Изобретение относится к микробиологической и медицинской промышленности и касается получения γ-аминомасляной кислоты из L-глутаминовой кислоты или ее солей с использованием биомассы бактерий Escherichia coli.
γ-Аминомасляная кислота (ГАМК), являясь эффективным нейромедиатором, применяется в медицине в составе ряда лекарственных средств (аминалон, гаммалон, пикамилон, пантогам) для лечения нарушений мозговой деятельности (расстройства памяти, речи, гемиплегии и гипертонии).
Известен химический способ получения ГАМК в ходе щелочного гидролиза пирролидонокарбоновой кислоты (Garmaise et al. Canad. J. Chemistry, 1956, 34, 743-748; А.с. СССР N 452196). Недостатком данного способа является высокая стоимость пирролидона, жесткие условия проведения синтеза, а также затрудненность гидролиза остаточного пирролидона в организме.
Известны способы получения ГАМК микробиологическим синтезом (Патенты Японии N 69 01197, N 70 15437). После 90 ч выращивания штаммов Bact. cadaveris ATCC 9760 и E. coli ATCC 9637 в культуральной жидкости накапливается до 4 г/л ГАМК. Продукт из отсепарированной при pH 2.0 культуральной жидкости выделяют методом ионообменной хроматографии на установке Diaion SKI (тип Н+). Выход кристаллического продукта из культуральной жидкости составляет 35%. Недостатком способов является низкий выход ГАМК и большая продолжительность процесса.
Известны способы получения ГАМК путем энзиматического декарбоксилирования L-глутаминовой кислоты, где в качестве катализатора реакции применяются клетки микроорганизмов, содержащие L-глутаматдекарбоксилазу (GAD, ЕС 4.1.1.15) (Патент Японии N 69 01196; Губарев E.М. и др. Биохимия. 1960. Т. 25, N 2, с. 261-263), а также иммобилизованная GAD (А.с. СССР N 799318). Недостатком способов является низкая глутаматдекарбоксилазная активность клеток и дополнительная процедура выделения и очистки GAD для иммобилизации.
Наиболее близким к предлагаемому изобретению является способ получения ГАМК декарбоксилированием L-глутаминовой кислоты клетками бактерий Arthrobacter simplex ATCC 15799 (Патент Японии 69 01196). Согласно этому способу бактериальную культуру выращивают при 30oC, pH 7.0 в течение 4-х суток на среде, содержащей 1% L-глутаминовой кислоты, 1% глюкозы, 0.05% фосфата калия двузамещенного, 0.025% сульфата магния семиводного, 0.2% сульфата аммония, 0.3% дрожжевого экстракта и 0.3 % мела. Концентрация биомассы в культуральной жидкости в конце процесса составляет 6 мг/мл. Для получения ГАМК 3 г сухой биомассы, обработанной ацетоном, вносят в водный раствор L-глутаминовой кислоты (10 г/300 мл) при pH 5.0 и температуре 37oC. Процесс декарбоксилирования ведут в течение 2-х ч до полного превращения L-глутаминовой кислоты в ГАМК при поддержании исходного значения pH автоматической подачей 3 н. соляной кислоты. По окончании процесса реакционную смесь освобождают от биомассы и выделяют ГАМК, добавляя этанол. Таким образом получают 7 г ГАМК, т.е. 1 г сухого ацетонированного порошка клеток синтезирует 2.3 г ГАМК.
Недостатками данного способа являются малое накопление биомассы, длительный период роста и высокий расход биомассы на получение ГАМК (0.43 г сухой биомассы на 1 г продукта) вследствие низкой глутаматдекарбоксилазной активности используемого штамма.
Нашей задачей было уменьшить расход биомассы на синтез ГАМК и увеличить прирост микроорганизмов, снижая тем самым удельный расход компонентов питательной среды на единицу продукта.
Задача решается созданием новых рекомбинантных штаммов бактерий Escherichia coli GAD1 и Escherichia coli GADK10, обладающих повышенной активностью глутаматдекарбоксилазы и интенсивным ростом в питательной среде.
В предлагаемом нами способе получения ГАМК расход биомассы составляет 0.04 - 0.05 г сырой биомассы на получение 1 г ГАМК.
Активность штаммов составляет 4 - 5 ед/мг сырой биомассы. (За единицу активности принимают такое количество фермента, которое образует 1 мкМ ГАМК за 1 мин при 37oC).
Продуктивность процесса культивирования микроорганизмов составляет 35 - 40 мг сырой биомассы/мл, продолжительность культивирования - 12 - 13 ч.
Штаммы Е. coli GAD1 и E. coli GADK10 получены методом генной инженерии и содержат плазмиды, несущие ген глутаматдекарбоксилазы и детерминанты устойчивости к антибиотикам. Штаммы депонированы во Всероссийской коллекции промышленных микроорганизмов и имеют регистрационные номера ВКПМ В-7460 и В-7452 соответственно.
Штаммы имеют следующие культурально-морфологические и физиолого-биохимические признаки.
Морфология клеток.
Грамотрицательные слабоподвижные палочки с закругленными концами, 1.5-2.0 мкм в длину.
Культуральные признаки.
1. Мясопептонный агар (МПА). Через 24 ч роста при 37oC образует круглые беловатые полупрозрачные колонии диаметром 1.5 - 3.0 мм; поверхность колоний гладкая, края ровные или слегка волнистые, центр колоний приподнят, структура однородная, консистенция пастообразная, легко эмульгируется.
2. Агар Лоури. Через 24 ч роста при 37oC образует круглые беловатые полупрозрачные колонии диаметром 1.5 - 3.0 мм; поверхность колоний гладкая, края ровные, структура однородная, консистенция пастообразная, легко эмульгируется.
3. Агаризованная среда Адамса. Через 36 - 48 ч роста при 37oC образует колонии диаметром 0.5 - 1.5 мм; серовато-белые, полупрозрачные, слегка выпуклые с блестящей поверхностью.
4. Рост в мясопептонном бульоне. Удельная скорость роста при 37oC - 1.2 ч-1; после 24 ч роста - сильное равномерное помутнение, характерный запах.
Физиолого-биохимические признаки.
Рост по уколу в МПА: хороший рост по всему уколу. Микроорганизм является факультативным анаэробом.
Желатину не разжижает.
Рост на молоке хороший с коагуляцией молока.
Индол не образует.
Отношение к температуре. Растет на мясопептонном бульоне при 43oC и ниже. Оптимальной является температура 37oC.
Отношение к pH среды. Растет на жидких средах с pH от 6.0 до 8.0. Оптимальное значение pH 7.0.
Отношение к источникам углерода. Хорошо растет на глюкозе, фруктозе, лактозе, маннозе, ксилозе, глицерине, манните.
Отношение к источникам азота. Усваивает азот в форме аммония, нитратов, а также азот некоторых органических соединений.
Штамм Е. coli GAD1 устойчив к ампициллину.
Штамм Е. coli GADK10 устойчив к ампициллину и канамицину.
Содержание плазмид. Клетки Е. coli GAD1 содержат многокопийную гибридную плазмиду pGAD1 (4.5 т.п.н.), обеспечивающую устойчивость к ампициллину и несущую ген gadA.
Клетки Е. coli GADK10 содержат многокопийную гибридную плазмиду pGADK10 (10.4 т.п.н.), обеспечивающую устойчивость к ампициллину и канамицину и несущую ген gadA.
Способ получения ГАМК осуществляют следующим образом. Выращивание культуры кишечной палочки ведут с использованием питательных сред, содержащих источник углерода, азота и минеральные соли. В качестве источника углерода используют глюкозу, фруктозу, сахарозу, глицерин, ацетат или субстраты, их содержащие. В качестве источника азота используют соли аммония, мочевину, гидролизаты белков и т.п. Для ускорения роста клеток и увеличения их количества в питательную среду могут быть внесены такие добавки, как пептон, кукурузный экстракт и другие принятые в микробиологии компоненты питательных сред. Для повышения активности GAD в состав сред может быть добавлен ее кофермент - пиридоксальфосфат. Выращивание клеток штаммов кишечной палочки проводят при температуре 24 - 40oC, преимущественно 36 - 38oC в течение 12-13 ч. pH среды поддерживают в пределах 5.5 - 8.0, преимущественно 6.8 - 7.0. После выращивания клетки отделяют от культуральной жидкости и используют в качестве катализатора в реакции получения ГАМК из L-глутаминовой кислоты или ее солей. Для повышения эффективности процесса трансформации клетки могут быть подвергнуты термической обработке (замораживание, нагревание) или обработке некоторыми растворителями (толуол, этилацетат и другими), а также могут быть разрушены обработкой ультразвуком или другими механическими способами.
Подготовленные, как указано выше, клетки помещают в раствор глутаминовой кислоты для проведения биотрансформации. Биотрансформацию ведут при pH 3.5 - 5.5, преимущественно 4.5 - 5.0, и температуре 30 - 50oC, оптимально 37 - 40oC. В ходе процесса концентрации субстрата и продуктов контролируют методом тонкослойной хроматографии на бумаге или пластинах, методом Варбурга или по количеству потребленного титранта (Salvadory C. et al., Ital. J. Biochem. , 1970, V. 19, N 3, 1). Процесс останавливают при полной переработке субстрата в ГАМК. Реакционную смесь сепарируют, отработанную биомассу отбрасывают, а нативный раствор передают на стадию выделения и очистки ГАМК.
Пример 1. Конструирование плазмид pGAD1 и pGAD10, содержащих ген gadA, и создание штаммов Е. coli GAD1 и Е. coli GADK10
Конструирование плазмид осуществляют следующим образом.
1. На первом этапе проводят клонирование гена gadA Е. coli при помощи метода полимеразной цепной реакции (ПЦР) на основании известной нуклеотидной последовательности (Smith et al. , J. Bacteriology, 1992, 174, 5820). Для амплификации гена используют пару праймеров GAD5 и GAD3, фланкирующих кодирующую часть гена gadA с 5' и 3'-концов соответственно.
GAD5: 5'-TTCCATATGGACCAGAAGCTGTTAACGG-3'
GAD3:5'-CAAGAGCTCTTATCAGGTGTGTTTAAAGCTGTTC-3'
Нуклеотидная последовательность праймеров включает рестрикционные сайты, необходимые для последующего клонирования в экспрессионном векторе: олигонуклеотид GAD5, содержит сайт рестрикции NdeI, а GAD3 - SacI.
В качестве матрицы используют хромосомную ДНК штамма Е. coli K12 TG1 (Maniatis T. et al., 1989, Molecular Cloning, A Laboratory Manual Cold Spring Harbor Laboratory, N. Y.).
Цикл амплификации включает в себя: денатурацию ДНК при 96oC в течение 1 минуты, отжиг праймера при 60oC в течение 1 минуты, синтез ДНК при 74oC в течение 1.5 минут. Всего проводят 35 циклов.
Фрагменты ДНК, синтезированные в ходе ПЦР, фракционируют в 1%-ном агарозном геле. ДНК, имеющую подвижность, соответствующую ожидаемому размеру гена gadA (≈ 1.4 т.п.н.), выделяют из агарозы, обрабатывают полинуклеотидкиназой фага T4 и субклонируют в векторе pUC19 (фирма "Stratagene") по SmaI сайту.
Плазмиды, содержащие необходимую вставку, отбирают при помощи рестрикционного анализа, а затем ДНК этих плазмид секвенируют методом Сэнгера (Sanger et al., Proc. Natl. Acad. Sci. USA, 1977, 74, 5463).
2. На втором этапе работы фрагмент ДНК с геном глутаматдекарбоксилазы встраивают в вектор pGEMEX1.
Экспрессионный вектор pGEMEX1 (фирма "Promega") обеспечивает устойчивость к ампициллину и несет полноразмерный ген 10 бактериофага T7. Непосредственно между стоп-кодоном и терминатором гена 10 встроен полилинкер с рядом уникальных сайтов рестрикции, в том числе и сайтом SacI. В месте инициирующего кодона, включая его, имеется сайт рестрикции NdeI. Замещение части гена 10, заключенного между рестрикционными сайтами NdeI и SacI, на ген gadA дает возможность использовать для экспрессии последнего все необходимые регуляторные элементы транскрипции и трансляции гена 10.
а) Отобранные на первом этапе плазмиды с геном gadA гидролизуют рестриктазами NdeI и SacI, затем выделяют фрагмент ДНК размером 1.4 т.п.н., содержащий указанный ген;
б) ДНК вектора pGEMEX1 рестрицируют рестриктазами AatII и SacI. Меньший из фрагментов ДНК длиной 1063 т.п.н. выделяют из агарозы;
в) ДНК вектора pGEMEX1 гидролизуют рестриктазами AatII и NdeI. Больший из фрагментов ДНК длиной 2098 т.п.н. выделяют из агарозы.
Полученные на этапах "а", "б" и "в" фрагменты ДНК смешивают, обрабатывают T4 ДНК-лигазой и трансформируют штамм Е. coli К-12 XL-1 Blue (фирма "Stratagene"). С помощью рестрикционного анализа среди трансформантов отбирают клоны с рекомбинантной плазмидой, содержащей вставку с геном gadA. Полученная плазмида (4.5 т.п.н.) получила название pGAD1.
Для экспрессии гена глутаматдекарбоксилазы рекомбинантную плазмиду pGAD1 переносят в специальный штамм Е. coli В BL21 (DE3) (фирма "Novagen"), содержащий хромосомную копию гена РНК-полимеразы фага T7. Таким образом получают рекомбинантный штамм Е. coli BL-21 (DE3) [pGAD1], в дальнейшем называемый Е. coli GAD1, устойчивый к ампициллину.
Активность штамма составляет 4.0 - 4.5 ед/мг сырой биомассы.
3. На этом этапе работы получают рекомбинантную плазмиду, содержащую ген gadA и несущую детерминанты устойчивости к двум антибиотикам - ампициллину и канамицину для удобства дальнейшей работы со штаммами-продуцентами ГАМК. Для этого:
а) плазмиду pGAD1 гидролизуют рестриктазой BamHI.
б) вектор pUC4K, содержащий ген neo, обеспечивающий устойчивость к канамицину и неомицину (фирма "Pharmacia"), гидролизуют той же рестриктазой. Меньший фрагмент размером 1.3 т.п.н. выделяют из агарозы.
Полученные на этапах "а" и "б" фрагменты ДНК смешивают, обрабатывают T4 ДНК-лигазой и полученной лигированной смесью проводят трансформацию штамма Е. coli K12 XL-Blue. Отбор трансформантов проводят по признаку устойчивости к ампициллину и канамицину.
Рестрикционный анализ показал, что устойчивые к двум антибиотикам трансформанты несут рекомбинантные плазмиды, содержащие исходную плазмиду pGAD1 и BamHI - BamHI фрагмент вектора pUC4K с геном neo в разных сочетаниях. Так, имелись плазмиды с одним фрагментом neo, встроенным в плазмиду pGAD1 в разных ориентациях, плазмиды, содержащие два BamHI - BamHI фрагмента с геном neo и одну копию pGAD1, а также структура с двумя последовательно расположенными плазмидами pGAD1 и одним фрагментом с геном neo. После введения полученных плазмид в экспрессионный штамм Е. coli В BL-21 (DE3) и оценки уровня продукции ГАМК отбирают штамм, несущий плазмиду pGADK10 (10.4 т.п.н.) с тандемным повтором исходной плазмиды pGAD1 и одним BamHI - BamHI фрагментом, содержащим ген neo, как обеспечивающий наиболее высокий уровень продукции. Таким образом получают рекомбинантный штамм Е. coli BL-21 (DE3) [pGADK10], в дальнейшем называемый Е. coli GADK10, устойчивый к ампициллину и канамицину.
Активность штамма составляет 4.5 - 5.0 ед/мг сырой биомассы.
Пример 2. Процесс получения γ-аминомасляной кислоты.
1. Получение биомассы клеток Е. coli GAD1 и Е. coli GADK10.
Односуточные колонии клеток Е. coli GAD1 и Е. coli GADK10, выращенные раздельно на МПА, вносят стерильно в колбы Эрленмейера, содержащие по 100 мл питательной среды (по 10 колб для каждой культуры). В качестве питательной среды используют L-бульон с внесенным антибиотиком: 0.01 г/л ампициллина для Е. coli GAD1 и 0.005 г/л канамицина для Е. coli GADK10. Посевные материалы выращивают 6 ч на лабораторной качалке при начальном значении pH 7.3, температуре 37oC и частоте вращения 200 об/мин. Затем культуры вносят в количестве 5 об.% в ферментеры фирмы "Marubishi" объемом 30 л, содержащие 20 л питательной среды следующего состава, г/л:
1. глюкоза - 1.00;
2. фосфат калия однозамещенный - 0.20;
3. аминокислотный гидролизат - 1.50;
4. сульфат магния - 0.05;
5. хлорид аммония - 0.10;
6. пептон - 0.50;
7. пиридоксальфосфат - 0.0025;
8. вода водопроводная - до 20 л.
Кроме того, в ферментационную среду для штамма Е. coli GAD1 входит 0.01 г/л ампициллина, в среду для штамма Е. coli GADK10 - 0.005 г/л канамицина.
Аминокислотный гидролизат, фосфат калия и пептон стерилизуют в ферментере при 0.8 ати в течение 45 мин; остальные компоненты стерилизуют отдельно.
Глюкозу, глюкозоаммиачную смесь, сульфат магния и хлорид аммония стерилизуют в автоклаве при 0.5 ати в течение 30 мин. Пиридоксальфосфат растворяют в 100 мл воды и проводят стерилизующую фильтрацию. Антибиотики растворяют в стерильной воде. Компоненты вносят в питательную среду непосредственно перед засевом.
Ферментацию ведут при 37oC, pH 7.0 ± 0.1, избыточном давлении 0.3 ати и аэрации с расходом воздуха 1 л/(л х мин) в течение 13 ч. Автоматическое поддержание pH осуществляется подачей стерильной глюкозоаммиачной смеси следующего состава:
1. глюкоза - 450 г/л;
2. аммиак - 62.5 г/л;
3. вода водопроводная - до 4-х л.
В конце процесса концентрация сырой биомассы Е. coli GAD1 составляет 35 г/л, Е. coli GADK10 - 38 г/л.
Культуральные жидкости охлаждают до 15oC и центрифугируют на молочном сепараторе АСГ-3М при 6000 об/мин. Сырая масса клеток обоих штаммов составляет 700 ± 50 г. Полученные биомассы распределяют по полиэтиленовым пакетам по 100 г и замораживают при -20oC в морозильной камере "Минск-17" с выдержкой 24 ч.
2. Подготовка биомассы.
С целью увеличения ферментативной активности биомассы проводят ее термоактивацию.
Активность глутаматдекарбоксилазы определяют по количеству образовавшейся ГАМК после 15-минутной инкубации клеток с концентрацией 0.25 мг сырой биомассы/мл в 1 мл 0.01%-ного раствора L-глутаминовой кислоты при 37oC и pH 4.6. Реакцию останавливают 30 мкл 30%-ного раствора едкого натра.
Концентрацию образовавшейся ГАМК анализируют методом тонкослойной хроматографии на бумаге FN-11 (Германия) с использованием системы бутанол - ледяная уксусная кислота - вода дистиллированная (4 : 1 : 2).
Элюцию ведут в течение 15 ч в хроматографической камере, после чего бумагу обрабатывают 0.5%-ным раствором нингидрина в ацетоне и помещают в термостат на 30 мин при 60oC.
Проявленное хроматографическое пятно вырезают и помещают в 5 мл 0.5%-ного раствора хлорида кадмия на 1 час при 60oC. Оптическую плотность раствора измеряют на спектрофотометре СФ-26 на длине волны 540 нм. Концентрацию ГАМК вычисляют как отношение оптической плотности пробы к оптической плотности стандартного 0.1%-ного раствора ГАМК.
Активность глутаматдекарбоксилазы у штамма Е. coli GAD1 составила 4 ед/мг сырой биомассы, у штамма Е. coli GADK10 - 5 ед/мг сырой биомассы.
12.5 г замороженной биомассы Е. coli GAD1 и 10 г Е. coli GADK10 разводят по отдельности в двух порциях по 60 мл 0.9%-ного раствора хлорида натрия и термостатируют 40 мин при 53oC в двух реакторах фирмы "Bioflo" объемом 500 мл. Затем температуру снижают до 37oC и поддерживают это значение в течение всего процесса биотрансформации.
3. Биотрансформация.
Процесс получения ГАМК ведут с использованием в качестве сырья глутамата натрия или L-глутаминовой кислоты.
Теоретический выход ГАМК при полной конверсии субстрата составляет:
- из глутамата натрия - 55 мас.%;
- из L-глутаминовой кислоты - 70 мас.%.
Практический выход ГАМК на стадии биотрансформации в обоих случаях очень близок к теоретическому. Остаточная концентрация субстрата не превышает 0.5 г/л.
При использовании глутамата натрия проводят предварительную модификацию сырья путем перевода глутамата натрия в L-глутаминовую кислоту с целью предотвращения образования натриевой соли ГАМК в ходе реакции трансформации. При использовании в качестве сырья L-глутаминовой кислоты эта стадия исключается из технологической схемы.
500 г глутамата натрия растворяют в 1500 мл водопроводной воды (pH 7.7) и добавляют 160 мл ледяной уксусной кислоты при интенсивном перемешивании. Образуется густая белая суспензия L-глутаминовой кислоты (pH 4.5). Реакция представлена уравнением 1.
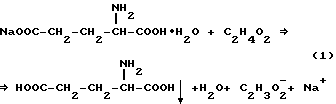
Далее суспензию фильтруют через фильтровальную бумагу, раствор сливают, а осадок L-глутаминовой кислоты передают на стадию биотрансформации. На стадии модификации сырья получают 470 г осадка L-глутаминовой кислоты с влажностью 30% (или 360 г L-глутаминовой кислоты). Потери L-глутаминовой кислоты в фильтрате определяются ее растворимостью и составляют 30 г. Процесс подготовки субстрата ведут аналогично для двух штаммов.
Биотрансформацию ведут в двух реакторах фирмы "Multigen" объемом 2 л каждый при pH 4.6 ± 0.1, температуре 37oC и минимальном перемешивании. Значение pH поддерживается ледяной уксусной кислотой.
Реакция представлена уравнением 2.

В реакторы загружают по 500 мл водопроводной воды и по 230 г полученной на предыдущей стадии сырой L-глутаминовой кислоты (первая порция). Вносят по 10 мл клеточных суспензий, приготовленных на стадии 2. В дальнейшем, начиная со второго часа, суспензии биомассы подают перистальтическими насосами фирмы "Verder" со скоростью 5 мл/час в течение процесса. Через 5 часов вносят еще по 230 г L-глутаминовой кислоты.
Полноту переработки субстрата определяют по стабилизации pH реакционной смеси и по результатам тонкослойной хроматографии. Конечные реакционные объемы составляют 1000 мл, pH 5.0. За 15 часов процесса в реакционных смесях накапливается 260 г/л ГАМК. Остаточная концентрация L-глутаминовой кислоты в растворах составляет 0.5 г/л.
Таким образом, на стадии биотрансформации из 500 г глутамата натрия получают 260 г ГАМК. То есть, по отношению к глутамату натрия выход составляет 52 мас.%. Расход биомассы Е. coli GAD1 составляет 0.05 г сырой биомассы/г ГАМК, Е. coli GADK10 - 0.04 г сырой биомассы/г ГАМК.
4. Выделение и очистка ГАМК.
Процесс выделения и очистки ГАМК состоит из следующих этапов:
1. микрофильтрация реакционной смеси;
2. ультрафильтрация раствора ГАМК;
3. упаривание раствора ГАМК в вакуумной сушилке;
4. кристаллизация ГАМК в этаноле и сушка кристаллов.
Микрофильтрацию реакционной смеси проводят на мембранном фильтре фирмы "Владипор" с диаметром пор 0.45 мкм.
Ультрафильтрацию раствора проводят на установке "Pellicon" фирмы "Millipor", снаряженной кассетой 5.0 SQ.FT с диаметром пор 10 kD, при температуре 10oC и давлении 2 ати.
Упаривание раствора ГАМК проводят на вакуум-выпарной установке LPH 03125, производства Венгрии, при остаточном давлении 100 мбар и температуре 60oC до остаточной влажности 6%.
Кристаллизацию ГАМК проводят в 94%-ном этаноле. 300 г упаренной смеси разводят в 1200 мл этанола при температуре 5oC. После выпадения кристаллов ГАМК маточный спиртовой раствор сливают и проводят отгонку этанола. Кристаллы ГАМК сушат на вакуум-выпарной установке LPH 03125 при остаточном давлении 100 мбар и температуре 60oC в течение 6 ч, взвешивают и проводят анализ полученного препарата согласно ФС 42-1903-82.
Результаты выделения и очистки ГАМК приведены в таблице.
Таким образом, получено по 200 г сухого препарата ГАМК, содержащего 98.7% основного вещества. Удельный расход сырой биомассы на единицу продукта составляет 0.05 г/г для штамма Е. coli GAD1 и 0.04 г/г для штамма Е. coli GADK10. При использовании в качестве сырья глутамата натрия общий выход ГАМК - 40 мас.%; при использовании в качестве сырья L-глутаминовой кислоты - 55.6 мас.%.

Предлагается способ получения γ-аминомасляной кислоты (ГАМК) биотрансформацией L-глутаминовой кислоты или ее солей в присутствии биомассы бактерий Escherichia coli ВКПМ В-7460 или Escherichia coli ВКПМ В-7452 с последующей очисткой и выделением продукта. Декарбоксилазная активность клеток составляет 5 мкМ ГАМК/(мин•мг сырой биомассы). Удельный расход клеток в процессе биотрансформации составляет 40 мг сырой биомассы на 1 г ГАМК. Выход конечного продукта из глутаминовой кислоты составляет 55,6 мас.%, из глутамата натрия - 40 мас.%. Содержание ГАМК в сухом препарате -98,7%. 1 табл.
Способ получения γ-аминомасляной кислоты биотрасформацией L-глутаминовой кислоты или ее солей в присутствии биомассы бактерий, обладающих глутаматдекарбоксилазной активностью, с последующим выделением и очисткой целевого продукта, отличающийся тем, что используют штаммы бактерий Escherichia coli ВКПМ В-7460 или Escherichia coli ВКПМ В-7452.
| Приспособление для плетения проволочного каркаса для железобетонных пустотелых камней | 1920 |
|
SU44A1 |
| Способ получения @ -аминомасляной кислоты | 1979 |
|
SU799318A1 |
| Приспособление для плетения проволочного каркаса для железобетонных пустотелых камней | 1920 |
|
SU44A1 |
| Canad | |||
| J | |||
| Chemistry, 1956, 34, 743-748. | |||

