Изобретение относится к улучшенной вакцине для иммунизации против TBE-вирусных инфекций, а также к способу ее получения.
TBE-Вирус (вирус клещевого энцефалита) распространен во многих европейских странах, бывшем Советском Союзе и Китае. В некоторых странах Центральной Европы, таких как Австрия, Чехия, Словакия, Словения или Венгрия, где каждый год регистрируется несколько сотен установленных в больницах случаев, заболевание представляет собой значительную проблему для общественного здравоохранения (WHO: EURO Reports and Studies 104; 1983).
TBE-Вирус, который существует в форме западного (европейского) подтипа и дальневосточного подтипа, относится к семейству флавивирусов, которые представляют собой сферические, окруженные липидной оболочкой РНК-вирусы (см. Monath T.P.: Flaviviruses, в Fields B.N. (изд.) Virology, Raven Press, N.Y. 1990, с. 763-814).
Вирион флавивируса в общем состоит из нуклеокапсида, который содержит геном плюс-нитьевой РНК в сочетании с вирусным капсидным (C)-протеином. Нуклеокапсид окружен липидной оболочкой, которая содержит ассоциированные с мембраной протеины E (50 - 60 кД) и M (7 - 8 кД) и (Heinz и Roehrig; в: van Regenmortel und Neurath (Hrsgb)". Immunochemistry of Viruses 11. The Basis for Seradiagnosis and Vaccines. Elsevier Sciences, Amsterdam, (1990), 289 - 305).
Основной протеин оболочки E играет центральную роль в биологии флавивирусов тем, что он выполняет значительные функции при проникновении (внедрении) вируса и индицирует защитный иммунный ответ у хозяина. Имеется уже значительное количество информации относительно структуры протеина оболочки E TBE-вируса и предложена структурная модель на основе множества биохимических и иммунологических данных (Mandl и др., J. Virol. 63, (1989), 564 - 571).
Заболевание можно эффективно предотвращать за счет вакцинации с помощью высокоочищенной вакцины, приготовленной инактивированием формалином полного вируса (см. Kunz и др., J. Med. Virol., 6 (1980), 103 - 109), которая индуцирует иммунный ответ против структурных протеинов вируса (Kunz, Ch. Acta leidensia 60, N 2, 1 - 14, 1992). Эта вакцина оказывается наиболее пригодной, однако в процессе ее получения нужно манипулировать с большими объемами инфекционных и потенциально опасных суспензий вируса. Поэтому необходимы дорогостоящие меры безопасности.
Способность антител нейтрализовать вирус зависит от того, насколько эффективно они распознают нативную структуру протеинов на поверхности вируса. В случае TBE-вирусов и других флавивирусов речь идет при этом в первую очередь о протеине E (см. Heinz и Mandl, APMIS, 101 (1994), 735 - 745). Для индукции по возможности эффективно действующих антител в процессе иммунизации желательно, чтобы вакцина содержала этот протеин в такой же форме, в которой он также находится на поверхности инфекционного вируса. Убитые вакцины имеют тот недостаток, что применяемый способ инактивации может приводить к частичному изменению нативной структуры протеина.
Техника рекомбинантных ДНК дает возможность заменять убитые вакцины на рекомбинантные протеины, которые содержат значительные доли индуцирующих иммуноответ протеинов. При этой генно-инженерной экспрессии отдельных протеинов вируса, однако, не гарантируется, что структура антигена этих рекомбинантных протеинов соответствует таковой соответствующих протеинов на поверхности вируса.
Экспрессия рекомбинантных поверхностных протеинов TBE-вирусов известна, например, из Allison и др., Gemeinsame Jahrestagung "OBG - "OGGGT (1993), р. 114. При этом установлено, что протеин E и протеин M при их рекомбинантной экспрессии при определенных условиях выделяются в различной форме, в том числе также в форме неинфекционных субвирусных частиц.
Такие субвирусные частицы известны для других членов семейства, особенно таких как вирус японского эмцефалита (JEV), вирус желтой лихорадки и вирус лихорадки Денге (см. Konishi и др., Virology, 188 (1992), 714 - 720, или ВОИС 92/03545).
Хотя у Konishi и др. описывается, что такие субвирусные частицы, которые эмульгированы в полном адъюванте Фрейнда и содержат весь протеин E, вызывают известный иммуноответ у мышей, однако, с другой стороны, обнаружено, что с помощью частично укороченного на C-конце протеина E можно достигать значительно более эффективной защиты, чем с помощью всего протеина E, против инфекции флавивирусом (ВОИС 92/03161).
Задача настоящего изобретения состоит в том, чтобы получить улучшенную вакцину против TBE-инфекций.
Эта задача согласно изобретению решается за счет вакцины для иммунизации против вируса клещевого энцефалита (TBE-вирус), которая включает неинфекционные, субвирусные частицы, содержащие по существу протеин E и в его полноценной нативной форме и в случае необходимости протеин prM/M, которые производятся от TBE-вируса. При этом существенно, чтобы протеин E находился в своей полноценной (полной) нативной форме, так как только с его помощью можно достигать эффективной защиты. В предлагаемой согласно изобретению вакцине нативную форму протеина E можно обнаруживать с помощью различных анализов, особенно:
а). Анализ антигенной структуры с помощью моноклональных антител;
б). Способность к индуцируемым кислотой изменениям конформации, и
в). Гемаглютинационная активность.
Тот факт, что предлагаемая согласно изобретению вакцина на основании своего состава не может быть инфекционной, представляет собой важный аспект для безопасности прививочного материала.
Предпочтительно протеин E и в случае необходимости протеин prM/M представляют собой рекомбинатные протеины.
Согласно особенно предпочтительной форме вакцины, протеин E производится от TBE-вируса, причем - в зависимости от области применения - используется как западный (европейский) подтип, так и дальневосточный подтип. Предпочтительно вакцина включает еще липидную компоненту, которая предпочтительно находится в везикулярной форме.
Оказалось, что - в противоположность мнению специалистов (см. ВОИС 92/03161) - рекомбинантный протеин E, который производится от TBE-вирусов, только в форме этих неинфекционных субвирусных частиц может обеспечивать достаточную иммунизацию против инфекций. Частично укороченная форма протеина E, при которой, как указано в ВОИС 92/03161, удален C-концевой мембранный якорь, не может применяться для получения эффективной вакцины. Предлагаемая согласно изобретению вакцина содержит предпочтительно неинфекционные частицы, которые по существу лишены определяемых посредством PCR, происходящих от TBE-вируса нуклеиновых кислот. Это можно показать, например, с помощью описанных у Konishi и др. методов.
Согласно другому аспекту, изобретение относится к способу получения TBE-вакцины, который отличается тем, что:
- используют систему культуры клеток, которая содержит кодирующие последовательности для протеинов prM и E, производимых от TBE-вируса;
протеин E экспрессируют в его полноценной, нативной форме; причем образуются субвирусные, неинфекционные частицы, которые содержат по существу рекомбинантный протеин E в его полноценной нативной форме и в случае необходимости рекомбинантный протеин prM/M; и частицы собирают, а также перерабатывают до пригодного для иммунизации состава.
Предлагаемый согласно изобретению способ имеет большое преимущество, заключающееся в том, что нет необходимости в стадии инактивирования вирусов, например, за счет формалина, что, с одной стороны, значительно улучшает качество вакцины (протеин E находится в нативной форме, а не в измененной за счет обработки формалином и по меньшей мере частично денатурированной форме) и, с другой стороны, отчетливо облегчает технологическое получение этой вакцины, так как частицы можно прямо (следовательно, без обработки формалином) перерабатывать в фармацевтический препарат.
При осуществлении способа особенно предпочтительно использование системы культуры клеток, которая содержит кодирующие последовательности для рекомбинантных протеинов prM и E из TBE-вируса в интегрированной форме в хромосоме.
Однако системы культуры клеток, в которых используют вирусные векторы, или системы культуры клеток, в которых работают без вируса, например, с плазмидным вектором, также пригодны в случае необходимости для получения предлагаемых согласно изобретению частиц.
Согласно предпочтительному варианту осуществления способа как экспрессию протеинов, так и образование частиц осуществляют непрерывно.
Согласно следующему аспекту, изобретение относится к применению неинфекционных субвирусных частиц, содержащих в основном протеин E в его полноценной, нативной форме и в случае необходимости протеин prM/M, которые производятся от TBE-вируса, для получения вакцины в целях активной иммунизации против вызываемых TBE-вирусом инфекций.
При этом протеин E и в случае необходимости протеин prM/M представляют собой предпочтительно рекомбинантные протеины.
Далее, изобретение относится к применению в медицине неинфекционных субвирусных частиц, содержащих по существу протеин E в его полноценной, нативной форме и в случае необходимости протеин prM/M, которые производятся от TBE-вируса, в особенности для получения анти-TBE-вирусных иммуноглобулиновых препаратов. Также здесь предпочтительно, чтобы протеин E и в случае необходимости протеин prM/M являлись рекомбинантными протеинами.
Неожиданно найдено, что нуклеиновая кислота, которая содержит упомянутые кодирующие последовательности как таковые, может применяться для иммунизации против TBE-вирусных инфекций.
Из уровня техники известно, что введение "голой" ДНК мышам может вызывать у них иммуноответ. Например, мыши, которым ввели путем инъекции плазмиду, содержащую геномную копию человеческого гормона роста (hGH), вырабатывают антитела против человеческого hGH (Nature, 356 (1992), 152 - 154).
Далее, описаны некоторые успешные "генетические иммунизации" благодаря "голой" ДНК. При этой генетической иммунизации вводят ДНК, которая кодирует один или несколько антигенов вируса, после чего ин виво синтезируются соответствующие вирусные антигены, которые, каждый отдельно, вызывают иммуноответ и таким образом в дальнейшем может вызываться защита от вирусных инфекций. Успешный защитный иммунитет у мышей за счет внутримышечной инъекции ДНК вируса гриппа описывается в PNAS 91 (1994), с. 9519 - 9523 (Raz и др.). Также успешная иммунизация крыс и мышей против вируса гепатита B (HBV) за счет внутримышечной инъекции плазмидной ДНК, которая содержит последовательность, кодирующую HBV-поверхностный антиген, описывается в Vaccine, 12 (16), (1994), с. 1503 - 1509 (Davis и др.). Многие опыты по иммунизации с помощью "голой" ДНК, кодирующей патогенные антигены, однако, остаются безуспешными. Например, хотя описаны опыты по иммунизации против ВИЧ (вирус иммунодефицита человека) посредством прямого переноса ДНК в клетки организма (ВОИС 93/17706), до сих пор, однако, с помощью этой методики не смогли получить удачную вакцину. В особенности оказывается, что для успешного иммунизирующего действия чистой ДНК-вакцины - наряду с введением ДНК в клетку - особенно предпочтительно, чтобы взывающий иммуноответ антиген находился в нативной форме в иммунной системе. Точная структура нативной формы, соответственно, биосинтетические процессы, которые необходимы для образования нативной формы и структуры, гарантированы только для немногих патогенов, так что эффективная иммунизация с помощью "голой" нуклеиновой кислоты во многих случаях крайне затруднительна, если вовсе - на основании недостаточно точных знаний антигенной структуры - не реализуема при использовании современного уровня знаний.
В рамках настоящего изобретения подчеркивается, что иммунизация с помощью последовательности нуклеиновой кислоты, которая кодирует протеин вызывающий значительный иммунный ответ к TBE-инфекции, недостаточна, чтобы получить иммуноответ, который защищает от заболевания.
Неожиданно смогли получить успешный иммуноответ только за счет введения последовательности нуклеиновой кислоты, которая наряду с кодирующей последовательностью для протеина E в его полноценной, нативной форме включает еще кодирующую последовательность для протеина prM/M. Далее, оказалось, что с помощью нуклеиновой кислоты, содержащей кодирующую протеин E последовательность, которая имеет делецию якорной области протеина E, также нельзя достигать никакой успешной иммунизации.
Настоящее изобретение поэтому, согласно следующему аспекту, относится к вакцине для иммунизации против инфекций, вызываемых вирусом клещевого энцефалита (TBE-вирусом), включающей нуклеиновую кислоту, которая кодирует протеин E и протеин prM/M, производимые от TBE-вируса, в их по крайней мере главным образом полноценной, нативной форме.
С помощью настоящего изобретения впервые, в общем, смогли показать иммунизацию за счет нуклеиновой кислоты для флавивирусов. Предлагаемая согласно изобретению иммунизирующая система поэтому в принципе применима не только для иммунизации против TBE-вируса, но и из-за большой гомологии всех флавивирусов в отношении протеина E и prM/M (см. Chambers и др., Annual Revies of Microbiology, т. 44, 1990, с. 649 - 688: Flavivirus Genome Organisation, Expression and Replication) - вообще для иммунизаций против флавивирусных инфекций.
Существенным для вакцины на основе нуклеиновой кислоты для иммунизации против TBE-вируса является то, что протеин E в своей по существу полноценной нативной форме кодируется нуклеиновой кислотой. Само собой разумеется, под настоящее изобретение также подпадают вакцины с нуклеиновыми кислотами, которые получаются за счет вырожденности генетического кода, или природных, или получаемых в лаборатории мутаций, пока с помощью этих модификаций нуклеиновой кислоты относительно кодирующей последовательности протеина E можно вызывать успешный иммуноответ.
Вакцины, содержащие нуклеиновые кислоты с делениями или инсерциями в кодирующей последовательности протеина E, которые по существу оставляют неизменной необходимую для иммунизации структуру протеина E в антигенной области, также нужно рассматривать как входящие в рамки изобретения, так как они имеют значение в качестве производных от TBE-вирусной последовательности. Соответственно это имеет значение, само собой разумеется, также для кодирующей последовательности протеина prM/M. Так как протеин prM/M имеет значение для надежного образования и секреции субвирусной частицы, отклонения в последовательности относительно протеина prM/M не имеют такого решающего значения, как в случае протеина E, пока обеспечивается успешная "сборка" частицы. Природа нуклеиновой кислоты согласно изобретению несущественна. В равной мере могут применяться РНК и ДНК, причем из-за повышенной стабильности, и противоположность РНК, в некоторых областях применения отдают предпочтение ДНК. Нуклеиновую кислоту можно получать в равной мере биологически или путем химического синтеза.
Предпочтительно кодирующая протеин E нуклеиновая кислота производится от европейского или дальневосточного подтипа TBE-вируса, так как это особенно распространенные подтипы.
Особенно предпочтительными вакцинами на основе нуклеиновой кислоты являются такие, в которых нуклеиновая кислота представляет собой плазмидный вектор. В качестве особенно пригодных плазмидных векторов следует особенно отметить такие, которые содержат сильные промоторы, как, например, HSV-; RSV-; EBV-; β- актин; адено-; MMTV-; hsp и hGH-промотор. Сильные промоторы обеспечивают эффективную экспрессию гена.
К пригодным векторам, соответственно, промоторам, причисляют производные от плазмиды pCMV β (Mac Gregor и др.) плазмиды, которые содержат ранний CMV-промотор.
Само собой разумеется, нуклеиновые кислоты предлагаемых согласно изобретению вакцин на основе нуклеиновой кислоты могут включать еще другие кодирующие или некодирующие последовательности, в особенности когда в распоряжении имеется нуклеиновая кислота в виде вектора нуклеиновой кислоты.
Примерами таких других последовательностей - наряду с промоторами - являются еще маркерные гены, другие регуляторы относительно транскрипции, трансляции или пострансляционных модификаций и т.д. В случае этих более сложных конструкций нуклеиновой кислоты, однако, всегда нужно обращать внимание на то, чтобы после введения нуклеиновой кислоты в целевую клетку путем трансляции нуклеиновой кислоты не мог образовываться никакой инфекционной вирус, следовательно, чтобы вакцина была "убитой", соответственно, оставалась убитой вакциной.
Предлагаемая согласно изобретению вакцина на основе нуклеиновой кислоты в самой простейшей форме может содержать "голую" нуклеиновую кислоту, в случае необходимости в пригодной буферной системе. Также, само собой разумеется, могут присутствовать другие составные части, как технологические в случае лекарственных средств добавки, носители или особые, специфические для введения вещества.
Введение предлагаемой согласно изобретению вакцины на основе нуклеиновой кислоты предпочтительно осуществляют путем инъекции, в особенности путем внутримышечной, внутрикожной или подкожной инъекции, однако, ее также можно вводить перорально.
Вводимое количество предлагаемой согласно изобретению вакцины на основе нуклеиновой кислоты зависит от рода введения и используемого вспомогательного средства. В случае внутримышечного введения это количество, как правило, выше, чем при внутрикожном введении, чтобы получить одинаковый защитный иммуноответ. Предпочтительно используемое количество в общем составляет 0,01 нг - 10 мг на дозу, чаще всего предпочтительно в пределах 1 нг - 1 мг на дозу.
Предлагаемая согласно изобретению вакцина на основе нуклеиновой кислоты имеет существенное преимущество в том, что не нужно использовать никаких систем культуры клеток для получения субвирусных частиц, а частицы образуются ин виво и, таким образом, могут прямо вызывать иммуноответ.
Предлагаемая согласно изобретению вакцина на основе нуклеиновой кислоты, далее, в противоположность вакцинам, которые основаны на протеинах, имеет преимущество в том, что она обладает большой стабильностью, в особенности, когда нуклеиновой кислотой является ДНК. В связи с этим вакцину можно хранить очень длительное время также без больших затрат на охлаждение, причем при этом хранении не нужно ожидать никаких значительных потерь активности. В особенности в лиофилизированной форме нуклеиновую кислоту можно хранить практически неограниченное время без всякого ущерба даже при комнатной температуре. Также реконструкцию лиофилизированной, предлагаемой согласно изобретению вакцины на основе нуклеиновой кислоты можно осуществлять намного проще, чем реконструкцию лиофилизированного протеинового раствора, который согласно изобретению часто на основании природы протеина может вызывать проблемы.
Следующее преимущество предлагаемой согласно изобретению вакцины на основе нуклеиновой кислоты заключается в том, что в противоположность обычным системам убитых прививочных материалов, иммунизирующий антиген синтезируется ин виво и приводит к возникновению эффективного клеточного иммуноответа (Science, т. 259, 19 марта 1993, с. 1691 - 1692, Research News: Naked DNA Points Way to Vaccines).
Согласно дальнейшему аспекту, изобретение относится к последовательностям нуклеиновых кислот, в особенности векторам нуклеиновых кислот, охватывающим всю кодирующую последовательность производимых от TBE-вируса протеинов E и prM/M, в качестве лекарственного средства. До сих пор в уровне техники еще не было сообщено ни о какой хотя бы одной успешной иммунизации нуклеиновой кислоты с помощью ДНК флавивируса. Применение в медицине этой нуклеиновой кислоты (векторов нуклеиновой кислоты) поэтому представляет собой неожиданную новую возможность использования. Под вектором согласно изобретению понимают любую индиферентную основу лекарственного препарата с нуклеиновой кислотой, как плазмиды, вирусы или транспозоны.
До сих пор описан лишь один, производимый от SN40, плазмидный вектор, который содержит последовательность производимых TBE-вируса протеинов prM/M и E (Allison и др. Virus-Genes, 8, (3) (1994), с. 187 - 198). Для этой SV-40-плазмиды, однако, не обсуждается возможность иммунизирующего действия.
Предлагаемые согласно изобретению плазмидные векторы отличаются прежде всего от до сих пор описанных плазмидных векторов своей пригодностью для иммунизации. Поэтому предлагаемые согласно изобретению плазмидные векторы находятся, в частности, в виде готовых к употреблению растворов или лиофилизатов в пригодном для введения шприце, соответственно, ампуле.
В случае плазмидных векторов оказались эффективными предлагаемые согласно изобретению промоторы, которые выбирают из группы CMV-, HSV-, RSV-, EBV-, β- актин-, адено-, MMTV-, hsp- и hGH-промоторов, причем как особенно эффективный оказался в особенности CMV-"Immediate Early" ("очень ранний") - промотор.
Согласно дальнейшему аспекту, настоящее изобретение относится к применению нуклеиновой кислоты, включающей всю кодирующую последовательность производимых от TBE-вируса протеинов prM/M и E, для получения вакцины.
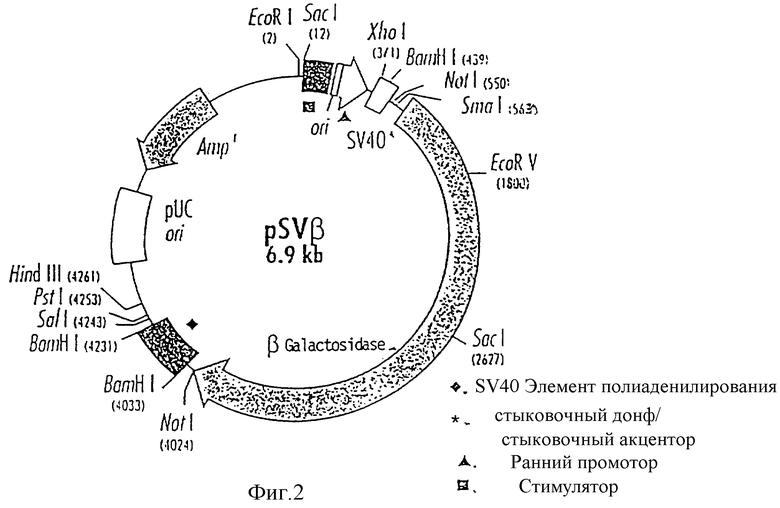
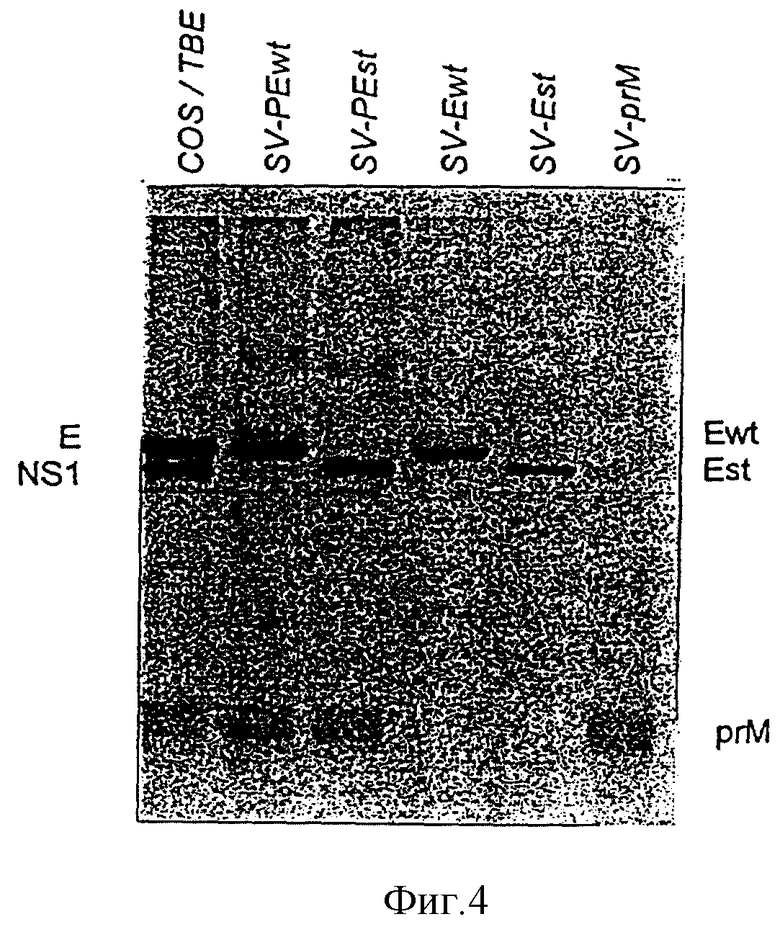
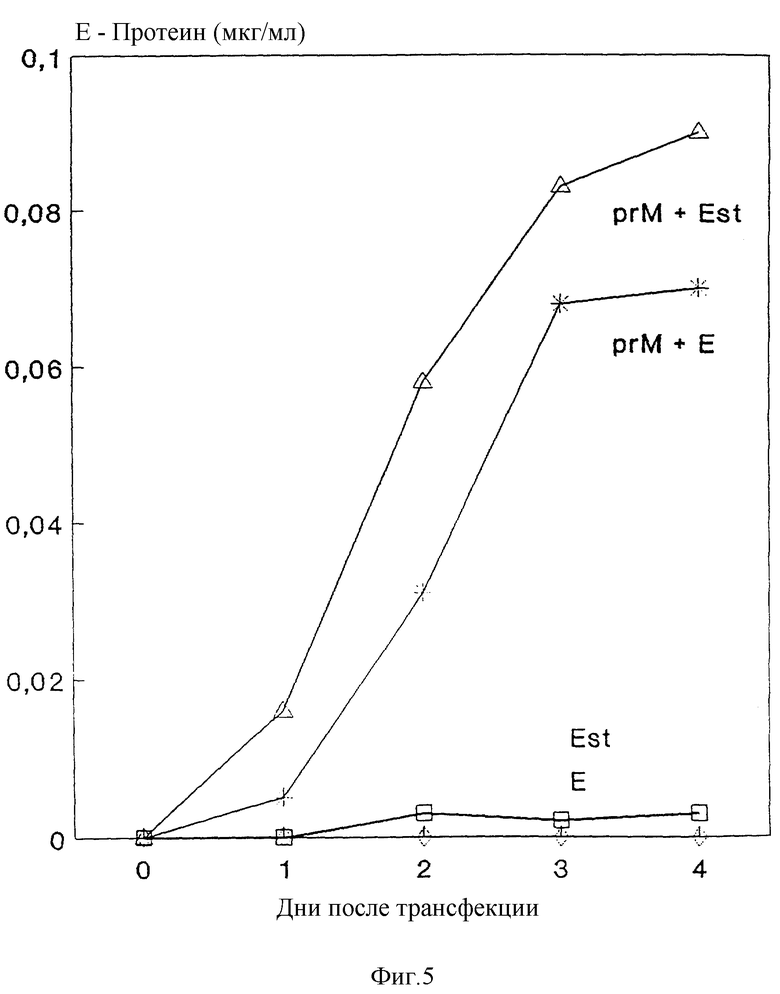
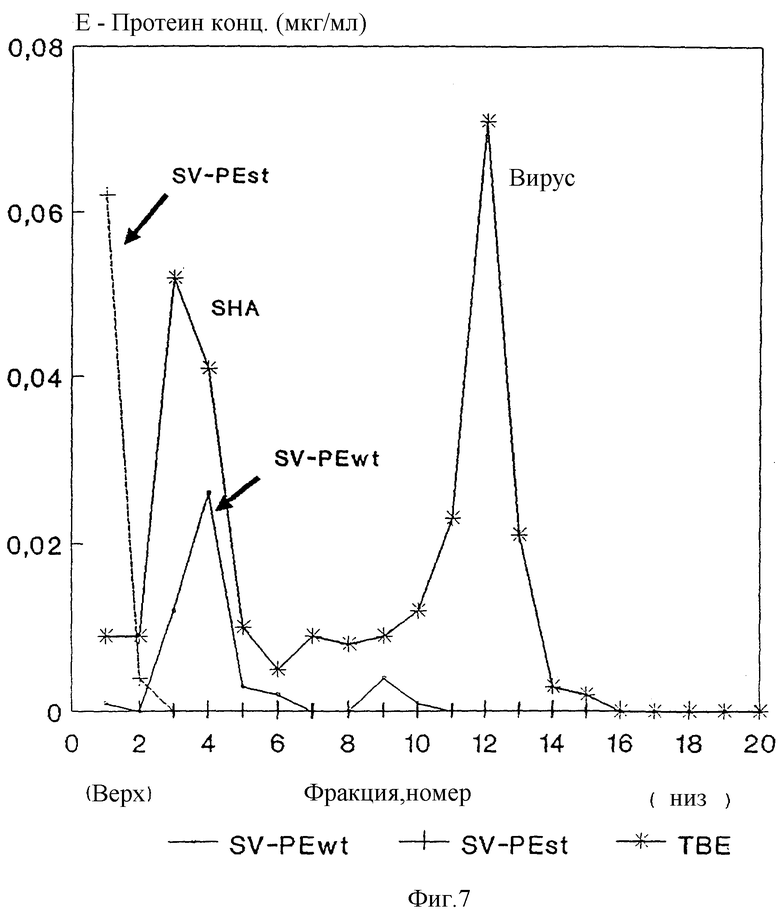
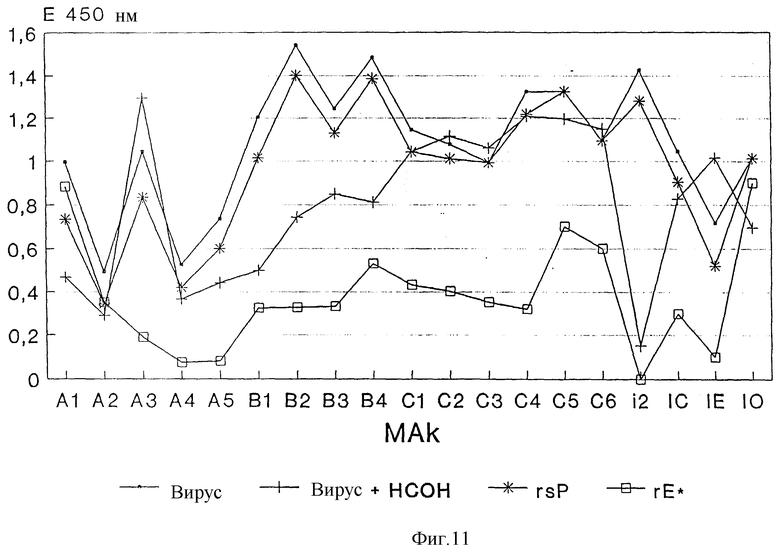
Изобретение поясняется еще подробнее ниже, руководствуясь примерами осуществления при ссылке на чертежи. Показано: на фиг. 1 - схематическое представление (изобретение) используемой в плазмиде экспрессии вставки; на фиг. 2 - схематическое представление плазмиды pSV β, которая применяется для экспрессии показанной на фиг. 1 конструкции; на фиг. 3a, b и c - полная нуклеотидная и аминокислотная последовательность представленной на фиг. 1 конструкции SV-PEwt; на фиг. 4 - иммунопреципитания солюбилизированных с помощью Triton X-100 лизатов клеток после трансфекции с помощью представленной на фиг. 1 конструкции, а также после инфекции с помощью TBE-вируса (COS/TBE); на фиг. 5 - диаграмма обнаружения протеина E с помощью 4-фазного метода ELISA в надосадочных жидкостях культуры COS-клеток, которые трансфецированы с помощью изображенных на фиг. 1 конструкций; на фиг. 6 - иммунопреципитация надосадочной жидкости культуры COS-клеток после трансфекции с помощью SV-PEwt, соответственно SV-PEst; на фиг. 7, на диаграмме, - седиментационный анализ надосадочной жидкости культуры COS-клеток после трансфекции с помощью SV-PEwt и SV-PEst, а также после инфекции с помощью TBE-вируса. Направление седиментации слева направо; на фиг. 8 представлен седиментационный анализ rSP без SV-PEwt), соответственно, после обработки с помощью 0,5% Triton X-100. Направление седиментации слева направо; на фиг. 9, на диаграмме, показано SDS-PAGE очищенного TBE-вируса и очищенного rSPs; окрашивание Coomassie-Blau; на фиг. 10 представлено сравнение rSPs из COS-клеток с таковым из стабильно трансфицированной линии CHO-клеток; на фиг. 11, на диаграмме, представлена реакционная способность 19 специфических к протеину E моноклональных антител при применении 4-фазного метода ELISA с TBE-вирусом, инактивированным формалином TBE-вирусом, rSP и rE*; на фиг. 12 представлена реакционная способность TBE-вируса, соответственно, rSP, при применении 4-фазного метода ELISA с mAks B3, i2 IC3 и C6, без (pH 8,0), соответственно, после обработки при pH 6,0; и на фиг. 13 представлена реакционная способность pE* при применении 4-фазного метода ELISA с mAKs B3, i2, IC 3 и С 6 без (pH 8,0), соответственно, после обработки при pH 6,0.
Изобретение подробнее описывается в нижеследующих примерах.
1. Приготовление частиц
Для этой цели конструируют 4 плазмиды экспрессии, которые содержат E-протеин, соответственно, лишенную мембранного якоря форму E-протеина одного, соответственно, вместе с prM (фиг. 1).
Пример получения этих плазмид описан у Allison и др., Virus Cenes, (3) (1994), с. 187 - 198.
Описанная там исходная плазмида pSV β представлена на фиг. 2.
В качестве вектора применяют pSV46 для конструкции клонов экспрессии. Этот вектор представляет собой производное базирующегося на SV40, эукариотного вектора экспрессии pSV β (Clontech), из которого путем расщепления с помощью Not 1 и регуляции удалена содержащая β- галактозидаза-ген вставка (Mac Gregor G.R. и Caskey C.T., Nucleic Acids Rec., 17, 2365, 1989). Часть полилинкера вниз от места прекращения (стоп-сайта) трансляции также удалена за счет переваривания с помощью XbaI и Hind III, заполнения по Кленову и регуляции.
В этот вектор встраивают PCR-продукты, которые получают следующим образом.
Синтетические олигонуклеотидные праймеры применяют для амплификации частей кДНК, соответственно, либо TBE-вирус - prM + E-области или одной E-области. Применяемые нуклеотидные координаты, согласно Mandl и др. (MandI C.W. , Heinz F. X. и Kunz C., Virology, 166, 197-205, 1988), в этом отношении перерабатывают так, что они содержат первые 20 нуклеотидов TBE-генома (Mandl C.W., Heinz F.X., Stockl E. и Kunz C., Virology, 173). 5'-Праймеры для rpM + E - и только для E-конструкций представляют собой 27-меры с последовательностями AAGCGGCCGCCATGGTTGGCTTGCAAA, соответственно, AAGCGGCCGCCATGGTTACCGTTGTGT. Первые 11 нуклеотидов каждой из последовательностей состоят из синтетической последовательности, содержащей сайт распознавания для фермента рестрикции Not 1 (GCGGCCGC), за которым следует последовательность из 16 нуклеотидов, соответственно, либо нуклеотидов 388 - 403 (SV-PE-ряд), либо 883 - 896 (SV-E-ряд). В каждом случае применяют естественно имеющийся ATC-кодон вверх от соответствующего гена в качестве стартового кодона, и праймер принимает такой вид, что он имеет ATG в пригодном контексте (GCCGCCATGG) для эффективного инициирования трансляции в COS-клетках (Kozak M., Cell, 44, 283 - 292, 1986, Kozak M., Mol. Biol., 196, 947 - 950, 1987). Такой же, длиной 28 нуклеотидов олигонуклеотид (ATGCGGCCGCTACTCATACCATACCATTTCTGAG) применяют в качестве 3'-праймера для обеих конструкций. Этот праймер на своем 3'-конце дополнен до нуклеотидов 2535 - 2550 и содержит на своем 5'-конце Not 1-сайт и комплемент (CTA) TAG - стоп-кодона в той же самой рамке считывания.
PCR - Амплификацию осуществляют по существу при стандартных условиях согласно Cetus-протоколу (Saiki R.K., Gelf and D.H., Stoffel S., Scharf S.J. , Higuchi R., Horn G.T., Mullis K.B. и Erlich H.A., Science, 239, 487 - 491, 1988). Реакционные смеси содержат 50 мМ KCl; 10 мМ (ммоль) ТРИС-HCl, pH 8; 3; 1,5 мМ M Cl2; смотря по обстоятельствам, 200 мкмоль dNTP, 10 нг каждого праймера, 10 мкл 1000-кратного разбавления кДНК-матрицы и 3 E AmpliTaq-полимеризы (Perkin Elmer Cetus) в общем объеме 100 мкл. Образцы покрывают с помощью 100 мкл парафинового масла и амплифицируют в аппарате с изменением температур (Temperaturwechsler) фирмы Перкин Элмер.
Образцы выдерживают 6,5 мин при 94oC, затем охлаждают до температуры ренатурации 40oC, прежде чем добавляют Taq-полимеразу. В целом осуществляют 35 циклов амплитфикации. Каждый цикл состоит из 1,5 мин при 94oC, 3 мин при 40oC и 7 мин при 68oC. Образцы очищают путем экстракции фенолом и хлороформом, осаждают этанолом и снова растворяют в стерильной, дважды перегнанной воде. Качество и количество PCP-продуктов определяют посредством электрофореза на агарозном геле, и образцы хранят при -20oC до тех пор, пока они не станут нужны для последующих стадий клонирования.
Полимеразную реакцию используют для того, что сконструировать минибанк кДНК-плазмидных клонов со вставками, которые соответствуют либо prM + E-области (SKPE-ряд), нуклеотиды 388 - 2550, либо E-области (SV-E-ряд), нуклеотиды 883 - 2550 генома TBE-вируса. (фиг. 1).
Так как структурные протеины TBE-вируса обычно получают путем процесса трансляции большого полипротеинового предшественника, по концам PCR-продуктов вводятся стартовые и стоп-сайты трансляции тем, что их встраивают в олегонуклеотидные праймеры. В каждую конструкцию вводится природный ATG-кодон, который располагается в пригодном контексте, чтобы служить в качестве инициирующего кодона, выше внутренней сигнальной последовательности, чтобы дать возможность протеканию естественного процесса за счет сигналазы хозяина и правильному отводу по секреторному пути. Подобным образом помещают TG-стоп-кодон в любой конструкции ниже сайта расщепления сигналазы, который образует карбокси - конец протеина E. Места распознавания для рестрикционного фермента Not 1 также встраиваются по концам, чтобы облегчить последующее клонирование PCR - продуктов.
PCR - ДНК разрезают с помощью Not 1 и по сайту Not 1-клонирования плазмидного вектора лигируют pSV 46, что позволяет корректно экспрессировать встроенные гены при регуляции предыдущего SV40-промотора (Mac Gregor G.P. и Caskey C. T. , Nuckeic Acids Res., 17, 2365, 1989). Вектор содержит также сигнал полиаденилирования для эффективной экспрессии и происхождения SV40-репликации, чтобы сделать возможной репликацию рекомбинантных плазмид в трансфектированных COS-клетках, которые конститутивно экспрессируют большой SV40-T-антиген (Gluzman Y., Cell, 23, 175 - 182, 1981).
Лигирующие смеси применяют для осуществления трансформации E.coli HB 101 и, выделяют отдельные, резистентные к ампициллину колонии, которые представляют собой отдельные клоны. Ориентацию вставки в каждой плазмиде определяют путем переваривания с помощью рестрикционных ферментов и электрофореза на агарозном геле. Клоны с правильно ориентированными вставками оставляют для дальнейшего анализа.
Плазмиды с правильно ориентированными вставками идентифицируют путем рестрикционного анализа и очищают путем CS C1-градиентного центрифугирования.
Для каждой плазмидной конструкции применяют очищенную двунитевую плазмидную ДНК из одного-единственного препарата для определения ДНК-последовательности. Реакции секвенирования осуществляют в аппарате с изменяющейся температурой фирмы Перкин Элмер при применении TBE-специфических праймеров, Ampli Taq-полимеразы (Perkin Elmer Cetus) и маркированных с помощью флуоресцирующего красителя дидезокситерминаторов, согласно инструкциям изготовителя (Applied Biosystems). Продукты реакций секвенирования анализируют на автоматическом ДНК-секвенаторе 377A Applied Biosystems (применяемой биосистемы).
Анализ, полученный путем полимеразной цепной реакции (PCR) реакции мутаций
Для того чтобы осуществить детальный анализ клонированных вставок и оценить эффективность мутагенеза с помощью Taq-полимеризы, выбирают 13 отдельных E.coli-клонов, которые содержат плазмиды SV-PE-серии, и 9 отдельных E. coli-клонов, которые содержат плазмиды SV-E-серии. Плазмидную ДНК из каждого клона очищают и анализируют путем полного секвенирования клонированных вставок в обеих нитях.
кДНК-последовательности затем сравнивают с соответствующей РНК-последовательностью родительского штамма TBE-вируса дикого типа Neudorfl (Mandl C. W., Heinz F.X. и Kunz C., Virology, 166, 197 - 205, 1988).
Как ожидают, каждый из плазмидных клонов содержит несколько различий в нуклеотидах по отношению к TBE-вирусной последовательности дикого типа. За некоторыми исключениями (см. ниже) большинство этих изменений представляют собой отдельные мутации, которые, очевидно, произошли во время PCR-амплификации и найдены только в одном - единственном клоне.
Как представлено в таблице 1, 22 секвенированных вставки (в целом 43 549 п. о. [пар оснований]) содержат 71 отдельное нуклеотидное изменение, причем эти изменения сводятся к недостатку Taq-полимеризы. Из этих изменений 42 (59%) приводят к замещениям в предсказанной аминокислотной последовательности, а 3 - к делециям, которые приводят к сдвигам рамок считывания (таблица 2).
Распределение мутаций сильно сдвинутого в пользу A/T ---> G/C и G/C ---> A/T переходных мутаций, как уже наблюдалось ранее другими (Dunning A.M., Talmud P. и Humphries S. E., Nucleic Acids Res., 16, 10393, 1988; Chen J., Sahota A. , Stambrook P. J. и Tischfield J.A., Mutat Res. 249, 169 - 176, 1991; Cadwell R.C. и Joyce G.F., PCR Methods Applic., 2, 28-33, 1992). Частота всех мутаций составляет 1,63 • 10-3 на пару оснований при амплификации в 35 циклов (4,7 • 10-5 на пару оснований и на цикл), что совпадает с опубликованными значениями для частоты недостатка Taq-полимеризы (15, 32). Средняя доля замены аминокислот составляет 0,46 на ген для prM (длина 164 аминокислоты) и 1,59 для E (длина 496 аминокислоты).
Спонтанная мутация в нуклеотиде 930 имеется во всех клонах, и предполагают, что она возникает как естественная мутация во время размножения вируса. Кроме того, установлено, что 5 клонов SV-PE-серия (SV-PE05, SV-PE07, SV-PE11, SV-PE12 и SV-PE13) вместе имеют 7 идентичных мутаций в нуклеотидах 849, 1285, 1346, 1404, 1909, 2247 и 2255. За исключением SV-PE10, который содержит 3 мутации (1909, 2247 и 2255), ни одной из них не найдено в других SV-PE-клонах или любых SV-E-клонах. Одно из этих изменений, нуклеотидная делеция в положении 1346, вызывает сдвиг рамки к кодону 125 в E-гене, и поэтому следовало бы ожидать, что они дают нефункциональный E-протеин. Так как содержащий эти же самые семь мутаций клон получается независимо от отдельных кДНК - и PCR-приготовлений (данные не приводятся), оказалось, что эти варианты уже имеются в вирусной РНК-популяции до амплификации. Эти особые мутации поэтому не включают в анализ полученных за счет PCR мутаций.
Конструкции, которые кодируют подвергнутые делеции и дикого типа E-протеины
Дополнительно к клонам, которые кодируют отдельные изменения аминокислот, нас интересовало также получение (prM + E) и только E-конструкций, которые кодируют подвергнутые делеции и дикого типа протеины. Так как, однако, все полученные согласно PCR клоны SV-PE-серии содержат мутации, которые ведут к изменениям в аминокислотной последовательности, применяют прямое субклонирование, чтобы получить конструкцию дикого типа, которая кодирует немодифицированные prM- и E-протеины, а также подвергнутую делеции форму только E-конструкции.
ДНК - Последовательность плазмиды SV-PE04, кроме двух спонтанных мутаций, показывает замещение A на T в нуклеотиде 2275, благодаря чему AAG-кодон для лизина 435 E-протеина изменяется до TAG-стопкодона (таблица 1). Поэтому предсказывается, что SV-PE04 кодирует prM-протеин дикого типа и подвергнутый делеции E-протеин, в котором отсутствуют карбокси-концевые в количестве 61 аминокислотные остатки, которые содержат предположительную закрепляемую мембраной (Membran-durchspannenden) область (Mandl C.W., Guirakhoo F., Holzmann H. , Heinz F.X. и Kunz C., J. Virol 63, 564 - 571, 1989). Оказалось, что плазмида SV-E01, в которой отсутствует большая часть prM-гена, содержит только спонтанную мутацию в E-гене, в связи с чем она кодирует E-протеин дикого типа.
Для создания prM + E-конструкции общей длины с аминокислотной последовательностью дикого типа и только E-конструкции без prM, однако, со стоп-кодоном в нуклеотиде 2275, обменивают рестрикционные фрагменты из SV-PE04 и SV-E01 (ниже называются SV-PEst, соответственно, SV-Ewt). Для этой цели применяют два участка расщепления для рестрикционного фермента Cfr101: один между нуклеотидами 960 и 961, вблизи границы prM - и E-генов, а другой - в резистентном к ампициллину гене вектора. Одна из полученных в результате плазмидных конструкций SV-PEwt, содержит prM-ген дикого типа из SV-PEst и E-ген дикого типа из SV-Ewt, в то время как другая, SV-Est, в которой отсутствует prM-ген, однако, содержит включающий стоп-кодон E-ген из SV-PEst. Эти изменения подтверждаются секвенированием вставок обеих плазмид. Эти обе конструкции дополняют серию из четырех плазмид экспрессии для сравнения структуры и переработки экспрессированных из конструкции дикого типа (SV-PEwt E-протеинов с такими, в которых отсутствует prM (SV-Ewt), E-якорная область (SV-PEst) или обе (SV-Est).
Полноценная последовательность PEwt-вставки представлена на фиг. 3a, b и c.
COS-Клетки трансфицируют с помощью пригодных плазмид и их анализируют на предмет экспрессии рекомбинантых протеинов следующим образом.
ДНК-Трансфекция
COS-1-Клетки (Cluzman Y. , Cell, 23, 175 - 182, 1981) выдерживают при 37oC и при 5% CO2 в "COS-среде", состоящей из Dulbecco's - MEM (Gibco - BRL), дополненной 10% зародышевой сыворотки крупного рогатого скота, пенициллином (100 E/мл) и стрептомицином (100 мкг на мл).
Плазмиды, содержащие клонированные TBE-гены, путем трансфекции через посредство липосом, при применении модификации способа Felgner и др. (Felgner P. L., Gadek T.R., Holm H., Roman R., Chan H.W., Wenz M., Northrop J.P., Ringold G. M. и Danielsen M., Proc. Natl. Acad. Sci. USA, 84, 7413 - 7417, 1987) с помощью аппарата BioRadGene Pulser вносят в COS-1-клетки. Липофектиновый реагент (Grbco - BRL) разбавляют в Opti - MEM-1 - редуцированной сывороточной среде (Gibco - BRL) до концентрации 20 мкг/мл и смешивают с равным объемом Opti MEM-1, содержащим 8 мкг/мл соответствующей плазмидной ДНК. На трансфекцию используют 3 мкг липофектина и 2 мкг плазмидной ДНК. Смесь оставляют стоять в течение 15 минут при комнатной температуре для образования комплексов ДНК-липофектин, и 0,5 мл этой смеси добавляют в каждое углубление пластины для культуры клеток (Nunc) с 24 углублениями, содержащими клетки, которые для удаления следов сыворотки из среды роста дважды промыты с помощью 1 мл Opti MEM.
Клетки инкубируют со смесью ДНК-липофектин в течение 5 ч при 37oC и в присутствии 5% CO2, с последующей добавкой 1 мл полной COS-среды, и инкубацию продолжают в течение ночи. Спустя 24 часа после трансфекции среду удаляют и заменяют свежей COS-средой, и инкубацию продолжают вплоть до примерно 46 часов после трансфекции, затем клетки либо фиксируют для иммунофлуоресцентного анализа, либо растворяют для анализа на внутрицеллюлярный антиген при помощи метода.
Способ инфекции COS-клеток с помощью TBE-вируса для иммунофлуоресцентного контроля по существу идентичен способу трансфекции, с тем исключением, что вместо плазмидной ДНК и липофектина TBE-вирус применяют в форме суспензии головного мозга мышат, которая 100-кратно разбавлена в Opti - MEM-1-среде.
Анализ экспрессированных протеинов
а. Иммунопреципитания лизатов клеток
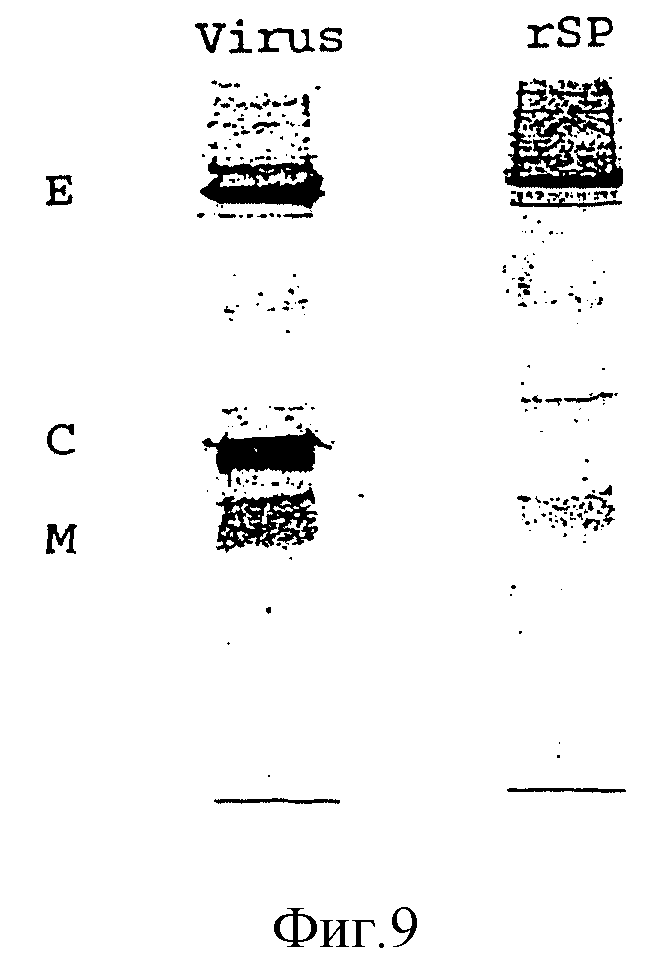
Трансфицированные клетки маркируют с помощью 35S-цистеина, солюбилизируют с помощью 1% Triton X-100 и подвергают иммунопреципитации с помощью поликлональной кроличьей сыворотки, которая специфична к TBE-вирусным E- и prM-протеинам. Как видно из фиг. 4, экспрессия конструкций SV-PEwt и SV-Ewt приводит к синтезу E-протеина аутентичной величины. Синтезированные из SV-PEt и SV-Est протеины, как ожидалось, вследствие своего C-концевого укорачивания, несколько меньше, чем E-протеин дикого типа.
б. Анализ на секрецию рекомбинантного протеина в надосадочной жидкости клеток
Надосадочные жидкости трансфецированных клеток количественно анализируют на наличие E-протеина в дни 0 - 4 после трансфекции при применении четырехфазного метода ELISA, как описано Heinz и др., J. Biol. Stand. (1986); 14: 133 - 141. Представленные на фиг. 5 результаты показывают, что только те конструкции, которые коэкспрессируют prM-протеин, приводят к секреции E-протеина. Иммунопреципитация из надосадочных жидкостей (фиг. 6) подтверждает, что отделенные протеины имеют такую же величину, как и соответствующие внутриклеточные протеины (см. фиг. 4).
Секреция рекомбинантных протеинов представляет собой огромное преимущество для производственных целей, так как таким образом отпадает необходимость лизиса клеток для получения желательного протеина и удаление заражающего клеточного материала. При пригодных условиях это также позволяет осуществлять непрерывный сбор рекомбинантного протеина, не разрушая клетки.
в. Характеристика отделенных рекомбинантных протеинов
Представленные на фиг. 5 надосадочные жидкости подвергают зональному центрифугированию с градиентом плотности по сахарозе при применении 5 - 30% (по массе) сахарозного градиента и ротора Beckman SW-40. Центрифугирование осуществляют при 3.000 об/мин и 4oC в течение 100 минут. Градиенты фракционируют и протеин E количественно определяют в каждой фракции с помощью четырехфазного метода (см. Heinz и др. J. Biol. Stand (1986) 14: 133 - 141).
В качестве контроля применяют надосадочную жидкость из инфицированных вирусом клеток, который дает два содержащих E-протеин пика (фиг. 7), из которых один соответствует полноценному вирусу, а другой так называемому неинфекционному "медленно седиментирующемуся гемаглютинину "slowly-sedimenting hemagglutinin" (SHA)) (см. Russel
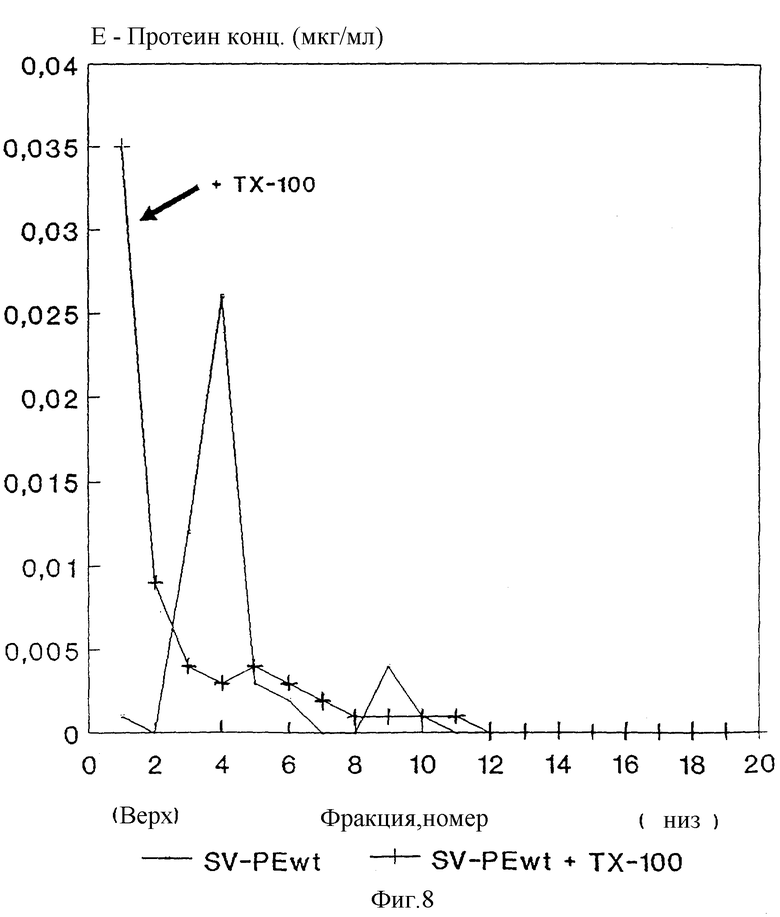
и др. , (1980), Chemical and antigenic structure of flaviviruses. B: Schlesinger, R.W. (изд.). The Togaviruses, 503 - 529, Academic Press). Надосадочная жидкость из трансфецированных с помощью SV-PEwt клеток содержит особую форму E со скоростью седиментации подобно вирусному SHA, в то время как укороченный на C-конце протеин E из SV-PEst не образует никаких стабильных частиц и находится сверху на градиенте.
Особую форму из SV-PEwt называют "рекомбинантная субвирусная частица" ("recombinant subviral particle", rSP"), а растворимую форму из SV-PEst называют "рекомбинантный E*" (rE*).
Обработка rSP с помощью 0,5%-ного Triton X-100 вызывает диссоциацию частиц, как показано путем седиментационного анализа на фиг. 8, что указывает на наличие липидной мембраны.
Приготовление rSP и rE*
а. Очистка rSP
Надосадочные жидкости трансфецированных с помощью SV-PEwt COS-клеток осветляют путем центрифугирования в течение 30 мин при скорости 10000 об/мин и при 4oC в высокоскоростной центрифуге Sorvall, и затем, путем центрифугирования при 44000 об/мин при 4oC и в течение 120 мин, частицы собирают, причем применяют ротор Beckman Ti 45. Содержание rSP в виде шариков фракцию снова суспендируют в TAN-буфере, с pH 8,0 и наносят на 5-20%-ный сахарозный градиент, который получают с помощью того же самого буфера. После центрифугирования в течение 90 мин при 3000 об/мин и 4oC при применении ротора Beckman SW 40 образцы фракционируют и "пиковые" фракции идентифицируют путем испытаний на HA-активность (Clarke и Casals, 1958, Aver. J. Trop. Med. Hyg. 7: 561 - 573). "Пиковые" фракции из зональных градиентов очищают далее посредством равновесного центрифугирования (35000 об/мин, 4oC, в течение ночи) и фракции снова идентифицируют с помощью HA. Общее содержание протеина E определяют количественно с помощью четырехфазного метода ELISA, а чистоту определяют при применении окрашивания Coomassie-Blau посредством SDS-PAGE (фиг. 9).
б. Приготовление rE*
Лишенные сыворотки надосадочные жидкости из SV-PEst - трансфецированных COS-клеток осветляют как описано выше и концентрируют путем ультрафильтрации примерно в 15 раз.
Получение линий клеток с помощью хромосомно-интегрированных prM и E-генов
Для получения стабильно трансфецированных клеток, которые продуцируют rSPs, применяют селекционную систему на основе дигидрофолатредуктазы (DHFR) (Current Protocols in Molecular Biology, John Wiley & Sons, Inc.), а также такую конструкцию, которая содержит полноценные гены для prM и Е-протеина (фиг. 1) и приводит к синтезу и секреции rSPs. Перенос DHFR-гена осуществляют с помощью плазмиды pSV2-dhfr (S. Subramani и др., Mol. Cell. Biol. 1, 854 - 864, 1981), перенос prM - E-генов осуществляют с помощью плазмиды CMV-PEwt, которую получают путем переклонирования вставки из плазмиды SV-PEwt в плазмиду pCMV β (Clontech).
DHFR-дефицитную линию клеток CHO-DG44 (Som. Cell. Mol. Get., 12, 555 - 666, 1986) выращивают на HAM-F 12-среде + 10% зародышевой телячьей сыворотки (FKS) и трансфицируют с помощью PCMV-PEwt, а также pSV2-dhfr в соотношении 20: 1 путем электропорации при применении аппарата BiORad Gene Pulser. Трансфецированные клетки сначала выращивают далее в течение 2-х дней в HAM - F-12-среде + 10% FKS, затем трипсинизируют и при высоком разбавлении, для получения отдельных, экспрессирующих DHFR и prM + E клонов клеток в селекционной среде (MEM -α без рибо- и дезоксипибонуклеотидов + 10% диализированной FKS), высевают на чашки Петри. Спустя примерно 10 дней отдельные видимые в микроскоп колонии клеток переносят в маленькие сосуды для культуры клеток и размножают далее. Те клоны клеток, которые продуцируют FSME-специфический антиген, идентифицируют путем анализа надосадочных жидкостей культуры клеток с помощью метода ELISA.
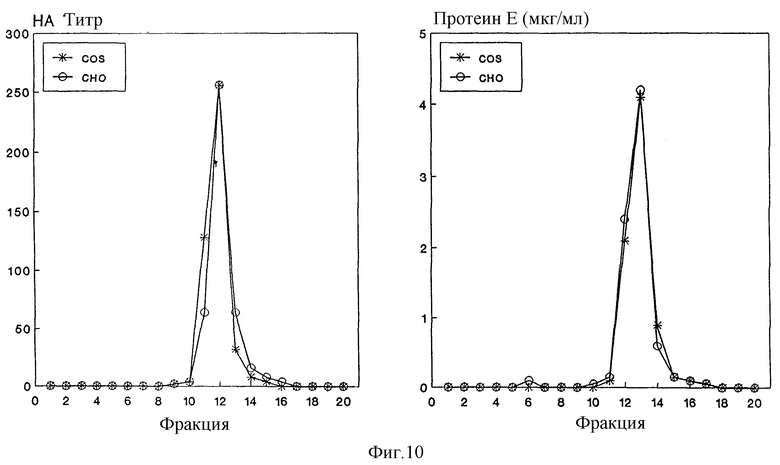
Один из этих клонов клеток (CHO-37) применяют для получения и охарактеризовывания продуцированного им rSPs. Для этой цели клетки выращивают в вышеуказанной селекционной среде и rSPs в надосадочной жидкости клеток подвергают такому же способу очистки, как и описанный для получаемых из COS-клеток rSPs. На фиг. 10 представлен результат центрифугирования с градиентом по плотности сахарозы, в случае которого rSPs из COS-клеток сравнивают с таковыми из стабильно трансфицированной линии клеток CHO-37. Применяют следующие условия центрифугирования: 20-50% сахарозы: Potop Beckman Ti-40; 35 000 об/мин; в течение ночи. После центрифугирования градиенты фракционируют и отдельные фракции анализируют по тесту на гемаглютинацию по методу ELISA на специфический к вирусу FSME антиген. Как следует из фигуры, оба препарата не обнаруживают никакого различия в седиментационной способности, соответственно, в удельной плотности, а также в специфической гемаглютинационной активности.
2. Характеристика в отношении нативной структуры протеина
Подобным образом полученные препараты подвергают следующим анализам:
a) Анализ антигенной структуры с помощью моноклональных антител;
b) Способность к индуцируемым кислотой конформационным изменениям;
c) Гемаглютинационная активность.
К a) Антигенная структура
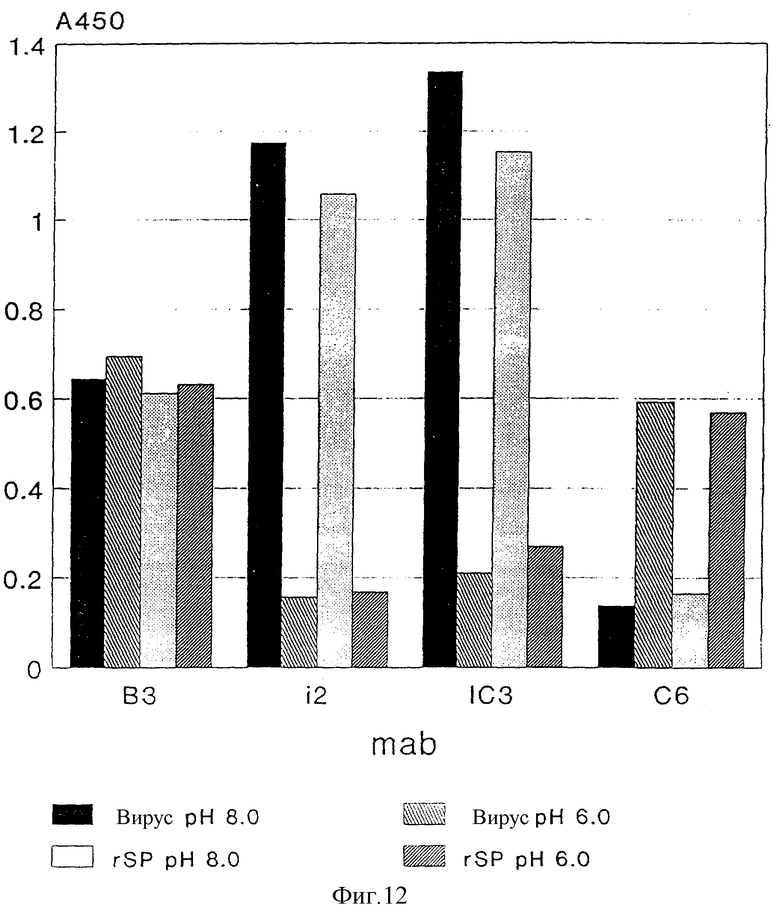
Антигенную структуру инактивированного формалином полноценного вируса, rSP и rE*, сравнивают с таковой нативного инфекционного вируса при применении 19 специфических к протеину E моноклональных антител (Mandl и др., 1989, J. Virology 63, 564 - 571; собственные испытания). Реактивность (реакционноспособность) каждого отдельного моноклонального антитела с каждым из вышеуказанных препаратов анализируют при применении 4-фазного метода ELISA, как описано Heinz и др. J. Biol. Stand. (1986), 14: 133 - 141. При этом используют все препараты антигенов с заранее определенной, постоянной концентрацией протеина E 1 мкг/мл и каждое из моноклональных антител применяют в разбавлении, при котором с нативным инфекционным вирусом получают величину экстинкции в пределах 0,5 - 1,6.
Как следует из фиг. 11, образец для испытания на реактивность практически идентичен с maks в случае rSP и инфекционным вирусом, так что можно исходить из того, что протеин E в rSPs находится в такой же нативной форме, как и в инфекционном вирусе.
Инактивация формалином, напротив, вызывает значительные изменения в антигенной структуре, к которой также относятся эпитопы, которые связывают нейтрализующие антитела. Это относится как к нейтрализующему mAk12 (Mandl и др. , 1989 J. Virol. 63: 564 - 571), эпитоп которого разрушается за счет обработки формалином, как и эпитопы в области домена A, а также в домене B (Mandl и др. , 1989, J. Virol. 63: 564 - 571), которые обладают по меньшей мере пониженной реакционностью. Этот анализ также показывает, что E* по своей структуре никоим образом не соответствует нативному протеину E на поверхности вируса.
К b) Индуцируемые кислотой конформационные изменения
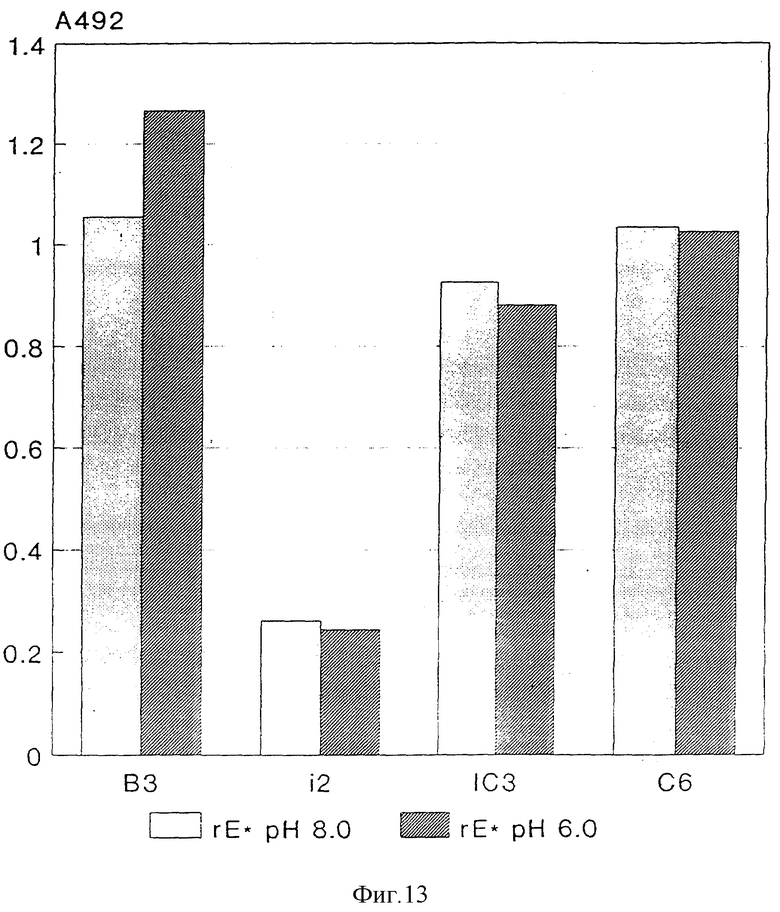
TBE - Вирус с помощью являющейся посредником к рецептору эндоцитозы проникает в клетки и благодаря этому попадает в эндосомы, кислое значение pH (< 6,4) которых индуцирует специфическое изменение конформации, которое необходимо для слияния вирусной мембраны с эндосомной мембраной и таким образом для инфекционности вируса. Эти структурные изменения могут иметь следствием изменение реактивности отдельных mAks (Heinz и др. 1994, Virology, 198: 109 - 117) и между прочим касаются эпитопов i2 и IC3 (сильное уменьшение реактивности), соответственно, C6 (усиленная реактивность), но не эпитопа B3.
Эти изменения существенны для функции протеина E. Способность реагировать описанным образом в кислой области pH поэтому также представляет собой критерий того, находится ли протеин E в нативной, соответствующей инфекционному вирусу форме. Препараты rSP, rE* и инфекционного вируса в триэталаминном буфере, pH 8,0 с помощью буфера, содержащего 0,05 М mES, 0,1 М NaCl и 0,1% бычьего альбумина в виде смеси, так что возникает pH - значение 6,0, инкубируют в течение 10 мин при 37oC и затем с помощью триэтаноламина снова устанавливают pH 8,0. Затем определяют реактивность этих препаратов до и после инкубации при кислом значении pH с помощью mAKsB 3 i2, IC 3 и C6 при применении 4-фазного метода ELISA, как описано в разделе "Антигенная структура". Результаты представлены на фиг. 12 и 13 и показывают, что кислое значение pH в протеине E из rSPs вызывает такие же структурные изменения, как и в инфекционном вирусе (см. фиг. 12), что, однако, при таком же анализе в случае E* не дает никоим образом указания на подобные перемещения (см. фиг. 13).
К c) Специфическая гемаглютинационная активность
Характерным свойством инфекционного FSME-вируса является его способность при определенных условиях агглютинировать гусиные эритроциты (гемалютинационная активность). Это свойство сообщается вирусу через протеин E и, таким образом, является дальнейшим индикатором для обнаружения нативной функциональной структуры этого протеина. Тесты на гемаглютинацию осуществляют как описано Clarke и Casals (1958), Amer. J. Trop. Hyg. 7: 561 - 573, при применении гусиных эритроцитов при pH 6,4.
В инфекционном вирусе, инактивированном формалином вирусе, rSP и rE*, устанавливают определенное по методу ELISA содержание антигена 5 мкг/мл и анализируют в тесте на гемаглютинацию. Результат представлен в нижеследующей таблице 3. Из этой таблицы следует, что rSP имеет такую же удельную гемаглютинирующую активность, как и инфекционный вирус, и что эта активность через посредство протеина E почти полностью теряется при инактивации формалином. Препарат rE* не обладает никакой измеримой гемаглютинирующей активностью (HA-активностью);
Таблица 3. Гемаглютинирующая активность
Препарат - HA-тест
Инфекционный вирус - 512
Инактивированный формалином вирус - 4
rSP - 512
eE* - < 2
3. Иммунногенность в случае мышей
Была проанализирована иммуногенность следующих антигенных препаратов путем иммунизации мышей:
- инактивированный формалином, очищенный вирус;
- rSP
- rE*
Содержание протеина E определяют путем ферментно-иммунного анализа (Heinz и др. , 1986, J. Biol.-Stand., 14: 133 - 141) и во всех препаратах устанавливают одну и ту же концентрацию антигена 5 мкг/мл. Соответственно используемой в настоящее время TBE-вакцине (FSME-Immun*) применяют в качестве буфера разбавления PBS pH 7,4, содержащий 0,1% человеческого альбумина, и добавляют 0,2% Al(OH)3 в качестве вспомогательного средства.
Иммуногенность rSP и rE* испытывают также без добавки адъюванта.
Протокол иммунизации
С помощью каждого из препаратов подкожно иммунизируют дважды, с интервалом в 14 дней, группы по 10 мышей Swiss albino весом 15 г (5 самок и 5 самцов), причем на одну мышь и одну иммунизацию вводят 0,2 мл антигена. Спустя неделю после второй иммунизации осуществляют отбор крови и всех мышей инфицируют интраперитонально с помощью 500 ЛД50 патогенного для мышей штамма TBE-вируса Hypr (Challenge тест). Мышей наблюдают в течение промежутка времени 14 дней на появление приводящего к гибели энцефалита.
Идентичные аликвоты сыворотки каждой отдельной мыши из 10-й группы, которые иммунизированы с помощью того же самого иммуногена, объединяют и анализируют на содержание специфических к TBE-вирусу антител в ферментном иммунотесте (ELISA), в тесте на подавление гемаглютинации (HHT) и тесте нейтрализации.
ELISA осуществляют при применении очищенного TBE-вируса в качестве антигена, как описано Heinz и др. (1984), J. Gen. Virol. 65: 1921 - 1929. HHT осуществляют как описано Clarke и Casals (1958). Amer. J. Trop. Med. Hyg. 7: 561 - 573, причем используют гусиные эритроциты при конечном значении pH 6,4. Для теста по нейтрализации применяют ВНК-21 клетки и концентрацию вируса 500 инфекционных единиц.
Результаты опыта по иммунизации представлены в таблице 4. Из этой таблицы следует, что rSP как без, так и адъювантами, точно так же, как инактивированный формалином полноценный вирус, защищает всех мышей от смертельной дозы TBE-вируса. В противоположность этому, после иммунизации с помощью rE* нельзя наблюдать никакого (без адъювантов) или только можно наблюдать минимальное защитное действие (с адъювантами).
В отношении индукции антител, rSPs даже превосходит инактивированный формалином вирус, особенно отчетливо при применении в качестве вспомогательного средства Al(OH)3. Это касается как обнаруживаемых по методу ELISA связывающих вирус антител, так и измеряемых в HHT и тесте на нейтрализацию, еще более существенных для защищающего иммуноответа, антител, которые, следовательно, блокируют специфические функции вируса (гемаглютинацию, соответственно, инфекционность).
В соответствии с отсутствующей незначительной защитной эффективностью также индукция антител за счет rE* очень низка, соответственно, не обнаруживается.
Следовательно, оказалось, что rSRs представляет собой превосходные иммуногены, которые защищают от, в противном случае приводящего к смерти, энцефалита и в отношении индукции функциональных, нейтрализующих вирус антител даже превосходят инактивированный формалином полноценный вирус.
4. Иммунизация с помощью "голой" ДНК
Для иммунизации с помощью "голой" ДНК применяют плазмиды, которые содержат представленные на фиг. 1 вставки при контроле раннего CMV-промотора. Эти плазмиды (CMV-PEwt, CMV-PEst, CMV-Ewt, CMV-Est) получают путем переклонирования вставок из описанных плазмид SV-PEwt, SV-PEst, SV-Ewt и SV-Est в плазмиду pCMVβ (Clontech) (Mac Gregor и др. Nucleic Acids Reseach, IRL Press, т. 17, N 6, 1989, с. 2365: "Construction of plasmids that express E. coli β- galactosidase in mammalian cells" и очищают путем центрифугирования по градиенту плотности CSC1 (Maniatis et al., Molecular Cloning: A Laboratory Manual. Cold Spring Harbor Laboratory, Cold Spring Harbor, N 4, 1982).
CMV-PEwt содержит последовательности протеинов дикого типа prM и E.
CMV-PEwt содержит последовательности для протеина дикого типа prM и делетированную последовательность протеина E, причем кодоны для 61-й карбоксиконцевой аминокислоты, которые образуют мембранный якорь протеина E, отсутствуют.
В CMV-Ewt отсутствует большая часть prM-последовательность, однако имеется полноценная последовательность протеина E дикого типа.
CMV-Est не содержит никакой prM-последовательности и делетированной протеин E-последовательности из CMV-PEst.
Каждую из этих четырех плазмид используют в двух концентрациях (60 мкг/мл и 300 мкг/мл в PSB) для иммунизации мышей (Swiss albino), причем для каждого препарата плазмиды используют 10 мышей (5 самок и 5 самцов). Каждой мыши внутрикожно инъецируют 2 раза, с интервалом в 2 недели, 100 мкл соответствующего ДНК-препарата.
В качестве контролей используют равным образом плазмиду pCMV β в концентрации 300 мкг/мл, а также PBS. Для сравнения с обычной иммунизацией группы по 10 мышей иммунизируют также 2 раза, с интервалом в две недели, с помощью инактивированного формалином вируса в концентрациях 1 мкг/мл - 5 мкг/мл, причем инъекцируют 0,2 мл на мышь подкожно и применяют 0,2%-ный Al(OH)3 в качестве адъюванта. Спустя одну неделю после второй иммунизации, для обнаружения специфических антител по методу ELISA, у мышей берут кровь и одновременно осуществляют интраперитонеальную Challenge-инфекцию с помощью 500 ЛД50 - TBE-вируса. Время наблюдения составляет 3 недели. Результаты обнаружения антител и испытания по защите представлены в таблице 5. Как следует из этой таблицы, в случае иммунизации ДНК достигается защита против смертельной инфекции с помощью TBE-вируса только с той плазмидой, которая содержит гены для полноценных prM - E-протеинов. Это, вероятно, нужно интерпретировать в том отношении, что необходимо присутствие prM/M-протеина для совместного конструирования подвирусных частиц, в которых находится протеин E в иммуногенной форме.
| название | год | авторы | номер документа |
|---|---|---|---|
| к-ДНК ДЛЯ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО ВИРУСА КЛЕЩЕВОГО ЭНЦЕФАЛИТА (TBEV) | 1995 |
|
RU2202612C2 |
| ЭФФЕКТИВНЫЕ ВИРУСОПОДОБНЫЕ ЧАСТИЦЫ (VLPS) ГРИППА | 2004 |
|
RU2369405C2 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК PC-NS3, ОБЕСПЕЧИВАЮЩАЯ ИНТЕГРАЦИЮ КОМПЛЕКСА ГЕНОВ C, PRM, E, NS1, NS2A, NS2B, NS3 ВИРУСА КЛЕЩЕВОГО ЭНЦЕФАЛИТА (ВКЭ) В ГЕНОМ ВИРУСА ОСПОВАКЦИНЫ (ВОВ) И РЕКОМБИНАНТНЫЙ ШТАММ ВИРУСА ОСПОВАКЦИНЫ, ЭКСПРЕССИРУЮЩИЙ В КЛЕТКАХ ИММУНИЗИРОВАННОГО ОРГАНИЗМА КОМПЛЕКС ГЕНОВ C, PRM, E, NS1, NS2A, NS2B, NS3 ВИРУСА КЛЕЩЕВОГО ЭНЦЕФАЛИТА | 1995 |
|
RU2112038C1 |
| РЕКОМБИНАНТНАЯ ВАКЦИНА ПРОТИВ ВИРУСА АФРИКАНСКОЙ КАТАРАЛЬНОЙ ЛИХОРАДКИ | 2007 |
|
RU2446823C2 |
| Рекомбинантная плазмида pHis6-flagG-protE, обеспечивающая синтез рекомбинантного химерного белка, включающего эпитопы гликопротеина Е вируса клещевого энцефалита и флагеллин G S.typhii и используемого в качестве основы для вакцины против вируса клещевого энцефалита | 2018 |
|
RU2702716C2 |
| СПОСОБ ПОЛУЧЕНИЯ ИММУНОГЕННОЙ КОМПОЗИЦИИ НА ОСНОВЕ ТРЕХ ГИБРИДНЫХ БЕЛКОВ ОБОЛОЧКИ ВИРУСА КЛЕЩЕВОГО ЭНЦЕФАЛИТА, ОПРЕДЕЛЯЮЩИХ ПРИНАДЛЕЖНОСТЬ К СИБИРСКОМУ (DBD2-D3S), ЕВРОПЕЙСКОМУ (DBD2-D3E) И ДАЛЬНЕВОСТОЧНОМУ (DBD2-D3D) ПОДТИПАМ ВИРУСА; РЕКОМБИНАНТНЫЕ ПЛАЗМИДЫ pDBD2-D3S, pDBD2-D3E И pDBD2-D3D; ШТАММЫ-ПРОДУЦЕНТЫ Escherichia coli M15 [pREP4]; ХИМЕРНЫЕ БЕЛКИ И ИХ ПРИМЕНЕНИЕ | 2014 |
|
RU2560588C1 |
| Иммунобиологическое средство для профилактики заболеваний, вызванных вирусом клещевого энцефалита на основе рекомбинантного вируса рода Flavivirus | 2022 |
|
RU2795800C1 |
| ВАКЦИНЫ ПРОТИВ ВИРУСОВ ГЕПАТИТА | 1995 |
|
RU2189254C2 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК PUR 291-HAV 22, КОДИРУЮЩАЯ СИНТЕЗ ГИБРИДНОГО ПОЛИПЕПТИДА АНТИГЕННЫХ ДЕТЕРМИНАНТ ВИРУСА ГЕПАТИТА А С β -ГАЛАКТОЗИДАЗОЙ | 1986 |
|
SU1380209A1 |
| Кассета, предназначенная для получения плазмидных векторов, используемых для создания клеток-продуцентов вирусоподобных частиц (ВПЧ) вируса гриппа | 2018 |
|
RU2680703C1 |











Изобретение относится к области медицины. В изобретении описана вакцина для иммунизации против ТВЕ-вирусных инфекций, включающая неинфекционные подвирусные частицы, которые охватывают протеин Е и в случае необходимости протеин prM/M; вакцина, включающая нуклеиновую кислоту, которая охватывает протеин Е и протеин prM/M, производимые от ТВЕ-вируса, причем протеин Е находится, соответственно кодируется по меньшей мере, по существу, в полноценной нативной форме. Изобретение расширяет арсенал средств для борьбы с флавивирусной инфекцией. 2 с. и 8 з.п.ф-лы, 13 фиг., 5 табл.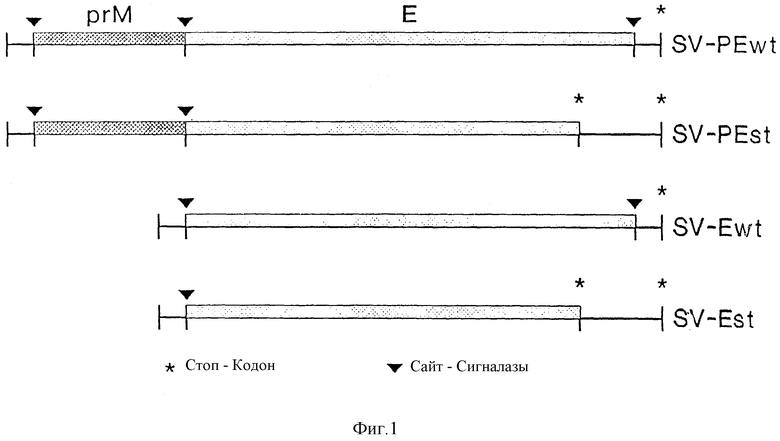
| Прибор для равномерного смешения зерна и одновременного отбирания нескольких одинаковых по объему проб | 1921 |
|
SU23A1 |
| Способ приготовления сернистого красителя защитного цвета | 1921 |
|
SU84A1 |
| Konishi S | |||
| et al | |||
| Virology | |||
| Поршень для воздушных тормозов с сжатым воздухом | 1921 |
|
SU188A1 |

