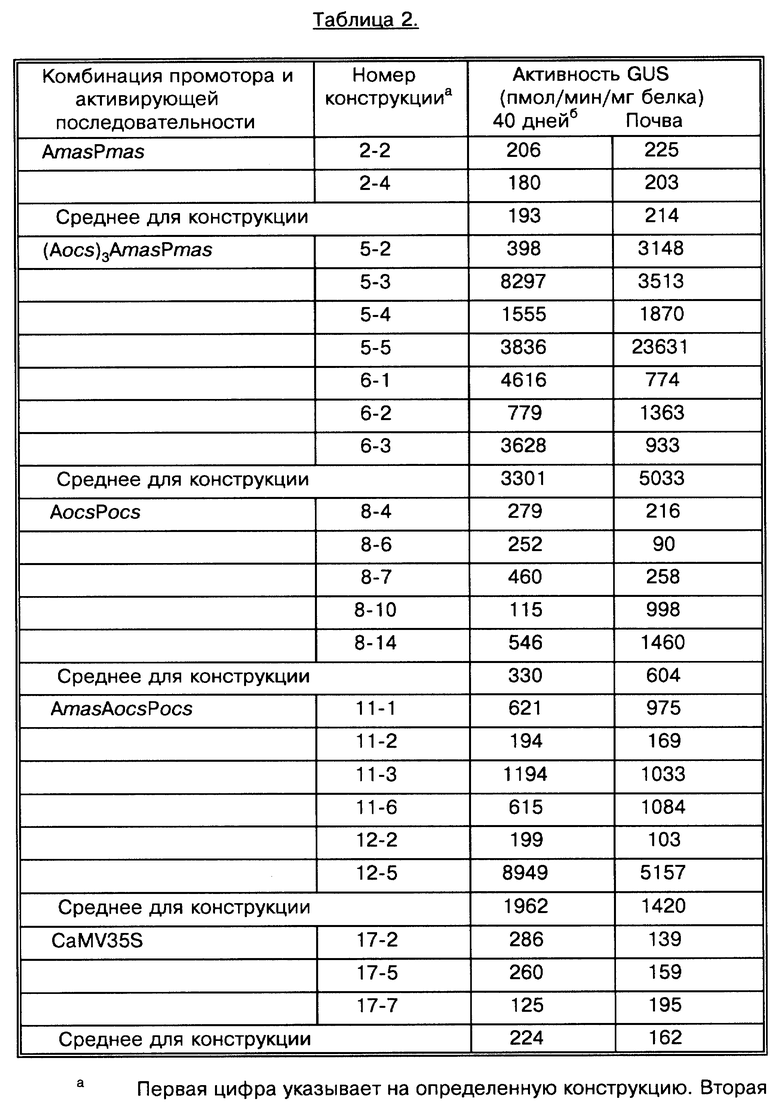
На фиг. 2 показана активность GUS конструкций на основе промотора mas в экстрактах первичных трансформантов табака. Активность GUS оценивали по общему белку, выделенному из тканей листьев (граф. А), стеблей (граф. Б) или корней (граф. В). Каждый столбец представляет активность отдельного трансформанта. Различные конструкции указаны внизу графиков. Число отдельных трансгенных растений, проанализированных в отношении каждой конструкции, представлено "n". Средняя активность GUS для каждой конструкции обозначена "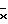 ". A - активирующая последовательность, P - промотор, (Aocs)3 - тример активирующей последовательности ocs.
". A - активирующая последовательность, P - промотор, (Aocs)3 - тример активирующей последовательности ocs.
На фиг. 3 показана активность GUS конструкций на основе промотора mas плюс активирующей последовательности в экстрактах первичных трансформантов табака. Активность GUS оценивали по общему белку, выделенному из тканей листьев (граф. А), стеблей (граф. Б) или корней (граф. В). Каждый столбец представляет активность отдельного трансформанта. Число отдельных трансгенных растений, проанализированных в отношении каждой конструкции, представлено "n". Различные конструкции указаны внизу графиков. Средняя активность GUS для каждой конструкции обозначена "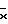 ". A - активирующая последовательность, P - промотор, AocsAmasPmas - мономер активирующей последовательности ocs, связанной с активирующей последовательностью mas и промотором; (Aocs)3AmasPmas - тример активирующей последовательности ocs, связанной с активирующей последовательностью mas и промотором.
". A - активирующая последовательность, P - промотор, AocsAmasPmas - мономер активирующей последовательности ocs, связанной с активирующей последовательностью mas и промотором; (Aocs)3AmasPmas - тример активирующей последовательности ocs, связанной с активирующей последовательностью mas и промотором.
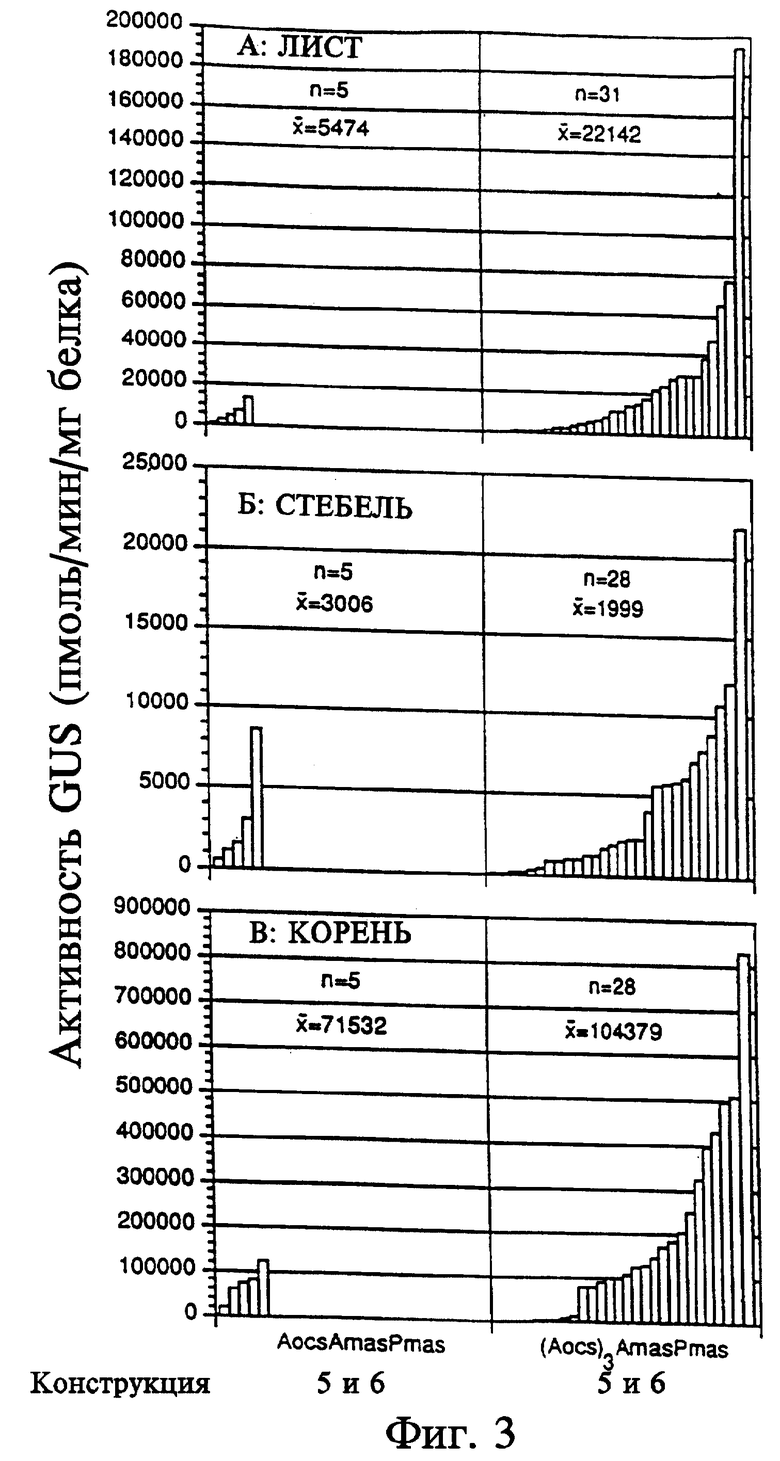
На фиг. 4 показана активность GUS конструкций на основе промотора ocs в экстрактах первичных трансформантов табака. Активность GUS оценивали по общему белку, выделенному из тканей листьев (граф. А), стеблей (граф. Б) или корней (граф. В). Каждый столбец представляет активность отдельного трансформанта. Различные конструкции указаны внизу графиков. Число отдельных трансгенных растений, проанализированных в отношении каждой конструкции, представлено "n". Средняя активность GUS для каждой конструкции обозначена "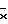 ". A - активирующая последовательность, P - промотор, Amas' - участок mas от -213 до -318, Amas'' - участок mas от -111 до -318.
". A - активирующая последовательность, P - промотор, Amas' - участок mas от -213 до -318, Amas'' - участок mas от -111 до -318.
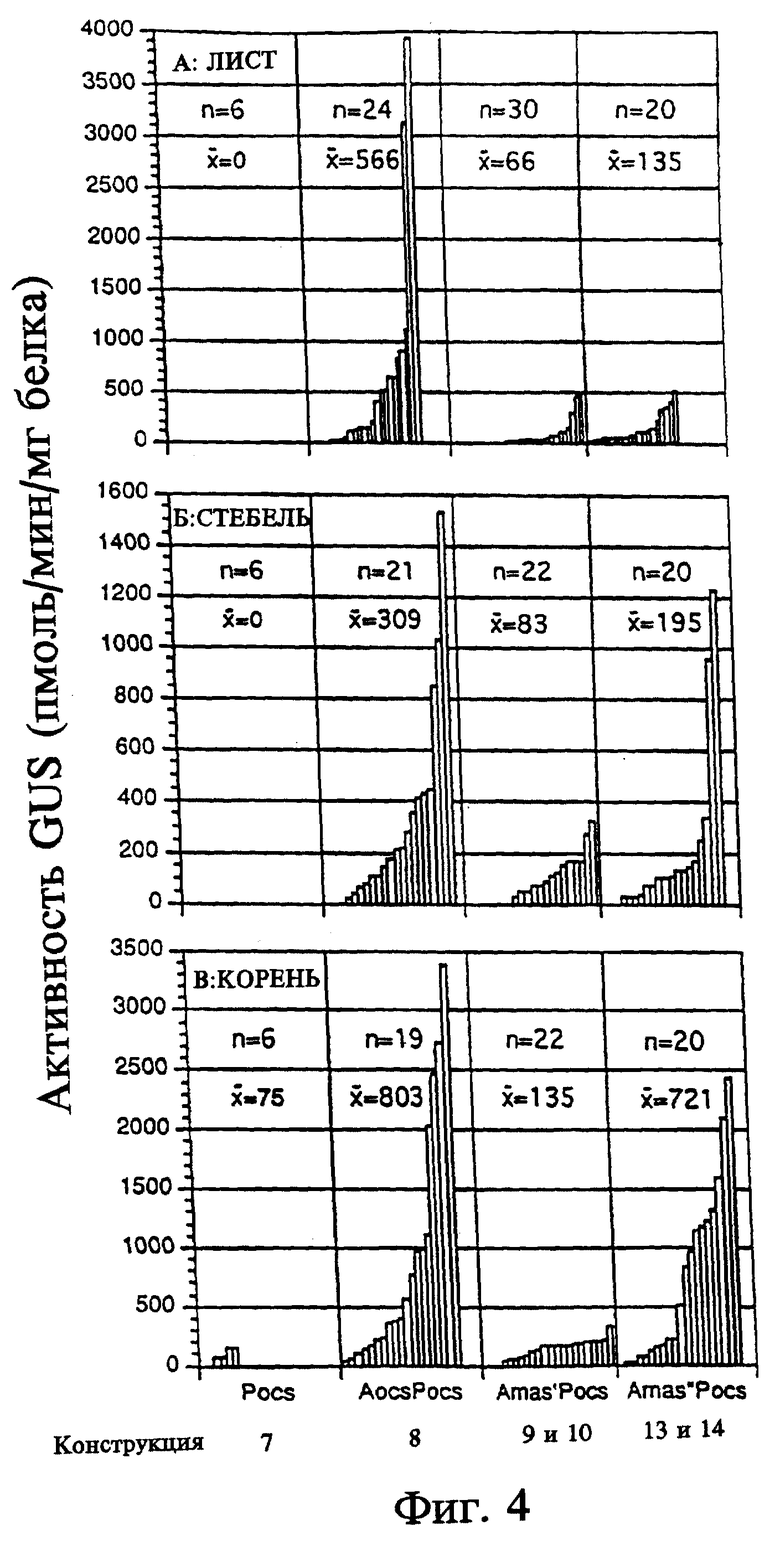
На фиг. 5 показана активность GUS конструкций на основе промотора ocs плюс активирующей последовательности в экстрактах первичных трансформантов табака. Активность GUS оценивали по общему белку, выделенному из тканей листьев (граф. А), стеблей (граф. Б) или корней (граф. В). Каждый столбец представляет активность отдельного трансформанта. Различные конструкции указаны внизу графиков. Число отдельных трансгенных растений, проанализированных в отношении каждой конструкции, представлено "n". Средняя активность GUS для каждой конструкции обозначена "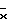 ". A - активирующая последовательность, P - промотор, Amas' - участок mas от -213 до -318, Amas'' - участок mas от -111 до -318.
". A - активирующая последовательность, P - промотор, Amas' - участок mas от -213 до -318, Amas'' - участок mas от -111 до -318.
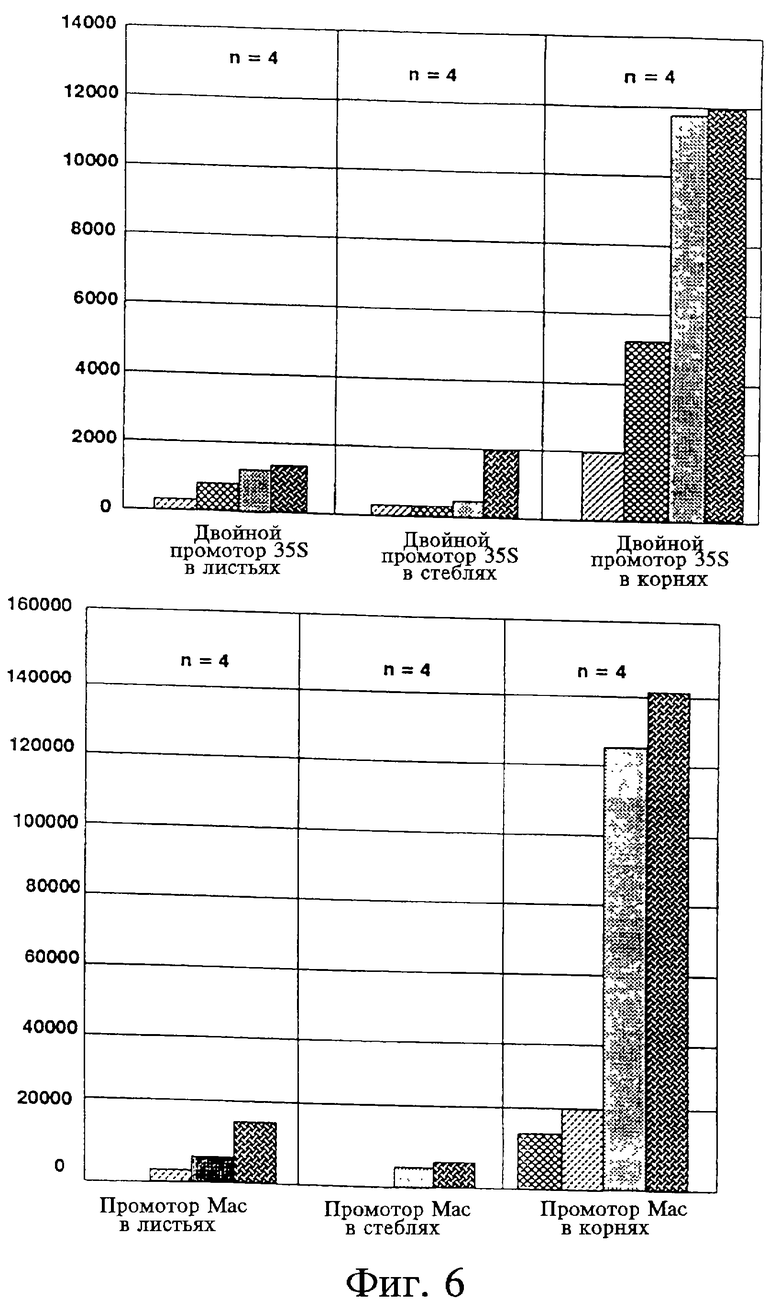
На фиг. 6 представлены приблизительные уровни экспрессии активности GUS двойных промоторов CaMV 35S и Mac в листьях (двойной 35SL, MacL), стеблях (двойной 35SS, MacS) и корнях (двойной 35SR, MacR) нескольких трансгенных растений табака.
На фиг. 7 схематично представлены конструкции, использованные при изучении индуцибельной экспрессии. UAS - расположенная против хода транскрипции активирующая последовательность, pmas - промотор mas, GUS - ген β -глюкуронидазы, NOS - сигналы полиаденилирования нопалин-синтазы.
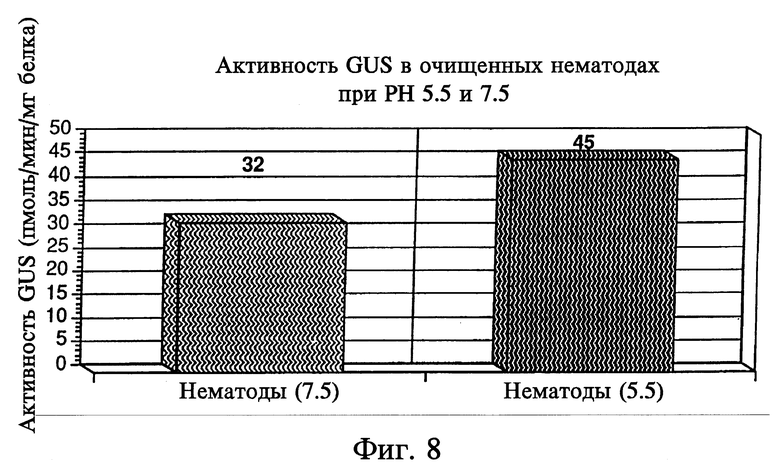
На фиг. 8 представлен уровень активности GUS в выделенных зрелых яйцах нематод. Анализы проводили при pH 5,5 и 7,5 для проверки активности GUS бактериального и животного происхождения.
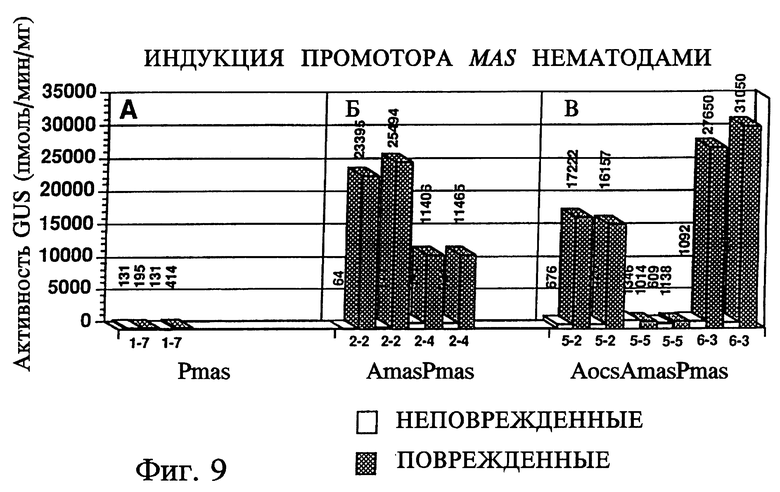
На фиг. 9 представлена активность индуцированной нематодами GUS в различных трансгенных растениях. На панели А представлены трансгенные растения, имеющие промотор Pmas, но не имеющие активирующих последовательностей, на панели Б - трансгенные растения, имеющие промотор AmasPmas, на панели В - трансгенные растения, имеющие промотор AocsAmasPmas.
Подробное описание предпочтительных вариантов изобретения.
Настоящее изобретение относится к химерным регуляторным участкам, содержащим различные расположенные против хода транскрипции активирующие последовательности и промоторы из генов опин-синтазы A. tumefaciens, которые применяют для контроля экспрессии чужеродных генов. Чужеродный ген содержит любую ДНК, планируемую для экспрессии в трансгенном растении. В данном контексте ген, независимо от его происхождения, вводят в геном растения, и он, таким образом, является чужеродным растению в локализации инсерции даже в случае, когда ген выделен из трансформируемого растения. Изобретенные конструкции в соответствии с настоящим изобретением также описаны в патентной заявке США N 08/155067, заявлено 19.11.93, включенной в настоящее описание путем ссылки.
Все типы продуктов, кодируемых генами, можно получить с помощью настоящего изобретения. Чужеродные гены, экспрессируемые в трансгенных растениях, согласно настоящему изобретению, включают, но не ограничиваются β -глюкуронидазой, генами, кодирующими инсектицидные и фунгицидные токсины, соединениями, обусловливающими устойчивость к патогенам, соединениями, обусловливающими сверхчувствительную реакцию, например пероксидазами, глюканазами и хитиназами, а также фитоалексинами; генами устойчивости к пестицидам, гербицидам и фунгицидам; ферментами растений, например связанными с содержанием белков, крахмала, сахаров и жиров; ингибиторами ферментов растений, например ингибиторами протеазы и амилазы; гормонами растений; гормонами и феромонами насекомых; фармацевтическими и пищевыми соединениями, например бета-каротином; витаминами и антителами, в том числе фрагментами и одноцепочечными производными антител, и антисмысловыми транскриптами, нарушающими нуклеотидные последовательности, присутствующие в растениях. См., например, Kung and Wu, Transgenic Plants, vol. 1 (Academic Press 1993).
В общем, природный промотор маннопин-синтазы с активирующей последовательностью или природный промотор октопин-синтазы с активирующей последовательностью являются относительно слабыми в тканях трансгенных растений табака (активность GUS, обусловленная связанными репортерными генами gusA, составляет несколько сотен, несколько тысяч единиц активности GUS). В соответствии с настоящим изобретением сочетание UAS из этих генов с промотором с активирующей последовательностью заметно повышает активность GUS связанного гена gusA (на два порядка), приводя к активностям порядка сотен тысяч единиц активности GUS. Гистохимическое исследование ткани трансгенных растений табака экспрессирующих эти слитые гены промотор-gusA показывает, что новые комбинации промотора и активирующей последовательности, представленные в изобретении, работают в большинстве типов клеток растений. Таким образом, эти комбинации (химерные регуляторные участки) могут быть более конститутивными по сравнению с известными промоторами.
Изобретение относится к улучшенным регуляторным участкам растений, обеспечивающим сильную и постоянную стимуляцию экспрессии генов. Дополнительно может быть получена тканенеспецифичная и/или повышенная в определенных тканях экспрессия. Выбор активирующих последовательностей и промоторов в соответствии с изобретением обусловливает желаемый тип экспрессии.
Представленные химерные регуляторные участки характеризуются более сильной экспрессией в табаке, чем любой другой известный регуляторный участок. Табак является широко используемой моделью для трансформации растений. Дополнительно описанные регуляторные участки могут быть близкими к конститутивным (постоянно включающимися) и потенциально могут быть использованы в большем числе тканей растений, чем известные регуляторные участки.
Изобретение относится к химерному регуляторному участку для экспрессии генов в растениях. Он может содержать расположенную против хода транскрипции активирующую последовательность, выделенную из первого гена опин-синтазы A. tumefaciens, оперативно связанную с последовательностью промотора, выделенной из второго гена опин-синтазы A. tumefaciens или с расположенной против хода транскрипции активирующей и промоторной последовательностью, выделенной из второго гена опин-синтазы A. tumefaciens.
Химерные регуляторные участки, согласно изобретению, могут быть оперативно связаны с последовательностью чужеродного гена, которая может быть оперативно связана с работающей в растении терминаторной последовательностью и далее оперативно связана с работающей в растении сигнальной последовательностью полиаденилирования. Химерный регуляторный участок в соответствии с изобретением может быть высоко конститутивным во многих случаях.
Расположенная против хода транскрипции активирующая последовательность является последовательностью, естественное положение которой обычно, по меньшей мере, на сто пар оснований предшествует положению природного сайта инициации транскрипции и которая может оказывать влияние на экспрессию. UAS генов октопин- и маннопин-синтазы используют, в частности, с этой целью. Эти UAS могут быть затем оперативно связаны с последовательностью промотора или с расположенной против хода транскрипции активирующей последовательностью и промотором, выделенными из иного гена опин-синтазы A. tumefaciens.
Термин "выделены", применяемый в отношении таких участков ДНК, как промоторы и расположенные против хода транскрипции активирующие последовательности, описывает ситуации, когда "выделенный" участок ДНК получают из участка ДНК или он имеет основой природный участок ДНК или участок ДНК из иного источника. "Выделенный" участок ДНК может отличаться мутацией от природного участка ДНК или участка ДНК, полученного из иного источника.
Выражение "оперативно связанные" относится к первой(ым) последовательности(ям), помещенной(ым) достаточно близко ко второй(ым) последовательности(ям) для того, чтобы первая(ые) последовательность(и) оказывала(и) влияние в области второй(ых) последовательности(ей) или участка, находящегося под контролем этой второй последовательности. Например, UAS может быть оперативно связана с промотором, при этом она повышает транскрипционную силу промотора. В такой ситуации типично 5'-расположение UAS относительно промотора. UAS и промотор, в свою очередь, могут быть оперативно связаны с генами, при этом экспрессия гена будет находиться под контролем комбинации UAS/промотор, для которой также типично 5'-положение относительно гена. Обычно промотор содержит 30 - 50 пар оснований от стартового сайта транскрипции и до нескольких сотен пар оснований от стартового сайта трансляции. Активирующая последовательность обычно составляет несколько сот пар оснований промотора. Например, большинство активирующих последовательностей содержат 300 - 400 пар оснований усиливаемого промотора. В вариантах изобретения, где используется более одной активирующей последовательности, они обычно содержат 100 - 200 пар оснований одна другой.
В сконструированном согласно изобретению химерном регуляторном участке источники генов опин-синтазы различны и предпочтительно выбраны из группы генов опин-синтазы, содержащей гены маннопин-, октопин-, нопалин- и агропин-синтазы.
Экспрессия активности GUS, направляемая промоторами mas и ocs с активирующими последовательностями, ограничена определенными типами клеток. Оперативное связывание активирующей последовательности ocs с промотором mas и с активирующей последовательностью, а также оперативное связывание активирующей последовательности mas с промотором ocs и с активирующей последовательностью показывает модуляцию экспрессии в сравнении с нативными конструкциями. Так, экспрессия GUS, а также иных генов может быть получена в большом числе типов клеток, включая сосуды ксилемы и эпидермальные клетки листьев. С другой стороны, ограниченные образцы экспрессии также могут быть получены в другом варианте изобретения. Химерный регуляторный участок, оперативно связывающий UAS ocs и минимальный промотор mas, дает сниженную экспрессию в сосудистой ткани листьев и экспрессию в стеблях, связанную с тканями флоэмы.
Изобретение также относится к рекомбинантному кластеру генов, кодирующему чужеродный полипептид. Рекомбинантный кластер генов может содержать находящуюся против хода транскрипции активирующую последовательность, выделенную из первого гена опин-синтазы A. tumefaciens и последовательность промотора или активирующую и промоторную последовательность, выделенные из иного, второго гена опин-синтазы A. tumefaciens. Все указанные последовательности оперативно связаны с последовательностью чужеродного гена. В конструкции последовательность чужеродного гена может быть оперативно связана с работающей в растении последовательностью терминатора и/или работающей в растении сигнальной последовательностью полиаденилирования. При этом последовательность терминатора и сигнал полиаденилирования могут оказывать влияние на ген или транскрипт гена. Последовательность терминатора и сигнал полиаденилирования имеют 3'-положение относительно гена.
Следующий аспект изобретения относится к комбинациям промотора и расположенной против хода транскрипции активирующей последовательности (регуляторным участкам), которые индуцируются повреждениями или вредителями. Первая из разработанных комбинаций (AmasPmas) предпочтительно экспрессируется в тканях корней и индуцируется патогенами, тогда как другая (AocsAmasPmas) экспрессируется во всем растении, но индуцируется вредителями. Эти экспрессирующие системы применимы для генов, направленных на корневых вредителей, например на нематоды или грибы, и могут использоваться против вредных насекомых и патогенов листьев. Например, гены, кодирующие нематоцидные токсины и белки, которые нарушают репродуктивный цикл нематод, могут быть применены в рамках настоящего изобретения.
Заражение патогенами индуцирует химерные регуляторные участки, в которых использован промотор маннопин-синтазы с активирующими последовательностями mas или ocs, как описано выше. Ряд генов, используемых для получения устойчивости против патогенов, описан Keen, Plant Molec. Biol. 19: 109-22 (1992).
Транскрипционные элементы, например промоторы и расположенные против хода транскрипции активирующие последовательности генов опин-синтазы, могут быть получены на основе имеющейся информации о последовательностях. Например, транскрипционные элементы генов октопин-синтазы описаны Leisner et al., Proc. Nat'l Acad. Sci. USA 85: 2553-57 (1988); Leisner et al., Plant Cell 1: 925-36 (1989). Транскрипционные элементы генов маннопин-синтазы описаны DiRita and Gelvin, supra, Fox et al., Plant Molec. Biol. 20: 219-33(1992); Leung et al., Mol. Gen. Genet. 230:463-74 (1991); Langridge et al., Proc. Nat'l Acad. Sci USA 86: 3219-23 (1989). Транскрипционные элементы генов нопалин-синтазы Ha et al., Nucl. Acids Res. 17: 215-23 (1989); Mitra et al., Mol. Gen. Genet. 215: 294-99 (1989); Ebert et al., Proc. Nat'l Acad. Sci USA 84: 57-49 (1987); An et al. , Mol. Gen. Genet. 203: 245-50 (1986). Элементы контроля транскрипции гена агропин-синтазы описаны Bandyopadhyay et al., J. Biol. Chem. 264: 19399-406 (1989). Дополнительно, полная последовательность Т-ДНК описана Barker et al., Plant Molec. Biol. 2: 335-50 (1983).
Различные уровни и образцы экспрессии были получены следующими приведенными способами. Количество и образец экспрессии оценивают с помощью маркерных систем, например, описанных здесь gusA. Изобретение проиллюстрировано следующими примерами, которые не ограничивают его.
Пример 1
Добавление расположенных против хода транскрипции активирующих последовательностей mas или ocs к промотору mas или ocs и активирующей последовательности значительно увеличивает экспрессию GUS.
Как показано на фиг. 1, были созданы новые комбинации промоторов mas и ocs. Были проанализированы различные субдомены UAS mas, поскольку предварительный делеционный анализ показал, что последовательности из 138 пар оснований, находящиеся перед сайтом инициации транскрипции, являются достаточными для точной инициации транскрипции слитого гена mas2'/nptll в тканях корончатых галл подсолнечника. Последовательности от -138 до -318, однако, могут также участвовать в регуляции количественного уровня активности промотора mas2'. DiRita and Gelvin, Mot. Gen. Genet. 207; 233-41 (1987). Проанализированные UAS mas представлены: (i) UAS от -318 до -138; (ii) UAS' от -318 до -213; (iii) UAS'' от -318 до -111. См. фиг. 1.
Первая группа химерных регуляторных участков была сконструирована и связана в качестве транскрипционных слияний с геном uidA (gusA) с помощью двух делеций -318 и -138 пар оснований от сайта инициации транскрипции (конструкции 1-6 фиг. 1) промотора mas. Первый сет химерных регуляторных участков содержит (в любой ориентации) мономер или тример активирующей последовательности ocs (от-116 до -333) перед делецией -138 промотора mas (конструкции 3 и 4 фиг. 1). Второй сет химерных регуляторных участков содержит подобные мономеры или тримеры активирующих последовательностей ocs перед делецией -318 промотора mas (конструкции 5 и 6 фиг. 1). Этот участок ocs содержит палиндром из 16 пар оснований, а также 5'- и 3'-последовательности модулятора, которые важны для активирования промотора ocs в каллюсе и растениях табака при стабильной инкорпорации в геном растений. Leisner and Gelvin, 1988 and 1989 supra; Kononowicz et al., Plant Cell 4: 17-27 (1992).
Вторая группа химерных регуляторных участков была сконструирована в виде трансляционных слияний с геном uidA на основе двух делеций ocs в положениях -333 и -116 от сайта инициации транскрипции (конструкция 8-16 фиг. 1). Было создано два участка расположенной против хода транскрипции активирующей последовательности mas. Короткий участок mas, содержащий последовательность от -213 до -318, был использован в конструкциях 9-12 фиг. 1. Длинный участок mas, содержащий последовательность от -111 до -318, был использован в конструкциях 13-16 фиг. 1. Короткий или длинный участки mas были введены перед двумя делециями ocs.
Химерные конструкции были получены следующим образом. Основой всех конструкций был бинарный вектор pBl101.2 из Clontech. Плазмида pBl101.2 имеет основой репликон pRK290. Плазмида pBl101.2 содержит границы Т-ДНК, химерный ген nos-nptll для селекции на фоне канамицина у растений и не имеющий промотора ген GUS после сигнала полиаденилирования. Регион промотора mas был получен из фрагмента EcoRV-XbaI, который содержит участок от -138 до +65, соответствующий сайту инициации транскрипции (пары оснований 20128 - 20343) из pKan2-138. Barker et al., Plant Mol. Biol. 2: 335-50 (1983): DiRita and Gelvin, supra. Активирующая последовательность и промоторный участок mas от -318 до +65, соответствующие сайту инициации транскрипции (пары оснований 20128 - 20513) из pKan2-318, были изначально клонированы в сайты SmaI-XbaI CUp31 (производное pUC13, имеющее основу pUC13, но содержащее полилинкер, считываемый в направлении 5' - 3' как HindIII, PstI, SstI, SmaI, BamH1, XbaI). Полученные фрагменты рестрикции HindIII-XbaI из CUp31 были последовательно реклонированы в сайты HindIII-XbaI мультилинкера pBl101.2, с образованием плазмид pNi1 и pNi2 (конструкции 1 и 2 соответственно).
Фрагмент энхансера ocs из плазмиды pEN Δ 1 (Leisner and Gelvin (1988) supra) был получен с помощью линкеров HindIII. Этот фрагмент соответствует от -333 до -116 сайта инициации транскрипции (пары оснований 13774 -13991, Barker et al., supra). Он был клонирован как тример в сайт HindIII pNi1 перед промотором mas в обеих ориентациях, что дало конструкции 3 и 4 (см. фиг. 1). Для создания конструкций 5 и 6 этот же фрагмент активирующей последовательности ocs был клонирован как тример или как мономер в сайт HindIII pNi2 перед активирующей последовательностью и промотором mas.
Фрагмент BamH1-EcoRI, содержащий участок промотора ocs с частью структурного гена ocs, который соответствует от -116 до +296 сайта инициации транскрипции (пары оснований 13774 - 13362) и фрагмент XbaI-EcoRI, содержащий активирующую последовательность и промоторный участок ocs, соответствующий участку от -333 до +296 сайта инициации транскрипции (пары оснований 13991 - 13362) из pEN1 (Leisner and Gelvin (1988), supra) были клонированы в сайты BamH1-EcoRI и XbaI-EcoRI соответственно, pBluescriptll SK+ (Стратаген). Фрагменты XbaI-EcoRV из полученных производных pBluescript были последовательно клонированы в сайты XbaI-SmaI pBl101.2 с созданием трансляционных слияний GUS в плазмидах pLH3 (конструкция 7) и pNi3 (конструкция 8). Фрагмент XhoI-HaeIII, содержащий короткую активирующую последовательность mas, соответствующую участку от -318 до -213 сайта инициации транскрипции (пары оснований 20513 - 20407) из pKan2-318, был клонирован в сайты XhoI-HincII pUX13 (производное pUC13 содержащее основу pUC13, но с сайтом SmaI, превращенным в сайт XhoI). Полученный XhoI-HindIII фрагмент был сделан непосредственно из фрагмента Klenow при введении линкеров XnaI. Фрагмент клонирован в обеих ориентациях в сайты XbaI pLH3 (конструкции 9 и 19) и pNi3 (конструкции 11 и 12). Аналогично длинный фрагмент активирующей последовательности masXhoI-MnII, соответствующей сайту инициации транскрипции (пары оснований 20513-20305) из pKan2-318, был сделан при непосредственном использовании фрагмента Klenow. Были введены линкеры XbaI и фрагмент был клонирован в обеих ориентациях в сайты XbaI pLH3 (конструкции 13 и 14) и pNi3 (конструкции 15 и 16).
Каждой из описанных конструкций последовательно трансформировали E. coli DH5a, выращенную при 37oC в среде LB с канамицином. Ориентации инсерций были определены рестрикционным картированием. Плазмида pBl121 (Clontech), содержащая фрагмент HindIII-BamHI из 800 пар оснований с промотором CaMV 35S была использована в качестве контроля для сравнения относительных длин химерных регуляторных участков.
Рекомбинантные плазмиды, содержащие инсерций, были введены в A. tumefaciens LBA4404 способом скрещивания с участием трех родителей с использованием E. coli MM294, несущей мобилизующую плазмиду pRK2013. Hoekema et al., Nature 303: 179-80 (1983); Ditta et al., Proc. Nat'l Acad. Sci. 77: 7347-51 (1980). В LBA4404 рекомбинантные плазмиды сохраняются как независимые репликоны (бинарные векторы), которые могут быть перенесены в растения с последующей интеграцией в ядерную ДНК растения. Иные способы, например электропорация, также могут быть использованы для трансформации клеток A. tumefaciens плазмидами.
Трансконьюганты A. tumefaciens были отселектированы на чашках с минимальной средой AB, содержащей 0,5% глюкозы, 10 мкг/мл рифампицина и 50 мкг/мл канамицина. Lichtenstein and Draper, DNA Cloning: a practical approach (Glover ed. , Oxford-IRL Press 1986). Интродукция мобилизованной плазмиды в реципиентный штамм A. tumefaciens была определена блоттингом ДНК.
Диски, вырезанные из листьев шестинедельных стерильных культур Nicotiana tabacum var. Wisconsin 38, были трансформированы с помощью A. tumefaciens несущей конструкции, с использованием способа трансформации листовых дисков. Horsh et al., Science 227: 1229-31 (1985). Инфицированные листовые диски в течение трех дней выращивали на среде MS3+ без антибиотиков, затем диски перенесли на свежую среду, индуцирующую образование проростков, которая содержала 1250 мг/л карбенициллина и 200 мг/л канамицина. Kononowicz et al., Plant Cell 4: 17-27 (1992). Через 4 - 5 недель по одному побегу с каждого листового диска перенесли в среду, индуцирующую образование корней, которая содержала 500 мг/л карбенициллина и 50 мг/л канамицина. Через две недели растения перенесли в среду BGS (среда MS, содержащая 1 мг/л фолиевой кислоты, 10 мг/л индол-уксусной кислоты и 30 мг/л кинетина), содержащую 50 мг/л канамицина, с целью поддержания культур каждой линии in vitro.
Регенерированные трансгенные растения табака, содержащие каждую из описанных конструкций, были проверены на активность GUS. Маленькие кусочки ткани растения были выделены из четвертого или пятого полностью развернувшегося листа, около стеблей и из активно растущих молодых корней, когда растения сформировали по 10 - 12 листьев. Ткани были растерты в 200 мкл экстракционного буфера и сохранялись при -70oC. Jefferson and Wilson, Plant Molecular Biology (Gelvin & Schilperoot eds., Kluwer Acad. Press 1991). Активность GUS определяли по Jefferson and Wilson, используя 10 мкл экстракта (20 - 30 мкг белка) и MUG (4-метилумбеллиферил -β- D-глюкуронид) в качестве субстрата. Концентрацию белка определяли по Patterson, Analyt Biochem. 83: 346-56 (1977).
Отдельные трансгенные растения, содержащие одну и ту же конструкцию, имели различия в активности GUS (см. фиг. 2 - 5). Однако, при определении активности GUS у большого числа растений, несущих одну и ту же конструкцию, можно установить относительную силу каждого химерного регуляторного участка. Поскольку не были установлены различия в уровне активности GUS при использовании конструкций, в которых элементы активирующих последовательностей были клонированы в противоположных ориентациях, данные по каждой двучленной подгруппе конструкций были суммированы (3 и 4, 5 и 6, 9 и 10, 11 и 12, 13 и 14, 15 и 16).
Во всех исследованных тканях экспрессия, направляемая делецией -138 mas, имела минимальный уровень активности GUS (фиг. 2 конструкция 1). Добавление природной активирующей последовательности mas к минимальному промотору mas приводило к образованию конструкции 2, которая обусловливала низкий уровень активности GUS в тканях листьев и стеблей. Он составлял около тысячи единиц (фиг. 2, граф. А и Б). Для этой конструкции была отмечена относительно высокая активность GUS в тканях корней. Она составляла около 12 тысяч единиц (фиг. 2, граф. В). Эти результаты показывают, что данные промотор и активирующий участок mas обеспечивают предпочтительную экспрессию в корнях промотора mas2', описанного выше. Замена активирующей последовательности mas гетерологичной активирующей последовательностью ocs (в виде тримера) перед делецией -138 mas (конструкции 3 и 4) практически не изменяет уровень активности GUS относительно комбинаций гомологичной активирующей последовательности mas и промотора mas в тканях листьев и стеблей (фиг. 2, граф. А и Б). Вместе с результатами, которые показывают, что экспрессия активирующей последовательности ocs и промотора незначительно выше в корнях, чем в листьях (см. ниже конструкция 8), данные результаты позволяют предположить, что предпочтительная экспрессия в корнях промотора mas обусловлена элементом из 138 пар оснований сайта инициации транскрипции mas. Количественный уровень тканеспецифичной экспрессии может быть еще более повышен активирующими последовательностями ocs или mas.
Идентифицирован гомолог тандемного повтора AS-1 (Lam et al., Proc. Nat'l Acad. Sci. 86: 7890-94 (1989) в участке промотора mas в положении -66. Этот элемент, находясь в промоторе CaMV 35S, взаимодействует с транс-действующим фактором ASF-1 (Lam et al., loc. cit.) и направляет тканеспецифичную экспрессию с высокой активностью в корнях (Benfey et al., supra).
Активность GUS, направляемую химерными конструкциями, сравнили с активностью, полученной при использовании промотора CaMV 35S (-800 pBl121). Большинство трансформантов 35S GUS имели активность, близкую или ниже таковой у растений, содержащих конструкции активирующей последовательности mas и промотора (фиг. 2). Lam et al., supra показана предпочтительная для корней экспрессия промотора 35S.
Результаты показывают, что промоторы ocs и mas содержат цис-элементы, которые направляют транскрипцию тканеспецифичным образом. Kononowicz et al., Plant Cell 4: 17-27 (1992); Leung et al., Mol. Gen. Genet. 230: 463-74 (1991). В свете этих результатов был проведен анализ комбинаций гетерологичных активирующих последовательностей с целью установления, может ли комбинация менять образец экспрессии, направляемый одним промотором ocs или mas, и повышать или снижать активность промотора в определенных тканях растения. Для проверки этой гипотезы конструкции 5 и 6 (см. фиг. 1), которые содержат тример или мономер активирующей последовательности ocs, расположенный перед активирующей последовательностью mas и промотором, были интродуцированы в табак. Активность GUS при этом была измерена в различных тканях (фиг. 3). Новые химерные участки, содержащие мономер активирующей последовательности ocs, повышали экспрессию активности GUS в 6,6 раза в листьях, в 3,0 раза в стеблях и в 3,4 в корнях в сравнении с промотором и активирующей последовательностью mas2' (см. фиг. 2 и 3). Конструкции, содержащие тример активирующей последовательности ocs, сильно повышали экспрессию активности GUS в листьях (в 22 раза), в стеблях (1,7 раза) и корнях (в 9 раз) в сравнении с промотором и активирующей последовательностью mas2' (конструкция 2 на фиг. 1 и 2), не содержащими дополнительной активирующую последовательность ocs. Эти результаты показывают, что по меньшей мере в тканях листьев и корней введение множественных копий активирующей последовательности ocs дает неожиданно мощный усиливающий эффект на соответствующую активность промотора и активирующей последовательности mas2'.
Анализы показывают, что активность промотора CaMV 35S в листьях трансгенных растений (в среднем 200 пмол/мин/мг белка) была сравнима с данными Comai et al., Plant Mol. Biol. 15: 373-81 (1990). Дупликация энхансера 35S привела к двукратному повышению активности GUS в листьях. Сравнивая результаты, приведенные Comai et al., loc. cit., с данными, полученными для предложенных конструкций, можно сказать, что химерные участки конструкций 5 и 6 обеспечивали соответственно 156- и 26-кратное усиление экспрессии GUS в листьях по сравнению с промотором 35S и усиленным двойным промотором 35S соответственно.
Были также проверены семена генерации T2 растений табака, содержащие конструкции двойной 35S-GUS или Mac-GUS. Определения активности GUS в листьях этих трансгенных растений подтвердили
относительные силы промоторов, как указано выше. В таблице 1 представлена средняя активность GUS в листьях трансгенных растений табака, несущих различные варианты слияний промотор-uidA.
В таблице 2 приведена средняя активность GUS в листьях в поколении F1 трансгенных растений табака, несущих различные варианты слияний промотор-uidA.
Серия химерных конструкций на основе минимального промотора ocs (от -116 до +296) была проанализирована с целью оценки силы и образцов тканеспецифичной экспрессии, как показано на фиг. 4, активирующая последовательность ocs и промотор определяют низкий и относительно универсальный уровень активности GUS (200 - 400 пмол/мин/мг белка) в тканях листьев, стебля и корней трансгенных растений табака. Замещение активирующей последовательности ocs коротким вариантом активирующей последовательности mas (от -213 до -318, конструкции 9 и 10) приводило к образованию химерной конструкции, определяющей низкий уровень активности GUS во всех исследованных тканях.
Длинный вариант активирующей последовательности mas (от -111 до -318), расположенный перед минимальным промотором ocs (конструкции 13 и 14), был также проанализирован. Эти конструкции обусловливали несколько повышенный уровень активности GUS, но только в тканях корней. В дополнение к гомологу AS-1 в положении -66 последовательность, близкая к элементу AS-1, была обнаружена в положении -290. Хотя эта последовательность присутствует в коротком варианте активирующей последовательности mas, иные последовательности между положениями -213 и -103 необходимы для предпочтительной экспрессии в корнях.
Короткий вариант активирующей последовательности mas (от -318 до -213, конструкции 11 и 12) был помещен перед активирующей последовательностью ocs и промотором. В сравнении с конструкцией, содержащей только активирующую последовательность ocs и промотор (конструкция 8), эти конструкции обеспечивали 6-, 2,5- и 15-кратное повышение активности GUS в тканях листьев, стебля и корней соответственно (см. фиг. 5). Экспрессия этих конструкций была несколько более предпочтительной в корнях, что предполагает возможность взаимодействия элемента, подобного AS-1, с активирующей последовательностью ocs. Конструкции, содержащие длинный вариант UAS mas (связанные с активирующей последовательностью ocs и промотором (конструкции 15 и 16)), обусловливали незначительно более низкий уровень активности GUS по сравнению с конструкциями 11 и 12, содержащими короткую активирующую последовательность mas (см. фиг. 5). Связывание активирующих последовательностей ocs и mas по 5'-концу с промотором mas приводило к значительному увеличению силы промотора по сравнению с конструкцией, состоящей из активирующей последовательности ocs и промотора.
Пример 2
Корреляция между числом копий Т-ДНК и активностью GUS в трансгенных растениях табака.
Для установления связи между числом копий Т-ДНК и активностью GUS была проанализирована геномная ДНК из 16 трансгенных растений, содержащих конструкцию 5 или 11. Геномную ДНК экстрагировали из тканей растений и расщепляли полностью с помощью HindIII. Rogers and Bendich, PLANT MOLECULAR BIOLOGY (Gelvin and Schilperoot eds., Kluwer Acad. Pub., 1992). 10 мкг ДНК обрабатывали HindIII, фрагменты разделяли электрофоретически в 1,0% агарозном геле и элюировали ДНК на нейлоновую мембрану способом капиллярного переноса. Maniatis et al. , MOLECULAR CLONING: A LABORATORY MANUAL (Cold Spring Harbor 1982). Нуклеиновые кислоты фиксировали на мембране нагреванием при 80oC под вакуумом в течение 2 часов. Предварительную гибридизацию проводили в течение 2 - 4 часов при 65oC в 1,5xSSC (раствор хлорида и цитрата натрия) (1xSSC - 0,15 М NaCl, 0,015 М цитрат натрия), 1,0% SDS (додецилсульфат натрия), 0,5% Blotto и 0,5 мг/мл ДНК измельченной молоки лосося.
Гибридизацию проводили при 65oC в течение ночи в свежем растворе с зондом, содержащим участок кодирующей последовательности GUS (рестрикционный фрагмент XbaI-SstI из pBl101.2) и меченным 32P-dCTP. После окончания гибридизации мембрану промывали в течение 15 минут при комнатной температуре следующими растворами: 2х SSC/0,1% SDS, 0,5х SSC/0,1% SDS, 0,1х SSC/0,1% SDS. Окончательное промывание проводили в течение 30 минут при 50oC 0,1х SSC/1,0% SDS.
С помощью этой комбинации рестриктаз и гибридизационного зонда число копий Т-ДНК устанавливали по числу и интенсивности гибридизационных бэндов. Число интегрированных копий варьировало от одного до нескольких в отдельных трансформантах. Активность GUS не коррелировала с числом интегрированных генов uidA. Например, растение, содержащее единственный интегрированный ген uidA, в ряде случаев обладало значительно более высокой активностью GUS в листьях, чем растение, содержащее множественные интегрированные копии гена uidA.
Пример 3
Корреляция между активностью GUS и мРНК uidA в трансгенных растениях табака.
Поскольку активность GUS была использована как мера силы экспрессии, было необходимо удостовериться, что эта активность отражает существующий уровень мРНК uidA. Эта корреляция является важной, потому, что существуют данные, что активность GUS не коррелируют с количеством мРНК uidA при использовании промотора mas2' и репортерного гена uidA. Hensgens et al., Plant Mol. Biol. 20: 921-38 (1992).
В соответствии с этим был изучен уровень мРНК uidA, выделенной из листьев отдельных трансгенных растений, содержащих 4 различных конструкции (конструкции 2, 5, 8, 11). Сначала выделили общую РНК по de Vries et al., PLANT MOLECULAR BIOLOGY (Gelvin and Schilperoot eds., Kluwer Acad. Pub., 1992). Образцы по 5 мг были фракционированы электрофорезом в формальдегидном геле через 1,2% агарозный гель в буфере МПСК/ЭДТА (50 мМ МПСК, 1 мМ ЭДТА, pH 7,0) с последующим блоттингом на нейлоновой мембране. Цельность РНК была проверена электрофорезом в агарозном геле и окрашиванием этидий-бромидом. Флюоресценция нуклеиновых кислот также служила для подтверждения того, что каждая дорожка содержала равные количества РНК. Условия гибридизации были описаны выше для анализа геномной ДНК. После окончания гибридизации мембрану промывали в течение 15 минут при комнатной температуре следующими растворами: 2х SSC/0,1% SDS, 0,5х SSC/0,1% SDS, 0,1х SSC/0,1% SDS. Окончательное промывание проводили в течение 30 минут при 60oC 0,1х SSC/1,0% SDS.
При блоттинг-анализе РНК с помощью гибридизационного зонда, выделенного из кодирующей последовательности GUS, был получен транскрипт ожидаемого размера (приблизительно 2300 нуклеотидов). Была отмечена тесная корреляция между активностью GUS и уровнем мРНК uidA, что противоречит данным Hensgens et al.
Пример 4
Модуляция специфичной для клеток экспрессии GUS различными комбинациями промоторов и активирующих последовательностей ocs и mas.
Гистологические исследования тканей трансгенного табака были проведены для установления специфичной для клеток активности GUS. Такие образцы экспрессии были получены при гистохимическом окрашивании тонких срезов тканей растения X-gluc.
Эти гистохимические исследования проводились по Jefferson and Wilson, supra. Растительные материалы предварительно фиксировали в течение 20-40 минут 0,1-0,3% формальдегидом, 0,1 М Тритоном Х-100, 0,1 М фосфатным буфером (pH 7,0), промывали 0,1 М фосфатным буфером и окрашивали 1 - 2 мМ X-gluc (в 0,1 М Тритоне Х-100, 0,01 М ЭДТА, 0,1 М фосфатном буфере) в течение 2 - 14 часов. После повторной фиксации в течение двух часов с использованием 3-5% формальдегида в 0,1 М фосфатном буфере образцы обрабатывали 70% этанолом, помещали в парафин и делали срезы (12 - 18 мм) на роторном микротоме. Срезы тканей контрастно окрашивали 1,0% периодной кислотой - 0,5% реактивом Шиффа ("PAS").
Ни в одной из исследованных типах клеток тканей листьев растений, несущих промотор mas, но не имеющих активирующую последовательность (конструкция 1), не было обнаружено активности GUS. Растения, содержащие химерный ген uidA под контролем природной активирующей последовательности mas и промотора (конструкция 2), имели средний уровень активности GUS в клетках мезофилла листьев (включая столбчатую и губчатую паренхиму) и замыкающих клетках, но не имели активности GUS в клетках эпидермиса. В тканях сосудов было отмечено умеренное окрашивание трахеид ксилемы при относительно более слабом окрашивании флоэмы и клеток лучевой паренхимы. Близкие значения активности GUS были отмечены в листовых пластинках, несущих тример активирующей последовательности ocs и промотора mas (конструкция 3 и 4). Однако в сосудистых тканях листьев активность GUS была сильно снижена во всех типах клеток. Связывание тримера активирующей последовательности ocs с активирующей последовательностью mas с промотором (конструкции 5 и 6) обусловило сильную активность GUS не только в мезофилле и замыкающих клетках листьев, но и в эпидермальных клетках. Это означает, что дистальная и проксимальная гетерологичные активирующие последовательности взаимодействуют, модулируя экспрессию. В сосудистых тканях листьев экспрессия не зависела от наличия тримерных активирующих последовательностей, связанных с активирующей последовательностью mas с промотором.
Активность GUS не была отмечена в тканях листьев, содержащих минимальный промотор ocs (конструкция 7). Экспрессия активности GUS, определяемой активирующей последовательностью ocs с промотором (конструкция 8), была близка к таковой, обусловленной активирующей последовательностью mas с промотором в поперечных срезах листовой пластинки. Этот образец активности GUS был отмечен в листовых пластинках растений, в которых ген uidA находился под контролем химерного промотора, состоящего из короткой активирующей последовательности mas и активирующей последовательности ocs с промотором (конструкции 11 и 12). В сосудистой ткани черешков листьев активирующая последовательность ocs с промотором также направляла экспрессию GUS, но только в трахеидах. В отличие от активирующей последовательности mas с промотором, активирующая последовательность ocs с промотором определяла умеренный уровень экспрессии активности GUS в клетках флоэмы и очень слабую экспрессию в трахеидах паренхимы и ксилемы в основных сосудистых тканях листьев. Близкий образец активности GUS был получен в сосудистой ткани листьев независимо от добавления короткой активирующей последовательности mas к активирующей последовательности ocs с промотором. Хотя активность была значительно более высокой в паренхиме, лучевых клетках и клетках флоэмы, активность GUS не определялась в растениях, содержащих активирующую последовательность mas, связанную с промотором ocs (конструкции 9 и 10).
Активирующая последовательность mas с промотором (конструкция 2) определяла слабую экспрессию активности GUS в ножках железистых трихом, но сильную экспрессию в головках. Оперативное связывание тримера активирующей последовательности ocs с активирующей последовательностью mas с промотором (конструкции 5 и 6) приводило к близкому образцу экспрессии. Однако относительный уровень активности GUS был сильно повышен в клетках ножек. Экспрессия активности GUS в трихомах, направляемая конструкциями, содержащими короткую активирующую последовательность mas связанную с активирующей последовательностью ocs с промотором (конструкции 11 и 12), была сильной в головках железистых трихом, но слабой в клетках ножек.
В стеблях трансгенных растений активирующая последовательность mas с промотором (конструкция 2) определяла слабую экспрессию активности GUS в корковых клетках. Сильная активность GUS была отмечена в лучевых клетках и клетках флоэмы, но экспрессия в трахеидных клетках ксилемы была относительно слабой. Этот образец экспрессии не изменялся даже при использовании тримера активирующей последовательности ocs перед активирующей последовательностью mas с промотором (конструкции 5 и 6). Когда активирующая последовательность mas была заменена тримером активирующей последовательности ocs (конструкции 3 и 4), был отмечен иной образец экспрессии. Экспрессия активности GUS оставалась сильной в клетках флоэмы, но была слабой в лучевых клетках и клетках трахеид ксилемы. В отличие от активирующей последовательности mas с промотором активирующая последовательность ocs с промотором (конструкция 8) определяла относительно более слабую экспрессию в лучевых клетках и клетках флоэмы при сильной экспрессии в клетках трахеид. Когда короткий промотор mas был введен перед активирующей последовательностью ocs с промотором (конструкция 11), образец экспрессии в основном сохранялся.
Активность GUS, определяемая активирующей последовательностью mas с промотором (конструкция 2) в тканях корней, была различной. Было получено регенерированное растение, в котором активность GUS определяли в клетках корневого чехлика, эпидермальных клетках, корневых волосках, корковых клетках корней, в клетках флоэмы и ксилемы зоны созревания корня. Однако, в зоне удлинения корня была зарегистрирована низкая активность GUS. Активность GUS часто, но не всегда определяли в каждом типе клеток зоны созревания корня в случае, когда тример активирующей последовательности ocs был добавлен к активирующей последовательности mas с промотором (конструкции 5 и 6). Замена активирующей последовательности mas тримером активирующей последовательности ocs (конструкции 3 и 4) обусловливала активность GUS только в корковых и эндодермальных клетках зоны удлинения периферических корней. Подобные образцы активности GUS были отмечены, когда промотор ocs и активирующая последовательность (конструкция 8) или короткая активирующая последовательность mas с активирующей последовательностью ocs с промотором (конструкции 11 и 12) направляли экспрессию гена uidA.
Пример 5
Сравнение предлагаемых химерных регуляторных участков с иными регуляторными участками.
Активность активирующей последовательности mas с промотором является наивысшей в тканях корней. Введение тримера активирующей последовательности ocs к активирующей последовательности mas с промотором повышало уровень активности GUS в 2-23 раза. Это повышение активности максимально в тканях листьев, но также отмечается в тканях стебля и корня. Активность активирующей последовательности ocs с промотором приблизительно одинакова в листьях, стеблях и корнях трансгенных растений табака. Введение короткого варианта активирующей последовательности mas к активирующей последовательности ocs с промотором повышало уровень экспрессии GUS в 2-20 раз. Это повышение активности было наиболее значительным в тканях листьев. Сочетание множественных копий активирующей последовательности ocs с активирующей последовательностью mas с промотором приводило к образованию транскрипционного регуляторного элемента, который в листьях был примерно в 156 раз сильнее, чем промотор 35S, в 26 раз сильнее, чем усиленный двойной промотор CaMV 35S и в 4,2 раза сильнее, чем промоторы "Mac" и "Big Mac", описанные Comai et al., Plant Mol. Biol. 15: 373-81 (1990) (см. фиг. 2 и 3 и табл. 1). Данные относительно двойного промотора 35S и промотора mas для листьев (L), стеблей (S) и корней (R) представлены на фиг. 6. Следует отметить, что большая активность химерных промоторов, представленная конструкциями 5 и 6 относительно двойного промотора CaMV 35S и промоторов Mac и Big Mac, оценивается минимально. Дополнительно гистохимический анализ клеток трансгенных растений, несущих эти конструкции, показал, что эти дополнительные активирующие последовательности направляли экспрессию активности GUS почти во всех типах клеток. Например, сильная экспрессия активности GUS была отмечена в ксилеме и эпидермальных клетках листьев, а также в мезофилле, замыкающих клетках, трихомах и клетках флоэмы листьев. В стебле активность была отмечена в клетках флоэмы и паренхимы, а также в корковых клетках. В корнях активность GUS была обнаружена в корневом чехлике и корневых волосках, а также в большинстве клеток зрелых частей корня.
Возможное влияние условий роста растений на различные химерные конструкции промотор-uidA должно быть принято во внимание. Hensgens et al., Plant Mol. Biol. 20: 921-38 (1992) показали, что активность GUS у растений, выращенных in vitro (укорененных в стерильном агаре), была в три раза больше, чем у растений, выращенных в почве в теплице. Дополнительно было показано, что листья растений, выращенных в почве в условиях внешней среды, экспрессировали активность GUS на более низком уровне, чем растения, выращенные in vitro. Однако относительные уровни активности GUS, определяемые различными регуляторными участками, не зависели от условий выращивания растений (табл. 2, supra). Дополнительно, относительная сила различных химерных регуляторных участков в листьях поколения F1 самоопыленных трансгенных растений табака была близка к таковой трансформированных и регенерированных растений (табл. 2).
Была обнаружена тесная корреляция между уровнем мРНК и активностью GUS у растений. Этот результат подтверждает достоверность использования данных анализа активности GUS как меры силы экспрессии. Эти данные противоречат таковым Hensgens et al., supra. Эти противоречия могут объясняться неудачным выбором условий денатурации для блоттинг-анализа РНК указанным автором.
Comai et al., supra показали, что, когда растения выращиваются в почве, участок активирующей последовательности mas2' и промотора короче, чем в наших исследованиях (-301 в сравнении с -318) и направляет только 10% активности GUS относительно силы промотора CaMV 35S. Введение последовательностей перед -301 повышало относительную активность GUS до 40% от активности промотора 35S. Comai et al. считают, что участок, расположенный против хода транскрипции, но близко к -300, необходим для реализации полной активности промотора mas2'. Активность этого промотора (-318), используемая в приведенных экспериментах, была несколько выше, чем активность, определяемая промотором 35S.
Langridge et al., supra показали, что активность слитого гена mas2'-lux может повышаться в несколько раз с помощью гормонов. Хотя были проведены исследования на растениях, выращенных in vitro в присутствии гормонов, эти условия роста не оказывали сильного влияния на результаты, которые продемонстрировали близкие относительные уровни активности GUS, полученные для растений, выращенных в почве.
Langridge et al. также показали, что в стеблях трансгенного табака активность промотора mas2' максимально экспрессируется в сосудистых тканях. Saito et al., supra продемонстрировали сильное окрашивание в корневом чехлике при более слабом окрашивании клеток флоэмы корней табака. Наши данные хорошо коррелируют с указанными. Однако Saito et al. определили активность GUS в сосудах, но не в клетках мезофилла листьев. В наших экспериментах активность GUS была обнаружена в клетках мезофилла листьев трансгенных растений, несущих химерный ген mas2'-uidA. Многие представленные нами результаты также коррелируют с результатами Leung et al. касательно экспрессии промотора mas2' в различных тканях трансгенных растений табака. Однако Leung et al. не отметили активность GUS в сосудистых клетках стеблей при использовании их конструкций.
Комбинирование гетерологичных активирующих последовательностей ocs или mas с активирующими последовательностями mas или ocs с промотором сильно повышало уровень активности GUS во всех исследованных тканях табака. Эти данные показывают, что повышенная экспрессия химерных регуляторных участков обусловлена совместным синергическим взаимодействием, а не аддитивными эффектами между положительными регуляторными элементами, которые находятся в химерных регуляторных участках.
Пример 6
Использование химерных регуляторных участков в индуцибельной экспрессии.
Другой аспект изобретения относится к комбинациям промотора с расположенной против хода транскрипции активирующей последовательностью, индуцируемым повреждениями или вредителями. Одна из полученных комбинаций (AmasPmas) предпочтительно экспрессируется в тканях корней и индуцируется патогеном, тогда как другая (AocsAmasPmas) экспрессируется более генерализованно, но индуцируется вредителями. Эти экспрессирующие системы применяют для генов с направленностью на вредителей корней, например нематод и грибы, они могут быть использованы против вредных насекомых и иных патогенов листьев.
Химерные регуляторные участки были сконструированы с использованием ядра участка промотора маннопин-синтазы с делецией до -138. Этот промотор был слит с кодирующей последовательностью GUS и терминирован сигналами полиаденилирования нопалин-синтазы (NOS). См. фиг. 7, конструкция 1.
UAS от -318 до -138 промотора маннопин-синтазы использовали в сочетании с ядром промотора маннопин-синтазы для создания конструкции 2 фиг. 7. Конструкции 5 и 6 фиг. 7 содержат тример UAS из гена октопин-синтазы от -333 до -116 в обеих ориентациях. Эти конструкции были введены в табак с помощью A. tumefaciens. Активность GUS определяли в листьях, стеблях и корнях большого числа отдельных трансгенных растений. Активность промотора mas и активирующей последовательности (фиг. 7 конструкция 2) сильнее в корнях и значительно слабее в листьях и стеблях. Введение активирующей последовательности ocs к активирующей последовательности mas и промотору (фиг. 7 конструкции и 6) повышали активность GUS в 10 раз в корнях и в 50 - 100 раз в стеблях и листьях по сравнению с конструкцией 2. Ориентация активирующей последовательности ocs эффекта не оказывала.
Индуцибельность повреждениями промотора mas с активирующей последовательностью возрастает до 30 раз в листьях, 17 раз в стеблях и до 3 раз в корнях трансгенных растений табака.
Для проверки индуцибельности различных химерных регуляторных участков патогенами отдельные трансгенные растения, содержащие конструкции 1, 2, 5 и 6, инфицировали нематодами, а затем проводили мониторинг индукции активности GUS. Чтобы удостовериться в отсутствии эндогенной экспрессии активности GUS в нематодах, были проанализированы очищенные препараты развитых яиц нематод. Были использованы нематоды Meloidogyne incognita расы 3, выделенные из корней томатов. Зрелые яйца получали, помещая зараженные корни в 10% раствор хлорокса на 4 мин при постоянном перемешивании. Затем раствор отмывали через ячеистое сито 200 для удаления корневого дебриса, а яйца собирали на ячеистом сите 500. Яйца промывали и подсчитывали с использованием Устройства для подсчета нематод Nematode Counting Slide (Olympic Equine Products). В качестве контроля для определения эндогенной активности GUS у нематод яйца анализировали при pH 5,5 и 7,5. Активность GUS при pH 5,5 обусловлена эндогенной экспрессией животных форм гена, которая дает фоновую активность. Активность GUS при pH 7,5 обусловлена бактериальной экспрессией гена. Активность GUS в зрелых яйцах нематод была минимальной при указанных значениях pH (фиг. 8). Таким образом, Meloidogyne incognita не несла бактериальной и животной активности GUS, которые могли бы создать проблемы фоновой активности в анализах с использованием яиц нематод.
Трансгенные растения были инфицированы 10000 очищенных яиц нематод и выращены на песчаной почве в тепличных условиях в течение нескольких месяцев. Были собраны корни зрелых растений, в которых визуально определили места нематодной инфекции по образованию корневых узелков, содержащих нематод и кладки яиц, тогда как неинфицированные места имели нормальный вид. Эти корневые узелки использовали для измерения активности GUS в инфицированных участках, а нормальные участки корней использовали в качестве контроля. Инфицированные и неинфицированные корни сравнивали у одного и того же растения, поэтому результаты не зависели от положения растения и вариаций экспрессии гена в различных растениях. Индукция активности GUS была определена в конструкции 1 с использованием растения номер 7 (1-7) при pH 5,5 и 7,5 (фиг. 9, панель А).
Уровни экспрессии были низкими в сравнении с трансгенными растениями, несущими конструкции 2, 5, 6. Это указывает на то, что один промотор Pmas экспрессирует только базовый уровень активности и не индуцируется инфекцией нематод.
Когда промотор Pmas содержит активирующую последовательность mas, Amas (конструкция 2), экспрессия активности GUS высокая и индуцируется нематодной инфекцией (фиг. 9, панель Б). Введение UAS промотора ocs (Aocs) дает подобную индукцию при нематодной инфекции (фиг. 9, панель В). Одно растение (5-5) не проявляло индуцибельную экспрессию. Это объясняется возрастом ткани корня или инсерцией гена в место на хромосоме, которое вызывало изменения в экспрессии, поскольку другой трансформант (5-2) давал реакцию индукции.
Эти результаты демонстрируют, что патоген индуцирует химерные регуляторные участки с помощью промотора маннопин-синтазы с активирующими последовательностями mas или ocs. Эти участки могут быть использованы для экспрессии генов нематоцидных токсинов или гормональных соединений, которые нарушают питание и размножение нематод. Например, ряд токсинов из Bacillus thuringiensis (токсины Bt) эффективны в отношении нематод. См. Adang et al., Plant Molec. Biol. 21: 1131-45 (1993). Аминокислотные последовательности этих токсинов и нуклеотидные последовательности кодирующих их генов представлены в Патентах США N 5281530 и 5322932, публикации PCT WO 92/04453 и Европейской патентной публикации 0517367 A1.
Применение этих индуцибельных регуляторных участков не ограничивается применением против нематод. Эти участки могут быть равно эффективными против иных патогенов, вызывающих повреждения.
Пример 7
Химерные регуляторные участки, контролирующие экспрессию инсектицидных токсинов.
Представленные в настоящем изобретении химерные регуляторные участки могут быть использованы для контроля экспрессии инсектицидных токсинов в трансгенных растениях. В качестве предпочтительного применения токсины Bt могут быть помещены под контроль представленных регуляторных участков.
С целью повышения уровней экспрессии в трансгенных растениях были разработаны модифицированные гены, кодирующие токсины Bt. Perlak et al., Proc. Nat'l Acad. Sci. USA 88: 3324-28 (1991) предложил модификации гена CryIA для замены последовательностей, которые не работают в растениях. Эти модификации повышают уровень активного токсина CryIA. Аналогично Adang et al., supra предложили модификации гена cryIIIA для улучшения экспрессии. Другие гены семейства токсина Bt также могут быть подвергнуты модификациям и использованы с настоящим изобретением. Возможно также применение генов, связанных с гиперчувствительной реакцией. См. Keen, supra. Настоящее изобретение не ограничено никаким специфичным типом токсина или соединения. Инсектицидные токсины или соединения любого типа, кодируемые генами или образуемые ферментами или иными веществами, кодируемыми генами, могут быть использованы с настоящим изобретением.
Пример 8
Химерные регуляторные участки, контролирующие экспрессию генов устойчивости к гербицидам.
Предложенные в настоящем изобретении химерные регуляторные участки могут быть использованы для экспрессии в трансгенных растениях генов, несущих устойчивость к гербицинам. Эта устойчивость определяется тремя первичными подходами: (i) связанная с растениями детоксикация гербицидов, (ii) повышенная экспрессия мишеней гербицидов и (iii) мутация сайтов связывания гербицидов. См. Schulz et al., Crit. Rev. Plant Sci. 9: 1-15 (1990). Любой из описанных подходов может быть использован с настоящим изобретением, хотя первые два подхода более соответствуют ему.
Известно несколько ферментов-детоксификаторов многих обычно применяемых гербицидов. Например, глутатион-S-трансферазы обусловливают устойчивость к s-триазиновым и хлорацетамидным гербицидам. См. например Schulz et al., supra; Shah et al., Plant Molec. Biol. 6:203-11 (1986); Weigand et al., Plant Molec. Biol. 7: 235-43 (1986). Фосфинотрицин был инактивирован с помощью гена из Streptomyces hygroscopicus. De Block et al., EMBO J. 6: 2513-18 (1987); Thompson et al., EMBO J. 6: 2519-23 (1987). Было показано, что гены нитрилазы детоксифицируют бромоксинил. Stalker et al., Science 242: 419-23 (1988). Остальные детоксифицирующие ферменты также могут быть использованы с настоящим изобретением.
Другой подход к первичной устойчивости основан на повышенной экспрессии мишеней гербицида. Например, глиофосфат является конкурентным ингибитором 5-енол-пирувилшикимат-3-фосфат-синтазы (EPSP-синтазы). Устойчивость к глиофосфату преодолевается повышенной экспрессией EPSP-синтазы, которую получают при использовании сильных конститутивных промоторов настоящего изобретения для контроля экспрессии последовательностей EPSP-синтазы. Дополнительно множественные копии последовательностей EPSP-синтазы с промоторами, предложенными в настоящем изобретении, могут быть помещены в трансгенные растения для получения еще более высоких уровней экспрессии. Известны последовательности, кодирующие EPSP-синтазу из различных источников. См. Duncan et al. , FEBS Lett. 170: 59 (1984); Stalker et al., J. Biol. Chem. 260: 4724-28 (1985); Shah et al., Science 233: 478-81 (1986).
И, наконец, изменение сайта связывания гербицида также может быть использовано для получения устойчивости к гербицидам. Stalker et al. и Shah et al. показали, что изменение аминокислот в EPSP-синтазе может определять устойчивость к глиофосфату. Аналогично, изменение в синтазе ацетогидроксикислот может обусловливать устойчивость к сульфанилмочевинам и имидазолинонам. См. Schulz et al., supra и Wek et al., Nucl. Acid. Res. 13: 3995-4010 (1985).
Пример 9
Химерные регуляторные участки, контролирующие экспрессию генов устойчивости к вирусам.
Устойчивость к вирусам в растениях может быть обусловлена экспрессией вирусных генов или антисмысловых последовательностей вирусных генов. Первичными подходами для получения устойчивости к вирусам являются: (i) устойчивость, связанная с белками, обычно белками оболочки и (ii) устойчивость, связанная с антисмысловой РНК. Beachy et al., Annu. Rev. Phytopathol. 28: 451-74 (1990) предложили обзор обоих указанных подходов. Трансгенные растения, экспрессирующие белок оболочки вируса, имеют большую устойчивость к вирусам, чем растения, содержащие антисмысловую РНК. Beachy et al., supra; Cuozzo et al., Bio/technology 6: 549-57 (1988).
Регуляторные участки, описанные в настоящем изобретении, могут быть использованы для экспрессии любого гена, определяющего устойчивость трансгенных растений к вирусам. Например, известны аминокислотные последовательности многих белков оболочки вирусов растений, что позволяет определить их нуклеотидные последовательности. Дополнительно известны нуклеотидные последовательности генов, кодирующих белки оболочки вирусов, что позволяет осуществить синтез точных последовательностей антисмысловых РНК. Возможна экспрессия этих последовательностей в трансгенных растениях с целью получения устойчивости.
Cuozzo et al., supra предложили последовательности белка оболочки вируса мозаики огурца. Были сконструированы трансгенные растения, содержащие иные вирусные последовательности, например Anderson et al. , Phytopath. 79: 1284-90 предложили трансгенные растения, которые экспрессируют белки оболочки вируса табачной мозаики и вируса мозаики люцерны. Hemenway et al., EMBO J. 7: 1273-80 (1988) предложили трансгенные растения, которые экспрессируют белок оболочки и антисмысловую РНК вируса X картофеля. Huisman et al., J. Gen. Biol. 69: 1789-98 (1988) описали последовательность этого вируса. Gerlach et al. , Nature 328: 802-05 (1987) представили трансгенные растения, которые экспрессируют сателлитную РНК вируса кольцевой пятнистости табака. Эта сателлитная РНК ослабляет симптомы кольцевой пятнистости. Eggenberger et al. , J. Gen. Virol. 70: 1853-60 представили последовательность из вируса мозаики сои. Однако применение настоящего изобретения не ограничено определенными типами вирусов. Последовательность из любого типа вируса может быть использована с настоящим изобретением.
Следует отметить, что описание, примеры, фиг. 1 - 9 и данные, представляющие предпочтительное использование, даны как иллюстрация и в качестве примеров, а не для ограничения настоящего изобретения. Различные изменения и модификации в рамках изобретения станут очевидными исследователю из приведенных дискуссии и открытия.




Группа изобретений может быть использована в селекции растений. Химерные регуляторные участки нуклеотидных последовательностей и генные конструкции на основе генов опин-синтазы Agrobacterium fumefaciens обеспечивают повышенные уровни экспрессии чужеродных генов в растениях. Для увеличения экспрессии генов различные "upstream"-расположенные активирующие последовательности, например, из генов маннопин- и октопин-синтазы операбельно соединяют с промоторами, которые, в свою очередь, операбельно соединяют с чужеродным геном. Конструкции вводят в растительные клетки с помощью плазмид. 19 с. и 13 з.п.ф-лы, 9 ил., 2 табл.
| Генная инженерия растений | |||
| Пер | |||
| с англ./Под ред | |||
| Дж.Дрейпера и др.-М.: Мир, 1991, с | |||
| Походная разборная печь для варки пищи и печения хлеба | 1920 |
|
SU11A1 |
| Способ получения промотора для конструирования вектора и способ конструирования вектора для экспрессии белков в клетках | 1990 |
|
SU1836424A3 |
