Изобретение касается растений с пониженной восприимчивостью к растительным паразитическим нематодам и способа их получения, отличающегося тем, что включает метод рекомбинантных ДНК.
Прототипы изобретения.
Распространенные по всему миру растительные паразитические нематоды вызывают заболевания почти всех экономически важных культурных растений, потери только в США оценивают в 5,8 миллиардов долларов в год (Sasser и Freckman 1987, Мировые перспективы в области нематодологии, в Vistas on Nematology под редакцией Veech и Dickson. Hyatts Vill, Maryland, стр. 7 - 14). В то время как в тропических регионах потери вызваны в основном клубеньковыми нематодами (root-knot nematodes) (Meloidogyne), в Европе серьезными вредителями и важными факторами, ограничивающими культивацию, например, картофеля, капусты и сахарной свеклы, считают нематоды родов Globodera и Heterodera соответственно. Только небольшое количество устойчивых сортов культур появилось в результате программ по селекции, например, картофеля, сахарной свеклы, помидоров, сои и масличной редьки (Dropkin, 1988, Ann. Rev. Phytopath. 26, 145-161. Trudgill, 1991, Ann. Rev. Phytopath. 29, 167 - 192). Часто эта устойчивость основана на одном P-гене (Rick и Fobes, 1974, Tomato Gen. Coop. 24, 25; Barone и др. 1990, Mol. Gen. Genet, 224, 177-182) и ведет к потере устойчивости через несколько поколений (Shidu и Webster, в Plant Parasitic Nematodes. Т. III, 1981, Zuckerman и др. (редакторы) Asad Press, New York, c. 61-87; Turner, 1990, Ann. Appl. Biol. 117, 385-397).
Растительные паразитические нематоды являются облигатными паразитами. Такие нематоды как цист-нематоды и клубеньковые полностью зависят от образования подходящих питательных структур в корнях растений. Эти питательные структуры появляются из отдельных клеток корня. В случае цист-нематод первичная питательная клетка (IFC) развивается в синцитий посредством расщепления клеточных оболочек и гипертрофии. После заражения клубеньковыми нематодами IFC развивается в гигантскую клетку посредством нескольких делений ядра без цитокинеза и становится метаболически очень активной. При образовании питательной структуры молодая нематода становится неподвижной и теряет способность перемещаться к другим местам питания, проявляя свою зависимость от питательной структуры для нематод (NFS).
Ясно, что существует большая потребность в растениях с пониженной восприимчивостью к растительным паразитическим нематодам. Представленные стратегии борьбы с болезнетворными организмами и вредителями включают экспрессию рекомбинантной ДНК, кодирующей продукт, непосредственно взаимодействующий с болезнетворными организмами или вредителями.
EP-A 159884 раскрывает экспрессию в растениях гена, кодирующего инсектицидный токсин Bacillus thuringiensis. Когда белок переваривается насекомым, он присоединяется к рецептору в желудочно-кишечном тракте, вызывая в конце концов смерть насекомого. Взаимодействие токсина и рецептора в насекомом является решающим для токсического действия, этим можно объяснить относительно частые случаи устойчивости к токсину.
EP-A 303426 сообщает, что множество штаммов B. thuringiensis действует на нематоды. Следовательно, неудивительно, что предлагается идентифицировать гены, кодирующие для этих нематицидных (nematicidal) токсинов, и осуществить экспрессию этих генов в растениях для их защиты от нематод (EP-A 352052).
Однако некоторое значение имеет тот факт, что прямое взаимодействие токсина и вредителя склонно к быстрому развитию устойчивости, как сообщается для B. thuringiensis эндотоксина (Peferoen M (1991) Agroindustry High - Tech, с. 5-9.
Следовательно, объектом настоящего изобретения является обеспечение растений с пониженной восприимчивостью к болезнетворным организмам и вредителям, что позволит совершенно избежать проблем приобретения устойчивости.
Настоящее изобретение обеспечивает растения, которые в результате экспрессии рекомбинантной ДНК обладают пониженной восприимчивостью к растительным паразитическим нематодам. В соответствии с предпочтительным вариантом указанная рекомбинантная ДНК вызывает разрушение питательной структуры для нематод. В другом предпочтительном варианте экспрессия указанной рекомбинантной ДНК по крайней мере замедляет образование питательной структуры для нематод. Согласно наиболее предпочтительному варианту указанная рекомбинантная ДНК включает:
1) ген-A, который при экспрессии разрушает питательную структуру для нематод, причем указанный ген-A контролируется промотором-A, который управляет экспрессией по крайней мере в питательной структуре для нематод; и
2) ген-B, который при экспрессии нейтрализует разрушающее действие гена-A, причем указанный ген-B контролируется промотором-B, который управляет экспрессией практически во всех клетках растения, где происходит экспрессия гена-A, при условии, что указанный промотор неэффективен в управлении экспрессией гена-B в питательной структуре для нематод. Предпочтительный ген-A кодирует barnase (барназу), а ген-B кодирует barstar (барстар), при этом промотор-A является постоянным промотором растения, а промотор-B можно получить из CaMV 35S-промотора или rolD-промотора. В других вариантах промотор-A выделяют из растения или он является производным указанного промотора. В соответствии с данным вариантом предпочтительным является промотор-A, который можно получить из Дельта-0.3Tob RB7-5A промотора или усеченного rolC промотора.
В соответствии с другим аспектом изобретения указанная рекомбинантная ДНК включает ген, ингибирующий эндогенный ген, который кодирует белок или полипептидный продукт, существенный для образования или поддержания питательной структуры для нематод. В соответствии с этим аспектом изобретения предпочтительным является ген-A, который продуцирует транскрипт РНК, который является комплементарным к транскрипту эндогенного гена, кодирующего белок или полипептидный продукт, существенный для образования или поддержания питательной структуры для нематод, в частности гена, существенного для жизнеспособности клеток, при этом указанный ген-B кодирует белок или полипептидный продукт, который заменяет функцию белка или полипептида, закодированного указанным эндогенным геном. Указанный нейтрализующий ген предпочтительно можно получать из гетерологичного гена различных видов, например, видов растений, видов животных или микробов, предпочтительны различные виды растений. Особо предпочтительными генами, которые существенны для жизнеспособности клеток, согласно настоящему изобретению являются гены, кодирующие белок или пептид, выбранный из группы, состоящей из АТФ-синтазы, транслокатора адениннуклеотида, транслокатора трикарбоксилата, транслокатора дикарбоксилата, транслокатора 2-оксо-глутарата, цитохрома C, пируваткиназы, глицеральдегид-3P-дегидрогеназы, NADPH-цитохром-p450-редуктазы, комплекса синтазы с жирной кислотой, глицерол-3P-ацилтрансферазы, оксиметилглутарил-CoA-редуктазы, аминоацилтрансферазы, фактора инициирования транскрипции и фактора элонгации транскрипции. Предпочтительными растениями в соответствии с изобретением являются растения из семейства Solanaceae, более предпочтительными из семейства Solanum tuberosum, картофель, еще более предпочтителен картофель, обладающий пониженной восприимчивостью к картофельным клеточным нематодам.
Изобретение также включает растительный материал, такой как цветы, фрукты, листья, пыльцу, семена или клубни, который можно получать из растений в соответствии с изобретением.
Изобретение также обеспечивает способы получения растений с пониженной восприимчивостью к растительным паразитическим нематодам, отличающиеся тем, что включают стадии:
(1) трансформации реципиентной растительной клетки с рекомбинантной ДНК, которая включает (а) ген, который при экспрессии в питательной структуре нематод вызывает ее разрушение, и (b) выбираемый ген-маркер растения;
(2) генерации растения из трансформированной клетки при давлении отбора;
(3) отбора трансформированного растения с пониженной восприимчивостью к растительным паразитическим нематодам.
Изобретение, кроме того, включает способ получения растения с пониженной восприимчивостью к растительным паразитическим нематодам, отличающийся тем, что включает стадии:
(1) трансформации реципиентной растительной клетки с рекомбинантной ДНК, которая включает (а) ген-A, который при экспрессии разрушает питательную структуру для нематод, причем указанный ген-A контролируется промотором-A, который управляет экспрессией по крайней мере в питательной структуре для нематод, и (b) ген-B, который при экспрессии нейтрализует разрушительное действие гена-A, причем указанный ген-B контролируется промотором-B, который управляет экспрессией практически всех клеток растений, в которых происходит экспрессия гена-A, при условии, что указанный промотор неэффективен в управлении экспрессией гена-B в питательной структуре для нематод, и (с) выбираемый ген-маркер растения,
(2) генерирования целого растения из трансформированной клетки под давлением отбора,
(3) идентифицирования трансформированного растения с пониженной восприимчивостью к указанным растительным паразитическим нематодам.
В соответствии с немного отличающимся вариантом обеспечивают способ получения растения с пониженной восприимчивостью к растительным паразитическим нематодам, отличающийся тем, что включает стадии:
(1) трансформации реципиентной растительной клетки рекомбинантной ДНК, которая включает (а) ген-B, который при экспрессии нейтрализует разрушающее действие гена-A, вводимого на стадии (3), причем указанный ген-B находится под контролем промотора-B, управляющего экспрессией практически во всех клетках растения, где происходит экспрессия гена-A, при условии, что указанный промотор неэффективен в управлении экспрессией гена-B в питательной структуре для нематод, и (b) выбираемый маркер растения,
(2) генерации целого растения из трансформированной клетки при давлении селекции,
(3) трансформации реципиентной клетки растения (полученного на стадии (2), или его потомства), которая содержит по крайней мере ген-B, за счет рекомбинантной ДНК, которая включает (с) ген-A, разрушающий при экспрессии питательную структуру для нематод, причем указанный ген-A находится под контролем промотора-A, управляющего экспрессией, по крайней мере, в питательной структуре для нематод корней зараженного растения,
(4) генерации растения из клетки, трансформированной на стадии (3) при подходящем давлении селекции,
(5) идентификации растения, обладающего пониженной восприимчивостью к растительным паразитическим нематодам.
Предпочтительным способом трансформации реципиентной клетки растения является совместная инкубация указанной клетки со штаммом Agrobacterium, который содержит указанную рекомбинантную ДНК.
Кроме того, изобретение обеспечивает рекомбинантную ДНК, содержащую растительный экспрессивный ген, который последовательно включает: промотор-A, управляющий экспрессией гена в питательной структуре для нематод; ген-A, который при экспрессии в питательной структуре для нематод разрушает эту структуру; и обычно терминатор транскрипции и сигнальную последовательность полиаденилирования; связанные таким образом, что экспрессия указанного гена-A осуществляется в указанной питательной структуре для нематод. Согласно изобретению в предпочтительной рекомбинантной ДНК указанный промотор можно получить из Дельта-0.3TobRВ7-5A или усеченного rolC промотора. Согласно изобретению в рекомбинантной ДНК предпочтителен также ген-A, который можно получить из Barnase гена Bacillus amyloliquefaciens.
В соответствии с другим предпочтительным вариантом обеспечивают рекомбинантную ДНК, в которой указанный ген-A можно получить из растения-хозяина и выбрать из группы, включающей гены, кодирующие АТФ-синтазу, транслокатор адениннуклеотида, транслокатор трикарбоксилата, транслокатор дикарбоксилата, транслокатор 2-оксоглутарата, цитохром C, пируваткиназу, глицеральдегид-3P-дегидрогеназу, NADPH-цитохром-p450-редуктазу, комплексы синтазы и жирных кислот, глицерол-3P-ацилтрансферазу, оксиметил-глутарил-CoA-редуктазу, аминоацилтрансферазу, фактор инициирования транскрипции и фактор элонгации транскрипции; при этом указанный ген встроен в указанный промотор с обратной ориентацией относительно природной ориентации в растении-хозяине.
Кроме этого, изобретение обеспечивает вектор трансформации растения, содержащий рекомбинантную ДНК, соответствующую изобретению, а также штаммы Agrobacterium, содержащие указанные векторы трансформации растений.
Особо предпочтительные векторы для использования в способе согласно изобретению можно выбрать из группы, состоящей из pMOG716, pMOG719, pMOG589, pMOG699, pMOG717, pMOG711, pMOG712, pMOG713, pMOG714, pMOG715, pMOG718 и pMOG720 и их производных, модифицированных несущественным для изобретения способом.
Кроме того, изобретение обеспечивает способы снижения ущерба урожаю, наносимого растительными паразитическими нематодами при росте растений согласно изобретению.
В соответствии с другим аспектом изобретения обеспечивают способ снижения ущерба урожаю, наносимого растительными паразитическими нематодами, отличающийся тем, что включает рост растения, содержащего ген гербицидной устойчивости, контролируемый промотором-B, экспрессия которого осуществляется практически во всех клетках корней растения, где происходит экспрессия гена-A, при условии, что он неэффективен в управлении экспрессии в питательной структуре для нематод (NFS); указанный способ включает стадии:
1) роста указанного растения;
2) контакта корней указанного растения с гербицидом.
Смысл использованных здесь экспрессий, наряду с применением и преимуществами изобретения будет ясен из следующего далее подробного описания изобретения.
Краткое описание чертежей.
Фиг. 1 (а, б) - схематическое изображение двухкомпонентной стратегии устойчивости, намеченной в этом изобретении.
Фиг. 2 - плазмида pMOG707, промежуточный вектор, сконструированный с целью клонирования.
Фиг. 3 - карта рестрикции фрагмента Ti-плазмиды pTiB6.
Фиг. 4 - промежуточный вектор pMOG579.
Фиг. 5 - бинарный вектор pMOG23.
Фиг. 6 - бинарный вектор pMOG22.
Фиг. 7 - бинарный вектор pMOG25. Плазмида, содержащая 35S-HPT и 35S-GUS между левой и правой границами T-ДНК.
Фиг. 8 - бинарные векторы pMOG452, pMOG630 и pMOG679.
Эти плазмиды являются производными от pMOG23 или pMOG22 и содержат либо GUS-конструкцию без промотора, либо GUS-конструкцию с rolD-промотором, либо GUS-конструкцию с усеченным rolD-промотором, и выбираемый маркер растения (NPT11 или HPT).
Фиг. 9 - бинарный вектор pMOG583. Эта плазмида является производной от pMOG22 и содержит 35S-промотор, встроенный в Barstar ген.
Фиг. 10 - бинарный вектор pMOG716. Эта плазмида является производной от pMOG22 и содержит 35S-промотор, встроенный в Barstar ген.
Фиг. 11 - бинарный вектор pMOG718. Эта плазмида является производной от pMOG22 и содержит два Barstar гена, один регулируется 35S-промотором, а другой регулируется rolD-промотором.
Фиг. 12 - плазмида pMOG587. Промежуточный вектор, полученный для инсерции Barnase гена и содержащий Barstar ген с бактериальным промотором и nos-терминатором.
Фиг. 13 - бинарные векторы pMOG589, pMOG591, pMOG717 и pMOG699. Эти плазмиды являются производными от pMOG23 и содержат либо Barnase ген без промотора сразу после правой границы (pMOG589), либо сайт мультиклонирования для инсерции набора регуляторных последовательностей различных типов промоторов-А (pMOG591), либо усеченный rolC-промотор и Barnase ген (pMOG717), либо усеченный Дельта-0.3TobRB7 промотор и Barnase ген (pMOG699). Для предотвращения возможных вредных эффектов Barnase гена во время процедур клонирования в бактерии с внешней стороны границы Т-ДНК можно либо вставить Barstar ген с бактериальным промотором, либо можно вставить интрон в кодирующую область Barnase гена. В последнем случае к номерам pMOG добавляют i (например, pMOG699i).
Фиг. 14 - бинарные векторы pMOG711 и pMOG715. Эти плазмиды являются производными от pMOG23 и содержат усеченный Дельта-0.3TobRB7 промотор и антисмысловую конструкцию гена, существенного для жизнеспособности клетки.
Фиг. 15 - бинарный вектор pMOG719, является общей иллюстрацией плазмиды, которую можно использовать для введения гетерологичного существенного гена, регулируемого промотором типа B.
Фиг. 16 - бинарный вектор pMOG720, является общей иллюстрацией плазмиды, которая содержит полную антисмысловую/смысловую двухкомпонентную систему для экспрессии в растениях с целью получения устойчивости к нематодам.
Было обнаружено, что растения можно сделать восприимчивыми к растительным паразитическим нематодам путем разрушения питательной структуры для нематод через экспрессию Barnase гена под контролем промотора, активного в питательной структуре для нематод (NFS), при этом было показано, что потенциальная активность по разрушению клеток растения вне NFS значительно ослаблена конкурентной экспрессией Barstar под контролем CaMV 35S-промотора. Эти растения не имеют неудобств, связанных с тем, что растительные паразитические нематоды могут оказаться устойчивыми относительно токсического вещества, примененного для защиты, так как прямое взаимодействие патогенного организма или вредителя с токсином не играет роли в защите растения.
Следующие удивительные находки иллюстрируют, как можно использовать изобретение различными способами. Когда CaMV 35S-промотор (Guilley и др., 1982, Cell 30, 763-773), который в здоровых растениях вызывает высокую степень экспрессии GUS-гена в клетках корней и, главным образом, сосудистом цилиндре (Benfey и др. (1990) EMBO J, 9, 1677-1684), используют для управления экспрессией GUS-гена в зараженных нематодами растениях, наблюдают заметное отсутствие GUS-активности в питательной структуре для нематод (NFS). Это было проверено на трех видах растений: Arabidopsis, табак и картофель, зараженных представителями видов растительных паразитических нематод (PPN) (H. schachtii, M. incognita, G. tabacum, G. rostochiensis соответственно), и в которых образовалась NFS. Другими промоторами, которые вызывают высокую степень экспрессии в ткани корней (Leach и Aoyagi, 1991, Plant Sci. 79, 69-76), включая клетки эпидермиса и корневые волоски, являются последовательности rolC и rolD промоторов плазмиды pRi A4 из Agrobacterium rhizogenes (5' фланкирующий участок открытых рамок считывания 12 и 15 соответственно в Slightom и др. (1986, J. Biol. Chem. 261, 108-121). С этими промоторами наблюдают также удивительно сильное подавление GUS-активности в NFS после заражения PPN.
В противоположность приведенным выше данным усеченная версия rolC-промотора, производного нуклеотидной последовательности 11286 до 12132 между открытыми рамками считывания 11 и 12 TL-ДНК из Agrobacterium rhizogenes плазмиды pRi A4 (Slightom и др. 1986, J. Biol. Chem. 261, 108-121) остается активной в NFS, как наблюдали после слияния усеченного rolC-промотора с GUS-геном и трансформации Arabidopsis с этой конструкцией. Очевидно, что часть промотора чувствительна к регуляции при развитии NFS, и когда эта часть удалена, промотор больше не подавляется в NFS.
Растения трансформируют конструкцией рекомбинантной ДНК на основе бинарного вектора из Agrobacterium, содержащем GUS-ген без промотора, клонированный прямо по соседству с правой границей в Т-ДНК в направлении от правой границы и второго выбираемого маркера. Регенеранты выращивают на избирательных средах до зрелости и позволяют завязаться семенам. Растения, полученные из семян, выращивают и после заражения и образования NFS проверяют корни на GUS-активность. Зарегистрировали несколько неожиданное количество линий растений с GUS-активностью в NFS. Однако, не обнаружили линий растений, обладающих GUS-активностью только в NFS; все линии клеток растений показывают, по крайней мере, некоторую GUS-активность в других (не NFS) частях растений. Кроме того, большинство линий растений с GUS-конструкцией без промотора, проявляющих GUS-активность в сосудистом цилиндре здоровых корней, также демонстрируют регуляцию GUS-активности при развитии NFS.
Авторы делают вывод, что с разумной частотой можно получать линии растений, которые дают ген без промотора в NFS зараженного растения, используя подход, проиллюстрированный GUS-геном без промотора. Из этих экспериментов также можно сделать вывод, что промоторы, действительно специфические для NFS, встречаются по крайней мере очень редко. Кроме того, недостаток GUS-ферментативной активности в NFS неизбежно вызывается понижающей регуляцией CaMV 35S-промотора в NFS.
Приведенные выше данные, например, о том, что промотор, активный в NFS, можно с разумной частотой прикреплять, используя конструкцию гена без промотора, и тот факт, что, как было показано, 35S промотор подвергается понижающей регуляции в NFS, приводят к выводу, что можно использовать особенности мозаичной экспрессии для получения растений с пониженной восприимчивостью к растительным паразитическим нематодам (PPN), применяя приведенный ниже подход.
Стадия 1.
Растения трансформируют за счет рекомбинантной ДНК, которая включает (а) ген-B, кодирующий нейтрализующее вещество как Barstar, встроенный после промотора-B, в основном, постоянного, но подвергающегося понижающей регуляции при развитии питательной структуры, и (b) выбираемый маркер растения. Регенеранты на селекционных средах можно отобрать для экспрессии гена-B, используя, например, способ Вестерн-детектирования. Растения, дающие экспрессию гена-B, выращивают до зрелости и дают завязаться семенам, из которых можно получить второе поколение (Т2).
Стадия 2.
Поколение Т2 используют для второго этапа трансформации теперь за счет рекомбинантной ДНК, которая включает ген-A без промотора, кодирующий разрушающее вещество, такое как Barnase, или ген, ингибирующий ген, существенный для жизнеспособности клеток в соответствии с изобретением, клонированный прямо по соседству с правой границей в T-ДНК в направлении от правой границы, и второй селектируемый маркер. (Селектируемый маркерный ген растения может отличаться от гена, используемого на первом этапе трансформации, в зависимости от того, присутствует ли еще этот ген или функционирует в Т2. Способы удаления или инактивации селектируемых маркеров растений известны специалистам (например, как раскрыто в WO 92/01370). Регенеранты выращивают на селекционной среде до зрелости и позволяют завязаться семенам, от которых выращивают следующее поколение растений (Т3).
Стадия 3.
Отбирают следующее поколение (Т3) для снижения восприимчивости к PPN и отсутствия роста и развития отклоняющихся форм. Вследствие случайности интеграции второй этап трансформации иногда дает интеграцию гена-A без промотора в участки генома растения, способного активно промотировать транскрипцию гена-A в NFS. Отбор после заражения дает в результате растения, обеспечивающие достаточную экспрессию гена-A в питательных структурах, достаточную для предотвращения полного развития этих структур, и которые неспособны поддерживать полный жизненный цикл PPN, при этом, с другой стороны, возможная экспрессия гена-A в других областях (кроме питательной структуры) нейтрализуется сопутствующей экспрессией гена-B.
Говоря более обобщенно, изобретение обеспечивает способ получения растений, которые демонстрируют пониженную восприимчивость к PPN, отличающийся тем, что включает стадии осуществления интеграции в геном указанного растения 1) гена-A, который при экспрессии разрушает питательную структуру для нематод, причем указанный ген находится под контролем промотора-A, экспрессия которого происходит, по крайней мере, в NFS, и 2) гена-B, который при экспрессии нейтрализует разрушающее действие гена-A, причем указанный ген-B находится под контролем промотора-B, управляющего экспрессией практически во всех клетках растения, но неэффективного в NFS, и стадию 3) селекции растений, демонстрирующих пониженную восприимчивость к PPN.
Преимущества и многочисленные пути осуществления изобретения будут понятны из последующего описания изобретения.
Для изобретения важно отметить, что не существует абсолютных требований для каждого из промоторов (A и B) в отдельности, то есть промотор-B не должен быть активным в каждой клетке растения вне NFS на всех стадиях развития, но должен быть активным по крайней мере в тех клетках растения вне питательной структуры, где промотор-A демонстрирует активность с неполным блокированием функций (leaky activity) в такой степени, что экспрессия разрушающего гена ухудшает жизнеспособность растения в областях вне питательной структуры для нематод. Подобным образом, промотор-A не должен быть совершенно специфическим для NFS до тех пор, пока активность в других клетках растения на любой стадии развития растения нейтрализуется конкурентной активностью продуктов гена-B в этих клетках. Промотор-B даже может проявлять некоторую активность по неполному блокированию функций (leaky activity) в NFS до тех пор, пока экспрессия гена-B достаточно низка, чтобы допустить вредное воздействие продуктов гена-A в NFS.
Подходящий промотор-A, который активен в NFS, но неактивен в возможно большем количестве других тканей, можно идентифицировать, применяя способы, описанные в заявке. После идентификации и выделения такого промотора-A его можно вставить перед разрушающим геном-A и использовать на втором этапе трансформации, как описано в стадии 2. Полученные линии растения, трансгенные и для конструкции промотор-A/ген-A, и для конструкции промотор-B/ген-B, можно проверить на пониженную восприимчивость к PPN, как указано в этом описании.
По-другому, обе составные конструкции (промотор-B/ген-B и промотор-A/ген-A) можно поместить в один вектор и использовать на одной стадии трансформации растения. Регенеранты выращивают на селективной среде до зрелости. Следующее поколение растений можно прямо отобрать для снижения восприимчивости к PPN и отсутствия роста и развития отклоняющихся форм.
В соответствии с другим аспектом изобретения NFS-специфическое подавление конструктивного промотора-B согласно изобретению можно использовать для получения растений, устойчивых к гербицидам или антибиотикам, получая растения, которые локально (например, в NFS) восприимчивы к соединениям с гербицидной или антибиотической активностью. Специалисты могут легко выбрать подходящие гены устойчивости к гербицидам, способные нейтрализовать гербицидное действие и которые встраивают к промотору-B согласно изобретению, эти гены включают гены устойчивости к гербицидам, указанные ниже как выбираемые маркеры, но не ограничены ими.
Подходящие выбираемые маркерные гены, которые можно использовать для отбора или поиска трансформированных клеток, можно выбрать из приведенного далее списка, не ограничивающего области изобретения: гены неомицин-фосфотрансферазы, сообщающие устойчивость к канамицину (EP-B 131623), ген устойчивости к гигромицину (EP 186425 A2), ген глутатион-S-трансферазы из печени крысы, сообщающий устойчивость к гербицидам, производным от глутатиона (EP-A 256223), ген глутаминсинтетазы, сообщающий при чрезмерной экспрессии устойчивость к ингибиторам глутаминсинтетазы, таким как фосфинотрицин (WO 87/05327), ген ацетилтрансферазы из Streptomyces viridochromogenes, сообщающий устойчивость к селективному средству фосфинотрипсину (EP-A 275957), ген, кодирующий 5-енолшикимат-3-фосфатсинтазу (5-enolshikimate-3- phosphate synthase) (EPSPS), сообщающий устойчивость к N-фосфонометилглицину, bar ген, сообщающий устойчивость к Bialaphos (биалафосу) (например, WO 91/02071) и подобные. Выбор маркера не является критичным до тех пор, пока он является функциональным (например, селективным) в комбинации с выбранными клетками растения.
Маркерный ген и интересующий ген необязательно должны быть связаны, так как совместная трансформация несвязанных генов (патент США 4399216) является также эффективным процессом в трансформации растений.
Настоящее изобретение также обеспечивает векторы, которые включают рекомбинантную ДНК для устойчивой трансформации растений и содержащие:
(а) отбираемый ген для изучения активности промотора и выделения подходящего промотора-A и/или промотора-B (pMOG452),
(b) подходящие гены нейтрализатора, согласно изобретению контролируемые промотором-B (pMOG583, pMOG716, pMOG719), либо
(с) разрушающие гены, согласно изобретению не имеющие промотора, которые можно использовать для второго этапа трансформации и которые ниже объединяются с подходящим промотором-A, согласно изобретению находящимся в геноме растения, который уже содержит нейтрализующий ген (pMOG589), либо
(d) разрушающие гены, которые согласно изобретению контролируются выделенным промотором-A, соответствующим изобретению (pMOG699, pMOG717, pMOG711, pMOG712, pMOG713, pMOG714, pMOG715), и которые можно использовать для второго этапа трансформации или совместной трансформации с вектором, содержащим нейтрализующий ген под контролем промотора-B согласно изобретению;
(е) подходящие комбинации разрушающего и нейтрализующего генов, в которых согласно изобретению разрушающий ген может не иметь промотора (pMOG718) или быть ниже выделенного промотора-A (pMOG718, pMOG720 и производные); указанные комбинации можно применять для трансформации растений в один этап, и многие из них имеют пониженную устойчивость к растительным паразитическим нематодам.
В контексте данного изобретения термины "вещества, разрушающие NFS" и "нейтрализующие вещества" охватывают серии выбранных соединений, которые кодируются ДНК, генные продукты которых (либо белок, либо РНК, либо антисмысловая РНК) вредны для метаболизма, и/или функционирования, и/или жизнедеятельности NFS или органелл в них, и для которых известно, что нейтрализующие вещества способны подавлять активность разрушающего вещества при одновременной экспрессии в одной клетке с разрушающим веществом. Предпочтительными комбинациями разрушающих и нейтрализующих веществ являются, например, barnase/barstar из Bacillus amyloliquefaciens (Hartley, 1988, J. Mol. Biol. 202, 913-915), рестриктазы/соответствующие метилазы, такие как EcoRI из Е. coli (Green и другие, 1981, J. Biol. Chem. 256, 2143-2153) и EcoRI-метилаза или похожие комбинации, как описано в обзоре модификационных систем для рестрикции типа II (Wilson, 1991, Nucl. Acid. Res. 19, 2539-2566), бактериоцины и соответствующие иммунные протеины, например колицин E3/иммунный протеин из E.coli (Lau и др., 1985, Nucl. Acid. Res. 12, 8733-8745) или любой ген, кодирующий разрушающее вещество, который можно нейтрализовать одновременным продуцированием противосмысловой РНК под контролем промотора-B, такой как последовательности ДНК, кодирующие цепь A токсина дифтерии (Czako и An. 1991, Plant Physiol., 95, 687-892), РНКазы, такие как РНКаза Т1, рибонуклеазы или протеазы, рибозимы против мРНК, которые кодируют фитотоксические протеины.
В соответствии с другим аспектом изобретения комбинации разрушающих и нейтрализующих веществ включают соответственно гены, ингибирующие эндогенный ген, кодирующий белок или полипептидный продукт, существенный для жизнеспособности клетки, и в качестве нейтрализующего гена ген, кодирующий белок или полипептидный продукт, способный выполнять функцию эндогенного белка или полипептидного продукта. Такие разрушающие гены можно выбрать из группы, состоящей из (а) генов, кодирующих рибозимы против транскрипта эндогенной РНК, (b) гены, продуцирующие при транскрипции транскрипты РНК, комплементарные или, по крайней мере, частично комплементарные транскриптам РНК эндогенных генов, существенных для жизнеспособности клетки, способом, известным как антисмысловое ингибирование экспрессии генов (раскрыт в EP-A 240208), или (с) генов, которые при транскрипции продуцируют транскрипты РНК, идентичные или по крайней мере очень похожие на транскрипты эндогенных генов, существенных для жизнеспособности клетки, неизвестным пока способом ингибирования экспрессии генов, относящимся к совместной супрессии (раскрыта Napoli C. и др., 1990, The Plant Cell, 2, 279-289).
В соответствии с предпочтительным вариантом изобретения применяют антисмысловые гены для ингибирования экспрессии эндогенных генов, существенных для жизнеспособности клеток, гены которых участвуют в экспрессии в питательных структурах для нематод, посредством промотора-A, встроенного выше указанного антисмыслового гена.
Разрушающее действие, производимое антисмысловым геном, ингибирующим по отношению к существенному эндогенному гену, нейтрализуют экспрессией нейтрализующего гена, контролируемого промотором-B согласно изобретению, причем указанный ген-B при экспрессии продуцирует белок или полипептидный продукт, идентичный или похожий на белок или полипептид, закодированный жизнеспособным эндогенным геном и способный выполнять функцию продукта эндогенного гена в растении-хозяине. Предпочтительно, чтобы последовательность нуклеотидов транскрипта РНК, закодированного согласно изобретению нейтрализующим геном, отклонялась от транскрипта РНК витального эндогенного гена, избегая возможного эффекта совместной супрессии. Следовательно, предпочтительно, чтобы нейтрализующий ген кодировал белок или полипептид по существу с такой же функцией, как эндогенный существенный ген, но посредством промежуточного продукта транскрипта РНК, который является дивергентным; нейтрализующие гены, наилучшим образом соответствующие этому описанию, можно удобно получить путем отбора из базы данных генов, получаемых из различных видов растений или даже из различных нерастительных видов, таких как дрожжи, эукариоты и прокариоты животных. Предпочтительно, чтобы идентичность нуклеотидных последовательностей транскриптов, закодированных разрушающим антисмысловым трансгеном и нейтрализующим смысловым трансгеном, составляла менее 90%, предпочтительно менее 80%, еще более предпочтительно, чтобы указанный нейтрализующий смысловой трансген кодировал белковый или полипептидный генный продукт, неидентичный по аминокислотной последовательности продукту разрушающего гена, в котором идентичность нуклеотидной последовательности транскриптам, закодированным нейтрализующим трансгеном, составляла менее 75%.
Заданные гены для антисмысловых разрушающих генов выбирают из генов, кодирующих ферменты, существенные для жизнедеятельности клеток, называемые также обязательными ферментами, эти гены должны быть закодированы ядрами, предпочтительно как однокопийные гены, хотя семейство генов небольшого размера должно также подходить для целей изобретения. Кроме того, эффект антисмысловой экспрессии указанных генов не должен сводиться к нулю диффузией или транслокацией от других клеток или органелл продуктов ферментов, обычно синтезированных такими ферментами. Предпочтительно выбирать гены, кодирующие для ферментов мембранных транслокаций, по тому, как они включаются в создание химических градиентов через мембраны органелл. Ингибирование таких белков по определению нельзя аннулировать диффузией субстратов через мембрану, в которой находятся эти белки. Перенесенные соединения не ограничены органическими молекулами, а могут иметь неорганическую природу, например P, H, OH или электроны.
Предпочтительно, чтобы ферменты мембранных транслокаций присутствовали в органеллах, количество которых увеличивается при паразитизме, что иллюстрирует существенную роль таких органелл в клетках, включающих NFS. Специфическими примерами таких органелл являются митохондрии, эндоплазматический ретикулум и плазмодесма (Hussey и др., 1992, Protoplasma, 167, 55-65; Magnusson и Colinowski, 1991, Can. J. Botany, 69, 44-52). Список усеченных ферментов приведен в табл. 1 в виде примеров, не ограничивающих изобретение упомянутыми в таблице ферментами. Более подробный перечень можно составить из таких серий, как Биохимия растений (редакторы Stumpf и Conn, 1988-1991, тома 1 - 16, Academic Press) или Энциклопедия физиологии растений (New Series, 1976, Springer-Verlag, Берлин).
Хотя гены, кодирующие эти ферменты, выделены только в некоторых случаях, и, следовательно, множество копий генов не известно, в этом изобретении описаны необходимые критерии.
Для увеличения до предела антисмысловых эффектов в растении-хозяине предпочтительно использовать гомологичные гены. "Гомологичные" означает получаемые из того же вида растения, что и растение-хозяин. "Гетерологичные" в данном описании будет означать получаемые из другого растения или нерастительных видов. Гетерологичные гены также будут включать синтетические аналоги генов, модифицированные в своих мРНК, кодирующих последовательность нуклеиновых кислот для отклонения от гена-хозяина по крайней мере на 5%. Так как обязательные гены являются в основном высококонсервативными для выделения соответствующего гена из культурного вида, который делают устойчивым, можно использовать гетерологичные зонды другого (растительного) вида. Такие выделения генов хорошо известны специалистам и не требуют чрезмерного экспериментирования с точки зрения настоящего изобретения.
Для дифференциации возможных заданных генов и избранных предпочтительных кандидатов для получения устойчивости к нематодам специалисты могут применять следующую процедуру: через интересующий ген промоторные последовательности можно выделить из геномной ДНК и использовать для клонирования перед маркерным геном, таким как GUS (Jefferson и др., 1987, EMBO J., 6, 3901-3907). Эту конструкцию для экспрессии можно затем поместить в бинарный вектор, подходящий для трансформации растения, активированный в Agrobacterium и перенесенный в геном растения.
Затем регенерированные растения можно заразить PPN и использовать для гистохимического GUS-анализа целых растений и, в частности, питательных структур. В случае, когда последовательность промотора изначально управляет геном, существенным для развития нематод, она не должна быть активной во многих частях растения (для управления обязательным геном), а может быть более активной в питательной структуре, чем в окружающих тканях. Один такой пример опубликован Cramer (1992, Proc. 31-й Ann. Meeting Amer. Soc. Nematologists, Vancouver, Canada) с промотором оксиметилглутарил-CoA-редуктазы (HMGR). Этот промотор становится более активным в зараженной ткани корней, особенно в тех клетках, которые включены в питание нематод. HMGR является ключевым (ограничивающим скорость) ферментом для большого количества таких соединений, как терпены и стеролы. Так как PPN не могут синтезировать свои собственные стеролы (Chitwood и Lusby, 1991, Lipids, 26, 619 - 627), они полностью зависят от питания растений, таким образом, высокий уровень HMGR благоприятен для паразитирующих нематод. Наоборот, понижающая регуляция этого фермента через антисмысловую экспрессию в питательных клетках, как в описании настоящего изобретения, будет оказывать разъединяющее действие на развитие нематод.
По-другому, для селекции предпочтительных кандидатов можно использовать как индикацию наличие мутантов в одноклеточных эукариотах, таких как дрожжи или Chlamydomonas. Если для отдельного фермента присутствует большое количество мутантов, то вероятно, что этот фермент является повторяемым, присутствует как семейства мультикопийных генов, или что существует другой путь метаболизма, чтобы обойти мутированный фермент (Strathern, Jones и Broach (редакторы) 1981. The molecular biology of the yeast Saccharomyces cerevisiae. Cold Spring Harbor Laboratory Press, New York). Такие гены меньше подходят для способа, описанного в этом изобретении. В противоположность этому мутации в ферментах, которые обычно являются летальными для реципиентной клетки и поэтому редко доступны, указывают, что антисмысловая дерегуляция таких генов будет ингибировать правильное развитие этой клетки и может быть использована для попытки получения пониженной восприимчивости к PPN, как раскрыто в этом изобретении. Способы генного разрушения доступны для контроля, если ген является существенным для жизнедеятельности клетки в случае, когда разрушение летально (Rothstein, 1983, Methods Enzym., 101, 202-211). Затем можно выделить гомологичный ген из полученного урожая с геном дрожжей в качестве зонда.
Настоящее изобретение не ограничено растительными видами, представленными примерами. Выбор вида растения в первую очередь определяется величиной ущерба вследствие заражения PPN, наносимого сельскому хозяйству, и подверженностью вида растения трансформации. Роды растений, которым PPN наносят ущерб в практике сельского хозяйства и которые можно сделать значительно менее восприимчивыми к PPN, используя настоящее изобретение, включают, роды, указанные в табл. 2, но не ограничены ими.
Виды нематод, как определено контекстом настоящего изобретения, принадлежат к суперсемейству Heteroderoidea и делятся на семейства Heteroderiae и Meloidogynidae и включают виды, указанные в табл. 2, но не ограничены ими.
Подходящий промотор-A определяют как промотор, который управляет экспрессией нижестоящего гена в NFS на таком уровне, который достаточен для нанесения вреда метаболизму, и/или функционированию, и/или жизнедеятельности NFS, при этом данный промотор предпочтительно (но не обязательно) должен быть неактивным в тканях вне NFS; по крайней мере он никогда не должен быть активным вне NFS на таком уровне, когда активность разрушающего вещества, закодированного геном-A, не могла бы быть нейтрализована в достаточной степени продуктами гена-B.
Подходящий промотор-A можно идентифицировать путем отбора растений, осуществляющих экспрессию гена-B под контролем подходящего промотора-B, причем указанные растения имеют конструкции, регенерированные после второго этапа трансформации за счет рекомбинантной ДНК и несущие конструкции гена-A без промотора и предпочтительно второй селектируемый маркер (отличный от маркера в конструкции промотор-B/ген-B), и анализа указанных растений на пониженную восприимчивость к PPN. Статистическая интеграция конструкции гена-A без промотора на участках с промотирующей активностью после заражения трансформированных регенерантов PPN допускает выбор растений с достаточной экспрессией гена-A в NFS при сохранении нормального фенотипа во всех других частях растения в результате нейтрализующего действия гена-B в тканях вне NFS. Как показано экспериментами, в принципе, нет необходимости выделять промотор-A со свойствами, описанными в изобретении, так как процедура интеграции работает относительно часто.
Этот способ, как показано в другом контексте Kertbundit и др. 1991, Proc. Nat. Acad. Sci. USA. 88, 5212-5216, является особенно подходящим для Arabidopsis растений, так как небольшой геном дает относительно высокую частоту интеграции на участках генома, активных в смысле транскрипции.
По-другому, подходящий промотор-A можно выделить через гены с повышенным уровнем экспрессии в NFS при заражении нематодами. Такие гены можно выделить путем дифференциального скрининга клонов кДНК, полученных из мРНК, экстрагированной из зараженных и здоровых корней, как показано для картофеля (Gurr S.J. и др. 1991, Mol. Gen. Genet., 226, 361-366). Хотя такие промоторы никогда не были подробно описаны, их можно выбрать и выделить из растения хорошо известным способом, который включает:
1) поиск мРНК, которая присутствует, главным образом (хотя не обязательно исключительно) в зараженной ткани корня,
2) выделение этой мРНК,
3) получение кДНК из этой мРНК,
4) использование этой кДНК в качестве зонда для идентификации участков в геноме растения, которые содержат ДНК, кодирующую для этой специфической мРНК,
5) идентификацию и выделение стоящих выше (5') последовательностей из ДНК, кодирующей эту специфическую мРНК и содержащую промотирный участок.
Предпочтительно чтобы использованные для выделения мРНК зараженные корни были бы обогащены NFS, например, путем синхронного заражения (Hammond-Kosack и др. 1989, Physiol. mol. Plant. Pathol., 35, 495-506) или посредством прямого выделения питательных структур из растений, в которых NFS является видимой при небольшом увеличении. Например, питательные структуры, которые развиваются в корнях Arabidopsis можно увидеть при небольшом увеличении и легко выделить с минимумом примесных клеток (Sijmons и др. 1991, Plant J. 1, 245-254). Это делает возможным выделение генов, соответствующих этим РНК, предпочтительно используя процедуры молекулярного обогащения (Dickinson и др. , 1991, Adv. Mol. Gen. Plant - Microbe Interact. 1, 276-279), и последующее выделение расположенных выше промотирных элементов. Идентифицированные однажды подобные гены можно отделить от других видов растений, затем идентифицированные гены используют в качестве зонда, как на стадии 4. Затем из этих других видов растений можно выделить специфические для вида последовательности, стоящие выше, для использования в стратегии, аналогичной описанной в этом изобретении. Стоящие выше по цепи последовательности идентифицированных клонов геномов можно вставить в ген-A для включения в подходящий вектор экспрессии (такой как pMOG22 или pMOG23) для трансформации растения.
По-другому, подходящие промоторы для экспрессии гена-A можно выделить посредством введения метки (Topping и др., 1991, Developm. 112, 1009-1019). При этом подходе регенерируют большое количество различных трансгенных растений после трансформации за счет T-ДНК из Agrobacterium, несущих GUS- конструкции без промотора, такие как описаны Topping и др. (1991, Developm. 112, 1009-1019) или pMOG452, как описанные в примерах. После заражения клубеньковыми или цист-нематодами и некоторого развития NFS корни можно окрасить для определения GUS-активности. Статистическая интеграция T-ДНК делает возможной идентификацию последовательностей промоторов, которые в первую очередь активны в NFS или, например, являются специфическими корневыми, но остаются активными в NFS. Этот тип мечения последовательностей промоторов особенно подходит для Arabidopsis (Kertbundit и др., 1991, Proc. Nat. Acad. Sci. USA 88, 5212-5216) и табака (Topping и др., Developm. 112, 1009-1019). Стоящие выше по цепи 5' последовательности, ответственные за GUS-экспрессию, можно выделить за счет инвертированной полимеразной цепной реакции (инвертированная PCR) (Does и др., 1991, Plant. Mol. Biol. 17, 151-153). Идентифицированные однажды подходящие регуляторные последовательности или гены, транскрипция которых происходит в NFS, можно использовать в качестве зондов для выделения гомологичных последовательностей из других видов растений. Эти последовательности из других видов можно по очереди встроить к гену-A для размещения в подходящем векторе для трансформации растений.
По-другому, подходящий промотор-A получают из усеченной версии rolC-промотора, производного нуклеотидной последовательности от 11286 до 12132 между открытыми рамками считывания 11 и 12 T-ДНК из Agrobacterium rhizogenes плазмиды pRi А4 (Slighton и др., 1986, J. Biol. Chem. 261, 108-121). Аналогично, промоторы, идентифицированные посредством описанного выше способа мечения, которые демонстрируют специфичность по отношению к сосудистой ткани корней, но подавляются при развитии NFS, можно сделать нечувствительными к NFS подавлению путем удаления тех частей промотора, которые ответственны за понижающую регуляцию, как показано на примере усеченной версии rolC-промотора. Такие мутированные промотирующие последовательности затем становятся подходящими для использования в качестве промотора-A.
По-другому, в качестве промотора-A можно использовать следующую последовательность промотора: усеченную версию специфического относительно корней табака промотора Δ 0.3TobRB7 (Yamamoto и др., 1991, Plant Cell 3, 371-382). Полная последовательность промотора TobRB7 высоко активна в NFS, и эта активность становится более специфической для NFS, когда используют усеченную Δ 0.3 версию промотора (Taylor и др. 1992, Proc. 31 Ann. Meeting Amer. Soc. Nematologists, Ванкувер, Канада).
По-другому, промоторы, активные в сосудистых тканях корней или других частях растений и становящиеся более активными именно в NFS (например, промотор окси-метил-глутарил-CoA-редуктазы; Gramer, 1992, Proc. 31 Ann. Meeting-Amer. Soc. Nematologists, Ванкувер, Канада) можно также использовать как промотор-A до тех пор, пока активность вне NFS не будет выше, чем активность промотора-B.
Подходящий промотор-B определяют как промотор, который управляет экспрессией практически во всех клетках, где происходит экспрессия гена-A при условии, что он не управляет экспрессией в питательной структуре для нематод или делает это неэффективно. (Выражение "практически во всех клетках" означает по крайней мере те клетки, которые должны быть жизнеспособны для нормального роста или развития растений, необходимого для коммерческого использования таких растений. В качестве иллюстрации растений, в которых разрушающее действие не нейтрализуют именно во всех клетках растения-хозяина и которые, однако, жизнеспособны и подходят для коммерческого использования, приводят растения, которые экспрессируют разрушающий ген согласно данному изобретению в клетках тычинок; в результате это может дать растения с мужской стерильностью, что в некоторых случаях считается коммерчески привлекательной чертой). Последовательности промотора являются регуляторными последовательностями, активными в управлении транскрипцией в растениях и которые можно получить из растений или вирусов растений или синтезировать химическими способами. Регуляторные последовательности могут также включать последовательности энхансеров, такие как обнаружены в CaMV 35S-промоторе (Kay и др. , 1987, Science 236, 1299-1302), и мРНК стабилизирующие последовательности, такие как лидерная последовательность Альфа-мозаичного вируса RNA4 (Brederode и др., 1980, Nucl. Acids Res. 8, 2213-2223) или любые другие последовательности, функционирующие подобным образом.
По-другому, чтобы обеспечить экспрессию во всех или практически во всех тканях растения, конструкцию промотор-B/ген-B можно дополнить второй конструкцией промотор-B'/ген-B, имеющей характер экспрессии, комплементарный с промотор-B/ген-B, при условии, что ни промотор-B, ни промотор-B' не управляют экспрессией в NFS. В область настоящего изобретения включены также гибридные промоторы, включающие различные промоторы (их части), объединенные таким образом, чтобы обеспечить необходимый характер экспрессии, определенный в данном изобретении.
Предпочтительно, чтобы промотор был промотором-35S мозаичного вируса цветной капусты или его производными, которые, как считают, обычно являются сильными образующими цепь промоторами в тканях растений (Odell и др. 1985, Nature 313, 810-812). Другим предпочтительным примером промотора-B является сильный корневой промотор rolD (Leach и Aoyagi) 1991, Plant Sci. 79, 69-76) из плазмиды pRiA4 Agrobacterium rhizogenes; 5' фланкирующий участок ORF15 (Slightom и др. 1986, J. Biol. Chem. 261, 108-121). Пригодность других образующих промоторов, таких как промотор нопалинсинтазы (Bevan, 1984, Nucl. Acids Res. 12, 8711-8721) или промотор мозаичного вируса норичника шишковатого (figwort) (EP-A 426641), для использования в качестве промотора-B можно проверить посредством присоединения к маркерным генам, таким как GUS (Jefferson, 1987, Plant Mol. Biol. Reporter, 5, 387-405), переносом этих конструкций в растения и гистохимическим анализом таких трансгенных растений после заражения PPN.
Другие регуляторные последовательности, такие как терминаторные последовательности или сигнальные последовательности полиаденилирования, включают любые последовательности, работающие в растении определенным образом, выбор таких последовательностей входит в компетенцию специалиста. Примером такой последовательности является 3' фланкирующий участок гена нопалинсинтазы (nos) Agrobacterium tumefaciens (Bevan, 1984, Nucl. Acids Res. 12, 8711-8721).
В соответствии с другим вариантом изобретения обеспечивают способ уменьшения ущерба урожаю, наносимого растительными паразитическими нематодами, который включает выращивание растений, которые содержат ген устойчивости к гербициду, помещенный под контроль промотора-B, экспрессия которого происходит по крайней мере в корнях растений, но неэффективна в питательной структуре для нематод; указанный способ включает стадии:
1) выращивания указанных растений,
2) взаимодействия корней указанных растений с гербицидом.
В этом варианте изобретения более строгими являются требования к экспрессии гена-B, то есть его экспрессия должна происходить во всех клетках, контактирующих с гербицидом, за исключением NFS. С этой целью можно дополнить промотор-B вторым образующим цепь промотором-B' (связанным с другой или той же копией гена-B), при этом в NFS также не происходит экспрессии промотора-B'. Подходящие для этого варианта изобретения примеры включают (но не ограничены этим) гербициды, например, класс соединений, называемых сульфонилкарбамидами (Mazur и Falcoо, 1989, Ann. Rev. Plant Physiol. 40, 441-470). Пример, использованный в этом изобретении (ген устойчивости к гигромицину (hygromicin), контролируемый 35S-промотором с последующим ростом и заражением в гигромицин-содержащей среде), приведен только для демонстрации возможности этого подхода. Выбор гербицида для сельскохозяйственного применения в первую очередь определяется доступностью гена устойчивости к гербициду, стоимостью и действием гербицида на окружающую среду, его пригодностью для составления композиций либо для введения в почву, либо для распыления, мобильностью в почве и способностью достигать и воздействовать на клетки NFS, мобильностью в тканях растения и способностью достигать NFS-клеток при распылении в воздухе.
Для некоторых видов нематод существует большой выбор носителей, и они могут паразитировать на большом количестве различных видов растений, а также вследствие высокой степени схожести питательных структур в различных видах растений полагают, что понижающая регуляция (то есть, отсутствие активности) таких промоторов, как CaMV 35S или другие промоторы соответствующие описанию образующих цепи промоторов, как описано в настоящем изобретении, будет иметь место в широком наборе видов растений, помимо тех, которые приведены в примерах этого изобретения. Изобретение не ограничено видами, указанными в примерах. Выбор видов растений в первую очередь определяется величиной оцененного ущерба вследствие заражения PPN и подверженностью видов растений трансформации. Роды растений, подверженные при сельскохозяйственном выращивании заражению PPN и которые можно получить значительно менее восприимчивыми к PPN при помощи настоящего изобретения, включают (но не ограничены) роды, указанные в табл. 2.
Виды нематод, как определено в контексте настоящего изобретения, включают все растительные паразитические нематоды, которые модифицируют клетки растения-хозяина в специально адаптированные питательные структуры и простираются от мигрирующих эктопаразитов (например, Xiphinema spp.) до более развитых оседлых эндопаразитов (например, Heteroderidae, Meloidogynae или Rotylen-Chulinae). Перечень паразитических нематод приведен в табл. 2, но изобретение не ограничено упомянутыми в этой таблице видами. Более подробные перечни присутствуют в Zuckerman и др. (ред., в Plant Parasitic Nematodes, т. 1, 1971, New York стр. 139-162).
Способы изобретения борьбы против ущерба урожаю вследствие заражения нематодами применимы также к нематодным вредителям и патогенным организмам всегда, когда указанный патогенный организм или вредитель локально понижающе регулирует промоторы растения на месте заражения (например, в вызванной грибком гаустории или в вызванном тлей злокачественном образовании). Принцип влияния продукции нейтрализующего вещества во всех или в большинстве частей незараженных растений по нейтрализации вещества, разрушающего клетки, на производство которого оказывается влияние, по крайней мере, в месте заражения, не зависит от типа или видов патогенных организмов или вредителей.
В контексте этого изобретения сказано, что растения демонстрируют пониженную восприимчивость к PPN, если можно наблюдать существенное статистическое снижение количества зрелых женских особей, развивающихся на поверхности корней растения по сравнению с контрольными растениями. Классификация восприимчивости/устойчивости в соответствии с количеством зрелых женских особей является стандартной практикой и для цист-нематод, и для клубеньковых нематод (например, La Mondia, 1991, Plant Disease 75, 453-454; Omwega и др., 1990, Phytopathol. 80, 745 - 748).
Основной принцип снижения восприимчивости растения к растительным паразитическим нематодам согласно изобретению - это манипуляции с питательной структурой для нематод. В первую очередь это манипуляции питательной структурой для нематод с той целью, чтобы это описание изобретения включало предупреждение или замедление образования NFS, так же как разрушение образовавшейся NFS.
Предпочтительно предупреждение или замедление образования NFS, то есть, на первых стадиях заражения нематодами; для этого ген-A, разрушающий NFS, должен контролироваться промотором-A, который управляет экспрессией сразу, когда начинает образовываться NFS.
Однако, в принципе, приемлемо также, если разрушающий ген-A находится под контролем промотора-A, который управляет экспрессией разрушающего гена-A на более ранней стадии образования NFS, вызывая ухудшение NFS или разрушение. Оба эти крайних случая сообщат зараженному растению пониженную восприимчивость к паразитическим нематодам. Выражение "разрушение Г" в данном изобретении включает замедление образования NFS, ухудшение образования уже образованной NFS или находящейся в процессе образования, так же как общее разрушение образованной NFS.
Пониженная восприимчивость к растительным паразитическим нематодам может быть результатом уменьшения количества NFS корней зараженного растения, снижения скорости образования NFS или комбинации обоих эффектов.
Согласно настоящему изобретению питательная структура для нематод включает начальную питательную клетку, под этим имеют в виду клетку или очень ограниченное количество клеток, предназначенных для образования питательной структуры для нематод при вовлечении паразитических нематод.
Согласно изобретению эффект разрушения NFS не ограничен только эффектами, вредящими NFS; предполагают также, что разрушающие эффекты, кроме того, оказывают вредное воздействие на развитие нематод путем прямого взаимодействия.
Существует несколько способов для введения в растения-хозяева рекомбинантной ДНК, содержащей последовательности ДНК, как описано в настоящем изобретении. Такие способы включают (но не ограничены этим) трансформацию протопластов с применением метода с использованием кальция/полиэтиленгликоля, электропорацию и микроинъекции или бомбардировку (покрытыми) частицами (Potrykus, 1990, Bio/Technol., 8, 535-542).
Вдобавок к этим, так называемым, способам прямой трансформации ДНК широко применимы трансформационные системы, включающие векторы, такие как вирусные векторы (например, из мозаичного вируса цветной капусты (CaMV) и бактериальные векторы (например, из гена Agrobacterium) (Potrylus, 1990, Bio/Technol, 8, 535-542). После отбора и/или скрининга трансформированные протопласты, клетки или части растений можно регенерировать в целые растения, используя способы, известные специалистам (Horsch и др., 1985, Science 225, 1229-1231). Выбор способов трансформации и/или регенерации несущественен для этого изобретения.
В соответствии с предпочтительным вариантом настоящего изобретения применяют так называемую систему бинарного вектора (раскрыта в Европейском патенте EP-A 120516), в которой используют штаммы Agrobacterium, которые содержат вспомогательную плазмиду с генами вирулентности и совместимую плазмиду, осуществляют перенос бинарного вектора, содержащего генную конструкцию. Этот вектор может осуществлять репликацию и в E.coli и в Agrobacterium; использованный здесь вектор является производным от бинарного вектора Bin 19 (Bevan, 1984, Nucl. Acids Res. 12, 8711-8721). Бинарные векторы, такие как использованы в этом примере, содержат между левой граничной и правой граничной последовательностями T-ДНК идентичный NPT11-ген, кодирующий устойчивость к канамицину (Bevan, 1984, Nucl. Acids Res. 12, 8711 - 8721), и сайт мультиклонирования для клонирования в требуемых генных конструкциях.
Трансформация и регенерация однодольных растений не является стандартной процедурой. Однако последние научные достижения показывают, что, в принципе, однодольные поддаются трансформации, и что плодоносящие трансгенные растения можно регенерировать из трансформированных клеток. Развитие репродуцируемых систем культур тканей для этих растений вместе с эффективными способами введения генетического материала в клетки растения способствуют трансформации. В настоящее время предпочтительными способами трансформации монодольных является бомбардировка микроснарядами эксплантатов или клеток суспензии и прямое поглощение ДНК или электропорация (Shimamoto и др., 1989, Nature 338, 274-276). Трансгенные растения маиса получают введением Streptomyces hygroscopicus bar гена, который кодирует фосфинотрицинацетилтрансферазу (фермент, который дезактивирует гербицид фосфинотрицин), в эмбриогенные клетки культуры суспензии маиса путем бомбардировки микрочастицами (Gordoh - Kamm, 1990, Plant Cell 2, 603-618). Введение генетического материала в протопласты алейронов других однодольных культур, таких как пшеница и ячмень, описано в (Lee, 1989, Plant Mol. Biol. 13, 21-30). Растения пшеницы регенерируют из культуры эмбриогенной суспензии путем отбора только старых плотных и узелковых эмбриогенных каллюсных тканей для образования культур эмбриогенных суспензий (Vasil, 1990 Bio/Technol. 8, 429-434). Комбинация с трансформационными системами для этих культур делает возможным применение настоящего изобретения к монодольным. Эти способы можно также применять для трансформации и регенерации двудольных.
Следующие примеры приведены только с целью иллюстрации и не призваны ограничить область этого изобретения. Если не указано отдельно, то все процедуры по манипулированию рекомбинантной ДНК в примерах осуществляли, используя стандартные способы, как описано в Sambrook и др. (Molecular Cloning, A Laboratory Manual, 2-ое издание, Cold Sopring Harbor Laboratory (1990)).
Пример I. Конструкция промежуточных векторов
а) Конструкция pMOG18
Детальное описание конструкции вектора экспрессии pMOG18 дано Pen и др. (1991), Eur. Pat. Appl. 0449375 A2. Это конструкция промотора 35S мозаичного вируса цветной капусты (CaMV) с двойной усиливающей транскрипцию последовательностью, лидерной последовательностью РНК4 из Alfalfa мозаичного вируса (AIMV), геном, кодирующим β-глюкуронидазу (из плазмиды pPAJ 275), (Jefferson, 1987, Plant Mol. Biol. Reporter 5, 387-405), и следующим терминатором транскрипции нопалинсинтазы (nos). Полная конструкция для экспрессии представлена в виде EcoRI/HindIII фрагмента в pMOG18.
b) Инструкция pMOG180
Вектор экспрессии pMOG180 является производным от pMOG18, где удален ген, кодирующий GUS; между AIMV RN A4 лидерной последовательностью и 3' nos терминатором можно вставить другие гены, как BamHI фрагмент.
Для этой цели синтезируют EcoRI/NcoI фрагмент из pMOG18, содержащей 35S-промотор и AIMV RNA4 лидерные последовательности, путем амплификации PCR, используя в качестве праймера наборы 5' GTTTCTACAGGACGGAGGATCCTGGAAGTATTTGAAAGA 3' (SEQ ID NO:1) и 5' CAGCTATGACCATGATTACG 3' (SEQ ID NO:2), производя, таким образом, мутацию NcoI сайта в BamHI сайт. Затем вектор pMOG18 разрезают EcoRI и BamHI, после чего между этими разрезанными сайтами вставляют вновь синтезированный фрагмент EcoRI/BamHI. Для того чтобы обойти вызванные PCR случайные мутации в последовательностях промоторов, фрагмент EcoRI/EcoRV в синтезированном за счет PCR фрагменте EcoRI/BamHI заменяют последовательностями немутантных типов из pMOG18. Короткий фрагмент EcoRV/BamHI проверяют на мутации, определяя последовательность аминокислотных остатков. П олучаемый вектор экспрессии является плазмидой pMOG180.
с) Конструкция pMOG707.
Конструируют вектор клонирования pMOG707, содержащий на правой границе последовательность T-ДНК, сайт мультиклонирования и терминатор для клонирования различных комбинаций промотор/ген на подходящем фрагменте. Этот вектор конструируют следующим образом: в векторе клонирования pMTL26 (Chambers и др. 1988 Gene 68, 139-149) удаляют сайт XhoI посредством XhoI-расщепления, резко прерываемого Klenow-полимеразой, с последующим выделением, получая pMTL 26/2. Этот модифицированный pMTL-вектор используют для клонирования EcoRI-BgIII фрагмента из pMOG23, содержащей сайт мультиклонирования и последовательности на правой границе, получая в результате pMOG584bis. Полилинкерную последовательность расширяют, вставляя синтетический линкер между сайтами BamHI и XhoI, создавая, таким образом, дополнительные сайты NcoI, XhoI и XbaI. Затем терминатор транскрипции нопалинсинтазы выделяют как фрагмент BamHI/HindIII из плазмиды ROK1 (Baulcombe и др. 1986, Nature 321, 446), сшитый с синтетическим адаптором таким образом, что сайт HindIII не восстанавливают, а вводят сайт EcoRI и затем клонируют в расширенной pMOG584bis как фрагмент BamHI-EcoRI, получая в результате плазмиду pMOG707 (фиг. 2).
Пример II. Конструкция штамма Agrobacterium MOG 101
Используют систему бинарного вектора для переноса генных конструкций в растения Arabidopsis. Вспомогательную плазмиду, содержащую Agrobacterium tumefacience функции вирулентности, получают из октопин-Ti-плазмиды рТiВ6. MOG101 является штаммом Agrobacterium tumefaciens, несущим неоктогенную Ti-плазмиду, из которой делетирован весь T-участок и заменен маркером устойчивости к бактериальному спектиномицину (Spectinomycin) из мигрирующего Tn 1831 (Hooykaas и др., 1980, Plasmid 4, 64-75).
Ti-плазмида pTiB6 содержит два соседних T-участка, TL (T-левый) и TR (T-правый). Для получения производного без TL- и TR-участков конструируют промежуточный вектор pMOG579. Плазмида pMOG579 является производной от pBR322, которая содержит 2 фрагмента Ti-плазмиды, которые расположены слева и справа от T-участков (фиг. 3). 2 Фрагмента (показаны темным) отделены в pMOG579 (фиг. 4) 2,5 тысячами BamHI-HindIII фрагментов от мигрирующего Tn1831 (Hooykaas и др., 1980, Plasmid 4, 64-75), несущего маркер устойчивости к спектиномицину (фиг. 4). Плазмиду вводят в Agrobacterium tumefaciens штамм L ВА1010 [С58-С9 (pTiB6) = удаленный из клетки С58-штамм, в который введена pTiB6 (Koekman и др. 1982, Plasmid 7, 119-132)] путем тройного скрещивания из E.coli, используя НВ101, содержащий pPK2013 в качестве помощника. Для устойчивости к рифампицину (Rifampicin) (20 мг/л) и спектиномицину (250 мг/л) выбирают трансконъюгаты. Двойная рекомбинация между pMOG579 и pTiB6 дает в результате потерю устойчивости к карбенициллину (Carbenicillin) (маркеру pBR322) и отсутствие целого T-участка. Из 5000 устойчивых к спектиномицину трансконъюгатов, копии которых помещали на содержащую карбенициллин (100 мг/л) среду, обнаружены два чувствительных. Саузерн-анализ (southern analysis) показал, что двойное скрещивание при проведении эксперимента делетирован целиком Т-участок (не показан). Полученный штамм назвали MOG101. Этот штамм и его конструкции аналогичны штамму GV 2260 (Deblaere и др. 1985, Nucl. Acids Res. 13, 4777 - 4783).
Пример III. Конструкция бинарных векторов
а) Конструкция pMOG23.
В этом примере описана конструкция бинарного вектора pMOG23 (в E.coli K-12 штамме DH5alpha), внесенного в Centraal Bureau voor Sdimmel-cultures - 29 января 1990 под номером CBS 102.90).
Бинарный вектор pMOG23 является производным вектора Bin 19 (Bevan, 1984, Nucl. Acids Res. 12, 8711-8721). Для получения pMOG23 вектор Bin 19 изменяют способом, несущественным для настоящего изобретения, используя технику, известную специалистам в области молекулярной биологии.
Во-первых, положения левой границы (LB) и правой границы (RB) изменяют относительно гена II неомицинфосфотрансферазы (NPTII ген). Во-вторых, изменяют на обратную ориентацию NPTII гена, что дает транскрипцию в направлении LB. В заключение, полилинкер Bin 19 заменяют полилинкером со следующими сайтами узнавания фермента рестрикции: EcoRI, SmaI, BamHI, XbaI, SacI, XhoI и HindIII (фиг. 5).
b) Конструкция pMOG22.
Бинарный вектор pMOG22 является производным от pMOG23, где NPTII-ген заменен геном устойчивости к гигроцину (HPT, гигромицинфосфотрансфераза из плазмиды PL G90; Van den Elzen и др. 1985, Plant Mol. Biol. 5, 299-302), используя приемы, известные специалистам в области молекулярной биологии, pMOG22 конструкции подробно описана (Cornelissen и др. (1991, Описание Европейского патента 0440304 A1) и внесена в E.Coli K-12 штамм DH5alpha в Centraal Bureau voor Scimmel-cultures 29 января 1990 под номером CBS 101.90 (фиг. 6).
c) Инструкция pMOG25 и pMOG28; бинарные векторы, содержащие 35S-промотор и GUS-ген.
Фрагмент EcoRI/HindIII из pMOG18 клонируют в полилинкер pMOG22 и pMOG23, получая в результате бинарные плазмиды pMOG25 (фиг. 7) и pMOG28 соответственно.
d) Конструкция pMOG452, бинарный вектор, содержащий GUS-ген без промотора.
Ген, кодирующий GUS, пристроенный к 3'nos последовательности терминатора, но без 5' регуляторных промотирующих последовательностей, клонируют как фрагмент EcoRI-BamHI из плазмиды pBI101 (Jefferson, 1987, Plant Mol. Biol. Reporter 5, 387-405) в сайт мультиклонирования бинарного вектора pMOG23, получая бинарную плазмиду pMOG452 (фиг. 8).
е) Конструкция pMOG630; бинарный вектор, содержащий rolD промотор и GUS-ген.
5' Фланкирующий участок (373 пар оснований) rolD плазмиды pRiA4 (Leach и Aoyagi 1991, Plant Sci. 79, 69-76) клонируют перед CUS геном плазмиды pMOG452, используя стандартную технику клонирования, получая в результате бинарную плазмиду pMOG630 (фиг. 8). Описаны очень похожие конструкции для анализа активности rolD-промотора.
f) Конструкция pMOG679; бинарный вектор, содержащий усеченный rolC-промотор и GUS-ген.
Усеченную версию rolC-промотора, выделенного из нуклеотидной последовательности от 11286 до 12132 между открытыми рамками считывания 11 и 12 TL-ДНК из Agrobacterium rhizogenes плазмиды pRiA4 (Slightom и др. 1986, J. Biol. Chem. 261, 108-121), выделяют из плазмиды pRiA4, используя стандартную PCR технику таким образом, что последовательность усеченного rolC-промотора амплифицируют за счет BamHI сайта, созданного с обеих сторон последовательности. Затем этот амплифицированный фрагмент встраивают к GUS-гену плазмиды pMOG452, получая в результате бинарную плазмиду pMOG679 (фиг. 8). Ориентацию усеченного rolC-промотора относительно GUS-гена проверяют при помощи рестрикционного анализа.
g) Конструкция pMOG583; бинарный вектор, содержащий 35S-промотор и Barstar ген.
Barstar ген из Bacillus amyloliquefaciens является производным плазмиды рМТ316 (Hartley, 1988, J. Mol. Biol. 202, 913-915). Плазмиду рМТ316 переводят в линейную форму за счет HindIII с последующей сшивкой в присутствии синтетического адаптора, замещая, таким образом, HindIII сайт рестрикции на сайт рестрикции BamHI. Затем barstar ген выделяют как фрагмент BamHI и клонируют в BamHI сайт плазмиды pMOG180. Затем целую конструкцию для экспрессии клонируют как EcoRI/HindIII фрагмент в сайт мультиклонирования бинарного вектора pMOG22, получая в результате плазмиду pMOG583 (фиг. 9).
h) Конструкция pMOG716, бинарный вектор, содержащий rolD-промотор и Barstar ген.
Barstar ген из Bacillus amyloliquefaciens является производным от плазмиды рМТ316 (Hartley, 1988, J. Mol. Biol. 202, 913-915). Плазмиду рМТ316 переводят в линейную форму за счет HindIII с последующей сшивкой в присутствии синтетического адаптора, замещая, таким образом, HindIII сайт рестрикции на BamHI сайт рестрикции. Затем barstar ген выделяют как BamHI фрагмент и клонируют перед терминатором нопалинсинтазы в BamHI сайт плазмиды pMOG707. Затем клонируют 373bp 5' фланкирующий участок rolD плазмиды pRiA4 (ORF15 в Slightom и др. 1986, J. Biol. Chem. 261, 108-121) перед barstar геном, после чего целую структуру переносят в pMOG22, получая в результате бинарный вектор pMOG716 (фиг. 10).
f) Конструкция pMOG718; бинарный вектор, содержащий два barstar гена, регулируемые 35S и rolD промоторами, соответственно.
Фрагменты 35S-Barstar и rolD-Barstar, как описано в примере IIIg и h, соответственно, также можно использовать для конструирования бинарного вектора, содержащего два barstar гена, каждый из которых регулируется разными промоторами типа B, оба пристроены к терминатору, действующему в растительных клетках. С этой целью фрагменты промотор-Barstar-терминатор могут использовать специалисты для совместного клонирования в pMOG23, получая в результате бинарный вектор pMOG718 (фиг. 11).
j) Конструкция pMOG589; бинарный вектор, содержащий Barnase-ген без промотора.
Из pMOG23 выделяют фрагмент EcoRI-BglII, содержащий сайт мультиклонирования и последовательности на правой границе, и клонируют в pMTL26/2 (пример 1с), получая в результате pMOG584. Затем выделяют nos-терминатор из pMOG18 как фрагмент BamHI - HindIII, присоединенный к синтетическому адаптору таким образом, что сайт HindIII не регенерируют, а вводят сайт EcoRI с последующим клонированием в pMOG584, как фрагмент BamHI-EcoRI, получая pMOG585.
Для преодоления потенциальной активности с блокированием функций (leaky activity) barnase гена в бактериях ген, кодирующий barstar и контролируемый бактериальным промотором, клонируют в бинарные векторы, снаружи от T-ДНК в случае бинарных векторов. Для этой цели плазмиду рМТ316 (Hartley, 1988, J. Mol. Biol. 202, 913-915) переводят в линейную форму при помощи EcoRI и подшивают синтетический адаптор, таким образом, что сайт EcoRI не регенерируют, а вводят сайт HindIII. Затем этот модифицированный фрагмент клонируют как фрагмент HindIII в вектор pMTL 24 (Chambers и др. 1988, Gene 68, 139-149), создавая, таким образом, pMOG586. Сайты XhoI и BamHI, присутствующие на участке tac промотора, удаляют, резко обрывая эти сайты Klenow полимеразой. Последовательность tac/Barstar выделяют как SphI фрагмент и включают в плазмиду pMOG585 в сайт SphI между правой границей и сайтом BglII, получая в результате плазмиду pMOG587 (фиг. 12).
Barnase ген из Bacillus amyloliquefaciens является производным плазмиды рМТ416 (Hartley, 1988, J. Mol. Biol. 202, 913-915). Ген, кодирующий зрелый barnase, усиливают, используя PCR технологию, за счет набора праймеров, который вводит сайт рестрикции XhoI и АТС в 5' сайт barnase и BamHI сайт в 3'сайт. Для этого фрагмента определяют последовательность мономерных звеньев для проверки правильности последовательности, кодирующей barnase, и подшивают в pMOG588. И pMOG23, и pMOG588 разлагаются при участии EcoRI и BglII, и получаемые фрагменты подшивают и переводят в линейную форму снова при помощи BglII. Фрагменты нужного размера подшивают в расщепляемую за счет BglII pMOG23 для образования бинарного вектора pMOG589 (фиг. 13).
По-другому, для преодоления потенциальной активности с блокированием функций (leaky activity) barnase гена в бактериях можно ввести интрон в последовательность, кодирующую barnase, как описано ниже; Barnase ген выделяют из плазмиды рМТ416 путем PCR, используя следующую комбинацию праймера:
5' праймер: 5' CTGCTCGACCTAGGCACAGGTTATCAACACGTTTG 3' (SEQ ID NO:3),
3' праймер: 5' CGGACTCTGGATCCGGAAAGTG 3' (SEQ ID NO:4), вводя, таким образом, оба сайта XhoI и Avr в 5'конец гена и сайт BamHI в 3'конец. Этот фрагмент субклонируют как фрагмент XhoI/BamHI в pMOG707. Второй интрон ST-LS1 из Solanum tuberosum выделяют путем PCR плазмиды p34S GUS INT (Vancanneyt и др. 1990, Mol. Gen. Genet. 220: 245-250), вводя посредством этого сайты AvrII с обоих концов интрона. Этот интрон вставляют как фрагмент AvrII в Barnase ген. Barnase ген, включающий интрон, повторно выделяют из этой плазмиды путем PCR, используя следующие праймеры:
5' праймер: 5' CTTACTCGAGCCATGGTAAGTTTCTGC 3' (SEQ ID NO:5)
3' праймер: 5' CGGACTCTGGATCCGGAAAGTG 3' (SEQ ID NO:6) вводя, таким образом, инициаторный кодон выше от последовательности ST-LS1 интрона. Затем этот фрагмент субклонируют как фрагмент XhoI/BamHI в pMOG707, которая переведена в линейную форму за счет BamHI и частично обрезана за счет XhoI (таким образом, сохраняют сайт XbaI в полилинкерной последовательности, получая в результате pMOG588i (фиг. 13). Обе плазмиды pMOG23 и pMOG588i расщепляются при действии EcoRI и BglII, и получаемые фрагменты подшивают и переводят в линейную форму снова при помощи BglII. Фрагменты нужного размера подшивают в BglII-расщепляемую pMOG23 для образования бинарного вектора pMOG689i (фиг. 13). Обозначение "pMOG---1" используют в этом описании патента для обозначения присутствия интрона в barnase гене.
Выбор между использованием barnase, содержащего интрон, или barstar гена снаружи от границ T-ДНК конструкции бинарных векторов в этом примере не является критичным для этого изобретения.
k) Конструкция pMOG591i; бинарный вектор, содержащий сайт мультиклонирования перед Barnase геном для промотора.
Эту конструкцию используют для введения набора различных регуляторных последовательностей, как идентифицировано в примере XII. Для этой цели синтетический сайт мультиклонирования плазмиды pMOG588i может размещаться между 5' barnase последовательностью и правой границей T-ДНК. Получаемую плазмиду pMOG591i используют для конструкции бинарного вектора pMOG591i (фиг. 13). Этот вектор затем используют для введения регуляторной последовательности промотора-A в транскрипционном слиянии с геном, кодирующим barnase.
l) Конструкция pMOG717i; бинарный вектор, содержащий усеченный rolC-промотор и barnase ген.
Последовательность усеченного rolC-промотора, описанного в примере IIIf, можно выделить из плазмиды pRi A4 (Slightom и др. 1986, J. Biol. Chem. 261, 108-121) и размножить за счет сайта XbaI, созданного на 5' конце и сайта NcoI на 3'конце. Этот фрагмент можно клонировать в pMOG588i, ведущую к транскрипционному слиянию между усеченным rolC-промотором и Barnase геном. Эту плазмиду используют для конструкции бинарного вектора pMOG717i (фиг. 13).
m) Конструкция pMOG699; бинарный вектор, содержащий усеченный промотор (Дельта 0.3TobRB7-5A) и barnase ген.
Последовательность промотора Дельта 0.3TobRB7-5A (Yamamoto и др. 1991, Plant Cell 3: 371-382) выделяют двухстадийной PCR на геномной ДНК, выделенной из табака. В первой PCR реакции часть гена TobRB7-5А выделяют, используя следующие праймеры:
5' праймер: 5' CTCCAAATACTAGCTCAAAACC 3' (SEQ ID NO:7)
3' праймер: 5' CCTCACCATGGTTAGTTCTC 3' (SEQ ID NO:8).
Получаемый PCR продукт используют для выделения фрагмента Дельта0.3TobRB7-5A, применяя следующие праймеры:
5' праймер: 5' CTTGAATTCTAGATAAGCTTATCTAAAT (SEQ ID NO:9)
3' праймер: 5' CCTCACCATGGTTAGTTCTC 3' (SEQ ID NO:10).
Получаемый PCR продукт выделяют из геля, затупляют конец и субклонируют в pUC9 (Vieira и Messing 1982 Gene 19, 259-268), которую затем переводят в линейную форму за счет SmaI. Расщепление получаемой плазмиды при помощи XbaI и частично NcoI дает правильный фрагмент Дельта0.3TobRB7-5A, который можно субклонировать в pMOG588i. Эту плазмиду используют для конструкции бинарного вектора pMOG699i (фиг. 13).
n) Конструкция pMOG711; бинарный вектор, содержащий усеченный промотор (Дельта0.3TobRB7-5A) и антисмысловой ген ATRI NADPH-CytP 450 редуктазы.
Клон NADPH-цитохром р450 редуктазы ATRI (EMBL - регистрационный номер Х66016) выделяют из Arabidopsis thaliana var. Landsberg eresta, используя PCR технику на кДНК, полученную из мРНК этого вида. Для размножения интересующей последовательности используют набор праймеров 5' GGCGGATCGGAGCGGGGAGCTGAAG3' (SEQ ID NO:11) и 5'GATACCATGGATCACCAGACATCTCTG 3' (SEQ ID NO: 12). Он вводит сайт NcoI на N-конец PCR фрагмента. Затем PCR фрагмент расщепляют за счет BamHI-NcoI и клонируют антисмысловым образом перед терминатором нопалинсинтазы в pMOG707. Последовательность усеченного промотора Дельта-0.3TobRB7-5A (Yamamoto и др. 1991, Plant Cell 3, 371-381), выделенную, как описано в примере IIIm, затем можно вставить как фрагмент XbaI-NcoI. Целую последовательность затем клонируют в бинарный вектор pMOG23 после расщепления за счет EcoRI и одного из оставшихся уникальных ферментов рестрикции, получая в результате бинарный вектор pMOG711 (фиг. 14).
о) Конструкция pMOG712; бинарный вектор, содержащий усеченный промотор (Дельта0.3TobRB7-5A) и антисмысловой ген ATR2 NADPH-Cyt P450 редуктазы.
Клон NADPH-цитохром p450 редуктазы ATR2 (EMBL регистрационный номер Х66017) выделяют из Arabidopsis thaliana var. Landsberg erecta, используя PCR технику на кДНК, полученной из мРНК этого вида. Для размножения используют набор праймеров:
5' GGTTCTGGGGATCCAAAACGTGTCGAG (SEQ ID NO:13) и
5' GGCTTCCATGGTTTCGTTACCATACATC 3' (SEQ ID NO:14). Он вводит оба фланкирующих PCR фрагмента BamI и NcoI. Затем PCR фрагмент расщепляют за счет BamI-NcoI и клонируют антисмысловым образом перед терминатором нопалинсинтазы в pMOG707. Последовательность усеченного промотора Дельта0.3TobRB7-5A (Yamamoto и др. 1991, Plant Cell 3, 371-382), выделенную, как описано в примере IIIm, затем можно вставить как фрагмент XbaI-NcoI. Затем целую последовательность клонируют в бинарный вектор pMOG23 после расщепления за счет XhoI и частичного расщепления за счет EcoRI, получая в результате бинарный вектор pMOG712 (фиг. 14).
р) Конструкция pMOG713; бинарный вектор, содержащий усеченный промотор (Дельта0.3TobRB7-5A) и антисмысловой ген глицерол-3-фосфатацилтрансферазы.
Клон глицерол-3-фосфатацилтрансферазы ATS1 (EMBL регистрационный номер D00673) выделяют из Arabidopsis thaliana, используя PCR технику на кДНК, полученной из мРНК этого вида. Для размножения применяют набор праймеров:
5' GCCCGGGATCCGGTTTATCCACTCG 3' (SEQ ID NO:15) и 5' GAGTATTTTCCATGGATTGTGTTTGTG 3' (SEQ ID NO:16). Он вводит SmaI, BamHI и NcoI фланкирующий ATS 1 клон. Затем PCR фрагмент расщепляют за счет SmaI-NcoI и субклонируют в pMOG445. (pMOG445 является производным pUC18, содержащим, благодаря инсерции олигоадаптора в сайт мультиклонирования, сайты экстра-рестрикции ClaI, NcoI и BglII между EcoRI и SstI). Затем ATS 1 клон выделяют после расщепления NcoI и частично BamHI и субклонируют антисмысловым образом перед терминатором нопалинсинтазы в pMOG707. Последовательность усеченного промотора Дельта0.3TobRB7-5A (Yamamoto и др. 1991, Plant Cell 3, 371-382), выделенную, как описано в примере IIIm, затем вставляют как фрагмент XbaI-NcoI. Целую последовательность затем клонируют в бинарный вектор pMOG23 после расщепления за счет EcoRI и одного из остающихся уникальных ферментов рестрикции, получая в результате бинарный вектор pMOG713 (фиг. 14).
g) Конструкция pMOG714; бинарный вектор, содержащий усеченный промотор (Дельта0.3TobRB7-5A) и антисмысловой ген транслокатора адениннуклеотида.
Клон для транслокатора митохондриального адениннуклеотида (PAN T1, EMBL регистрационный номер Х57557; Winning и др. 1992 Plant J. 2, 763-773) выделяют из Solanium tuberosum, используя PCR технику на кДНК, полученной из мРНК этого вида. Для размножения используют набор праймеров:
5'GCTAGCCGGATCCATCTGAGCTCCAG 3' (SEQ ID NO:17) и 5' GACGTCCATGGCTGAATTAGCCACCACCG3' (SEQ ID NO:18). Он вводит оба BamHI и NcoI фланкирующих PAN T1 клон. Затем PCR фрагмент расщепляют за счет BamHI-NcoI и клонируют антисмысловым образом перед терминатором нопалинсинтазы в pMOG707. Последовательность усеченного промотора Дельта0.3TobRB7-5A (Yamamoto и др. 1991, Plant Cell 3, 371-382), выделенную, как описано в примере IIIm, затем вставляют как фрагмент XbaI-NcoI. Целую последовательность затем клонируют в бинарный вектор pMOG23 после расщепления за счет EcoRI и одного из остающихся уникальных сайтов рестрикции, получая в результате бинарный вектор pMOG714 (фиг. 14).
r) Конструкция pMOG715; бинарный вектор, содержащий усеченный промотор (Дельта0.3TobRB7-5A) и антисмысловой ген АТФ-синтазы.
Клон для бета субъединицы АТФ-синтазы (Boutry и Chua 1985 EMBO J, 4., 2159-2165) выделяют из табака (Nicotiana plumbaginifolia), используя PCR технику на кДНК, полученной из мРНК этого вида. Для размножения применяют набор праймеров:
5' CCCTCCAGGATCCCTTCTCGGAGGCTTC 3' (SEQ ID NO:19) и 5' GAAAAGAAAGCCATGGAACTTTATAATC 3' (SEQ ID NO:20). Он вводит оба BamHI и NcoI фланкирующих клон АТФ синтазы. Затем PCR фрагмент расщепляют за счет BamHI-NcoI и клонируют антисмысловым образом перед терминатором нопалинсинтазы в pMOG707. Последовательность усеченного промотора Дельта0.3TobRB7-5A (Yamamoto и др. 1991, Plant Cell 3, 371-382), выделенная, как описано в примере IIIm, вставляют как фрагмент XbaI-NcoI. Целую последовательность затем клонируют в бинарный вектор pMOG23 после расщепления за счет EcoRI и одного из остающихся уникальных сайтов рестрикции, получая в результате бинарный вектор pMOG715 (фиг. 14).
s) Описание pMOG719, бинарной плазмиды, содержащей промотор типа В и смысловые комбинации гетерологичного жизнеспособного гена.
Это описание дают с помощью основной иллюстрации; pMOG719 является производной pMOG22, и ее используют для первого этапа трансформации растения с последующей трансформацией за счет бинарного вектора, как описано в примере IIIn-r. Конструкции pMOG719 содержат промотор-B, слитый со смысловым геном, кодирующим белок или полипептид, функционально похожий на антисмысловой ген из примера IIIn-r, но являющийся гетерологичным в последовательности нуклеотидов (фиг. 15). Предпочтительно, чтобы идентичность нуклеотидных последовательностей транскриптов, закодированных антисмысловым трансгеном и смысловым трансгеном, составляла менее 90%, предпочтительно менее 80% и еще более предпочтительно менее 75%. Смысловой трансген можно получить из видов, отличных от рассматриваемого вида. В качестве примеров таких гетерологичных генов сделаны следующие конструкции:
- кассету экспрессии pMOG180 (пример I, 35S-промотор/nos-терминатор) переносят в pMOG22 как фрагмент EcoRI - HindIII, получая в результате pMOG22bis.
- pMOG719-1: ген, кодирующий глицерол-3P-ацилтрансферазу (Ishizaki и др. 1988 FEBS Lett 238, 424-430, EMBL регистрационный номер Y00771), выделяют из кДНК тыквы, применяя PCR амплификацию со следующими праймерами:
5' GCTTCCAGATCTATGGCGGAG 3' (SEQ ID NO:21) и 5' GATTACCAAAAGATCTGATGTTG 3' (SEQ ID NO:22).
Размноженный фрагмент расщепляют за счет BglII и клонируют в бинарный вектор pMOG22bis, который переводят в линейную форму при помощи BamHI, получая в результате бинарный вектор pMOG719-1. Этот вектор проверяют на правильную смысловую ориентацию G3PAT, проводя рестрикционный анализ,
- pMOG719-2: ген, кодирующий транслокатор адениннуклеотида (Baker и Leaver 1985 Nucl. Acids Res. 13, 5857-5867, EMBL регистрационный номер Х02842), выделяют из кДНК маиса, применяя PCR амплификацию со следующими праймерами:
5' GAATTCGGATCCTGATGCCAGACCCCGCTCTGTG3' (SEQ ID NO:23) и 5' GGACCGGGATCCCACACTGCTCTTG 3' (SEQ ID NO:24).
Размноженный фрагмент расщепляют при помощи BamHI и клонируют в бинарный вектор pMOG22bis, который переводят в линейную форму за счет BamHI, получая в результате бинарный вектор pMOG719-2. Этот вектор проверяют на правильную смысловую ориентацию ANT путем рестрикционного анализа,
- pMOG719-3: ген, кодирующий бета субъединицу АТФ синтазы (Winning и др. 1990 Nucl. Acids Res. 18, 5885, EMBL регистрационный номер Х54233), выделяют из кДНК маиса, применяя PCR амплификацию со следующими праймерами:
5'CCCTGGATCCGGACCGGCCATGGC 3' (SEQ ID NO:25) и 5' CAAGCAAGGATCCTCCTTATGAAGC 3' (SEQ ID NO:26).
У размноженного фрагмента затупляют конец и субклонируют в pUC9 (Vieira и Messing 1982 Gene 19, 259 - 268), которую переводят в линейную форму за счет SmaI и HinCII. Получаемый вектор частично расщепляют при помощи BamHI и клонируют в бинарный вектор pMOG22bis, получая бинарный вектор pMOG719-3. Этот вектор проверяют на правильную смысловую ориентацию АТФ синтазы посредством рестрикционного анализа.
Пример IV. Мобилизация бинарных векторов в Agrobacterium.
Бинарные векторы, описанные в примере III, мобилизуют в тройном скрещивании за счет E.coli K-2 штамма НВ101 (содержащего плазмиду KR-2013) (Ditta и др., 1980, Proc. Nat. Acad. Sci. USA 77, 7347-7351) в Agrobacterium tumefacience штамм MOG101 (пример II) или LBA4404 (Hoekema и др. 1983, Nature 303, 179-180), который содержит плазмиду с генами вирулентности, необходимыми для переноса T-ДНК в растения.
Пример V. Трансформация растений
а) Трансформация Arabidopsis thaliana
Arabidopsis трансформируют путем совместного культивирования ткани растения с Agrobacterium tumefaciens штаммом MOG101, содержащим один из бинарных векторов, описанных в примере III. Трансформацию осуществляют, применяя совместное культивирование сегментов корней Arabidopsis thaliana (экотип С24), как описано Valvekens и др. (1988, Proc. Nat. Acad. Sci. USA 85, 5536-5540). Трансгенные растения регенерируют из ростков, которые вырастают на селекционной среде (содержащей либо канамицин, либо гигромицин в зависимости от исходной бинарной плазмиды, pMOG23 или pMOG22 соответственно), укореняют и переносят в среду проращивания или на грунт. Молодые растения выращивают до зрелости и дают им самоопылиться и завязать семена.
b) Трансформация картофеля (Solanum tuberosum spp.).
Картофель трансформируют путем совместного культивирования ткани растения с Agrobacterium tumefaciens штаммом L ВА4404, содержащим один из бинарных векторов, описанных в примере III. Трансформацию осуществляют, применяя совместное культивирование дисков клубней картофеля (Solanum tuberosum var. Desiree), как описано Hoekema и др. 1989, Bio/Techn. 7, 273 - 278). Трансгенные растения регенерируют из ростков, которые вырастают на селекционной среде (содержащей либо канамицин, либо гигромицин в зависимости от исходной бинарной плазмиды pMOG22 или pMOG23), укореняют, размножают аксенически, разрезая меристему, и переносят на грунт. Молодые растения выращивают до зрелости и дают им образовать клубни.
с) Трансформация табака (Nicotiana tabacum SRI)
Табак трансформируют совместным культивированием ткани растения с Agrobacterium tumefaciens штаммом LBA4404 (Hoekema и др. 1980, Nature 303, 179-180), содержащим один из бинарных векторов, описанных в примере III. Трансформацию осуществляют, применяя совместно культивирование дисков листьев табака (Nicotiana tabacum SRI), как описано Horsch и др. 1985, Science 227, 1229-1231). Трансгенные растения регенерируют из ростков, которые вырастают на селекционной среде (содержащей либо канамицин, либо гигромицин в зависимости от исходной бинарной плазмиды pMOG22 или pMOG23), укореняют и переносят на грунт. Молодые растения выращивают до зрелости и дают им самоопылиться и завязать семена.
Пример VI. Исследование трансгенных растений Arabidopsis на восприимчивость к PPN.
Трансгенные растения Arabidopsis исследуют in vitro или на грунте на устойчивость против M. incognita или цистнематод H. schachtii. Для анализа in vitro стерилизуют поверхность семян, семена проращивают и высевают, как описано Sijmons и др. (1991, Plant J. 1, 245-254). Через две недели после заражения можно визуально наблюдать, что корневые системы отмечены многочисленной, успешно развивающейся инфекцией, и сравнить эти растения с растениями диких видов Arabidopsis. Линии растений считают устойчивыми, когда они демонстрируют значительное снижение восприимчивости к PPN (то есть значительное уменьшение количества женских особей, обнаруженных на проверяемых корнях). Для растений, выращиваемых на грунте, ростки проращивают на селективной среде (10 мг/л гигромицина или 50 мг/л канамицина). Устойчивые ростки переносят на смесь земли с песком (1:3 по объему) в прозрачные пластиковые трубки 1 х 1 х 6 см. Когда розетки (rozettes) хорошо разовьются (примерно 14 дней), контейнеры засевают выведенными J2H. schachtii, примерно 300 в каждый контейнер. Выведение H. schachtii стимулируют, погружая цисты на несколько дней в 3 мМ раствор ZnCl2 при температуре около 20oC. Через восемнадцать дней после засева корни осторожно вынимают из смеси земли с песком и окрашивают кислым фуксином (Dropkin, 1989 в: Introduction to plant nematology, 2-ое издание, Wiley & Sons, New York). В этом опыте восприимчивые растения дают 17 цист на корневую систему (разброс значений 4-40 цист на корневую систему). Генотип считают устойчивым, когда количество цист снижается до 2 на корневую систему). Аналогично, растения можно засевать выведенными J2 M. incognita или яйцекладками, которые смешивают со смесью земли и песка. Затем растения исследуют на присутствие галлов, которые ясно видны после удаления с корней смеси земли и песка.
Пример VII. Исследование трансгенных растений табака на восприимчивость к PPN.
Для проверки устойчивости к нематодам предварительно заражают землю яйцекладками M. incognita. Этот посевной материал можно получить, выдерживая исходную культуру M. incognita на растениях сельдерея (Apium graveolens), растущих на грунте в стандартных тепличных условиях при температуре ниже 25oC. Корневые системы зрелых растений сельдерея, содержащие большое количество корневых наростов и зрелых женских особей M. incognita, аккуратно очищают для удаления земли, быстро гомогенизируют в миксере (Waring blendor) (2 с) и развешивают по порции по 60 г. Эти образцы корней смешивают с 1 кг смеси песок : земля для горшков (1:1) и используют для выращивания трансгенных растений табака в течение 6 недель. Для каждого генотипа в каждом тесте используют по крайней мере 100 индивидуальных растений. Смесь земли с песком затем тщательно смывают и при помощи бинокуляра подсчитывают количество галлов на корневой системе. В этом эксперименте контрольные растения имеют 25 ± 15 галлов. Генотип считают устойчивым, когда количество галлов снижено до 2 на корневую систему.
Пример VIII. Исследование трансгенных растений картофеля на восприимчивость к PPN.
Трансгенные растения картофеля исследуют на устойчивость к M. incognita, используя землю, предварительно зараженную яйцекладками M. incognita, в смеси с песком (1: 3 по весу), выращивая на этой смешанной почве растения картофеля в течение 6 недель и подсчитывая количество образовавшихся на корневых системах галлов после удаления почвы. По-другому, для проверки устойчивости к Globodera ssp. используют закрытый контейнер. Для этого опыта три одинаковых клубня (2-4 см) переносят на грунт, который предварительно засеян цистами G. rostochiensis иди G. pallida в прозрачных контейнерах. Периферийные корневые системы исследуют визуально на присутствие цист через 7-8 недель после начала развития. Генотип считают устойчивым, если ни в одном из повторных опытов нет цист, и восприимчивым, если по крайней мере в одном из экспериментов появляются цисты.
Пример IX. Исследование GUS-активности в питательных структурах растений, трансгенных для chimaeric 35S/GUS- конструкции.
Линии растений, трансгенных для T-ДНК конструкций pMOG25 или pMOG28 (35S/GUS), используют для заражения цист- или клубеньковыми нематодами и последующего исследования GUS-активности (Jefferson, 1987 Plant Mol. Biol. Reporter 5, 387-405) в питательных структурах. Arabidopsis специально подходит для этой цели, так как эти растения легко использовать в культурах с Heterodera schachtii и Meloidogyne incodnita, паразитирующими только в одном растении-хозяине (Sijmons и др. 1991 Plant J., 1., 245-254). Развитие питательной структуры исключительно хорошо видно у этого вида растений при выращивании на пластинах агара. Куски агара, содержащие части зараженных корней, срезают прямо с чашек Петри, не нарушая взаимодействия нематоды - растение, и напрямую используют для гистохимического GUS окрашивания при 37oC при помощи субстрата 5-бром-4-хлор-3-индолил-глукуронида (X-Glu) в буфере, содержащей следующие компоненты: 100 мМ NaPi pH 7,0 10 мM Na2EDTA, 0,1% Triton X100, 2,1 μг/мл феррицианида, 1,6 μг/мл ферроцианида. На этой модельной растительной системе ясно демонстрируют понижающую регуляцию активности 35S-промотора на клеточном уровне. В любых клетках, включающих питательную структуру, отсутствует детектируемая GUS-активность (следовательно, нет активности 35S-промотора) даже после длительного окрашивания X-Glu. С другой стороны, в незараженных корнях обнаруживают GUS-активность во всех частях корней, она особенно высока в тканях молодых корней и сосудистом цилиндре. На последних стадиях заражения Arabidopsis нематодами M. incognita понижающая регуляция 35S-промотора распространяется даже за пределы питательной структуры.
Почву, на которой растут трансгенные растения табака, засевают Globodera tabacum или M. incognita и анализируют после надлежащего развития питательных структур. Для этой цели корни осторожно промывают для удаления почвы, инкубируют с буфером, содержащим X-Glu, и дважды окрашивают кислым фуксином для определения нематод (Dropkin, 1989).
Аналогичные эксперименты проводят с трансгенным картофелем после засевания почвы Globodera rostochiensis, G. pallida или M. incognita. Во всех случаях активность не детектируют в питательных структурах, тогда как незараженные части корней показывают экспрессию, ожидаемую для 35S-промотора. Таким образом, промотор CaMV 35S соответствует требованиям настоящего описания, предъявляемым к промотору типа В.
Пример X. Исследование GUS-активности в питательных структурах растений, трансгенных для химерной (chimaeric) rolD/GUS-конструкции.
Линии растений, трансгенных для T-ДНК конструкции pMOG630 (rolD/GUS), используют для заражения цист- или клубеньковыми нематодами и последующего анализа GUS-активности, как описано в предыдущем примере. Здесь опять ясно демонстрируют понижающую регуляцию активности rolD промотора на клеточном уровне. В любых клетках, включающих питательную структуру, отсутствует детектируемая GUS-активность (следовательно, нет активности rolD-промотора) даже после продолжительного окрашивания X-Glu. С другой стороны, в незараженных корнях GUS-активность обнаруживают во всех частях корней, она особенно высока в тканях молодых корней, включая сосудистый цилиндр, эпидермис и корневые волоски. Таким образом, Agrobacterium rhizogenes rolD-промотор соответствует требованиям, предъявляемым в данном описании промотору типа B.
Пример XI. Исследование GUS-активности в питательных структурах растений, трансгенных для химерной усеченный rolC/GUS-конструкции.
Линии растений, трансгенных для T-ДНК конструкции pMOG679 (усеченный rolC/GUS), используют для заражения цист- или клубеньковыми нематодами и последующего анализа GUS- активности, как описано в примере VI. В противоположность 35S или rolD промотору, а также в противоположность некоторым другим промоторам, исследованным в аналогичных условиях, слияние 5'усеченного rolC-промотора/GUS увеличивает синий осадок в питательной структуре, когда добавляют подходящий субстрат. Таким образом, Agrobacterium rhizogenes 5'усеченный rolC-промотор соответствует требованиям настоящего описания, предъявляемым к промотору типа А.
Пример XII. Идентификация регуляторных последовательностей типа промотора-A в NFS трансгенных растений.
а) Трансформация Arabidopsis за счет pMOG452
Arabidopsis трансформируют при помощи Agrobacterium штамма MOG101, содержащего бинарный вектор pMOG452 (GUS без промотора, присоединенный к правой границе T-ДНК), используя процедуру, описанную в примере V.
b) Исследование GUS-активности в питательных структурах растений трансгенных для GUS-конструкции без промотора
Линии трансгенных растений (pMOG452, Т2 или более поздние генерации) используют для заражения PPN и последующего GUS-анализа, как описано в примере IX. После засевания H. schachtii результаты таких экспериментов по скринингу демонстрируют удивительно большое количество растений с GUS-активностью в синцитии и низкую GUS-активность или ее отсутствие в других частях растений, показывая, таким образом, что регуляторные последовательности, которые управляют активностью генов в NFS, связаны с GUS-геном и поэтому их можно исправить для быстрого выделения таких регуляторных последовательностей. Одна из 13 независимых трансгенных линий растений указывает присоединенный промотор, активный только в корнях, и который остается активным в NFS. Это можно подтвердить Arabidopsis растениями, трансгенными для GUS-конструкции без промотора, как описано в (Topping и др., 1991, Developm. 112, 1009-1019). Две из 25 трансгенных линий растений демонстрируют GUS-активность в клетках, включающих NFS. Эта высокая частота иллюстрирует способность прикреплять и, следовательно, выделять регуляторные последовательности, которые выполняют требования промотора-A. Обратную ситуацию (GUS-активность в здоровом сосудистом цилиндре, но регулируется с понижением в развивающихся питательных структурах) наблюдают даже с большей частотой и, следовательно, метод в принципе подходит также для идентификации регуляторных последовательностей, которые соответствуют требованиям промотора типа B.
с) Выделение регуляторных последовательностей типа промотора-A из трансгенных растений, экспрессирующих GUS в NFS.
Выбирают линии трансгенных растений, которые проявляют GUS-активность в NFS и предпочтительно низкую активность или ее отсутствие в других тканях, и используют их для выделения геномной ДНК. Регуляторные последовательности выше (5') интегрированного GUS-гена выделяют на следующих стадиях, используя обратную PCR (Does и др. 1991, Plant Mol. Biol. 17, 151-153) с праймерами: 5' CCA GAC TGA ATG CCC ACA GGC 3' (SEQ ID NO:27) и 5' GGT GAC GCA TGT CGC GCA AG 3' (SEQ ID NO:28).
Размноженный фрагмент используют для отбора геномной библиотеки Arabidopsis для выделения геномных клонов, содержащих регуляторные последовательности, которые соответствуют требованиям промотора-A. По-другому, первые 200 пар оснований GUS-гена можно использовать для исследования банка геномов, полученного из выбранных растений.
Пример XIII. Синтез Barstar в трансгенных растениях, контролируемых регуляторными последовательностями промоторов типа B.
Для вестерн (western)-анализа растений, экспрессирующих Barstar ген, используют антисыворотку против Barstar, которую получают следующим образом: тремя различными способами прогнозирования, основанными на гидрофильности, пластичности и β-поворотах, определяют большинство антигенных детерминант Barstar пептида. На основании этого анализа синтезируют три пептида, используя автоматический синтезатор пептидов со следующей последовательностью аминокислот:
a) Ala-Glu-Ser-Val-Leu-Gln-Val-Phe-Arg-Glu-Ala-Lys-Ala-Glu-Gly (SEQ ID NO:29)
b) Arg-Gln-Phe-Glu-Gln-Ser-Lys-Gln-Leu-Thr-Glu-Asn-Gly-Ala-Glu (SEQ ID NO:30)
c) Leu-Lys-Lys-Glu-Leu-Ala-Leu-Pro-Glu-Tyr-Tyr-Gly- Asn-Leu-Asp (SEQ ID NO:31).
Эти пептиды чистят и 1,5 мг каждого помещают в 7,5 мг бычьего сывороточного альбумина (BSA), используя глутаральдегид. Полученные конъюгаты анализируют, они содержат 7,5 молей пептида (а) на моль BSA, 10,0 молей пептида, (b) на моль BSA и 6,7 молей пептида (с) на моль BSA. Конъюгаты растворяют в 50 мМ Hepes, 100 мМ NaCl с финальной концентрацией 1 мг/мл и перемешивают. Кроликам делают инъекцию 0,15 мл смешанных конъюгатов в полных Freunds адъювантах и стимулируют три раза дополнительными инъекциями по 0,15 мл смешанных конъюгатов в полных Freunds адъювантах. Получаемую сыворотку частично чистят, выделяя фракцию антител и хранят при -80oC. Эту сыворотку используют при 5000-10000 кратном разбавлении в присутствии 5% обезжиренного сухого молока в физиологическом растворе с фосфатным буфером, 1% Tween 20, 0,5% Triton - X100. Для вестерн-анализа сырые растительные экстракты разделяют на 10-27% градиентном SDS - PAGE (электрофорез на додецилсульфате натрия-полиакриламидном геле) с последующим электроблотированием на нитроцеллюлозу. Barstar антитела детектируют при помощи конъюгатов goat-anti-rabbit-пероксидазы и наблюдают, применяя Amersham (UK) ECL-систему детектирования.
Выбор Arabidopsis, табака и картофеля для введения конструкций для экспрессии barnase и barstar основан на превосходных свойствах Arabidopsis анализировать взаимодействие растение-нематоды (Sijmons и др. 1991, Plant J. 1, 245-254) или превосходных условиях трансформации, имеющихся в табаке и картофеле, что не ограничивает набор растений, к которым применимо это изобретение.
Arabidopsis трансформируют за счет Agrobacterium штамма MOG101, содержащего бинарный вектор pMOG583, или pMOG716, или pMOG718 (все описаны в примере III), применяя процедуру, описанную в примере Va. Трансформанты регенерируют на селективной среде, содержащей 10 мг/л гигромицина. Всем растениям, устойчивым к гигромицину, дают самоопылиться и полученные сорта семян проращивают на гигромицине и проверяют на экспрессию barstar гена посредством вестерн-анализа, используя антитела против конъюгатов BSA-Barstar. Линии представленных растений с высокой, средней и низкой экспрессией используют для следующего этапа трансформации.
Табак и картофель трансформируют за счет Agrobacterium штамма LBA4404, содержащего бинарный вектор pMOG583, или pMOG716, или pMOG718 (все описаны в примере III), применяя процедуру, описанную в примере Vb и Vc. Трансформанты регенерируют на селективной среде и устойчивым растениям дают самоопылиться или размножают клонально. В случае табака полученные сорта семян проращивают на селективной среде и проверяют на экспрессию barstar гена посредством вестерн-анализа, используя антитела против barstar. Клонально размноженные клоны картофеля исследуют на экспрессию barstar после переноса растений на грунт. Линии представленных растений с высокой, средней и низкой экспрессией используют для второго этапа трансформации.
Пример XIV. Введение последовательностей ДНК Barnase без промотора в растения, экспрессирующие Barstar, из примера XIII и исследование пониженной восприимчивости к PPN
а) Трансформация Arabidopsis за счет pMOG589i.
Выбранные линии растений Arabidopsis из примера XIII, которые экспрессируют Barstar ген, используют для трансформации за счет Agrobacterium штамма MOG101, содержащего бинарный вектор pMOG589i (пример III), применяя процедуру, описанную в примере VI. Регенерацию трансформантов производят на среде, содержащей 50 мг/л канамицина. Все реагенты, устойчивые к канамицину, выращивают до зрелости, дают им самоопылиться и анализируют на отклоняющиеся фенотипы. Семена от всех растений с нормальным фенотипом проращивают на канамицине и выращивают, получая следующее поколение семян. Следующий этап проращивания на среде, содержащей канамицин, позволяет идентифицировать линии растений, гомологичные для NPTII гена.
b) Исследование трансгенных Arabidopsis на восприимчивость к PPN.
Растения, полученные по процедуре, описанной в примере XIVa, используют для создания их степени восприимчивости к PPN, как описано в примере VI. Из этих исследований идентифицируют несколько линий растений, которые показывают пониженную восприимчивость к PPN (то есть, уменьшение количества цист с количественной характеристикой менее 10 цист на корневую систему), было обнаружено несколько устойчивых линий (то есть, значение меньше 2 цист на корневую систему), особенно, среди тех линий, которые выбирают по их высокой экспрессии barstar в примере XIII. Наблюдают также много abbarent фенотипов (то есть, со сниженной жизнедеятельностью, стерильных, не цветущих, с маленькими корневыми системами), возможно, в результате локальной экспрессии barnase гена. Используя обратную PCR (Does и др. 1991, Plant Mol. Biol. 17, 151-153), выделяют регуляторные последовательности выше (5') интегрированного Barnase гена. Размноженный фрагмент используют для отбора геномной библиотеки Arabidopsis для выделения геномных клонов, содержащих регуляторные последовательности, которые соответствуют требованиям промотора-A. По-другому, barnase ген используют для исследования банка геномов, полученного из выбранных растений.
с) Трансформация табака и картофеля за счет pMOG591bis
Регуляторные последовательности, которые идентифицированы по процедуре, описанной в этом примере, можно использовать для введения в сайт мультиклонирования pMOG591i (пример III), создавая, таким образом, бинарный вектор pMOG591i. Этот вектор затем используют для второго этапа трансформации растений, например, растений табака или картофеля, выбранных по высокой экспрессии Barstar в примере XIII. Полученные двойные трансформанты можно исследовать на снижение восприимчивости к PPN, как описано в примерах VII и VIII.
Пример XV. Введение "усеченный rolC-промотор/Barnase ДНК" последовательностей в растения, экспрессирующие Barstar.
Выбранные линии растений из примера XIII используют для трансформации за счет Agrobacterium штамма MOG101 или L ВА4404, содержащего бинарный вектор pMOG717i (усеченный rolC/Barnase; пример III), применяя процедуру примера VI. Регенерацию трансформантов проводят на среде, содержащей канамицин. Все регенеранты, устойчивые к канамицину, выращивают до зрелости, дают им самоопылиться или образовать клубни (в зависимости от вида) и анализируют на отклоняющиеся фенотипы. Для самоопыляющихся видов все растения с нормальным фенотипом проращивают на канамицине и выращивают, получая следующее поколение семян. Следующий этап проращивания на среде, содержащей канамицин, позволяет идентифицировать линии растений, гомозиготные для NPTII гена. В случае картофеля трансгенные линии растений размножают клонально.
b) Исследование дважды трансгенных линий растений на восприимчивость к PPN.
В данном случае исследуют растения, трансгенные для Barstar гена (снабженного pMOG583, или pMOG716 или pMOG718) и Barnase гена (pMOG717i), на степень их восприимчивости к PPN, как описано в примерах VI - VIII. Из этих исследований идентифицируют линии растений, которые показывают значительное снижение восприимчивости к PPN (то есть, значительное уменьшение количества цист или галлов по сравнению с контрольными корнями), особенно, среди тех линий, которые выбраны по их высокой экспрессии barstar в примере XIII.
Пример XVI. Введение "Дельта0.3TobRB7-5A-промотор/Barnase ДНК" последовательностей в растения, экспрессирующие Barstar.
Выбранные линии растений из примера XIII используют для трансформации за счет Agrobacterium штамма MOG101 или L BA4404, содержащего бинарный вектор pMOG699i (пример III), применяя процедуру, описанную в примере V. Регенерацию трансформантов производят на среде, содержащей канамицин. Все регенеранты, устойчивые к канамицину, выращивают до зрелости, дают им самоопылиться или образовать клубни (в зависимости от вида) и анализируют на отклоняющиеся фенотипы. Для самоопыляющихся образцов все растения с нормальным фенотипом проращивают на канамицине и выращивают, получая следующее поколение семян. Следующий этап проращивания на среде, содержащей канамицин, позволяет идентифицировать линии растений, гомозиготные для NPTII гена. В случае картофеля трансгенные линии растений размножают клонально.
b) Исследование дважды трансгенных линий растений на восприимчивость к PPN.
Растения, трансгенные и для Barstar (pMOG583, или pMOG716, или pMOG718), и для Barnase (pMOG699i) конструкции, можно исследовать на степень их восприимчивости к PPN, как описано в примерах VI - VIII. Из этих исследований идентифицируют линии растений, которые показывают значительное снижение восприимчивости к M. incognita (то есть, значительное уменьшение количества галлонов по сравнению с контрольными корнями), особенно, среди тех линий, которые выбраны по их высокой экспрессии barstar в примере XIII.
Пример XVII. Инженерия пониженной восприимчивости к PPN за счет одностадийной трансформации.
По-другому, можно конструировать бинарные векторы, которые содержат обе кассеты экспрессии для осуществления регулированной экспрессии Barstar и Barnase, из фрагментов ДНК, описанных в примере III (g)-(m) способами, применимыми для основного описания этого патента, как показано на фиг. 1. Такие бинарные векторы можно затем использовать для инженерии растений с пониженной восприимчивостью к PPN посредством одностадийной трансформации и исследования линий растения, как описано в примерах VI - VIII.
Пример XVIII. Инженерия пониженной восприимчивости к PPN, используя бинарные конструкции: промотор-A/антисмысловой гомологичный существенный ген и промотор-B/смысловой гетерологичный существенный ген.
Эти примеры основаны на той же концепции для двухкомпонентной системы, как в основном показано на фиг. 1. Она включает локальную (в питательной структуре) антисмысловую экспрессию гомологичного гена (предпочтительно, выделенного из интересующего вида), который является существенным для метаболизма клеток растения, и экспрессию второго трансгена, который при экспрессии способен нейтрализовать или частично нейтрализовать эффект, вызванный первым трансгеном, и является, в действительности, смысловой конструкцией гетерологичного гена, его экспрессия происходит в клетках вне питательной структуры.
а) Введение химерных ДНК последовательностей Дельта0.3TobRB7-промотора и антисмыслового NADPH-цитохром-P450-ATR1-гена для специфического подавления индуцируемой нематодами питательной структуры в Arabidopsis
Растения Arabidopsis, трансгенные для конструкции типа B-промотора и гетерологичного гена для NADPH-Cyt P450 редуктазы, используют для второго этапа трансформации посредством совместного культивирования ткани растения с Agrobacterium tumefaciens штаммом MOG101, содержащим бинарный вектор pMOG711, как описано в примере V. Трансгенные растения регенерируют из отростков, которые выращивают на селекционной среде, укореняют и переносят на среду для проращивания или на грунт. Молодые растения выращивают до зрелости и дают им самоопылиться и завязать семена. Растения, трансгенные для обеих конструкций, смысловой и антисмысловой, исследуют на пониженную восприимчивость к PPN, как описано.
b) Введение химерных ДНК последовательностей Дельта-0.3TobRB7-промотора и антисмыслового NADPH-цитохром-P450-ATR2-гена для специфического подавления индуцируемых нематодами питательных структур в Arabidopsis
Растения Arabidopsis, трансгенные для конструкции типа B-промотора и гетерологичного гена для NADPH-Cyt-P-450 редуктазы, используют для второго этапа трансформации посредством совместного культивирования ткани растения с Agrobacterium tumefaciens штаммом MOG101, содержащим бинарный вектор pMOG712, как описано в примере V. Трансгенные растения регенерируют из ростков, которые проращивают на селекционной среде, укореняют и переносят на среду для проращивания или на грунт. Молодые растения выращивают до зрелости и дают им самоопылиться и завязать семена. Растения, трансгенные для обеих конструкций, смысловой и антисмысловой, анализируют на пониженную восприимчивость к PPN, как описано.
с) Введение химерных ДНК последовательностей 35S-промотора и Squash-гена, кодирующего глицерол-3-фосфатацилтрансферазу
Растения Arabidopsis трансформируют за счет pMOG719-1 посредством совместного культивирования ткани растения с Agrobacterium tumefaciens штаммом MOG101, содержащим бинарный вектор, как описано в примере V. Трансгенные растения регенерируют из ростков, которые проращивают на селекционной среде, укореняют и переносят на среду для проращивания или грунт. Молодые растения можно выращивать до зрелости и дать им самоопылиться и завязать семена. Гомозиготные линии выбирают, исследуя потомство при проращивании на селективной среде. Уровень экспрессии гетерологичного гена исследуют посредством нозерн-блотирования.
d) Введение химерных ДНК последовательностей Дельта-0.3TobRB7 промотора и антисмыслового гена глицерол-3-фосфатацилтрансферазы для специфического подавления индуцируемых нематодами питательных структур в Arabidopsis
Растения Arabidopsis, трансгенные для T-ДНК конструкции pMOG719-1, используют для второго этапа трансформации посредством совместного культивирования ткани растения с Agrobacterium tumefaciens штаммом MOG101, содержащим бинарный вектор pMOG713, как описано в примере V. Трансгенные растения регенерируют из отростков, которые выращивают на селекционной среде, укореняют и переносят на среду проращивания или грунт. Молодые растения можно вырастить до зрелости, дать им самоопылиться и завязать семена. Растения, трансгенные для обеих конструкций, смысловой и антисмысловой, исследуют на пониженную восприимчивость к PPN, как описано.
е) Введение химерных ДНК последовательностей 35S-промотора и гена маиса, кодирующего для транслокатора адениннуклеотида.
Растения табака трансформируют за счет pMOG719-2 при совместном культивировании ткани растения с Agrobacterium tumefaciens штаммом L BA4404, содержащим бинарный вектор, как описано в примере V. Трансгенные растения регенерируют из ростков, которые выращивают на селекционной среде, укореняют и переносят на среду проращивания или грунт. Молодые растения можно выращивать до зрелости, дать им самоопылиться и завязать семена. Гомозиготные линии выбирают, анализируя потомство при проращивании на селективной среде. Уровень экспрессии гетерологичного гена исследуют посредством нозерн-блотирования.
f) Введение химерных ДНК последовательностей Дельта-0.3TobRB7 промотора и антисмыслового гена транслокатора адениннуклеотида для специфического подавления вызываемых нематодами питательных структур в картофеле.
Растения картофеля, трансгенные для T-ДНК конструкции pMOG719-2, используют для второго этапа трансформации посредством совместного культивирования ткани растения с Agrobacterium tumefaciens штаммом L BA4404, содержащим бинарный вектор pMOG714, как описано в примере V. Трансгенные растения регенерируют из отростков, которые проращивают на селекционной среде, укореняют, размножают клонально и переносят на грунт. Растения, трансгенные для обеих конструкций, смысловой и антисмысловой, анализируют на пониженную восприимчивость к PPN, как описано.
g) Введение химерных ДНК последовательностей 35S-промотора и гена маиса, кодирующего для β-субъединицы АТФ-синтазы.
Растения картофеля трансформируют за счет pMOG719-3 при совместном культивировании ткани растения с Agrobacterium tumefaciens штаммом L ВА4404, содержащим бинарный вектор, как описано примере V. Трансгенные растения регенерируют из ростков, которые проращивают на селекционной среде, укореняют и переносят на грунт или клонально размножают in vitro. Уровень экспрессии гетерологичного гена исследуют способом нозерн-блотирования.
h) Введение химерных ДНК последовательностей Дельта-0.3TobEB7 промотора и антисмыслового гена АТФ синтазы для специфической репрессии индуцированных нематодами питательных структур в табаке.
Растения табака, трансгенные для конструкции pMOG717-3, используют для второго этапа трансформации при совместном культивировании ткани растения с Agrobacterium tumefaciens штамма L ВА4404, содержащего бинарный вектор pMOG715, как описано в примере V. Трансгенные растения регенерируют из ростков, которые проращивают на селекционной среде, укореняют и переносят на среду для выращивания или грунт. Молодые растения можно выращивать до зрелости и дать им самоопылиться и завязать семена. Растения, трансгенные для обеих конструкций, смысловой и антисмысловой, анализируют на пониженную восприимчивость к PPN, как описано.
Пример XIX. Исследование трансгенных растений Arabidopsis, устойчивых к токсину, на восприимчивость к PPN в присутствии токсических соединений.
Выбранные линии растений из примера XI, трансгенные для конструкции pMOG25, несущей гены, кодирующие для GUS и HPT, оба находящиеся под контролем CaMV-промотора, выращивают на среде агара, содержащего нематоды (Sijmons и др. 1991, Plant. J. 1, 245-254), с добавлением 10-20 мкг/мл гигромицина (добавляя по 1 мкг/мл) для образования максимальных сублетальных концентраций гигромицина для каждой линии растений, так как позиционные эффекты сайта интегрирования T-ДНК влияют на уровень экспрессии HPT-гена и, следовательно, уровень устойчивости к гигромицину. Новые семена от выбранных линий затем проращивают на свежих чашках Петри со средой, содержащей агар с нематодами, при максимальной сублетальной концентрации гигромицина и засевают PPN (Sijmons и др. 1991, Plant J. 1, 245-254). Ежедневно следят за поведением нематод при помощи обратного микроскопа и наблюдают нормальное проникание и поведение в корнях на ранних стадиях заражения в присутствии гигромицина. На более поздних стадиях заражения при росте в гигромицине развивается значительно меньшее количество женских особей. Развитию NFS препятствует локальная восприимчивость к гигромицину, как результат понижающей регуляции 35S-промотора.

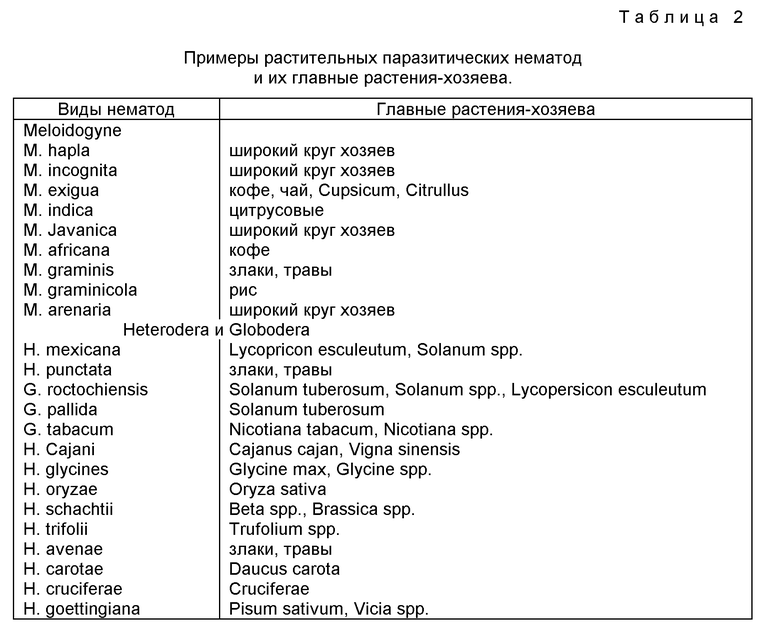









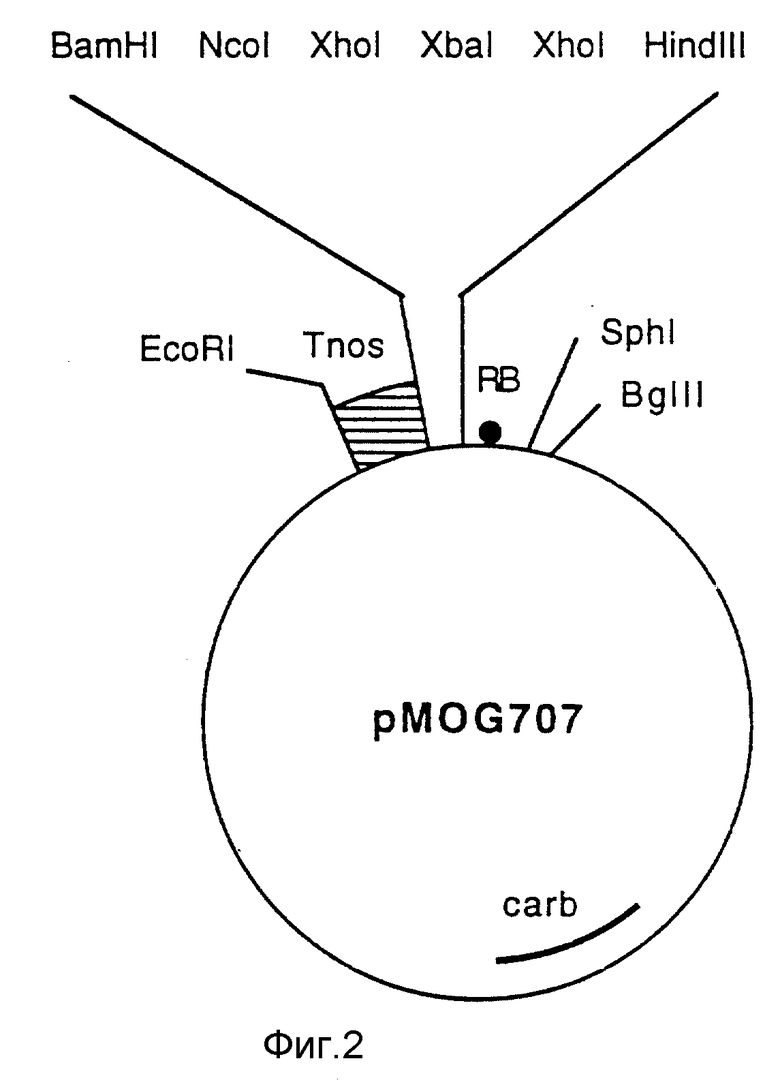
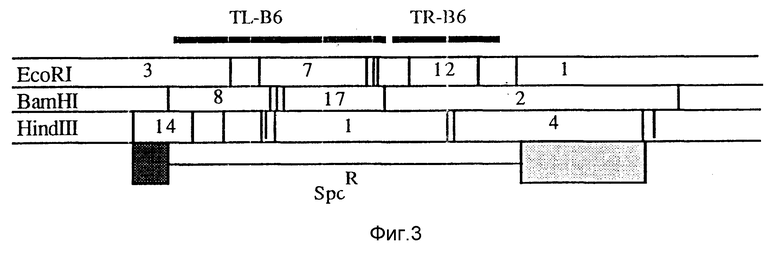
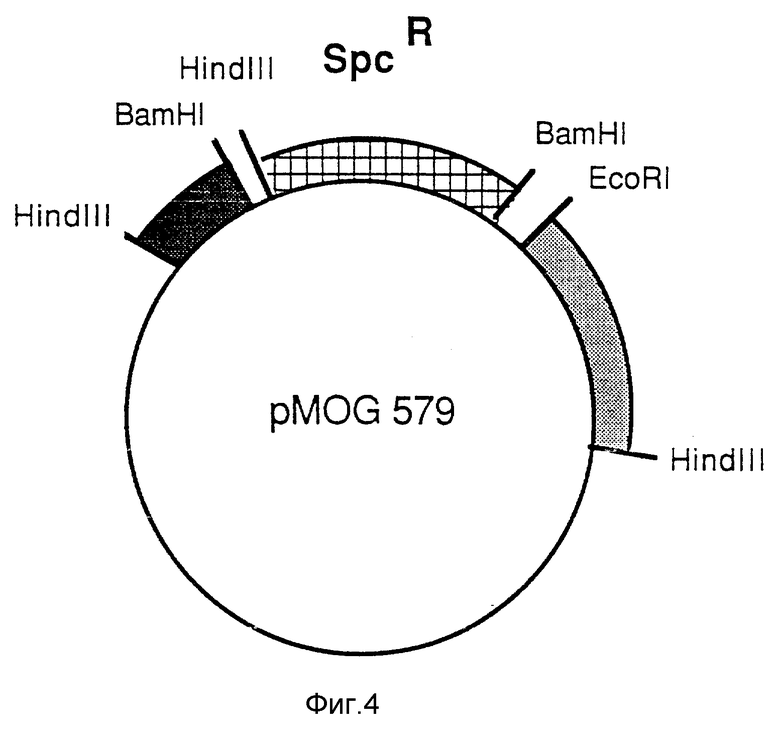
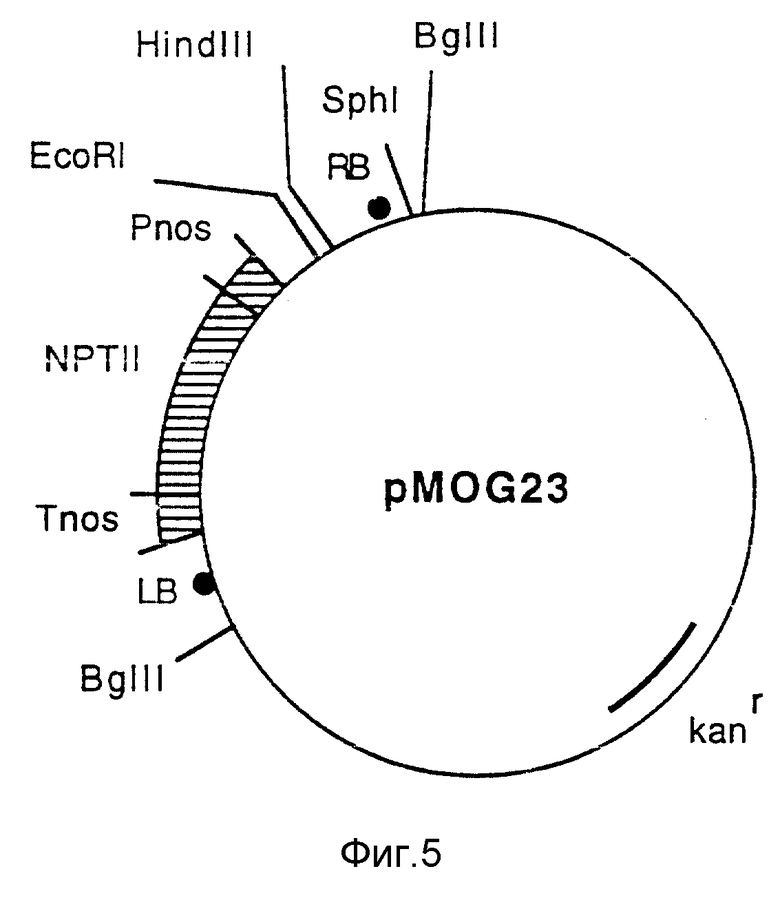
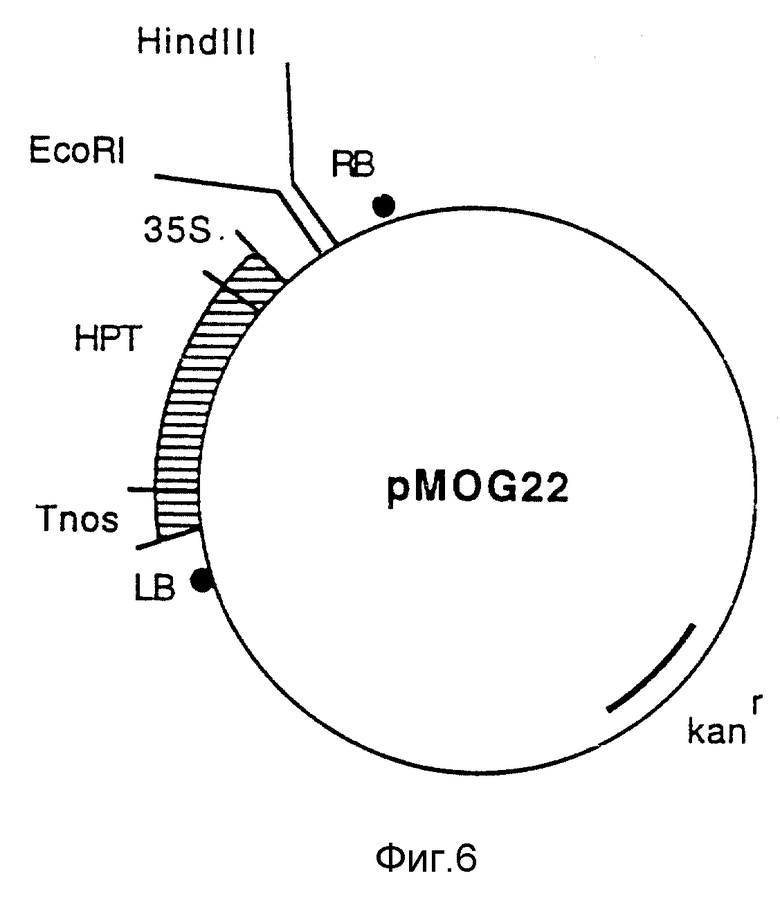
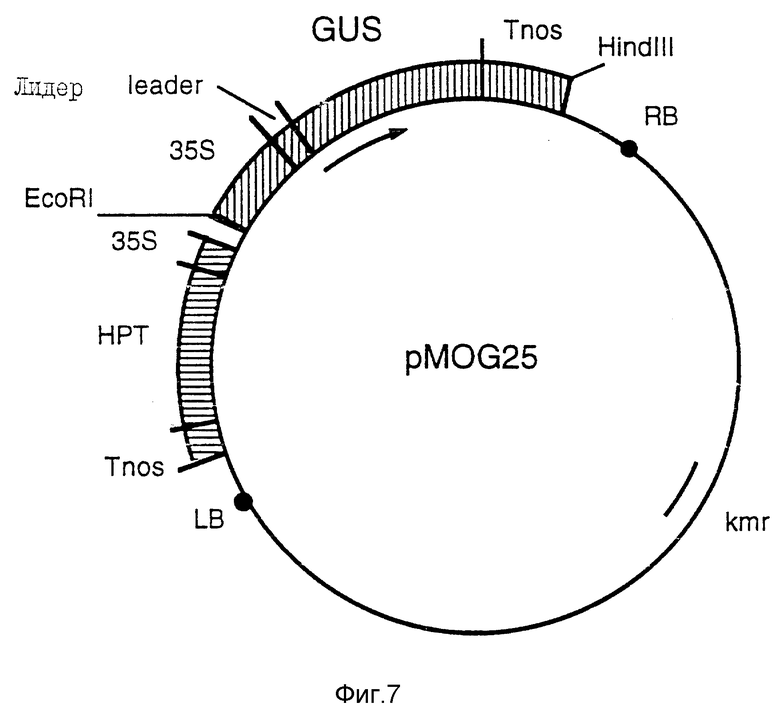
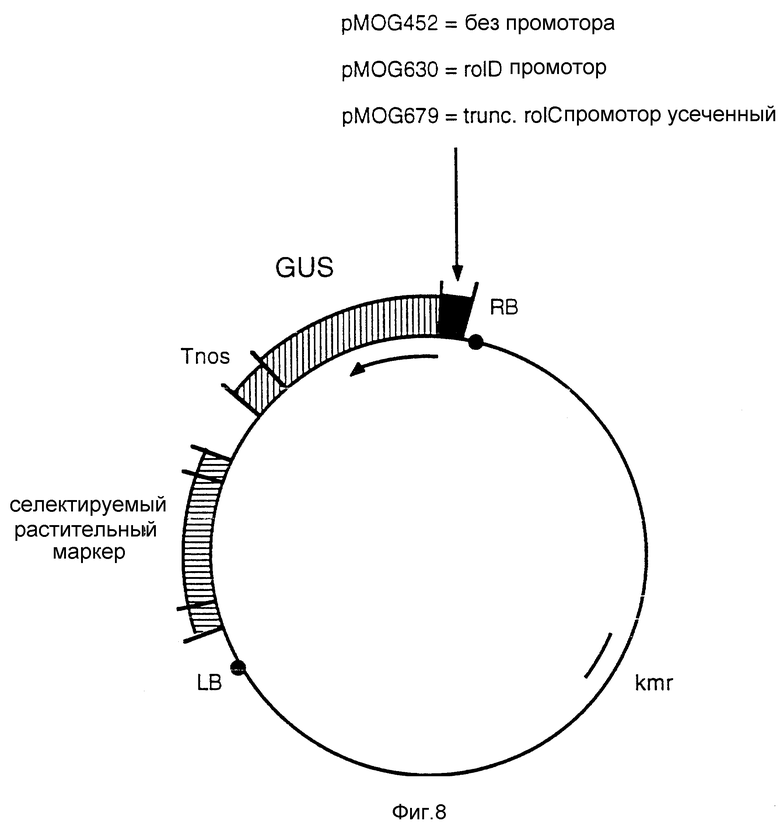
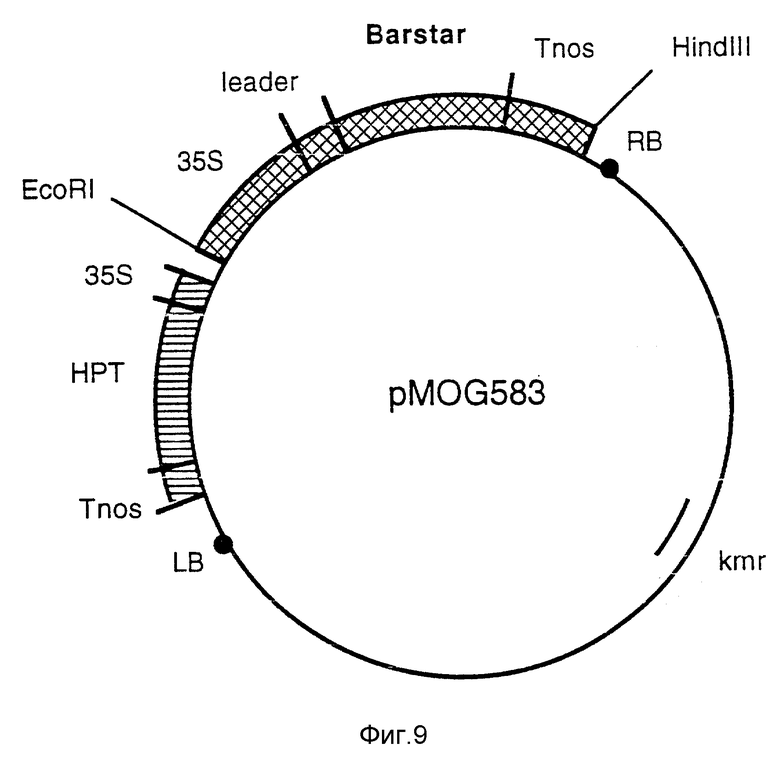
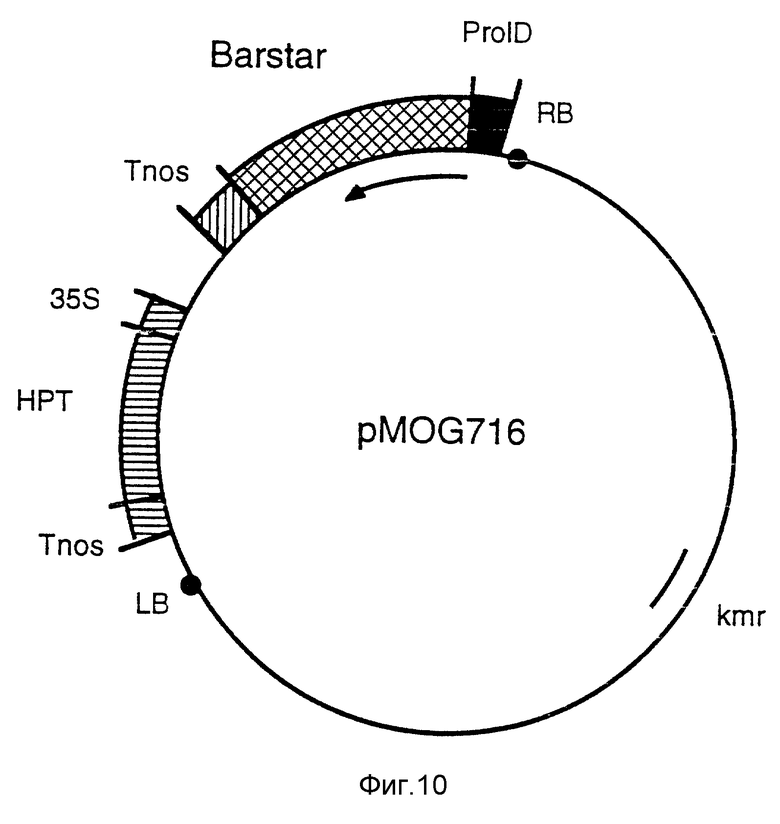
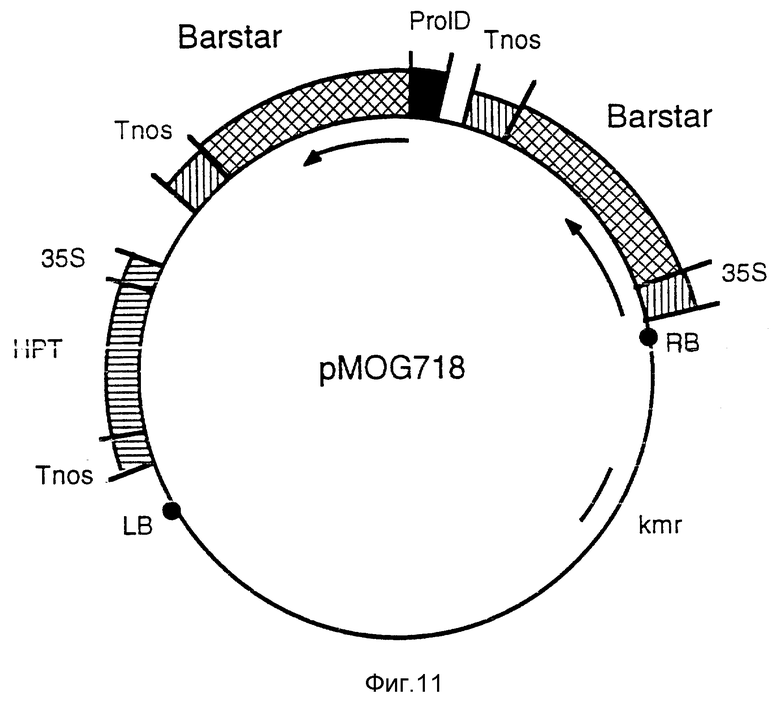
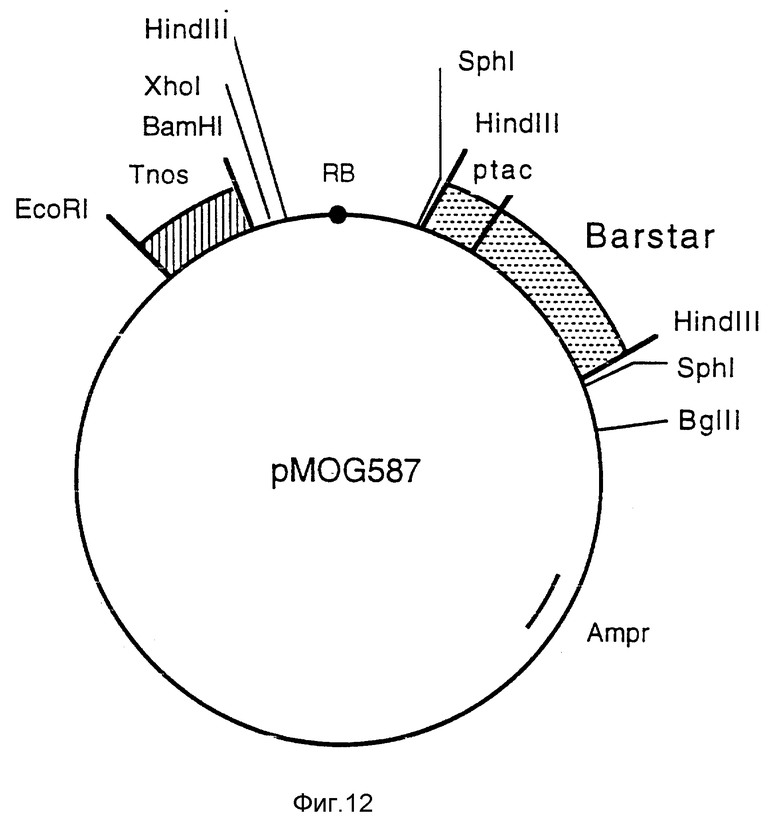
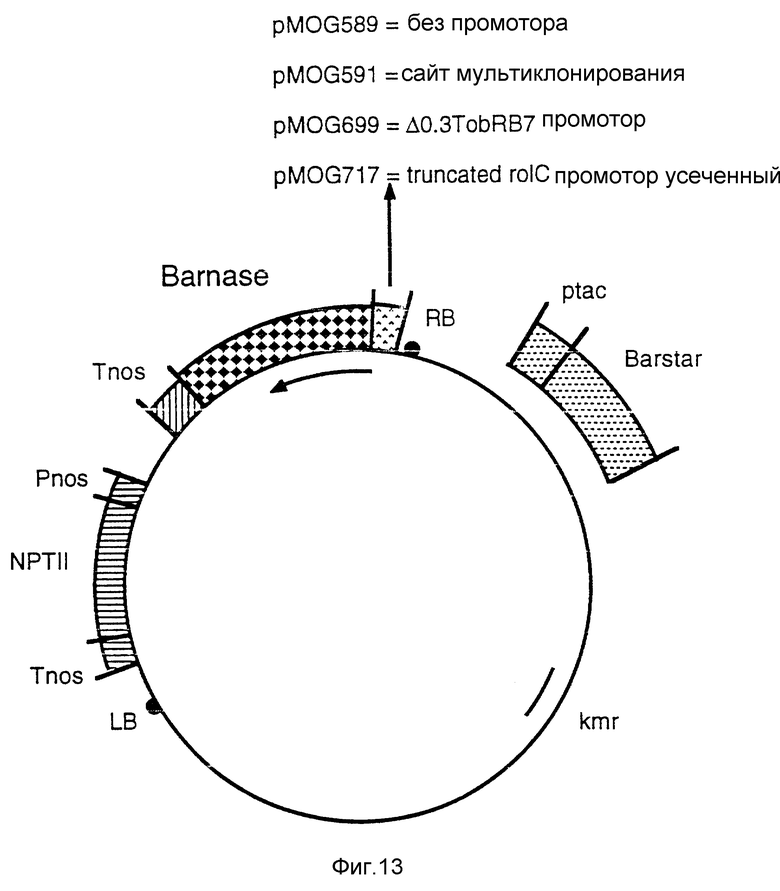
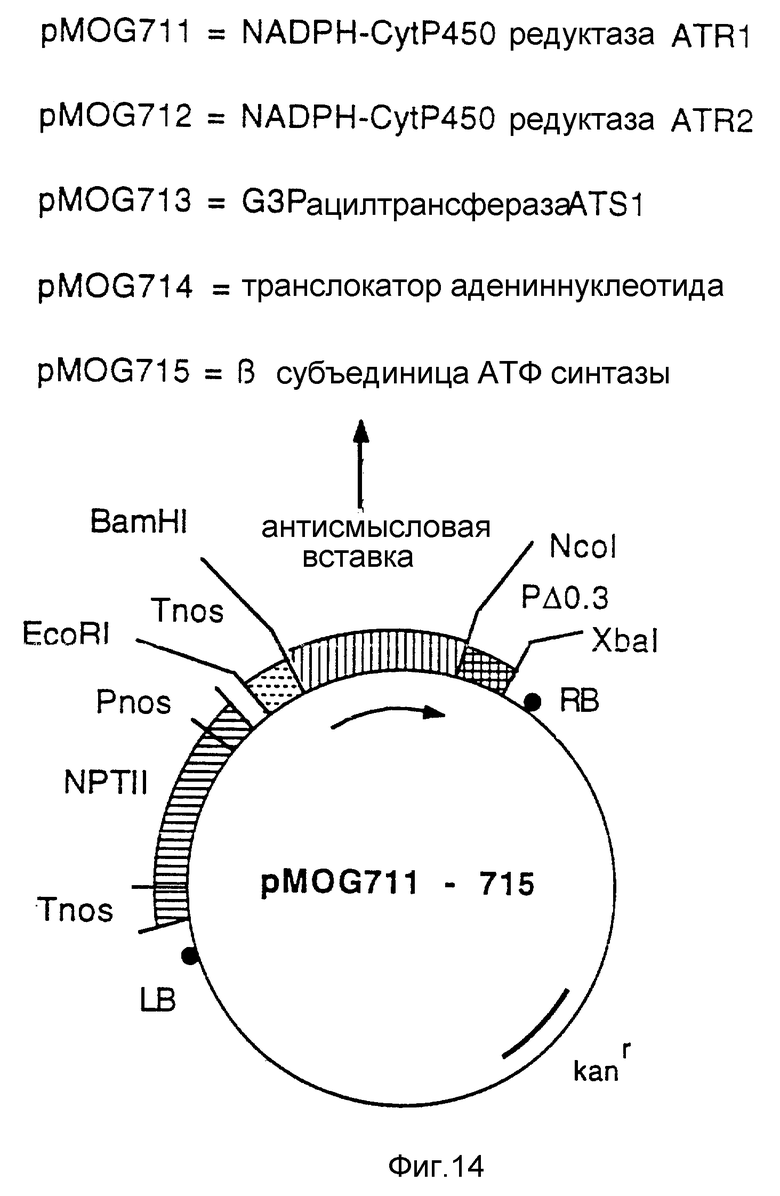
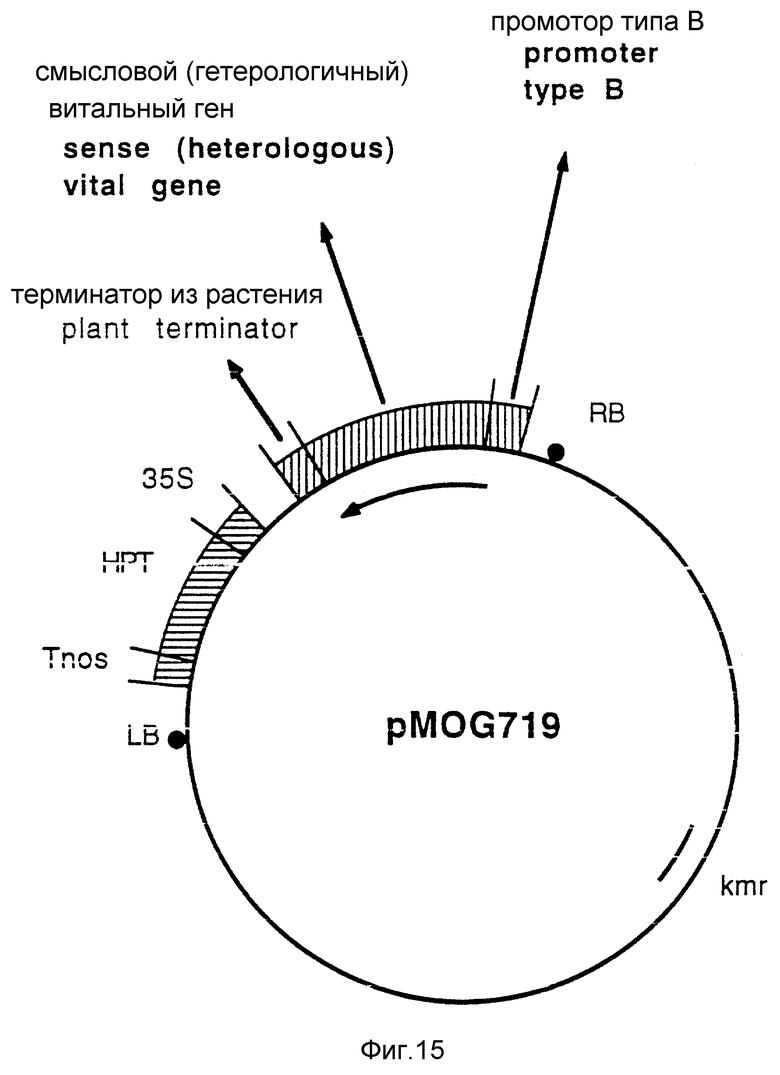
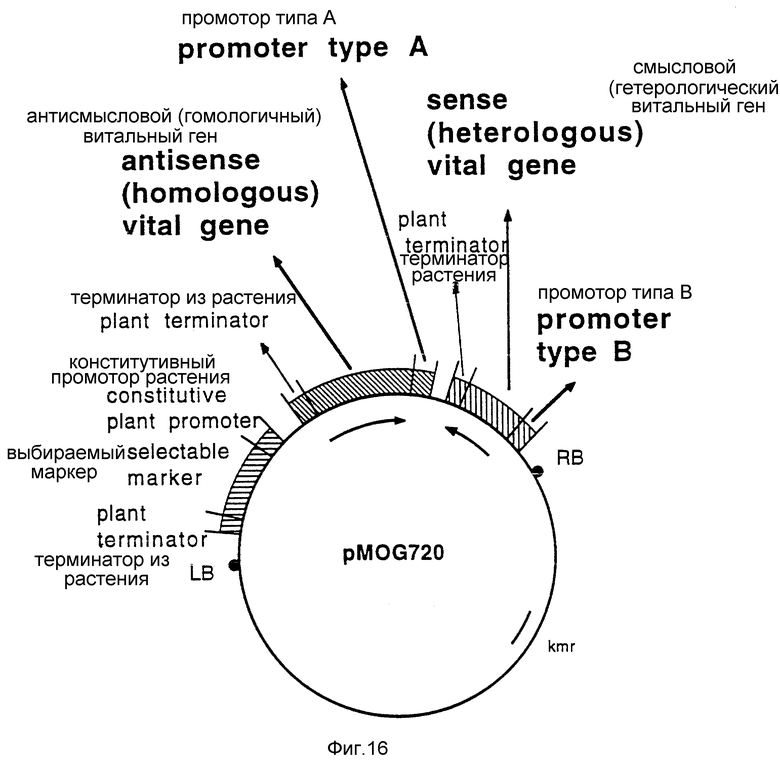
Изобретение предназначено для использования в селекции растений. Рекомбинантная ДНК содержит два гена, один из которых продуцирует вещество, разрушающее питательную среду для нематод, а другой - вещество, которое нейтрализует действие первого. Введение такой ДНК с помощью вектора или штамма Agrobacterium в растение обеспечивает снижение чувствительности трансформированного растения к нематоде за счет разрушения или торможения образования питательной среды в этом растении для нематод. 7 с. и 9 з.п.ф-лы, 16 ил. , 2 табл.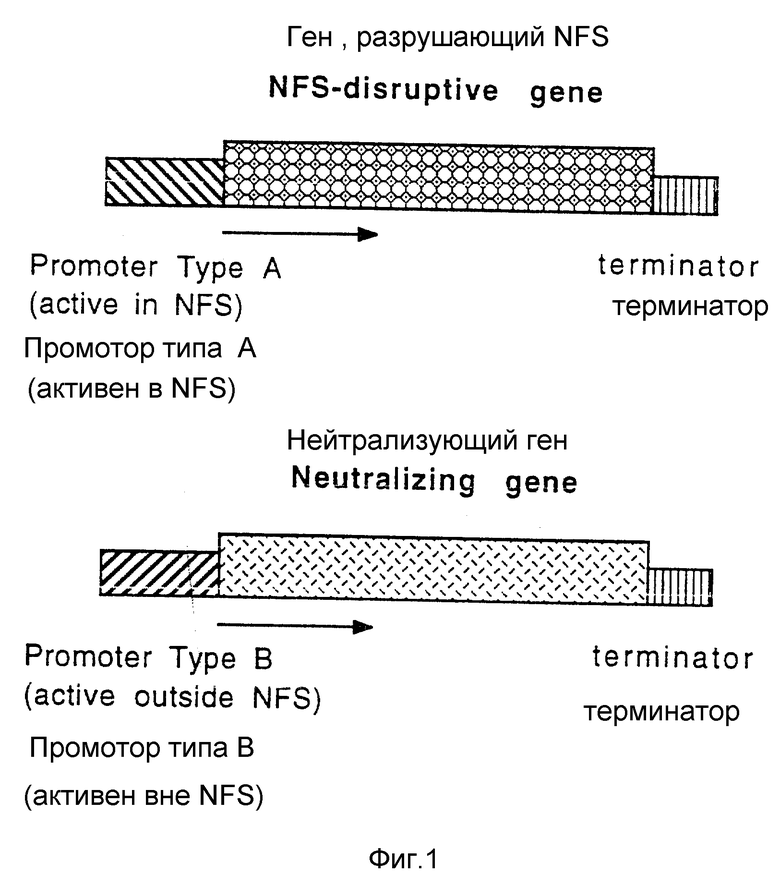
Приоритет по пунктам:
20.11.91 по пп.1, 2, 3, 4, 10, 15 и 16;
02.11.92 по пп.5, 6, 7, 8, 9, 11, 12, 13 и 14.
| СПОСОБ СОЗДАНИЯ МГНОВЕННОГО ПОДЛИВА В СКВАЖИНЕ ПРИ ИССЛЕДОВАНИИ ПОГЛОЩАЮЩИХ ПЛАСТОВ | 0 |
|
SU303426A1 |
| 1969 |
|
SU412911A1 | |
| Способ защиты растений от галловой нематоды | 1987 |
|
SU1533040A1 |
| Штамм гриба продуцент нематотоксина | 1975 |
|
SU552052A1 |

